Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui
|
|
|
- Loreen Avice Whitehead
- 6 years ago
- Views:
Transcription
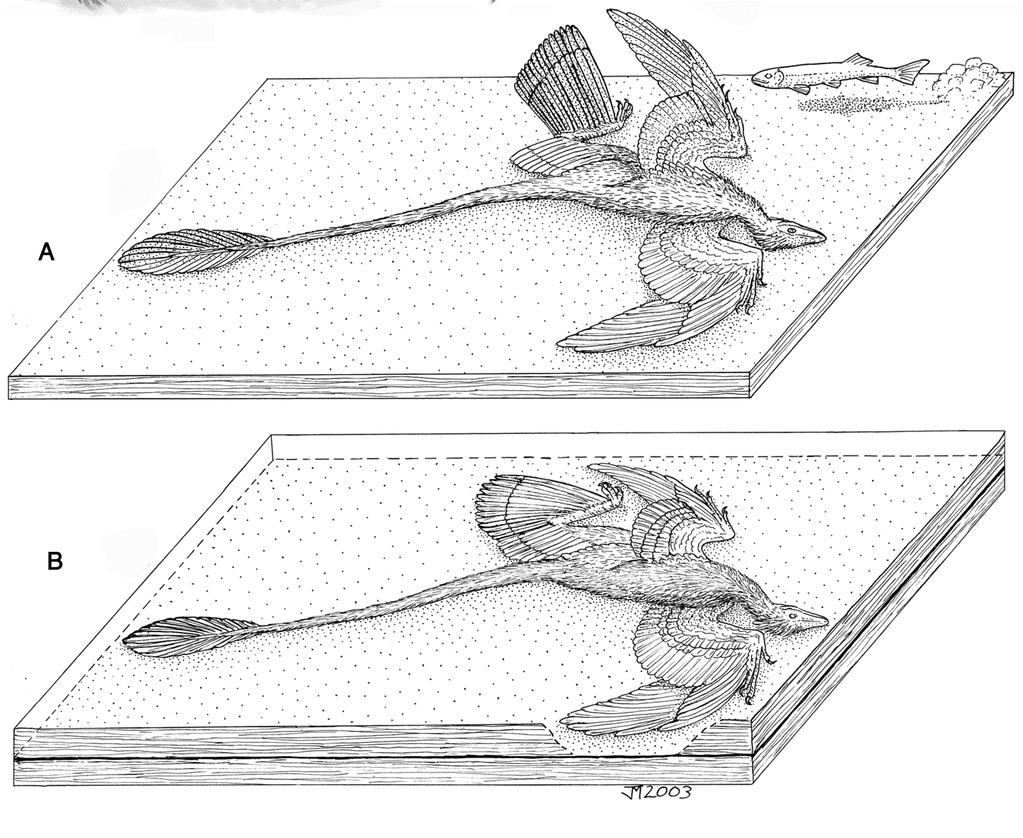
1 Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui Sankar Chatterjee* and R. Jack Templin *Department of Geosciences, Museum of Texas Tech University, P.O. Box 43191, Lubbock, TX ; and 2212 Aster Street, Ottawa, ON, Canada K1H 6R6 Communicated by Lynn Margulis, University of Massachusetts, Amherst, MA, November 17, 2006 (received for review October 24, 2005) Microraptor gui, a four-winged dromaeosaur from the Early Cretaceous of China, provides strong evidence for an arboreal-gliding origin of avian flight. It possessed asymmetric flight feathers not only on the manus but also on the pes. A previously published reconstruction shows that the hindwing of Microraptor supported by a laterally extended leg would have formed a second pair of wings in tetrapteryx fashion. However, this wing design conflicts with known theropod limb joints that entail a parasagittal posture of the hindlimb. Here, we offer an alternative planform of the hindwing of Microraptor that is concordant with its feather orientation for producing lift and normal theropod hindlimb posture. In this reconstruction, the wings of Microraptor could have resembled a staggered biplane configuration during flight, where the forewing formed the dorsal wing and the metatarsal wing formed the ventral one. The contour feathers on the tibia were positioned posteriorly, oriented in a vertical plane for streamlining that would reduce the drag considerably. Leg feathers are present in many fossil dromaeosaurs, early birds, and living raptors, and they play an important role in flight during catching and carrying prey. A computer simulation of the flight performance of Microraptor suggests that its biplane wings were adapted for undulatory phugoid gliding between trees, where the horizontal feathered tail offered additional lift and stability and controlled pitch. Like the Wright 1903 Flyer, Microraptor, a gliding relative of early birds, took to the air with two sets of wings. arboreal origin of flight Chinese feathered dinosaurs phugoid gliding Wright 1903 Flyer The evolution of powered flight in birds from theropod dinosaurs is recognized as the key adaptive breakthrough that contributed to the biological success of this group. The transformation of wing design from nonavian dinosaurs to early birds is beginning to unravel in recent times from a wealth of fossil record from China. Hundreds of small, exquisitely preserved, feathered theropods were discovered in the Early Cretaceous Jehol Group of northeastern China as they died some 125 million years ago, smothered in the Cretaceous Pompeii. Both anatomy and phylogeny strongly suggest that these theropods, including Sinosauropteryx (1), Caudipteryx and Protarchaeopteryx (2), Microraptor (3), Sinornithosaurus (4), Cryptovolans (5), and the early bird Confuciusornis (6), show constructions ranging from small winged, arboreal theropods to fully winged, active flying birds. They offer new insights into the origins of feathers and flight, favoring the arboreal ( trees-down ) over the cursorial ( ground-up ) hypothesis (3 5, 7 11). Among these recent finds, Microraptor gui offers the best evidence that arboreal dromaeosaurs might have acquired powered flight through a gliding stage where both forelimbs and hindlimbs were involved (12). With two sets of winged limbs, each having long, asymmetric flight feathers at the distal segments of both forelimb and hindlimb, Microraptor developed broad airfoil surfaces and was probably an efficient glider. There are 12 manual and 14 pedal primary feathers, which are long and asymmetric to create aerodynamic forces for lift. The longest primary on the metatarsus would be 19 cm (Fig. 1A). The hooked, interlocking barbs gave strength and flexibility to the asymmetric feathers and prevented air from passing through it in flight (Fig. 1B). In the proximal part of the wing and hindleg, the contour feathers have symmetric vanes (Fig. 1C). A typical contour feather is composed of a long, tapering central rachis with a broad, flexible vane on either side. Vanes are asymmetrical in flight feathers in relation to the central rachis, where the leading edge is narrower and stronger than the trailing edge. This asymmetry provides an airfoil-shaped crosssection of the feather because air pressure is greater along the leading edge. The aerodynamic function of the asymmetric feather is attributable to aeroelastic stability in the lifting feather. The forward location of the rachis suppresses a tendency for the rachis to twist elastically in response to the aerodynamic force on the lifting feather because lift is concentrated in the forward quarter of the feather s area. The asymmetry is a sure sign that the feather has been adapted for lifting. Xu et al. (12) (Fig. 1C) reconstructed limbs of four-winged Microraptor as tandem wings similar to those of insects and gliding fish, where all wings are spread horizontally in tetrapteryx fashion. They argued that Microraptor was clumsy on the ground; because it had these long feathers on its feet, it would have had trouble walking or running and would have been vulnerable on the ground. It was probably a tree-dweller, thus supporting the arboreal theory of flight, where gravity was the main source of flying energy. The holotype specimen of Microraptor, with an estimated live weight of 1 kg and measuring 77 cm in length, has a long bony tail that bears asymmetric retrices on two sides that could provide additional lift and control pitch (Table 1). However, Xu et al. (12) did not discuss why Microraptor was a glider and how it used its wings during flight. The life restoration of the hindlimb of Microraptor in a laterally extended position by Xu et al. (12) (Fig. 1C) appears to be aerodynamically inefficient and so anatomically anomalous that it generated lively debate and speculation (13). In our view, the leading edge of the asymmetric flight feathers on the metatarsus should face forward against the direction of the airflow like those of the hand section, not sidewise, as they reconstructed. In all theropods (including birds), the hindlimb is held in an erect and parasagittal gait, unlike the reconstruction of Microraptor in a laterally extended position. Because of the critical importance of this fossil in the early evolutionary history of avian flight, we propose here a second restoration of the wing planform and estimate the flight Author contributions: S.C. designed research; S.C. and R.J.T. performed research; R.J.T. contributed new reagents/analytic tools; S.C. and R.J.T. analyzed data; and S.C. and R.J.T. wrote the paper. The authors declare no conflict of interest. Freely available online through the PNAS open access option. Abbreviation: M, mass. To whom correspondence should be addressed. sankar.chatterjee@ttu.edu. This article contains supporting information online at /DC by The National Academy of Sciences of the USA PNAS January 30, 2007 vol. 104 no. 5 cgi doi pnas
![Fig. 1. Feathers of M. gui. (A) Holotype of M. gui [Institute of Vertebrate Paleontology and Paleoanthropology (IVPP) V13352] as preserved [modified from Xu et al. (12)]. (Scale bar, 5 cm.](/docs-images/80/81292113/images/2-0.jpg ") (B) The long feathers on the hand and metatarsal sections had evolved for flight; they were asymmetric with interlocking barbules.")
2 Fig. 1. Feathers of M. gui. (A) Holotype of M. gui [Institute of Vertebrate Paleontology and Paleoanthropology (IVPP) V13352] as preserved [modified from Xu et al. (12)]. (Scale bar, 5 cm.) (B) The long feathers on the hand and metatarsal sections had evolved for flight; they were asymmetric with interlocking barbules. (C) In the rest of the wing and hindleg, the feathers are symmetric (10). performance of Microraptor in a manner fundamentally different from the conclusions reached by Xu et al. (12). Hindlimb Posture and Orientation of Metatarsal Feathers In all theropods (including birds), the hip, knee, and ankle joints are stable and fully congruent during parasagittal motion of the hindlimbs, permitting a wide range of flexion and extension but little abduction and adduction. The femur head is cylindrical, fitting into a perforated acetabulum, which allows little transverse deviation from the parasagittal plane (14). The hip joint becomes quickly incongruent as the femur is abducted horizontally from the parasagittal plane. The parasagittal hindlimb posture of Microraptor is beautifully preserved in the holotype specimen (Fig. 1A), which differs strikingly from the horizontal restoration (12) but closely resembles that of the Berlin Archaeopteryx specimen, which also displays tibial feathers (15) (see Fig. 4). The most unusual feature in Microraptor is the presence of long, asymmetric flight feathers on the entire length of the metatarsus, which are unknown in Archaeopteryx, feathered dromaeosaurs, and modern birds. The orientation of the metatarsal feathers as reconstructed by Xu et al. (12) is problematic because these feathers extend below the level of the feet, thus hindering terrestrial locomotion. In this reconstruction, the leading edge of these primary feathers on the metatarsus would face sidewise during gliding without producing any lift. Because they are asymmetric flight feathers like those of the forewings, the narrow, leading edge should face forward against the direction of airflow to gain lift. We suggest that these feathers were oriented in life in a transverse horizontal plane like those of the hand section, to be an effective airfoil during flight, but that they collapsed backward during fossilization. The symmetrical tibial feathers also are preserved in a backward fashion. In life, these contour feathers should be projected posteriorly for streamlining. Three biological and aerodynamic constraints provide important clues to the hindwing design of Microraptor:(i) The hindlimb should be oriented in a parasagittal plane as in all theropods Chatterjee, S., Templin, R. J. (2005) Geol Soc Am Abstr Prog 37:88 (abstr.). (including birds) and could not be splayed sidewise. (ii) The hindlimb wings should be folded neatly into a compact package during walking in such a fashion that metatarsal feathers would not project ventrally beyond the foot to prevent damage to the feathers. (iii) The leading edge of the primary feathers on the metatarsus should face forward as in the manus; this arrangement makes each feather capable of acting as an individual airfoil and also is needed to maintain the entire wing s camber and to carry the aerodynamic load. The first two constraints contradict the original hindlimb reconstruction of Xu et al. (12) (Fig. 1C). We present several possible hindlimb orientations (Fig. 2 A D) in Microraptor during flight. We have discarded the bird-like pedal configuration in flight (Fig. 2A) because in that pose, the primaries would face backward to produce lift in an unproductive direction (ventrally) that would interfere with gliding. Among other alternatives, we prefer Fig. 2D, a reconstruction in which the hindlimb is held in a z-fashion resembling the pouncing posture of modern raptors when catching prey from the air and carrying it (Fig. 2H). In this pose, the femur would be kept in a subhorizontal position close to the body, directed forward, with its feathers oriented backward and merging with the body contour to form a continuous surface, allowing this airfoil to move smoothly through air with the least drag. We propose that the symmetric feathers on the tibia were arranged like those in modern raptors, streamlining the circular shaft of the tibia by stretching it backward to maintain a smooth flow of air (Fig. 2 E G). Without tibial streamlining, the cylindrical leading edge of the vertical tibia could increase the total drag by nearly 40%. Biplane Wing Configuration Because Microraptor could not extend its hindwings directly behind the forewings in the same plane, it probably held its feet lower than its arms, a more anatomically and aerodynamically stable configuration. Once the parasagittal posture and feather orientation of the hindlimb are corrected based on both anatomical and aerodynamic modeling, the wings of Microraptor resemble those of a staggered biplane from the side, where the forewing forms the dorsal wing and the metatarsal wing forms EVOLUTION Table 1. Aerodynamic data of M. gui, Nyctosaurus gracilis, and F. magnificens Specimen (holotype) M, kg Wing area (forewing plus hindwing), S (m 2 ) Tailwing area, S(m 2 ) Wingspan, b (m) Aspect ratio, A b 2 /S WL, mg/s (N/m 2 ) Gliding speed (m/s) Microraptor IVPP V13352 Nyctosaurus YPM Fregata M, mass; N, Newton; S, wing area; WL, wing loading; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology; YPM, Yale Peabody Museum. Chatterjee and Templin PNAS January 30, 2007 vol. 104 no
3 Fig. 3. Flight performance of Microraptor. (A) Power curves (steady level flight) for Microraptor. The horizontal line represents the estimated maximum continuous power available. Two curves are shown for the level flight power required. Curve 1 is from streamtube theory (20), and curve 2 is based on the simpler aircraft theory (22). They converge for speeds of greater than 6 m/s. (B) Glide polars for Microraptor, compared with a seabird (frigatebird, M 1.5 kg) and a pterosaur (Nyctosaurus,M 1.85 kg) (ref. 22; see Table 1 for aerodynamic data). (C) Glide paths of Microraptor from a perch. Curve 1 shows phugoid gliding. Curve 2 shows a final rapid pitchup with high drag. Curve 3 shows gliding path with pitch damper on. Curve 4 shows a parachuting trajectory. the ventral one (Fig. 2 I and J). The metatarsal wing of Microraptor is unique among vertebrates and needs further comment. In the z-shaped orientation of the hindlimb, the feathers on the metatarsus become horizontal, laterally extended, and form the ventral wing. The ventral wing tilts slightly upward from the horizontal position, allowing the lower wing to have a few degrees greater incidence (upward angle of attack) than the upper wing, known as decalage in biplane theory (an unstable situation, which is compensated by the long tail) (16) Fig. 2. Wing planform of Microraptor. (A D) Different possible hindlimb postures during flight. (A) Hindlimb backwardly directed as in modern birds. (B D) Biplane configuration. (B) Hindlimb backwardly sloping position. (C) Hindlimb forwardly sloping in predatory strike position. (D) Hindlimb in z-fashion with a body silhouette showing the animal in lateral view with an upwardly tilted tail for pitch control. (E) Cross-section of the tibia fibula showing a streamlining and stretching effect of the cylindrical tibia by adding feathers caudally. (F) Cylindrical structure offers maximum resistance to the airstream as the airflow behind it becomes broken up into eddies, creating turbulence. (G) Filling the spaces in a cylindrical structure in front and behind improves streamlining, as in the case of the feathered tibia of raptors. (H) Pouncing posture of a raptor, Falco.(I) A typical staggered biplane (Stearman 75) for comparison with Microraptor; in biplane aircraft of the 1920s, there was a large additional drag of wires, struts, etc. between the two wings, which eventually made the biplane obsolete except for a niche application; such drag-induced structures were absent in Microraptor.(J) Life reconstruction of M. gui (IVPP V13352) in dorsal view showing the morphology and distribution of hindlimb feathers (Left) and orientation of the hindlimb (Right) during gliding, based on Fig. 1A; proximal feathers on the humerus and femur are inferred (data are from ref. 12). (Scale bar, 5 cm.) cgi doi pnas Chatterjee and Templin
 with unequal wing sections and different chords (16) and a ventral wing area (0.042 m 2 ) approximately one-half the extent of the dorsal wing (0.089 m 2 ) (Fig. 2 J).")
4 (Fig. 2D). The lower wing is placed somewhat posterior and ventral in relation to the dorsal wing (on the hand) like a staggered biplane (Fig. 2G) with unequal wing sections and different chords (16) and a ventral wing area (0.042 m 2 ) approximately one-half the extent of the dorsal wing (0.089 m 2 ) (Fig. 2 J). Merely by stretching both of its wings, Microraptor would have been able to glide in much the same way as a mechanical glider without muscle power. The stiff mesotarsal ankle joint would prevent torsion of the ventral wing on the metatarsus. A ligament, analogous to the avian vinculum, could hold the feathers of the hindwing in place during flight and aid in folding when not in use. Using Fig. 2D as a guide, we reconstruct the dorsal view of Microraptor in a gliding pose to show the biplane configuration of the wing planform (Fig. 2 J). Flight Performance Microraptor displays several anatomical features that suggest it could become airborne (12): elongate and asymmetric vanes in the flight feathers (17) at the distal segment of each limb; a scapulocoracoid whose ends are oriented at an acute angle to each other (18); a laterally facing glenoid for gentle dorsoventral movement of the wing (19); a single, enlarged sternum for attachment of the flight muscles; ossified sternal ribs and well developed uncinate processes for resisting compression force on the thoracic cavity imposed during downstroke; strongly bowed outer metacarpal; and a flattened central digit for attachment of primaries. To analyze the flight performance of Microraptor, we used two computer algorithms, ANFLTPWR (animal flight power) and ANFLTSIM (animal flight simulation), which are based on the streamtube model (20) using several flight parameters (Table 1). For any flying animal or fixed-wing aircraft or helicopter, the plot of power required for level flight against airspeed is a U-shaped curve, higher at both ends than at the middle. Using the ANFLTPWR program (20), we generated the power curve of Microraptor (Fig. 3A), where the U-shaped curve is the power required for steady level flight and the horizontal line is the estimated maximum steady power available. Because the two curves, power required and power available, nearly coincide at flight speeds of 9 15 m/s, limited steady flight may have been theoretically possible. Gliding performance is shown as a polar curve, which is a plot of horizontal speed versus sinking speed. We used the AN- FLTSIM program (20) to calculate these values. Fig. 3B shows the potential gliding performance of Microraptor in terms of sinking speed versus horizontal speed. By this plot, Microraptor was possibly a moderate glider, not as efficient as highperformance gliders, such as the long-span frigatebird (Fregata magnificens, M 1.5 kg) or a Cretaceous pterosaur (Nyctosaurus, M 1.86 kg) (21), of comparable size. Anatomical evidence indicates that Microraptor was not capable of ground or running takeoff, because it lacked the supracoracoideus pulley to elevate the wings. Moreover, running takeoff would damage the ventral metatarsal wings (12). We have calculated the takeoff capability of Microraptor from a perch using both streamtube (20) and aircraft (22) models (Fig. 3C). When birds take off from a perch, they do not seem to use excess power; they lose height at first and then swoop up with a large-amplitude undulation to swing between two perches, known as phugoid gliding (8, 20). In Fig. 3C, we have plotted several glide paths for Microraptor, starting from horizontal launches at 3 m/s. Considerable height losses are involved, as shown in Fig. 3C (curve 1), but they are minimized by the use of a simulated pitch damper, which may imitate the tail motion of Microraptor to control the phugoid oscillations. The pitch damper adjusts the wing lift in synchrony with speed changes to reduce or eliminate the oscillations. By using phugoid gliding, Microraptor could potentially have traveled from one tree to another tree by undulating flight covering a horizontal distance of 40 m. This mode of transportation would have been energetically very efficient for Microraptor. The long feathered tail would provide pitch damping as well as general stability. The minimum speed during the pitchup is 4.5 m/s, which may have been safe for landing. The combined wing appears to be too small to serve as a parachute that would withstand a fatal fall. We plotted curve 4 in Fig. 3C, a trajectory for a flat parachute descent in which Microraptor has high drag with outstretched wings, while generating no lateral force. The trajectory is not simply a vertical line because of the horizontal jump-off. The terminal velocity is 8.7 m/s, a potentially crashing speed against a solid surface but perhaps safe for landing on padded ground cover or on a flexible tree branch. Drag could be dramatically reduced in a gliding Microraptor by streamlining the vertically held tibia with feathers so that the turbulent spaces behind are filled in and the front areas are rounded or tapered (Fig. 2 E G). Without leg feathers, the cross-section of the tibia would be circular, so the airflow behind the bone would break up into eddies, producing turbulence. With the presence of tibial feathers, Microraptor could reduce 40% of drag when the legs were held in z-fashion [see supporting information (SI)]. EVOLUTION Fig. 4. A simple cladogram of eumaniraptoran theropods showing the distribution of leg feathers in selected taxa (modified from refs. 23, 25, and 26). In Microraptor, the contour feathers are present on the femur, tibia, and metatarsus, but only the metatarsal feathers are asymmetric and form the ventral wing of the biplane design; the feathers on the femur and tibia are symmetric (12). In Pedopenna, long metatarsal feathers are present to form the ventral wing of the biplane layout, but they appear to be symmetrical (25). In Archaeopteryx, long contour feathers are present on the femur and tibia, but they appear to be lost on the metatarsus (15). At this stage, the evolution of monoplane design probably took place. In an unnamed enantiornithine bird, long contour feathers are present on the femur and tibia but absent in the metatarsal region (26). In modern raptors such as the falcon Falco, similar contour leg feathers persist on the femur and tibia for streamlining, but metatarsal feathers are generally reduced or absent. Chatterjee and Templin PNAS January 30, 2007 vol. 104 no
5 Discussion and Conclusion Phylogenetically, Microraptor belongs to eumaniraptorans, which also include a series of feathered theropods and Archaeopteryx (23), possessing long arms and hands for the attachment of vaned, barbed feathers (Fig. 4A). Although Microraptor appears to have been a glider, there are two phylogenetic and biomechanical interpretations for this unusual biplane wing configuration of Microraptor: either (i) avian flight went through a Microraptor-like biplane stage to become a monoplane configuration when the hindlimb became decoupled from its gliding function (with the loss of ventral wing); or (ii) the biplane wing configuration may represent a failed or temporary experiment in the deployment of aerodynamic feathers among one branch of deinonychosaurs that dallied with gliding. Both scenarios are equally possible. However, the former view finds support from current fossil and recent evidence in a broad phylogenetic context indicating a gradual shift in locomotory dominance from the hindlimb to forelimb during the evolution of avian flight (Fig. 4). Several Chinese maniraptorans including Caudipteryx (24), Sinornithosaurus (4), and Cryptovolans (5), as well as modern raptors, show contour feathers on the hindlimbs, where tibial feathers were probably used for streamlining. The recent discovery of Pedopenna (25) from the Middle or Late Jurassic of China, another feathered maniraptoran with long metatarsal feathers, may support the biplane wing configuration of gliding dromaeosaurs before the Archaeopteryx stage (Fig. 4). Archaeopteryx shows long contour feathers on the hindlimb, especially in the tibial region in the Berlin specimen (15); it apparently lacked the metatarsal feathers, which were compensated with larger forewing and long asymmetric retrices on the tail for additional lift. Long contour tibial feathers also are known in an unnamed Early Cretaceous enantiornithine bird from China, but metatarsal feathers appear to be absent (26). Symmetric contour feathers occur on the femur and tibia in living raptors, even on the proximal part of the tarsometatarsus (Fig. 4). Unlike other birds, raptors keep their hindlegs in a z-configuration during preparation for aerial attack and carrying prey, dangling their tibiae in a vertical plane (Fig. 2H). The feathered trousers are a conspicuous costume of predatory birds, keeping their prey-catching legs streamlined during aerial attack. Microraptor provided the crucial clue about the role of leg feathers in the flight of living raptors. Aircraft designers have mimicked many of nature s flight inventions, usually inadvertently. Leading edge slats delay stalling, as does the alula of birds; birds feet act as airbrakes, and streamlining reduces drag. Now, it seems likely that Microraptor invented the biplane 125 million years before the Wright 1903 Flyer. Methodology Various flight parameters such as wingspan (b), body length, and forewing and tailwing areas (S) were calculated from a high-fidelity cast of the holotype specimen at the National Museum of Rio de Janeiro in Brazil. The hindwing area was estimated from a modified dorsal reconstruction of Microraptor (Fig. 2 J). We have digitized the body outline in dorsal aspect with a computer program by Rohlf ( and estimated the combined wing area (forewing plus hindwing) as well as the area from tail feathers (Table 1). Our method of estimating the mass of Microraptor used a multivariate analysis proposed by Atanassov and Strauss. The predicted mass of Microraptor was calculated to be 0.95 kg, which is approximately the mass of medium-sized extant predatory birds [i.e., the common black hawk (Buteogallus anthracinus) or the northern goshawk (Accipiter gentilis)] (27). To analyze the flight performance of Microraptor, we used two computer algorithms, ANFLTPWR and ANFLTSIM, which are based on the streamtube model (20) and also described above. In all cases, the body and wing drag coefficients are computed as functions of the Reynolds number (see, ref. 20, and SI for detailed methodology). Atanassov, M. N., Strauss, R. (2002) J Vertebr Paleontol 22:33A (abstr.). We thank Lynn Margulis for her suggestions, support, and sponsorship; Jeff Martz for stimulating discussions on the wing planform and for illustrations; M. Atanassov for estimating the wing area and mass of Microraptor; Alexander Kellner for the high-fidelity cast and life mount of Microraptor at the National Museum of Rio de Janeiro, Brazil; J. Martz, M. Atanassov, J. Barrick, Soumya Chatterjee, and several anonymous reviewers for insightful comments and suggestions for the improvement of the manuscript; Xu Xing for providing images of Microraptor and Pedopenna; and Thomas Moore for permission to use the sketch of Microraptor in Fig. 1. Funding was provided by Texas Tech University. 1. Chen PJ, Dong Z, Zhen S (1998) Nature 391: Ji Q, Currie PJ, Norell MA, Ji S (1998) Nature 393: Xu X, Zhou Z, Wang X (2000) Nature 408: Xu X, Wang X, Wu X (1999) Nature 401: Czerkas SA, Zhang D, Li J, Li Y (2002) Mus J 1: Hou L, Zhou Z, Martin LD, Feduccia A (1995) Nature 377: Chatterjee S (1997) The Rise of Birds (Johns Hopkins Univ Press, Baltimore). 8. Chatterjee S, Templin RJ (2003) Naturwissenschaften 90: Chatterjee S, Templin RJ (2004) in Feathered Dragons: Studies on the Transition from Dinosaurs to Birds, eds Currie PJ, Koppelhus EB, Shugar MA, Wright JL (Indiana Univ Press, Bloomington, IN), pp Chatterjee S, Templin RJ (2005) Nat Hist 114: Zhang F, Zhou Z, Xu X, Wang X (2002) Naturwissenschaften 89: Xu X, Zhou Z, Wang X, Kuang X, Zhang F, Du X (2003) Nature 421: Padian K, Dial KP (2005) Nature 438:E3 E Hotton N, III (1980) in A Cold Look at the Warm-Blooded Dinosaurs, eds Thomas RDK, Olson EC (Westview, Boulder, CO), pp Longrich N (2006) Palaeobiology 32: Munk MM (1923) Nat Adv Aero Rep 151: Feduccia A, Tordoff HB (1979) Science 203: Feduccia A (1996) The Origin and Evolution of Birds (Yale Univ Press, New Haven, CT). 19. Jenkins FA, Jr (1993) Am J Sci 293A: Templin RJ (2000) Prog Aeronaut Sci 36: Chatterjee S, Templin RJ (2004) Geol Soc Am Sp Pap 376: Pennycuick CJ (1975) Mechanics of Flight in Avian Biology, Vol 5, eds Farner DS, King JR (Academic, New York), pp Holtz TR (2005) Nat Hist 114: Jones TD, Farlow JO, Ruben JA, Henderson DM, Hillenius WJ (2000) Nature 406: Xu X, Zhang F (2005) Naturwissenschaften 92: Zhang F, Zhou Z (2004) Nature 431: Sibley DA (2000) The Sibley Guide to Birds (Alfred A. Knopf, New York) cgi doi pnas Chatterjee and Templin

6 1 Chatterjee, S, and Templin, R. J. Supporting Information Files in this Data Supplement Supporting Figure 5 Supporting Figure 6 Supporting Figure 7 Supporting Text Fig. 5. Holotype of Microraptor gui (IVPP V13352) as preserved, scale bar 5 cm; inset, enlarged view of the leg feathers (After Xu et al., 1).
 Articulation of pelvic girdle and femur (length of femur = 12 cm) of Bambiraptor in anterior view showing normal vertical limb posture; (B) the same in anterior view; but the femur is put in a")
7 2 Fig. 6. (A) Articulation of pelvic girdle and femur (length of femur = 12 cm) of Bambiraptor in anterior view showing normal vertical limb posture; (B) the same in anterior view; but the femur is put in a horizontal pose, so that it becomes unstable at the joint and dislocated from the hip socket; no dinosaur could assume this horizontal leg posture; in the reconstruction of Microraptor by Xu et al. (1, Fig. 1c), the femur is placed in this horizontal fashion, which is anatomically incorrect. (C) Life restoration of Microraptor at the National Museum of Rio de Janeiro, Brazil, where the hindlimb is shown in a vertical pose. However, the leading edge of the asymmetric flight feathers on the metatarsals faces downward (shown by arrow), which would fail to generate thrust and lift. The orientation of the metatarsal feathers in this pose is incorrect from an aerodynamic point of view.
8
9
Accepted Manuscript. News & Views. Primary feather vane asymmetry should not be used to predict the flight capabilities of feathered fossils
 Accepted Manuscript News & Views Primary feather vane asymmetry should not be used to predict the flight capabilities of feathered fossils Xia Wang, Robert L. Nudds, Colin Palmer, Gareth J. Dyke PII: S2095-9273(17)30453-X
Accepted Manuscript News & Views Primary feather vane asymmetry should not be used to predict the flight capabilities of feathered fossils Xia Wang, Robert L. Nudds, Colin Palmer, Gareth J. Dyke PII: S2095-9273(17)30453-X
Barney to Big Bird: The Origin of Birds. Caudipteryx. The fuzzy raptor. Solnhofen Limestone, cont d
 Barney to Big Bird: The Origin of Birds Caudipteryx The fuzzy raptor The discovery of feathered dinosaurs in Liaoning, China, has excited the many paleontologists who suspected a direct link between dinosaurs
Barney to Big Bird: The Origin of Birds Caudipteryx The fuzzy raptor The discovery of feathered dinosaurs in Liaoning, China, has excited the many paleontologists who suspected a direct link between dinosaurs
The wing of Archaeopteryx as a primary thrust generator
 Page 1 of 5 The wing of Archaeopteryx as a primary thrust generator Nature 399, pp. 60-62 (1999) Macmillan Publishers Ltd. PHILLIP BURGERS* AND LUIS M. CHIAPPE * San Diego Natural History Museum, PO Box
Page 1 of 5 The wing of Archaeopteryx as a primary thrust generator Nature 399, pp. 60-62 (1999) Macmillan Publishers Ltd. PHILLIP BURGERS* AND LUIS M. CHIAPPE * San Diego Natural History Museum, PO Box
A new maniraptoran dinosaur from China with long feathers on the metatarsus
 Naturwissenschaften (2005) 92:173 177 DOI 10.1007/s00114-004-0604-y SHORT COMMUNICATION Xing Xu Fucheng Zhang A new maniraptoran dinosaur from China with long feathers on the metatarsus Received: 18 March
Naturwissenschaften (2005) 92:173 177 DOI 10.1007/s00114-004-0604-y SHORT COMMUNICATION Xing Xu Fucheng Zhang A new maniraptoran dinosaur from China with long feathers on the metatarsus Received: 18 March
The Origin of Birds. Technical name for birds is Aves, and avian means of or concerning birds.
 The Origin of Birds Technical name for birds is Aves, and avian means of or concerning birds. Birds have many unusual synapomorphies among modern animals: [ Synapomorphies (shared derived characters),
The Origin of Birds Technical name for birds is Aves, and avian means of or concerning birds. Birds have many unusual synapomorphies among modern animals: [ Synapomorphies (shared derived characters),
A juvenile coelurosaurian theropod from China indicates arboreal habits
 Naturwissenschaften (2002) 89:394 398 DOI 10.1007/s00114-002-0353-8 SHORT COMMUNICATION Fucheng Zhang Zhonghe Zhou Xing Xu Xiaolin Wang A juvenile coelurosaurian theropod from China indicates arboreal
Naturwissenschaften (2002) 89:394 398 DOI 10.1007/s00114-002-0353-8 SHORT COMMUNICATION Fucheng Zhang Zhonghe Zhou Xing Xu Xiaolin Wang A juvenile coelurosaurian theropod from China indicates arboreal
Origin and Evolution of Birds. Read: Chapters 1-3 in Gill but limited review of systematics
 Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
The Evolution of Birds & the Origin of Flight
 The Evolution of Birds & the Origin of Flight Archaeopteryx Solnhofen quarry Oldest known bird, but not ancestral to modern birds Inhabited coastal habitats where it probably glided between conifers, cycads,
The Evolution of Birds & the Origin of Flight Archaeopteryx Solnhofen quarry Oldest known bird, but not ancestral to modern birds Inhabited coastal habitats where it probably glided between conifers, cycads,
What is evolution? Transitional fossils: evidence for evolution. In its broadest sense, evolution is simply the change in life through time.
 Transitional fossils: evidence for evolution http://domain- of- darwin.deviantart.com/art/no- Transitional- Fossils- 52231284 Western MA Atheists and Secular Humanists 28 May 2016 What is evolution? In
Transitional fossils: evidence for evolution http://domain- of- darwin.deviantart.com/art/no- Transitional- Fossils- 52231284 Western MA Atheists and Secular Humanists 28 May 2016 What is evolution? In
Name: GEOL 104 Dinosaurs: A Natural History Video Assignment. DUE: Wed. Oct. 20
 GEOL 104 Dinosaurs: A Natural History Video Assignment DUE: Wed. Oct. 20 Documentaries represent one of the main media by which scientific information reaches the general public. For this assignment, you
GEOL 104 Dinosaurs: A Natural History Video Assignment DUE: Wed. Oct. 20 Documentaries represent one of the main media by which scientific information reaches the general public. For this assignment, you
Origin and Evolution of Birds. Read: Chapters 1-3 in Gill but limited review of systematics
 Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Recall: The Earliest Thoughts about Flying Took place before the days of science.
 Recall: The Earliest Thoughts about Flying Took place before the days of science. Before man began to investigate with carefully planned experiments, and to figure things out in an orderly fashion. Men
Recall: The Earliest Thoughts about Flying Took place before the days of science. Before man began to investigate with carefully planned experiments, and to figure things out in an orderly fashion. Men
From Reptiles to Aves
 First Vertebrates From Reptiles to Aves Evolutions of Fish to Amphibians Evolution of Amphibians to Reptiles Evolution of Reptiles to Dinosaurs to Birds Common Ancestor of Birds and Reptiles: Thecodonts
First Vertebrates From Reptiles to Aves Evolutions of Fish to Amphibians Evolution of Amphibians to Reptiles Evolution of Reptiles to Dinosaurs to Birds Common Ancestor of Birds and Reptiles: Thecodonts
The Fossil Record of Vertebrate Transitions
 The Fossil Record of Vertebrate Transitions The Fossil Evidence of Evolution 1. Fossils show a pattern of change through geologic time of new species appearing in the fossil record that are similar to
The Fossil Record of Vertebrate Transitions The Fossil Evidence of Evolution 1. Fossils show a pattern of change through geologic time of new species appearing in the fossil record that are similar to
Video Assignments. Microraptor PBS The Four-winged Dinosaur Mark Davis SUNY Cortland Library Online
 Video Assignments Microraptor PBS The Four-winged Dinosaur Mark Davis SUNY Cortland Library Online Radiolab Apocalyptical http://www.youtube.com/watch?v=k52vd4wbdlw&feature=youtu.be Minute 13 through minute
Video Assignments Microraptor PBS The Four-winged Dinosaur Mark Davis SUNY Cortland Library Online Radiolab Apocalyptical http://www.youtube.com/watch?v=k52vd4wbdlw&feature=youtu.be Minute 13 through minute
SUPPLEMENTARY INFORMATION
 1. Preservation of STM4-1 and STM22-6 STM4-1 represents a complete skeleton with integumentary structures preserved on part and counterpart slabs. STM22-6 is a nearly complete skeleton with integumentary
1. Preservation of STM4-1 and STM22-6 STM4-1 represents a complete skeleton with integumentary structures preserved on part and counterpart slabs. STM22-6 is a nearly complete skeleton with integumentary
From Dinosaurs to Birds: Puzzles Unraveled while Evidence Building up
 From Dinosaurs to Birds: Puzzles Unraveled while Evidence Building up CHEN Pingfu 1 and SONG Jianlan 2 1 Institute of Vertebrate Paleontology and Paleoanthropology 2 BCAS Staff Reporter Rejuvenation of
From Dinosaurs to Birds: Puzzles Unraveled while Evidence Building up CHEN Pingfu 1 and SONG Jianlan 2 1 Institute of Vertebrate Paleontology and Paleoanthropology 2 BCAS Staff Reporter Rejuvenation of
8/19/2013. Topic 14: Body support & locomotion. What structures are used for locomotion? What structures are used for locomotion?
 Topic 4: Body support & locomotion What are components of locomotion? What structures are used for locomotion? How does locomotion happen? Forces Lever systems What is the difference between performance
Topic 4: Body support & locomotion What are components of locomotion? What structures are used for locomotion? How does locomotion happen? Forces Lever systems What is the difference between performance
Anatomy. Name Section. The Vertebrate Skeleton
 Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported
 Drawing of early ankle development of Alligator mississippiensis, as reported") Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported by a previous study 1. The intermedium is formed at
Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported by a previous study 1. The intermedium is formed at
'Rain' of dead birds on central NJ lawns explained; Federal culling program killed up to 5,000 Associated Press, January 27, 2009
 'Rain' of dead birds on central NJ lawns explained; Federal culling program killed up to 5,000 Associated Press, January 27, 2009 Study May Give Hope That Ivory-billed Woodpeckers Still Around Science
'Rain' of dead birds on central NJ lawns explained; Federal culling program killed up to 5,000 Associated Press, January 27, 2009 Study May Give Hope That Ivory-billed Woodpeckers Still Around Science
Comparative Physiology 2007 Second Midterm Exam. 1) 8 pts. 2) 14 pts. 3) 12 pts. 4) 17 pts. 5) 10 pts. 6) 8 pts. 7) 12 pts. 8) 10 pts. 9) 9 pts.
 8 pts. 2) 14 pts. 3) 12 pts. 4) 17 pts. 5) 10 pts. 6) 8 pts. 7) 12 pts. 8) 10 pts. 9) 9 pts.") Name: Comparative Physiology 2007 Second Midterm Exam 1) 8 pts 2) 14 pts 3) 12 pts 4) 17 pts 5) 10 pts 6) 8 pts 7) 12 pts 8) 10 pts 9) 9 pts Total 1. Cells I and II, shown below, are found in the gills
Name: Comparative Physiology 2007 Second Midterm Exam 1) 8 pts 2) 14 pts 3) 12 pts 4) 17 pts 5) 10 pts 6) 8 pts 7) 12 pts 8) 10 pts 9) 9 pts Total 1. Cells I and II, shown below, are found in the gills
A new species of Confuciusornis from Lower Cretaceous of Jianchang Liaoning China
 29 2 2010 6 GLOBAL GEOLOGY Vol. 29 No. 2 Jun. 2010 1004-5589 2010 02-0183 - 05 1 2 2 2 1. 110004 2. 110034 Confuciusornis jianchangensis sp. nov. 蹠 V 蹠 Q915. 865 A doi 10. 3969 /j. issn. 1004-5589. 2010.
29 2 2010 6 GLOBAL GEOLOGY Vol. 29 No. 2 Jun. 2010 1004-5589 2010 02-0183 - 05 1 2 2 2 1. 110004 2. 110034 Confuciusornis jianchangensis sp. nov. 蹠 V 蹠 Q915. 865 A doi 10. 3969 /j. issn. 1004-5589. 2010.
Discovery of an Avialae bird from China, Shenzhouraptor sinensis gen. et sp. nov.
 Discovery of an Avialae bird from China, Shenzhouraptor sinensis gen. et sp. nov. by Qiang Ji 1, Shuan Ji 2, Hailu You 1, Jianping Zhang 3, Chongxi Yuan 3, Xinxin Ji 4, Jinglu Li 5, and Yinxian Li 5 1.
Discovery of an Avialae bird from China, Shenzhouraptor sinensis gen. et sp. nov. by Qiang Ji 1, Shuan Ji 2, Hailu You 1, Jianping Zhang 3, Chongxi Yuan 3, Xinxin Ji 4, Jinglu Li 5, and Yinxian Li 5 1.
Shedding Light on the Dinosaur-Bird Connection
 Shedding Light on the Dinosaur-Bird Connection This text is provided courtesy of the American Museum of Natural History. When people think of dinosaurs, two types generally come to mind: the huge herbivores
Shedding Light on the Dinosaur-Bird Connection This text is provided courtesy of the American Museum of Natural History. When people think of dinosaurs, two types generally come to mind: the huge herbivores
On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds
 and the origin of birds") On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds by Qiang Ji and Shu an Ji Chinese Geological Museum, Beijing Chinese Geology Volume 233 1996 pp.
On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds by Qiang Ji and Shu an Ji Chinese Geological Museum, Beijing Chinese Geology Volume 233 1996 pp.
TAXONOMIC HIERARCHY. science of classification and naming of organisms
 TAXONOMIC HIERARCHY Taxonomy - science of classification and naming of organisms Taxonomic Level Kingdom Phylum subphylum Class subclass superorder Order Family Genus Species Example Animalae Chordata
TAXONOMIC HIERARCHY Taxonomy - science of classification and naming of organisms Taxonomic Level Kingdom Phylum subphylum Class subclass superorder Order Family Genus Species Example Animalae Chordata
New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia
 1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China
 SUPPLEMENTARY INFORMATION A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China Ya-Ming Wang 1, Hai-Lu You 2,3 *, Tao Wang 4 1 School of Earth Sciences and Resources, China
SUPPLEMENTARY INFORMATION A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China Ya-Ming Wang 1, Hai-Lu You 2,3 *, Tao Wang 4 1 School of Earth Sciences and Resources, China
A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers
 A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers Fucheng Zhang, Zhonghe Zhou, Xing Xu, Xiaolin Wang, Corwin Sullivan Laboratory of Evolutionary Systematics of Vertebrates,
A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers Fucheng Zhang, Zhonghe Zhou, Xing Xu, Xiaolin Wang, Corwin Sullivan Laboratory of Evolutionary Systematics of Vertebrates,
Bird evolution. Primer
 R350 other to achieve their goal. There are also heroes and heroines and Ashburner identifies these as including Gerry Rubin and Suzi Lewis in particular. Heroes come in all shapes and sizes and, despite
R350 other to achieve their goal. There are also heroes and heroines and Ashburner identifies these as including Gerry Rubin and Suzi Lewis in particular. Heroes come in all shapes and sizes and, despite
An Archaeopteryx-like theropod dinosaur newly
 BCAS Vol.25 No.4 2011 Archaeopteryx: Dinosaur or Bird? By SONG Jianlan (Staff Reporter) An Archaeopteryx-like theropod dinosaur newly found from western Liaoning Province in northeastern China would make
BCAS Vol.25 No.4 2011 Archaeopteryx: Dinosaur or Bird? By SONG Jianlan (Staff Reporter) An Archaeopteryx-like theropod dinosaur newly found from western Liaoning Province in northeastern China would make
Vertebrate Locomotion: Aquatic
 Vertebrate Locomotion: Aquatic Swimming Nearly all vertebrates can swim Sole form of locomotion for fish and larval amphibians Primary swimmers Terrestrial vertebrates that readapt to aquatic life still
Vertebrate Locomotion: Aquatic Swimming Nearly all vertebrates can swim Sole form of locomotion for fish and larval amphibians Primary swimmers Terrestrial vertebrates that readapt to aquatic life still
For every purpose of dog, there are specific builds that give superior performance.
 LAURIE EDGE-HUGHES, BScPT, MAnimSt, (Animal Physio), CAFCI, CCRT Four Leg Rehab Inc The Canine Fitness Centre Ltd For every purpose of dog, there are specific builds that give superior performance. Huskies,
LAURIE EDGE-HUGHES, BScPT, MAnimSt, (Animal Physio), CAFCI, CCRT Four Leg Rehab Inc The Canine Fitness Centre Ltd For every purpose of dog, there are specific builds that give superior performance. Huskies,
PERSPECTIVES IN ORNITHOLOGY ARCHAEOPTERYX 2007: QUO VADIS?
 PERSPECTIVES IN ORNITHOLOGY ARCHAEOPTERYX 2007: QUO VADIS? Author(s): Alan Feduccia, Larry D. Martin, Sam Tarsitano Source: The Auk, 124(2):373-380. Published By: The American Ornithologists' Union https://doi.org/10.1642/0004-8038(2007)124[373:pioaqv]2.0.co;2
PERSPECTIVES IN ORNITHOLOGY ARCHAEOPTERYX 2007: QUO VADIS? Author(s): Alan Feduccia, Larry D. Martin, Sam Tarsitano Source: The Auk, 124(2):373-380. Published By: The American Ornithologists' Union https://doi.org/10.1642/0004-8038(2007)124[373:pioaqv]2.0.co;2
Early diversification of birds: Evidence from a new opposite bird
 Early diversification of birds: Evidence from a new opposite bird ZHANG Fucheng 1, ZHOU Zhonghe 1, HOU Lianhai 1 & GU Gang 2 1. Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy
Early diversification of birds: Evidence from a new opposite bird ZHANG Fucheng 1, ZHOU Zhonghe 1, HOU Lianhai 1 & GU Gang 2 1. Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy
Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida. Evo-Devo Revisited. Development of the Tetrapod Limb
 Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida Evo-Devo Revisited Development of the Tetrapod Limb Limbs whether fins or arms/legs for only in particular regions or LIMB FIELDS. Primitively
Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida Evo-Devo Revisited Development of the Tetrapod Limb Limbs whether fins or arms/legs for only in particular regions or LIMB FIELDS. Primitively
Origins of avian flight a new perspective
 Origins of avian flight a new perspective Larry D. Martin Department of Ecology and Evolutionary Biology; Museum of Natural History and Biodiversity Research Center, University of Kansas, Lawrence, KS
Origins of avian flight a new perspective Larry D. Martin Department of Ecology and Evolutionary Biology; Museum of Natural History and Biodiversity Research Center, University of Kansas, Lawrence, KS
1/9/2013. Divisions of the Skeleton: Topic 8: Appendicular Skeleton. Appendicular Components. Appendicular Components
 /9/203 Topic 8: Appendicular Skeleton Divisions of the Skeleton: Cranial Postcranial What makes up the appendicular skeleton? What is the pattern of serial homology of the limbs? Tetrapod front limb morphology
/9/203 Topic 8: Appendicular Skeleton Divisions of the Skeleton: Cranial Postcranial What makes up the appendicular skeleton? What is the pattern of serial homology of the limbs? Tetrapod front limb morphology
Fossilized remains of cat-sized flying reptile found in British Columbia
 Fossilized remains of cat-sized flying reptile found in British Columbia By Washington Post, adapted by Newsela staff on 09.06.16 Word Count 768 An artist's impression of the small-bodied, Late Cretaceous
Fossilized remains of cat-sized flying reptile found in British Columbia By Washington Post, adapted by Newsela staff on 09.06.16 Word Count 768 An artist's impression of the small-bodied, Late Cretaceous
Walking Like Dinosaurs: Chickens with Artificial Tails Provide Clues about Non-Avian Theropod Locomotion
 Walking Like Dinosaurs: Chickens with Artificial Tails Provide Clues about Non-Avian Theropod Locomotion Bruno Grossi 1,2, José Iriarte-Díaz 3,4 *, Omar Larach 2, Mauricio Canals 2, Rodrigo A. Vásquez
Walking Like Dinosaurs: Chickens with Artificial Tails Provide Clues about Non-Avian Theropod Locomotion Bruno Grossi 1,2, José Iriarte-Díaz 3,4 *, Omar Larach 2, Mauricio Canals 2, Rodrigo A. Vásquez
the Feather or the Bird?
 Which Came First, the Feather or the Bird? FEATHERS EVOLVED in carnivorous, bipedal dinosaurs before the origin of birds. The creatures depicted here are reconstructions of fossils found recently in northern
Which Came First, the Feather or the Bird? FEATHERS EVOLVED in carnivorous, bipedal dinosaurs before the origin of birds. The creatures depicted here are reconstructions of fossils found recently in northern
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A.
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
First Flightless Pterosaur
 First Flightless Pterosaur David Peters no affiliation 9 Greenfield Court, Saint Charles, MO 63303 USA Pterosaur fossils have been discovered all over the world [1], but so far no flightless pterosaurs
First Flightless Pterosaur David Peters no affiliation 9 Greenfield Court, Saint Charles, MO 63303 USA Pterosaur fossils have been discovered all over the world [1], but so far no flightless pterosaurs
Juehuaornis gen. nov.
 34 1 2015 3 GLOBAL GEOLOGY Vol. 34 No. 1 Mar. 2015 1004 5589 2015 01 0007 05 Juehuaornis gen. nov. 1 1 1 2 1. 110034 2. 110034 70% Juehuaornis zhangi gen. et sp. nov Q915. 4 A doi 10. 3969 /j. issn. 1004-5589.
34 1 2015 3 GLOBAL GEOLOGY Vol. 34 No. 1 Mar. 2015 1004 5589 2015 01 0007 05 Juehuaornis gen. nov. 1 1 1 2 1. 110034 2. 110034 70% Juehuaornis zhangi gen. et sp. nov Q915. 4 A doi 10. 3969 /j. issn. 1004-5589.
Biology 1B Evolution Lecture 11 (March 19, 2010), Insights from the Fossil Record and Evo-Devo
, Insights from the Fossil Record and Evo-Devo") Biology 1B Evolution Lecture 11 (March 19, 2010), Insights from the Fossil Record and Evo-Devo Extinction Important points on extinction rates: Background rate of extinctions per million species per year:
Biology 1B Evolution Lecture 11 (March 19, 2010), Insights from the Fossil Record and Evo-Devo Extinction Important points on extinction rates: Background rate of extinctions per million species per year:
University of Bristol - Explore Bristol Research. Early version, also known as pre-print
 Godefroit, P., Sinitsa, S. M., Dhouailly, D., Bolotsky, Y. L., Sizov, A. V., McNamara, M. E.,... Spagna, P. (2014). Dinosaur evolution. A Jurassic ornithischian dinosaur from Siberia with both feathers
Godefroit, P., Sinitsa, S. M., Dhouailly, D., Bolotsky, Y. L., Sizov, A. V., McNamara, M. E.,... Spagna, P. (2014). Dinosaur evolution. A Jurassic ornithischian dinosaur from Siberia with both feathers
Diapsida. BIO2135 Animal Form and Function. Page 1. Diapsida (Reptilia, Sauropsida) Amniote eggs. Amniote egg. Temporal fenestra.
 Amniote eggs. Amniote egg. Temporal fenestra.") Diapsida (Reptilia, Sauropsida) Vertebrate phylogeny Mixini Chondrichthyes Sarcopterygii Mammalia Pteromyzontida Actinopterygii Amphibia Reptilia! 1! Amniota (autapomorphies) Costal ventilation Amniote
Diapsida (Reptilia, Sauropsida) Vertebrate phylogeny Mixini Chondrichthyes Sarcopterygii Mammalia Pteromyzontida Actinopterygii Amphibia Reptilia! 1! Amniota (autapomorphies) Costal ventilation Amniote
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes)
") Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
Diapsida. BIO2135 Animal Form and Function. Page 1. Diapsida (Reptilia, Sauropsida) Amniote egg. Membranes. Vertebrate phylogeny
 Amniote egg. Membranes. Vertebrate phylogeny") Diapsida (Reptilia, Sauropsida) 1 Vertebrate phylogeny Mixini Chondrichthyes Sarcopterygii Mammalia Pteromyzontida Actinopterygii Amphibia Reptilia!! Amniota (autapomorphies) Costal ventilation Amniote
Diapsida (Reptilia, Sauropsida) 1 Vertebrate phylogeny Mixini Chondrichthyes Sarcopterygii Mammalia Pteromyzontida Actinopterygii Amphibia Reptilia!! Amniota (autapomorphies) Costal ventilation Amniote
WHY ORNITHOLOGISTS SHOULD CARE ABOUT THE THEROPOD ORIGIN OF BIRDS
 The Auk A Quarterly Journal of Ornithology Vol. 119 No. 1 January 2002 The Auk 119(1):1 17, 2002 PERSPECTIVES IN ORNITHOLOGY WHY ORNITHOLOGISTS SHOULD CARE ABOUT THE THEROPOD ORIGIN OF BIRDS RICHARD O.
The Auk A Quarterly Journal of Ornithology Vol. 119 No. 1 January 2002 The Auk 119(1):1 17, 2002 PERSPECTIVES IN ORNITHOLOGY WHY ORNITHOLOGISTS SHOULD CARE ABOUT THE THEROPOD ORIGIN OF BIRDS RICHARD O.
Your web browser (Safari 7) is out of date. For more security, comfort and the best experience on this site: Update your browser Ignore
 is out of date. For more security, comfort and the best experience on this site: Update your browser Ignore") Your web browser (Safari 7) is out of date. For more security, comfort and the best experience on this site: Update your browser Ignore Activityapply ADAPTIVE RADIATIO N How do species respond to environmental
Your web browser (Safari 7) is out of date. For more security, comfort and the best experience on this site: Update your browser Ignore Activityapply ADAPTIVE RADIATIO N How do species respond to environmental
Effects of size and behavior on aerial performance of two species of flying snakes (Chrysopelea)
") The Journal of Experimental Biology 2, 135-147 Published by The Company of Biologists 25 doi:.1242/jeb.15 135 Effects of size and behavior on aerial performance of two species of flying snakes (Chrysopelea)
The Journal of Experimental Biology 2, 135-147 Published by The Company of Biologists 25 doi:.1242/jeb.15 135 Effects of size and behavior on aerial performance of two species of flying snakes (Chrysopelea)
Introduction and methods will follow the same guidelines as for the draft
 Locomotion Paper Guidelines Entire paper will be 5-7 double spaced pages (12 pt font, Times New Roman, 1 inch margins) without figures (but I still want you to include them, they just don t count towards
Locomotion Paper Guidelines Entire paper will be 5-7 double spaced pages (12 pt font, Times New Roman, 1 inch margins) without figures (but I still want you to include them, they just don t count towards
Effects of size and behavior on aerial performance of two species of flying snakes (Chrysopelea)
") The Journal of Experimental Biology 2, 135-1 Published by The Company of Biologists 25 doi:.122/jeb.15 135 Effects of size and behavior on aerial performance of two species of flying snakes (Chrysopelea)
The Journal of Experimental Biology 2, 135-1 Published by The Company of Biologists 25 doi:.122/jeb.15 135 Effects of size and behavior on aerial performance of two species of flying snakes (Chrysopelea)
Ceri Pennington VELOCIRAPTOR
 Ceri Pennington VELOCIRAPTOR The Velociraptor - meaning swift seizer - lived during the late Cretaceous period - 75-71 million years ago. They were a genus of dromaeosaurid theropod dinosaur and there
Ceri Pennington VELOCIRAPTOR The Velociraptor - meaning swift seizer - lived during the late Cretaceous period - 75-71 million years ago. They were a genus of dromaeosaurid theropod dinosaur and there
.56 m. (22 in.). COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition
. COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition") Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Dilong Paradoxus. (DIE-long) Early Tyrannosaurid Found in the Middle Jurassic to the Early Cretaceous. Claudia Montilla
 Early Tyrannosaurid Found in the Middle Jurassic to the Early Cretaceous. Claudia Montilla") Dilong Paradoxus (DIE-long) Early Tyrannosaurid Found in the Middle Jurassic to the Early Cretaceous Claudia Montilla William Parker BRY Trilobites to T-Rex Dilong Paradoxus 2 Abstract Dilong paradoxus,
Dilong Paradoxus (DIE-long) Early Tyrannosaurid Found in the Middle Jurassic to the Early Cretaceous Claudia Montilla William Parker BRY Trilobites to T-Rex Dilong Paradoxus 2 Abstract Dilong paradoxus,
Name. Compare the bones found in the foot, as well as the number of digits.
 MAMMALOGY LAB 4 LIMBS & LOCOMOTION Today s exercise focuses on the variation in limbs and lifestyles of mammals. You will be interpreting the lifestyles of a number of mammals based on various aspects
MAMMALOGY LAB 4 LIMBS & LOCOMOTION Today s exercise focuses on the variation in limbs and lifestyles of mammals. You will be interpreting the lifestyles of a number of mammals based on various aspects
University of Bristol - Explore Bristol Research. Peer reviewed version. Link to published version (if available): /pala.
: /pala.") Saitta, E. T., Gelernter, R., & Vinther, J. (2018). Additional information on the primitive contour and wing feathering of paravian dinosaurs. Palaeontology, 61(2), 273-288. DOI: 10.1111/pala.12342 Peer
Saitta, E. T., Gelernter, R., & Vinther, J. (2018). Additional information on the primitive contour and wing feathering of paravian dinosaurs. Palaeontology, 61(2), 273-288. DOI: 10.1111/pala.12342 Peer
Modeling and Control of Trawl Systems
 Modeling and Control of Trawl Systems Karl-Johan Reite, SINTEF Fisheries and Aquaculture Supervisor: Professor A. J. Sørensen * Advisor: Professor H. Ellingsen * * Norwegian University of Science and Technology
Modeling and Control of Trawl Systems Karl-Johan Reite, SINTEF Fisheries and Aquaculture Supervisor: Professor A. J. Sørensen * Advisor: Professor H. Ellingsen * * Norwegian University of Science and Technology
AMERICAN NATURALIST. Vol. IX. -DECEMBER, No. 12. OR BIRDS WITH TEETH.1 OI)ONTORNITHES,
ONTORNITHES,") AMERICAN NATURALIST. Vol. IX. -DECEMBER, 1875.-No. 12. OI)ONTORNITHES, OR BIRDS WITH TEETH.1 BY PROFESSOR 0. C. MARSH. REMAINS of birds are amono the rarest of fossils, and few have been discovered except
AMERICAN NATURALIST. Vol. IX. -DECEMBER, 1875.-No. 12. OI)ONTORNITHES, OR BIRDS WITH TEETH.1 BY PROFESSOR 0. C. MARSH. REMAINS of birds are amono the rarest of fossils, and few have been discovered except
Evolution as Fact. The figure below shows transitional fossils in the whale lineage.
 Evolution as Fact Evolution is a fact. Organisms descend from others with modification. Phylogeny, the lineage of ancestors and descendants, is the scientific term to Darwin's phrase "descent with modification."
Evolution as Fact Evolution is a fact. Organisms descend from others with modification. Phylogeny, the lineage of ancestors and descendants, is the scientific term to Darwin's phrase "descent with modification."
Field Trip: Harvard Museum of Natural History (HMNH)
") Field Trip: Harvard Museum of Natural History (HMNH) Objectives To observe the diversity of animals. To compare and contrast the various adaptations, body plans, etc. of the animals found at the HMNH.
Field Trip: Harvard Museum of Natural History (HMNH) Objectives To observe the diversity of animals. To compare and contrast the various adaptations, body plans, etc. of the animals found at the HMNH.
Evolution on Exhibit Hints for Teachers
 1 Evolution on Exhibit Hints for Teachers This gallery activity explores a variety of evolution themes that are well illustrated by gallery specimens and exhibits. Each activity is aligned with the NGSS
1 Evolution on Exhibit Hints for Teachers This gallery activity explores a variety of evolution themes that are well illustrated by gallery specimens and exhibits. Each activity is aligned with the NGSS
What is the evidence for evolution?
 What is the evidence for evolution? 1. Geographic Distribution 2. Fossil Evidence & Transitional Species 3. Comparative Anatomy 1. Homologous Structures 2. Analogous Structures 3. Vestigial Structures
What is the evidence for evolution? 1. Geographic Distribution 2. Fossil Evidence & Transitional Species 3. Comparative Anatomy 1. Homologous Structures 2. Analogous Structures 3. Vestigial Structures
TWO NEW PINE-FEEDING SPECIES OF COLEOTECHNITES ( GELECHIIDAE )
") Journal of the Lepidopterists' Society 32(2), 1978, 118-122 TWO NEW PINE-FEEDING SPECIES OF COLEOTECHNITES ( GELECHIIDAE ) RONALD W. HODGES l AND ROBERT E. STEVENS2 ABSTRACT. Two new species of moths,
Journal of the Lepidopterists' Society 32(2), 1978, 118-122 TWO NEW PINE-FEEDING SPECIES OF COLEOTECHNITES ( GELECHIIDAE ) RONALD W. HODGES l AND ROBERT E. STEVENS2 ABSTRACT. Two new species of moths,
Tetrapod Similarites The Origins of Birds
 Tetrapod Similarites The Origins of Birds Birds Reptiles Mammals Integument Feathers, scales Scales Hair Digestive Horny bill Teeth Teeth Skeletal Fusion of bones Some fusion Some fusion Reduction in number
Tetrapod Similarites The Origins of Birds Birds Reptiles Mammals Integument Feathers, scales Scales Hair Digestive Horny bill Teeth Teeth Skeletal Fusion of bones Some fusion Some fusion Reduction in number
Darwin and the Family Tree of Animals
 Darwin and the Family Tree of Animals Note: These links do not work. Use the links within the outline to access the images in the popup windows. This text is the same as the scrolling text in the popup
Darwin and the Family Tree of Animals Note: These links do not work. Use the links within the outline to access the images in the popup windows. This text is the same as the scrolling text in the popup
Evolution of Birds. Summary:
 Oregon State Standards OR Science 7.1, 7.2, 7.3, 7.3S.1, 7.3S.2 8.1, 8.2, 8.2L.1, 8.3, 8.3S.1, 8.3S.2 H.1, H.2, H.2L.4, H.2L.5, H.3, H.3S.1, H.3S.2, H.3S.3 Summary: Students create phylogenetic trees to
Oregon State Standards OR Science 7.1, 7.2, 7.3, 7.3S.1, 7.3S.2 8.1, 8.2, 8.2L.1, 8.3, 8.3S.1, 8.3S.2 H.1, H.2, H.2L.4, H.2L.5, H.3, H.3S.1, H.3S.2, H.3S.3 Summary: Students create phylogenetic trees to
CHIHUAHUA (Chihuahueño)
") FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) SECRETARIAT GENERAL: 13, Place Albert 1 er B 6530 Thuin (Belgique) FCI-Standard N 218 / 21.10.2009 / GB CHIHUAHUA (Chihuahueño) 2 TRANSLATION: C. Seidler.
FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) SECRETARIAT GENERAL: 13, Place Albert 1 er B 6530 Thuin (Belgique) FCI-Standard N 218 / 21.10.2009 / GB CHIHUAHUA (Chihuahueño) 2 TRANSLATION: C. Seidler.
As a rebuttal to Darwin s (1859) explanation of the
 explanation of the") What Use Is Half a Wing in the Ecology and Evolution of Birds? KENNETH P. DIAL, ROSS J. RANDALL, AND TERRY R. DIAL The use of incipient wings during ontogeny in living birds reveals not only the function
What Use Is Half a Wing in the Ecology and Evolution of Birds? KENNETH P. DIAL, ROSS J. RANDALL, AND TERRY R. DIAL The use of incipient wings during ontogeny in living birds reveals not only the function
Outline 17: Reptiles and Dinosaurs
 Outline 17: Reptiles and Dinosaurs Evolution of Reptiles The first reptiles appeared in the Mississippian. They evolved from amphibians, which first appeared in the Devonian. The evolutionary jump was
Outline 17: Reptiles and Dinosaurs Evolution of Reptiles The first reptiles appeared in the Mississippian. They evolved from amphibians, which first appeared in the Devonian. The evolutionary jump was
Do the traits of organisms provide evidence for evolution?
 PhyloStrat Tutorial Do the traits of organisms provide evidence for evolution? Consider two hypotheses about where Earth s organisms came from. The first hypothesis is from John Ray, an influential British
PhyloStrat Tutorial Do the traits of organisms provide evidence for evolution? Consider two hypotheses about where Earth s organisms came from. The first hypothesis is from John Ray, an influential British
Remains of the pterosaur, a cousin of the dinosaur, are found on every continent. Richard Monastersky reports
 Reading Practice Remains of the pterosaur, a cousin of the dinosaur, are found on every continent. Richard Monastersky reports PTEROSAURS Remains of the pterosaur, a cousin of the dinosaur, are found on
Reading Practice Remains of the pterosaur, a cousin of the dinosaur, are found on every continent. Richard Monastersky reports PTEROSAURS Remains of the pterosaur, a cousin of the dinosaur, are found on
What is a dinosaur? Reading Practice
 Reading Practice What is a dinosaur? A. Although the name dinosaur is derived from the Greek for "terrible lizard", dinosaurs were not, in fact, lizards at all. Like lizards, dinosaurs are included in
Reading Practice What is a dinosaur? A. Although the name dinosaur is derived from the Greek for "terrible lizard", dinosaurs were not, in fact, lizards at all. Like lizards, dinosaurs are included in
ALFRED GILLETT AND FOSSILS FROM STREET
 ALFRED GILLETT AND FOSSILS FROM STREET This collection of local fossils was formerly in the Crispin Hall, Street. Most of these fossils came from Alfred Gillett (1814-1904), a retired ironmonger who lived
ALFRED GILLETT AND FOSSILS FROM STREET This collection of local fossils was formerly in the Crispin Hall, Street. Most of these fossils came from Alfred Gillett (1814-1904), a retired ironmonger who lived
Fish 2/26/13. Chordates 2. Sharks and Rays (about 470 species) Sharks etc Bony fish. Tetrapods. Osteichthans Lobe fins and lungfish
 Sharks etc Bony fish. Tetrapods. Osteichthans Lobe fins and lungfish") Chordates 2 Sharks etc Bony fish Osteichthans Lobe fins and lungfish Tetrapods ns Reptiles Birds Feb 27, 2013 Chordates ANCESTRAL DEUTEROSTOME Notochord Common ancestor of chordates Head Vertebral column
Chordates 2 Sharks etc Bony fish Osteichthans Lobe fins and lungfish Tetrapods ns Reptiles Birds Feb 27, 2013 Chordates ANCESTRAL DEUTEROSTOME Notochord Common ancestor of chordates Head Vertebral column
2 nd Term Final. Revision Sheet. Students Name: Grade: 11 A/B. Subject: Biology. Teacher Signature. Page 1 of 11
 2 nd Term Final Revision Sheet Students Name: Grade: 11 A/B Subject: Biology Teacher Signature Page 1 of 11 Nour Al Maref International School Riyadh, Saudi Arabia Biology Worksheet (2 nd Term) Chapter-26
2 nd Term Final Revision Sheet Students Name: Grade: 11 A/B Subject: Biology Teacher Signature Page 1 of 11 Nour Al Maref International School Riyadh, Saudi Arabia Biology Worksheet (2 nd Term) Chapter-26
CLADISTICS Student Packet SUMMARY Phylogeny Phylogenetic trees/cladograms
 CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin
 Chinese Science Bulletin 2008 SCIENCE IN CHINA PRESS ARTICLES Springer A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin XU Xing 1, ZHAO Qi 1, NORELL Mark 2, SULLIVAN
Chinese Science Bulletin 2008 SCIENCE IN CHINA PRESS ARTICLES Springer A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin XU Xing 1, ZHAO Qi 1, NORELL Mark 2, SULLIVAN
NREM/ZOOL 4464 Ornithology Dr. Tim O Connell Lectures February, 2015
 NREM/ZOOL 4464 Ornithology Dr. Tim O Connell Lectures 12 14 9 13 February, 2015 Modern hierarchy of life on earth: Domain Kingdom Phylum (plural phyla ) Class Order Family Genus (plural genera ) Species
NREM/ZOOL 4464 Ornithology Dr. Tim O Connell Lectures 12 14 9 13 February, 2015 Modern hierarchy of life on earth: Domain Kingdom Phylum (plural phyla ) Class Order Family Genus (plural genera ) Species
Birds & Mammals. Chapter 15
 Birds & Mammals Chapter 15 What is a Bird? Vertebrate Endothermic Feathered 4 chambered heart Egg laying Fore-limbs adapted for flight Bones nearly hollow (allow for lighter weight) Bird Internal Anatomy
Birds & Mammals Chapter 15 What is a Bird? Vertebrate Endothermic Feathered 4 chambered heart Egg laying Fore-limbs adapted for flight Bones nearly hollow (allow for lighter weight) Bird Internal Anatomy
GROUP III WORKING DOGS III-29. Siberian Husky
 GROUP III WORKING DOGS III-29 Siberian Husky Origin and Purpose The Siberian Husky was developed untold centuries ago by the Chukchis of Northern Siberia. The Chukchis, a nomadic people, utilized their
GROUP III WORKING DOGS III-29 Siberian Husky Origin and Purpose The Siberian Husky was developed untold centuries ago by the Chukchis of Northern Siberia. The Chukchis, a nomadic people, utilized their
Title: Phylogenetic Methods and Vertebrate Phylogeny
 Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Lower Cretaceous Kwanmon Group, Northern Kyushu
 Bull. Kitakyushu Mus. Nat. Hist., 11: 87-90. March 30, 1992 A New Genus and Species of Carnivorous Dinosaur from the Lower Cretaceous Kwanmon Group, Northern Kyushu Yoshihiko Okazaki Kitakyushu Museum
Bull. Kitakyushu Mus. Nat. Hist., 11: 87-90. March 30, 1992 A New Genus and Species of Carnivorous Dinosaur from the Lower Cretaceous Kwanmon Group, Northern Kyushu Yoshihiko Okazaki Kitakyushu Museum
古脊椎动物学报 VERTEBRATA PALASIATICA. Corwin SULLIVAN
 第 52 卷第 1 期 2014 年 1 月 古脊椎动物学报 VERTEBRATA PALASIATICA pp. 3-30 figs. 1-9 Reinterpretation of the Early Cretaceous maniraptoran (Dinosauria: Theropoda) Zhongornis haoae as a scansoriopterygid-like non-avian,
第 52 卷第 1 期 2014 年 1 月 古脊椎动物学报 VERTEBRATA PALASIATICA pp. 3-30 figs. 1-9 Reinterpretation of the Early Cretaceous maniraptoran (Dinosauria: Theropoda) Zhongornis haoae as a scansoriopterygid-like non-avian,
A new carnosaur from Yongchuan County, Sichuan Province
 A new carnosaur from Yongchuan County, Sichuan Province by Dong Zhiming Institute of Vertebrate Palaeontology and Palaeoanthropology, Academia Sinica Zhang Yihong, Li Xuanmin, and Zhou Shiwu Chongqing
A new carnosaur from Yongchuan County, Sichuan Province by Dong Zhiming Institute of Vertebrate Palaeontology and Palaeoanthropology, Academia Sinica Zhang Yihong, Li Xuanmin, and Zhou Shiwu Chongqing
muscles (enhancing biting strength). Possible states: none, one, or two.
. Possible states: none, one, or two.") Reconstructing Evolutionary Relationships S-1 Practice Exercise: Phylogeny of Terrestrial Vertebrates In this example we will construct a phylogenetic hypothesis of the relationships between seven taxa
Reconstructing Evolutionary Relationships S-1 Practice Exercise: Phylogeny of Terrestrial Vertebrates In this example we will construct a phylogenetic hypothesis of the relationships between seven taxa
Fossils explained 53
 Fossils explained 53 Titans of the skies: azhdarchid pterosaurs Pterosaurs, the flying reptiles of the Mesozoic, often play second fiddle in popularity to their contemporaries, the dinosaurs. Such treatment
Fossils explained 53 Titans of the skies: azhdarchid pterosaurs Pterosaurs, the flying reptiles of the Mesozoic, often play second fiddle in popularity to their contemporaries, the dinosaurs. Such treatment
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Examining Bird Adaptations
 Biology I Unit V: Zoology Chapter 25-28 & DOL: Vertebrates Examining Bird Adaptations Lab Name: Date: Hour: Examining Bird Adaptations Pre-Lab Discussion There are many different types of birds. Each type
Biology I Unit V: Zoology Chapter 25-28 & DOL: Vertebrates Examining Bird Adaptations Lab Name: Date: Hour: Examining Bird Adaptations Pre-Lab Discussion There are many different types of birds. Each type
Phylogeny Reconstruction
 Phylogeny Reconstruction Trees, Methods and Characters Reading: Gregory, 2008. Understanding Evolutionary Trees (Polly, 2006) Lab tomorrow Meet in Geology GY522 Bring computers if you have them (they will
Phylogeny Reconstruction Trees, Methods and Characters Reading: Gregory, 2008. Understanding Evolutionary Trees (Polly, 2006) Lab tomorrow Meet in Geology GY522 Bring computers if you have them (they will
Abstract. The pterosaur Quetzalcoatlus is analyzed and discussed. Bone structure is looked
 Abstract The pterosaur Quetzalcoatlus is analyzed and discussed. Bone structure is looked at as part of this analysis. Wing structure gives insight on to how Quetzalcoatlus may have flown, and how different
Abstract The pterosaur Quetzalcoatlus is analyzed and discussed. Bone structure is looked at as part of this analysis. Wing structure gives insight on to how Quetzalcoatlus may have flown, and how different
Theoretical Morphology and Development of Flight Feather Vane Asymmetry with Experimental Tests in Parrots
 RESEARCH ARTICLE Theoretical Morphology and Development of Flight Feather Vane Asymmetry with Experimental Tests in Parrots TERESA J. FEO 1,2 * AND RICHARD O. PRUM 1,2 1 Department of Ecology and Evolutionary
RESEARCH ARTICLE Theoretical Morphology and Development of Flight Feather Vane Asymmetry with Experimental Tests in Parrots TERESA J. FEO 1,2 * AND RICHARD O. PRUM 1,2 1 Department of Ecology and Evolutionary
Crocs and Birds as Dino models Crocs and birds united with dinos by morphology Both also have parental care and vocal communication between offspring
 Chapter 16. Mesozoic Diapsids Phylogenetic relationships Earliest from late carboniferous stem diapsids Petrolacosaurus Lineage split into two: Archosauromorpha Crocs, birds, dinos, pterosaurs Lepidosauromorpha
Chapter 16. Mesozoic Diapsids Phylogenetic relationships Earliest from late carboniferous stem diapsids Petrolacosaurus Lineage split into two: Archosauromorpha Crocs, birds, dinos, pterosaurs Lepidosauromorpha
A new species of sauropod, Mamenchisaurus anyuensis sp. nov.
 A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
Pre-Archaeopteryx coelurosaurian dinosaurs and their implications for understanding avian origins
 Invited Review Geology December 2010 Vol.55 No.35: 3971 3977 doi: 10.1007/s11434-010-4150-z SPECIAL TOPICS: Pre-Archaeopteryx coelurosaurian dinosaurs and their implications for understanding avian origins
Invited Review Geology December 2010 Vol.55 No.35: 3971 3977 doi: 10.1007/s11434-010-4150-z SPECIAL TOPICS: Pre-Archaeopteryx coelurosaurian dinosaurs and their implications for understanding avian origins
Modern taxonomy. Building family trees 10/10/2011. Knowing a lot about lots of creatures. Tom Hartman. Systematics includes: 1.
 Modern taxonomy Building family trees Tom Hartman www.tuatara9.co.uk Classification has moved away from the simple grouping of organisms according to their similarities (phenetics) and has become the study
Modern taxonomy Building family trees Tom Hartman www.tuatara9.co.uk Classification has moved away from the simple grouping of organisms according to their similarities (phenetics) and has become the study
ARE CURRENT CRITIQUES OF THE THEROPOD ORIGIN OF BIRDS SCIENCE? REBUTTAL TO FEDUCCIA (2002)
") Commentary The Auk 120(2):550 561, 2003 ARE CURRENT CRITIQUES OF THE THEROPOD ORIGIN OF BIRDS SCIENCE? REBUTTAL TO FEDUCCIA (2002) Department of Ecology and Evolutionary Biology, and Natural History Museum,
Commentary The Auk 120(2):550 561, 2003 ARE CURRENT CRITIQUES OF THE THEROPOD ORIGIN OF BIRDS SCIENCE? REBUTTAL TO FEDUCCIA (2002) Department of Ecology and Evolutionary Biology, and Natural History Museum,