AND THE POSSIBLE ROLE OF THE INTEGUMENT
|
|
|
- August Carter
- 5 years ago
- Views:
Transcription
1 COMPARATTVE BIOLOGY OF S ALINITY DISCRIMINATION IN CROCODILIANS AND THE POSSIBLE ROLE OF THE INTEGUMENT Katherine Jackson A thesis submitted in conformity with the requirements for the degree of Master of Science Graduate Department of Zoology University of Katherine Jackson 1995
2 National Library 1*1 of Canada Acquisitions and Bibliographie Services Bibliothèque nationale du Canada Acquisitions et services bibliographiques 395 Wdiington Street 395, rue Wdlingtm OttawaON KlAON4 OttawaON KlAON4 canada Canada The author has granted a nonexclusive licence aüowing the National Library of Canada to reproduce, loan, distribute or sell copies of this thesis in microform, paper or electronic formats. The author retaias ownership of the copyright in this thesis. Neither the thesis nor substantial extracts fiom it may be printed or otherwise reproduced without the author's permission. L'auteur a accordé une Licence non exclusive permettant à la Bibliothèque nationale du Canada de reproduire, prêter, distribuer ou vendre des copies de cette thèse sous la forme de microfiche/nlm, de reproduction sur papier ou sur format électronique. L'auteur conserve la propriété du droit d'auteur qui protège cette thèse. Ni la thèse ni des extraits substantiels de celle-ci ne doivent être imprimés ou autrement reproduits sans son autorisation.
3 TABLE OF CONTENTS TABLE OF CONTENTS... i ABSTRACT... iv LIST OF FIGURES... vii LISTOFTABLES... x LIST OF ABBREVIATIONS... xi CHAPTER 1 : General Introduction... 1 Osmoregul atory S trategies of Marine Animais... 2 Marine Adaptation versus Estuarine Adaptation... 6 Osmoregulation in Estuarine Crocodilians... 9 Permeability of the Integument... 9 The renalcloacal system Lingual salt-secreting glands The role of selective drinking Phylogenetic trends... 15
4 Objectives of the thesis CHAPTER 2: Habitat and Phylogeny Influence Salinity Discrimination Abstract Introduction Lack of Salinity Discrimination in Caimm crocodilus Materials and Methods Resuits and Discussion Influence of Habitat Materials and Methods Results and Discussion Influence of Phylogeny Materials and Methods Results and Discussion General Discussion CHAPTER 3: Evidence for Integumenial Chemoreception in C. porosus Abstract Introduction... 49
5 Materials and Methods Results Discussion CHAPTER 4: Morphology and Ultrastructure of a Putative Integumentary Sense Organ Abstract Introduction Materials and Methods SEM Light microscopy and TEM Results Gross morphology SEM Light rnicroscopy and TEM Discussion C'EbWER 5: Conclusions and Directions for Future Research... 77
6 ABSTRACT COMPARATIVE BIOLOGY OF SALINITY DISCRIMINATION IN CROCODILIANS AND THE POSSIBLE ROLE OF THE INTEGUMENT Master of Science, 1995 Katherine Jackson Department of Zoology, University of Toronto This study investigated the mechanisms underlying salinity discrimination in crocodilians. The results are as follows: (1) In crocodilians, the capacity for salinity discrimination has a phylogenetic component. A11 crocodylids tested discriminated between salinities and did not drink hyperosmotic sea water. In alligatorids, however, only individuals collected in an estuarine area were capable of salinity discrimination. (2) The salinity discrimination organ of the estuarine crocody 1 id, Crocodylus porosus, is not located in the mouth. Dehy drated crocodiles discriminate very precisely between salinities and do not drink hyperosmotic sea water when they are immersed in it. However, they drink strongly hyperosmotic sea water when it is dripped directly ont0 their tongues. Crocodiles must therefore have a salinity discrimination organ on another part of the body. (3) Crocodylids but not alligatorids have sensory organs on their post-cranial scales. The morphoiogy and ultrastructure of pits on the scales of C. porosus was studied using SEM, TEM, and light rnicroscopy. Their structure is consistent with a sensory function. Physiological shidy will be required to determine whether their function is mechanosensory or chemosensory.
7 It has been an exciting year. Now in this totally inadequate space 1 will attempt to thank the many people who have helped me out dong the way. First of ail 1 wish to thank my supervisor, Prof. D. G. Butler, for providing suppon while keeping me on a long leash when it came to research. 1 am especially grateful to him for sending me ail the way to Singapore to follow up what was really just a hunch, and for tolerating the encroachment of dip-nets and crocodiles arnong the glassware and eels in his lab. As my project dnfted îùrther and further away from comparative endocrinology, 1 came to rely more and more on my three adjunct supervisors. 1 wish to thank Dr. D. R. Brooks for an inspiring introduction to the field of comparative biology, Dr. J. J. B. Smith for his insights into sensory biology and experirnental design, and Dr. J. H. Youson for advice on the interpretation of TEMs and for playing the useful role of devil's advocate in the chemoreceptor-mechanoreceptor debate. Many experts --herpetologists, morphologists, and physiologists --took time from their own work to offer advice, suggest techniques, send me offprints, and answer my letters and . 1 am especially grateful to T. H. Fritts, C. Gans, G. C. Grigg, S. Hillyard, H. Hong, J. Lang, P. Maderson, N. Mrosovsky, 1. Orchard, T. Parsons, C. A. Ross, R. Wassersug, R, Stephenson, H. D. Sues, A. Summers and G. Zug. Toronto is not the ideal location for a crocodile biologist, and 1 have been fortunate in king able to visit places where crocodilians can be found. In Singapore, Prof. T. J. Lam and his students generously provided laboratory space at the National University of Singapore. Mr. Yap Boon Chark, Pnmary Production Department, helped me through the bureaucratie maze associated with exporting crocodiles. Most of dl, 1 would like to thank Mr. and Mrs. Lee Bak Kuan, owners of the Long Kuan Hung Crocodile Farm, and their daughters Peilin and Peihui who welcomed me like one of the family. Mr. Lee showed me more crocodiles, from eggs to nesting adults, than 1 would have thought possible in a month. Mrs. LRe taught me the secret to rearing hatchlings successfully, and Peilin and Peihui devoted most of their summer holiday to helping with
8 my experiments and showing me Singapore. Also a big thankyou to Peilin, Mabel. and the girls who helped get me and the crocs past the daunting Mr. Kwon and ont0 Our flight home. At the St. Augustine Alligator Farm, in St. Augustine, Florida, Kent Vliet let me experiment on valuable and exotic hatchlings. At Sape10 Isiand, Dr. J. Alberts, Director of the University of Georgia Marine Institute, made an exception for me to his usuai rule about herpetologists, Charles Durant and Mary Price provided a warm welcome to the island, and Greg Balckom and the Wildlife Management Students from Archibaid Baldwin Agricul tural College heiped me coilect alligators. In spite of al1 this travelling, 1 acnially spent most of the ycar in Toronto. Essential technical assistance was provided by N. White, S. Norwood, R. Villadiego, E. Lin, and E. Knapp, often at very short notice. My labmates, Gavin, Donald, and Colin were very good Company (even though they won? stop teasing me about chiggers). 1 want to thank al1 rny friends in the Deparment. 1 know I'm going to miss you ail next year. 1 especially want to thank Tuhin, Stevie and Jose for coffee, beer. moral support, ruthless proofreading, tiring runs through deep snow, and (a mal test of friendship) reading my entire thesis. This research was funded by an NSERC operating grant to D. G. Butler. Persona1 support was provided by an Ann Sheppard Memorial Scholarship in Biology, from University College. Finally, 1 wish to thank the crocodiles. We've been through a lot together this year -Singapore customs, the baggage scanner in the Seoul airport, vaseline. minnows, CoCl,, and bags and bags of Instant Ocean Sea Salt. 1 wish them long and happy lives in their new home at the St. Augustine Alligator Fm.
 osmoconfoming, so that the M y fluids have an ionic composition similar to that of sea water, (B)")
9 LIST OE FIG- Fig Distribution of osmoreplatory strategies across taxa The three strategies are as follows: (A) osmoconfoming, so that the M y fluids have an ionic composition similar to that of sea water, (B) osmoregulating by concentrating organic osmons in the body fluids, and (C) osmoregulating by excreting NaCl and maintaining a plasma osmolality below that of sea water Fig Independent derivations of salt-secreting glands in marine and estuarine lineages of non-mamrnalian amniotes Fig Mass of water ingested by Cairnan crocodilus (n=6) foiiowing dehydration. Values are mean +/- s.e.m. Differences in arnount dnink at different salinities are not significant (P>0.05, ANOVA, Scheffe test) Fig Body mass in gram (mean +/- s.e.rn.) of Ca. crocodilus (n=9), and mass of 20 ppt sea water ingested as a percentage of initial body mass (mean +/- s.e.m.): initial mass (prior to dehydration), dehydrated mass (following dehydration), mass after 15 minutes exposure to sea water, and after 75 minutes exposure. ' hdicates significant increase in mass (P4.001, rrnanova) Fig. 23. Body mass and water ingested (mean +/- s.e.m.) by dehydrated ALLigator mississipiensis (n=10) from a captive, freshwater population before and after 15 min exposure to (a) 30 ppt sea water and (b) fresh water. ' Indicates a significant mass increase from the dehydrated condition (Pd.OO1, t-test). The amount drunk by the group exposed to 30 ppt sea water is not significantly different from the amount dmnk by the group exposed to fresh water (t-test, PzO.05) Fig Body mass and water ingested (mean +/- s.e.m.) by dehydrated A. mississipiensis (n=3) from an estuarine population, before exposure to water, after 15 min exposure to 30 ppt sea water, and after 15 min exposure to fresh water. ' Indicates a significant mass increase relative to the mass increase in sea water (paired t-test, P<0.025) Fig Body mass and water ingested (mean +/- s.e.m.) by dehydrated Crocodylus porosus (n=6), before exposure to water, after 15 min exposure to 30 ppt sea water, and after 15 min exposure to fresh water. ' Indicates a significantly pater mass increase relative to the mass increase in sea water (paired t-test, P<0.005) Fig Body mass and water ingested (mean +/- s.e.m.) by dehydrated CrocodyLus siamiensis (n=lo) from a captive, freshwater population (a) before and after 15 min exposure to 30 ppt sea water, and (b) before and after 15 min exposure to fresh water. ' Indicates a significantly greater mass increase in fresh water relative to the mass increase in 30 ppt sea water (t-test, P4.005) vii
10 Fig Body mass and water ingested (mean 4- s.e.m.) by dehydrated Osteolaemus tetraspis (n=3), before exposure to water, after 15 min exposure to 30 ppt sea water, and after 15 min exposure to fresh water. ' Indicates a significantly pater mass increase relative to the mass increase in sea water (paired t-test, P4.05) Fig The phylogenetic distribution of salinity preference among crocodilian species examined. (F/S) indicates animals which drank both fresh water and sea water, and (F/-) indicates those which drank fresh water but not sea water Fig Two equally parsimonious hypotheses for the evolutionary significance of salinity preference in crocodilians. In (a) lack of salinity discrimination is the plesiomorphic condition for crocodilians, in (b) it is the derived condition in freshwater alligatorids. (F/S) indicates drinking of both fresh water and sea water; (F/-) indicates drinking of frcsh water but not sea water Fig Methodology for Experiment 1. F=fresh water, S=30 ppt sea water. Dehydrateci crocodiles drink when immersed in fresh water (a). do not dnnk when immersed in 30 ppt sea water (b), drink 30 ppt sea water from a bottle (c), drink 30 ppt sea water from a bottle while immersed in fresh water (d), and drink fresh water from a bottle when immersed in 30 ppt sea water Fig A dehydrated C. porosus drinking hyperosmotic sea water from a wash bonle Fig (A) Ventral scales of a crocodylid, C. porosus, showing the pits (arrow). (B) Ventral scales of an alligatorid, A. mississipiensis, which lacks pits Fig Touch papillae (sensu von During, 1973) from the cranid scales of C. porosus: (A) head of a crocodile with touch papillae indicated (arrow), (B, C, D) SEMs of touch papillae from the mandibular region Fig SEMs of the pst-cranial pits rom the ventral scdes of C. porosus: (A) ventro-lateral view, (B, C, D) ventral view Fig Light micrographs of cross-sections a pst-cranial pit (A) at l4ox magnification, showing diffuse pit region of the dermis (pr) and collagen-rich non-pit region (nr), and (B) at 640X magnification, showing stratum germinativum (sg) and stratum comeum (SC) layers of the epidermis (ep) Fig TEM (2400X) of the apex of pit region of the dermis, showing high concentration of cells (bl=basal lamina of the stratum germinativum of the epidermis, f=fibroblast cell, ir=iridocyte) viii
11 Fig TEM (7400X) of fibroblast cells frorn the pit region of the demis (c-collagen fibres) Fig TEM (8900X) of nerve tenninals (nt) supported by a fibroblast ce11 ( Fig TEM (135ûûX) of an iridocyte from the pit region of the demis Fig TEM (2700X) of epidermis from (A) the pit region, and (B) from another area of the same scale (sc=stratum comeum, sg=stratum germinativum, ddermis) Fig Summary illustration of the integumentary sense organ (sc=stratum comeum, sg=straturn germinativum, pr=pit region of the dermis, mon-pit region of the demis. d=derrnis) Fig Altemate interpretations of the hinction of the integumentary sense organ: (a) mec hanoreceptor, (b) chemoreceptor, (c) osrnoreceptor
12 LIST OF TABLES Table 1.1. Osmolality. Na'. CI.. and K+ concentrations of plasma and cloacal urine of C. porasirs and A. mississipiensis exposed to 20 ppt sea water... Il Table 1.2. Phylogenetic trends in osmoregulatory strategies of crocodiiians Table 2.1. Osmolaiity. Na' and K+. and blood hematocrit in cairnan plasma before and after dehydration by IO% of body mass. ' Indicates a significant increase (P4.05, paired t.test. corrected for mu1 tiple comparisons)... 26
13 LIST OF ABBREVIATIONS ANOVA... Anaiysis of variance OC...Deg reecelcius Ca2'... Calcium cation (divalent) COCI,... Cobaltchloride Cl'... Chloride anion (monovalent) cm... Centimetre ECM... Extracellular matrix F... Freshwater g... Gravitational force g... Gram hr ISO K' KCl Hour Integumentary sense organ Potassium cation (monovalent) Potassium chloride kg... Kilogram krn... Kilometre kv... min... mosm... MC... Kilovolt Minute Milliosmole Magnesium cation (divalent)
14 pi Microlitre pm... Micrometre pol... Micromole mm... Millimetre rnm... Millimolar n... Samplesize NaCl... Sodium chloride OsO,... Osmiumtetroxide ppt... rrnanova... Parts per thousand Repeated measures analysis of variance S... s.e.m. SEM TEM Seawater Standard error of the mean Scanning electron micrograph Transmission electron micrograph xii
15 CHAPTER ONE General Introduction: Osmoregulation in Marine and Estuarine Environments
16 . 1: Generai Introduction Osmoregulatory strategies of marine animais Animals which live in marine environments face the challenge of maintaining - physiologicd function in a highly saline extemal environment A wide variety of osmoregulatory mechanisms are employed, but these fa11 into three basic categories: (A) tolerating high concentrations of inorganic osmons in the body fluids, (B) eliminating the osmotic gradient by concentrating organic osmons in the body fluids, and (C) actively excreting inorganic osmons so as to maintain an interna1 osmolality below that of the surrounding sea water. The term "sea water" will be used throughout this thesis to indicate ionic composition but not concentration. Full sea water has a concentration of approximately 30 ppt. Thus "15 ppt sea water" means brackish water. The term "salt water" is avoided because it is ambiguous and could mean any sait in soiution (e.g. NaCi. KCI, CoCI,, etc.). The more precise term "NaCI solution" is also avoided, on the grounds that it is misleading. Although sea water is mostly an NaCl solution, it also contains traces of other physiologically important ions (e.g. Ca2', K*, ~g2', etc.). The distribution of different osmoregulatory strategies across taxa is summarised in figure 1.1. The first and simplest strategy (A) is simply to osmoconform, so that the body fluids have an ionic composition similar to that of sea water. This strategy is employed by the marine invertebrates, as well as by the presumably basal vertebrate group, the hagfishes (Myxinoidea) (Bentley 1971). With the exception of the hagfishes, however, al1 marine vertebrates employ some sort of mechanism to keep the concentration of NaCl
17 1 : General Introduction Fig Distribution of osmoregulatory strategies across taxa. The three strategies are as follows: (A) osmoconforming, so that the body fluids have an ionic composition similar to that of sea water, (B) osmoregulating by concentrating organic osmons in the body fluids, and (C) osmoregulating by excreting NaCl and maintaining a plasma osmolality below that of sea water.
18
19 1: Gened Introduction in the body fluids lower than that of the surrounding sea water. The elasmobrmchs have adopted strategy (B), and concentrate urea in their tissues. Their plasma osmolality is. thus comparable to that of sea water while their plasma NaCl concentration is kept much lower than that of their surroundings. They obtain water from sea water by drinking and then excreting the NaCl by rneans of a gland ("digitifoxm" or "rectal" gland) which concentrates NaCI fiom the bloodstream and empties it into the posterior intestine (Bentley 1971, Kirschner 1979). Teleost fishes maintain a low plasma osmolality relative to their surroundings (strategy C), and excrete excess NaCl by actively transporting Cl- out across the giil membranes. This system can work equally well as an osmoregulatory mechanism in fresh water, by actively transporting Cl' into the body, thereby allowing the retention of NaCl. The larnprey, Petromyzon, uses the same osmoregulatory mechanism (J. H. Youson; pers. cornrn.). The coelocanth, Lutimeriu osmoregulates by hyperuraemia of the interstitial fluids (Bentley 1970, Kirschner 1979). The Amphibia have not colonised the sea as other vertebrate groups have. The closest thing to a marine amphibian is the Crab-eating frog, Ranu cancrivora, which lives in southeast Asian estuariss. Interestingly, this species adopts two different osmoregulatory strategies, depending on its ontogenetic stage. Tadpoles maintain a low plasma osmolality relative to their 1977). presumably by actively transporting Cl- out across the gills, as teleost fishes do. Adults osmoregulate like elasmobranchs, using hyperuraernia of the interstitial fluids (Gordon et al. 1961).
20 - 1 : Gcncral Introduction Regardless of habitat type (marine, aquatic. terresuial), al1 amniotes maintain a plasma osmolality of about 300 mosm kg", and excrete excess NaCl under hyperosmotic. conditions. Marine and terrestrial habitats have similar osmotic stresses (the need to conserve water and excrete salt) and as a result, marine and desert amniotes often use the same mechanisms. Mamrnals are able to use the kidney for osmoregulation, since the mamrnalian nephron has lmps of Henle which allow the concentration of NaCl in the urine (Romer and Parsons 1986). Marine mammals are therefore able to excrete excess NaCl renaily. Non-rnammalian amniotes excrete excess salt extra-renally by means of specialised salt-secreting glands. These glands have ken independently derived in several lineages (Fig. 1.2). Among the Chelonia, the lachrymal gland has become modified for salt- secretion in marine-adapted lineages (Peaker and Linzell 1975). The tnily marine representatives of this order, the sea aides (Cheloniidae, Dermochelyidae) dnnk sea water, consume osmoconforming prey, and have lachrymal glands capable of high secretory rates (up to pmoles of Na' 100g" hi1) (Mazzotti and Dunson 1989) for the excretion of excess salt. In the marine lizard, Amblyrhynchus, the nasai gland has ken adapted for secreting salt (Peaker and Linzell 1975). In marine and estuarine snakes salt glands have evolved independently at lest three times, through the modification of either the premaxillary or the posterior sublingual glands in the mouth (Heatwole and Guinea 1993). Birds excrete NaCl through supra-orbital salt glands which drain into the nares (Peaker
21 - 1: General Introduction and Linzell, 1975). Marine adaptation versus estuarine adaptation The physiological stresses of estuarine environments are often discussed as though they were simply those of marine environments manifested to a lesser degree. This view, however, is rnisleading. Salinity in marine environments is high and constant. In an estuarine environment, by contrat, the salinity fluctuates, and may change in a few hours from highly saline to almost fresh. Estuarine habitats therefore have two important characteristics with respect to osmoregulatory physiology: (1) the stress of a constantly changing environmental osmolality, and (2) the opportunity to make use of intermittently available fresh or brackish water. Many of the physiologicai adaptations of estuarine animals are the same as those of marine animals, but there are other adaptations which are specific to estuarine habitats. Osmoconforrning invertebrates require special modifications to ailow them to survive fluctuating environmental salinities without becorning flooded with water. Protozoans actively remove water by means of water excreting vesicles (Brusca and Brusca 1990). Osmoconforming metazoan invertebratess use osmoregulatory organs such as nephridia, and a variety of mechanisms such as manipulating the concentrations of other inorganic osmons normally present in sea water in their body fluids (e.g. ca2+. M$) so as to modify osmotic gradients (Nicol 196 1).
22 1 : General Introduction Fig Independent denvations of salt-secreting glands in marine and estuarine lineages of non-mammalian amniotes.
23
24 1: Generai Introduction For animals which maintain a plasma osmolality lower than that of sea water by actively removing NaCl, adapting to estuarine life presents less of a chailenge than adapting to a fully marine existence, since there is simply less NaCl to actively remove. This is especially true of estuarine animals which take advantage of the fluctuating salinity by selectively drinking when the water is hypo-osmotic and avoiding drinking when it is hyper-osmotic. The selective drinking of only hypo-osmotic sea water is an important osmoregulatory mechanism for estuarine reptiles. Estuarine populations of normally freshwater species such as the turtle, Chelydra, and the water snake Nerodia, tack salt glands but are able to survive brief periods of exposure to hyperosmotic salinities by only drinking when the salinity is hypo-osmotic. Individuals from freshwater populations of these species lack this adaptation and will drink al1 salinities indiscrirninately (Dunson 1980, 1986). Other estuarine reptiles such as the turtle Mahclemys and the snake Cerberrcs have sait glands of low secretory capacity ( poles of Na' i00g-' hi1) and combine this adaptation with selectively avoiding drinking hyperosmotic sea water (Dunson 1985, Mazzotti and Dunson 1989). It is important to note that salinity discrimination and selective drinking of only hypo-osmotic salinities are specialisations for estuarine life and are completely useless as osmoregulatory mechanisms in marine environments when the salinity is hyperosmotic and unchanging. Fully marine osmoregulating animals must thetefore dnnk hyperosmotic sea water to obtain water, and have a good mechanism for excreting the salt. 8
25 1 : General Introduction Osmoregulatory strategies of estuarine crocodilians The living crocodilians are primarily freshwater animais, some populations of which survive in estuarine areas. Two species, Crocodylus acuzus and C. poroslcs are largely estuarine, and large individuais are often seen far out at sea (Taplin 1988). However, the frequently used cornmon name "Salt-water crocodile" for C. porosus is misleading. None of the living crocodilians is as specialised for marine life as the tntiy marine reptiles, the sea snakes and sea turtles (discussed above). Crocodilians do, however, possess some morphological and physiologicai specialisations for estuarine life. Penneabiliry of the integwnent Reducing integumental pemeability so as to minimise osmotic water loss is probably an important osmoregulatory mechanism for crocodilians living in hyperosmotic water. Unfortunately, however, Our understanding of the importance of integumental permeability as an osmoregulatory mechanism has suffered from a lack of comprehensive studies and from differences in methodology. Methodological differences include in vivo studies versus studies of isolated pieces of skin, and studies of restrained versus unrestrained animals. Distinguishing between dnnking and osmotic uptake of water through the buccal epithelium can be very difficult, and as a resuit, much confusion surrounds integumental permeability. The integumental pemeabilities of Crocodylus and of Alligator, for example, have been contrastingly descnbed as approximately equal
26 . 1 : General Introduction (Dunson and Mazzotti 1988) and as differing by an order of magnitude (Taplin 1988). Ellis and Evans (1984) measured integumentary Na' and water flux in Alligator mississipiensis in fresh water, and noted that the rate of Na' efflux was 0.02 v ol cmj hr- ' in the head region. compared with 0.01 po1 cm-2 hr-' for the ~ sof t the integument. The buccal epithelium of crocodylids and gavialids has a smooth, keratinised surface, while in alligatonds the buccal epitheliurn is characterised by the presence of mucous zones and dense papillae. This difference is thought to represent an adaptation by crocodylids to reduce osmotic water loss through the buccal epithelium (Taplin and Grigg 1989). It would be interesting to measure the difference in permeability of the buccal epithelium between crocodylids and alligatonds so as to detennine exactly how much of an osmotic advantage a keratinised buccal epithelium confers on a crocodilian in a hyperosmotic habitat. No data are currently available. The renal-cloaca1 sysrem The non-mammalian nephron lacks loops of Henle (Romer and Parsons 1986). and the crocodilian kidney is therefore an unlikely route for the excretion of excess salt. Schmidt-Nieslen and Skadhauge (1967) confinned this experimentally in C. acutus by comparing osmolality and electrolyte composition of ureteral urine with that of cloaca1 urine. As Na' load was increased, ureteral urine showed no significant change in electrolyte composition. In the cloaca, resorption of Na' and Cr was found to occur under hypo-osmotic conditions. However, increased Na' loading led to a decrease in 10
27 - 1 : General Introduction Table 1.1. Osmolality, Na', Cld, and K+ concentrations of plasma and cloaca1 urine of C. - porosus and A. mississippiensis exposed to 20 ppt sea water. Osmolality (mosm kg") Crocodylus porosud 1 Alligator mississippiensi~ Plasma Urine Plasma Urine from Grigg (1981) from Lauren (1985)
28 - 1: Generai Introduction cloacal resorption of these ions. Grigg (198 1) collected cloacai urine from C. porosus captured dong a saiinity gradient in order to investigate the possible role of the renal-cloacai system as an osmoregulatory mechanism. NaCl composition of cloaca1 urine remained constant as environmental salinity increased. However, urine osmolality increased, largely as a result of increasing K+ concentration. There was also an increase in the solid component (K4, ~ g ", and ca2' salis) of the urine. suggesting that cloacal water resorption was occumng. Lauren (1985) measured plasma and cloacal urine osmolality and ion concentrations in A. mississippiensis which had been kept in water of different salinities ranging from fresh water to 20 ppt (no data were recorded from higher salinities because mortalities started to occur at 20 ppt). Baxd on their own observations of C. porosus and C. n iloticus and compari son w ith Lauren' s ( 1985) observations of A. mississippiensis, Taplin (1988) and Taplin and Lovendge (1988) point out that the concentration of Na' in the cloacal urine of Alligator is markedly higher than that of Crocodylus (Table 1.1). This may indicate that Alligator but not Crocodylus uses the renalcloacal system for Na' excretion.
29 - 1: General Introduction Lingual salt-secreting glands With the exception of the mammds, al1 marine amniotes studied to date excrete excess Na+ extra-renally through specialised glands. Given this precedent, the presence of some sort of gland capable of secreting a hyperosmotic NaCl solution seemed a likety answer to the question of how crocodiles survive in hyperosrnotic sea water. The osmolalities of nasal and lachrymal secretions were duly measured in search of a possible route for Na' excretion (Peaker and Linzell 1975). The lingual salt-secreting glands were identified by Taplin and Grigg (1981) in C. porosus. These consist of highly vascularised glands, each with a duct leading to the surface of the tongue. Each gland is divided into lobular subunits, and blood flow from capillaries of the lingual vein is from the centre of the gland outward to the periphery, countercumnt to the flow of secretion as a mechanism for producing a concentrated NaCl solution (Taplin and Grigg 1981, Franklin and Grigg 1993). Intra-peritoneal injection of methacholine chloride stimulates secretion by the lingual glands of a NaCl solution 3-5 times more concentrated than the plasma (Taplin and Grigg 1981). The secretory rate is comparable to that of estuarine reptiles (e.g. Malaclemys) rather than marine reptiles (e.g. Chelonia) (Mazzotti and Dunson 1989). Lingual glands capable of secreting a hyperosmotic solution of NaCl (ma'] = 365 to 740 mm01 La') have been identified in dl crocodylids studied (12 species) including freshwater species, while alligatorids and gavialids appear to lack saltsecreting glands (Taplin et al. 1985). Surprisingly, the lingual salt glands do not secrete NaCl in response to Na+ loading 13
30 . 1 : Gencral Introduction (Taplin and Grigg 1981). Al1 rneasurements of secretory rate must therefore be made by stimulating secretion through intra-peritoneal injection of the parasympathetic nerve stimulant methacholine chloride. The failure of the lingual salt-secreting glands to respond to Na' loading has been interpreted as evidence that these glands are not involved in osmoregulation in estuarine environments (Mazzotti and Dunson 1989). However, the same phenornenon has recently been reported in ostriches (Gray and Brown 1995). Further study will be required to determine why some sait-secreting glands do not respond to Na4 loading. Other experiments have demonstrated indirectly the function of crocodile salt glands. Taplin and Loveridge (1988) observed that C. niloticus became hypernatraemic more rapidly when kept in sea water with lingual glands sealed with glue than the control goup in which the glands wen not sealed. Franklin and Grigg (1993) found that in C. porosus the volume of the vasculature supplying the lingual tands increased when animals were kept in hyperosmotic sea water. The lingual salt glands of crocodylids have generally been interpreted as an osmoregulatory adaptation for estuarine life. The presence of lingual glands in freshwater crocodylids has therefore ken considered vestigial and interpreted as evidence of a marine ancestry (Taplin et al Taplin and Grigg 1989). An alternative hypothes is is that the lingual glands of freshwater crocodylids are adaptations allowing the animal to deal with the osmotic stress of aestivation (Mazzotti and Dunson 1989). The osmoregulatory physiology of aestivating crocodiles has yet to be studied, however.
31 . 1 : General Introduction The role of selective drinking Selective drinking of only hypo-osmotic sea water has been shown to be an important osmoregulatory mechanism for many estuarine reptiles (Dunson 1980, 1985, 1986, Dunson and Mazzotti 1989). Taplin (1984) demonstrated that estuarine C. porosus wiii not drink hyperosmotic sea water even when severely dehydrated. Mazzotti and Dunson ( 1984) found that C actrtus dehydrated by 10% of body mass could distinguish very precisely between sea water of hyper- and hypo-osmotic saiinities, and would only drink salinities of 9 ppt or less (these king hypo-osmotic). Moreover, when the plasma osmoldity was increased by further dehydrating them (by 20% of body mass) the crocodiles drank sea water of salinities up to 15 ppt. With the exception of these two studies, the study of salinity discrimination and selective dnnking has been much neglected. Although it has been studied as a possible osrnoregulatory mechanism for two species of estuarine crocodylid, neither salinity discrimination nor selective drinking has been considered in a phylogenetic context. Their underlying mechanism has yet to be studied, and they have been virtually ignored as a possible confounding factor in skin permeability experiments comparing different crocodilian species (e.g. Lauren 1985, Mazzotti and Dunson 1984). Phylogenetic trends Perhaps the most surprising result of recent research into the osmoregulatory adaptations of crocodilians is the extent of the differences between alligatorids and 15
32 crocodylids. Although the farnilies Alligatoridae and Croçodylidae both have estuarine representatives, the crocodylids possess a suite of morphological and physiological modifications which make them bener adapted to estuarine life than alligatorids. These are summarised in Table 1.2. The best-studied of these. especidly in tem of phylogenetic trends, are the lingual salt-secreting glands in crocodylids, which are absent in alligatorids (Taplin et al. 1985). Another modification is the keratinised buccal epithelium of crocodylids, which is presumabiy less permeable to salt and water than that of alligatorids (Taplin and Grigg 1989). Less well-studied are the differences in the renal-cloacal system of alligatorids and crocodylids (Table 1.1). Taplin (1988) and Taplin and Loveridge (1988) interpret the difference in cloaca1 urine Na' concentration between A. mississippiensis and two species of Crocodylus kept in 20 ppt sea water as a phylogenetic difference between alligatorids and crocodylids. They propose that alligatorids (which lack lingual sait-secreting glands) are able to excrete Na' in their urine when exposed to hyperosmotic saiinities. The presence of estuarine adaptations (e.g. lingual sait-secreting glands) in freshwater crocodylids is of paiticular interest since these adaptations appear to have no function in freshwater habitats. These have been interpreted as evidence for a marine phase in the recent evolutionary history of crocodylids (Taplin et al. 1985, Taplin Taplin and Grigg 1989).
33 . 1 : Gened Introduction Table 1.2. Phy logenetic trends in osmoregulatory strategies of crocodilians. Croeod y lidae Mainiy freshwater, some estuarine populations Some species primarily estuarine Americas (except for one Asian species) Integumental permeability Sai t-secreting glands3 Yes Buccal epithelium' Mucous zones, dense papillae * Some Na' excreted Selective drinking SUBJECT OF THIS THESIS Keratinised to reduce osmotic water loss Not used for Na' excretion 'Taplin (1988), 'Taplin and Grigg (1989). qaplin et al. (1985). 'Lauren ( 1985). %rigg (198 1)
34 - 1 : General Introduction Objectives of the thesis The objective of this thesis was to study the role of salinity discrimination and selective drinking as an osmoregulatory mechanism used by estuarine crocodilians. S pecificail y, 1 attempted to determine (1) whether crocody lids and alligatorids differ in their capacity for saiinity discrimination, and (2) what mechanism crocodilians use to discriminate between salinities. Chapter 2 documents a series of experiments undertaken to test for salinity preference in a variety of crocodilian species from freshwater and estuarine habitats, in search of phylogenetic and ecological trends. Chapters 3 and 4 explore possible mechanisms underlying the phylogenetic difference revealed by the experirnents in chapter 2. Specificaliy, chapter 3 describes an expriment undertaken to determine the role of the buccal epithelium in salinity discrimination. Chapter 4 is a detailed morphological study of a possible salinity discrimination organ. Finally, chapter 5 is an attempt to integrate the phylogenetic, morphologicai, and physiological data, and to discuss areas of particular interest for future research.
35 C-R TWO Habitat and Phylogeny Influence Salinity Discrimination (Adapted from: Jackson. K., Butler, D. G., and D. R. Brooks. Habitat and phylogeny influence salinity discrimination in crocodilians: implications for osmoregulatory physiology and historical biogeography. Biological Journal of the Linnaean Society (In press))
36 2 Habitat and Phylogeny Influence Salinity Discrimination Abstract The ability to discriminate fresh water from hyperosmotic sea water, and to avoid drinking the latter, is known to be an important osmoregulatory mechanism for estuarine crocody 1 ids. The crocodilian families Crocody l idae and Alligatoridae both include species in w hich some populations inhabit brackish or coastal areas. Estuarine crocodylids, however, are more common than estuarine alligatorids, and members of the crocodylid farnily possess morphological specialisations (e-g. lingual salt-secreting glands, keratinised buccal epithelium) which confer an advantage to them over alligatorids in adapting to hyperosmotic conditions. This study was undertaken to determine whether the ability to discriminate between hyper- and hypo-osmotic salinities is determined by habitat, as it is in other normally freshwater reptiles, or whether, like morphological adaptations associated with estuarine life, it has a phylogenetic bais. Two species of freshwater alligatorid were found to drink fresh water and hyperosrnotic sea water indiscriminately, while an estuarine population of a normally freshwater ailigatorid species drank only fresh water. This indicated that salinity discrimination is determined by habitat. However, al1 three crocodylid species tested drank fresh water but not hyperosmotic sea water, suggesting that, in crocodilians, the ability to distinguish between fresh water and sea water is influenced by phylogeny as well as by habitat.
37 2: Habitat and Phyiogeny Influence Salinity Discrimination Introduction Reptiles inhabiting marine and estuarine environments face the challenge of maintaining a constant plasma osmolality while living in a hyperosmotic medium. They employ a variety of osmoregulatory strategies, including behavioural modifications such as avoiding drinking hyperosmotic sea water together with morphological adaptations associated with osrnoregulation such as salt-secreting glands and reduced integumental permeability. Dunson (1980, 1985, 1986) and Mazzotti and Dunson (1989) studied the physiological bais of putative marine adaptations in reptiles using snakes and turtles as models. Dunson and Mazzotti (1989) recognised a number of conditions which they interpreted as a gradient of evolutionary specialisations. In their model, the presumed plesiomorphic condition is represented by aquatic (freshwater) snakes and turtles (e.g. Nerodia, Chelydra). The second stage is represented by estuarine populations of the same species, which differ from freshwater populations in king able to tolerate limited exposure to hyperosmotic sea water by selectively drinking only hypo-osmotic water (Dunson 1980, 1986). Reptilian nephrons lack loops of Henle, and are therefore not capable of producing a hypertonic urine. Like marine birds, therefore, some reptiles possess extrarend salt-secreting glands capable of secreting a hypertonic NaCl solution. The third stage of marine adaptation is marked by the appearance of salt-secreting glands of low secretory capaci ty (volume and concentration) (e.g. Acrochordus, Malaclemys), which allow a constant plasma osmolality to be maintained when used in conjunction with selective drinking of only hypo-osmotic 1985). The fourth and final stage
38 2; Habitat and Phylogeny Influence Salinity Discrimination is represented by the tmly marine reptiles, the sea snakes (Hydrophiinae, Laticaudinae), sea turtles (Cheloniidae, Derrnochelyidae), and the marine iguana (Amblyrhychus). In these species the sait-secreting glands are well developed and ailow the maintenance of a constant plasma osmolality even when hyperosmotic sea water and osmo-conforming prey such as jellyfish are ingesteci. Sait glands have been independently derived several times in reptiles. Salt glands are sub-lingual in snakes, lachrymai in turtles, and nasal in iizards (Peaker and Linzell 1975). Crocodylids but not alligatorids have lingual salt glands of low secretory capacity (Taplin and Grigg 1981, Taplin et al. 1982, 1985). Crocodilians include species in which some populations inhabit brackish or es tuari ne habitats. Crocodylus acutrcs and C. porosus (Crocody 1 idae) are primari 1 y estuarine, while several other normally freshwater croçodylid species (e.g. C. cala ph ractus, C. johnstoni, C. more letii, C. niloticus, and C. palustris) have some estuarine populatio~s. Estuarine populations of Alligator mississippiensis, Caiman crocodilus and Ca. lutirostris (Alligatondae) are also known to exist (reviewed by Taplin, 1988). Estuarine crocodylids are more common than estuarine alligatorids, perhaps because they possess morphological specialisations, independent of habitat, which confer an advantage to them over alligatorids in adapting to hyperosmotic conditions. The lingual glands of crocodylids, for example, secrete an hyperosmotic solution of NaCl in response to stimulation with methacholine chloride, while those of alligatorids secrete an iso-osmotic solution (Taplin and Grigg 1981, Taplin et al. 1985). Additionally, crocodylids possess a keratinised buccal epithelium, so that osmotic water loss under
39 2: Habitat and Phylogcny Influenee Sdinity Discrimination hyperosmotoc conditions is less than for alligatorids, which lack protection for the highly permeable buccal epithefium (Taplin and Gngg, 1989). Opportunistic drinking of fresh water or hypo-osmotic sea water is thought to be an important mechanism allowing crocodilians inhabiting estuarine areas to maintain a constant plasma osmolality in a fluctuating hyperosmotic environment. The estuarine crocodyiids, C. acutus and C. porosus, as well as estuarine populations of C. johnstoni, will not drink hyperosmotic sea water even when severely dehydrated (Mazzotti and Dunson 1984, Taplin 1984, 1988, Taplin et al. 1993). Moreover, these species can distinguish very precisely between brackish water of hyper- and hypo-osmotic salinities (Mazzotti and Dunson 1984, Taplin 1984). No experimental data exist on the ability of alligatorids to selectively avoid drinking hyperosmotic sea water. Lauren (1985) found that juvenile A. mississipiensis died after three weeks of continuous exposure to salinities of 15 ppt or greater, and Mazzotti and Dunson (1984) observed that the mortality rate for A. mississippiensis was higher than chat of C. acutus when both species were exposed to the same regime of altemating hyper- and hypo-osmotic salinities. Bentley and Schmidt-Nielsen (1965) observed in the course of an experiment on skin permeability in the freshwater alligatorid Ca. crocodilur that 20% of their experimental animals died hours after king placed in a 33 ppt NaCl solution. apparently as a result of drinking the medium. The following study was undertaken to obtain experimental data on drinking of h y perosmotic sea water by a freshwater alligatorid, Cu. crocodilus, for cornparison w ith
40 2; Habitat and Phylogeny Influence Salinity Discrimination existing data from estuarine crocodylids, and to obtain data from representative alligatorid and crocodylid species from freshwater and estuarine habitats, to determine whether the - reported difference in capacity for salinity discrimination represented (1) a difference between freshwater and estuanne populations, similar to those observed in other normally freshwater reptiles in which some populations are estuarine, or (2) a difference between crocodylids and alligatorids anaiogous to morphological differences associated with marine adaptation between these two families. Lack of salinity discrimination in Caiman crocodilrrs Materials and Methods Captive-raised juvenile ( g) Ca. crocodilus were imported from Venezuela, housed in a tank of dechlorinated tap water with a land/water choice, at 30 C (air temperature), and fed live minnows ad libirum. Blood was sampled in order to determine which sea water dilutions were hyper- and hypo-osmotic to cairnan plasma. Blood samples of 0.2 ml were withdrawn from the caudal vein and centrifuged (4000 g) at 5OC for 10 minutes. Plasma was collected from caimans before and after dehydration (10% of initial body mass), and stored at -80 C until analysis. Na' and K+ concentrations were measured by flarne photometry (Instrument Laboratories, Model 9431, and osmolality by freezing point depression (Advanced Instruments micro-osmorneter, Model 3MO). Two experiments were conducted to determine whether dehydrated caimans would drink sea water. The first experiment measured the amount of water ingested by
41
42 - 2: Habitat and Phylogcny Influence Salinity Preference Table 2.1. Osmolality, Na' and K4, and blood hematocrit in caiman plasma before and after dehydration by 10% of body mas. ' Indicates a significant increase (P4.05, paired t-test, corrected for multiple comparisons). Before dehydration After dehydration % change n - Na' (mm) /- 1.4 Plasma 1 ~loocî I Osmolality (m0s-g) 289 +/- 1.6 hematocrit (%) / /- 1.6' / /- 2.4' 23.û+/- 1.3' +15% K' (mm) / no change +13% ' +42%,
43 2: Habitat and Phylogeny Influence Salinity Discrimination Fig Mass of water ingested by Caiman crocodil~~ (n=6) following dehydration. Values are mean +/- s.e.m. Differences in amount dmnk at different salinities are not significant (b0.05, ANOVA, Scheffe test).
44 10 15 salinity (ppt)
45 2: Habitat and Phylogeny Influence Salinity Discrimination Fig Body mass in grarns (mean +/- s.e.m.) of Ca. crocodilus (n=9), and mass of 20 ppt sea water ingested as a percentage of initial body mass (mean +/- s.e.m.): initiai mass (prior to dehydration), dehydrated mass (following dehydration), mass after 15 minutes exposure to sea water, and after 75 minutes exposure. ' Indicates significant increase in mass (P rmanova).
46 initial dehydrated 15 min 75 min
47 2; Habitat and Phylogeny Influence Sdinity Discrimination dehydrated caimans at different sea water dilutions. Sea water solutions in both experiments were made using Instant Ocean sea salt (Aquarium Systems, Mentor, Ohio 44060, USA). Six unfed caimans were seiected from a group of nine and weighed. They were then dehydrated by 10% of their initial body mas, in a current of air (30 C), re-weighed and placed in a 50-gallon plastic tank containing fresh water 10 cm in depth. After 15 minutes, they were re-weigned to determine, by difference, the amount of water ingested. This procedure was repeated for 5, 10, 15, 20, and 30 ppt sea water to determine whether or not increases in salinity would affect drinking by dehydrated caimans. Integumentary osmotic uptake of water in this species is known to be 1.l pl hfl (Bentley and Schmidt-Nielsen 1965). In a 200g animal immersed in fresh water for 15 minutes this represents a gain of only 95 pl (Sudace ma= 11.7~assO-", Dunson and Mazzotti 1988), so this was not an important factor in measuring drinking by increased mas. Between experiments, the test caimans were retumed to the fresh water holding tank for a period of at least seven days to allow time for rehydration. The second experiment was used to determine the amount of water ingested by dehydrated caimans during a longer period of exposure to hyperosmotic sea water. Body mass of nine unfed caimans was measured before and after dehydration, following a 15-minute exposure, and finally, a 75-minute exposure to 20 ppt sea water. Results and Discussion Following dehydration there was a significant increase in plasma Na+ concentration
48 2; Habitat and Phylogcny Influence Salinity Discrimination (15%) and osmolality (13%) but no significant change in plasma K* (Table 2.1). Blood hematocrit increased by 42%, indicating that the vascular cornpartment had become smailer in response to dehydration. Cairnan plasma was found to be hyprosmotic to 10 ppt sea water and hypo-osmotic to 15 ppt sea water, both before and after dehydration. Following dehydration, caimans were transferred to fresh water or to one of a series of diluted sea waters for an observation period of 15 minutes. At dl sea water concentrations tested, dehydrated caimans drank a significant volume of water, regaining of 2-5% of their initial body mass (Fig. 2.1). No overall statistical difference was found in the arnount drunk of the different sea water dilutions (ANOVA, PM.05). and a Scheffe test showed no significant difference in arnount drunk between any two dilutions (PM.05). When the 15-minute observation period was increased to 75 minutes (Fig. 2.2) caimans transferred to 20 ppt sea water continued to drink water and to increase in mass so that the final mass was 99.6% of the original. This indicated that the caimans continued to drink hyperosmotic sea water when given access to it for more than 15 minutes. These results (Figs. 2.1, 2.2) show that caimans drink water of ail salinities tested when dehydrated by 10% of body mass. That the caimans regained 24% of their initial body mass during 15 minutes of exposure to water over the range of salinities, is interpreted as an indication of drinking, as opposed to integumental uptake by dehydrated animals, because (1) the exposure period was short (15 minutes) in order to minimise the possible effects of osmotic uptake, and (2) the animals increased in mass during exposure to water of hyperosmotic salinities in which the osmotic gradient should have led to a
49 2: Habitat and Phylogeny Influence Salinity Discrimination decrease rather than an increase in mass if diffusion across the integument had been an important factor. Results from the first experiment (Fig. 2.1) indicated a slight. though statisticaily nonsignificant. decrease in the amount drunk at the two highest salinities (20 ppt and 30 ppt). It seemed possible that this result rnight indicate that although the caimans were drinking at al1 salinities, they stopped dnnking after an initial mouthful at strongly hyperosmotic salinities. However. when the experiment was repeated using 20 ppt sea water, rhis time weighing the caimans twice and extending the period of exposure to 75 minutes, the mean increase in mass after 15 minutes was 6.5% of initial body mass (Fig. 2.2). This is the largest mass increase recorded at any salinity, suppoxting the view that the amount drunk does not decrease at the highest salinities. When re-weighed after 75 minutes, the caimans were found to have further increased their mass (Fig. 2.2). This result is evidence against the idea that the caimans stopped drinking after an initiai mouthfül at hyperosmotic salinities. These data provide quantitative evidence of the inability of the freshwater alligatorid. Ca. crocodilus to osmoregulate by selectively drinking only water of hypo-osmotic salinities. These results are strikingly different from those obtained in studies of the estuarine crocodylids. C. ocutus (Mazzotti and Dunson 1984) and C. porosus (Taplin 1984). These species will not drink sea water of hyperosmotic salinities and are capable of distinguishing very precisely between hyper- and hypo-osmotic sea water. What Our experimental data for a freshwater alligatorid, and those of other researchers from
50
51 2: Habitat and Phylogeny Influence Salinity Discrimination Fig Body mass and water ingested (mean +/- s.e.m.) by dehydrated Alligator mississippiensis (n= 10) from a captive, freshwater population before and after 15 min exposure to (a) 30 ppt sea water and (b) fresh water. ' Indicates a significant mass increase from the dehydrated condition (P<0.001, t-test). The amount dmnk by the group exposed to 30 ppt sea water is not significantly different from the amount dnink by the group exposed to fresh water (t-test, P>0.05).
52 (6) ssou Kpoq (6) ssou Kpaa
53 2: Habitat and Phylogeny Influence Salinity Discrimination Fig Body mass and water ingested (mean +/- s.e.m.) by dehydrated A. rnississippiensis (n=3) from an estuarine population, More exposure to water, after 15 min exposure to 30 ppt sea water, and after 15 min exposure to fresh water. ' indicates a significant mass increase relative to the mass increase in sea water (paired t-test, PcO.025).
54 dehydrated seo water fresh water
55 2: Habitat and Phylogeny Influence Salinity Discrimination estuarine crocodylids fail to reveal, however, is whether this difference in capacity for behavioural osmoregulation represents (1) a phylogenetic constraint on the capacity of aliigatorids to adapt to estuarine conditions (e.g. inability of alligatorids to taste salt), (2) a behavioural modification with the potential to evolve independently in any crocodilian population exposed to fluctuating sa1 inities, or (3) a behaviour learned by individual crocodilians exposed to fluctuating saiinities (Le. without a genetic basis). Infïuence of Habitat Materials and Meth& This experiment assessed the effect of habitat in determining the capacity for salinity discrimination in crocodilians, by comparing Alligator mississippiensis from freshwater and estuarine populations. Experiments on freshwater A. mississippiensis were perforrned on captive-bred hatchlings (52-68g) (n=20) at the St. Augustine Alligator Farm in Florida. Estuarine A. mississippiensis juveniles ( g) (n=3) were collected from a freshwater pond on Sapelo Island, one of a string of barrier islands off the Coast of Georgia. Sapelo Island is approximately 20 km long by 6 km wide and is separated from the shore by a salt marsh 10 km wide, although it formed part of the mainland as recently as five to ten thousand years ago (Martof, 1963). Alligators from this population live in the freshwater ponds and salt marshes of the island. Ailigators ranging in size from large juveniles (>lkg) to adults have ken observed on the beach facing the Atlantic Ocean, and large individuals are often seen several kilometres from shore. Captured alligators were
56 2: Habitat and Phylogeny Influence Sdinity Discrimination housed temporarily in fibreglass tanks (approx. M)cm3), one alligator per tank, in an outbuilding where they were exposed to outdoor temperatures but protected frorn rain and - direct sunlight. In preparation for the expenment, alligators were dehydrated by 10% of initial body mass, to stimulate thirst, by keeping them out of the water for 24-4û hours. Once dehydrated, the animals were weighed and transferred to a tank containing either fresh water or 30 ppt sea water 10 cm in depth. After minutes they were rernoved frorn the water, blotted dry, and reweighed in order to determine, by difference, the amount of water ingested. For the freshwater alligators, in which the sample size was large, the animals were separated into two groups of ten. One group was exposed to sea water and the other to fresh water. For the estuarine alligators, the sample size was much smaller. Al1 the dehydrated animals were therefore exposed initially to 30 ppt sea water and then, if they did not drink, transferred to fresh water, since response (dnnking versus not drinking) to sea water would be more informative than response to fresh water. Moreover, animals which did not dnnk 30 ppt sea water in the course of 15 minutes exposure were still dehydrated by 10% of initial body mas, and could therefore be transferred to fresh water afterward and used to test response to fresh water. Sea water solutions were prepared as described above for Ca. crocodiius. Results and Discussion Freshwater A. mississippiensis increased in mass following exposure to fresh water
57 2: Habitat and Phylogeny Influence Salinity Discrimination Fig Body mass and water ingested (mean +/- s.e.m.) by dehydrated Crocodylus porosus (n=6), before exposure to water, after 15 min exposure to 30 ppt sea water, and after 15 min exposure to fresh water. ' Indicates a significantly greater mass increase relative to the mass increase in sea water (paired t-test, P<0.005).
58 dehydrated seo water fresh water
59 2: Habitat and Phylogeny Influence Salinity Discrimination Fig Body mass and water ingested (mean +/- s.e.m.) by dehydrated Crocodylus siamiensis (n= 10) from a captive, freshwater population (a) before and after 15 min exposure to 30 ppt sea water, and (b) before and after 15 min exposure to fresh water. ' Indicates a significantly greater mass increase in fresh water relative to the mass increase in 30 ppt sea water (t-test, P4.005).
60 (ssouj Apoa Z) palsa6ui ~a10m (6) ssow Aooc (6) ssou Apoa
61 2: Habitat and PhyIogeny Influence Sdinity Discrimination Fig Body mass and water ingested (mean +/- s.e.m.) by dehydrated Osteolaemus tetmspis (n=3), before exposure to water, after 15 min exposure to 30 ppt sea water, and after 15 min exposure to fresh water. ' Indicates a significantly greater rnass increase relative to the mas increase in sea water (paired t-test, Pe0.05).
62 body mass 1 b- water ingested dehydrated seo water fresh woter
63 2; Habitat and Phylogeny Influence Salinity Discrimination and to 30 ppt sea water (Fig. 2.3). There was no significant difference in amount of water drunk in fresh water versus sea water. By contrast, the estuarine A. mississippiemis drank fresh water but not sea water (Fig. 2.4). This result supports the hypothesis that the ability to selectively drink only water of hypo-osmotic salinities is determined at least in part by habitat. G. C. Grigg (pers. comm.) has observed that estuarine populations of Ca. latirostris also avoid drinking hyperosmotic sea water. Other estuarine populations of normally freshwater reptiles follow the same pattern. In the turtle, Chelydra, and the aquatic snake, Nerodia, individuds from freshwater populations drink al1 salinities, whereas those from estuarine populations selectively avoid drinking hyperosmotic sea water (Dunson 1980, 1986). Influence of Phylogeny Materials and Methodr In order to evaluate the possible role of phylogeny in determining the capacity for salinity discrimination in crocodilians, an attempt was made to obtain salinity discrimination data from a large number of crocodilian taxa so as to reveal phylogenetic patterns. Experiments were therefore undertaken to test for salinity preference in an addi tional t hree crocody lid species, C. porosus, C. siamiensis. and Osreolaernus tetraspis. Of these, two were exclusively freshwater crocodylid species: C. siamiensis (Asian) and 0. tetraspis (African). Data from 0steolaemu.s were especially interesting from a phylogenetic perspective, since this genus is the sister group of Crocodylus (Fig. 2.8).
64 2: Habitat and Phylogeny Influence Salinity Discrimination Although it is known that estuarine C. porosus and C. acutus hatchlings are able to distinguish very precisely between hyper- and hypo-osrnotic salinities (Taplin 1984, - Mazzotti and Dunson 1984), we included C. porosus in our study and used captive-bom, freshwater-raised hatchlings which had never previously been exposed to sea water, so as to rule out the possibility that avoidance of drinking hyperosmotic sea water is a behaviour tearned by individual crocodilians in response to exposure to sea water. Experirnents with C. siamiensis (n=20), and 0. tetraspis (n=3) were carried out at the S t. Augustine Alligator Farm, using captive-bom, freshwater-raised hatchlings (61-78g. 0. tetraspis; , C. siamiensis). Freshwater-raised C. porosus hatchlings ( g) (n=7) were obtained from the Long Kuan Hung Crocodile Farm in Singapore, and experiments were performed under laboratory conditions at the University of Toronto. The experimental procedure was the sarne as that described above for the estuarine and freshwater alligatoriàs. For C. simiensis, in which the sample size was relatively large (n=20), the animals were divided into two groups of ten. One group was exposed to fresh water and the other to 30 ppt sea water. The sample sizes of 0. tetraspis (n=3) and C. porosus (n=6) were smaller. Animals were therefore first exposed to 30 ppt sea water and then, if they did not drink, transferred to fresh water, as described above for estuarine A. mississippiensis. Results and Discussion Al1 three crocodylid species drank fksh water but not 30 ppt sea water. C.
65 2: Habitat and Phylogeny Influence Salinity Discrimination Fig The phylogenetic distribution of salinity preference among crocodilian species exarnined. (WS) indicates animals which drank both fresh water and sea water, and (FI-) indicates those which drank fresh water but not sea water.
66
67 2: Habitat and Phylogcny Influence Salinity Discrimination porosus (Fig. 2.5) and 0. tetrmpis (Fig. 2.7) increased in mass significantly more in fresh water than in sea water. The mass increase in the freshwater group of C. siamiensis was significantly greater than that for the sea water group (Fig. 2.6). The cornparison of fresh water and estuarine populations of A. mississipiensis indicated that for alligatonds at least, the capacity to discriminate between fresh water and hyper-osmotic sea water, and to avoid drinking the latter, is an adaptation found only in populations inhabiting areas where they are exposed to sea water. However, the results of the experiments on the freshwater crocodylids, C. siamiensis and 0. terraspis. and on freshwater-raised hatchlings of the estuarine species, C. porosus, suggest that phylogeny is also involved. Al1 crocodylids, whether freshwater or estuarine md with or without previous experience of hyperosmotic sea water, discriminate between fresh water and hyperosmotic sea water and will not drink the latter. Although previous studies have not tested freshwater crocodylids, it has previously been shown that estuarine C. acuzus (Mazzotti and Dunson, 1984) as well as estuarine populations of the normaily freshwater C. johnstoni (Taplin et al. 1993) selectively avoid drinlcing hyperosmotic sea water. General Discussion Al1 crocodylid species, whether estuarine or freshwater, drank only fresh water. Among the alligatorids, however, only those from an estuarine population distinguished between salinities. These results are presented in Fig superimposed on a phylogenetic tree depicting the evolutionary relationships of the taxa involved (Norell, fide
68 2; Habiîat and Phylogcny Innuencc Saihity Discrimination - -- Benton and Clark 1988). In crocodilians. therefore, the capacity for salinity discrimination has a strong phylogenetic component, analogous to morphological - adaptations associated with estuarine life. such as lingual salt-secreting glands and a keratinised buccal epithelium, which are present in crocodylids and absent from alligatorids. A search for the physiological mechanism underlying salinity discrimination in crocodylids may shed new light on the evolutionary significance of this adaptation. It would be useful, for example. to know whether crofodylids and estuarine alligators use the same mechanism to distinguish between fresh water and sea water. Di fferences in osmoregulatory phy siology between crocody lid and alligatorid crocodilians are of particular interest in the context of hwo conflicting hypotheses to explain the cumnt global distribution of crocodilians. The transsceanic migration hypothesis (Densmore 1983). explains the distribution of living cracodilian species as the result of a post-pliocene tram-oceanic migration on the part of a marine-adapted ancestral crocodylid. This hypothesis depends on an upper Cretaceoudearly Tertiary divergence between crocodylid and alligatorid lineages, which is more ment than indicated by the fossil record (Buffetaut 1982, Si , Steel 1968), but supporteci by molecular clock calculations based on haemoglobin sequence data (Densmore 1983). Lingual sait-secreting glands have been interpreted as crocodylid synapomorphies associated with adaptation to marine conditions on the part of a crocodylid ancestor, and consistent with the transoceanic migration hypothesis (Taplin et al 1985, Taplin and Grigg 1989). as has the keratinised (and consequently less permeable) buccal epithelium of crocodylids and
69 2: Habitat and Phylogcny Influence Salinity Discrimination Fig Altemate hypootheses for the evolutionary significance of salinity preference in crocodilians. In (a) lack of salinity discrimination is the plesiomorphic condition for crocodilians, in (b) it is the derived condition in freshwater alligatorids. (F/S) indicates drinking of both fresh water and sea water. (F/-) indicates dnnking of fresh water but not sea water.
70
71 2; Habitat and Phylogeny Influence Salinity Discrimination gavialids and the non-keratinised buccal epithelium of alligatorids. The presence of saitsecreting glands in freshwater crocodiles is thus considered vestigiai. An alternative hypothesis ex plains the distribution of living crocodilians as the result of speciation and upstream migration by a widely distributed, estuarine ancestral group. Systematic evidence from the cocvolving digenean parasites of crocodilians indicates a cosmopolitan distribution in the early Cretaceous, which supports the fossil data suggesting an ancient origin (Brooks 1978, Brooks and O'Grady 1989, Brooks and MacLennan 1993). However, the parasite data also show a mixture of freshwater (e.g. digeneans of the family Proterodiplostomidae) and estuarine-derived parasite groups (e-g. digeneans of the subfamily Acanthostominae), and could therefore be consistent with either a freshwater or an estuarine origin. The phyiogenetic significance of salinity discrimination in crocodilians can be interpreted in four different ways, depending on whether the capacity for salinity discrimination is assumed to be the plesiomorphic or the derived condition, and on whether the Crocodilia had a cosmopoliîan distribution in the Cretaceous (as suggested by the parasite data) or diverged more recently (as suggested by the molecular data). The first possibility (Fig. 2.9.a.) is that lack of salinity preference is plesiomorphic, and that the ability to distinguish between salinities has evolved independently in alligatorid populations exposed to sea water, and in an ancestral crocodylid. This interpretation is consistent with both the trans-oceanic migration hypothesis and the estuarine origin hypothesis depending on the time scaie involved. If the marine adaptation by the
72 2; Habitat and Phylogeny Influence Salinity Discrimination ancestral crocodylid is recent (after the separation of the continents), the tram-oceanic migration hypothesis is supported. However, if the adaptation at the base of the - crocodylid lineage occurred prior to the break-up of Pangaea, it could also be interpreted as consistent with the hypothesis that modem crocodilians arose from widely distributed estuarine ancestors and that upstrearn migration and Ereshwater adaptation occurred secondarily. An alternative interpretation (Fig. 2.9.b.), is that the ability to distinguish between salinities is plesiomorphic and has been secondarily lost in fieshwater alligatorids. This view is consistent with the estuarine origin hypothesis, although it still does not necessarily require a marine or estuarine ancestor. At this point, each of these trees requires exactly thme steps. Obtaining data from more taxa, especially from Gavialis and from basal alligatorids (e.g. Palaeosuchur) should help to determine which of these two trees more accurately reflects evolutionary history. Although these two alkrnatives each require an equal number of evolutionary events, it is not known whether or not the derivation of salinity preference from lack of salinity preference oçcurs more readily than the opposite, lack of salinity preference from salinity preference. It is also not known whether it is possible for a population to secondarily lose its capacity to avoid drinlcing hyperosmotic sea water. In contrast to the reverse situation, in which lack of salinity preference confers an obvious selective disadvantage on an individual inhabiting an estuarine are& there is no immediately obvious disadvantage associated with the latent capacity for sea water avoidance in an
73 2; Habitat and Phy logtny Influence Salinity Discrimination -- - individual living in fresh water. The same question applies to morphological speciaiisations associated with marine adaptation. The presence of lingual salt-secreting - glands in crocody lids, for exarnple. has been interpreted as a crocodylid synapomorphy consistent with the trans-oceanic migration hypothesis (Taplin and Grigg 1989). An alternative interpretation is that the presence of lingual salt-secreting glands is plesiomorphic, reflecting the estuarine ancestry of the group, and that their absence from alligatorids represents a secondary loss associated with upstrearn migration and speciation. Once again, it is not known whether Ioss of salt-secreting glands is a likely result of freshwater adaptation by estuarine reptiles. Part of the problern is that models of marine adaptation in reptiles are based on studies of snakes and turtles, and assume that the direction of the evolutionary trend is aiways from fresh water to sea water (e.g. Dunson and Mazzotti 1989). Such models are therefore not necessari1 y transferable to crocodilians, for which bth historical biogeogqhical sceniuios (trans-oceanic migration and estuarine origin hypotheses) involve estuarine to fresh water adaptation at some point. Further study of physiological adaptation to fresh water by marine or estuarine reptiles may provide answers to some of these questions and a better bais from which to evaiuate alternative reconstructions of the historical biogeography of the Crocodilia
74 CWU"ER THREE Evidence for Integumental Chemoreception in C. porosus
75 3: Evidence for Intcgumental Chemoreccption Abstract Crocodylids possess a suite of morphological and physiological specialisations - which make them better-adapted than alligatorids to the osmotic stresses of estuarine habitats. One of these modifications is a coating of keratin on the buccal epithelium to reduce osmotic water loss. An expected side-effect of a coating of this sort might be reduced chemosensory capacity on the buccal epithelium. However, another phylogenetic di fference associated w ith osmoregulation is that al1 crocody lids are able to distinguish very precisely between sea water of hyper- and hyposmotic saiinities, whereas in alligatorids this adaptation is only found in individuals from estuarine populations. This study investigated the role of the buccal epithelium in salinity discrimination in the Estuarine crocodile, Crocodylus porosus. When immersed in water, dehydrated crocodiles drink fresh water but not hyperosmotic sea water. However, crocodiles do drink hyperosmotic sea water when it is dripped directly into their mouths. Crocodiles drink hyperosmotic sea water dripped into their mouths while their bodies are immersed in fresh water, as well as fresh water dripped into their mouths while they are immersed in sea water. These results suggest that C. porosus does not use the buccal epithelium for salinity discrimination, and that one or more chemosensory organs must be located on another part of the body.
76 -- 3: Evidenct for Intcgumental Chemottception ~ -- In traduction Salinity discrimination and selective drinking are important osmoreplatory - mechanisms for esniarine crocodilians (chaptcr 1). This mechanism has a phylogenetic component, as al1 crocodylids, regardless of habitat (freshwater or estuarine) discriminate between fresh water and hyperosmotic sea water and will not drink the latter. Ailigatorids, however, are only capable of salinity discrimination if they belong to an estuarine population (chapter 2). Crocodylids possess a suite of estuarine adaptations which are absent in alligatorids (chapter 1). The ability to discriminate between salinities (regardless of habitat or experience) is one of these (chapter 2). Paradoxically, another of these adaptations is a modified buccal epithelium. In crocodylids the buccal epithelium is smooth and covered by a yellowish layer of keratin, presumably to reduce osmotic water loss under hyperosmotic conditions (Taplin and Grigg 1989). In alligatorids, by contrast, the buccal epithelium is characterised by the presence of mucous zones and dense papillae. Taplin and Grigg (1989) have speculated that the keratin coating on the buccal epithelium of crocodylids may interfere with sensory transduction. It seems surprising, therefore, that crocodylids are better at tasting salinity differences than alligatorids. Given these phylogenetic differences between crocodylids and alligatorids in salinity discrimination capacity and in buccal morphology, it seems possible that crocodylids and alligatorids may use different mechanisms to distinguish between sea water of hyper- and hypo-osmotic salinities. The following study was undertaken to
77 3: Evidcncc for Intcgumental Chcmomxption - investigate the mechanism used to distinguish between salinities by the Estuarine crocodile, Crocodylus porosus, especially the role of the keratinised buccal epi thelium in saiinity discrimination and selective drinking. Although the capacity for salinity discrimination is well-donimented in C. porosus (Chapter 2, Taplin 1984, Mazzotti and Dunson 1984). al1 studies of selective dnnking have used animals immersed in tanks of water, so that al1 parts of the crocodile are exposed to the sarne salinity. The purpose of the following study was to study selective drinking in animals in which the buccal epithelium and the post-cranial integument were exposed to different salinities, in order to determine what part of the crocodile tastes salt. Materials and Methods JuveniIe C. porosus (n4) (250g) were obtained from the Long Kuan Hung Crocodile Fam in Singapore. Crocodiles were housed in a tank of dechlonnated tap water with a ladwater choice, at 30 C (air temperature), and fed live rninnows ad libitum. In preparation for the expriment, crocodiles were dehydrated by 10% of their body mass, in order to stimulate thirst. This was achieved by preventing access to water for 48 to 72 hours. Crocodiles were weighed every 1-6 hours until 10% dehydration was reached. Two plastic tanks (approx. 20cm X 60cm) were filled to a depth of approx. 5 cm, one with fresh water (dechlonnated tap water), the other with 30 ppt sea water. Two wash bottles were also filled, one with frcsh water, the other with 30 ppt sea water. The
78 3: Evidence for Integumcntal Chemorcception. Fig Methodology for Expriment 1. F=fresh water, S=30 ppt sea water. Dehydrated crocodiles drink when immersed in fresh water (a), do not drink when immersed in 30 ppt sea water (b), drink 30 ppt sea water from a bottle (c), drink 30 ppt sea water from a bottle while irnmersed in fresh water (d), and drink fresh water from a bottle when immersed in 30 ppt sea water.
79 DRINKS DOES NOT DRINK DRINKS DRINKS
80 3: Evidcncc for Intcgumental Chcmorcception 30 ppt sea water solution was made using the rnethod described in chapter 2. Following dehydration, crocodiles were allowed access to drinking water (either fresh water or 30 ppt sea water) in the form of water dripped into their mouths with a wash bottle. To test whether the crocodiles would drink when the salinity of the water in contact with their buccal epithelium differed from that in contact with the rest of the integument, crocodiles were offered drinking water from the wash bottle while sitting in one of the tanks of water. Crocodiles were not restrained during these experiments. They showed little inclination to submerge their heads in the water however, perhaps because of the shallowness of the water, the small size of the tanks, and the presence of the researcher. Three permutations were tested: (1) 30 ppt sca water from a wash bottle while out of the water (n=6) (Fig. 3. lx), (2) 30 ppt sta watcr from a wash bottle while sitting in fresh water (n=3) (Fig. 3.l.d). and (3) fresh water from a wash bonle while sitting in 30 ppt sea water (n=3) (Fig. 3.1.e). Following the 30 min period of exposure, crocodiles were removed from the water and blotted dry. As in chapter 2, an increasc in body mass following exposure to water was considered evidence of drinking. The methodology of this experiment differed from chapter 2 in that the crocodiles had water dripped into their mouths from a wash bottle rather than king immersed in it. Since drinking from a wash bonle requires more time, the tirne Iimit was increased to 30 min. This introduced the problem of allowing for different rates of osmotic and evaporative water loss for crocodiles in fresh water, in sea water, and out of the water. Because of these problems, and the problems of comparing
81 3: Evidence for Integumcntal Chcmorcception - Fig A dehydrated C. porosus drinking hyperosmotic sea water from a wash bottle.
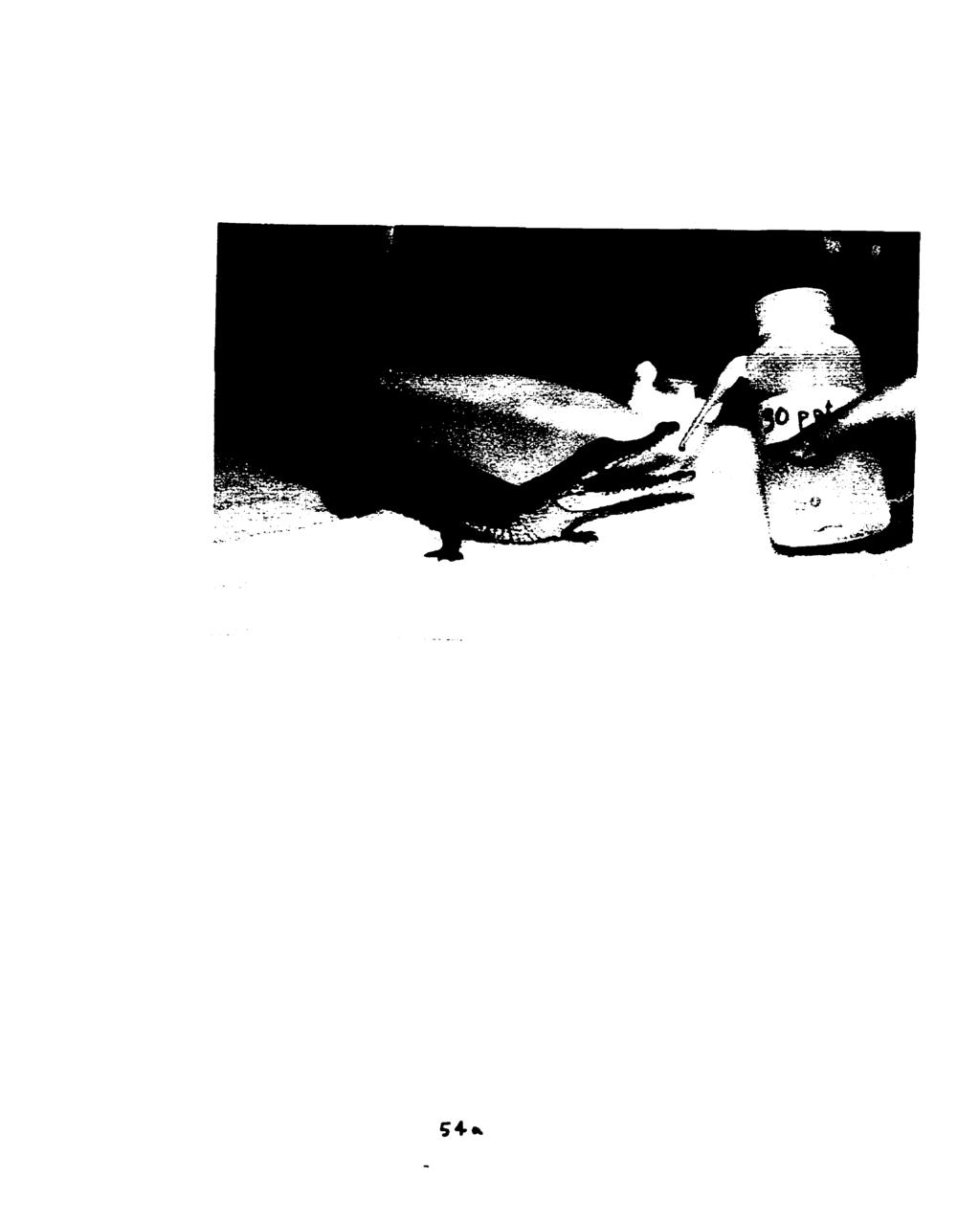
82
83 3: Evidcnct for Intcgurncntal Chemorcccption amounts drunk from this experiment with amounts from prcvious experiments (chapter 2) in which the animals were immened in the water, this expriment tested only wheiher the crocodiles drank or did not drink, rather than comparing amounts of water ingested. An increase in body mass of 2g in 30 min indicated that water had been ingested. This a conservative estimate. as Mavotti and Dunson (1984) used an increase of Ig in one hour as indicating drinking in C. acurus, even though these were largcr animals (20e70g) and immersed in water rather than having it dripped into their mouths. Resuits The results are surnmarised in figure 3.1. Revious experiments (chapter 2) have shown that dehydrated C. porosur drink when immersed in fresh water (Fig. 3.1.a) and do not drink when immersed in 30 ppt sea water (Fig. 3.1.b). However, in this study, dehydrated crocodiles (n=6) did drink 30 ppt sea water when it was dripped directly into their mouths (Fig. 3.1.c). Crocodiles (n=3) also drank 30 ppt sea water when it was dripped into their mouths while they were irnmersed in a tank of fresh water (Fig. 3.l.d). The dehydrated crocodiles (n=3) also drank fresh water when it was dripped into their mouths while they were immersed in a tank of sea water (Fig. 3.1.e). Discussion Previous experiments (chapter 2) showed that dehydrated crocodiles drink when
84 3: Evidence for Intcgumcntal Chcmoreccpùon immersed in fresh water and do not drink when immersed in hyperosmotic sea water. In this expenment, however, it was found that dehydrated crocodiles do drink hyperosmotic - sea water when it is dripped directly ont0 the buccal epithelium. The fact that crocodiles immersed in water drink fresh water but do not drink hyperosmotic sea water indicates that crocodiles have a mtchanism for distinguishing between salinities. This presumably allows hem to avoid drinking when it is osmotically disadvan tageous to do so. Although dehydrated crocodiles do not drink hyperosmotic sea water when irnmersed in it, they do drink it when it is dripped dirrcdy into their mouths (Fig. 3.2). This suggests that the salinity discrimination mechanism must be located somewhere other than on the buccal epithelium. The apparent lack of chemosensation in the mouth may be the rtsult of the keratin coating of the buccal epitheiium interfenng w i th sensory transduction. To test the hypothesis that the salinity discrimination organ of crocodiles is located on the integument of the body rather than in the mouth, two further experiments were undertaken in which crocodiles had water of one salinity dripped into their mouths while imrnersed in waîer of another salinity. Dehydrateci crocodiles drank hyperosmotic sea water dnpped directly into their mouths while immersed in fresh water (Fig. 3.l.d), supporting the hypothesis that crocodiles rely on scnsory information h m their skin rather than from their mouths to detennine salinity. In this case, the crocodiles were presumably drinking in response to the prcsencc of fresh watcr in contact with their skin. in spite of the fact that the water king drippcà into their mouths was strongly hyperosmotic.
85 3: Evidcnce for Intcgumcntal Chcmorcccption Dehydrated crocodiles do dnnk f ~sh water dripped into their mouths when they are imrnersed in sea water (Fig. 3.1.e). Following the same reasoning as above, if the - salinity of available drinking water is detennined by an organ on the skin rather than in the mouth, a crocodile ought not to drink if the water in contact with its skin is hyperosmotic, even if the water actually king dripped into its mouth is fmsh. One possible explanation for this.apparently anornalous result is that the crocodiles recognise that the contents of the wash bottle belongs to a different body of water than the one they are imrnersed in. The faft that the crocodiles drank hyperosmotic sea water ciripped into their mouths indicates that they are unable to discriminate salinities using only the buccal epithelium. It may be thercfore, that since the crocodiles are unable to taste the saiinity of water drïpped into their mouths from above, they assume that it is fresh water. Although under laboratory conditions this may ôe a dangerous assumption to make, in an ecologicai context it makes a certain amount of sense, as rainwater is never hyperosmotic. This idea could be ttsted by rcpeating the experiments outlined above, in which the salinity of the water in contact with the integument differs from the water in contact with the buccal epithelium. However, instead of having the two saiinities divided into two separate bodies of watcr (the wash bottle and the tank), the experiments should be carried out under conditions in which the intcgumcnt and the buccal epithelium can be ex posed to di fferent salinities, w ithou t the crocodiles perceiving them as separate bodies of water. One possible way of acheiving this might be in a flow tank in which waters of
86 3: Evidtnce for Inwgumcntal Ckrnoreception p~ different salinities could bc separated by cumnu. One important expcrimcnt is notably absent from this chapter What mechanism do estuarine alligatorids use to discriminate ktwecn salinities? My hypothesis is that crocodylids have adapted to estuarine habitats by devcloping a keratinised buccal epithelium, and then made up for the loss of chemosensory function in the mouth by developing integumentai chemorcctption. Aiiigatorids lack a keratinised buccal epithelium. and ought therefore to be able to use the buccal epitheliurn for sensoy transduction. Although freshwatcr alligatorids drink sea water of al1 salinities i ndiscriminatel y (Chapter 2). alligatorids from estuaxine populations arc able to discriminate between salinities, like crocodylids. If my hypothesis is correct, alligatorids and crocody 1 ids use dif fertnt mechanisms to discriminate between salinities: estuarine alligatorids taste sea water using the buccal epithelium, while crocodyüds use the integument. If this expriment were repeated using estuarine aiiigatorids, therefore, the alligatorids should not drinlc hyperosmotic sea water when it is dripped directiy into their mouths.
87 CHAPTER FOUR Morphology and Ultrastmcture of a Putative Integumentary Sense Organ
88 Abstract The structure of darkiy pigmented pits which arc prrsent, one per scale, on the. post-cranial scales of crocodylids was studied using light microxopy and scanning and transmission electron microscopy. The stratum comeum of the epidermis in the area of the pit is thinner than on the rcst of the scale. The outer surface of the seanirn comeum covering the pit is continuous with a dcepr layer of the strarim comeum covering the rest of the scaie, and is rcvealed through a circular opening in the outer surface of the stratum comeum proper. Widel y disperscd fibroc ytes, ne rve terminais and chromatophores are found throughout the pit region of the dermis, but these elements are concentrated in the area immediateiy beneath the stratum germinativum. The stratum germinativum of the epidennis is thicker in the area of the pit than in other regions of the integument, and bcneath the epidermai layer the pit region is fret of collagen fibres relative to the rest of the demis. The morphology of the pits suggests that they are sensory organs of some sort. Sensory organs on the amniote integument have traditionally been assumed to be mechanosensory. In this study, however, alternate interpretations of the stnicture (mechanosensory, chemosensory, osmosensory) are discussed.
89 Introduction Integumental pits an present, one per scale, on the post-cranial scales of some. crocodilians, and have been used extensivcly as taxonomic characters in the identification of crocodilian skins (King and Brazaitis 1971, Brazaitis 1987, Wennuth and Fuchs 1978). They are pksent on the post-cranial scales of crarodylids and gavialids (Fig. 4.1.a). but absent from the post-cranial scales of alligatorids (Fig. 4.1.b). The prcsence or absence of pits can therefore be useû to distinguish ailigatorids from the other two crocodilian families (even when the specimen involved is a smdl skin fragment such those used in a wallet or a belt). The pits have been rcfcrrcd to in the taxonomic Iiterature as "follicle pores" (King and Brazaitis 1971), "Porcn" (Wermuth and Fuchs 1978). "follicle glands". "follicle pits" (BraLaitis 1987). and "integumcntary sense organs" or "ISOs"(Brazaitis 1987). Although the pits have been well studied as taxonornic characters, their structure and function are not known. Brazaitis (1987) says that they are thought to be mechanosensory, while Grigg and Gans (1993) speculate that they may be either sensory structures or secretory porcs. An extensive search of the literahire reveals no detailed study of their structure or function. In contrast, detailed morphological and ultrastructural studies of mechanoreceptors or "touch papillae" on the cranial scales of the alligatorid, Cairnan crocodilus, have ken done (von During 1973, 1974, von During and Miller 1979). These "touch papillae" are confined to the cranial scales and are found in al1 crocodilians (Fig. 4.2). Alligatorids lack pits on the pst-cranid scales. The touch
90 Fig (A) Ventral scales of a crocodylid, C. porosus. showing the pits (arrow). (B) Ventral scales of an alligatorid, Ca. crocodilus, which lacks pits.
91
92 Fig Touch papillae (sensu von hiring, 1973) from the cranial scales of C. porosur: (A) head of a crocodile with touch papillae indicated (arrow), (B. C, D) SEMs of touch papillae from the mandibular region.
93
94 - -- papillae on the cranial wales of Ca. crocodiius are obviousiy not the pst-cranial pits referred to in the taxonomie literature, therefore. Von During's work may nonetheless be the source of the idea that the pits have a mechanosensory function, however. Guibe (1970) reports that the abundance of pits decreases as the animal ages. The source of this information, however, is a study which compared juvenile Crocodylus with adul t Alligator (Hulanicka, 1913), and it therefore seems Iikely that this difference may have more to do with phylogeny than with ontogeny. Evidence from previous experiments (chapter 3) indicates that the Crocodylid, Crocodylus porosus discriminates between sea water salinities using an organ located sornewhere other than in the mouth. The objective of this study was to investigate the general morphology and the ultrastructure of the post-cranial pits of C. porosus, in order to determine whether these might be salinity discrimination organs. Materials and Methods Animals Captive-bred juvenile ( g) crocodiles (Crocodylus porosus) were obtained from the Long Kuan Hung Crocodile F m in Singapore, where they had been housed in fresh water in an outdoor enclosure. A total of five specimens were obtained, of which four were frozen specimens for gross morphological snidies. in addition, one live specimen was euthanised by cervical dislocation in order to fix tissues for electron microscopie examination.
95 4: Intcgumentary Sense Organs Scanning electron microscopy Integument was dissected from the ventral surface of the freshly killed crocodile. and transferred immediately to Bouin solution. FolIowing 24 h fixation, tissue was transferred to 70% ethanol. Cubes of tissue (lmm3), each with a pit at its centre, were cut out of the ventral integument. Specirnens were dehydrated in a graded series of ethanol, dried in a Sowall critical point drying system, mounted on metai stubs, sputtercoated with gold, and observed in a Hitachi H-2500 scanning electron microscope at an acceleration voltage of 15 kv. Light microscopy and transmission electron microscopy Integument was dissected from the ventral surface of the freshly killed crocodile and transferred immediately to a solution of ice-cold 2.5% giutaraldehyde in 0.1 M Millonig's phosphate buffer at ph 7.3. The tissue was cut into lrnm cubes, each with a pit at its centre, and fixed in the abve fixative for 3 hours. The tissue cubes were then rinsed in the buffer, stored in the buffer for 3 days, and postfixed for 2 hours in 1% OsO, in the same buffer. Tissues were dehydrated in ethanol and propylene oxide, and embedded in Spurr's resin. Tissue blocks were sectioned with glass knives, using a Sorvall MT2 ultra-microtome. Semithin (0.5 um thickness) sections were piaced on glass slides and stained with 1% toluidine blue in saturated sodium tetraborate. Thin (silver) sections were cut using a diamond knife, and mounted on copper grids. The specirnens were stained with saturated uranyl acetate and lead citrate, and examined using a Hitachi
96 4: Integumentary Sense Organs Fig SEMs of the posi-cranial pits from the ventral scales of C. porosus: (A) ventro- lateral view, (B, C, D) ventral view.
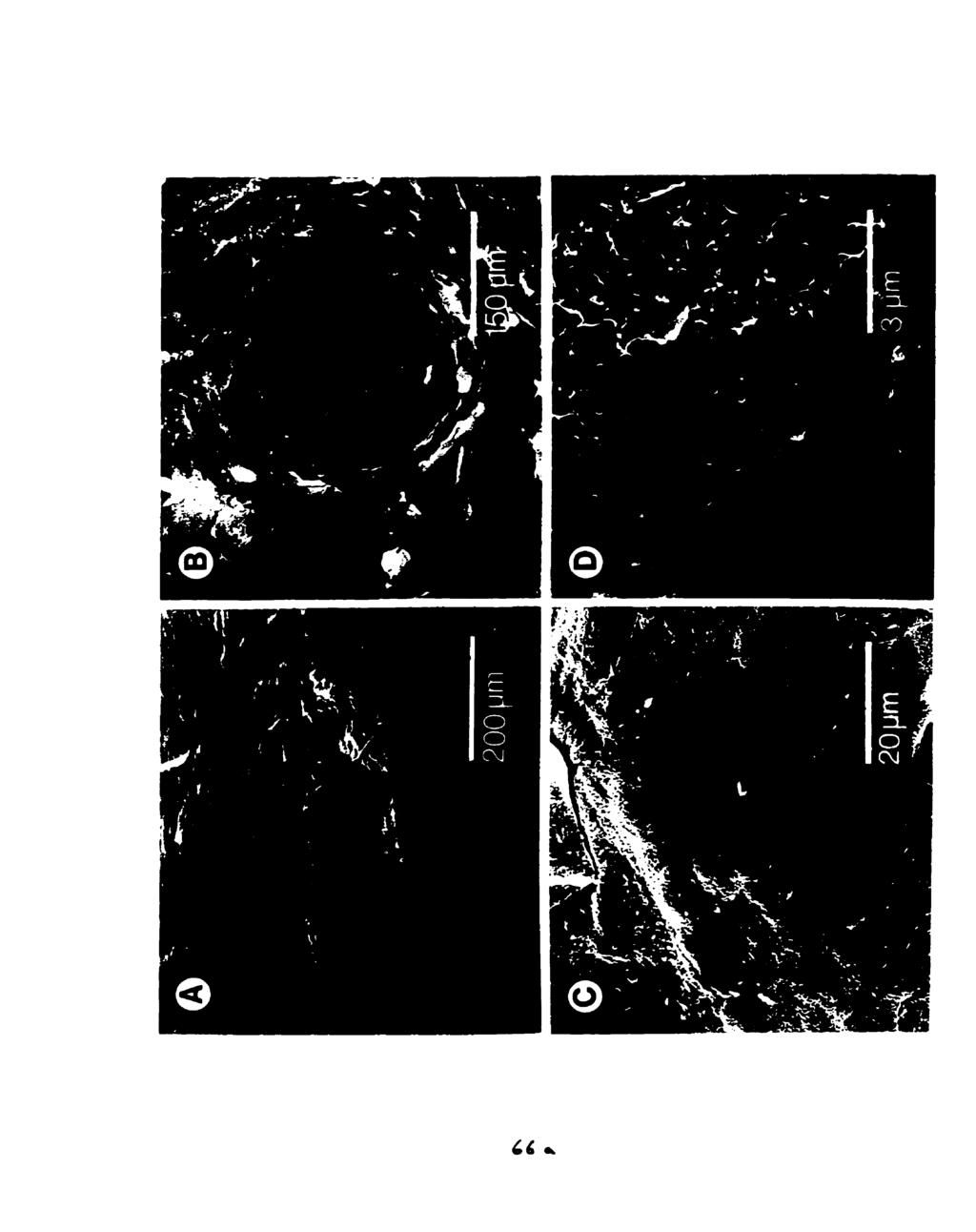
97
98 H-7000 transmission electron microscope. Ail fixation and staining procedures were those of Bozzola and Russell (1992). Resul ts Gross morphology Pits are present on al1 the post-cranial scales. The pits arc darkly pigmented and are therefore most clearly visible on the large and relatively unpigrnented ventral scales (Fig. 4.1.a). However, they are also present on the darkly pigmented dorsal scales and on the very small scales surrounding the proximal ends of the limbs. There is usually one pit on each scale, although the number occasionally varies from zero to the. The pit is usually centred (sagitally) in the caudal third of the scale. When more than one pit is present on a single scale, the two (or three) pits are positioned in line dong the same transverse plane. Scanning electron microscopy In SEM, the pit is revealed frorn the outer surface as a roughly circular opening, approx. 300 um in diameter, in the stiff outer layer of the stratum comeum that fonns a protective coating over the scales. A slightiy convex surface of the epidermis of the pit is revealed through the opening (Fig. 4.3.a,b). At the edges of the circular opening, the outer layer of keratin is seen to be flaking off in stiff sheets. The convex surface of the pit, when viewed at high magnification (Fig. 4.3.c), shows the margins between adjacent
99 Fig Light micrographs of cross-sections a pst-cranid pit (A) at 140X magnification, showing diffuse pit region of the demis (pr) and collagen-rich non-pit region (nr), and (B) at 640X magnification, showing (couectively) stratum germinativum, stratum spinosum, and straturn granulosum (sg) and stratum comeum (SC) layers of the epidermis (ep).
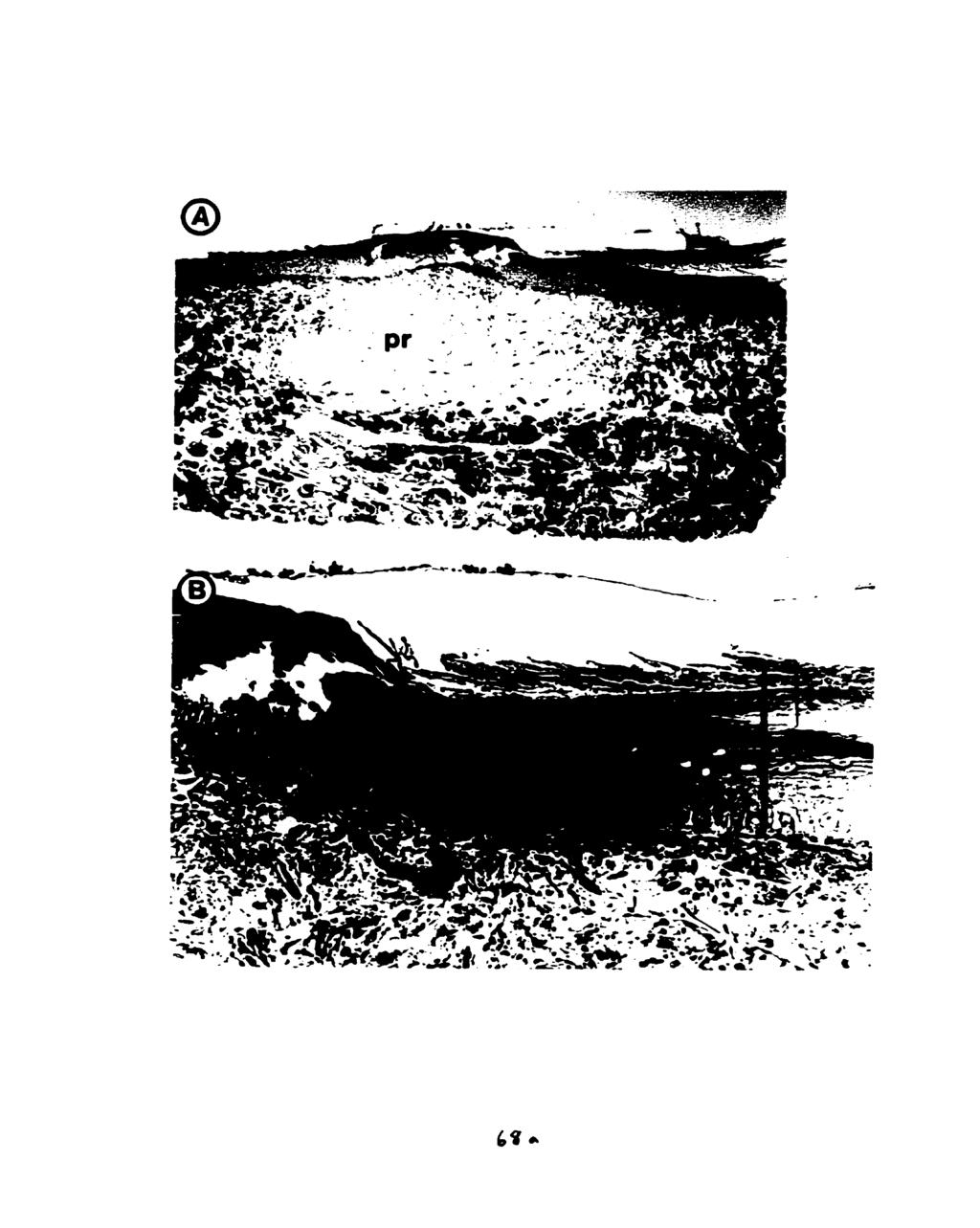
100
101 Fig TEM (2400X) of the apex of pit region of the demis, showing high concentration of cells (bl=basal lamina of the stranim germinativum of the epidexmis, f=fibroblast ce11, ir=iridocyte). Fig TEM (7400X) of fibroblast cells from the pit region of the dermis (c=collagen fibres). Fig TEM (8900X) of nerve terminais (nt) supported by a fibroblast ce11 (0. Fig TEM (135ûûX) of an indocyte from the pit region of the dermis.

102
103 4: Integumentary Sense Organs Fig TEM (27ûûX) of epidennis from (A) the pif region, and (B) from another area of the sarne scde (sc=stratum corneum, sg=collectively: stratum germinativurn, straturn s pinosum, stratum granulosum, d=dennis).
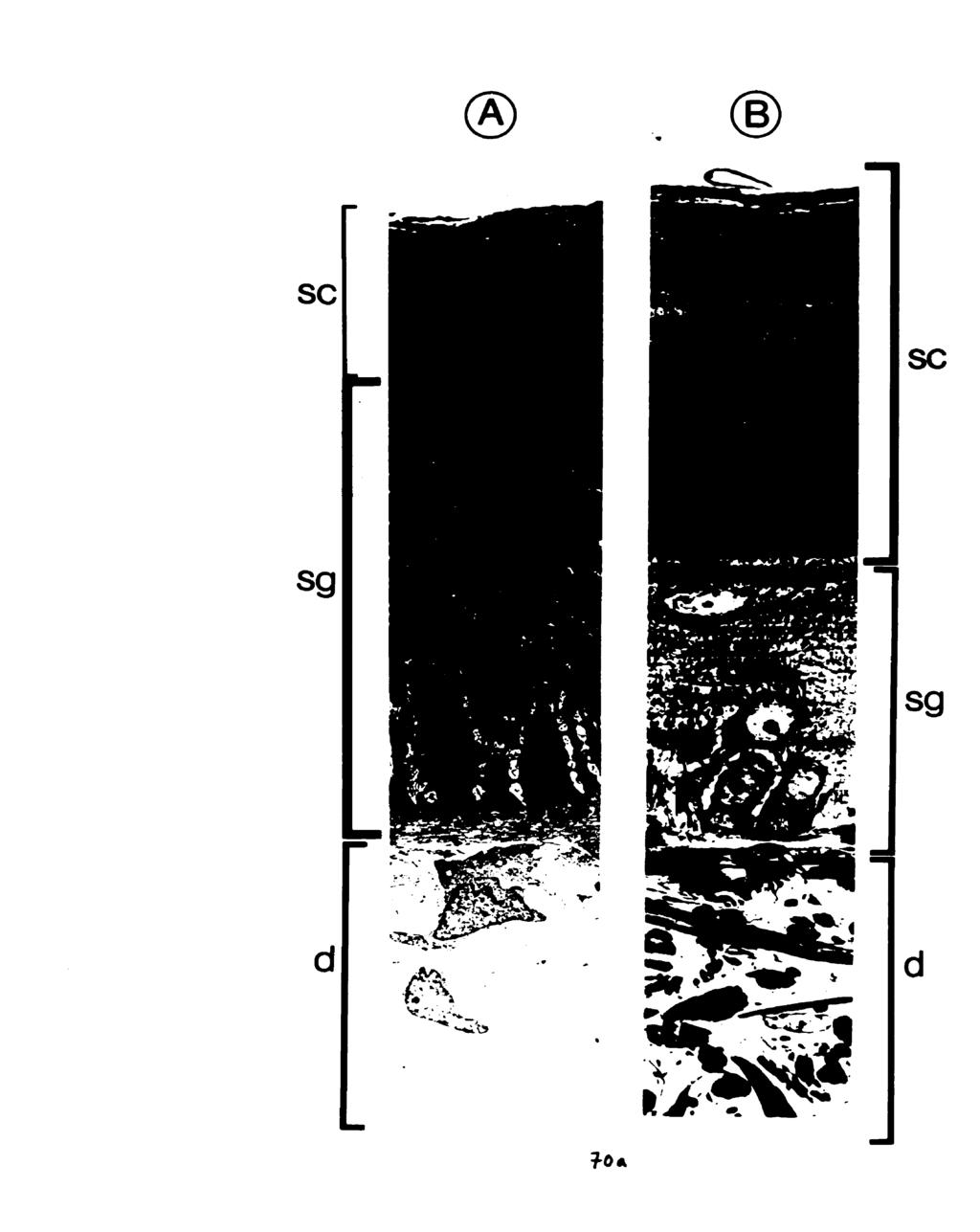
104
105 -- epidermal cells. The surface of these cells is pitted and possibly porous (Fig. 4.3.d). Light microscopy and transmission electron microscopy In cross-section and at low magnification (Fig. 4.4) the pit is revealed as a diffuse, lightly stained pket in the darkly stained. collagen-rich surrounding dermal tissue. The pit occupies an approximately spherical space in the dermis imrnediaîely underlying the circular opening in the stratum corneum proper of the epidennis. Celis in the dermal portion of the pit are widely separated, and there are very few collagen fibres compared with the surrounding dermis. Those cells that are present are usually densely concentrated at the apex of the demal sphere, in the area immediately underlying the stratum germinativum. The stratum corneum appears to have two distinct layers: an outer layer through which the underlying second layer protrudes through a circular opening. The outer layer tended to separate from the underlying layer and to break off the tissue block during sectioning. The cells of the pit beneath the epidermal layer are widely dispersed among a few collagen fibres and extensive ground substance of extracellular matrix. Three types of cells were observed in the dermal region of the pit, and al1 of these are most numerous at the apex of the dermal ngion of the pit. in the ana irnmediately underlying the stratum germinativum of the epidecmis (Figs ). Fibroblast cells (Fig. 4.6) are the most abundant ce11 type, but melanocytes are also cornmon. Al1 cells are often found close to nerve terminais (Fig. 4.7). but structural support for the nerve terminais may be provided by the attenuated processes of fibroblasts (Fig. 4.7). There are two types of
106 chromatophores: melanocytes and iridocytes (Fig. 4.8). The former contain many melanosornes, and the cytoplasm of the latter has many indophores or guanine crystals. - The epidemis in the pit region (Fig. 4.9.a) differs from the epidermis of non-pit regions (Fig. 4.9.b). Although the total thickness of the epidermis is equal in both regions, the pit region has a thimer stratum corneum than the non-pit region. Hence, the germinativa, spinosa, and granulosa strata are more prominent in the pit region. Discussion The location and general morphology of the poskrania.1 pits in C. porosus suggest that they are sensory structures of some sort. The pit is an opening in the stiff outer layer of the stratum corneum, through which a thinner, underlying layer of the epidermis is revealed. Immediately beneath this exposed region of the epidemiis is a fluid-filled area in the demis. Nerve terrninals are found in the fluid-filled region, immediately beneath the epidermis. The post-cranial pits of C. porosus differ from known cranial touch papillae of Ca. crocodilus (von During 1973, 1974) in several ways. The pits are present on only the post-cranial scales of crocodylids and gavialids, while the touch papillae are present on the cranial but not the post-cranial scales of dl crocodilians. There is usuaily only one pit on each post-cranial scale (with occasionai exceptions) and the pi& are not concentrated in any one particular region of the skin. The number of touch papillae on the cranial scales, by contrast, is much more variable (4-16). Papillae are most numerous on the scales
107 4: Integumentary Stnx Organs Fig Summary illustration of the integumentary sense organ (sc=stratum corneum, sg=collectively: stratum germinativum, stratum spinosum, stratum granulosum, p vit region of the dermis, nr=non-pit region of the demis, ddennis).

108
109 4: Integumentary Sense Organs Fig Altemate interpretations of the function of the integumentary sense organ: (a) rnechanoreceptor, (b) chemoreceptor, (c) osmoreceptor.
 chemoreceptor sea water (c) osmoreceptor A fresh water Hz0 ttt hyperosrnotic sea water The epidennis is Grmeable to water.")
110 (a) mechanoreceptor resting state phy sical pressure Physical pressure from outside changes the shape of the convex surface of the sensory organ. Nerve terminals are stimulated by the change in shape. (b) chemoreceptor sea water (c) osmoreceptor A fresh water Hz0 ttt hyperosrnotic sea water The epidennis is Grmeable to water. Osmotic changes in theextemal medium cause changes in the volume of the fluid-filled region, changing the shape of the convex surfrice. Nerve terrninals are stimulated by the change in shape >A *
Lingual Salt Glands in Crocodylus acutus and C. johnstoni and their absence from Alligator mississipiensis and Caiman crocodilus
 Lingual Salt Glands in Crocodylus acutus and C. johnstoni and their absence from Alligator mississipiensis and Caiman crocodilus Laurence E. Taplin 1, Gordon C. Grigg 1, Peter Harlow 1, Tamir M. Ellis
Lingual Salt Glands in Crocodylus acutus and C. johnstoni and their absence from Alligator mississipiensis and Caiman crocodilus Laurence E. Taplin 1, Gordon C. Grigg 1, Peter Harlow 1, Tamir M. Ellis
2/11/2015. Body mass and total Glomerular area. Body mass and medullary thickness. Insect Nephridial Structure. Salt Gland Structure
 Body mass and medullary thickness Thicker medulla in mammals from dry climate Negative allometry why? Body mass and total Glomerular area Glomerular area is a measure of total ultrafiltration rate Slope
Body mass and medullary thickness Thicker medulla in mammals from dry climate Negative allometry why? Body mass and total Glomerular area Glomerular area is a measure of total ultrafiltration rate Slope
Osmoregulation. 31 st Lecture Fri 03 April Chapter 26 & 27. Research Proposal Meetings 1
 31 st Lecture Fri 03 April 2009 Vertebrate Physiology ECOL 437 (MCB/VetSci 437) Univ. of Arizona, spring 2009 Kevin Bonine & Kevin Oh Osmoregulation Chapter 26 & 27 Research Proposal Meetings 1 Housekeeping,
31 st Lecture Fri 03 April 2009 Vertebrate Physiology ECOL 437 (MCB/VetSci 437) Univ. of Arizona, spring 2009 Kevin Bonine & Kevin Oh Osmoregulation Chapter 26 & 27 Research Proposal Meetings 1 Housekeeping,
Osmoregulation Chapter 26 & 27
 31 st Lecture Fri 03 April 2009 Vertebrate Physiology ECOL 437 (MCB/VetSci 437) Univ. of Arizona, spring 2009 Kevin Bonine & Kevin Oh Housekeeping, Wed 01 April 2009 Readings Today, Mon 30 Mar: Ch 26 (Ionic
31 st Lecture Fri 03 April 2009 Vertebrate Physiology ECOL 437 (MCB/VetSci 437) Univ. of Arizona, spring 2009 Kevin Bonine & Kevin Oh Housekeeping, Wed 01 April 2009 Readings Today, Mon 30 Mar: Ch 26 (Ionic
Summary. Introduction
 Grigg GC, LE Taplin, P Harlow and J Wright 1980 Survival and growth of hatchling Crocodylus porosus in salt water without access to fresh drinking water. Oecologia 47:264-6. Survival and Growth of Hatchling
Grigg GC, LE Taplin, P Harlow and J Wright 1980 Survival and growth of hatchling Crocodylus porosus in salt water without access to fresh drinking water. Oecologia 47:264-6. Survival and Growth of Hatchling
Morphology and Ultrastructure of Possible Integumentary Sense Organs in the Estuarine Crocodile (Crocodylus porosus)
") JOURNAL OF MORPHOLOGY 229:315-324 (1996) Morphology and Ultrastructure of Possible Integumentary Sense Organs in the Estuarine Crocodile (Crocodylus porosus) KATE JACKSON, DAVID G. BUTLER, AND JOHN H.
JOURNAL OF MORPHOLOGY 229:315-324 (1996) Morphology and Ultrastructure of Possible Integumentary Sense Organs in the Estuarine Crocodile (Crocodylus porosus) KATE JACKSON, DAVID G. BUTLER, AND JOHN H.
UREA AND OSMOREGULATION IN THE DIAMONDBACK TERRAPIN MALACLEMYS CENTRATA CENTRATA (LATREILLE)*
*") J. Exp. Biol. (1970), 5a, 691-697 691 Printed in Great Britain UREA AND OSMOREGULATION IN THE DIAMONDBACK TERRAPIN MALACLEMYS CENTRATA CENTRATA (LATREILLE)* BY M. GILLES-BAILLIENf Laboratory of Marine
J. Exp. Biol. (1970), 5a, 691-697 691 Printed in Great Britain UREA AND OSMOREGULATION IN THE DIAMONDBACK TERRAPIN MALACLEMYS CENTRATA CENTRATA (LATREILLE)* BY M. GILLES-BAILLIENf Laboratory of Marine
8/19/2013. Topic 12: Water & Temperature. Why are water and temperature important? Why are water and temperature important?
 Topic 2: Water & Temperature Why are water and temperature important? Why are water and temperature important for herps? What are adaptations for gaining water? What are adaptations for limiting loss of
Topic 2: Water & Temperature Why are water and temperature important? Why are water and temperature important for herps? What are adaptations for gaining water? What are adaptations for limiting loss of
Biology. Slide 1of 50. End Show. Copyright Pearson Prentice Hall
 Biology 1of 50 2of 50 Phylogeny of Chordates Nonvertebrate chordates Jawless fishes Sharks & their relatives Bony fishes Reptiles Amphibians Birds Mammals Invertebrate ancestor 3of 50 A vertebrate dry,
Biology 1of 50 2of 50 Phylogeny of Chordates Nonvertebrate chordates Jawless fishes Sharks & their relatives Bony fishes Reptiles Amphibians Birds Mammals Invertebrate ancestor 3of 50 A vertebrate dry,
Jeff Baier MS DVM Birds of Prey Foundation Broomfield, CO
 Jeff Baier MS DVM Birds of Prey Foundation Broomfield, CO drjeffbaier@gmail.com Squamates Chelonians Snakes Lizards Varanids Monitor Lizards Crocodilians Reptilian adaptations Anaerobic glycolysis Low
Jeff Baier MS DVM Birds of Prey Foundation Broomfield, CO drjeffbaier@gmail.com Squamates Chelonians Snakes Lizards Varanids Monitor Lizards Crocodilians Reptilian adaptations Anaerobic glycolysis Low
Question Set 1: Animal EVOLUTIONARY BIODIVERSITY
 Biology 162 LAB EXAM 2, AM Version Thursday 24 April 2003 page 1 Question Set 1: Animal EVOLUTIONARY BIODIVERSITY (a). We have mentioned several times in class that the concepts of Developed and Evolved
Biology 162 LAB EXAM 2, AM Version Thursday 24 April 2003 page 1 Question Set 1: Animal EVOLUTIONARY BIODIVERSITY (a). We have mentioned several times in class that the concepts of Developed and Evolved
Biology Slide 1 of 50
 Biology 1 of 50 2 of 50 What Is a Reptile? What are the characteristics of reptiles? 3 of 50 What Is a Reptile? What Is a Reptile? A reptile is a vertebrate that has dry, scaly skin, lungs, and terrestrial
Biology 1 of 50 2 of 50 What Is a Reptile? What are the characteristics of reptiles? 3 of 50 What Is a Reptile? What Is a Reptile? A reptile is a vertebrate that has dry, scaly skin, lungs, and terrestrial
REQUEST FOR STATEMENTS OF INTEREST SOUTH FLORIDA-CARIBBEAN CESU NETWORK NUMBER W912HZ-16-SOI-0007 PROJECT TO BE INITIATED IN FY 2016
 REQUEST FOR STATEMENTS OF INTEREST SOUTH FLORIDA-CARIBBEAN CESU NETWORK NUMBER W912HZ-16-SOI-0007 PROJECT TO BE INITIATED IN FY 2016 Project Title: Evaluating Alligator Status as a System-wide Ecological
REQUEST FOR STATEMENTS OF INTEREST SOUTH FLORIDA-CARIBBEAN CESU NETWORK NUMBER W912HZ-16-SOI-0007 PROJECT TO BE INITIATED IN FY 2016 Project Title: Evaluating Alligator Status as a System-wide Ecological
Alligator & Reptile Culture
 Alligator & Reptile Culture Chapter 8 Management Practices for Alligators, Frogs, and Plants Origin of the Alligator name el largato the lizard lagato alligator Photo 2001 by Kent Vliet Alligator mississippiensis
Alligator & Reptile Culture Chapter 8 Management Practices for Alligators, Frogs, and Plants Origin of the Alligator name el largato the lizard lagato alligator Photo 2001 by Kent Vliet Alligator mississippiensis
Class Reptilia Testudines Squamata Crocodilia Sphenodontia
 Class Reptilia Testudines (around 300 species Tortoises and Turtles) Squamata (around 7,900 species Snakes, Lizards and amphisbaenids) Crocodilia (around 23 species Alligators, Crocodiles, Caimans and
Class Reptilia Testudines (around 300 species Tortoises and Turtles) Squamata (around 7,900 species Snakes, Lizards and amphisbaenids) Crocodilia (around 23 species Alligators, Crocodiles, Caimans and
From Slime to Scales: Evolution of Reptiles. Review: Disadvantages of Being an Amphibian
 From Slime to Scales: Evolution of Reptiles Review: Disadvantages of Being an Amphibian Gelatinous eggs of amphibians cannot survive out of water, so amphibians are limited in terms of the environments
From Slime to Scales: Evolution of Reptiles Review: Disadvantages of Being an Amphibian Gelatinous eggs of amphibians cannot survive out of water, so amphibians are limited in terms of the environments
Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve,
 at Sungei Buloh Wetland Reserve,") Author Title Institute Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve, Singapore Thesis (Ph.D.) National
Author Title Institute Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve, Singapore Thesis (Ph.D.) National
Interpreting Evolutionary Trees Honors Integrated Science 4 Name Per.
 Interpreting Evolutionary Trees Honors Integrated Science 4 Name Per. Introduction Imagine a single diagram representing the evolutionary relationships between everything that has ever lived. If life evolved
Interpreting Evolutionary Trees Honors Integrated Science 4 Name Per. Introduction Imagine a single diagram representing the evolutionary relationships between everything that has ever lived. If life evolved
REPTILES. Scientific Classification of Reptiles To creep. Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Reptilia
 Scientific Classification of Reptiles To creep Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Reptilia REPTILES tetrapods - 4 legs adapted for land, hip/girdle Amniotes - animals whose
Scientific Classification of Reptiles To creep Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Reptilia REPTILES tetrapods - 4 legs adapted for land, hip/girdle Amniotes - animals whose
08 alberts part2 7/23/03 9:10 AM Page 95 PART TWO. Behavior and Ecology
 08 alberts part2 7/23/03 9:10 AM Page 95 PART TWO Behavior and Ecology 08 alberts part2 7/23/03 9:10 AM Page 96 08 alberts part2 7/23/03 9:10 AM Page 97 Introduction Emília P. Martins Iguanas have long
08 alberts part2 7/23/03 9:10 AM Page 95 PART TWO Behavior and Ecology 08 alberts part2 7/23/03 9:10 AM Page 96 08 alberts part2 7/23/03 9:10 AM Page 97 Introduction Emília P. Martins Iguanas have long
SALT GLAND FUNCTION IN THE GREEN SEA TURTLE CHELONIA MYDAS
 / exp. Biol. 144, 171-184 (1989) 171 Wrinted in Great Britain The Company of Biologists Limited 1989 SALT GLAND FUNCTION IN THE GREEN SEA TURTLE CHELONIA MYDAS BY SARAH W. NICOLSON AND PETER L. LUTZ Division
/ exp. Biol. 144, 171-184 (1989) 171 Wrinted in Great Britain The Company of Biologists Limited 1989 SALT GLAND FUNCTION IN THE GREEN SEA TURTLE CHELONIA MYDAS BY SARAH W. NICOLSON AND PETER L. LUTZ Division
Animal Form and Function. Amphibians. United by several distinguishing apomorphies within the Vertebrata
 Animal Form and Function Kight Amphibians Class Amphibia (amphibia = living a double life) United by several distinguishing apomorphies within the Vertebrata 1. Skin Thought Question: For whom are integumentary
Animal Form and Function Kight Amphibians Class Amphibia (amphibia = living a double life) United by several distinguishing apomorphies within the Vertebrata 1. Skin Thought Question: For whom are integumentary
Vertebrates. skull ribs vertebral column
 Vertebrates skull ribs vertebral column endoskeleton in cells working together tissues tissues working together organs working together organs systems Blood carries oxygen to the cells carries nutrients
Vertebrates skull ribs vertebral column endoskeleton in cells working together tissues tissues working together organs working together organs systems Blood carries oxygen to the cells carries nutrients
Phylogeny of Animalia (overview)
") The Diversity of Animals 2 Chapter 23 Phylogeny of Animalia (overview) Key features of Chordates Phylum Chordata (the Chordates) includes both invertebrates and vertebrates that share (at some point in
The Diversity of Animals 2 Chapter 23 Phylogeny of Animalia (overview) Key features of Chordates Phylum Chordata (the Chordates) includes both invertebrates and vertebrates that share (at some point in
Page # Diversity of Arthropoda Crustacea Morphology. Diversity of Arthropoda. Diversity of Arthropoda. Diversity of Arthropoda. Arthropods, from last
 Arthropods, from last time Crustacea are the dominant marine arthropods Crustacea are the dominant marine arthropods any terrestrial crustaceans? Should we call them shellfish? sowbugs 2 3 Crustacea Morphology
Arthropods, from last time Crustacea are the dominant marine arthropods Crustacea are the dominant marine arthropods any terrestrial crustaceans? Should we call them shellfish? sowbugs 2 3 Crustacea Morphology
Conservation (last three 3 lecture periods, mostly as a led discussion). We can't cover everything, but that should serve as a rough outline.
. We can't cover everything, but that should serve as a rough outline.") Comments on the rest of the semester: Subjects to be discussed: Temperature relationships. Echolocation. Conservation (last three 3 lecture periods, mostly as a led discussion). Possibly (in order of importance):
Comments on the rest of the semester: Subjects to be discussed: Temperature relationships. Echolocation. Conservation (last three 3 lecture periods, mostly as a led discussion). Possibly (in order of importance):
Reptilian Physiology
 Reptilian Physiology Physiology, part deux The study of chemical and physical processes in the organism Aspects of the physiology can be informative for understanding organisms in their environment Thermoregulation
Reptilian Physiology Physiology, part deux The study of chemical and physical processes in the organism Aspects of the physiology can be informative for understanding organisms in their environment Thermoregulation
Animal Diversity wrap-up Lecture 9 Winter 2014
 Animal Diversity wrap-up Lecture 9 Winter 2014 1 Animal phylogeny based on morphology & development Fig. 32.10 2 Animal phylogeny based on molecular data Fig. 32.11 New Clades 3 Lophotrochozoa Lophophore:
Animal Diversity wrap-up Lecture 9 Winter 2014 1 Animal phylogeny based on morphology & development Fig. 32.10 2 Animal phylogeny based on molecular data Fig. 32.11 New Clades 3 Lophotrochozoa Lophophore:
Reproductive physiology and eggs
 Reproductive physiology and eggs Class Business Reading for this lecture Required. Gill: Chapter 14 1. Reproductive physiology In lecture I will only have time to go over reproductive physiology briefly,
Reproductive physiology and eggs Class Business Reading for this lecture Required. Gill: Chapter 14 1. Reproductive physiology In lecture I will only have time to go over reproductive physiology briefly,
Return to the sea: Marine birds, reptiles and pinnipeds
 Figure 34.14 The origin of tetrapods Return to the sea: Marine birds, reptiles and pinnipeds Phylum Chordata Free swimmers Nekton Now we move to reptiles (Class Reptilia) and birds (Class Aves), then on
Figure 34.14 The origin of tetrapods Return to the sea: Marine birds, reptiles and pinnipeds Phylum Chordata Free swimmers Nekton Now we move to reptiles (Class Reptilia) and birds (Class Aves), then on
University of Canberra. This thesis is available in print format from the University of Canberra Library.
 University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
Using a Spatially Explicit Crocodile Population Model to Predict Potential Impacts of Sea Level Rise and Everglades Restoration Alternatives
 Using a Spatially Explicit Crocodile Population Model to Predict Potential Impacts of Sea Level Rise and Everglades Restoration Alternatives Tim Green, Daniel Slone, Michael Cherkiss, Frank Mazzotti, Eric
Using a Spatially Explicit Crocodile Population Model to Predict Potential Impacts of Sea Level Rise and Everglades Restoration Alternatives Tim Green, Daniel Slone, Michael Cherkiss, Frank Mazzotti, Eric
Lesson 7. References: Chapter 6: Chapter 12: Reading for Next Lesson: Chapter 6:
 Lesson 7 Lesson Outline: Embryonic Origins of the Dermis Specializations of the Dermis o Scales in Fish o Dermal Armour in Tetrapods Epidermal/Dermal Interactions o Feathers o Hair o Teeth Objectives:
Lesson 7 Lesson Outline: Embryonic Origins of the Dermis Specializations of the Dermis o Scales in Fish o Dermal Armour in Tetrapods Epidermal/Dermal Interactions o Feathers o Hair o Teeth Objectives:
JoJoKeKe s Herpetology Exam
 ~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~~*~*~*~*~*~*~*~*~*~*~*~*~*~*~ JoJoKeKe s Herpetology Exam (SSSS) 2:30 to be given at each station- B/C Station 1: 1.) What is the family & genus of the shown
~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~~*~*~*~*~*~*~*~*~*~*~*~*~*~*~ JoJoKeKe s Herpetology Exam (SSSS) 2:30 to be given at each station- B/C Station 1: 1.) What is the family & genus of the shown
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes)
") Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
SOUTH AFRICAN NATIONAL STANDARD
 ISBN 978-0-626-35881-5 SOUTH AFRICAN NATIONAL STANDARD Pet food Nutritional and manufacturing requirements WARNING This document references other documents normatively. Published by the South African Bureau
ISBN 978-0-626-35881-5 SOUTH AFRICAN NATIONAL STANDARD Pet food Nutritional and manufacturing requirements WARNING This document references other documents normatively. Published by the South African Bureau
Field Herpetology Final Guide
 Field Herpetology Final Guide Questions with more complexity will be worth more points Incorrect spelling is OK as long as the name is recognizable ( by the instructor s discretion ) Common names will
Field Herpetology Final Guide Questions with more complexity will be worth more points Incorrect spelling is OK as long as the name is recognizable ( by the instructor s discretion ) Common names will
Name Date When you put food away in the kitchen, you sort the food into groups. You put foods that are alike in certain ways into the same
 1 Name Date When you put food away in the kitchen, you sort the food into groups. You put foods that are alike in certain ways into the same group. Scientists do the same thing with animals, plants and
1 Name Date When you put food away in the kitchen, you sort the food into groups. You put foods that are alike in certain ways into the same group. Scientists do the same thing with animals, plants and
Vertebrates. Vertebrates are animals that have a backbone and an endoskeleton.
 Vertebrates Vertebrates are animals that have a backbone and an endoskeleton. The backbone replaces the notochord and contains bones called vertebrae. An endoskeleton is an internal skeleton that protects
Vertebrates Vertebrates are animals that have a backbone and an endoskeleton. The backbone replaces the notochord and contains bones called vertebrae. An endoskeleton is an internal skeleton that protects
CLADISTICS Student Packet SUMMARY Phylogeny Phylogenetic trees/cladograms
 CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
Vertebrates. Vertebrate Characteristics. 444 Chapter 14
 4 Vertebrates Key Concept All vertebrates have a backbone, which supports other specialized body structures and functions. What You Will Learn Vertebrates have an endoskeleton that provides support and
4 Vertebrates Key Concept All vertebrates have a backbone, which supports other specialized body structures and functions. What You Will Learn Vertebrates have an endoskeleton that provides support and
INDIVIDUAL IDENTIFICATION OF GREEN TURTLE (CHELONIA MYDAS) HATCHLINGS
 HATCHLINGS") INDIVIDUAL IDENTIFICATION OF GREEN TURTLE (CHELONIA MYDAS) HATCHLINGS Ellen Ariel, Loïse Corbrion, Laura Leleu and Jennifer Brand Report No. 15/55 Page i INDIVIDUAL IDENTIFICATION OF GREEN TURTLE (CHELONIA
INDIVIDUAL IDENTIFICATION OF GREEN TURTLE (CHELONIA MYDAS) HATCHLINGS Ellen Ariel, Loïse Corbrion, Laura Leleu and Jennifer Brand Report No. 15/55 Page i INDIVIDUAL IDENTIFICATION OF GREEN TURTLE (CHELONIA
Brine Shrimp Investigation AP Biology Name: Per:
 Brine Shrimp Investigation AP Biology Name: Per: Background Have you ever gone on a hike and come across an animal that blends in so well with its surroundings that you almost did not notice it? Camouflage
Brine Shrimp Investigation AP Biology Name: Per: Background Have you ever gone on a hike and come across an animal that blends in so well with its surroundings that you almost did not notice it? Camouflage
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and
 Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
Phylum Chordata. Fish, Amphibians, Reptiles
 Phylum Chordata Fish, Amphibians, Reptiles Chordates Three different groups Vertebrates Lancelets Tunicates At some point in their lives, they all have four special body parts Notocord Hollow nerve cord
Phylum Chordata Fish, Amphibians, Reptiles Chordates Three different groups Vertebrates Lancelets Tunicates At some point in their lives, they all have four special body parts Notocord Hollow nerve cord
13. Swim bladder function: A. What happens to the density of a fish if the volume of its swim bladder increases?
 Ch 11 Review - Use this worksheet as practice and as an addition to your Chapter 11 Study Guide. Test will only be over Ch 11.1-11.4. (Ch 11.5 Fossil and Paleontology section will not be on your test)
Ch 11 Review - Use this worksheet as practice and as an addition to your Chapter 11 Study Guide. Test will only be over Ch 11.1-11.4. (Ch 11.5 Fossil and Paleontology section will not be on your test)
muscles (enhancing biting strength). Possible states: none, one, or two.
. Possible states: none, one, or two.") Reconstructing Evolutionary Relationships S-1 Practice Exercise: Phylogeny of Terrestrial Vertebrates In this example we will construct a phylogenetic hypothesis of the relationships between seven taxa
Reconstructing Evolutionary Relationships S-1 Practice Exercise: Phylogeny of Terrestrial Vertebrates In this example we will construct a phylogenetic hypothesis of the relationships between seven taxa
Comparison of the osmoregulatory capabilities among three amphibious sea snakes (Laticauda spp.) in Taiwan
 in Taiwan") Cheng et al. Zoological Studies 2013, 52:28 RESEARCH Open Access Comparison of the osmoregulatory capabilities among three amphibious sea snakes (Laticauda spp.) in Taiwan Yuan-Cheng Cheng 1, Ming-Chung
Cheng et al. Zoological Studies 2013, 52:28 RESEARCH Open Access Comparison of the osmoregulatory capabilities among three amphibious sea snakes (Laticauda spp.) in Taiwan Yuan-Cheng Cheng 1, Ming-Chung
Physiological Ecology. Water and Salt Balance Respiratory Gas Exchange Respiration and Metabolism Thermoregulation Dormancy Energetics
 Physiological Ecology Water and Salt Balance Respiratory Gas Exchange Respiration and Metabolism Thermoregulation Dormancy Energetics Importance Amphibians and reptile physiology is directly tied to the
Physiological Ecology Water and Salt Balance Respiratory Gas Exchange Respiration and Metabolism Thermoregulation Dormancy Energetics Importance Amphibians and reptile physiology is directly tied to the
Reptile Regulations Training. Florida Fish and Wildlife Conservation Commission September 29, 2012 Division of Law Enforcement
 Reptile Regulations Training Florida Fish and Wildlife Conservation Commission September 29, 2012 Division of Law Enforcement Objective To provide an overview of the regulations relative to the commercialization
Reptile Regulations Training Florida Fish and Wildlife Conservation Commission September 29, 2012 Division of Law Enforcement Objective To provide an overview of the regulations relative to the commercialization
A brief report on the 2016/17 monitoring of marine turtles on the São Sebastião peninsula, Mozambique
 A brief report on the 2016/17 monitoring of marine turtles on the São Sebastião peninsula, Mozambique 23 June 2017 Executive summary The Sanctuary successfully concluded its 8 th year of marine turtle
A brief report on the 2016/17 monitoring of marine turtles on the São Sebastião peninsula, Mozambique 23 June 2017 Executive summary The Sanctuary successfully concluded its 8 th year of marine turtle
Characteristics of a Reptile. Vertebrate animals Lungs Scaly skin Amniotic egg
 Reptiles Characteristics of a Reptile Vertebrate animals Lungs Scaly skin Amniotic egg Characteristics of Reptiles Adaptations to life on land More efficient lungs and a better circulator system were develope
Reptiles Characteristics of a Reptile Vertebrate animals Lungs Scaly skin Amniotic egg Characteristics of Reptiles Adaptations to life on land More efficient lungs and a better circulator system were develope
Reptile Round Up. An Educator s Guide to the Program
 Reptile Round Up An Educator s Guide to the Program GRADES: K-3 PROGRAM DESCRIPTION: This guide provided by the Oklahoma Aquarium explores reptiles and their unique characteristics. The Reptile Round Up
Reptile Round Up An Educator s Guide to the Program GRADES: K-3 PROGRAM DESCRIPTION: This guide provided by the Oklahoma Aquarium explores reptiles and their unique characteristics. The Reptile Round Up
FACULTATIVE AESTIVATION IN A TROPICAL FRESHWATER TURTLE CHELODINA RUGOSA
 FACULTATIVE AESTIVATION IN A TROPICAL FRESHWATER TURTLE CHELODINA RUGOSA G. C. GRIGG, * K. JOHANSEN, P. HARLOW, * L. A. BEARD* and L. E. TAPLIN *Zoology A.08, The University of Sydney, NSW 2006, Australia.
FACULTATIVE AESTIVATION IN A TROPICAL FRESHWATER TURTLE CHELODINA RUGOSA G. C. GRIGG, * K. JOHANSEN, P. HARLOW, * L. A. BEARD* and L. E. TAPLIN *Zoology A.08, The University of Sydney, NSW 2006, Australia.
2019 Herpetology (B/C)
") 2019 Herpetology (B/C) Information shared by: Emily Burrell - Piedmont Herpetology Coach Maya Marin - NC State Herpetology Club Corina Mota - Piedmont Head Coach Adapted from KAREN LANCOUR - National Bio
2019 Herpetology (B/C) Information shared by: Emily Burrell - Piedmont Herpetology Coach Maya Marin - NC State Herpetology Club Corina Mota - Piedmont Head Coach Adapted from KAREN LANCOUR - National Bio
Investigating Fish Respiration
 CHAPTER 31 Fishes and Amphibians Section 31-1 SKILL ACTIVITY Interpreting graphs Investigating Fish Respiration It is well known that a fish dies from lack of oxygen when taken out of water. However, water
CHAPTER 31 Fishes and Amphibians Section 31-1 SKILL ACTIVITY Interpreting graphs Investigating Fish Respiration It is well known that a fish dies from lack of oxygen when taken out of water. However, water
Name Class Date. After you read this section, you should be able to answer these questions:
 CHAPTER 14 4 Vertebrates SECTION Introduction to Animals BEFORE YOU READ After you read this section, you should be able to answer these questions: How are vertebrates different from invertebrates? How
CHAPTER 14 4 Vertebrates SECTION Introduction to Animals BEFORE YOU READ After you read this section, you should be able to answer these questions: How are vertebrates different from invertebrates? How
Title of Project: Distribution of the Collared Lizard, Crotophytus collaris, in the Arkansas River Valley and Ouachita Mountains
 Title of Project: Distribution of the Collared Lizard, Crotophytus collaris, in the Arkansas River Valley and Ouachita Mountains Project Summary: This project will seek to monitor the status of Collared
Title of Project: Distribution of the Collared Lizard, Crotophytus collaris, in the Arkansas River Valley and Ouachita Mountains Project Summary: This project will seek to monitor the status of Collared
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1
 Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
Title: Phylogenetic Methods and Vertebrate Phylogeny
 Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
EXTERNAL FEATURES TEACHER RESOURCE BOOKLET
 EXTERNAL FEATURES TEACHER RESOURCE BOOKLET Koala, Phascolarctos cinereus. Image: QM. Saltwater crocodile, Crocodylus porosus. Image: QM. Poinciana Longicorn Beetle, Agrianome spinicollis. Image: Jeff Wright,
EXTERNAL FEATURES TEACHER RESOURCE BOOKLET Koala, Phascolarctos cinereus. Image: QM. Saltwater crocodile, Crocodylus porosus. Image: QM. Poinciana Longicorn Beetle, Agrianome spinicollis. Image: Jeff Wright,
DISTRIBUTION, ABUNDANCE AND HABITAT CONSERVATION OF CROCODYLUS POROSUS IN REMBAU-LINGGI ESTUARY, PENINSULAR MALAYSIA
 3 DISTRIBUTION, ABUNDANCE AND HABITAT CONSERVATION OF CROCODYLUS POROSUS IN REMBAU-LINGGI ESTUARY, PENINSULAR MALAYSIA Mohd Fazlin Nazli*, Nor Rasidah Hashim and Mohamed Zakaria M.Sc (GS265) 3 rd Semester
3 DISTRIBUTION, ABUNDANCE AND HABITAT CONSERVATION OF CROCODYLUS POROSUS IN REMBAU-LINGGI ESTUARY, PENINSULAR MALAYSIA Mohd Fazlin Nazli*, Nor Rasidah Hashim and Mohamed Zakaria M.Sc (GS265) 3 rd Semester
Sec KEY CONCEPT Reptiles, birds, and mammals are amniotes.
 Thu 4/27 Learning Target Class Activities *attached below (scroll down)* Website: my.hrw.com Username: bio678 Password:a4s5s Activities Students will describe the evolutionary significance of amniotic
Thu 4/27 Learning Target Class Activities *attached below (scroll down)* Website: my.hrw.com Username: bio678 Password:a4s5s Activities Students will describe the evolutionary significance of amniotic
! Three things needed to survive on land were: ! 1. Have lungs and breathe air. ! 2. Have a body resistant to drying out.
 Marine Reptiles, Birds and Mammals Vertebrates! Invaded the land and are descendants from the bony fish and were able to withstand the conditions on the land.! They evolved two sets of limbs (even snakes)
Marine Reptiles, Birds and Mammals Vertebrates! Invaded the land and are descendants from the bony fish and were able to withstand the conditions on the land.! They evolved two sets of limbs (even snakes)
Name Class Date. After you read this section, you should be able to answer these questions:
 CHAPTER 14 2 The Animal Kingdom SECTION Introduction to Animals BEFORE YOU READ After you read this section, you should be able to answer these questions: What is diversity? What are vertebrates? What
CHAPTER 14 2 The Animal Kingdom SECTION Introduction to Animals BEFORE YOU READ After you read this section, you should be able to answer these questions: What is diversity? What are vertebrates? What
Frisch s Outreach: Reptiles and Amphibians (Gr.1-3) Extensions
 Extensions") Frisch s Outreach: Reptiles and Amphibians (Gr.1-3) Extensions At a glance This program will allow students to explore Reptiles and Amphibians and their unique adaptations. Goal This class is designed
Frisch s Outreach: Reptiles and Amphibians (Gr.1-3) Extensions At a glance This program will allow students to explore Reptiles and Amphibians and their unique adaptations. Goal This class is designed
Frog Dissection Information Manuel
 Frog Dissection Information Manuel Anatomical Terms: Used to explain directions and orientation of a organism Directions or Positions: Anterior (cranial)- toward the head Posterior (caudal)- towards the
Frog Dissection Information Manuel Anatomical Terms: Used to explain directions and orientation of a organism Directions or Positions: Anterior (cranial)- toward the head Posterior (caudal)- towards the
Fishes, Amphibians, Reptiles
 Fishes, Amphibians, Reptiles Section 1: What is a Vertebrate? Characteristics of CHORDATES Most are Vertebrates (have a spinal cord) Some point in life cycle all chordates have: Notochord Nerve cord that
Fishes, Amphibians, Reptiles Section 1: What is a Vertebrate? Characteristics of CHORDATES Most are Vertebrates (have a spinal cord) Some point in life cycle all chordates have: Notochord Nerve cord that
Reptilian Requirements Created by the North Carolina Aquarium at Fort Fisher Education Section
 Essential Question: North Carolina Aquariums Education Section Reptilian Requirements Created by the North Carolina Aquarium at Fort Fisher Education Section What physical and behavioral adaptations do
Essential Question: North Carolina Aquariums Education Section Reptilian Requirements Created by the North Carolina Aquarium at Fort Fisher Education Section What physical and behavioral adaptations do
The study of nasal gland secretions in the lizard Uromastix loricatus (Agamidae: Reptilia) in Iran
 in Iran") AENSI Journals Journal of Applied Science and Agriculture Journal home page: www.aensiweb.com/jasa/index.html The study of nasal gland secretions in the lizard Uromastix loricatus (Agamidae: Reptilia)
AENSI Journals Journal of Applied Science and Agriculture Journal home page: www.aensiweb.com/jasa/index.html The study of nasal gland secretions in the lizard Uromastix loricatus (Agamidae: Reptilia)
Estimating radionuclide transfer to reptiles
 Estimating radionuclide transfer to reptiles Mike Wood University of Liverpool What are reptiles? Animals in the Class Reptilia c. 8000 species endangered (hence protected) Types of reptile Snakes Lizards
Estimating radionuclide transfer to reptiles Mike Wood University of Liverpool What are reptiles? Animals in the Class Reptilia c. 8000 species endangered (hence protected) Types of reptile Snakes Lizards
Ocean Teens. Water Quality Worksheet SECTION 1 SECTION 2. Tidal Touch Pools & Seahorse Sanctuary - Temperature. Jellyfish Kingdom - Light
 SECTION 1 Tidal Touch Pools & Seahorse Sanctuary - Temperature Feel the temperature of the water in the touch and tell tank. It is water from the ocean! Therefore it has the same temperature as the ocean.
SECTION 1 Tidal Touch Pools & Seahorse Sanctuary - Temperature Feel the temperature of the water in the touch and tell tank. It is water from the ocean! Therefore it has the same temperature as the ocean.
LABORATORY EXERCISE 6: CLADISTICS I
 Biology 4415/5415 Evolution LABORATORY EXERCISE 6: CLADISTICS I Take a group of organisms. Let s use five: a lungfish, a frog, a crocodile, a flamingo, and a human. How to reconstruct their relationships?
Biology 4415/5415 Evolution LABORATORY EXERCISE 6: CLADISTICS I Take a group of organisms. Let s use five: a lungfish, a frog, a crocodile, a flamingo, and a human. How to reconstruct their relationships?
Taxonomy. Chapter 20. Evolutionary Development Diagram. I. Evolution 2/24/11. Kingdom - Animalia Phylum - Chordata Class Reptilia.
 Taxonomy Chapter 20 Reptiles Kingdom - Animalia Phylum - Chordata Class Reptilia Order Testudines - turtles Order Crocodylia - crocodiles, alligators Order Sphenodontida - tuataras Order Squamata - snakes
Taxonomy Chapter 20 Reptiles Kingdom - Animalia Phylum - Chordata Class Reptilia Order Testudines - turtles Order Crocodylia - crocodiles, alligators Order Sphenodontida - tuataras Order Squamata - snakes
Animal Diversity III: Mollusca and Deuterostomes
 Animal Diversity III: Mollusca and Deuterostomes Objectives: Be able to identify specimens from the main groups of Mollusca and Echinodermata. Be able to distinguish between the bilateral symmetry on a
Animal Diversity III: Mollusca and Deuterostomes Objectives: Be able to identify specimens from the main groups of Mollusca and Echinodermata. Be able to distinguish between the bilateral symmetry on a
SOAR Research Proposal Summer How do sand boas capture prey they can t see?
 SOAR Research Proposal Summer 2016 How do sand boas capture prey they can t see? Faculty Mentor: Dr. Frances Irish, Assistant Professor of Biological Sciences Project start date and duration: May 31, 2016
SOAR Research Proposal Summer 2016 How do sand boas capture prey they can t see? Faculty Mentor: Dr. Frances Irish, Assistant Professor of Biological Sciences Project start date and duration: May 31, 2016
Today s Class. Go over viewfinder lab A closer look at the Animal Kingdom Taxonomy Worksheet
 Kingdom Animalia Today s Class Go over viewfinder lab A closer look at the Animal Kingdom Taxonomy Worksheet Viewfinder Kingdom: Animalia Cells: multicellular Food: eats other organisms Movement: moves
Kingdom Animalia Today s Class Go over viewfinder lab A closer look at the Animal Kingdom Taxonomy Worksheet Viewfinder Kingdom: Animalia Cells: multicellular Food: eats other organisms Movement: moves
Testing Phylogenetic Hypotheses with Molecular Data 1
 Testing Phylogenetic Hypotheses with Molecular Data 1 How does an evolutionary biologist quantify the timing and pathways for diversification (speciation)? If we observe diversification today, the processes
Testing Phylogenetic Hypotheses with Molecular Data 1 How does an evolutionary biologist quantify the timing and pathways for diversification (speciation)? If we observe diversification today, the processes
The Divergence of the Marine Iguana: Amblyrhyncus cristatus. from its earlier land ancestor (what is now the Land Iguana). While both the land and
. While both the land and") Chris Lang Course Paper Sophomore College October 9, 2008 Abstract--- The Divergence of the Marine Iguana: Amblyrhyncus cristatus In this course paper, I address the divergence of the Galapagos Marine
Chris Lang Course Paper Sophomore College October 9, 2008 Abstract--- The Divergence of the Marine Iguana: Amblyrhyncus cristatus In this course paper, I address the divergence of the Galapagos Marine
4 Many species of mammals, birds, reptiles, amphibians and fish 940L. Source 1 Habitats
 Source 1 Habitats 1 American Alligators can be found in fresh water environments like rivers, lakes, ponds, swamps and marshes. They also like to live in areas that are brackish, which means the water
Source 1 Habitats 1 American Alligators can be found in fresh water environments like rivers, lakes, ponds, swamps and marshes. They also like to live in areas that are brackish, which means the water
Writing: Lesson 31. Today the students will be learning how to write more advanced middle paragraphs using a variety of elaborative techniques.
 Top Score Writing Grade 4 Lesson 31 Writing: Lesson 31 Today the students will be learning how to write more advanced middle paragraphs using a variety of elaborative techniques. The following passages
Top Score Writing Grade 4 Lesson 31 Writing: Lesson 31 Today the students will be learning how to write more advanced middle paragraphs using a variety of elaborative techniques. The following passages
Liver and Gallbladder Morphology of the juvenile Nile crocodile, Crocodylus niloticus (Laurenti, 1768)
") Liver and Gallbladder Morphology of the juvenile Nile crocodile, Crocodylus niloticus (Laurenti, 1768) by ERNA VAN WILPE Submitted in partial fulfilment of the requirements for the degree MSc DEPARTMENT
Liver and Gallbladder Morphology of the juvenile Nile crocodile, Crocodylus niloticus (Laurenti, 1768) by ERNA VAN WILPE Submitted in partial fulfilment of the requirements for the degree MSc DEPARTMENT
Objectives: Outline: Idaho Amphibians and Reptiles. Characteristics of Amphibians. Types and Numbers of Amphibians
 Natural History of Idaho Amphibians and Reptiles Wildlife Ecology, University of Idaho Fall 2005 Charles R. Peterson Herpetology Laboratory Department of Biological Sciences, Idaho Museum of Natural History
Natural History of Idaho Amphibians and Reptiles Wildlife Ecology, University of Idaho Fall 2005 Charles R. Peterson Herpetology Laboratory Department of Biological Sciences, Idaho Museum of Natural History
1 Describe the anatomy and function of the turtle shell. 2 Describe respiration in turtles. How does the shell affect respiration?
 GVZ 2017 Practice Questions Set 1 Test 3 1 Describe the anatomy and function of the turtle shell. 2 Describe respiration in turtles. How does the shell affect respiration? 3 According to the most recent
GVZ 2017 Practice Questions Set 1 Test 3 1 Describe the anatomy and function of the turtle shell. 2 Describe respiration in turtles. How does the shell affect respiration? 3 According to the most recent
SALT WATER CROCODILE LIFE CYCLE FOR KIDS. Download Free PDF Full Version here!
 SALT WATER CROCODILE LIFE CYCLE FOR KIDS Download Free PDF Full Version here! SALTWATER CROCODILE FACTS FOR KIDS WITH PICTURES EHOW Saltwater crocodile facts for kids the saltwater crocodile is the largest
SALT WATER CROCODILE LIFE CYCLE FOR KIDS Download Free PDF Full Version here! SALTWATER CROCODILE FACTS FOR KIDS WITH PICTURES EHOW Saltwater crocodile facts for kids the saltwater crocodile is the largest
Diversity of Animals
 Classifying Animals Diversity of Animals Animals can be classified and grouped based on similarities in their characteristics. Animals make up one of the major biological groups of classification. All
Classifying Animals Diversity of Animals Animals can be classified and grouped based on similarities in their characteristics. Animals make up one of the major biological groups of classification. All
Course # Course Name Credits
 Curriculum Outline: Course # Course Name Credits Term 1 Courses VET 100 Introduction to Veterinary Technology 3 ENG 105 English Composition 3 MATH 120 Technical Mathematics 3 VET 130 Animal Biology/ Anatomy
Curriculum Outline: Course # Course Name Credits Term 1 Courses VET 100 Introduction to Veterinary Technology 3 ENG 105 English Composition 3 MATH 120 Technical Mathematics 3 VET 130 Animal Biology/ Anatomy
OBSERVATIONS ON THE QUALITATIVE AND QUANTITATIVE STRUCTURAL CHARACTERISTICS OF THE REPTILIAN KIDNEYS.
 OBSERVATIONS ON THE QUALITATIVE AND QUANTITATIVE STRUCTURAL CHARACTERISTICS OF THE REPTILIAN KIDNEYS. ~B~SI"Y OF Nmlll,.tpj,Tb 1.11.,,)' A Thesis submitted to the university of Nairobi in partial fulfillment
OBSERVATIONS ON THE QUALITATIVE AND QUANTITATIVE STRUCTURAL CHARACTERISTICS OF THE REPTILIAN KIDNEYS. ~B~SI"Y OF Nmlll,.tpj,Tb 1.11.,,)' A Thesis submitted to the university of Nairobi in partial fulfillment
Monitoring marine debris ingestion in loggerhead sea turtle, Caretta caretta, from East Spain (Western Mediterranean) since 1995 to 2016
 since 1995 to 2016") 6th Mediterranean Conference on Marine Turtles 16 19 October 2018, Poreč, Croatia Monitoring marine debris ingestion in loggerhead sea turtle, Caretta caretta, from East Spain (Western Mediterranean) since
6th Mediterranean Conference on Marine Turtles 16 19 October 2018, Poreč, Croatia Monitoring marine debris ingestion in loggerhead sea turtle, Caretta caretta, from East Spain (Western Mediterranean) since
5 pt. 10 pt. 15 pt. 20 pt. 25 pt
 Final Jeopardy Characteristics of Vertebrates Characteristics of Fish Amphibians Reptiles Chapter 16 Vocabulary 5 pt 5 pt 5 pt 5 pt 5 pt 10 pt 10 pt 10 pt 10 pt 10 pt 15 pt 15 pt 15 pt 15 pt 15 pt 20 pt
Final Jeopardy Characteristics of Vertebrates Characteristics of Fish Amphibians Reptiles Chapter 16 Vocabulary 5 pt 5 pt 5 pt 5 pt 5 pt 10 pt 10 pt 10 pt 10 pt 10 pt 15 pt 15 pt 15 pt 15 pt 15 pt 20 pt
Living Dinosaurs (3-5) Animal Demonstrations
 Animal Demonstrations") Living Dinosaurs (3-5) Animal Demonstrations At a glance Students visiting the zoo will be introduced to live animals and understand their connection to a common ancestor, dinosaurs. Time requirement One
Living Dinosaurs (3-5) Animal Demonstrations At a glance Students visiting the zoo will be introduced to live animals and understand their connection to a common ancestor, dinosaurs. Time requirement One
How Often Should You Replace Your Fish Tank Filter
 How Often Should You Replace Your Fish Tank Filter Do you clean your filter often enough? By Stephen G. Noble How Often do You Clean a Filter? All things In-tank power filters should be serviced every
How Often Should You Replace Your Fish Tank Filter Do you clean your filter often enough? By Stephen G. Noble How Often do You Clean a Filter? All things In-tank power filters should be serviced every
Unit 19.3: Amphibians
 Unit 19.3: Amphibians Lesson Objectives Describe structure and function in amphibians. Outline the reproduction and development of amphibians. Identify the three living amphibian orders. Describe how amphibians
Unit 19.3: Amphibians Lesson Objectives Describe structure and function in amphibians. Outline the reproduction and development of amphibians. Identify the three living amphibian orders. Describe how amphibians
30-3 Amphibians Slide 1 of 47
 1 of 47 What Is an Amphibian? What Is an Amphibian? An amphibian is a vertebrate that, with some exceptions: lives in water as a larva and on land as an adult breathes with lungs as an adult has moist
1 of 47 What Is an Amphibian? What Is an Amphibian? An amphibian is a vertebrate that, with some exceptions: lives in water as a larva and on land as an adult breathes with lungs as an adult has moist
 SUSTAINABLE TRADE: EXPLORING RELIABLE TRACEABILITY SYSTEMS FOR MANAGING TRADE OF PYTHON SKINS A. Participatory and Inclusive B. Transparent, Credible and Practical C. Acknowledge A review of the trade
SUSTAINABLE TRADE: EXPLORING RELIABLE TRACEABILITY SYSTEMS FOR MANAGING TRADE OF PYTHON SKINS A. Participatory and Inclusive B. Transparent, Credible and Practical C. Acknowledge A review of the trade
Catalogue. August 2014 PRODUCT GUIDE
 August 2014 Catalogue PRODUCT GUIDE KENT Marine is committed to providing effective ways to keep beautiful, healthy aquariums. For over 15 years, we have been offering solutions that help the hobbyist
August 2014 Catalogue PRODUCT GUIDE KENT Marine is committed to providing effective ways to keep beautiful, healthy aquariums. For over 15 years, we have been offering solutions that help the hobbyist
History of Lineages. Chapter 11. Jamie Oaks 1. April 11, Kincaid Hall 524. c 2007 Boris Kulikov boris-kulikov.blogspot.
 History of Lineages Chapter 11 Jamie Oaks 1 1 Kincaid Hall 524 joaks1@gmail.com April 11, 2014 c 2007 Boris Kulikov boris-kulikov.blogspot.com History of Lineages J. Oaks, University of Washington 1/46
History of Lineages Chapter 11 Jamie Oaks 1 1 Kincaid Hall 524 joaks1@gmail.com April 11, 2014 c 2007 Boris Kulikov boris-kulikov.blogspot.com History of Lineages J. Oaks, University of Washington 1/46
Station 1 1. (3 points) Identification: Station 2 6. (3 points) Identification:
 Identification: Station 2 6. (3 points) Identification:") SOnerd s 2018-2019 Herpetology SSSS Test 1 SOnerd s SSSS 2018-2019 Herpetology Test Station 20 sounds found here: https://drive.google.com/drive/folders/1oqrmspti13qv_ytllk_yy_vrie42isqe?usp=sharing Station
SOnerd s 2018-2019 Herpetology SSSS Test 1 SOnerd s SSSS 2018-2019 Herpetology Test Station 20 sounds found here: https://drive.google.com/drive/folders/1oqrmspti13qv_ytllk_yy_vrie42isqe?usp=sharing Station
Most amphibians begin life as aquatic organisms and then live on land as adults.
 Section 3: Most amphibians begin life as aquatic organisms and then live on land as adults. K What I Know W What I Want to Find Out L What I Learned Essential Questions What were the kinds of adaptations
Section 3: Most amphibians begin life as aquatic organisms and then live on land as adults. K What I Know W What I Want to Find Out L What I Learned Essential Questions What were the kinds of adaptations
Evolution as Fact. The figure below shows transitional fossils in the whale lineage.
 Evolution as Fact Evolution is a fact. Organisms descend from others with modification. Phylogeny, the lineage of ancestors and descendants, is the scientific term to Darwin's phrase "descent with modification."
Evolution as Fact Evolution is a fact. Organisms descend from others with modification. Phylogeny, the lineage of ancestors and descendants, is the scientific term to Darwin's phrase "descent with modification."