The Thermal Ecology of the European Grass Snake, Natrix natrix, in southeastern England. Leigh Anne Isaac B.E.S., York University, 1997
|
|
|
- Peter Bradley
- 5 years ago
- Views:
Transcription
1 The Thermal Ecology of the European Grass Snake, Natrix natrix, in southeastern England Leigh Anne Isaac B.E.S., York University, 1997 A Thesis Submitted in Partial Fulfillment of the Requirements for the Degree of MASTER OF SCIENCE In the Department of Biology O Leigh Anne Isaac, 2003 University of Victoria All rights reserved. This thesis may not be reproduced in whole or in part, by photocopy or other means, without the permission of the author.
2 Supervisor: Dr. P. T. Gregory ABSTRACT What factors influence the biology and distribution of animals? The ecology of ectotherms is inevitably about how these animals interact with their thermal environment because body temperatures (Tb; plural, Tbs) are determined principally by ambient conditions. Temperature is important because it influences rates of basic biological processes (e.g., membrane transport), thus affecting physiological functions. Terrestrial ectotherms, such as reptiles, are therefore compromised when they cannot maintain Tbs that optimize performance. This has potential behavioural and ecological consequences that can ultimately influence fitness. High-latitude environments are particularly challenging for ectotherms because they experience wide variations in temperature both spatially and temporally. Here I study the thermal ecology of a high-latitude natricine, the European Grass Snake, Natrix natrix, using field and laboratory methods, at a site near Fordwich, UK. I collected spot Tb measurements of Grass Snakes caught in the field. Grass Sakes were active over a broad range of Tbs and snakes maintained relatively constant Tbs when environmental temperatures were high. In order to monitor Tbs in a more controlled environment, I used radiotelemetry to continuously record Tbs of Grass Snakes in an outdoor enclosure in June-July I used physical models to estimate the environmental temperatures, or operative temperatures (T,; plural, T,s) available to snakes. I then determined if snakes thermoregulated when given the opportunity (high T,s). Snakes achieved high Tbs during the hottest part of the day, although these opportunities were limited ( 45 % of the days).
3 From piecewise regression, I showed that Grass Snakes initiated thermoregulation at relatively high T,s (T,= OC, Tb= 27.7 OC) and maintained fairly stable Tbs (30 OC) as T,s increased. Body temperatures similar to T,s at lower T,s, however, suggested only moderate thermoregulatory behaviour at these temperatures. Contrary to expectation, nongravid females maintained significantly higher Tbs than gravid females at lower T,s, however, this difference was nonsignificant at high T,s. Maintaining optimal Tbs in the field might be compromised by risks such as predation. In order to determine what Tbs snakes would select in the absence of these risks, I used radiotelemetry to determine the preferred or selected temperatures (TSet) of Grass Snakes in the laboratory. Body temperatures of snakes were monitored in a thermogradient over a 24-hr period during June and August Despite significant variation among individuals, all snakes maintained high Tbs (Tbs>30 OC). These high Tset values exceed those reported in the literature for other natricines. I then asked whether Tbs maintained by snakes in the field and chosen in the laboratory coincided with Tbs where performance is optimized. I tested performance of 3 functions (crawling, swimming & tongue-flick rate) over a range of Tbs in the laboratory. Mean and maximum speeds of crawling and swimming were highly temperaturedependent and maximum speeds occurred over a broad range of high Tbs that included temperatures maintained in the field and in the laboratory. Tongue-flick rates, however, appeared to be relatively independent of temperature but I cannot rule out experimental design flaws in this case. Overall, the data I have collected suggest that the thermal ecology of Grass Snakes in Fordwich, UK is similar to that of other temperate-zone natricines.
4
5 TABLE OF CONTENTS ABSTRACT ii... TABLE OF CONTENTS...v LIST OF TABLES LIST OF FIGURES..vill... ACKNOWLEDGEMENTS...x... GENERAL INTRODUCTION.. 1 CHAPTER ONE: NATURAL HISTORY AND THERMAL ECOLOGY OF THE EUROPEAN GRASS SNAKE, NATRIX NATRIX, IN SOUTHEASTERN ENGLAND Introduction Study Species Study Site Methods Results...I5 Discussion Literature Cited CHAPTER TWO: THERMOREGULATION AND THERMAL LIMITATION AT A HIGH LATITUDE: COMPARISON OF GRAVID AND NONGRAVID FEMALES OF AN OVIPAROUS ECTOTHERM, THE GRASS SNAKE (NATRIX NATRIX) Introduction Methods Results Discussion Literature Cited vii
6 CHAPTER THREE: THERMAL PREFERENCE AND PERFORMANCE OF THE EUROPEAN GRASS SNAKE. NATRIX NATRIX Introduction Methods Results Discussion Literature Cited GENERAL CONCLUSIONS... 82
7 vii LIST OF TABLES CHAPTER 1: Table 1. Morphometric data for Grass Snakes at Fordwich, UK. Females were larger than males, on average. Females with stump tails were excluded from the tail length analysis. Only snakes weighed with no food item were included in the weight summary Table 2. Summary of food items found in stomachs of Grass Snakes at Fordwich, UK. One animal was severely injured and therefore not included in the analysis CHAPTER 2: Table 1. Summary of operative temperatures (Tes) measured by the snake models (copper-pipe models and TidbiTBs) at various locations within the enclosure from June-July Temperatures vary by location, but overall, the warmest spot was in full sun. All Tes were calibrated with the snake model (see text). The discrepancy in sample size was due to equipment malfunction Table 2. Summary of operative temperatures (Tes) measured by the copper-pipe model exposed to full sun. Thermoregulatory opportunities were greatest, when Te>300C, during midday. Given are the number of days and percentage of the total days when each time period met the temperature criteria. Uneven sample size is attributable to equipment malfunction CHAPTER 3: Table 1. a) Summary of operative temperatures (Tes) measured in various locations throughout the outdoor enclosure during the day ( ). Snakes could reach high Tbs (i.e. Tes228.6OC) in 4 of the 6 locations ('*'). b) Summary of Tes measured in the same locations during the night ( ). Snakes had no opportunities to raise their Tbs to high levels (i.e. Tes228.6"C) in any microhabitat Table 2. Summary of the number of daytime periods when snake Tbs could reach at least 28.6"C and 38.4"C (two estimates of Tset, see text). Shown are the warmest microhabitats measured in the outdoor enclosure. Tset could be achieved more frequently in full sun
8 LIST OF FIGURES CHAPTER 1: Fig. 1. The Grass Snake, Natrix natrix. (Photo: R. A. Griffiths) Fig. 2. Map of England, Kent County and aerial view of the study site in Fordwich, UK. (Maps: Aerial photograph: Fig. 3. Typical microhabitats used by Grass Snakes. a) Branch piles and blackberry bushes, and b) stinging nettles make ideal retreat sites. (Photos: P. T. Gregory) Fig. 4. Unique colouration pattern of the ventral side of a Grass Snake. (Photo: P. T. Gregory).... Fig. 5. a) Body temperatures (Tbs) of snakes caught in Fordwich, UK, and b) operative temperatures (T,s) at point of capture. There was a significant difference between Tbs and Tes. c) Operative temperatures (Tes) measured when snakes escaped capture. There was no significant difference between T,s of captured snakes vs. Tes of snakes seen but not captured Fig. 6. Scatterplot of body temperatures (Tbs) versus operative temperatures (T,s) at Fordwich, UK. Each dot represents a single field measurement. The fitted quadratic line is indicated by red '*'s and the reference line (slope=l) is indicated by the straight black line CHAPTER 2: Fig. 1. a) Photograph of the outdoor enclosure at the University of Kent at Canterbury in Canterbury, UK. b) Aerial schematic of the various microhabitats within the 2 64 m enclosure Fig. 2. Box-and-whisker plot of operative temperatures (Tes) collected over the study period that were measured by the snake model in full sun. The data are divided into eight 3-hr time periods. Thermoregulatory periods when Tes>30 "C, are limited to midday ( ). Boxes represent the interquartile range (50 % of the data) divided by the median. Arithmetic mean is presented by the dot. Whiskers extend from the minimum to the maximum data values Fig. 3. Scatterplot of mean body temperatures (Tbs) VS. mean operative temperatures (Tes) measured in full sun for gravid and non-gravid females during the study period. Each point represents the mean Tb for one snake in a particular time period on a particular day. The black dashed line indicates slope= 1 and points falling along this line imply perfect thermoconformity (i.e. Tb=T,). Dots indicate mean Tbs for gravid females and stars indicate mean Tbs for nongravid females...i6
9 The solid red line is the 'predicted' thermoregulatory pattern from piecewise regression for all snakes regardless of reproductive condition CHAPTER 3: Fig. 1. Boxplot of mean crawling speeds (dsec) measured at three different body temperatures (cool: 16 "C, room: 21 "C, warm: 30 "C). Mean crawling speed increases with rising body temperature. Boxes represent the interquartile range (50 % of the data) divided by the median. Arithmetic mean is presented by the dot. Whiskers extend from the minimum to the maximum data values Fig. 2. Boxplot of maximum crawling speeds (dsec) measured at three different body temperatures (cool: 16 "C, room: 21 "C, warm: 30 "C). Maximum crawling speed increases with rising body temperature Fig. 3. Boxplot of mean crawling speeds over a higher range of body temperatures (room: 25 "C, warm: 30 "C, hot: 34 "C, very hot: 38 "C). Mean crawling speed increased slightly, levelled off, and was followed by a nonsignificant reduction in speed. Data collected by P. T. Gregory Fig. 4. Boxplot of maximum crawling speeds over a higher range of body temperatures (room: 25 "C, warm: 30 "C, hot: 34 "C, very hot: 38 "C). Maximum crawling speed did not vary significantly with temperature. Data collected by P. T. Gregory Fig. 5. Boxplot of mean swimming speeds (dsec) measured at three different body temperatures (cool: 16 "C, room: 21 "C, warm: 30 "C). Mean swimming speed increases with rising body temperature Fig. 6. Boxplot of maximum swimming speeds (dsec) measured at three different body temperatures (cool: 16"C, room: 21 C, warm: 30 C). Maximum swimming speed increases with rising body temperature Fig. 7. Boxplot of mean tongue flick frequency over a 30-sec interval at different body temperatures. Mean frequency is greater at lower temperatures Fig. 8. Boxplot of maximum tongue flick frequency over a 30-sec interval at different temperatures. Maximum frequency is greater at lower temperatures Fig. 9. Boxplot of body temperatures selected (T,,,) by three snakes in a thermogradient in a) June, 2002, and b) the same three snakes in a thermogradient in August, Fig. 10. Histogram of operative temperatures (T,s) measured in a) in an underground pipe, b) in full shade, c) in a rock pile, d) in the pond, e) in full sun and f) under a metal sheet. Note different scales in some of the graphs....75
10 ACKNOWLEDGEMENTS I would like to thank my supervisor, Pat Gregory, for showing me how important it is to always keep your question in mind when doing research. He has helped me develop both my critical thinking and writing skills and I am grateful. I am indebted to my husband, Doug Schneider, who was instrumental in catching animals, completing laboratory experiments, being extremely patient, and for being an overall awesome guy. I am grateful to Dr. R.A. Griffiths of the University of Kent at Canterbury, Canterbury, Kent, UK for his logistical support of my project. Thank you to Pat Kerfoot of the Biology Electronics Laboratory for his technical assistance and to Colin May of Faversham, Kent, UK for generously providing all weather data. Eleanore Floyd of the Biology Department provided invaluable support throughout my graduate career. This research was financially supported by an N.S.E.R.C. PGS A Scholarship held by L. A. Isaac and an N.S.E.R.C. Research Grant held by P. T. Gregory. Funding was also provided by the University of Victoria.
11 GENERAL INTRODUCTION Temperature is a major physical factor influencing the biology and distribution of animals (Root, 1988; Humphries et al., 2002). Because of its influence on the rates of biological processes (e.g., membrane function: Gracey, et al., 1996), temperature affects physiological functions and, by extension, when and where animals can be active (Chappell & Bartholomew, 1981; Grant & Dunham, 1988). Animals are compromised when they cannot maintain body temperatures (Tb; plural, Tbs) that optimize performance. This has potential behavioural and ecological consequences that can ultimately influence fitness (Christian & Tracy, 1981; Beaupre, 1995). High latitudes and high altitudes are therefore challenging environments because they experience wide variations in temperature, both temporally and spatially (Huey & Kingsolver, 1989; Blouin-Demers & Weatherhead, 2001). Such environments are particularly challenging for ectotherms, or cold-blooded animals, because they have a relatively limited physiological capacity for generating significant heat metabolically (Hutchison et al., 1985). Rather, ectotherms rely on external sources of heat to maintain Tbs. Terrestrial ectotherms, such as reptiles, potentially can meet the challenges of a in various ways that are not mutually exclusive. They can: 1. avoid lethally extreme surface temperatures by hibernating or aestivating (Gregory, 1984; Grant & Dunham, 1988), 2. tolerate a wide range of Tbs allowing activity over a wider range of T,s (Kingsbury, 1994; Dorcas, 1998), and 3. thermoregulate, when possible, to maintain optimum Tbs. In reptiles, behavioural thermoregulation, such as microhabitat selection and postural adjustments, is typically more important than physiological mechanisms in controlling Ths (Stevenson, 1985). Because the main thermal challenges facing reptiles are in temperate zones rather than the tropics (Shine &
12 Madsen, 1996), reptiles with northerly distributions provide an opportunity to investigate issues in thermal biology where potential 'costs' are perhaps more extreme. An important question is the extent to which temperate-zone reptiles are able to thermoregulate. In order to evaluate thermoregulatory patterns, three different temperature measurements should be collected: 1. body temperature, Tb, describes the temperature of an animal in a given environment, 2. operative temperatures (T,; plural, T,s) describe the environmental temperatures that are available for an animal to exploit, and finally, 3. preferred temperature, Tse,, describes the Tt, an animal would select when given a wide range of temperature options. Ultimately, thermoregulation should have fitness consequences. One explicitly fitness-oriented model of thermoregulation is that of Huey and Slatkin (1976), who argue that the extent to which reptiles thermoregulate reflects the relative 'benefits' and 'costs' associated with such behaviour as well as the constraints imposed by the environment (Grant & Dunham, 1988; Christian & Bedford, 1995). Benefits include physiological or ecological advantages that arise from thermoregulating, such as accelerated rates of development and increased copulatory frequency that ultimately influence the fitness of an individual. Thermoregulatory costs, on the other hand, include disadvantages that an individual incurs, such as increased predation risk and increased energy expenditure that have potential negative fitness consequences. Huey and Slatkin (1976) suggest that reptiles should stop thermoregulating, and thermoconform to the thermal environment, when these costs outweigh the benefits. It is generally assumed that thermoregulatory costs are relatively high in the temperate zone, largely because environmental temperatures are highly variable and thermoregulatory opportunities may be somewhat limited. Recent
13 work (Blouin-Demers & Weatherhead, 2001), however, suggests that reptiles in thermally challenging temperate-zone environments can be moderately effective thermoregulators overall. More effective thermoregulators maintain Tbs in the field that are closer, on average, to preferred Tbs that are measured in the laboratory. Another factor influencing thermoregulatory behaviour is the physiological state of an individual (Charland & Gregory, 1990; Daut & Andrews, 1993; Burns et al., 1996). Much attention has been focused on reproductive state because developmental rate, viability of offspring and phenotype of offspring are all markedly affected by temperature (van Damme et al., 1992; Rock & Cree, 2003). Thus, gravid females of viviparous snakes usually are found to thermoregulate more precisely around a given Tb than do non-gravid females in the same environmental conditions (Beuchat, 1986; Charland, 1995; Brown & Weatherhead, 2000). Although females of oviparous species retain eggs for a shorter period of time than do viviparous species, they also might be expected to thermoregulate in a similar fashion while gravid. Few studies have been done of oviparous species, but recent work (Blouin-Demers & Weatherhead, 2001) supports this prediction. Although most species of snakes are oviparous (=80%), viviparous species predominate at high latitudes and altitudes. This is presumably because viviparous females can maintain the best possible thermal conditions for their developing offspring by retaining them throughout the entire pregnancy ('cold-climate' hypothesis; Shine, 1985, 1987a, b). Oviparous species, by contrast, can use thermoregulation to optimize development of offspring only while eggs are in utero; following oviposition, further development depends on ambient temperatures in the nest site (Vinegar, 1974; Gutzke & Packard, 1987), which may be quite variable at high latitudes. Thus, oviparous species are rare or absent at high latitudes.
14 The European Grass Snake, Natrix natrix, is one of the most common and widespread snakes in Europe, ranging from northern Africa to near the Arctic Circle in Scandinavia. Thus, it occurs in a wide range of thermal environments making it ideal for comparative geographic studies. It also is ecologically similar and phylogenetically related to North American snakes of the genus Thamnophis (garter snakes), which have been widely used in studies of thermal ecology (Hawley & Aleksiuk, 1975; Gregory, 1977; Gibson & Falls, 1979; Lysenko & Gillis, 1980; Gibson et al., 1989), including tests of the cold-climate hypothesis (Charland, 1995). By contrast, the thermal ecology of Natrix natrix is relatively unstudied, except for occasional field observations (Mertens, 1994; Gentilli & Zuffi, 1995). Natrix also differs from Thamnophis in reproductive mode (Natrix: egg-laying; Tharnnophis: livebearing), and thus offers an opportunity to test a logical corollary of the cold-climate hypothesis: that high-latitude oviparous snakes should thermoregulate in a similar fashion to viviparous snakes while gravid. In this thesis, I focus on several aspects of the thermal ecology of Natrix natrix in southern England. The specific questions I ask are: How does performance vary with temperature? What Tbs do snakes select when given a wide range of temperatures from which to choose? How often can snakes reach their presumed optimal Tbs? What thermoregulatory patterns do snakes display in a semi-natural environment? Do gravid females thermoregulate differently from nongravid females?
15 LITERATURE CITED Beuchat, C. A Reproductive influences on the thermoregulatory behaviour of a live-bearing lizard. Copeia 1986(4): Beaupre, S. J Effects of geographically variable thermal environment on bioenergetics of Mottled Rock Rattlesnakes. Ecology 76(5): Blouin-Demers, G. & Weatherhead, P. J Thermal ecology of black rat snakes (Elaphe obsolete) in a thermally challenging environment. Ecology 82(11): Brown, G. P. & Weatherhead, P. J Thermal ecology and sexual size dimorphism in Northern Water Snakes, Nerodia sipedon. Ecol. Mono. 70(2): Burns, G., Ramos, A. & Muchlinski, A Fever response in North American snakes. J. Herp. 30 (2): Chappell, M. A. & Bartholomew, G. A Activity and thermoregulation of the Antelope Ground Squirrel, Ammospermophilus leucurus, in winter and summer. Physiol (2): Charland, M. B Thermal consequences of reptilian viviparity: Thermoregulation in gravid and nongravid Garter Snakes (Thamnophis). J. Herp. 29(3) Charland, M. B. & Gregory, P. T The influence of female reproductive status on thermoregulation in viviparous snake, Crotalus viridis. Copeia 1990: Christian, K. A. & Tracy, C. R. 198 I. The effect of the thermal environment on the ability of hatchling Galapagos Land Iguanas to avoid predation during dispersal. Oecologia (Berl) 49: Christian, K. A. & Bedford, G. S Seasonal changes in thermoregulation by the Frillneck Lizard, Chlamydosaurus kingii, in tropical Australia. Ecology 76 (1): Daut, E. F. & Andrews, R. M The effect of pregnancy of thermoregulatory behaviour of the viviparous lizard Chalcides ocellatus. J. Herp. 27(1): Dorcas, M. E Daily temperature variation in free-ranging rubber boas. Herpetologica 54(1): Gentilli, A. & Zuffi, M. A. L Thermal ecology of a Grass Snake (Natrix natrix) population in Northwestern Italy. Amphibia-Reptilia 16:
16 Gibson, A. R. & Falls, J. B Thermal biology of the common garter snake Thamnophis sirtalis. I. Temporal variation, environmental effects and sex differences. Oecologia 43: Gibson, A. R., Smucny, D. A,, & Kollar, J The effects of feeding and ecdysis on temperature selection by young Garter Snakes in a simple thermal mosaic. Can. J. Zool. 67: Gracey, A. Y., Logue, J., Tiku, P. E. & Cossins, A. R Adaptation of biological membranes to temperature: biophysical perspectives and molecular mechanisms. In Animals and Temperature: Phenotypic and Evolutionary Adaptation. (Johnston, A. & Bennett, A. F. eds.). Cambridge University Press, New York. pp: Grant, B. W. & Dunham, A. E Thermally imposed time constraints on the activity of the desert lizard Sceloporous merriami. Ecology 69(1): Gregory, P. T Life-history parameters of the Red-Sided Garter Snake (Thamnophis sirtalis parietalis) in an extreme environment, the Interlake region of Manitoba. Natl. Mus. Can. Publ. Zool. 13: Gregory, P. T Communal denning in snakes. In Vertebrate Ecology and Systematics: A Tribute to Henry S. Fitch. (Seigel, R. A., Hunt, L. E., Knight, J. L., Malaret L., & Zuschlag, N. L. eds). Univ. Kans. Mus. Nat. Hist. Spec. Pub. 10: Gutzke, W. H. N. & Packard, G. C Influence of the hydric and thermal environments on eggs and hatchlings of Bull Snakes, Pituophis melanoleucus. Physiol. Zool. 60(1): Hawley, A. & Aleksiuk, M Thermal regulation of spring mating behaviour in the Red-Sided Garter Snake (Thamnophis sirtalis parietalis). Can. J : Huey, R. B. & Kingsolver, J. G Evolution of thermal sensitivity of ectotherm performance. TREE 4(5): Huey, R. B. & Slatkin, M Cost and benefits of lizard thermoregulation. Quart. Rev. Biol. 51(3): Humphries, M. M., Thomas, D. W., & Speakman, J. R Climate-mediated energetic constraints on the distribution of hibernating mammals. Nature 418: Hutchison, V. H., Dowling, H. G., & Vinegar, A Thermoregulation in a brooding female python, Python molurus bivittat~is. Science 151: Kingsbury, B. A Thermal constraints and eurythermy in the lizard Elgaria multicarinata. Herpetologica 50(3):
17 Lysenko, S. & Gillis, J. E The effect of ingestive status on the thermoregulatory behaviour of Tharnnophis sirtalis sirtalis and Tharnnophis sirtalis parietalis. J. Herp. 14(2): Mertens, D Some aspects of thermoregulation and activity in free-ranging Grass Snakes (Natrix natrix L.). Arnphibia-Reptilia 15: Rock, J. & Cree, A Intraspecific variation in the effect of temperature on pregnancy in the viviparous gecko, Hoplodactylus rnaculatus. Herpetologica. 59(1): Root, T Environmental factors associated with avian distributional boundaries. J. Biogeogr. 15: Shine, R The evolution of viviparity in reptiles: An ecological analysis. In Biology of the Reptilia, vol. 15. (Gans, C. & Billet, F. eds.). John Wiley and Sons, New York. pp a. Reproductive mode may determine geographic distributions in Australian venomous snakes (Pseudechis, Elapidae). Oecologia (Berl) 71: b. The evolution of viviparity: ecological correlates of reproductive mode within a genus of Australian snakes (Pseudechis, Elapidae). Copeia 1987: Shine, R. & Madsen, T. E Is thermoregulation unimportant for most reptiles? An example using water pythons (Liasis fuscus) in tropical Australia. Physiol (2): Stevenson, R. D The relative importance of behavioural and physiological adjustments controlling body temperature in terrestrial ectotherms. Am. Nut. 126(3): van Damme, R., Bauwens, D., Brana, F., & Verheyen, R.F Incubation temperature differentially affects hatching time, egg survival, and hatchling performance in the lizard Podarcis muralis. Herpetologica 48(2): Vinegar, A Evolutionary implications of temperature induced anomalies of development in snake embryos. Herpetologica 30:
18 CHAPTER 1: Natural History and Thermal Ecology of the European Grass Snake, Natrix natrix, in southeastern England INTRODUCTION The design and interpretation of experimental results in biology requires an appropriate context. That is, are the conditions imposed in the experiment within or outside the range of those that the organism might be expected to encounter in the wild? Natural history, the study of organisms in their natural settings, provides that context (Greene, 1986). Natural history also plays a key role in the development of theory and in testing hypotheses derived from theory (Greene, 1986). The questions addressed by natural historians pervade most disciplines of contemporary biology - ecology, evolutionary biology and population genetics (Bartholomew, 1986); natural history thus is the vehicle by which these disparate areas can be integrated. The thermal ecology of ectotherms, such as snakes, is inevitably about how animals interact with their physical environment because body temperatures (Tb; plural, Tbs) are determined principally by ambient conditions rather than by metabolic generation of heat. Determination of the conditions under which such animals are active in the field thus provides essential background for designing and interpreting experiments to test ideas about temperature choice and thermoregulation. In this thesis, I address various questions about the thermal ecology of the Grass Snake, Natrix natrix, a widespread and abundant species in Europe. Wideranging species are especially interesting because they are likely to be "broad-niched" (Brown, 1995), tolerating, for example, a wide range of thermal conditions. Despite its abundance, however, no comprehensive study has been done of the thermal ecology of the Grass Snake. Such study should complement and test the generality of work done on other high-latitude taxa of snakes, especially the related and
19 ecologically similar, and well-studied Garter Snakes (Thamnophis) of North America (Hawley & Aleksiuk, 1975; Gregory, 1977; Gibson & Falls, 1979; Lysenko & Gillis, 1980; Gibson et al., 1989). Here I describe the general natural history of the Grass Snake, with a particular focus on characterizing the body temperatures of free-ranging animals at a site in southeastern England. STUDY SPECIES The Grass Snake is widely distributed across much of Europe. It ranges from Lake Baikal (Russia) in the east to western Portugal and from northern Africa to southern Scandinavia, including England and various Mediterranean islands (Beebe & Griffiths, 2000). Grass Snakes are slate-green in colour with a distinctive yellow and black band around the neck and black marks extending down the dorsal side (Fig. I). Adult Grass Snakes range in body size from 500 to 800 mm. As in other natricine snakes (Garter Snakes, Gregory, 1977; Water Snakes, Weatherhead et al., 1995), females are significantly larger than males (Luiselli et al., 1997). Fecundity is strongly related to female body size, with larger females producing larger clutches (between 4-24 eggs; Madsen, 1987; Luiselli et al., 1997). Grass Snakes are generalist predators preying upon anurans, small mammals, birds and fish (Reading & Davies, 1996; Luiselli et al., 1997; Gregory & Isaac, in press). There is relatively little geographic variation in the major prey categories of reported diets; amphibians, particularly anurans (i.e., frogs and toads), are typically the most frequent prey type (Brown, 1991; Reading & Davies, 1996; Luiselli et al., 1997). Seasonal activity begins with emergence from hibernation typically by males in early spring (Luiselli et al., 1997). Mating takes place in early to late spring and

20 Fig. 1. The Grass Snake, Natrix natrix. (Photo: R. A. Griffiths)
21 male movements during this period can be extensive (mean distance travelled: m SD per day; Madsen, 1984). Female movements are typically most extensive immediately prior to and after oviposition (mean distance travelled: m SD per day), which presumably occurs in mid-summer (Madsen, 1984). Hatching occurs in late summer to early fall and hibernation is thought to begin by late fall (Luiselli & Shine, 1997). Grass Snakes exhibit an unusual behaviour called death-feigning. This behaviour has been observed in other species such as the Western Hognose Snake (Burghardt & Greene, 1988) and the Western Whip Snake (Rugiero, 1999). Deathfeigning may include all or some of the following components: limp body, rolled eyes, everted cloaca, extended tongue, and/or spontaneous bleeding from the mouth. One possible hypothesis for this behaviour is that it is an anti-predator strategy but it has not been studied in any detail. STUDY SITE I studied a population of Grass Snakes in southern England, near Fordwich, 3.25 krn NE of Canterbury, Kent, UK (58ON 17.58'N, 1 N 08.19'E, 5 m elevation) (Fig. 2). Although there is considerable variability in climatic conditions between months, the temperate climate of this region is characterized by cool, wet winters and warm, dry summers. My study area encompassed habitats on either side of the River Stour and included predominantly second-growth deciduous woods and open fields with walking paths and fishing spots. The principal vegetation was Stinging Nettles (Urtica dioica) and Common Reed (Phragmites australis) as well as various species of brambles (Rubus sp.). Large portions of the site were once used for gravel extraction, these quarries have since filled with water to form small lakes. This network of ponds provided ideal habitat for Grass Snakes and their prey (Fig. 3 a, b).
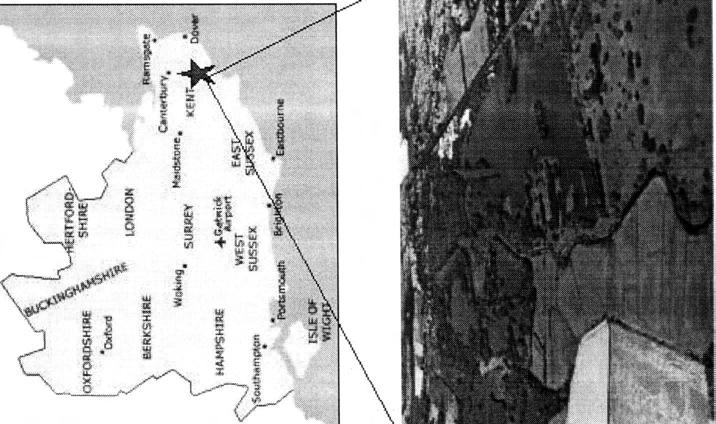
22

 Branch pi des and blackberry bushes")
23 Fig. 3. Typical microhabitats used by Grass Snakes. a) Branch pi des and blackberry bushes and b) stinging nettles make ideal retreat sites. (Photos: P. T. Gregory)
24 METHODS I caught Grass Snakes opportunistically by hand and by flipping cover objects. Immediately following each capture, I measured the snake's cloaca1 temperature (body temperature, Tb) using a mercury thermometer. I also measured operative temperatures (T,; plural, T,s) at the snake's capture site. Operative temperature provides an estimate of the potential Tbs a snake could achieve if it thermoconformed to its thermal environment. Thus, a thermoconforming snake will have Tb=T,, whereas a thermoregulating snake will have Tb+T,. Physical models, such as copper pipe models with thermocouples inserted inside, are usually used to measure T, (Peterson et al., 1993). To measure T, of field-captured snakes, I placed the mercury thermometer, inserted inside its metal casing, in the same location the snake was caught and I allowed it to equilibrate for at least one minute before reading the temperature. Next, I measured a set of standard variables. I determined sex by examining the shape of the tail (female tails taper immediately following the vent) or by using a probe (probe extends down tail if a male). I measured snout-vent-length (SVL) and tail length to the nearest 1 mm by stretching the snake along a metre stick. Head diameter was determined by placing the snake's head into the different sized holes in an inking template. Body mass was measured using a hand-held Pesola spring scale. I palpated female snakes to determine the presence or absence of ovarian follicles or eggs and to estimate their numbers if present. Presence of stomach contents was obtained in a similar fashion, by gentle palpation. I scored whether injuries were present or absent and whether snakes exhibited any signs of death-feigning behaviour. In order to compare injury rates and death-feigning behaviour between different sizes,
25 I divided the sample into two size groups (small and large snakes) by the median SVL (699 mm). Finally, I took a photograph of the unique patterns on the ventral side of each snake to be used for identification of individual animals (Fig. 4). Because snakes that are warm also are faster, they might elude capture more easily. If so, spot-temperature data taken in the field will be biased towards cooler individuals. This idea can be tested, at least partially, by comparing T,s of snakes that were caught vs. those that were seen but not captured. I collected T, data for snakes that were not captured by placing the mercury thermometer, inserted inside its metal casing, in the same location each snake was originally seen. I again allowed the thermometer to equilibrate for at least one minute before reading the temperature. RESULTS In 2002, the mean annual temperature ( O C SD) and the mean summer temperature (May to August; "C SD) did not significantly differ from yearly and summer averages respectively, from 1999,2000 & 2001 (annual: F(3,471= 0.14, P= 0.94, summer: Fo, 15)= 0.17, P= 0.92). Similar patterns were also observed for rainfall (annual: mm SD; F(3,47)= 0.76, P= 0.53, summer: mm SD; F(3. IS)= 0.04, P= 0.99). All weather data were collected in Faversham, Kent County, approximately 50 krn from my study site (Colin May: h~p:nwww.ci4386.den1on.co.uk/weather/weaher.htm). Thus, the weather to which snakes were exposed during my study was broadly typical of the study site. From May to August 2002, I caught 63 Grass Snakes (28 males, 35 females) in a variety of different microhabitats such as open patches of vegetation, fallen tree trunks, and piles of dead blackberry bushes. Grass Snakes are semi-aquatic and I saw them retreat to water when threatened, but I did not observe them in open water.

26 Fig. 4. Unique colouration pattern of the ventral side of a Grass Snake. (Photo: P. T. Gregory)
27 I caught snakes with a wide range (18 OC to OC) of Tbs (mean Tb: OC SD), but I rarely encountered snakes when conditions were extremely hot (Tes>35 OC; Fig. 5). Tbs and T,s were significantly correlated (r= 0.63, P<0.0001, n=43). However, the relationship between Tb and T, was best described by a polynomial (i.e., quadratic) regression, which was significant (r2= 0.47, F(2,'$2)= 17.71, P<0.0001; Fig. 6). Tbs matched T,s closely at low T,s, however, Tbs were independent of T,s at higher T,s, suggesting thermoregulation. Contrary to my expectation, the frequency of low and high T,s (divided at the median T,, 24.0 "C) was similar for snakes that I caught and snakes that were seen but not caught (Chi-square: X2= , d+ 1, P=0.6158). There was, therefore, no evidence that captures were biased towards snakes active under cooler conditions. Females were significantly longer than males (ANOVA: F[I, 62)= 9.89, P=0.003; Table 1). When adjusted for SVL, tail length also significantly differed between the sexes (ANCOVA: F(1,58) = 7.84, P=0.007; slopes equal). Males had relatively longer tails than females. Females had significantly larger heads than males (ANCOVA: F(1, 59)= , P<0.0001; slopes equal) and were consistently heavier than males relative to their body size (ANCOVA: (F(,, 57)= 54.54, Pe0.0001; slopes equal). Approximately half of the Grass Snakes (30 of 62 animals) captured had food in their stomachs. The most frequent prey were amphibians, all anurans (Table 2). The introduced Marsh Frog, Rana ridibunda, was the prey item most frequently eaten. Approximately 52 % of the animals captured (33 of 63 animals) had evidence of injuries of some type, including healed bone breaks, scars in various regions of the body as well as recent injuries (i.e., open wounds). I did not witness any direct predation attempts but presumably they are avian (Madsen, 1987). There was no
28 Body Temperature Vb) (C) Opefatlve Temperature (Te) (C) Operative Temperatures m) (C) 4 Fig. 5. a) Body temperatures (Tbs) of snakes caught in Fordwich, UK, and b) operative temperatures (T,s) at point of capture. c) Operative temperatures (T,s) measured when snakes escaped capture.
29 Operative Temperature (Te) (C) Fig. 6. Scatterplot of body temperatures (Tbs) versus operative temperatures (T,s) at Fordwich, UK. Each dot represents a single field measurement. The fitted quadxakic line is indicated by red '*'s and the reference line (slope=l) is indicated by the straight black line.
30 Table 1. Morphometric data for Grass Snakes at Fordwich, UK. Females were larger than males, on average. Females with stump tails were excluded from the tail length analysis. Only snakes weighed with no food item were included in the weight summary. Variable Sex n Mean SD Min Max SVL Males Females Tail Length Males Females Weight Males Females Head Males Diameter Females
31 Table 2. Summary of food items found in stomachs of Grass Snakes at Fordwich, UK. One animal was severely injured and therefore not included in the analysis. Prey Type Number of Snakes Containing Prey Type Amphibians Toads Frogs Small Mammals 7 Birds I Unknown item No food 34 TOTAL SNAKES 62
32 significant difference in injury rate between the sexes (Chi-Square: X2= 0.36, dfi 1, P=0.55). There was, however, a significant different in injury rates between size groups. Larger snakes (22 of 33 animals) had significantly more injuries than smaller snakes (Chi-Square: x2= 8.45, dfi 1, P=0.004). Only 25 of 62 animals (40 %) death-feigned upon capture. There was no significant difference in the occurrence of death-feigning either between the sexes (Chi-Square: x2= 1.43, df= 1, P=0.23) or between different sized snakes (Chi-Square: x2= 2.57, df= 1, P=O.11). DISCUSSION Overall, the data I collected suggest that Grass Snakes at Fordwich had a similar natural history to that described for populations elsewhere. Common features of this species' natural history include a diet principally of anurans (Reading & Davies, 1996), pronounced sexual size dimorphism (Madsen & Shine, 1997), and a broad range of activity temperatures (Mertens, 1994). Injury rates have not previously been reported in this species, but the pattern I observed is similar to that reported for other species of natricines, in which larger, presumably older, animals have a higher frequency of injuries (Fitch, 2003). Death-feigning was observed in a high proportion of the animals captured, but was not related to either sex or size of animal. Interpreting body temperature variation of field-active animals in the context of an animal7 s natural history, including available environmental temperatures, can provide insights into thermal strategies used by reptiles (Peterson et al., 1993). One important generalization concerning body temperature (Tb) variation is that when possible, many species maintain Tbs within a relatively narrow range (stenothermy), using behavioural mechanisms (Brattstrom, 1965; Avery 1982; Stevenson, 1985).
33 Although the necessity of thermoregulation for most reptiles has been debated in the literature (Shine & Madsen, 1996; Akani et al., 2002; Luiselli & Akani, 2002), the benefits of such behaviour include enhanced locomotory abilities (Stevenson, et al., 1985), increased rates of digestion (Skocylas, 1970; Lillywhite, 1987), and accelerated growth and developmental rates (van Damme et al., 1992; Autumn & de Nardo, 1995). Despite the benefits of stenothermy, some species have highly variable Tbs during periods of activity (eurythermy), particularly species living in thermally variable environments (e.g.,, high latitudes or high altitudes; Garter Snakes, Peterson, 1987; Rubber Boas, Dorcas & Peterson, 1998; Water Snakes, Brown & Weatherhead, 2000, Rat Snakes: Blouin-Demers & Weatherhead, 2001). In this study, I have shown that Grass Snakes are active over a broad range of Tbs (18 OC to OC), consistent with previous work on populations in Germany and Italy (Mertens, 1994; Gentilli & Zuffi, 1995). Despite variation in Tbs, it is evident that Grass Snakes are not simply thermally passive; Tbs plateau at high T,s as snakes thermoregulate and presumably seek refuge from lethally high ambient temperatures. Thus, it is likely that Tbs of snakes I caught did not differ significantly from those that I did not catch. This conclusion is supported by the lack of significant differences between the T,s of each group. Presumably, my failure to catch some snakes depended on other details of the encounter. What are the benefits of being active over a wide range of Tbs? The most apparent benefit is the ability to be active during periods when ambient temperatures are relatively low. This is particularly important for temperate-zone species, such as the Grass Snake, that experience wide fluctuations in temperature (T, range in the enclosure from May to August 2002: 10.7 OC to 70.5 OC; Mertens, 1994). This ability
34 to tolerate and perform at lower temperatures may allow the use of thermally marginal microhabitats, such as continuous forest (Kingsbury, 1994). By exploiting such cooler environments, Grass Snakes could gain access to different food resources. Even in eurythermal species, however, there are limits to the range of temperatures allowing activity. These limits are principally physiological and include lower digestion rates, reduced locomotory abilities, etc. Skocylas (1970), for example, tested digestion rates of Grass Snakes over different ambient temperatures and observed that digestion was optimized over a fairly broad range of moderate to high Tbs (25 "C and 35 "C), but was arrested at very low Tbs (5 "C). Similarly, I found that Grass Snakes crawled consistently well over a wide range of higher Tbs (25 C to 38 "C; see Chapter 3), but performance was hindered at low Tbs (15 "C). How might activity over a broad range of Tbs influence the fitness of Grass Snakes? In this study, I have shown that Grass Snakes maintain a wide range of Tbs in the field, but more important, that Grass Snakes can perform well across this temperature range (see Chapter 3). The relatively high incidence of injuries in this population suggests that encounters with predators are common, so that snakes must be able to move quickly at any Tbs at which they are active. However, the true fitness consequences of different thermal strategies have yet to be studied.
35 LITERATURE CITED Autumn, K. & de Nardo, D. F Behavioural thermoregulation increases growth rate in a nocturnal lizard. J. Herp. 29(2): Avery, R. A Field studies of body temperatures and thermoregulation. In Biology of the Reptilia v. 12. (Gans, C. & Pough, F. H. eds.). Academic Press, Toronto. pp Bartholomew, G. A The role of natural history in contemporary biology. Bioscience 36: Beebe, T. J. C. & Griffiths, R. A Amphibians and Reptiles. London: Harper Collins Publishers. Brown, J. H Macroecology. University of Chicago Press, Chicago. Brown, P. R Ecology and vagility of the Grass Snake, Natrix natrix helvetica LacCpede. Unpubl. PhD Diss., University of Southampton, UK. Brattstrom, B. H Body temperatures of reptiles. Am. Midl. Nut. 73(2): Burghardt, G. M. & Greene, H. W Predator simulation an duration of death feigning in neonate Hognose Snakes. Anim. Behav. 36: Fitch H. S Tail loss in garter snakes. Herpetologica 34(3): Gentilli, A. & Zuffi, M. A. L Thermal ecology of a grass snake (Natrix natrix) population in Northwestern Italy. Amphibia-Reptilia 16: Gibson, A. R. & Falls, J. B Thermal biology of the Common Garter Snake, Thamnophis sirtalis (L.). Oecologia (Berl.) 43: Gibson, A. R., Smucny, D. A., & Kollar, J The effects of feeding and ecdysis on temperature selection by young Garter Snakes in a simple thermal mosaic. Can. J. Zool. 67: Greene, H. W Natural history and evolutionary biology. In Predator-Prey Relationships. (Feder, M. E., & Lauder, G. V. eds.). Univ Chicago Press, Chicago. pp: Gregory, P. T Life-history parameters of the red-sided garter snake (Thamnophis sirtalis parietalis) in an extreme environment, the Interlake region of Manitoba. Natl. Mus. Can. Publ. Zool. 13: Gregory, P. T. & Isaac, L. A. In Press. Food habits of the Grass Snake in southeastern England: Is Natrix natrix a generalist predator? J. Herp.
36 Hawley, A. W. L. & Aleksiuk, M Thermal regulation of spring mating behaviour in the Red-Sided Garter Snake (Thamnophis sirtalis parietalis). Can. J : Luiselli, L., Capula, M., & Shine, R Food habits, growth rates, and reproductive biology of Grass Snakes, Natrix natrix (Colubridae) in the Italian Alps. J Lond. 241: Lysenko, S. & Gillis, J. E The effect of ingestive status on the thermoregulatory behaviour of Thamnophis sirtalis sirtalis and Thamnophis sirtalis parietalis. J. Herp. 14(2): Madsen, T Growth rates, maturation and sexual size dimorphism in a population of Grass Snakes, Natrix natrix, in southern Sweden. Oikos 40: Madsen, T Movements, home range size and habitat use of radio-tracked Grass Snakes (Natrix natrix) in southern Sweden. Copeia 1984(3): Madsen, T Cost of reproduction and female life-history tactics in a population of Grass Snakes, Natrix natrix, in southern Sweden. Oikos 49: Madsen, T. & Shine, R Phenotypic plasticity in body sizes and sexual size dimorphism in European Grass Snakes. Evolution 47(1): Mertens, D Some aspects of thermoregulation and activity in free-ranging Grass Snakes (Natrix natrix L.). Amphibia-Reptilia 15: Peterson, C. R., Gibson, A. R., & Dorcas, M. E Snake thermal ecology: causes and consequences of body temperature variation. In Snakes: Ecology and Behaviour. (Seigel, R. A. & Collins, J. T. eds.). McGraw-Hill, New York. Pp: Reading, C.J. & Davies, J. L Predation by Grass Snakes (Natrix natrix) at a site in southern England. J Lond. 239: Rugiero, L Death feignin in the Western Whip Snake, Coluber viridiflavus. Amphibia-Reptilia 20(4): Schaefer, W. H Diagnosis of sex in snakes. Copeia 1934: 181. Stevenson, R. D The relative importance of behavioural and physiological adjustments controlling body temperature in terrestrial ectotherms. Am. Nut. 126(3): Stevenson, R. D., Peterson, C. R., & Tsuji, J. S The thermal dependence of locomotion, tongue flicking, digestion, and oxygen consumption in the Wandering Garter Snake. Physiol (1):
37 Van Damme, R., Bauwens, D., Brana, F., & Verheyen, R. F Incubation temperature differentially affects hatching time, eggs survival, and hatchling performance in the lizard Pocarcis muralis. Herpetologica 48(2): Weatherhead, P. J., Barry, F. E., Brown, G. P., & Forbes, M. R. L Sex ratios, mating behaviour and sexual size dimorphism of the Northern Water Snake, Nerodia sipedon. Behav. Ecol. Sociobiol. 56:
38 CHAPTER 2: Thermoregulation and Thermal Limitation at a High Latitude: Comparison of Gravid and Nongravid Females of an Oviparous Ectotherm, the European Grass Snake (Natrix natrix) INTRODUCTION All else being equal, when given the opportunity, ectotherms should behave so as to maintain body temperatures (Tb; plural, Tbs) that are favourable to essential functions such as locomotion and digestion (Stevenson et al., 1985; Lillywhite, 1987; Beck, 1996). Terrestrial ectotherms frequently maintain relatively constant body temperatures despite variation in environmental temperatures (Peterson, 1987; Brown & Weatherhead, 2000; Blouin-Demers & Weatherhead, 2001), although animals in different physiological states may maintain different body temperature profiles (Gibson et al., 1989; Charland & Gregory, 1990). But how common is thermoregulatory behaviour in ectotherms? Shine and Madsen (1996) argue that thermoregulation actually is unimportant for most reptiles because the majority of them live in the tropics where the environment is relatively invariant. At most, they must avoid extremely hot conditions (Akani et al., 2002; Luiselli & Akani, 2002). At the other extreme, however, temperate-zone species encounter not only hot conditions during the active season, but also cool conditions. Even in hot weather, night-time temperatures often will be too low for reptiles to reach high Tbs. Thus, high-latitude environments are thermally challenging for ectotherms (Blouin-Demers & Weatherhead, 2001). 'Effective' thermoregulation (i.e., Tbs close to preferred levels; Hertz et al., 1993) may be uncommon in such environments, not because it is unimportant, but because it is difficult and potentially costly (Huey & Slatkin, 1976).
39 The European Grass Snake, Natrix natrix, is a medium-sized, oviparous, diurnal natricine snake that ranges from extreme northern Africa to near the Arctic Circle in Scandinavia (Beebe & Griffiths, 2000). Here, I present the results of a study of the thermal ecology of the Grass Snake at a high-latitude site in England. I used a semi-natural enclosure, radio-telemetry, and a novel regression analysis to address two main questions: 1. How often are environmental conditions suitable for thermoregulation? I predicted that such conditions often would be limited to mid-day, even in warm weather. 2. How well can snakes thermoregulate when conditions allow it? I predicted that, at low environmental temperatures, snakes should be thermoconformers or, at best, weak thermoregulators. Above some threshold temperature, however, snakes should be able to maintain Tbs that are independent of environmental temperatures. An ancillary question that I addressed is whether gravid female snakes maintain different or less variable Tbs than non-gravid females. Other studies have shown this to be the case in viviparous snakes (Charland & Gregory, 1990; Charland, 1995; Brown & Weatherhead, 2000) and in at least one oviparous species (Blouin- Demers & Weatherhead, 2001). The most likely explanation for these observations is that, via careful thermoregulation, gravid females can maintain the best possible temperature for their developing offspring; thus, it should be an especially important behaviour for gravid females in cool temperate-zone sites. Oviparous snakes are still relatively unstudied in this respect and the Grass Snake, with its high-latitude distribution, is a particularly suitable study species.
40 METHODS I conducted this study during June-July 2002 in an outdoor semi-natural field enclosure at the University of Kent at Canterbury (UKC) in Canterbury, Kent, UK. The enclosure was located 3.25 km NE of my field site on the River Stour (58"N 17.58'N, 1 ON 08.19'E, 5 m elevation) and thus experienced similar weather. I caught Grass Snakes opportunistically by hand and by flipping artificial cover objects. I retained gravid and nongravid females in a laboratory at UKC and chose three of each for the enclosure experiment (reduced to five snakes later- see Results). Snakes were maintained in the laboratory in individual plastic cages (45 cm X 25 cm X 25 cm) lined with newsprint. Water was provided ad libidurn and snakes were each fed 5 goldfish approximately twice a week. I chose the largest healthy snakes in each reproductive group for implantation of 1.5 g radio-transmitters (maximum ratio of transmitter mass: body mass = 0.05: 1 ; Model BD-2GT, Holohil Systems, Carp, Ontario, Canada; 12-week battery life at 25 "C). All animal maintenance and experimental procedures complied with guidelines for live reptiles and were approved by the University of Victoria Animal Care Committee. The pulse rate of each radiotransmitter was proportional to temperature, and calibration curves were supplied by the manufacturer (pulse rate range: 0-40 "C in 10 "C increments). I used second-order polynomial (quadratic) regressions of the 5 calibration points for each transmitter to derive an equation to predict temperature based on pulse rate. All calibration equations provided a high degree of fit (r in all cases). Surgery and implantation (and later removal) of transmitters were performed by a veterinary surgeon, using a modified version of Reinert and Cundall's (1982) method. Isofluorane was used to anaesthetize the snakes. Transmitters and their antennae were implanted subcutaneously with sterile techniques and 3-0 absorbable
41 sutures were used to close the 2 cm incision. All snakes recovered rapidly; tongueflicking and movement resumed within 2 hours of surgery. I kept the snakes in the laboratory for 5 days before releasing them in the enclosure and did not begin monitoring Tb until 48 hours after their release. I constructed an 8 m X 8 m enclosure at UKC in late May 2002, roughly following the design of Lee and Mills (2000). I placed the enclosure in an open area fully exposed to the sun, so that snakes would be able to achieve high Tbs in warm, sunny weather. Snakes could retreat to underground tunnels, rock crevices, and artificial cover objects for protection or cooling (Fig. 1). I also provided numerous basking sites (e.g., compost pile and bramble pile), as well as spots where snakes could raise their Tb without being exposed (e.g., under metal sheet). The vegetation was allowed to grow unhindered to provide additional cover, which I have observed snakes retreating to in the field. I created aquatic and semi-aquatic habitats by immersing vegetation into the pond and laying it on the pond edge. The 64 m2 enclosure was a compromise between making construction manageable and providing sufficient room for snakes to behave naturally. I have observed Grass Snakes in close proximity to each other in the field, so I did not consider the six snakes to be too crowded, and I saw no obviously abnormal behaviour of snakes during the experiment. The enclosure simulated natural conditions as closely as possible by including a variety of microhabitats (e.g., burrows, ponds, retreat sites, basking sites) and thermal conditions (e.g., full sun, full shade, surdshade mosaic). An artificial pond provided constant access to water and was also stocked with goldfish for food. Grass Snakes eat fish in the field (Gregory & Isaac, in press) and the laboratory (pers. obs.) and it was a convenient way to provide food.
 Photograph of the outdoor enclosure at the")
 Aerial schematic of the various microhabitats")
42 Metal Sheet / - Full Sun Pond / Fig. 1. a) Photograph of the outdoor enclosure at the University of Kent at Canterbury in Canterbury, UK. b) Aerial schematic of the various microhabitats within the 64 m2 enclosure.
THE concept that reptiles have preferred
 Copeia, 2000(3), pp. 841 845 Plasticity in Preferred Body Temperature of Young Snakes in Response to Temperature during Development GABRIEL BLOUIN-DEMERS, KELLEY J. KISSNER, AND PATRICK J. WEATHERHEAD
Copeia, 2000(3), pp. 841 845 Plasticity in Preferred Body Temperature of Young Snakes in Response to Temperature during Development GABRIEL BLOUIN-DEMERS, KELLEY J. KISSNER, AND PATRICK J. WEATHERHEAD
Bio4009 : Projet de recherche/research project
 Bio4009 : Projet de recherche/research project Is emergence after hibernation of the black ratsnake (Elaphe obsoleta) triggered by a thermal gradient reversal? By Isabelle Ceillier 4522350 Supervisor :
Bio4009 : Projet de recherche/research project Is emergence after hibernation of the black ratsnake (Elaphe obsoleta) triggered by a thermal gradient reversal? By Isabelle Ceillier 4522350 Supervisor :
A test of the thermal coadaptation hypothesis in the common map turtle (Graptemys geographica) Elad Ben-Ezra. Supervisor: Dr. Gabriel Blouin-Demers
 Elad Ben-Ezra. Supervisor: Dr. Gabriel Blouin-Demers") A test of the thermal coadaptation hypothesis in the common map turtle (Graptemys geographica) by Elad Ben-Ezra Supervisor: Dr. Gabriel Blouin-Demers Thesis submitted to the Department of Biology in partial
A test of the thermal coadaptation hypothesis in the common map turtle (Graptemys geographica) by Elad Ben-Ezra Supervisor: Dr. Gabriel Blouin-Demers Thesis submitted to the Department of Biology in partial
Thermal quality influences effectiveness of thermoregulation, habitat use, and behaviour in milk snakes
 Oecologia (2006) 148: 1 11 DOI 10.1007/s00442-005-0350-7 ECOPHYSIOLOGY Jeffrey R. Row Æ Gabriel Blouin-Demers Thermal quality influences effectiveness of thermoregulation, habitat use, and behaviour in
Oecologia (2006) 148: 1 11 DOI 10.1007/s00442-005-0350-7 ECOPHYSIOLOGY Jeffrey R. Row Æ Gabriel Blouin-Demers Thermal quality influences effectiveness of thermoregulation, habitat use, and behaviour in
BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS
 Nov., 1965 505 BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS Lack ( 1954; 40-41) has pointed out that in species of birds which have asynchronous hatching, brood size may be adjusted
Nov., 1965 505 BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS Lack ( 1954; 40-41) has pointed out that in species of birds which have asynchronous hatching, brood size may be adjusted
Conflicts between Courtship and Thermoregulation: The Thermal Ecology of Amorous Male Garter Snakes (Thamnophis sirtalis parietalis, Colubridae)
") 508 Conflicts between Courtship and Thermoregulation: The Thermal Ecology of Amorous Male Garter Snakes (Thamnophis sirtalis parietalis, Colubridae) R. Shine 1,* P. S. Harlow 1 M. J. Elphick 1 M. M. Olsson
508 Conflicts between Courtship and Thermoregulation: The Thermal Ecology of Amorous Male Garter Snakes (Thamnophis sirtalis parietalis, Colubridae) R. Shine 1,* P. S. Harlow 1 M. J. Elphick 1 M. M. Olsson
MATERNAL NEST-SITE CHOICE AND OFFSPRING FITNESS IN A TROPICAL SNAKE (TROPIDONOPHIS MAIRII, COLUBRIDAE)
") Ecology, 85(6), 2004, pp. 1627 1634 2004 by the Ecological Society of America MATERNAL NEST-SITE CHOICE AND OFFSPRING FITNESS IN A TROPICAL SNAKE (TROPIDONOPHIS MAIRII, COLUBRIDAE) G. P. BROWN AND R. SHINE
Ecology, 85(6), 2004, pp. 1627 1634 2004 by the Ecological Society of America MATERNAL NEST-SITE CHOICE AND OFFSPRING FITNESS IN A TROPICAL SNAKE (TROPIDONOPHIS MAIRII, COLUBRIDAE) G. P. BROWN AND R. SHINE
Reptilian Physiology
 Reptilian Physiology Physiology, part deux The study of chemical and physical processes in the organism Aspects of the physiology can be informative for understanding organisms in their environment Thermoregulation
Reptilian Physiology Physiology, part deux The study of chemical and physical processes in the organism Aspects of the physiology can be informative for understanding organisms in their environment Thermoregulation
Seasonal Shifts in Reproductive Investment of Female Northern Grass Lizards ( Takydromus septentrionalis
 Seasonal Shifts in Reproductive Investment of Female Northern Grass Lizards (Takydromus septentrionalis) from a Field Population on Beiji Island, China Author(s): Wei-Guo Du and Lu Shou Source: Journal
Seasonal Shifts in Reproductive Investment of Female Northern Grass Lizards (Takydromus septentrionalis) from a Field Population on Beiji Island, China Author(s): Wei-Guo Du and Lu Shou Source: Journal
Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve,
 at Sungei Buloh Wetland Reserve,") Author Title Institute Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve, Singapore Thesis (Ph.D.) National
Author Title Institute Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve, Singapore Thesis (Ph.D.) National
Short-term Water Potential Fluctuations and Eggs of the Red-eared Slider Turtle (Trachemys scripta elegans)
") Zoology and Genetics Publications Zoology and Genetics 2001 Short-term Water Potential Fluctuations and Eggs of the Red-eared Slider Turtle (Trachemys scripta elegans) John K. Tucker Illinois Natural History
Zoology and Genetics Publications Zoology and Genetics 2001 Short-term Water Potential Fluctuations and Eggs of the Red-eared Slider Turtle (Trachemys scripta elegans) John K. Tucker Illinois Natural History
Northern Copperhead Updated: April 8, 2018
 Interpretation Guide Northern Copperhead Updated: April 8, 2018 Status Danger Threats Population Distribution Habitat Diet Size Longevity Social Family Units Reproduction Our Animals Scientific Name Least
Interpretation Guide Northern Copperhead Updated: April 8, 2018 Status Danger Threats Population Distribution Habitat Diet Size Longevity Social Family Units Reproduction Our Animals Scientific Name Least
Effects of nest temperature and moisture on phenotypic traits of hatchling snakes (Tropidonophis mairii, Colubridae) from tropical Australia
 from tropical Australia") Blackwell Publishing LtdOxford, UKBIJBiological Journal of the Linnean Society24-466The Linnean Society of London, 26? 26 891 159168 Original Article INCUBATION EFFECTS IN A SNAKE G. P. BROWN and R. SHINE
Blackwell Publishing LtdOxford, UKBIJBiological Journal of the Linnean Society24-466The Linnean Society of London, 26? 26 891 159168 Original Article INCUBATION EFFECTS IN A SNAKE G. P. BROWN and R. SHINE
University of Canberra. This thesis is available in print format from the University of Canberra Library.
 University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
*Using the 2018 List. Use the image below to answer question 6.
 Herpetology Test 1. Hearts in all herps other than consists of atria and one ventricle somewhat divided by a septum. (2 pts) a. snakes; two b. crocodiles; two c. turtles; three d. frogs; four 2. The food
Herpetology Test 1. Hearts in all herps other than consists of atria and one ventricle somewhat divided by a septum. (2 pts) a. snakes; two b. crocodiles; two c. turtles; three d. frogs; four 2. The food
NOTES ON THE ECOLOGY AND NATURAL HISTORY OF CTENOPHORUS CAUDICINCTUS (AGAMIDAE) IN WESTERN AUSTRALIA
 IN WESTERN AUSTRALIA") NOTES ON THE ECOLOGY AND NATURAL HISTORY OF CTENOPHORUS CAUDICINCTUS (AGAMIDAE) IN WESTERN AUSTRALIA By ERIC R. PIANKA Integrative Biology University of Texas at Austin Austin, Texas 78712 USA Email: erp@austin.utexas.edu
NOTES ON THE ECOLOGY AND NATURAL HISTORY OF CTENOPHORUS CAUDICINCTUS (AGAMIDAE) IN WESTERN AUSTRALIA By ERIC R. PIANKA Integrative Biology University of Texas at Austin Austin, Texas 78712 USA Email: erp@austin.utexas.edu
CHOOSING YOUR REPTILE LIGHTING AND HEATING
 CHOOSING YOUR REPTILE LIGHTING AND HEATING What lights do I need for my pet Bearded Dragon, Python, Gecko or other reptile, turtle or frog? Is specialised lighting and heating required for indoor reptile
CHOOSING YOUR REPTILE LIGHTING AND HEATING What lights do I need for my pet Bearded Dragon, Python, Gecko or other reptile, turtle or frog? Is specialised lighting and heating required for indoor reptile
Objectives: Outline: Idaho Amphibians and Reptiles. Characteristics of Amphibians. Types and Numbers of Amphibians
 Natural History of Idaho Amphibians and Reptiles Wildlife Ecology, University of Idaho Fall 2005 Charles R. Peterson Herpetology Laboratory Department of Biological Sciences, Idaho Museum of Natural History
Natural History of Idaho Amphibians and Reptiles Wildlife Ecology, University of Idaho Fall 2005 Charles R. Peterson Herpetology Laboratory Department of Biological Sciences, Idaho Museum of Natural History
Field Herpetology Final Guide
 Field Herpetology Final Guide Questions with more complexity will be worth more points Incorrect spelling is OK as long as the name is recognizable ( by the instructor s discretion ) Common names will
Field Herpetology Final Guide Questions with more complexity will be worth more points Incorrect spelling is OK as long as the name is recognizable ( by the instructor s discretion ) Common names will
Sec KEY CONCEPT Reptiles, birds, and mammals are amniotes.
 Thu 4/27 Learning Target Class Activities *attached below (scroll down)* Website: my.hrw.com Username: bio678 Password:a4s5s Activities Students will describe the evolutionary significance of amniotic
Thu 4/27 Learning Target Class Activities *attached below (scroll down)* Website: my.hrw.com Username: bio678 Password:a4s5s Activities Students will describe the evolutionary significance of amniotic
NOTES ON THE ECOLOGY AND NATURAL HISTORY OF TWO SPECIES OF EGERNIA (SCINCIDAE) IN WESTERN AUSTRALIA
 IN WESTERN AUSTRALIA") NOTES ON THE ECOLOGY AND NATURAL HISTORY OF TWO SPECIES OF EGERNIA (SCINCIDAE) IN WESTERN AUSTRALIA By ERIC R. PIANKA Integrative Biology University of Texas at Austin Austin, Texas 78712 USA Email: erp@austin.utexas.edu
NOTES ON THE ECOLOGY AND NATURAL HISTORY OF TWO SPECIES OF EGERNIA (SCINCIDAE) IN WESTERN AUSTRALIA By ERIC R. PIANKA Integrative Biology University of Texas at Austin Austin, Texas 78712 USA Email: erp@austin.utexas.edu
reproductive life History and the effects of sex and season on morphology in CRoTALus oreganus (northern PaCifiC RATTLESNAKES)
") reproductive life History and the effects of sex and season on morphology in CRoTALus oreganus (northern PaCifiC RATTLESNAKES) Benjamin Kwittken, Student Author dr. emily n. taylor, research advisor abstract
reproductive life History and the effects of sex and season on morphology in CRoTALus oreganus (northern PaCifiC RATTLESNAKES) Benjamin Kwittken, Student Author dr. emily n. taylor, research advisor abstract
Station 1 1. (3 points) Identification: Station 2 6. (3 points) Identification:
 Identification: Station 2 6. (3 points) Identification:") SOnerd s 2018-2019 Herpetology SSSS Test 1 SOnerd s SSSS 2018-2019 Herpetology Test Station 20 sounds found here: https://drive.google.com/drive/folders/1oqrmspti13qv_ytllk_yy_vrie42isqe?usp=sharing Station
SOnerd s 2018-2019 Herpetology SSSS Test 1 SOnerd s SSSS 2018-2019 Herpetology Test Station 20 sounds found here: https://drive.google.com/drive/folders/1oqrmspti13qv_ytllk_yy_vrie42isqe?usp=sharing Station
Care For Us Re#culated Python (Python re/culatus)
") Care For Us Re#culated Python (Python re/culatus) Animal Welfare Animal welfare refers to an animal s state or feelings. An animal s welfare state can be positive, neutral or negative. An animal s welfare
Care For Us Re#culated Python (Python re/culatus) Animal Welfare Animal welfare refers to an animal s state or feelings. An animal s welfare state can be positive, neutral or negative. An animal s welfare
Nest-site selection in Eastern hognose snakes (Heterodon platirhinos) Casey Peet-Paré
 Casey Peet-Paré") Nest-site selection in Eastern hognose snakes (Heterodon platirhinos) by Casey Peet-Paré Thesis submitted to the Department of Biology in partial fulfillment of the requirements for the B.Sc. Honours degree,
Nest-site selection in Eastern hognose snakes (Heterodon platirhinos) by Casey Peet-Paré Thesis submitted to the Department of Biology in partial fulfillment of the requirements for the B.Sc. Honours degree,
Biology. Slide 1of 50. End Show. Copyright Pearson Prentice Hall
 Biology 1of 50 2of 50 Phylogeny of Chordates Nonvertebrate chordates Jawless fishes Sharks & their relatives Bony fishes Reptiles Amphibians Birds Mammals Invertebrate ancestor 3of 50 A vertebrate dry,
Biology 1of 50 2of 50 Phylogeny of Chordates Nonvertebrate chordates Jawless fishes Sharks & their relatives Bony fishes Reptiles Amphibians Birds Mammals Invertebrate ancestor 3of 50 A vertebrate dry,
BODY size and temperature affect nearly every
 Copeia, 2004(1), pp. 145 151 Effects of Body Mass and Temperature on Standard Metabolic Rate in the Eastern Diamondback Rattlesnake (Crotalus adamanteus) MICHAEL E. DORCAS, WILLIAM A. HOPKINS, AND JOHN
Copeia, 2004(1), pp. 145 151 Effects of Body Mass and Temperature on Standard Metabolic Rate in the Eastern Diamondback Rattlesnake (Crotalus adamanteus) MICHAEL E. DORCAS, WILLIAM A. HOPKINS, AND JOHN
Rubber Boas in Radium Hot Springs: Habitat, Inventory, and Management Strategies
 : Habitat, Inventory, and Management Strategies ROBERT C. ST. CLAIR 1 AND ALAN DIBB 2 1 9809 92 Avenue, Edmonton, AB, T6E 2V4, Canada, email rstclair@telusplanet.net 2 Parks Canada, Box 220, Radium Hot
: Habitat, Inventory, and Management Strategies ROBERT C. ST. CLAIR 1 AND ALAN DIBB 2 1 9809 92 Avenue, Edmonton, AB, T6E 2V4, Canada, email rstclair@telusplanet.net 2 Parks Canada, Box 220, Radium Hot
Intraspecific relationships extra questions and answers (Extension material for Level 3 Biology Study Guide, ISBN , page 153)
") i Intraspecific relationships extra questions and answers (Extension material for Level 3 Biology Study Guide, ISBN 978-1-927194-58-4, page 153) Activity 9: Intraspecific relationships extra questions
i Intraspecific relationships extra questions and answers (Extension material for Level 3 Biology Study Guide, ISBN 978-1-927194-58-4, page 153) Activity 9: Intraspecific relationships extra questions
Notes on Road-Killed Snakes and Their Implications on Habitat Modification Due to Summer Flooding on the Mississippi River in West Central Illinois
 Transactions of the Illinois State Academy of Science (1995), Volume 88, 1 and 2, pp. 61-71 Notes on Road-Killed Snakes and Their Implications on Habitat Modification Due to Summer Flooding on the Mississippi
Transactions of the Illinois State Academy of Science (1995), Volume 88, 1 and 2, pp. 61-71 Notes on Road-Killed Snakes and Their Implications on Habitat Modification Due to Summer Flooding on the Mississippi
Incubation temperature and phenotypic traits of Sceloporus undulatus: implications for the northern limits of distribution
 DOI 10.1007/s00442-006-0583-0 ECOPHYSIOLOGY Incubation temperature and phenotypic traits of Sceloporus undulatus: implications for the northern limits of distribution Scott L. Parker Æ Robin M. Andrews
DOI 10.1007/s00442-006-0583-0 ECOPHYSIOLOGY Incubation temperature and phenotypic traits of Sceloporus undulatus: implications for the northern limits of distribution Scott L. Parker Æ Robin M. Andrews
Reproductive physiology and eggs
 Reproductive physiology and eggs Class Business Reading for this lecture Required. Gill: Chapter 14 1. Reproductive physiology In lecture I will only have time to go over reproductive physiology briefly,
Reproductive physiology and eggs Class Business Reading for this lecture Required. Gill: Chapter 14 1. Reproductive physiology In lecture I will only have time to go over reproductive physiology briefly,
Writing: Lesson 23. Today the students will practice planning for informative/explanatory prompts in response to text they read.
 Top Score Writing Grade 4 Lesson 23 Writing: Lesson 23 Today the students will practice planning for informative/explanatory prompts in response to text they read. The following passages will be used in
Top Score Writing Grade 4 Lesson 23 Writing: Lesson 23 Today the students will practice planning for informative/explanatory prompts in response to text they read. The following passages will be used in
Like mother, like daughter: inheritance of nest-site
 Like mother, like daughter: inheritance of nest-site location in snakes Gregory P. Brown and Richard Shine* School of Biological Sciences A0, University of Sydney, NSW 00, Australia *Author for correspondence
Like mother, like daughter: inheritance of nest-site location in snakes Gregory P. Brown and Richard Shine* School of Biological Sciences A0, University of Sydney, NSW 00, Australia *Author for correspondence
4 Many species of mammals, birds, reptiles, amphibians and fish 940L. Source 1 Habitats
 Source 1 Habitats 1 American Alligators can be found in fresh water environments like rivers, lakes, ponds, swamps and marshes. They also like to live in areas that are brackish, which means the water
Source 1 Habitats 1 American Alligators can be found in fresh water environments like rivers, lakes, ponds, swamps and marshes. They also like to live in areas that are brackish, which means the water
JoJoKeKe s Herpetology Exam
 ~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~~*~*~*~*~*~*~*~*~*~*~*~*~*~*~ JoJoKeKe s Herpetology Exam (SSSS) 2:30 to be given at each station- B/C Station 1: 1.) What is the family & genus of the shown
~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~~*~*~*~*~*~*~*~*~*~*~*~*~*~*~ JoJoKeKe s Herpetology Exam (SSSS) 2:30 to be given at each station- B/C Station 1: 1.) What is the family & genus of the shown
B-Division Herpetology Test. By: Brooke Diamond
 B-Division Herpetology Test By: Brooke Diamond Rules: - Play each slide for 2 minutes and answer the questions on the test sheet. - Use only pages attached to your binder, you may not use stray pages.
B-Division Herpetology Test By: Brooke Diamond Rules: - Play each slide for 2 minutes and answer the questions on the test sheet. - Use only pages attached to your binder, you may not use stray pages.
Habitats and Field Methods. Friday May 12th 2017
 Habitats and Field Methods Friday May 12th 2017 Announcements Project consultations available today after class Project Proposal due today at 5pm Follow guidelines posted for lecture 4 Field notebooks
Habitats and Field Methods Friday May 12th 2017 Announcements Project consultations available today after class Project Proposal due today at 5pm Follow guidelines posted for lecture 4 Field notebooks
Weaver Dunes, Minnesota
 Hatchling Orientation During Dispersal from Nests Experimental analyses of an early life stage comparing orientation and dispersal patterns of hatchlings that emerge from nests close to and far from wetlands
Hatchling Orientation During Dispersal from Nests Experimental analyses of an early life stage comparing orientation and dispersal patterns of hatchlings that emerge from nests close to and far from wetlands
Investigating Fish Respiration
 CHAPTER 31 Fishes and Amphibians Section 31-1 SKILL ACTIVITY Interpreting graphs Investigating Fish Respiration It is well known that a fish dies from lack of oxygen when taken out of water. However, water
CHAPTER 31 Fishes and Amphibians Section 31-1 SKILL ACTIVITY Interpreting graphs Investigating Fish Respiration It is well known that a fish dies from lack of oxygen when taken out of water. However, water
FEMALE PHENOTYPE, LIFE HISTORY, AND REPRODUCTIVE SUCCESS IN FREE-RANGING SNAKES (TROPIDONOPHIS MAIRII)
") Ecology, 86(10), 2005, pp. 2763 2770 2005 by the Ecological Society of America FEMALE PHENOTYPE, LIFE HISTORY, AND REPRODUCTIVE SUCCESS IN FREE-RANGING SNAKES (TROPIDONOPHIS MAIRII) G. P. BROWN AND R.
Ecology, 86(10), 2005, pp. 2763 2770 2005 by the Ecological Society of America FEMALE PHENOTYPE, LIFE HISTORY, AND REPRODUCTIVE SUCCESS IN FREE-RANGING SNAKES (TROPIDONOPHIS MAIRII) G. P. BROWN AND R.
Who Cares? The Evolution of Parental Care in Squamate Reptiles. Ben Halliwell Geoffrey While, Tobias Uller
 Who Cares? The Evolution of Parental Care in Squamate Reptiles Ben Halliwell Geoffrey While, Tobias Uller 1 Parental Care any instance of parental investment that increases the fitness of offspring 2 Parental
Who Cares? The Evolution of Parental Care in Squamate Reptiles Ben Halliwell Geoffrey While, Tobias Uller 1 Parental Care any instance of parental investment that increases the fitness of offspring 2 Parental
Writing: Lesson 31. Today the students will be learning how to write more advanced middle paragraphs using a variety of elaborative techniques.
 Top Score Writing Grade 4 Lesson 31 Writing: Lesson 31 Today the students will be learning how to write more advanced middle paragraphs using a variety of elaborative techniques. The following passages
Top Score Writing Grade 4 Lesson 31 Writing: Lesson 31 Today the students will be learning how to write more advanced middle paragraphs using a variety of elaborative techniques. The following passages
Social and Thermal Cues Influence Nest-site Selection in a Nocturnal Gecko, Oedura lesueurii
 RESEARCH PAPER Social and Thermal Cues Influence Nest-site Selection in a Nocturnal Gecko, Oedura lesueurii David A. Pike*, Jonathan K. Webb* & Robin M. Andrews * School of Biological Sciences A08, University
RESEARCH PAPER Social and Thermal Cues Influence Nest-site Selection in a Nocturnal Gecko, Oedura lesueurii David A. Pike*, Jonathan K. Webb* & Robin M. Andrews * School of Biological Sciences A08, University
Corn Snake Care Sheet
 Corn Snake Care Sheet Temperament With the odd exception, Corn Snakes are calm, docile, placid snakes that are hardy and thrive very well in captivity. Due to their temperament Corn Snakes are a recommended
Corn Snake Care Sheet Temperament With the odd exception, Corn Snakes are calm, docile, placid snakes that are hardy and thrive very well in captivity. Due to their temperament Corn Snakes are a recommended
Thermal adaptation of maternal and embryonic phenotypes in a geographically widespread ectotherm
 International Congress Series 1275 (2004) 258 266 www.ics-elsevier.com Thermal adaptation of maternal and embryonic phenotypes in a geographically widespread ectotherm Michael J. Angilletta Jr. a, *, Christopher
International Congress Series 1275 (2004) 258 266 www.ics-elsevier.com Thermal adaptation of maternal and embryonic phenotypes in a geographically widespread ectotherm Michael J. Angilletta Jr. a, *, Christopher
WHAT ARE HERPTILES? WHICH IS WHICH? 1. Vertebrates are animals that have 2. Complete the following chart of vertebrate groups: EGGS LAID WHERE?
 WHAT ARE HERPTILES? 1. Vertebrates are animals that have 2. Complete the following chart of vertebrate groups: SKIN COVERING? GILLS OR LUNGS? EGGS LAID WHERE? ENDOTHERMIC OR ECTOTHERMIC Fish AMPHIBIANS
WHAT ARE HERPTILES? 1. Vertebrates are animals that have 2. Complete the following chart of vertebrate groups: SKIN COVERING? GILLS OR LUNGS? EGGS LAID WHERE? ENDOTHERMIC OR ECTOTHERMIC Fish AMPHIBIANS
Biology Slide 1 of 50
 Biology 1 of 50 2 of 50 What Is a Reptile? What are the characteristics of reptiles? 3 of 50 What Is a Reptile? What Is a Reptile? A reptile is a vertebrate that has dry, scaly skin, lungs, and terrestrial
Biology 1 of 50 2 of 50 What Is a Reptile? What are the characteristics of reptiles? 3 of 50 What Is a Reptile? What Is a Reptile? A reptile is a vertebrate that has dry, scaly skin, lungs, and terrestrial
Silence of the Frogs Lexile 1040L
 daptation Silence of the Frogs Lexile 1040L 1 mphibians require specific habitats. They need a moist environment to be active and standing water to breed in. They need food for both tadpoles and adults.
daptation Silence of the Frogs Lexile 1040L 1 mphibians require specific habitats. They need a moist environment to be active and standing water to breed in. They need food for both tadpoles and adults.
Maturity and Other Reproductive Traits of the Kanahebi Lizard Takydromus tachydromoides (Sauria, Lacertidae) in Mito
 in Mito") Japanese Journal of Herpetology 9 (2): 46-53. 1981. Maturity and Other Reproductive Traits of the Kanahebi Lizard Takydromus tachydromoides (Sauria, Lacertidae) in Mito Sen TAKENAKA SUMMARY: Reproduction
Japanese Journal of Herpetology 9 (2): 46-53. 1981. Maturity and Other Reproductive Traits of the Kanahebi Lizard Takydromus tachydromoides (Sauria, Lacertidae) in Mito Sen TAKENAKA SUMMARY: Reproduction
Reptiles and amphibian behaviour
 Reptiles and amphibian behaviour Understanding how a healthy reptile and amphibian should look and act takes a lot of observation and practice. Reptiles and amphibians have behaviour that relates to them
Reptiles and amphibian behaviour Understanding how a healthy reptile and amphibian should look and act takes a lot of observation and practice. Reptiles and amphibians have behaviour that relates to them
This is the book of David, the son of Abraham and the father of Jesus Christ Matthew 1.1. Declaration
 ! ii This is the book of David, the son of Abraham and the father of Jesus Christ Matthew 1.1 Declaration This thesis is my original work and no part has been previously submitted for a degree. Chapters
! ii This is the book of David, the son of Abraham and the father of Jesus Christ Matthew 1.1 Declaration This thesis is my original work and no part has been previously submitted for a degree. Chapters
Consequences of Extended Egg Retention in the Eastern Fence Lizard (Sceloporus undulatus)
") Journal of Herpetology, Vol. 37, No. 2, pp. 309 314, 2003 Copyright 2003 Society for the Study of Amphibians and Reptiles Consequences of Extended Egg Retention in the Eastern Fence Lizard (Sceloporus
Journal of Herpetology, Vol. 37, No. 2, pp. 309 314, 2003 Copyright 2003 Society for the Study of Amphibians and Reptiles Consequences of Extended Egg Retention in the Eastern Fence Lizard (Sceloporus
Maternal Effects in the Green Turtle (Chelonia mydas)
") Maternal Effects in the Green Turtle (Chelonia mydas) SUBMITTED BY SAM B. WEBER TO THE UNIVERSITY OF EXETER AS A THESIS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY IN BIOLOGY; 8 TH JUNE 2010 This thesis is
Maternal Effects in the Green Turtle (Chelonia mydas) SUBMITTED BY SAM B. WEBER TO THE UNIVERSITY OF EXETER AS A THESIS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY IN BIOLOGY; 8 TH JUNE 2010 This thesis is
HUMAN APPENDIX BATS & TROPICAL FLOWERS
 HUMAN APPENDIX In humans, the appendix is a short piece of tissue off the large intestine. It is not used by humans for digestive functions. In other mammals, like rabbits and deer, the cecum is a large
HUMAN APPENDIX In humans, the appendix is a short piece of tissue off the large intestine. It is not used by humans for digestive functions. In other mammals, like rabbits and deer, the cecum is a large
Animal Adaptations. Structure and Function
 Name period date assigned date due date returned 1. What is a variation 2. What is an adaptation omplete the chart with the examples from the power point. List adaptations that help animals do the following:
Name period date assigned date due date returned 1. What is a variation 2. What is an adaptation omplete the chart with the examples from the power point. List adaptations that help animals do the following:
Influence of meal size on postprandial thermophily in cornsnakes (Elaphe guttata)
") TRANSACTIONS OF THE KANSAS ACADEMY OF SCIENCE Vol. 109, no. 3/4 p. 184-190 (2006) Influence of meal size on postprandial thermophily in cornsnakes (Elaphe guttata) LYNETT R. BONTRAGER, DAPHNE M. JONES,
TRANSACTIONS OF THE KANSAS ACADEMY OF SCIENCE Vol. 109, no. 3/4 p. 184-190 (2006) Influence of meal size on postprandial thermophily in cornsnakes (Elaphe guttata) LYNETT R. BONTRAGER, DAPHNE M. JONES,
Natural history of Xenosaurus phalaroanthereon (Squamata, Xenosauridae), a Knob-scaled Lizard from Oaxaca, Mexico
, a Knob-scaled Lizard from Oaxaca, Mexico") Natural history of Xenosaurus phalaroanthereon (Squamata, Xenosauridae), a Knob-scaled Lizard from Oaxaca, Mexico Julio A. Lemos-Espinal 1 and Geoffrey R. Smith Phyllomedusa 4():133-137, 005 005 Departamento
Natural history of Xenosaurus phalaroanthereon (Squamata, Xenosauridae), a Knob-scaled Lizard from Oaxaca, Mexico Julio A. Lemos-Espinal 1 and Geoffrey R. Smith Phyllomedusa 4():133-137, 005 005 Departamento
Squamates of Connecticut
 Squamates of Connecticut Reptilia Turtles are sisters to crocodiles and birds Yeah, birds are reptiles, haven t you watched Jurassic Park yet? Lizards and snakes are part of one clade called the squamates
Squamates of Connecticut Reptilia Turtles are sisters to crocodiles and birds Yeah, birds are reptiles, haven t you watched Jurassic Park yet? Lizards and snakes are part of one clade called the squamates
DOES VIVIPARITY EVOLVE IN COLD CLIMATE REPTILES BECAUSE PREGNANT FEMALES MAINTAIN STABLE (NOT HIGH) BODY TEMPERATURES?
 BODY TEMPERATURES?") Evolution, 58(8), 2004, pp. 1809 1818 DOES VIVIPARITY EVOLVE IN COLD CLIMATE REPTILES BECAUSE PREGNANT FEMALES MAINTAIN STABLE (NOT HIGH) BODY TEMPERATURES? RICHARD SHINE School of Biological Sciences,
Evolution, 58(8), 2004, pp. 1809 1818 DOES VIVIPARITY EVOLVE IN COLD CLIMATE REPTILES BECAUSE PREGNANT FEMALES MAINTAIN STABLE (NOT HIGH) BODY TEMPERATURES? RICHARD SHINE School of Biological Sciences,
Class Reptilia Testudines Squamata Crocodilia Sphenodontia
 Class Reptilia Testudines (around 300 species Tortoises and Turtles) Squamata (around 7,900 species Snakes, Lizards and amphisbaenids) Crocodilia (around 23 species Alligators, Crocodiles, Caimans and
Class Reptilia Testudines (around 300 species Tortoises and Turtles) Squamata (around 7,900 species Snakes, Lizards and amphisbaenids) Crocodilia (around 23 species Alligators, Crocodiles, Caimans and
Vertebrates. Vertebrate Characteristics. 444 Chapter 14
 4 Vertebrates Key Concept All vertebrates have a backbone, which supports other specialized body structures and functions. What You Will Learn Vertebrates have an endoskeleton that provides support and
4 Vertebrates Key Concept All vertebrates have a backbone, which supports other specialized body structures and functions. What You Will Learn Vertebrates have an endoskeleton that provides support and
Conservation (last three 3 lecture periods, mostly as a led discussion). We can't cover everything, but that should serve as a rough outline.
. We can't cover everything, but that should serve as a rough outline.") Comments on the rest of the semester: Subjects to be discussed: Temperature relationships. Echolocation. Conservation (last three 3 lecture periods, mostly as a led discussion). Possibly (in order of importance):
Comments on the rest of the semester: Subjects to be discussed: Temperature relationships. Echolocation. Conservation (last three 3 lecture periods, mostly as a led discussion). Possibly (in order of importance):
Grade Level: 3-5. Next Generation Sunshine State Standards SC.3.L.15.1 SC.4.L.16.2; SC.4.L.17.4 SC.5.L.15.1; SC.5.L.17.1
 Grade Level: 3-5 Next Generation Sunshine State Standards SC.3.L.15.1 SC.4.L.16.2; SC.4.L.17.4 SC.5.L.15.1; SC.5.L.17.1 Program Overview Discover the realm of reptiles, amazing creatures adapted to land
Grade Level: 3-5 Next Generation Sunshine State Standards SC.3.L.15.1 SC.4.L.16.2; SC.4.L.17.4 SC.5.L.15.1; SC.5.L.17.1 Program Overview Discover the realm of reptiles, amazing creatures adapted to land
Parthenogenesis in Varanus ornatus, the Ornate Nile Monitor.
 Parthenogenesis in Varanus ornatus, the Ornate Nile Monitor. Parthenogenesis in varanids has been reported in two other species of monitor, the Komodo dragon, Varanus komodiensis (Watts et al) and the
Parthenogenesis in Varanus ornatus, the Ornate Nile Monitor. Parthenogenesis in varanids has been reported in two other species of monitor, the Komodo dragon, Varanus komodiensis (Watts et al) and the
Influence of Incubation Temperature on Morphology, Locomotor Performance, and Early Growth of Hatchling Wall Lizards (Podarcis muralis)
") JEZ 0774 422 F. BRAÑA JOURNAL AND OF X. JI EXPERIMENTAL ZOOLOGY 286:422 433 (2000) Influence of Incubation Temperature on Morphology, Locomotor Performance, and Early Growth of Hatchling Wall Lizards (Podarcis
JEZ 0774 422 F. BRAÑA JOURNAL AND OF X. JI EXPERIMENTAL ZOOLOGY 286:422 433 (2000) Influence of Incubation Temperature on Morphology, Locomotor Performance, and Early Growth of Hatchling Wall Lizards (Podarcis
Red-Tailed Hawk Buteo jamaicensis
 Red-Tailed Hawk Buteo jamaicensis This large, dark headed, broad-shouldered hawk is one of the most common and widespread hawks in North America. The Red-tailed hawk belongs to the genus (family) Buteo,
Red-Tailed Hawk Buteo jamaicensis This large, dark headed, broad-shouldered hawk is one of the most common and widespread hawks in North America. The Red-tailed hawk belongs to the genus (family) Buteo,
Lecture 9 - Avian Life Histories
 Lecture 9 - Avian Life Histories Chapters 12 16 Read the book many details Courtship and Mating Breeding systems Sex Nests and Incubation Parents and their Offspring Outline 1. Pair formation or other
Lecture 9 - Avian Life Histories Chapters 12 16 Read the book many details Courtship and Mating Breeding systems Sex Nests and Incubation Parents and their Offspring Outline 1. Pair formation or other
Impact of colour polymorphism and thermal conditions on thermoregulation, reproductive success, and development in Vipera aspis
 Impact of colour polymorphism and thermal conditions on thermoregulation, reproductive success, and development in Vipera aspis Sylvain Dubey, Johan Schürch, Joaquim Golay, Briséïs Castella, Laura Bonny,
Impact of colour polymorphism and thermal conditions on thermoregulation, reproductive success, and development in Vipera aspis Sylvain Dubey, Johan Schürch, Joaquim Golay, Briséïs Castella, Laura Bonny,
ACTIVITY #6: TODAY S PICNIC SPECIALS ARE
 TOPIC What types of food does the turtle eat? ACTIVITY #6: TODAY S PICNIC SPECIALS ARE BACKGROUND INFORMATION For further information, refer to Turtles of Ontario Fact Sheets (pages 10-26) and Unit Five:
TOPIC What types of food does the turtle eat? ACTIVITY #6: TODAY S PICNIC SPECIALS ARE BACKGROUND INFORMATION For further information, refer to Turtles of Ontario Fact Sheets (pages 10-26) and Unit Five:
Seasonality provokes a shift of thermal preferences in a temperate lizard, but altitude does not
 ARTICLE IN PRESS Journal of Thermal Biology 31 (2006) 237 242 www.elsevier.com/locate/jtherbio Seasonality provokes a shift of thermal preferences in a temperate lizard, but altitude does not Jose A. Dı
ARTICLE IN PRESS Journal of Thermal Biology 31 (2006) 237 242 www.elsevier.com/locate/jtherbio Seasonality provokes a shift of thermal preferences in a temperate lizard, but altitude does not Jose A. Dı
Australian Journal of Zoology
 CSIRO PUBLISHING Australian Journal of Zoology Volume 47, 1999 CSIRO Australia 1999 A journal for the publication of the results of original scientific research in all branches of zoology, except the taxonomy
CSIRO PUBLISHING Australian Journal of Zoology Volume 47, 1999 CSIRO Australia 1999 A journal for the publication of the results of original scientific research in all branches of zoology, except the taxonomy
A REPTILE SURVEY AT THE LAND AT HILL ROAD AND ELM TREE DRIVE, ROCHESTER, KENT,
 A REPTILE SURVEY AT THE LAND AT HILL ROAD AND ELM TREE DRIVE, ROCHESTER, KENT, Commissioned by: King & Johnston Homes Ltd and Medway Council Report Number: October 2013 Regent s Place, 3 rd Floor, 338
A REPTILE SURVEY AT THE LAND AT HILL ROAD AND ELM TREE DRIVE, ROCHESTER, KENT, Commissioned by: King & Johnston Homes Ltd and Medway Council Report Number: October 2013 Regent s Place, 3 rd Floor, 338
A Population Analysis of the Common Wall Lizard Podarcis muralis in Southwestern France
 - 513 - Studies in Herpetology, Rocek Z. (ed.) pp. 513-518 Prague 1986 A Population Analysis of the Common Wall Lizard Podarcis muralis in Southwestern France R. BARBAULT and Y. P. MOU Laboratoire d'ecologie
- 513 - Studies in Herpetology, Rocek Z. (ed.) pp. 513-518 Prague 1986 A Population Analysis of the Common Wall Lizard Podarcis muralis in Southwestern France R. BARBAULT and Y. P. MOU Laboratoire d'ecologie
Impact of colour polymorphism in free ranging asp vipers
 Impact of colour polymorphism in free ranging asp vipers Sylvain Dubey, Daniele Muri, Johan Schuerch, Naïke Trim, Joaquim Golay, Sylvain Ursenbacher, Philippe Golay, Konrad Mebert 08.10.15 2 Background
Impact of colour polymorphism in free ranging asp vipers Sylvain Dubey, Daniele Muri, Johan Schuerch, Naïke Trim, Joaquim Golay, Sylvain Ursenbacher, Philippe Golay, Konrad Mebert 08.10.15 2 Background
WATER plays an important role in all stages
 Copeia, 2002(1), pp. 220 226 Experimental Analysis of an Early Life-History Stage: Water Loss and Migrating Hatchling Turtles JASON J. KOLBE AND FREDRIC J. JANZEN The effect of water dynamics is well known
Copeia, 2002(1), pp. 220 226 Experimental Analysis of an Early Life-History Stage: Water Loss and Migrating Hatchling Turtles JASON J. KOLBE AND FREDRIC J. JANZEN The effect of water dynamics is well known
Motuora island reptile monitoring report for common & Pacific gecko 2017
 Motuora island reptile monitoring report for common & Pacific gecko 7 Prepared by Su Sinclair August 7 Work on this monitoring project was carried out under a Wildlife Act Authority issued by the Department
Motuora island reptile monitoring report for common & Pacific gecko 7 Prepared by Su Sinclair August 7 Work on this monitoring project was carried out under a Wildlife Act Authority issued by the Department
Reproductive strategies in snakes
 Received 10 October 2002 Accepted 4 December 2002 Published online 1 April 2003 Review Paper Reproductive strategies in snakes Richard Shine School of Biological Sciences A08, University of Sydney, Sydney,
Received 10 October 2002 Accepted 4 December 2002 Published online 1 April 2003 Review Paper Reproductive strategies in snakes Richard Shine School of Biological Sciences A08, University of Sydney, Sydney,
Is Parental Care the Key to Understanding Endothermy in Birds and Mammals?
 vol. 162, no. 6 the american naturalist december 2003 Is Parental Care the Key to Understanding Endothermy in Birds and Mammals? Michael J. Angilletta, Jr., * and Michael W. Sears Department of Life Sciences,
vol. 162, no. 6 the american naturalist december 2003 Is Parental Care the Key to Understanding Endothermy in Birds and Mammals? Michael J. Angilletta, Jr., * and Michael W. Sears Department of Life Sciences,
BUMPUS IN THE SNAKE DEN: EFFECTS OF SEX, SIZE, AND BODY CONDITION ON MORTALITY OF RED-SIDED GARTER SNAKES
 Evolution, 55(3), 2001, pp. 598 604 BUMPUS IN THE SNAKE DEN: EFFECTS OF SEX, SIZE, AND BODY CONDITION ON MORTALITY OF RED-SIDED GARTER SNAKES R. SHINE, 1,2 M. P. LEMASTER, 3 I. T. MOORE, 3 M. M. OLSSON,
Evolution, 55(3), 2001, pp. 598 604 BUMPUS IN THE SNAKE DEN: EFFECTS OF SEX, SIZE, AND BODY CONDITION ON MORTALITY OF RED-SIDED GARTER SNAKES R. SHINE, 1,2 M. P. LEMASTER, 3 I. T. MOORE, 3 M. M. OLSSON,
Sec KEY CONCEPT Amphibians evolved from lobe-finned fish.
 Wed 4/26 Activities Learning Target Class Activities *attached below (scroll down)* Website: my.hrw.com Username: bio678 Password:a4s5s Students will describe the adaptations of amphibians that help them
Wed 4/26 Activities Learning Target Class Activities *attached below (scroll down)* Website: my.hrw.com Username: bio678 Password:a4s5s Students will describe the adaptations of amphibians that help them
ABSTRACT THE IMPORTANCE OF PRE- AND POSTNATAL THERMAL CONDITIONS IN DETERMINING GROWTH TRAJECTORIES IN THREE VIVIPAROUS GRASSLAND SNAKES
 ABSTRACT THE IMPORTANCE OF PRE- AND POSTNATAL THERMAL CONDITIONS IN DETERMINING GROWTH TRAJECTORIES IN THREE VIVIPAROUS GRASSLAND SNAKES Tanya K. O Brien, M.S. Department of Biological Sciences Northern
ABSTRACT THE IMPORTANCE OF PRE- AND POSTNATAL THERMAL CONDITIONS IN DETERMINING GROWTH TRAJECTORIES IN THREE VIVIPAROUS GRASSLAND SNAKES Tanya K. O Brien, M.S. Department of Biological Sciences Northern
Australian Journal of Zoology
 Publishing Australian Journal of Zoology Volume 49, 2001 CSIRO 2001 A journal for the publication of the results of original scientific research in all branches of zoology, except the taxonomy of invertebrates
Publishing Australian Journal of Zoology Volume 49, 2001 CSIRO 2001 A journal for the publication of the results of original scientific research in all branches of zoology, except the taxonomy of invertebrates
WildlifeCampus Advanced Snakes & Reptiles 1. Vipers and Adders
 Advanced Snakes & Reptiles 1 Module # 4 Component # 9 Viperidae - Hinged Front Fang Snakes This Family is divided into two sub-families. These are Old World and Modern / New World Adders. The predominant
Advanced Snakes & Reptiles 1 Module # 4 Component # 9 Viperidae - Hinged Front Fang Snakes This Family is divided into two sub-families. These are Old World and Modern / New World Adders. The predominant
Breeding White Storks( Ciconia ciconia at Chessington World of Adventures Paul Wexler
 Breeding White Storks(Ciconia ciconia) at Chessington World of Adventures Paul Wexler The White Stork belongs to the genus Ciconia of which there are seven other species incorporated predominantly throughout
Breeding White Storks(Ciconia ciconia) at Chessington World of Adventures Paul Wexler The White Stork belongs to the genus Ciconia of which there are seven other species incorporated predominantly throughout
LIFE history tradeoffs are prevalent in nature because
 Copeia 2012, No. 1, 100 105 Do Sidewinder Rattlesnakes (Crotalus cerastes, Viperidae) Cease Feeding During the Breeding Season? Michael M. Webber 1, Xavier Glaudas 1, and Javier A. Rodríguez-Robles 1 Seasonal
Copeia 2012, No. 1, 100 105 Do Sidewinder Rattlesnakes (Crotalus cerastes, Viperidae) Cease Feeding During the Breeding Season? Michael M. Webber 1, Xavier Glaudas 1, and Javier A. Rodríguez-Robles 1 Seasonal
08 alberts part2 7/23/03 9:10 AM Page 95 PART TWO. Behavior and Ecology
 08 alberts part2 7/23/03 9:10 AM Page 95 PART TWO Behavior and Ecology 08 alberts part2 7/23/03 9:10 AM Page 96 08 alberts part2 7/23/03 9:10 AM Page 97 Introduction Emília P. Martins Iguanas have long
08 alberts part2 7/23/03 9:10 AM Page 95 PART TWO Behavior and Ecology 08 alberts part2 7/23/03 9:10 AM Page 96 08 alberts part2 7/23/03 9:10 AM Page 97 Introduction Emília P. Martins Iguanas have long
phenotypes of hatchling lizards, regardless of overall mean incubation temperature
 Functional Ecology 2004 Seasonal shifts in nest temperature can modify the Blackwell Publishing, Ltd. phenotypes of hatchling lizards, regardless of overall mean incubation temperature R. SHINE* Biological
Functional Ecology 2004 Seasonal shifts in nest temperature can modify the Blackwell Publishing, Ltd. phenotypes of hatchling lizards, regardless of overall mean incubation temperature R. SHINE* Biological
VIRIDOR WASTE MANAGEMENT LIMITED. Parkwood Springs Landfill, Sheffield. Reptile Survey Report
 VIRIDOR WASTE MANAGEMENT LIMITED Parkwood Springs Landfill, Sheffield July 2014 Viridor Waste Management Ltd July 2014 CONTENTS 1 INTRODUCTION... 1 2 METHODOLOGY... 3 3 RESULTS... 6 4 RECOMMENDATIONS
VIRIDOR WASTE MANAGEMENT LIMITED Parkwood Springs Landfill, Sheffield July 2014 Viridor Waste Management Ltd July 2014 CONTENTS 1 INTRODUCTION... 1 2 METHODOLOGY... 3 3 RESULTS... 6 4 RECOMMENDATIONS
A SURVEY FOR THREATENED AND ENDANGERED HERPETOFAUNA IN THE LOWER MARAIS DES CYGNES RIVER VALLEY
 ('. A SURVEY FOR THREATENED AND ENDANGERED HERPETOFAUNA IN THE LOWER MARAIS DES CYGNES RIVER VALLEY KELLYJ. IRWIN JOSEPH T. COLLINS F.inal Report to the Kansas Department of Wildlife & Parks Pratt, Kansas
('. A SURVEY FOR THREATENED AND ENDANGERED HERPETOFAUNA IN THE LOWER MARAIS DES CYGNES RIVER VALLEY KELLYJ. IRWIN JOSEPH T. COLLINS F.inal Report to the Kansas Department of Wildlife & Parks Pratt, Kansas
Carphophis amoenus Family Colubridae Subfamily Xenodontidae
 Carphophis amoenus Family Colubridae Subfamily Xenodontidae Small snakes adapted for fossorial life Reduced eyes with a narrow head Tail short and sharply pointed Dorsal scales smooth Anal plate divided
Carphophis amoenus Family Colubridae Subfamily Xenodontidae Small snakes adapted for fossorial life Reduced eyes with a narrow head Tail short and sharply pointed Dorsal scales smooth Anal plate divided
Amniote Relationships. Reptilian Ancestor. Reptilia. Mesosuarus freshwater dwelling reptile
 Amniote Relationships mammals Synapsida turtles lizards,? Anapsida snakes, birds, crocs Diapsida Reptilia Amniota Reptilian Ancestor Mesosuarus freshwater dwelling reptile Reptilia General characteristics
Amniote Relationships mammals Synapsida turtles lizards,? Anapsida snakes, birds, crocs Diapsida Reptilia Amniota Reptilian Ancestor Mesosuarus freshwater dwelling reptile Reptilia General characteristics
VERTEBRATE READING. Fishes
 VERTEBRATE READING Fishes The first vertebrates to become a widespread, predominant life form on earth were fishes. Prior to this, only invertebrates, such as mollusks, worms and squid-like animals, would
VERTEBRATE READING Fishes The first vertebrates to become a widespread, predominant life form on earth were fishes. Prior to this, only invertebrates, such as mollusks, worms and squid-like animals, would
Species Fact Sheets. Order: Gruiformes Family: Cariamidae Scientific Name: Cariama cristata Common Name: Red-legged seriema
 Order: Gruiformes Family: Cariamidae Scientific Name: Cariama cristata Common Name: Red-legged seriema AZA Management: Green Yellow Red None Photo (Male): Red-legged seriemas are identical in plumage although
Order: Gruiformes Family: Cariamidae Scientific Name: Cariama cristata Common Name: Red-legged seriema AZA Management: Green Yellow Red None Photo (Male): Red-legged seriemas are identical in plumage although
10/03/18 periods 5,7 10/02/18 period 4 Objective: Reptiles and Fish Reptile scales different from fish scales. Explain how.
 10/03/18 periods 5,7 10/02/18 period 4 Objective: Reptiles and Fish Reptile scales different from fish scales. Explain how. Objective: Reptiles and Fish Reptile scales different from fish scales. Explain
10/03/18 periods 5,7 10/02/18 period 4 Objective: Reptiles and Fish Reptile scales different from fish scales. Explain how. Objective: Reptiles and Fish Reptile scales different from fish scales. Explain
Evolution of viviparity in warm-climate lizards: an experimental test of the maternal manipulation hypothesis
 doi:10.1111/j.1420-9101.2006.01296.x Evolution of viviparity in warm-climate lizards: an experimental test of the maternal manipulation hypothesis X. JI,* C.-X. LIN, à L.-H. LIN,* Q.-B. QIUà &Y.DU à *Jiangsu
doi:10.1111/j.1420-9101.2006.01296.x Evolution of viviparity in warm-climate lizards: an experimental test of the maternal manipulation hypothesis X. JI,* C.-X. LIN, à L.-H. LIN,* Q.-B. QIUà &Y.DU à *Jiangsu
Maritime Shipping on the Great Lakes and the Lake Erie Water Snake
 Activity for Biology Lesson #2 Name Period Date Maritime Shipping on the Great Lakes and the Lake Erie Water Snake Background Information on Lake Erie water snake and round goby: Lake Erie water snake:
Activity for Biology Lesson #2 Name Period Date Maritime Shipping on the Great Lakes and the Lake Erie Water Snake Background Information on Lake Erie water snake and round goby: Lake Erie water snake:
Disappearing Marine Iguanas: A Case of Population Collapse
 WLHS/Marine Biology/Oppelt Name Disappearing Marine Iguanas: A Case of Population Collapse Directions: Read the following scenarios and answer the corresponding questions Part 1: Disappearing Marine Iguanas
WLHS/Marine Biology/Oppelt Name Disappearing Marine Iguanas: A Case of Population Collapse Directions: Read the following scenarios and answer the corresponding questions Part 1: Disappearing Marine Iguanas
Lacerta vivipara Jacquin
 Oecologia (Berl.) 19, 165--170 (1975) 9 by Springer-Verlag 1975 Clutch Size and Reproductive Effort in the Lizard Lacerta vivipara Jacquin R. A. Avery Department of Zoology, The University, Bristol Received
Oecologia (Berl.) 19, 165--170 (1975) 9 by Springer-Verlag 1975 Clutch Size and Reproductive Effort in the Lizard Lacerta vivipara Jacquin R. A. Avery Department of Zoology, The University, Bristol Received
Lecture 9 - Avian Life Histories
 Lecture 9 - Avian Life Histories Chapters 12 16 Many details in book, esp know: Chpt 12 pg 338-345, 359-365 Chpt 13 pg 367-373, 377-381, 385-391 Table 13-1 Chpt 14 pg 420-422, 427-430 Chpt 15 pg 431-438,
Lecture 9 - Avian Life Histories Chapters 12 16 Many details in book, esp know: Chpt 12 pg 338-345, 359-365 Chpt 13 pg 367-373, 377-381, 385-391 Table 13-1 Chpt 14 pg 420-422, 427-430 Chpt 15 pg 431-438,