ACCEPTED. Parasitology Unit, Max Planck Institute for Infection Biology, Berlin, Germany
|
|
|
- Cori Ferguson
- 6 years ago
- Views:
Transcription
1 EC Accepts, published online ahead of print on 30 January 2009 Eukaryotic Cell doi: /ec Copyright 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved Disruption of Plasmodium sporozoite transmission by depletion of the Sporozoite Invasion-Associated Protein-1 Sabine Engelmann 1,#, Olivier Silvie 1,2,#, and Kai Matuschewski 1,2 * 1 Department of Parasitology, Heidelberg University School of Medicine, Heidelberg, Germany 2 Parasitology Unit, Max Planck Institute for Infection Biology, Berlin, Germany * Corresponding author: Phone: # , Fax: # , matuschewski@mpiib-berlin.mpg.de # These authors contributed equally. Running head: Role for Plasmodium SIAP-1 in sporozoite transmission Abstract: 178 words Total word count: 6, Key words: Plasmodium, sporozoite, gliding motility, Anopheles vector, malaria transmission Abbreviations: ag17, antigen 17; CSP, circumsporozoite protein; S5, sporozoite-specific gene 5; SIAP-1, sporozoite invasion-associated protein 1.
2 24 25 ABSTRACT Accumulation of infectious Plasmodium sporozoites in the Anopheles salivary glands 26 marks the final step of the complex development of the malaria parasite in the insect vector. Sporozoites are formed inside midgut-associated oocysts and actively egress into the mosquito hemocoel. Traversal of the salivary gland acinar cells correlates with the sporozoite s capacity to perform continuous gliding motility. Here, we characterized the cellular role of the Plasmodium berghei sporozoite invasion- associated protein-1 (SIAP-1). Intriguingly, SIAP-1 orthologs are found exclusively in apicomplexan hemoprotozoa, parasites that are transmitted by arthropod vectors, i.e. Plasmodium, Babesia and Theileria. By fluorescent tagging with mcherry we show that SIAP-1 is expressed in oocyst-derived and salivary gland-associated sporozoites, where it accumulates at the apical tip. Targeted disruption of SIAP-1 does not affect sporozoite formation but causes a partial defect in sporozoite egress from oocysts and abolishes sporozoite colonization of mosquito salivary glands. siap-1(-) parasites are blocked in their capacity to perform continuous gliding motility. We propose that arthropod-transmitted apicomplexan parasites specifically express secretory factors, 40 such as SIAP-1, that mediate efficient oocyst exit and migration to the salivary glands. 2
3 INTRODUCTION Protozoan parasites of the phylum Apicomplexa adopted an obligate intracellular life style in a wide range of animal hosts. Apicomplexan parasites share many characteristics, including a set of apical specialized secretory organelles, active substrate-dependent locomotion, and compartimentalization of biosynthetic pathways in apicoplasts. Transmission to the vertebrate hosts occurs via tailor-made motile parasite stages, termed sporozoites, which are formed inside oocysts, the only extracellular replication phase during the complex apicomplexan life cycles. Apicomplexan parasites differ, however, fundamentally in their transmission modes. Coccidian parasites, such as Toxoplasma gondii, the causative agent of toxoplasmosis, and Cryptosporidia, which can cause life-threatening diarrheas, form oocysts that are taken up orally by contaminated food or water, respectively. After accidental ingestion sporozoites are liberated from the oocysts and breach or directly invade the intestinal endothelium to commence an infection. In marked contrast, sporozoites of apicomplexan hemoprotozoa are inoculated intradermally by arthropod vectors, i.e. Anopheles mosquitoes or ticks. Sporozoites then actively leave the inoculation site and enter the blood circulation (4). In the case of Plasmodium, the causative agent of malaria, sporozoites are formed in midgut-associated oocysts (15), actively exit the oocyst (1, 23), and penetrate the distal portion of the lateral salivary gland lobes, where they eventually accumulate rendering the infected mosquito infectious to the vertebrate host (3). Colonization of the salivary glands by Plasmodium parasites is driven by a number of stage-specific surface proteins (9), including (i) the major sporozoite surface protein circumsporozoite protein (CSP) (18), (ii) the sporozoite adhesin apical membrane antigen/ erythrocyte binding-like protein (MAEBL) (8), and (iii) the sporozoite invasin 3
4 thrombospondin-related anonymous protein (TRAP) (7, 10, 17, 24). However, none of these proteins are conserved across apicomplexan hemoprotozoa, indicating a high degree of parasite / invertebrate host co-adaptation. Towards a better understanding of the basic molecular mechanisms of sporozoite motility and arthropod-mediated transmission, we searched for sporozoite-specific candidates that are shared between Plasmodium, Babesia, and Theilaria. SIAP-1/ag17/S5 (PFD0425w) was first discovered in a systematic screen for P.falciparum sporozoite antigens recognized by sera from individuals that were immunized with irradiated sporozoites and termed antigen 17 (ag17) (2). Subsequently, the P. yoelii ortholog was isolated in a suppression subtractive hybridization screen for genes that are specifically upregulated in sporozoites as compared to blood stages (6). More recently, the P. falciparum ortholog was identified as a gene that is transiently upregulated in activated P.falciparum sporozoites (14). In this study, we fluorescently tagged the PbSIAP-1 gene and show enrichment of the protein in the apical pole of the sporozoite. Targeted gene deletion resulted in viable blood stage parasites that displayed a specific and complete block in sporozoite transmission to the vertebrate host. 81 4
5 MATERIAL AND METHODS Experimental animals. Animals were from Charles River Laboratories. All animal work was conducted in accordance with European regulations and approved by the state authorities (Regierungspräsidium Karlsruhe). Reverse transcriptase-pcr. Total RNA was purified from P. berghei ANKA (GFP507cl1)-infected mosquito midguts (oocysts), salivary glands (sporozoites), and from infected HepG2 cells (early liver stages) or mouse erythrocytes (mixed blood stages) using the RNeasy kit (Qiagen). Reverse transcription was performed using the RETROscript kit (Ambion). Real time qpcr was performed on cdna preparations using the ABI 7500 sequence detection system and Power SYBR Green PCR Master Mix (Applied Biosystems), according to the manufacturer s instructions. qpcr was performed in triplicates, with 1 cycle of 95 C for 15 min, followed by 40 cycles of 95 C for 15 s, 55 C for 15 s, and 60 C for 45 s. Standard curves were generated for all primers using WT cdna serial dilutions and gave amplification efficiencies of %. Data were analyzed with the SDS software (Applied Biosystems). Relative transcript abundance was normalized to the GFP gene, which is constitutively expressed in P. berghei ANKA GFP507cl1 (5). The following primers were used for real time PCR: GFP forward, 5 - GATGGAAGCGTTCAACTAGCAGACC-3 ; GFP reverse, 5 - AGCTGTTACAAACTCAAGAAGGACC-3 ; SIAP-1 forward, 5 - CAGCAATTAGGGACAGTGATGG-3 ; SIAP-1 reverse, 5 - AATAATGGACACCTCCGTGTGG-3 ; HSP70 forward, 5-5
6 TGCAGCAGATAATCAAACTC-3 ; HSP70 reverse, 5 -ACTTCAATTTGTGGAACACC- 3. Generation of the mcherry-tagged SIAP-1 parasite line. For targeted fluorescent tagging of PbSIAP-1, a fragment corresponding to the 3 end of PbSIAP-1 ORF without the stop codon was amplified by PCR using P.berghei genomic DNA as template and primers mcherry-siap-1for (5 - ATAAGAATGCGGCCGCGCATCTGAAAGTAAAGGAAAATGGGTCGC-3 ; NotI site is underlined) and mcherry- SIAP-1rev (5 - GGGCTAGCTGAATTGTCCACGAAAACACTGTAACTATAGG-3 ; NheI site is underlined). The PCR product was cloned into the B3D+mCherry vector (16), immediately upstream of the mcherry sequence and PbDHFR/TS 3 UTR, resulting in the SIAP- 1/mCherry targeting plasmid. The SIAP-1/mCherry plasmid was linearized with SpeI, and parasite transfection, positive selection, and parasite cloning were performed as described previously (5). Integration-specific PCR amplification of the mcherry-tagged SIAP-1 was generated using specific primer combinations. We obtained one parasite population that was used for a systematic expression and localization analysis. Expression of the mcherry fusion proteins was analyzed through direct detection of the red fluorescence of mcherry by confocal microscopy. Images were acquired on a Zeiss LSM510 confocal system (Zeiss, Germany) and processed with Adobe Photoshop software (Adobe Systems Inc.) Generation of the siap-1(-) parasites. For targeted disruption of the SIAP-1 gene two PCRfragments, flanking the SIAP-1 open reading frame, were amplified from genomic Plasmodium berghei DNA. For the 5 UTR amplification with the primer combination 6
7 SIAP-1-Rep_for (5 GGGGTACCGTGTTTTCATGCTATATGTACATTTGC 3 ; KpnI site is underlined) and 5 SIAP-1-Rep_rev (5 CCCAAGCTTGATGATATTGACCGTAAAATCC 3 ; HindIII site is underlined) resulted in a 634-bp 5 UTR fragment. Cloning into the standard P. berghei transfection vector b3d.dt^h.^d resulted in pse-41. Next, a 788-bp 3 UTR fragment was amplified with primers 5 SIAP-1-Rep_ for (5 CGGGATCCCCTATAGTTACAGTGTTTTCG 3 ; BamHI site is underlined) and 5 SIAP-1-Rep_ rev (5 - GGACTAGTGTTCGATATACCTTGCAGATTCC-3 ; SpeI site is underlined) and cloned into pse-41. This resulted in the SIAP-1 targeting vector pse-65, which contained both fragments flanking the positive selection marker T. gondii dihydrofolate reductase/thymidylate synthase (dhfr/ts), allowing replacement of the endogenous SIAP-1 locus upon a double cross-over event and subsequent selection with the antifolate pyrimethamine. P. berghei transfection was done with 10 µg KpnI/SpeI-digested pse-65 and gradient-purified P. berghei NK65 and P. berghei ANKA (clon GFP507cl1; ref. 5) schizonts. Positive selection for successful integration of the targeting plasmid was by providing 70 µg/ml pyrimethamine with the drinking water for a period of 8 days. Transfer of the emerging P. berghei population into naive animals confirmed pyrimethamine resistance. Genomic DNAs for selected parasite populations were genotyped by an integration specific PCR using primers Tgfor (5 CCCGCACGGACGAATCCAGATGG 3 ) and SIAP-1_int_check2_rev (5 CTGTGAATGTGTATATTGTGCATATGCC 3 ). Clonal siap1(-) parasite populations were obtained by limiting dilution into 15 naive NMRI mice and confirmed by integration specific PCR. As a complementary approach, we generated SIAP-1 knockout parasites specifically expressing GFP in sporozoites. For this purpose, we first amplified by PCR a fragment 7
8 comprising the CSP promoter region and the GFP coding sequence, using primers GFPfor (5 -ATAAGAATGCGGCCGCGACATGCATATGTGTTGGTTGTAATTGAGG-3 ; NotI site is underlined) and GFPrev (5 -CGCGGATCCTTATTTGTATAGTTCATCCATGCC- 3 ; BamHI site is underlined), and genomic DNA from the PbFluspo parasite line (11) as a template. Cloning of this fragment into the B3D+ vector (16) using NotI/BamHI resulted in the vector B3D+CSPGFP. 5 and 3 fragments of the PbSIAP-1 gene were then amplified using primers SIAP-1_REP1 (5 -TCCCCGCGGAGTTAACTCGAAAAAAATAAGGACG- 3 ; SacII site is underlined) and SIAP-1_REP2 (5 ATAAGAATGCGGCCGCTTTTAAGATATACCCCTTAATGAGAGC-3 ; NotI site is underlined), and primers SIAP-1_REP3 (5 - CCCAAGCTTGATGTATGTATAATACCGAAAATTGG-3 ; HindIII site is underlined) and SIAP-1_REP4 (5 -CGGGGTACCCGCCTTATTTAAGGGATAAAGAATGGG-3 ; KpnI site is underlined), respectively, and genomic DNA from wild type P. berghei. These two fragments were then cloned into the B3D+CSPGFP vector, using the respective restriction endonucleases to generate the targeting construct SIAP-1-GFPrep. Transfection, selection and cloning were then performed as described above. Genotyping PCR using specific primer combinations confirmed the expected recombination event Phenotypical analysis during the Plasmodium life cycle in vivo. Blood stage development was analyzed in vivo in asynchronous infections using NMRI mice. Gametocyte differentiation and exflagellation of microgametes were detected in mice before mosquito feedings. Anopheles stephensi mosquito rearing and maintenance was carried out under a 14h light/10h dark cycle, 75% humidity and at 28 C. Once infected, Anopheles mosquitoes were kept at 80% humidity and 20 C. Sporozoite populations were separated and analyzed 8
9 as described previously (21). Briefly, for determination of sporozoite infectivity, and numbers of midgut- and salivary gland-associated sporozoites, infected mosquitos were dissected at days 10, 14, and 17 after feeding, respectively. Hemocoel sporozoites were recovered at days 14, 16 and 18 after feeding. For determination of infectivity, sporozoites isolated from mosquito midguts, hemocoel or salivary glands were injected intravenously at the numbers indicated into young Sprague/Dawley (SD) rats. Patency was checked daily by microscopic examination of Giemsa-stained blood smears. In vitro experiments. For analysis of gliding motility, sporozoites isolated from infected mosquito salivary glands were deposited on glass slides coated with bovine serum albumin, and incubated at 37 C for 30 min. Trails left behind gliding parasites were then visualized using anti-csp antibodies (12), followed by anti-mouse Alexa-Fluor 488 antibodies (Molecular Probes). For analysis of EEF development, we used HuH7 cells cultured in DMEM supplemented with 10% FCS and antibiotics. P. berghei sporozoites were added in triplicate wells, incubated for 2 hours at 37 C, and washed off. After 48 hours, exoerythrocytic forms (EEFs) were revealed using primary antibodies against Plasmodium heat shock protein 70 (HSP70) (20), followed by the anti-mouse Alexa Fluor 488 secondary antibody. In vitro analysis of the PbSIAP-1/mCherry parasites was performed in HepG2 cells, as described before (16). 9
10 RESULTS SIAP-1 is conserved in arthropod-transmitted Apicomplexa and is specifically expressed in P. berghei mosquito stages. We initiated our characterization of SIAP-1/ag17/S5 by an in silico analysis of orthologous genes in Plasmodium species and related apicomplexan parasites (Fig. 1A). The SIAP-1 genes from the model rodent Plasmodium species, P. yoelii and P. berghei, encode proteins of 999 amino acid residues each. SIAP-1 members appear to be potentially secreted proteins and are readily retrieved from annotated Plasmodium genomes. Remarkably, SIAP- 1 orthologs are present in the Babesia and Theileria genomes (Suppl. Fig. 1), but absent in Toxoplasma and Cryptosporidium, indicating that its presence may be restricted to arthropod-transmitted apicomplexan parasites (Fig. 1A). Analysis of SIAP-1 protein sequence indicates the presence of a signal peptide, but no identified conserved domain (Suppl. Fig. 1). We next analyzed SIAP-1 transcript abundance during sporozoite maturation in the rodent malaria model parasite P. berghei. cdnas generated from asexual blood stages, liver stages and oocyst and salivary gland-associated sporozoites were used as templates for quantitative RT-PCR (Fig. 1B). PbSIAP-1 expression was highest in young oocysts and sporozoites, and rapidly decreased after sporozoite invasion of hepatocytes. In good agreement with data from P. yoelii (6), expression of PbSIAP-1 was very low in blood stages. Collectively, SIAP-1 expression correlates with a presumed role in the arthropod vector PbSIAP-1 is localized at the apical tip and the surface of sporozoites 10
11 In order to investigate the cellular localization and the expression timing of PbSIAP- 1, we generated parasite lines expressing the endogenous SIAP-1 protein fused to the mcherry red fluorescent protein (13). This was achieved by transfection of targeting vectors that contain an amino-terminally truncated PbSIAP-1 copy and in-frame fusion of the mcherry coding region, followed by the DHFR/TS 3 untranslated region (Suppl. Fig. 2A). Upon a single crossover event, integration of this construct is predicted to result in an allelic duplication, resulting in a mcherry tagged full-length copy and a non-transcribed 5 truncated version of the PbSIAP-1. Transfection was performed in P. berghei ANKA parasites expressing GFP (5), leading to green fluorescent parasites that express a red fluorescent SIAP-1 protein. Genotyping by PCR using specific primer combinations confirmed the desired integration event (Suppl. Fig. 2B). Importantly, PbSIAP-1/mCherry parasites completed the parasite life cycle normally. They were indistinguishable from WT parasites in development and growth of asexual stages (data not shown). PbSIAP-1/mCherry parasites produced gametocytes and exflagellation of male gametocytes was similar to WT parasites (data not shown). Transmission to Anopheles stephensi mosquitoes and oocyst development were also normal when compared to WT parasites. PbSIAP-1/mCherry oocysts produced sporozoites, which invaded mosquito salivary glands. In two independent infection experiments, we found 20,000 and 16,700 salivary gland-associated sporozoites per mosquito, respectively, numbers which are similar to those obtained with WT parasites (Figure 4D). More importantly, PbSIAP-1/mCherry sporozoites were infective as demontrated by complete liver stage development in vitro (see Fig. 2E) and in vivo. C57/BL6 mice inoculated intravenously with 5,000 PbSIAP-1/mCherry sporozoites (n=5) or exposed to the bites of 5 infected mosquitoes (n=5) all developed a patent parasitemia, at day 3 or day 4 after inoculation, respectively, similar to WT parasites. Collectively, these 11
12 data demonstrate that the C-terminal tagging of SIAP-1 had no detrimental effect on P. berghei infectivity. We first performed live cell imaging of blood stages of the transgenic PbSIAP- 1/mCherry. In good agreement with our transcription analysis and published data (6), we could not detect a red signal throughout blood stage development of the malaria parasite (data not shown). In contrast, the fusion protein was abundantly expressed in oocyst-derived and salivary gland sporozoites (Fig. 2A-B). Remarkably, in both immature and mature sporozoites, PbSIAP-1/mCherry showed a polarized distribution predominant at the apical tip of the parasites, as confirmed by examination of motile sporozoites (Fig. 2C). Interestingly, in addition to this apical concentration, PbSIAP-1/mCherry was also detected as distinct patches distributed along the parasite (Fig. 2C). These patches were sometimes clearly distributed at the periphery of the parasite, suggesting a possible association of SIAP- 1 with the sporozoite pellicle (Fig. 2D). This pattern is consistent with immunofluorescence data generated with antibodies against P. falciparum SIAP-1 (14). Interestingly, we observed no change in the distribution of PbSIAP-1/mCherry during the gliding of motile sporozoites (Fig. 2C). We further examined the fate of PbSIAP-1/mCherry after sporozoite invasion of HepG2 hepatocytic cells. In transforming sporozoites 4 hours post-infection, PbSIAP- 1/mCherry was detected as distinct patches, without clear apical polarization (Fig. 2E). These dotted structures were still detected 24 hours post-infection, although at a lower intensity, but had completely disappeared in mature liver stages 72 hours post-infection (Fig. 2E), consistent with the absence of PbSIAP-1/mCherry expression observed in blood stages Generation of PbSIAP-1 knockout parasites 12
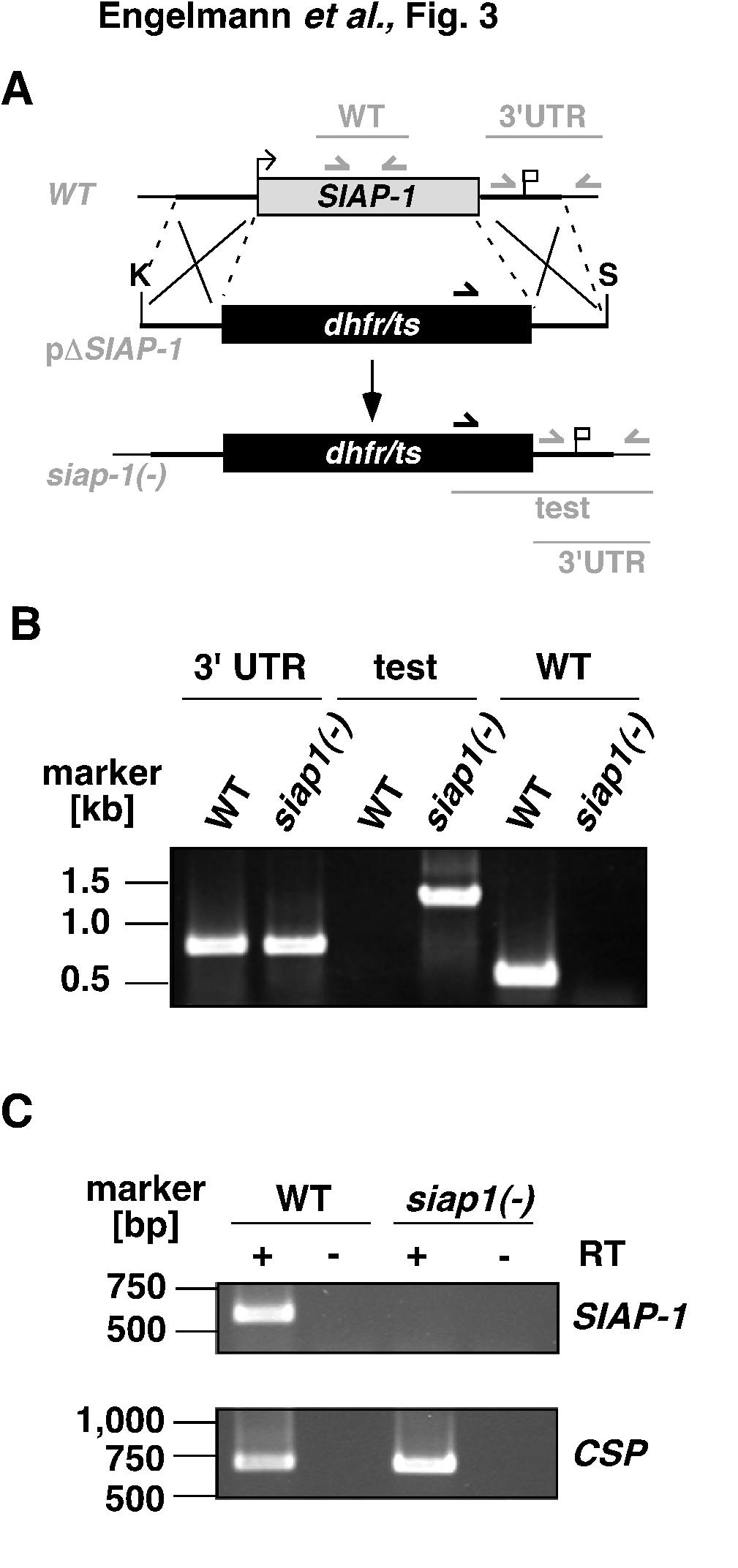
13 To address the function of SIAP-1 during the Plasmodium life cycle, we generated loss-of-function P. berghei parasites. We used a replacement strategy (Fig. 3A) to disrupt the endogenous SIAP-1 gene copy by double crossover homologous recombination. A targeting construct comprising 5 and 3 fragments of SIAP-1 flanking the pyrimethamine- resistance cassette was used for positive selection after parasite transfection. The parental blood stage population from a successful transfection was used to isolate one disruptant clone, which was subsequently used for phenotypical analysis. Occurrence of the double crossover was confirmed by PCR in pyrimethamine-resistant parasites, using primers specific for the recombination event (Fig. 3B). The wild type locus was not detected in siap- 1(-) parasites, confirming the homogeneity of the expected recombination. While CSP control transcripts were detected in WT as well as siap-1(-) sporozoites by RT-PCR, SIAP-1 transcripts were detected only in WT, but not in siap-1(-) sporozoites, confirming the successful depletion of SIAP-1 in the mutants (Fig. 3C). PbSIAP-1 is required for infection of mosquito salivary glands and transmission to the mammalian host We next examined the phenotype of siap-1(-) parasites during the P. berghei life cycle. As expected from successful disruption of the SIAP-1 gene in blood stages, siap-1(-) parasites were indistinguishable from WT parasites in development and growth of asexual stages (data not shown). SIAP-1-deficient parasites produced gametocytes and exflagellation of male gametocytes was similar to WT parasites (data not shown). These data demonstrate that SIAP-1 is not required during asexual blood stage development and sexual differentiation. 13
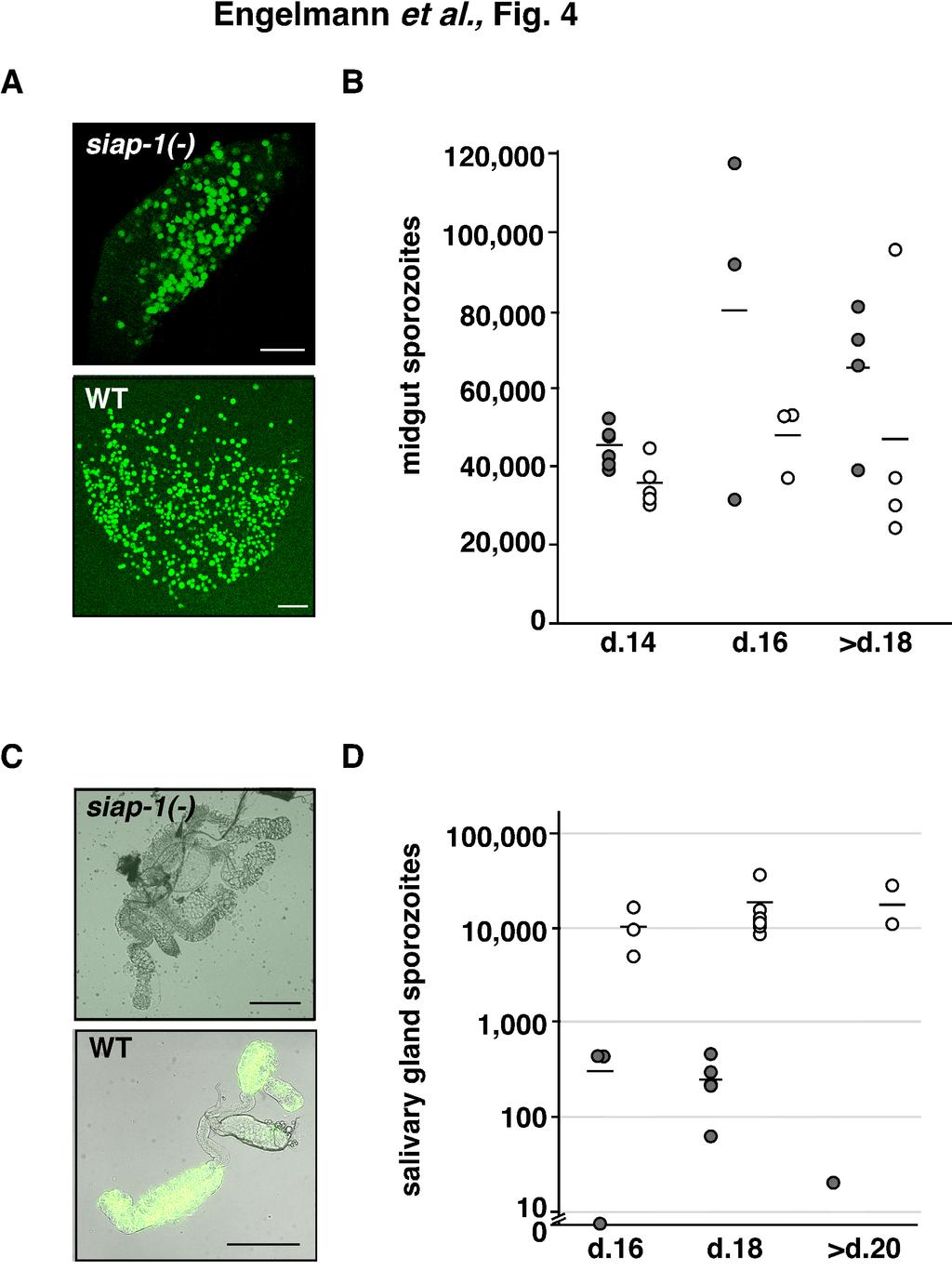
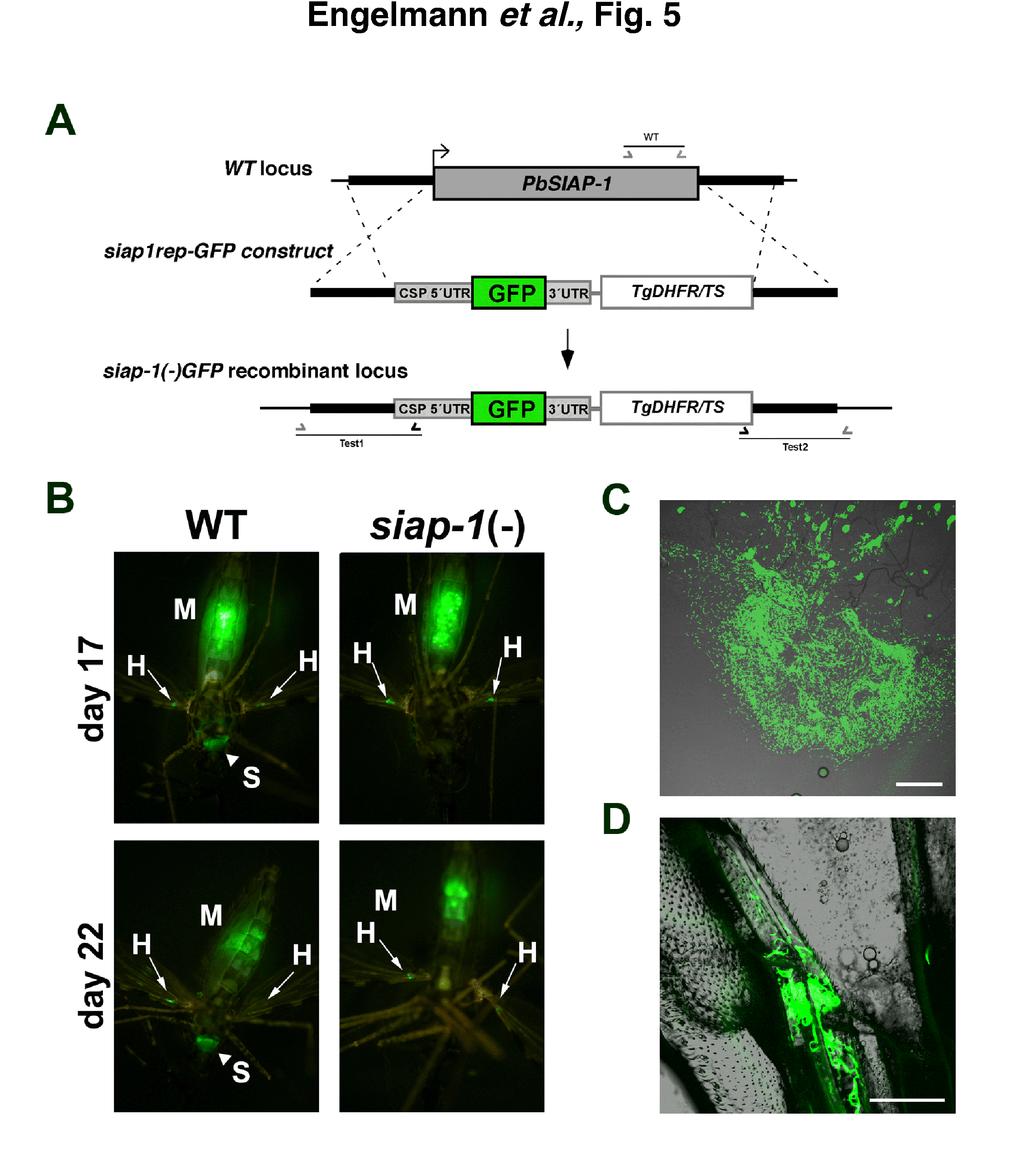
14 We next investigated transmission of SIAP-1 knockout parasites to Anopheles stephensi mosquitoes. We let mosquitoes feed on mice infected with siap-1(-) or WT parasites, and at different time points after the blood meal mosquitoes were dissected and examined for parasite development. Examination at days 10 and 14 demonstrated that siap- 1(-) parasites formed high numbers of oocysts and oocyst-derived sporozoites, respectively, as did WT parasites (Fig. 4 A,B). We noticed persistence of high numbers of midgut- associated sporozoites per infected mosquito beyond day 18 after infection (Fig. 4B). Strikingly, at day 16 after mosquito feeding, the number of siap-1(-) sporozoites associated with salivary glands was dramatically reduced as compared to WT parasites (Fig. 4 C,D). This defect was also observed when salivary glands were examined later, beyond day 20. These results indicate that siap-1(-) sporozoites have a defect in egress from oocysts and/or invasion of salivary glands. To discriminate between these two possibilities, we performed in vivo imaging of whole infected mosquitoes. For this purpose, we generated an additonal SIAP-1-deficient line using a replacement cassette containing the GFP coding sequence under control of the CSP promoter, cloned in tandem with the positive selection marker Tgdhfr/ts (Fig. 5A). After integration of this construct in recombinant P. berghei parasites, we obtained a siap- 1(-)/GFP parasite line, characterized by deletion of the SIAP-1 gene (Suppl. Fig. 3) and expression of high levels of GFP in sporozoites, similar to the Pbfluspo line (11), which was used as a WT control. We first examined WT (Pbfluspo) at day 17 and day 22 after mosquito infection (Fig. 5B). Parasites could be detected in three different compartments, the midgut, the salivary glands, and the wings, the latter apparently reflecting the presence of parasites in the hemocoel (Fig. 5B). In sharp contrast, siap-1(-)/gfp parasites were only observed in the 14
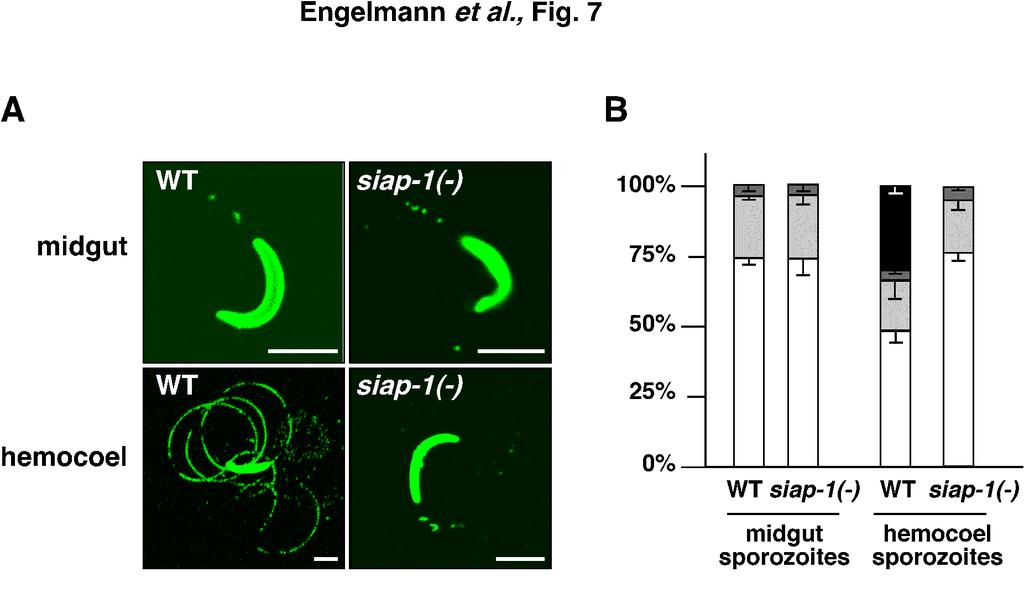
15 midgut and the wings of infected mosquitoes, but not in the salivary glands (Fig. 5B). Closer examination of the midgut of siap-1(-)/gfp-infected mosquitoes day 21 post-infection revealed the presence of numerous oocysts, most of which quickly ruptured after dissection of the midgut, releasing high numbers of sporozoites (Fig. 5C). This is consistent with a defect in egress from oocysts in SIAP-1-deficient parasites. However, this defect is not complete, since siap-1(-)/gfp sporozoites could be found in the hemocoel, as demonstrated by closer examination of the wings of infected mosquitoes (Fig. 5D). These sporozoites were either non-motile or displayed a limited non-productive motility (data not shown), as typically observed with oocyst-derived sporozoites. To provide a quantitative assessment of siap-1(-) sporozoite egress, we isolated hemolymph from infected mosquitoes and determined the numbers of hemocoel sporozoites (Fig. 6). We consistently recovered siap-1(-) sporozoites from hemocoel, but the numbers were typically lower when compared to WT-infected mosquitoes. This result differs markedly from TRAP knockout sporozoites, which egress normally but are impaired in their capacity to invade salivary glands, resulting in a significant accumulation of hemocoel sporozoites (10). Lower hemocoel sporozoite numbers of the siap-1(-) parasites indicate a defect in exiting oocysts. However, the siap-1(-) phenotype is intermediary between WT and the ones observed for egress mutants, i.e. the CSP region II mutant and the Pbsera5/ecp1(-) parasites (1, 23). Collectively, these data confirm that deletion of SIAP-1 causes a partial defect in egress from oocysts, and indicate an additional effect on invasion of salivary glands. We next examined gliding locomotion of the siap-1(-) sporozoites (Fig. 7). Midgutassociated sporozoites isolated from siap-1(-) infected mosquitoes are indistinguishable from WT sporozoites and display infrequent, non-productive motility. Continuous gliding 15
16 locomotion, as seen in a proportion of WT hemocoel sporozoites was completely absent from siap-1(-) sporozoites, indicating an important role for SIAP-1 in this process. Finally, we investigated the role of SIAP-1 in transmission of sporozoites to the mammalian host (Table 1). Rats were exposed to the bites of mosquitoes infected with WT or siap-1(-) parasites. Subsequently, animals were checked daily by blood smear examination to detect the appearance of a blood stage infection. In contrast to WT parasites, which induced parasitemia in all exposed animals, none of the siap-1(-) exposed rats developed a patent blood stage infection (Table 1). We obtained the same result in C57/BL6 mice. All mice exposed to the bites of 20 siap-1(-) (n = 2 mice) or siap-1(-)/gfp-infected mosquitoes (n = 5 mice) remained blood smear-negative. Furthermore, we bypassed natural transmission and injected mice with siap-1(-) sporozoites isolated from salivary glands, the hemocoel or midguts of infected mosquitoes. Infected animals typically remained blood smear-negative (Table 1). As observed previously with the trap(-) mutant (17) only very high inocula, i.e. 50,000 salivary gland sporozoites or 1 Mio. midgut-associated sporozoites, induced patent animals. Collectively, our results demonstrate that SIAP-1 plays a dual role in parasite egress from midgut-associated oocysts and sporozoite colonization of salivary glands. Both cellular functions are directly linked to sporozoite locomotion, a vital parasite activity that is abolished in SIAP-1 loss-of-function lines. These defects result in a complete block of sporozoite transmission from highly infected mosquitoes. 16
17 DISCUSSION In this study we identified the first conserved gene among apicomplexan hemoprotozoa that plays an essential role in sporozoite transmission to the mammalian host. SIAP-1 loss-of- function sporozoites display a specific defect in sporozoite maturation. Anopheles mosquitoes infected with siap-1(-) parasites cannot transmit malaria to susceptible rodents, despite normal formation of sporozoites in their midguts. Mutant parasites fail to mature, as seen by the lack of continuous gliding locomotion in hemocoel sporozoites and colonization of the final target organ, the salivary glands. This phenotype correlates with a transient accumulation in midgut oocysts and is reminiscent of the previously described egress mutants (1, 23). However, a direct role in oocyst rupture, is unlikely because siap-1(-) sporozoites were consistently recovered outside the midgut. One possible function of SIAP-1 is a role in parasite locomotion. This notion is supported by the complete absence of trail formation and entry into salivary glands. In addition, tagging of the endogenous SIAP-1 copy revealed a striking distribution with apical polarization and accumulation to additional specific areas. Intriguingly, these SIAP-1- positive patches remain constant upon sporozoite motility. Therefore, an immediate role, for instance in association with TRAP family-invasins, in rearward distribution of tight junctions between sporozoite receptors and the substratum is unlikely. Because of the paucity of typical microfilaments in extracellular stages of apicomplexan parasites we could not address whether the composition of the motor machinery in vivo is altered in siap-1(-) sporozoites. The distribution pattern of PbSIAP-1/mCherry combined with the absence of trail formation suggest that SIAP-1 may localize to structures located under the sporozoite surface and may be involved in gliding motility, possibly by interacting with the inner 17
18 membrane complex. Alternatively, we cannot exclude an extracellular function, perhaps in preparation for active locomotion. In P. falciparum SIAP-1 is processed and released into the supernatant upon incubation at 37C (14), a feature shared with the apical membrane antigen 1 (AMA-1) and TRAP. Presently, it is unknown where SIAP-1 is clipped. Due to the presence of a predicted cleavable signal sequence, we fluorescently tagged SIAP-1 at the carboxy-terminus. Hence, we may only be able to visualize the unprocessed full-length protein and localization of the mature form may differ, including dissociation into the extracellular milieu and deposition onto trails. At this stage, we cannot formally exclude a protective role of SIAP-1 against clearance in the mosquito. Sporozoites that fail to reach and penetrate the salivary glands are quickly cleared by the mosquito immune system (22). Previous studies with trap(-) and maebl (-) parasites showed that a block in life cycle progression translates into accumulation of hemocoel sporozoites. Absence of SIAP-1 on the sporozoite surface may render sporozoites more accessible to the mosquito innate immune system, resulting in enhanced clearance from the mosquito hemocoel. Our results with siap-1(-) parasites clearly demonstrate that SIAP-1 is required for efficient egress from oocysts and also for invasion of the mosquito salivary glands. In particular, using a novel approach that allows sensitive detection of fluorescent parasites in whole infected mosquitoes, we could show that a proportion of siap-1(-) sporozoites could exit from oocysts and access the hemocoel. Nevertheless, these parasites were not able to efficiently invade salivary glands. We hypothesize that the defect in motility in siap-1(-) sporozoites is responsible for the dual defect in egress from oocysts and invasion of salivary glands, resulting in a complete block in sporozoite transmission to the vertebrate host. 18
19 Apicomplexan sporogony in ticks differs largely from Plasmodium sporozoite formation in Anopheles mosquitoes. For instance, Babesia and Theileria sporozoite formation is initiated after kinete invasion of salivary glands and occurs inside a multinucleated sporoblast/sporont residing in a hypertrophied salivary gland cell. After exiting the sporoblast, sporozoites can be directly injected with the saliva into the vertebrate host, largely omitting the need for sporozoite-encoded proteins that function in the tick vector. Therefore, functional orthologs of Plasmodium TRAP and/or MAEBL may rather be expressed by the motile kinete. SIAP-1 stands out as it appears to be structurally conserved across apicomplexan hemoprotozoa and may play similar roles in the final maturation steps of Babesia and Theileria sporozoites. Potential roles in sporozoite exit from the sporoblast or sporont and gliding locomotion will be difficult to address by genetic approaches in these parasites. Expression profiling during the parasite life cycles and complementation of the siap1(-) phenotypes described in this study will reveal the degree of similarity between the orthologous genes. Ultimately, identifying the conserved cellular roles of SIAP-1 will provide important insights into basic principles of sporozoite biology and of arthropod-borne parasitic diseases. Acknowledgements We thank Marion Steinbuechel for assistance with WT-infected mosquito experiments, Diana Scheppan for assistance with whole mosquito pictures, and Kristin Götz for technical assistance. This work was supported in part by grants from the research focus 'Tropical Medicine Heidelberg' of the Medical Faculty of Heidelberg University, the Deutsche Forschungsgemeinschaft (SFB 544, #B10), the European Commission (BioMalPar, #23), the 19
20 Joachim Siebeneicher Foundation and the Chica and Heinz Schaller Foundation. OS is a recipient of a Marie Curie Intra-European fellowship and an EMBO Long Term Fellowship. REFERENCES 1. Aly, A.S. and K. Matuschewski A malarial cysteine protease is necessary for Plasmodium sporozoite egress from oocysts. J. Exp. Med. 202: Doolan, D.L., S. Southwood, D.A. Freilich, J. Sidney, N.L. Graber, L.Shatney, L. Bebris, L. Florens, C. Dobano, A.A. Witney, E. Appella, S.L. Hoffman, J.R. Yates III., D.J. Carucci, and A. Sette Identification of Plasmodium falciparum antigens by antigenic analysis of genomic and proteomic data. Proc. Natl. Acad. Sci. U.S.A. 100: Frischknecht, F., P. Baldacci, B. Martin, C. Zimmer, S. Thiberge, J.C. Olivo-Marin, S.L. Shorte, and R. Ménard Imaging movement of malaria parasites during transmission by Anopheles mosquitoes. Cell. Microbiol. 6: Frischknecht, F The skin as interface in the transmission of arthropod-borne pathogens. Cell. Microbiol. 9: Janse, C.J., B. Franke-Fayard, G.R. Mair, J. Ramesar, C. Thiel, S. Engelmann, K. Matuschewski, G.J. van Gemert, R.W. Sauerwein, and A.P. Waters High efficiency transfection of Plasmodium berghei facilitates novel selection procedures. Mol. Biochem. Parasitol. 145: Kaiser, K., K. Matuschewski, N. Camargo, J. Ross, and S.H. Kappe Differential transcriptome profiling identifies Plasmodium genes encoding preerythrocytic stage-specific proteins. Mol. Microbiol. 51:
21 Kappe, S., T. Bruderer, S. Gantt, H. Fujioka, V. Nussenzweig, and R. Ménard Conservation of a gliding motility and cell invasion machinery in Apicomplexan parasites. J. Cell Biol.147: Kariu, T., M. Yuda, K. Yano, and Y. Chinzei MAEBL is essential for malarial sporozoite infection of the mosquito salivary gland. J. Exp. Med. 195: Matuschewski, K Getting infectious: formation and maturation of Plasmodium sporozoites in the Anopheles vector. Cell. Microbiol. 8: Matuschewski, K., A.C. Nunes, V. Nussenzweig, and R. Ménard Plasmodium sporozoite invasion into insect and mammalian cells is directed by the same dual binding system. EMBO J. 21: Natarajan, R., Thathy, V., Mota, M.M., Hafalla, J.C., Ménard, R., and Vernick, K Fluorescent Plasmodium berghei sporozoites and pre-erythrocytic stages: a new tool to study mosquito and mammalian host interactions with malaria parasites, Cell. Microbiol. 3: Potocnjak, P., N. Yoshida, R.S. Nussenzweig, and V. Nussenzweig Monovalent fragments (Fab) of monoclonal antibodies to a sporozoite surface antigen (Pb44) protect mice against malarial infection. J. Exp. Med. 151: Shaner, N.C., R.E. Campbell, P.A. Steinbach, B.N. Giepmans, A.E. Palmer, and R.Y. Tsien (2004) Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 22: Siau, A., Silvie, O., Franetich, J.-F., Yalaoui, S., Marinach, C., Hannoun, L., van Gemert, G.-J., Luty, A.J.F., Bischoff, E., David, P.H., Snounou, G., Vaquero, C., Froissard, P., and Mazier, D Temperature shift and host cell contact up- 21
22 regulate sporozoite expression of Plasmodium falciparum genes involved in hepatocyte invasion. PloS Pathog. 4: e Sinden, R.E., and K. Matuschewski The sporozoite. In Molecular Approaches to Malaria. Sherman I. (ed.). Washington DC: American Society for Microbiology Press, pp Silvie, O., K. Goetz, and K. Matuschewski A sporozoite asparagine-rich protein controls initiation of Plasmodium liver stage development. PLoS Pathog. 4: e Sultan, A.A., V. Thathy, U. Frevert, K.J. Robson, A. Crisanti, V. Nussenzweig, R.S. Nussenzweig, and R. Ménard TRAP is necessary for gliding motility and infectivity of Plasmodium sporozoites. Cell 90: Tewari, R., D. Rathore, and A. Crisanti Motility and infectivity of Plasmodium berghei sporozoites expressing avian Plasmodium gallinaceum circumsporozoite protein. Cell. Microbiol. 7: Thathy, V., and R. Ménard Gene targeting in Plasmodium berghei. Methods Mol. Med. 72: Tsuji, M., D. Mattei, R.S. Nussenzweig, D. Eichinger, and F. Zavala Demonstration of heat-shock protein 70 in the sporozoite stage of malaria parasites. Parasitol. Res. 80: Vanderberg, J.P Development of infectivity by the Plasmodium berghei sporozoite. J. Parasitol. 61: Vaughan, J.A., L. Hensley, and J.C. Beier Sporogonic development of Plasmodium yoelii in five anopheline species. J. Parasitol. 80:
23 Wang, Q., H. Fujioka, and V. Nussenzweig Exit of Plasmodium sporozoites from oocysts is an active process that involves the circumsporozoite protein. PLoS Pathog. 1: e Wengelnik, K., R. Spaccapelo, S. Naitza, K.J.H. Robson, C.J. Janse, F. Bistoni, A.P. Waters, and A. Crisanti The A-domain and the thrombospondin-related motif of Plasmodium falciparum TRAP are implicated in the invasion process of mosquito salivary glands. EMBO J. 18:
24 501 FIGURE LEGENDS FIG 1. Plasmodium sporozoite invasion-associated protein-1 (SIAP-1). (A) Schematic diagram of the primary structures of the apicomplexan SIAP-1 proteins. The primary structure of P.berghei SIAP-1 (PB ; re-annotated), and the orthologs of P. yoelii (PY00455), P.falciparum (PFD0425w), P.vivax (Pv000815), Babesia bovis (XP_ ), and Theileria parva (XP_765499) are indicated as white boxes. The predicted cleavable signal peptide is represented in grey. Amino acid sequence identities are indicated as percentage of identical residues compared to the P. berghei sequence. (B) Quantitative RT-PCR analysis of P. berghei SIAP-1 gene expression in midgut oocysts (day 12 post-infection of mosquitoes), salivary gland sporozoites (day 18 post-infection of mosquitoes), early liver stages (6h post-infection of HepG2 cells) and mixed blood stages. Relative gene expression was normalized to GFP expression level, and is shown as the mean of two independent experiments (+/- SD). FIG. 2. Expression and localization of PbSIAP-1/mCherry in sporozoites. Expression of the mcherry tagged SIAP-1 (red) was analyzed by confocal fluorescence microscopy of SIAP-1/mCherry P. berghei parasites constitutively expressing GFP (green). (A) Midgut sporozoites. Bars, 20 µm. (B) Salivary gland-associated sporozoites. Bars, 10 µm. (C) Fluorescent imaging of a motile sporozoite. Sequential acquisition of the red fluorescence and DIC confirms SIAP-1 accumulation mainly at the apical tip. Note that the distribution of SIAP-1/mCherry is not modified as the sporozoite glides. Bar, 5 µm. (D) Higher magnification of a fixed salivary gland-associated sporozoite. (E) Confocal microscopy analysis of HepG2 cell cultures 4 h, 24 h and 72 h after infection with P. berghei parasites 24
25 expressing GFP (green) and SIAP-1/mCherry (red). Nuclei were stained with Hoechst (blue). Bars, 10 µm. FIG. 3. Targeted deletion of the P. berghei SIAP-1 gene. (A) Replacement strategy for targeted gene disruption of PbSIAP-1. The wild-type SIAP-1 locus (WT) is targeted with a KpnI (K)/ SpeI (S)-linearized replacement plasmid (p SIAP-1) containing the 5 and 3 UTR of PbSIAP-1 and the positive selection marker TgDHFR-TS. After double cross over homologous recombination, the SIAP-1 open reading frame is substituted by the selection marker, resulting in the mutant siap-1(-) allele. Replacement- and WT-specific test primer combinations and expected fragments are shown as lines. (B) Replacement-specific PCR analysis. Confirmation of the predicted gene targeting is done with primer combinations that only amplify a signal in the recombinant locus (test). The absence of a WT-specific signal in the clonal siap-1(-) population confirms the purity of the mutant parasite line. (C) Depletion of SIAP-1 transcripts in siap-1(-) parasites. cdna from WT and siap-1(-) sporozoites was used as template for SIAP-1-specific PCR reactions (upper panel). Amplification of the circumsporozoite protein (CSP) transcripts was used as a positive control (lower panel). FIG. 4. siap-1(-) sporozoites are impaired in egress from oocysts. (A) Representative immunofluorescence pictures of infected A. stephensi midguts at day 14 after infection. (B) Quantification of oocyst-associated sporozoites per infected mosquito for siap-1(-) (grey circles) and WT (white circles) parasites. The bars represent the mean values. (C) Representative merged bright field and immunofluorescence pictures of infected A. stephensi salivary glands at day 17 after infection. (D) Quantification of salivary gland- 25
26 associated sporozoites per infected mosquito for siap-1(-) (grey circles) and WT (white circles) parasites. Note the logarithmic scale. Bars, 200 µm. FIG. 5. siap-1(-) sporozoites fail to colonize the mosquito salivary glands. (A) Replacement strategy for targeted gene disruption of PbSIAP-1 and insertion of a GFP expression cassette. The wild-type SIAP-1 locus is targeted with a SacII/KpnI-linearized replacement plasmid (siap1rep-gfp construct) containing the 5 and 3 UTR of PbSIAP-1, the positive selection marker TgDHFR-TS, and the GFP coding sequence under control of the CSP promoter region. After double cross-over homologous recombination, the SIAP-1 open reading frame is substituted by the selection marker and the GFP cassette, resulting in the mutant siap-1(-)gfp allele. (B) Fluorescence imaging of whole mosquitoes infected with wild type (WT) or siap-1(-)/gfp parasites, day 17 and day 22 after the infectious blood meal. Note the presence of green fluorescent WT parasites in the mosquito midgut (M), hemocoel (H, best visualized in the wings with arrows), and salivary glands (S, arrowheads). siap-1(-)/gfp parasites were only detected in the midgut (M) and the hemocoel (H, arrows), but not in salivary glands of infected mosquitoes. (C) Fluorescence microscopy of a dissected mosquito midgut 21 days after infection with siap1(-)/gfp parasites. Note the presence of oocysts and the release of high numbers of sporozoites. Bar, 200 µm. (D) Fluorescence microscopy of a dissected mosquito wing 21 days after infection with siap-1(- )/GFP parasites, demonstrating the presence of sporozoites in the hemocoel. Bar, 50 µm
27 FIG. 6. Quantitative assessment of siap-1(-) sporozoite release into the mosquito hemocoel. Shown are quantifications of hemocoel sporozoites per infected mosquito for siap-1(-) (grey circles) and WT (white circles) parasites. The bars represent the mean values. FIG. 7. Defective gliding locomotion in siap-1(-) sporozoites. (A) Representative immunofluorescence pictures of sporozoites isolated from Anopheles midguts or hemocoel. Sporozoites were deposited onto coated cover slips, and trails were visualized with the anti- circumsporozoite protein (CSP) antibody. Note, that siap-1(-) hemocoel sporozoites lack productive motility and are indistinguishable from midgut-associated sporozoites. (B) Quantification of the WT and siap1(-) motility patterns. Shown is the percentage of non- motile, i.e. detached (white) and attached (light grey), sporozoites and sporozoite that display non-productive (dark grey) and productive (black) motility. In contrast to WT sporozoites that mature upon egress to the mosquito hemocoel siap-1(-) parasites remain non-gliding. 27
28 TABLE 1: In vivo infectivity of siap-1(-) parasites. sporozoite Number of animals Prepatency population parasite Inoculumr a infected (days) salivary gland siap-1(-) mosquito bite 0 / 4 N/A Wild type mosquito bite 8 / siap-1(-) 50,000 i.v. 1 / ,000 i.v. 0 / 5 N/A Wild type 10,000 i.v. 2 / hemocoel siap-1(-) 10,000 i.v. 1 / 4 (6) 5,000 i.v. 0 / 3 N/A Wild type 1,000 i.v. 10 / midgut siap-1(-) 1,000,000 i.v. 1 / ,000 i.v. 1 / 2 (7) 100,000 i.v. 1 / 7 (7) Wild type 100,000 i.v. 10 / a Young Sprague/Dawley rats were injected intravenously (i.v.) with indicated numbers of sporozoites or exposed to the bites of 5-10 infected mosquitoes at days after mosquito infection. N/A: not applicable
29

30 D E T P C A E C
31 CCEPTE
32
33
34
35
Arrested oocyst maturation in Plasmodium parasites. lacking type II NADH:ubiquinone dehydrogenase
 Supplemental Information for: Arrested oocyst maturation in Plasmodium parasites lacking type II NADH:ubiquinone dehydrogenase Katja E. Boysen and Kai Matuschewski Contents: - Supplemental Movies 1 and
Supplemental Information for: Arrested oocyst maturation in Plasmodium parasites lacking type II NADH:ubiquinone dehydrogenase Katja E. Boysen and Kai Matuschewski Contents: - Supplemental Movies 1 and
Identification of an AP2-family Protein That Is Critical for Malaria Liver Stage Development
 Identification of an AP2-family Protein That Is Critical for Malaria Liver Stage Development Shiroh Iwanaga, Izumi Kaneko, Tomomi Kato, Masao Yuda* Department of Medical Zoology, Mie University School
Identification of an AP2-family Protein That Is Critical for Malaria Liver Stage Development Shiroh Iwanaga, Izumi Kaneko, Tomomi Kato, Masao Yuda* Department of Medical Zoology, Mie University School
A Cysteine Protease Inhibitor of Plasmodium berghei Is Essential for Exo-erythrocytic Development
 A Cysteine Protease Inhibitor of Plasmodium berghei Is Essential for Exo-erythrocytic Development Christine Lehmann 1, Anna Heitmann 1, Satish Mishra 2, Paul-Christian Burda 3, Mirko Singer 4, Monica Prado
A Cysteine Protease Inhibitor of Plasmodium berghei Is Essential for Exo-erythrocytic Development Christine Lehmann 1, Anna Heitmann 1, Satish Mishra 2, Paul-Christian Burda 3, Mirko Singer 4, Monica Prado
Infecting Anopheles stephensi With Rodent Malaria Parasites Alida Coppi & Photini Sinnis
 Infecting Anopheles stephensi With Rodent Malaria Parasites Alida Coppi & Photini Sinnis A. Reagents: 1. DMEM or RPMI DMEM (4.5g/L glucose) RPMI 1640 Cellgro #MT-10-017-CM Cellgro #MT-10-040-CM 2. Giemsa
Infecting Anopheles stephensi With Rodent Malaria Parasites Alida Coppi & Photini Sinnis A. Reagents: 1. DMEM or RPMI DMEM (4.5g/L glucose) RPMI 1640 Cellgro #MT-10-017-CM Cellgro #MT-10-040-CM 2. Giemsa
Marissa Vignali*, Cate Speake* and Patrick E Duffy*
 Minireview Malaria sporozoite proteome leaves a trail Marissa Vignali*, Cate Speake* and Patrick E Duffy* Addresses: *Malaria Program, Seattle Biomedical Research Institute, Seattle, Washington 98109,
Minireview Malaria sporozoite proteome leaves a trail Marissa Vignali*, Cate Speake* and Patrick E Duffy* Addresses: *Malaria Program, Seattle Biomedical Research Institute, Seattle, Washington 98109,
Quantitative Dynamics of Plasmodium yoelii Sporozoite Transmission by Infected Anopheline Mosquitoes
 INFECTION AND IMMUNITY, July 2005, p. 4363 4369 Vol. 73, No. 7 0019-9567/05/$08.00 0 doi:10.1128/iai.73.7.4363 4369.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Quantitative
INFECTION AND IMMUNITY, July 2005, p. 4363 4369 Vol. 73, No. 7 0019-9567/05/$08.00 0 doi:10.1128/iai.73.7.4363 4369.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Quantitative
Plasmodium yoelii Sporozoites with Simultaneous Deletion of P52 and P36 Are Completely Attenuated and Confer Sterile Immunity against Infection
 INFECTION AND IMMUNITY, Aug. 2007, p. 3758 3768 Vol. 75, No. 8 0019-9567/07/$08.00 0 doi:10.1128/iai.00225-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Plasmodium yoelii Sporozoites
INFECTION AND IMMUNITY, Aug. 2007, p. 3758 3768 Vol. 75, No. 8 0019-9567/07/$08.00 0 doi:10.1128/iai.00225-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Plasmodium yoelii Sporozoites
A n estimated 3.3 billion people were at risk of malaria infection in There is as of yet no licensed
 OPEN SUBJECT AREAS: PARASITOLOGY MOLECULAR BIOLOGY Received 27 March 2014 Accepted 23 June 2014 Published 11 July 2014 Correspondence and requests for materials should be addressed to A.S.I.A. (aaly@tulane.
OPEN SUBJECT AREAS: PARASITOLOGY MOLECULAR BIOLOGY Received 27 March 2014 Accepted 23 June 2014 Published 11 July 2014 Correspondence and requests for materials should be addressed to A.S.I.A. (aaly@tulane.
The Transmembrane Isoform of Plasmodium falciparum MAEBL Is Essential for the Invasion of Anopheles Salivary Glands
 The Transmembrane Isoform of Plasmodium falciparum MAEBL Is Essential for the Invasion of Anopheles Salivary Glands Fabian E. Saenz 1,2, Bharath Balu 1, Jonah Smith 2, Sarita R. Mendonca 1,2, John H. Adams
The Transmembrane Isoform of Plasmodium falciparum MAEBL Is Essential for the Invasion of Anopheles Salivary Glands Fabian E. Saenz 1,2, Bharath Balu 1, Jonah Smith 2, Sarita R. Mendonca 1,2, John H. Adams
Motility precedes egress of malaria parasites from oocysts
 RESEARCH ARTICLE Motility precedes egress of malaria parasites from oocysts Dennis Klug*, Friedrich Frischknecht* Integrative Parasitology, Center for Infectious Diseases, Heidelberg University Medical
RESEARCH ARTICLE Motility precedes egress of malaria parasites from oocysts Dennis Klug*, Friedrich Frischknecht* Integrative Parasitology, Center for Infectious Diseases, Heidelberg University Medical
PLASMODIUM MODULE 39.1 INTRODUCTION OBJECTIVES 39.2 MALARIAL PARASITE. Notes
 Plasmodium MODULE 39 PLASMODIUM 39.1 INTRODUCTION Malaria is characterized by intermittent fever associated with chills and rigors in the patient. There may be enlargement of the liver and spleen in the
Plasmodium MODULE 39 PLASMODIUM 39.1 INTRODUCTION Malaria is characterized by intermittent fever associated with chills and rigors in the patient. There may be enlargement of the liver and spleen in the
Chimeric Plasmodium falciparum parasites expressing Plasmodium vivax circumsporozoite protein fail to produce salivary gland sporozoites
 https://doi.org/10.1186/s12936-018-2431-1 Malaria Journal RESEARCH Open Access Chimeric Plasmodium falciparum parasites expressing Plasmodium vivax circumsporozoite protein fail to produce salivary gland
https://doi.org/10.1186/s12936-018-2431-1 Malaria Journal RESEARCH Open Access Chimeric Plasmodium falciparum parasites expressing Plasmodium vivax circumsporozoite protein fail to produce salivary gland
Understanding Epidemics Section 3: Malaria & Modelling
 Understanding Epidemics Section 3: Malaria & Modelling PART B: Biology Contents: Vector and parasite Biology of the malaria parasite Biology of the anopheles mosquito life cycle Vector and parasite Malaria
Understanding Epidemics Section 3: Malaria & Modelling PART B: Biology Contents: Vector and parasite Biology of the malaria parasite Biology of the anopheles mosquito life cycle Vector and parasite Malaria
Epigenetic regulation of Plasmodium falciparum clonally. variant gene expression during development in An. gambiae
 Epigenetic regulation of Plasmodium falciparum clonally variant gene expression during development in An. gambiae Elena Gómez-Díaz, Rakiswendé S. Yerbanga, Thierry Lefèvre, Anna Cohuet, M. Jordan Rowley,
Epigenetic regulation of Plasmodium falciparum clonally variant gene expression during development in An. gambiae Elena Gómez-Díaz, Rakiswendé S. Yerbanga, Thierry Lefèvre, Anna Cohuet, M. Jordan Rowley,
CelTOS, a novel malarial protein that mediates transmission to mosquito and vertebrate hosts
 Blackwell Publishing LtdOxford, UKMMIMolecular Microbiology0950-382X 2005 The Authors; Journal compilation 2005 Blackwell Publishing Ltd? 200559513691379Original ArticleA protein that mediates malarial
Blackwell Publishing LtdOxford, UKMMIMolecular Microbiology0950-382X 2005 The Authors; Journal compilation 2005 Blackwell Publishing Ltd? 200559513691379Original ArticleA protein that mediates malarial
Downloaded from:
 Al-Khattaf, FS; Tremp, AZ; El-Houderi, A; Dessens, JT (2016) The Plasmodium alveolin IMC1a is stabilised by its terminal cysteine motifs and facilitates sporozoite morphogenesis and infectivity in a dosedependent
Al-Khattaf, FS; Tremp, AZ; El-Houderi, A; Dessens, JT (2016) The Plasmodium alveolin IMC1a is stabilised by its terminal cysteine motifs and facilitates sporozoite morphogenesis and infectivity in a dosedependent
PRINCIPAL INVESTIGATOR: Dr. Jetsumon (Sattabongkot) Prachumsri
 Prachumsri") AD (Leave blank) Award Number: W81XWH-07-2-0090 TITLE: Proteomic Study of Human Malaria Parasite Plasmodium Vivax Liver Stages for Development of Vaccines and Drugs PRINCIPAL INVESTIGATOR: Dr. Jetsumon
AD (Leave blank) Award Number: W81XWH-07-2-0090 TITLE: Proteomic Study of Human Malaria Parasite Plasmodium Vivax Liver Stages for Development of Vaccines and Drugs PRINCIPAL INVESTIGATOR: Dr. Jetsumon
Blood protozoan: Plasmodium
 Blood protozoan: Plasmodium Dr. Hala Al Daghistani The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans: four species are associated The Plasmodium spp.
Blood protozoan: Plasmodium Dr. Hala Al Daghistani The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans: four species are associated The Plasmodium spp.
A:Malaria (Plasmodium species) Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign
 Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign") A:Malaria (Plasmodium species) Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign tertian malaria P. ovale: causes benign tertian malaria
A:Malaria (Plasmodium species) Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign tertian malaria P. ovale: causes benign tertian malaria
Malaria in the Mosquito Dr. Peter Billingsley
 Malaria in the Mosquito Senior Director Quality Systems and Entomology Research Sanaria Inc. Rockville MD. 1 Malaria: one of the world s foremost killers Every year 1 million children die of malaria 250
Malaria in the Mosquito Senior Director Quality Systems and Entomology Research Sanaria Inc. Rockville MD. 1 Malaria: one of the world s foremost killers Every year 1 million children die of malaria 250
Phylum:Apicomplexa Class:Sporozoa
 Phylum:Apicomplexa Class:Sporozoa The most characteristic features of sporozoa are 1-unique appearance of most protozoa makes it possible for knowledge able person to identifiy them to level of genus and
Phylum:Apicomplexa Class:Sporozoa The most characteristic features of sporozoa are 1-unique appearance of most protozoa makes it possible for knowledge able person to identifiy them to level of genus and
PDF hosted at the Radboud Repository of the Radboud University Nijmegen
 PDF hosted at the Radboud Repository of the Radboud University Nijmegen The following full text is a publisher's version. For additional information about this publication click this link. http://hdl.handle.net/2066/70169
PDF hosted at the Radboud Repository of the Radboud University Nijmegen The following full text is a publisher's version. For additional information about this publication click this link. http://hdl.handle.net/2066/70169
Parasitology Departement Medical Faculty of USU
 Malaria Mechanism of infection Parasitology Departement Medical Faculty of USU Introduction Malaria parasites Phylum Order Suborder Family Genus Species : : Apicomplexa : Eucoccidiida : Haemosporida :
Malaria Mechanism of infection Parasitology Departement Medical Faculty of USU Introduction Malaria parasites Phylum Order Suborder Family Genus Species : : Apicomplexa : Eucoccidiida : Haemosporida :
A. Effect upon human culture 1. Control of malaria has contributed to world=s population explosion 2. Africans brought to U.S.
 VI. Malaria A. Effect upon human culture 1. Control of malaria has contributed to world=s population explosion 2. Africans brought to U.S. because they were resistant to malaria & other diseases 3. Many
VI. Malaria A. Effect upon human culture 1. Control of malaria has contributed to world=s population explosion 2. Africans brought to U.S. because they were resistant to malaria & other diseases 3. Many
Malaria. This sheet is from both sections recording and includes all slides and diagrams.
 Malaria This sheet is from both sections recording and includes all slides and diagrams. Malaria is caused by protozoa family called plasmodium (Genus) mainly affect blood system specially RBCs and each
Malaria This sheet is from both sections recording and includes all slides and diagrams. Malaria is caused by protozoa family called plasmodium (Genus) mainly affect blood system specially RBCs and each
Plasmodium sporozoites acquire virulence and. immunogenicity during mosquito hemocoel transit
 IAI Accepts, published online ahead of print on 30 December 2013 Infect. Immun. doi:10.1128/iai.00758-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 Plasmodium sporozoites
IAI Accepts, published online ahead of print on 30 December 2013 Infect. Immun. doi:10.1128/iai.00758-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 Plasmodium sporozoites
Gliding Motility Assay for P. berghei Sporozoites
 Gliding Motility Assay for P. berghei Sporozoites Important Notes: 1. For all dilutions (including antibodies and sporozoites), always make slightly more than needed. For instance, if you need 200 µl sporozoites
Gliding Motility Assay for P. berghei Sporozoites Important Notes: 1. For all dilutions (including antibodies and sporozoites), always make slightly more than needed. For instance, if you need 200 µl sporozoites
INVESTIGATING THE MOTILITY OF PLASMODIUM
 INVESTIGATING THE MOTILITY OF PLASMODIUM by Natasha Vartak A thesis submitted to Johns Hopkins University in conformity with the requirements for the degree of Master of Science Baltimore, Maryland April,
INVESTIGATING THE MOTILITY OF PLASMODIUM by Natasha Vartak A thesis submitted to Johns Hopkins University in conformity with the requirements for the degree of Master of Science Baltimore, Maryland April,
Malaria parasites: virulence and transmission as a basis for intervention strategies
 Malaria parasites: virulence and transmission as a basis for intervention strategies Matthias Marti Department of Immunology and Infectious Diseases Harvard School of Public Health The global malaria burden
Malaria parasites: virulence and transmission as a basis for intervention strategies Matthias Marti Department of Immunology and Infectious Diseases Harvard School of Public Health The global malaria burden
Automated classification of Plasmodium sporozoite movement patterns reveals a shift towards productive motility during salivary gland infection
 Automated classification of Plasmodium sporozoite movement patterns reveals a shift towards productive motility during salivary gland infection Stephan Josef Hegge, Mikhail Kudryashev, Ashley Smith, Friedrich
Automated classification of Plasmodium sporozoite movement patterns reveals a shift towards productive motility during salivary gland infection Stephan Josef Hegge, Mikhail Kudryashev, Ashley Smith, Friedrich
Blood protozoan: Plasmodium
 Blood protozoan: Plasmodium The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans:four species are associated The Plasmodium spp. life cycle can be divided
Blood protozoan: Plasmodium The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans:four species are associated The Plasmodium spp. life cycle can be divided
THE TRANSMISSION EFFICIENCY OF PLASMODIUM YOELII INFECTED MOSQUITOES
 THE TRANSMISSION EFFICIENCY OF PLASMODIUM YOELII INFECTED MOSQUITOES by Maya A. Aleshnick A thesis submitted to Johns Hopkins University in conformity with the requirements for the degree of Master of
THE TRANSMISSION EFFICIENCY OF PLASMODIUM YOELII INFECTED MOSQUITOES by Maya A. Aleshnick A thesis submitted to Johns Hopkins University in conformity with the requirements for the degree of Master of
Developmental Biology of Sporozoite-Host. Malaria: Implications for Vaccine Design. Javier E. Garcia, Alvaro Puentes and Manuel E.
 Developmental Biology of Sporozoite-Host Interactions in Plasmodium falciparum Malaria: Implications for Vaccine Design Javier E. Garcia, Alvaro Puentes and Manuel E. Patarroyo Clin. Microbiol. Rev. 2006,
Developmental Biology of Sporozoite-Host Interactions in Plasmodium falciparum Malaria: Implications for Vaccine Design Javier E. Garcia, Alvaro Puentes and Manuel E. Patarroyo Clin. Microbiol. Rev. 2006,
BIO Parasitology Spring 2009
 BIO 475 - Parasitology Spring 2009 Stephen M. Shuster Northern Arizona University http://www4.nau.edu/isopod Lecture 10 Malaria-Life Cycle a. Micro and macrogametocytes in mosquito stomach. b. Ookinete
BIO 475 - Parasitology Spring 2009 Stephen M. Shuster Northern Arizona University http://www4.nau.edu/isopod Lecture 10 Malaria-Life Cycle a. Micro and macrogametocytes in mosquito stomach. b. Ookinete
alaria Parasite Bank Collection sites of P. falciparum isolates PARASITE BIOLOGY
 M alaria Parasite Bank established in 1992 is a supporting unit for research activities on different aspects of malaria. The main objective of establishing this facility is to strengthen researches at
M alaria Parasite Bank established in 1992 is a supporting unit for research activities on different aspects of malaria. The main objective of establishing this facility is to strengthen researches at
Malaria Parasite Pre-Erythrocytic Stage Infection: Gliding and Hiding
 Malaria Parasite Pre-Erythrocytic Stage Infection: Gliding and Hiding Ashley M. Vaughan, 1 Ahmed S.I. Aly, 1 and Stefan H.I. Kappe 1,2, * 1 Seattle Biomedical Research Institute, Seattle, WA 98109, USA
Malaria Parasite Pre-Erythrocytic Stage Infection: Gliding and Hiding Ashley M. Vaughan, 1 Ahmed S.I. Aly, 1 and Stefan H.I. Kappe 1,2, * 1 Seattle Biomedical Research Institute, Seattle, WA 98109, USA
9 Parasitology 9 EXERCISE EQA. Objectives EXERCISE
 0696T_c09_81-90.qxd 07/01/2004 23:19 Page 81 EXERCISE 9 Parasitology Exercise Pre-Test Attempt to answer the following questions before starting this exercise. They will serve as a guide to important concepts.
0696T_c09_81-90.qxd 07/01/2004 23:19 Page 81 EXERCISE 9 Parasitology Exercise Pre-Test Attempt to answer the following questions before starting this exercise. They will serve as a guide to important concepts.
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12234 Supplementary Figure 1. Embryonic naked mole-rat fibroblasts do not undergo ECI. Embryonic naked mole-rat fibroblasts ( EF) were isolated from eight mid-gestation embryos. All the
doi:10.1038/nature12234 Supplementary Figure 1. Embryonic naked mole-rat fibroblasts do not undergo ECI. Embryonic naked mole-rat fibroblasts ( EF) were isolated from eight mid-gestation embryos. All the
The silent path to thousands of merozoites: the Plasmodium liver stage
 The silent path to thousands of merozoites: the Plasmodium liver stage Miguel Prudêncio*, Ana Rodriguez and Maria M. Mota* Abstract Plasmodium sporozoites are deposited in the skin of their vertebrate
The silent path to thousands of merozoites: the Plasmodium liver stage Miguel Prudêncio*, Ana Rodriguez and Maria M. Mota* Abstract Plasmodium sporozoites are deposited in the skin of their vertebrate
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
Giardia and Apicomplexa. G. A. Lozano UNBC
 Giardia and Apicomplexa G. A. Lozano UNBC NINE Protozoan diseases/parasites Ciliphora, Ichthyophthirius, Ick Sarcomastigophora, Giardia, giardiasis Apicomplexa: Eimeria, Toxoplasma, Sarcocystis, Cryptosporidium.
Giardia and Apicomplexa G. A. Lozano UNBC NINE Protozoan diseases/parasites Ciliphora, Ichthyophthirius, Ick Sarcomastigophora, Giardia, giardiasis Apicomplexa: Eimeria, Toxoplasma, Sarcocystis, Cryptosporidium.
Comparative Plasmodium gene overexpression reveals distinct perturbation of sporozoite transmission by profilin
 MBoC ARTICLE Comparative Plasmodium gene overexpression reveals distinct perturbation of sporozoite transmission by profilin Yuko Sato a,b, *, Marion Hliscs a,c, Josefine Dunst a,d, Christian Goosmann
MBoC ARTICLE Comparative Plasmodium gene overexpression reveals distinct perturbation of sporozoite transmission by profilin Yuko Sato a,b, *, Marion Hliscs a,c, Josefine Dunst a,d, Christian Goosmann
THE ROLE OF RHOMBOID PROTEASES AND A OOCYST CAPSULE PROTEIN IN MALARIA PATHOGENESIS AND PARASITE DEVELOPMENT PRAKASH SRINIVASAN
 THE ROLE OF RHOMBOID PROTEASES AND A OOCYST CAPSULE PROTEIN IN MALARIA PATHOGENESIS AND PARASITE DEVELOPMENT BY PRAKASH SRINIVASAN Submitted in partial fulfillment of the requirements For the degree of
THE ROLE OF RHOMBOID PROTEASES AND A OOCYST CAPSULE PROTEIN IN MALARIA PATHOGENESIS AND PARASITE DEVELOPMENT BY PRAKASH SRINIVASAN Submitted in partial fulfillment of the requirements For the degree of
Diagnosis, treatment and control: dealing with coccidiosis in cattle
 Vet Times The website for the veterinary profession https://www.vettimes.co.uk Diagnosis, treatment and control: dealing with coccidiosis in cattle Author : Adam Martin Categories : Vets Date : January
Vet Times The website for the veterinary profession https://www.vettimes.co.uk Diagnosis, treatment and control: dealing with coccidiosis in cattle Author : Adam Martin Categories : Vets Date : January
Protozoan parasites of the genus Plasmodium are the causative
 Exploring the transcriptome of the malaria sporozoite stage Stefan H. I. Kappe*, Malcolm J. Gardner, Stuart M. Brown, Jessica Ross*, Kai Matuschewski*, Jose M. Ribeiro, John H. Adams, John Quackenbush,
Exploring the transcriptome of the malaria sporozoite stage Stefan H. I. Kappe*, Malcolm J. Gardner, Stuart M. Brown, Jessica Ross*, Kai Matuschewski*, Jose M. Ribeiro, John H. Adams, John Quackenbush,
Malaria Sporozoites Traverse Host Cells within Transient Vacuoles
 Malaria Sporozoites Traverse Host Cells within Transient Vacuoles Veronica Risco-Castillo, Selma Topçu, Carine Marinach, Giulia Manzoni, Améliee. Bigorgne, Sylvie Briquet, Xavier Baudin, Maryse Lebrun,
Malaria Sporozoites Traverse Host Cells within Transient Vacuoles Veronica Risco-Castillo, Selma Topçu, Carine Marinach, Giulia Manzoni, Améliee. Bigorgne, Sylvie Briquet, Xavier Baudin, Maryse Lebrun,
Apicomplexans Apicomplexa Intro
 Apicomplexans Apicomplexa Intro Cryptosporidium Apicomplexan Select Characteristics Gliding motility Apical Complex organelle for invasion of host cell Life cycle alternates b/w sexual and asexual phases
Apicomplexans Apicomplexa Intro Cryptosporidium Apicomplexan Select Characteristics Gliding motility Apical Complex organelle for invasion of host cell Life cycle alternates b/w sexual and asexual phases
Exposure of Plasmodium sporozoites to the intracellular concentration of potassium enhances infectivity and reduces cell passage activity
 Molecular & Biochemical Parasitology 156 (2007) 32 40 Exposure of Plasmodium sporozoites to the intracellular concentration of potassium enhances infectivity and reduces cell passage activity Kota Arun
Molecular & Biochemical Parasitology 156 (2007) 32 40 Exposure of Plasmodium sporozoites to the intracellular concentration of potassium enhances infectivity and reduces cell passage activity Kota Arun
The color and patterning of pigmentation in cats, dogs, mice horses and other mammals results from the interaction of several different genes
 The color and patterning of pigmentation in cats, dogs, mice horses and other mammals results from the interaction of several different genes 1 Gene Interactions: Specific alleles of one gene mask or modify
The color and patterning of pigmentation in cats, dogs, mice horses and other mammals results from the interaction of several different genes 1 Gene Interactions: Specific alleles of one gene mask or modify
Developmentally Regulated!nfectivity of Malaria Sporozoites for Mosquito Salivary Glands and the Vertebrate Host
 Developmentally Regulated!nfectivity of Malaria Sporozoites for Mosquito Salivary Glands and the Vertebrate Host By Musa G. Touray, Alon Warburg, Andre Laughinghouse, Antoniana U. Krettli,* and Louis H.
Developmentally Regulated!nfectivity of Malaria Sporozoites for Mosquito Salivary Glands and the Vertebrate Host By Musa G. Touray, Alon Warburg, Andre Laughinghouse, Antoniana U. Krettli,* and Louis H.
Parasitology Amoebas. Sarcodina. Mastigophora
 Parasitology Amoebas Sarcodina Entamoeba hisolytica (histo = tissue, lytica = lyse or break) (pathogenic form) o Trophozoite is the feeding form o Life Cycle: personfeces cyst with 4 nuclei with thicker
Parasitology Amoebas Sarcodina Entamoeba hisolytica (histo = tissue, lytica = lyse or break) (pathogenic form) o Trophozoite is the feeding form o Life Cycle: personfeces cyst with 4 nuclei with thicker
Department of Immunology and Infectious Diseases, Harvard School of Public Health, Boston, Massachusetts, USA, and 2
 scientific report scientificreport A mitogen-activated protein kinase regulates male gametogenesis and transmission of the malaria parasite Plasmodium berghei Radha Rangarajan 1,AmyK.Bei 1, Deepa Jethwaney
scientific report scientificreport A mitogen-activated protein kinase regulates male gametogenesis and transmission of the malaria parasite Plasmodium berghei Radha Rangarajan 1,AmyK.Bei 1, Deepa Jethwaney
Cyclic GMP Balance Is Critical for Malaria Parasite Transmission from the Mosquito to the Mammalian Host
 RESEARCH ARTICLE crossmark Cyclic GMP Balance Is Critical for Malaria Parasite Transmission from the Mosquito to the Mammalian Host Viswanathan Lakshmanan, a Matthew E. Fishbaugher, a Bob Morrison, a Michael
RESEARCH ARTICLE crossmark Cyclic GMP Balance Is Critical for Malaria Parasite Transmission from the Mosquito to the Mammalian Host Viswanathan Lakshmanan, a Matthew E. Fishbaugher, a Bob Morrison, a Michael
CIRCUMSPOROZOITE PROTEINS OF HUMAN MALARIA PARASITES PLASMODIUM FALCIPARUM AND PLASMODIUM VIVA,F*
 CIRCUMSPOROZOITE PROTEINS OF HUMAN MALARIA PARASITES PLASMODIUM FALCIPARUM AND PLASMODIUM VIVA,F* BY ELIZABETH H. NARDIN, VICTOR NUSSENZWEIG, RUTH S. NUSSENZWEIG, WILLIAM E. COLLINS, K. TRANAKCHIT HARINASUTA,
CIRCUMSPOROZOITE PROTEINS OF HUMAN MALARIA PARASITES PLASMODIUM FALCIPARUM AND PLASMODIUM VIVA,F* BY ELIZABETH H. NARDIN, VICTOR NUSSENZWEIG, RUTH S. NUSSENZWEIG, WILLIAM E. COLLINS, K. TRANAKCHIT HARINASUTA,
Transmission success of the malaria parasite Plasmodium mexicanum into its vector: role of gametocyte density and sex ratio
 Transmission success of the malaria parasite Plasmodium mexicanum into its vector: role of gametocyte density and sex ratio 575 J. J. SCHALL* Department of Biology, University of Vermont, Burlington, Vermont
Transmission success of the malaria parasite Plasmodium mexicanum into its vector: role of gametocyte density and sex ratio 575 J. J. SCHALL* Department of Biology, University of Vermont, Burlington, Vermont
Malaria remains the most important parasitic disease. Review Article
 Review Article Pre-erythrocytic Stage of Malaria Infection and the Molecular Targets Available for Interventions Dickson Adah 1,2, Yi Jun Yang 1,2, Quan Liu 1,2, Limei Qin 1, Li Qin 1, Xiaoping Chen 1
Review Article Pre-erythrocytic Stage of Malaria Infection and the Molecular Targets Available for Interventions Dickson Adah 1,2, Yi Jun Yang 1,2, Quan Liu 1,2, Limei Qin 1, Li Qin 1, Xiaoping Chen 1
Malaria parasites of rodents of the Congo (Brazzaville) :
 :") Annales de Parasitologie (Paris), 1976, t. 51, n 6, pp. 637 à 646 Malaria parasites of rodents of the Congo (Brazzaville) : Plasmodium cbabaudi adami subsp. nov. and Plasmodium vinckei lentum Landau, Michel,
Annales de Parasitologie (Paris), 1976, t. 51, n 6, pp. 637 à 646 Malaria parasites of rodents of the Congo (Brazzaville) : Plasmodium cbabaudi adami subsp. nov. and Plasmodium vinckei lentum Landau, Michel,
BioSci 110, Fall 08 Exam 2
 1. is the cell division process that results in the production of a. mitosis; 2 gametes b. meiosis; 2 gametes c. meiosis; 2 somatic (body) cells d. mitosis; 4 somatic (body) cells e. *meiosis; 4 gametes
1. is the cell division process that results in the production of a. mitosis; 2 gametes b. meiosis; 2 gametes c. meiosis; 2 somatic (body) cells d. mitosis; 4 somatic (body) cells e. *meiosis; 4 gametes
Biotecnologicas (IIB-INTECH), Universidad Nacional de San Martin, Av. General Paz 5445, Predio INTI, edificio 24 (1650), Buenos Aires, Argentina
, Universidad Nacional de San Martin, Av. General Paz 5445, Predio INTI, edificio 24 (1650), Buenos Aires, Argentina") [Frontiers in Bioscience 17, 726-744, January 1, 2012] Plasmodium sporozoite motility: an update Georgina N. Montagna 1, Kai Matuschewski 1, Carlos A. Buscaglia 2 1 Parasitology Unit, Max Planck Institute
[Frontiers in Bioscience 17, 726-744, January 1, 2012] Plasmodium sporozoite motility: an update Georgina N. Montagna 1, Kai Matuschewski 1, Carlos A. Buscaglia 2 1 Parasitology Unit, Max Planck Institute
Molecular diagnosis of Theileria infections in wildlife from Southern Africa ~ implications for accurate diagnosis.
 Molecular diagnosis of Theileria infections in wildlife from Southern Africa ~ implications for accurate diagnosis. Ronel Pienaar Parasites Vectors and Vector-borne Diseases Onderstepoort Veterinary Institute
Molecular diagnosis of Theileria infections in wildlife from Southern Africa ~ implications for accurate diagnosis. Ronel Pienaar Parasites Vectors and Vector-borne Diseases Onderstepoort Veterinary Institute
Plasmodium Pre-Erythrocytic Stages: Biology, Whole Parasite Vaccines and Transgenic Models
 American Journal of Immunology, 2012, 8 (3), 88-100 ISSN 1553-619X 2012 Science Publication doi:10.3844/ajisp.2012.88.100 Published Online 8 (3) 2012 (http://www.thescipub.com/aji.toc) Plasmodium Pre-Erythrocytic
American Journal of Immunology, 2012, 8 (3), 88-100 ISSN 1553-619X 2012 Science Publication doi:10.3844/ajisp.2012.88.100 Published Online 8 (3) 2012 (http://www.thescipub.com/aji.toc) Plasmodium Pre-Erythrocytic
Was the Spotted Horse an Imaginary Creature? g.org/sciencenow/2011/11/was-the-spotted-horse-an-imagina.html
 Was the Spotted Horse an Imaginary Creature? http://news.sciencema g.org/sciencenow/2011/11/was-the-spotted-horse-an-imagina.html 1 Genotypes of predomestic horses match phenotypes painted in Paleolithic
Was the Spotted Horse an Imaginary Creature? http://news.sciencema g.org/sciencenow/2011/11/was-the-spotted-horse-an-imagina.html 1 Genotypes of predomestic horses match phenotypes painted in Paleolithic
Anti-tick vaccines: A potential tool for control of the blacklegged ticks and other ticks feeding on whitetailed deer
 Anti-tick vaccines: A potential tool for control of the blacklegged ticks and other ticks feeding on whitetailed deer Andrew Y. Li USDA-ARS Invasive Insect Biocontrol and Behavior Laboratory (IIBBL) Beltsville,
Anti-tick vaccines: A potential tool for control of the blacklegged ticks and other ticks feeding on whitetailed deer Andrew Y. Li USDA-ARS Invasive Insect Biocontrol and Behavior Laboratory (IIBBL) Beltsville,
Reverse genetics screen identifies six proteins important for malaria development in the mosquito
 Molecular Microbiology (2008) 70(1), 209220 doi:.1111/j.1365-2958.2008.06407.x First published online 27 August 2008 Reverse genetics screen identifies six proteins important for malaria development in
Molecular Microbiology (2008) 70(1), 209220 doi:.1111/j.1365-2958.2008.06407.x First published online 27 August 2008 Reverse genetics screen identifies six proteins important for malaria development in
Evaluation of the hair growth and retention activity of two solutions on human hair explants
 activity of two solutions on human hair explants Study Directed by Dr E. Lati of Laboratoire Bio-EC, Centre de Recherches Biologiques et d Experimentations Cutanees, on behalf of Pangaea Laboratories Ltd.
activity of two solutions on human hair explants Study Directed by Dr E. Lati of Laboratoire Bio-EC, Centre de Recherches Biologiques et d Experimentations Cutanees, on behalf of Pangaea Laboratories Ltd.
Parasites of Small Mammals in Grand Teton National Park: Babesia and Hepatozoon
 University of Wyoming National Park Service Research Center Annual Report Volume 19 19th Annual Report, 1995 Article 13 1-1-1995 Parasites of Small Mammals in Grand Teton National Park: Babesia and Hepatozoon
University of Wyoming National Park Service Research Center Annual Report Volume 19 19th Annual Report, 1995 Article 13 1-1-1995 Parasites of Small Mammals in Grand Teton National Park: Babesia and Hepatozoon
Systemic Apicomplexans. Toxoplasma
 Systemic Apicomplexans Toxoplasma Protozoan Groups Historically, protozoa have been grouped by mode of motility. Flagellates Hemoflagellates Trypanosoma cruzi Leishmania infantum Mucoflagellates Tritrichomonas
Systemic Apicomplexans Toxoplasma Protozoan Groups Historically, protozoa have been grouped by mode of motility. Flagellates Hemoflagellates Trypanosoma cruzi Leishmania infantum Mucoflagellates Tritrichomonas
Developmental expression of synthetic cis-regulatory systems composed of spatial control elements from two different genes
 Proc. Natl. Acad. Sci. USA Vol. 93, pp. 13849 13854, November 1996 Developmental Biology Developmental expression of synthetic cis-regulatory systems composed of spatial control elements from two different
Proc. Natl. Acad. Sci. USA Vol. 93, pp. 13849 13854, November 1996 Developmental Biology Developmental expression of synthetic cis-regulatory systems composed of spatial control elements from two different
A role for apical membrane antigen 1 during invasion of hepatocytes
 JBC Papers in Press. Published on December 15, 2003 as Manuscript M311331200 A role for apical membrane antigen 1 during invasion of hepatocytes by Plasmodium falciparum sporozoites. Olivier Silvie a,
JBC Papers in Press. Published on December 15, 2003 as Manuscript M311331200 A role for apical membrane antigen 1 during invasion of hepatocytes by Plasmodium falciparum sporozoites. Olivier Silvie a,
PCR detection of Leptospira in. stray cat and
 PCR detection of Leptospira in 1 Department of Pathology, School of Veterinary Medicine, Islamic Azad University, Shahrekord Branch, Shahrekord, Iran 2 Department of Microbiology, School of Veterinary
PCR detection of Leptospira in 1 Department of Pathology, School of Veterinary Medicine, Islamic Azad University, Shahrekord Branch, Shahrekord, Iran 2 Department of Microbiology, School of Veterinary
Diurnal variation in microfilaremia in cats experimentally infected with larvae of
 Hayasaki et al., Page 1 Short Communication Diurnal variation in microfilaremia in cats experimentally infected with larvae of Dirofilaria immitis M. Hayasaki a,*, J. Okajima b, K.H. Song a, K. Shiramizu
Hayasaki et al., Page 1 Short Communication Diurnal variation in microfilaremia in cats experimentally infected with larvae of Dirofilaria immitis M. Hayasaki a,*, J. Okajima b, K.H. Song a, K. Shiramizu
NOTES. Received 28 August 2001/Returned for modification 17 October 2001/Accepted 3 December 2001
 INFECTION AND IMMUNITY, Mar. 2002, p. 1599 1603 Vol. 70, No. 3 0019-9567/02/$04.00 0 DOI: 10.1128/IAI.70.3.1599 1603.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. NOTES Babesia
INFECTION AND IMMUNITY, Mar. 2002, p. 1599 1603 Vol. 70, No. 3 0019-9567/02/$04.00 0 DOI: 10.1128/IAI.70.3.1599 1603.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. NOTES Babesia
23 Plasmodium coatneyi Eyles, Fong, Warren, Guinn, Sandosham, and Wharton, 1962
 23 Plasmodium coatneyi Eyles, Fong, Warren, Guinn, Sandosham, and Wharton, 1962 IN the course of studies on simian malaria begun by the late Dr. Don Eyles in Malaya, he and his co-workers isolated a new
23 Plasmodium coatneyi Eyles, Fong, Warren, Guinn, Sandosham, and Wharton, 1962 IN the course of studies on simian malaria begun by the late Dr. Don Eyles in Malaya, he and his co-workers isolated a new
Corallopyronin A: a new anti-filarial drug. Kenneth Pfarr Institute for Medical Microbiology, Immunology and Parasitology
 Corallopyronin A: a new anti-filarial drug Kenneth Pfarr Institute for Medical Microbiology, Immunology and Parasitology PEG, Weimar, 17 th October, 2014 Filariasis ~150 million people infected >1.3 billion
Corallopyronin A: a new anti-filarial drug Kenneth Pfarr Institute for Medical Microbiology, Immunology and Parasitology PEG, Weimar, 17 th October, 2014 Filariasis ~150 million people infected >1.3 billion
both are fatal diseases. In babesiosis blood comes out with the urine and hence it is also known as Red water disease. Theileria vaccines are not
 1.1 INTRODUCTION Animal husbandry plays an important role in Indian agriculture. Indians by large are vegetarian and as such the only source of animal protein is milk and milk products. With the increasing
1.1 INTRODUCTION Animal husbandry plays an important role in Indian agriculture. Indians by large are vegetarian and as such the only source of animal protein is milk and milk products. With the increasing
Protozoa. Apicomplexa Sarcomastigophora Ciliophora. Gregarinea Coccidia Piroplasma
 Protozoa Apicomplexa Sarcomastigophora Ciliophora Gregarinea Coccidia Piroplasma Coccidia characterized by thick-walled oocysts excreted in feces In Humans Cryptosporidium Isospora Cyclospora Sarcocystis
Protozoa Apicomplexa Sarcomastigophora Ciliophora Gregarinea Coccidia Piroplasma Coccidia characterized by thick-walled oocysts excreted in feces In Humans Cryptosporidium Isospora Cyclospora Sarcocystis
HISTOPATHOLOGY. Introduction:
 Introduction: HISTOPATHOLOGY Goats and sheep are the major domestic animal species in India. Much of the economy of the country has been depend upon the domestication of these animals. Especially economy
Introduction: HISTOPATHOLOGY Goats and sheep are the major domestic animal species in India. Much of the economy of the country has been depend upon the domestication of these animals. Especially economy
UNDERSTANDING THE TRANSMISSION OF TICK-BORNE PATHOGENS WITH PUBLIC HEALTH IMPLICATIONS
 UNDERSTANDING THE TRANSMISSION OF TICK-BORNE PATHOGENS WITH PUBLIC HEALTH IMPLICATIONS A. Rick Alleman, DVM, PhD, DABVP, DACVP Lighthouse Veterinary Consultants, LLC Gainesville, FL Tick-transmitted pathogens
UNDERSTANDING THE TRANSMISSION OF TICK-BORNE PATHOGENS WITH PUBLIC HEALTH IMPLICATIONS A. Rick Alleman, DVM, PhD, DABVP, DACVP Lighthouse Veterinary Consultants, LLC Gainesville, FL Tick-transmitted pathogens
Coccidia. Nimit Morakote, Ph.D.
 Coccidia Nimit Morakote, Ph.D. 1 Learning objectives After class, students will be able to: Describe morphology, life cycle, signs and symptoms, prevention and control, laboratory diagnosis and treatment
Coccidia Nimit Morakote, Ph.D. 1 Learning objectives After class, students will be able to: Describe morphology, life cycle, signs and symptoms, prevention and control, laboratory diagnosis and treatment
Running title: Model to down-select human malaria vaccines
 CVI Accepts, published online ahead of print on 27 March 2013 Clin. Vaccine Immunol. doi:10.1128/cvi.00066-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10
CVI Accepts, published online ahead of print on 27 March 2013 Clin. Vaccine Immunol. doi:10.1128/cvi.00066-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10
Genome 371; A 03 Berg/Brewer Practice Exam I; Wednesday, Oct 15, PRACTICE EXAM GENOME 371 Autumn 2003
 PRACTICE EXAM GENOME 371 Autumn 2003 These questions were part of the first exam from Autumn 2002. Take the exam in a quiet place and only when you are sure you will have time to complete the exam uninterrupted.
PRACTICE EXAM GENOME 371 Autumn 2003 These questions were part of the first exam from Autumn 2002. Take the exam in a quiet place and only when you are sure you will have time to complete the exam uninterrupted.
Bi156 Lecture 1/13/12. Dog Genetics
 Bi156 Lecture 1/13/12 Dog Genetics The radiation of the family Canidae occurred about 100 million years ago. Dogs are most closely related to wolves, from which they diverged through domestication about
Bi156 Lecture 1/13/12 Dog Genetics The radiation of the family Canidae occurred about 100 million years ago. Dogs are most closely related to wolves, from which they diverged through domestication about
Novel ELISA method as exploratory tool to assess immunity induced by radiated attenuated sporozoites to decipher protective immunity
 DOI 10.1186/s12936-017-2129-9 Malaria Journal METHODOLOGY Open Access Novel ELISA method as exploratory tool to assess immunity induced by radiated attenuated sporozoites to decipher protective immunity
DOI 10.1186/s12936-017-2129-9 Malaria Journal METHODOLOGY Open Access Novel ELISA method as exploratory tool to assess immunity induced by radiated attenuated sporozoites to decipher protective immunity
Antimalarial Activity of Allicin, a Biologically Active Compound from Garlic Cloves
 ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, May 2006, p. 1731 1737 Vol. 50, No. 5 0066-4804/06/$08.00 0 doi:10.1128/aac.50.5.1731 1737.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved.
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, May 2006, p. 1731 1737 Vol. 50, No. 5 0066-4804/06/$08.00 0 doi:10.1128/aac.50.5.1731 1737.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved.
T Mike Lo 1,2 and Maureen Coetzee 1,2*
 Lo and Coetzee Parasites & Vectors 2013, 6:184 RESEARCH Open Access Marked biological differences between insecticide resistant and susceptible strains of Anopheles funestus infected with the murine parasite
Lo and Coetzee Parasites & Vectors 2013, 6:184 RESEARCH Open Access Marked biological differences between insecticide resistant and susceptible strains of Anopheles funestus infected with the murine parasite
A Role for Apical Membrane Antigen 1 during Invasion of Hepatocytes by Plasmodium falciparum Sporozoites*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 10, Issue of March 5, pp. 9490 9496, 2004 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. A Role for Apical
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 10, Issue of March 5, pp. 9490 9496, 2004 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. A Role for Apical
EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS WORK-PROGRAMME PROPOSAL Version 2 VISAVET. Universidad Complutense de Madrid
 FOR BOVINE TUBERCULOSIS WORK-PROGRAMME PROPOSAL Version 2 VISAVET. Universidad Complutense de Madrid") EUROPEAN COMMISSION HEALTH & CONSUMERS DIRECTORATE-GENERAL Directorate D Animal Health and Welfare Unit D1- Animal health and Standing Committees EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS
EUROPEAN COMMISSION HEALTH & CONSUMERS DIRECTORATE-GENERAL Directorate D Animal Health and Welfare Unit D1- Animal health and Standing Committees EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS
Neither Mosquito Saliva nor Immunity to Saliva Has a Detectable Effect on the Infectivity of Plasmodium Sporozoites Injected into Mice
 INFECTION AND IMMUNITY, Jan. 2010, p. 545 551 Vol. 78, No. 1 0019-9567/10/$12.00 doi:10.1128/iai.00807-09 Copyright 2010, American Society for Microbiology. All Rights Reserved. Neither Mosquito Saliva
INFECTION AND IMMUNITY, Jan. 2010, p. 545 551 Vol. 78, No. 1 0019-9567/10/$12.00 doi:10.1128/iai.00807-09 Copyright 2010, American Society for Microbiology. All Rights Reserved. Neither Mosquito Saliva
Phenotype Observed Expected (O-E) 2 (O-E) 2 /E dotted yellow solid yellow dotted blue solid blue
 2 (O-E) 2 /E dotted yellow solid yellow dotted blue solid blue") 1. (30 pts) A tropical fish breeder for the local pet store is interested in creating a new type of fancy tropical fish. She observes consistent patterns of inheritance for the following traits: P 1 :
1. (30 pts) A tropical fish breeder for the local pet store is interested in creating a new type of fancy tropical fish. She observes consistent patterns of inheritance for the following traits: P 1 :
The impact on the routine laboratory of the introduction of an automated ELISA for the detection of Cryptosporidium and Giardia in stool samples
 The impact on the routine laboratory of the introduction of an automated ELISA for the detection of Cryptosporidium and Giardia in stool samples Nigel Stephenson BMS 3 Department of Medical Microbiology
The impact on the routine laboratory of the introduction of an automated ELISA for the detection of Cryptosporidium and Giardia in stool samples Nigel Stephenson BMS 3 Department of Medical Microbiology
Alveolar proteins stabilize cortical microtubules in Toxoplasma gondii
 Alveolar proteins stabilize cortical microtubules in Toxoplasma gondii Clare R. Harding 1,*, Matthew Gow 2, Joon Ho Kang 3,, Emily Shortt 1, Scott R. Manalis,5,6, Markus Meissner 2,7, and Sebastian Lourido
Alveolar proteins stabilize cortical microtubules in Toxoplasma gondii Clare R. Harding 1,*, Matthew Gow 2, Joon Ho Kang 3,, Emily Shortt 1, Scott R. Manalis,5,6, Markus Meissner 2,7, and Sebastian Lourido
THE ABUNDANCE AND INFECTION STATUS OF ANOPHELES MOSQUITOES IN LOUDOUN COUNTY, VIRGINIA
 THE ABUNDANCE AND INFECTION STATUS OF ANOPHELES MOSQUITOES IN LOUDOUN COUNTY, VIRGINIA Andrew Lima Clarke (Manassas, VA) Priya Krishnan ODU M.S. candidate (Richmond, VA) Objectives To determine: 1) the
THE ABUNDANCE AND INFECTION STATUS OF ANOPHELES MOSQUITOES IN LOUDOUN COUNTY, VIRGINIA Andrew Lima Clarke (Manassas, VA) Priya Krishnan ODU M.S. candidate (Richmond, VA) Objectives To determine: 1) the
XXI. Malaria [MAL = bad; ARIA = air] (Chapter 9) 2008 A. Order Haemosporida, Family Plasmodiidae 1. Live in vertebrate tissues and blood 2.
![XXI. Malaria [MAL = bad; ARIA = air] (Chapter 9) 2008 A. Order Haemosporida, Family Plasmodiidae 1. Live in vertebrate tissues and blood 2.](/thumbs/80/81826026.jpg "XXI. Malaria [MAL = bad; ARIA = air] (Chapter 9) 2008 A. Order Haemosporida, Family Plasmodiidae 1. Live in vertebrate tissues and blood 2.") XXI. Malaria [MAL = bad; ARIA = air] (Chapter 9) 2008 A. Order Haemosporida, Family Plasmodiidae 1. Live in vertebrate tissues and blood 2. SCHIZOGONY (asexual reproduction) in vertebrates 3. SPOROGONY
XXI. Malaria [MAL = bad; ARIA = air] (Chapter 9) 2008 A. Order Haemosporida, Family Plasmodiidae 1. Live in vertebrate tissues and blood 2. SCHIZOGONY (asexual reproduction) in vertebrates 3. SPOROGONY
National Research Center
 National Research Center Update of immunodiagnosis of cystic echinococcosis cysts Global distribution of zoonotic strains of Echinococcus granulosus (Adapted from Eckert and Deplazes, 2004) Echinococcus
National Research Center Update of immunodiagnosis of cystic echinococcosis cysts Global distribution of zoonotic strains of Echinococcus granulosus (Adapted from Eckert and Deplazes, 2004) Echinococcus
INHERITANCE OF BODY WEIGHT IN DOMESTIC FOWL. Single Comb White Leghorn breeds of fowl and in their hybrids.
 440 GENETICS: N. F. WATERS PROC. N. A. S. and genetical behavior of this form is not incompatible with the segmental interchange theory of circle formation in Oenothera. Summary.-It is impossible for the
440 GENETICS: N. F. WATERS PROC. N. A. S. and genetical behavior of this form is not incompatible with the segmental interchange theory of circle formation in Oenothera. Summary.-It is impossible for the
Outline 1/13/15. Range is mostly surrounding Puerto Rico Important for Tourism and ecological balance
 1/13/15 Prevalence of Toxoplasma gondii in Antillean manatees (Trichechus manatus manatus) and investigating transmission from feral cat feces in Puerto Rico Heidi Wyrosdick M.S. Candidate University of
1/13/15 Prevalence of Toxoplasma gondii in Antillean manatees (Trichechus manatus manatus) and investigating transmission from feral cat feces in Puerto Rico Heidi Wyrosdick M.S. Candidate University of
FACULTY OF VETERINARY MEDICINE
 FACULTY OF VETERINARY MEDICINE DEPARTMENT OF VETERINARY PARASITOLOGY AND ENTOMOLOGY M.Sc. AND Ph.D. DEGREE PROGRAMMES The postgraduate programmes of the Department of Veterinary Parasitology and Entomology
FACULTY OF VETERINARY MEDICINE DEPARTMENT OF VETERINARY PARASITOLOGY AND ENTOMOLOGY M.Sc. AND Ph.D. DEGREE PROGRAMMES The postgraduate programmes of the Department of Veterinary Parasitology and Entomology
Malaria parasite exit from the host erythrocyte: A two-step process requiring extraerythrocytic proteolysis
 Malaria parasite exit from the host erythrocyte: A two-step process requiring extraerythrocytic proteolysis Brandy L. Salmon, Anna Oksman, and Daniel E. Goldberg* Howard Hughes Medical Institute, Departments
Malaria parasite exit from the host erythrocyte: A two-step process requiring extraerythrocytic proteolysis Brandy L. Salmon, Anna Oksman, and Daniel E. Goldberg* Howard Hughes Medical Institute, Departments
COMMITTEE FOR VETERINARY MEDICINAL PRODUCTS
 The European Agency for the Evaluation of Medicinal Products Veterinary Medicines and Information Technology EMEA/CVMP/005/00-FINAL-Rev.1 COMMITTEE FOR VETERINARY MEDICINAL PRODUCTS GUIDELINE FOR THE TESTING
The European Agency for the Evaluation of Medicinal Products Veterinary Medicines and Information Technology EMEA/CVMP/005/00-FINAL-Rev.1 COMMITTEE FOR VETERINARY MEDICINAL PRODUCTS GUIDELINE FOR THE TESTING
ELISA assays for parasitic and tick-borne diseases
 ELISA assays for parasitic and tick-borne diseases We are passionate about the health and well-being of humans and animals. Immunodiagnostics from contribute to a global, adequate supply of safe and nutritious
ELISA assays for parasitic and tick-borne diseases We are passionate about the health and well-being of humans and animals. Immunodiagnostics from contribute to a global, adequate supply of safe and nutritious