University of Bristol - Explore Bristol Research. Peer reviewed version. Link to published version (if available): /pala.
|
|
|
- Clarissa Lyons
- 5 years ago
- Views:
Transcription
: 10.1111/pala.")
1 Saitta, E. T., Gelernter, R., & Vinther, J. (2018). Additional information on the primitive contour and wing feathering of paravian dinosaurs. Palaeontology, 61(2), DOI: /pala Peer reviewed version Link to published version (if available): /pala Link to publication record in Explore Bristol Research PDF-document This is the author accepted manuscript (AAM). The final published version (version of record) is available online via Wiley at Please refer to any applicable terms of use of the publisher. University of Bristol - Explore Bristol Research General rights This document is made available in accordance with publisher policies. Please cite only the published version using the reference above. Full terms of use are available:
2 Additional information on the primitive contour and wing feathering of paravian dinosaurs Evan T. Saitta 1, Rebecca Gelernter 2, & Jakob Vinther 1,3 1 School of Earth Sciences, University of Bristol, Bristol, United Kingdom; s: evansaitta@gmail.com (ORCiD ID: orcid.org/ ), jakob.vinther@bristol.ac.uk (ORCiD ID: orcid.org/ ) 2 Near Bird Studios, New Haven, Connecticut, USA; rebecca@nearbirdstudios.com 3 School of Biological Sciences, University of Bristol, Bristol, United Kingdom ABSTRACT: Identifying feather morphology in extinct dinosaurs is challenging due to dense overlapping of filaments within fossilized plumage and the fact that some extinct feather morphologies are unlike those seen in extant birds or those predicted from an evo-devo model of feather evolution. Here, comparisons are drawn between a range of dinosaur taxa with preserved integumentary appendages using high resolution photographs to better understand fossil feather morphology and gain insight into their function and evolution. A specimen of the basal paravian Anchiornis possesses contour feathers disarticulated from the plumage, revealing a novel feather type much simpler than the contour feathers of most extant birds a shaggy, open-vaned, bifurcated feather with long barbs attached to a short rachis. In contrast, the contour feathers of the Sinosauropteryx contour feathers are likely simpler than those seen in Anchiornis; a tuft morphology of multiple barbs connected at their bases (e.g. via a shared follicle), but lacking a rachis, is tentatively preferred. However, unless isolated Sinosauropteryx contour feathers are discovered in the manner of the Anchiornis specimen, conclusive morphological descriptions will
3 remain difficult. In addition to contour feathers, preserved paravian wing feathers also show potentially plesiomorphic traits. Comparison with Confuciusornis suggests that Anchiornis wing feathers were at least partially open-vaned. Combined with the interpretation of Anchiornis contour feathers, this suggests that differentiated barbicels are relatively derived compared to pennaceous feathers and the appearance of wings. Shaggy contour feathers likely influenced thermoregulatory and water repellence abilities, and in combination with open-vaned wing feathers, would have decreased aerodynamic efficiency. Simplified, open-vaned wing feathers were also observed on the oviraptorosaur Caudipteryx, consistent with, but not necessarily diagnostic of, its suggested flightlessness. Taken together, these observations have broad implications for how we depict a wide variety of dinosaurs and how we view the function and evolution of feathers in these taxa. KEY WORDS: dinosaur, theropod, paravian, contour feathers, wing feathers, plesiomorphy, barbules
4 INITIAL descriptions of feathered non-avian dinosaurs (e.g. Chen et al. 1998; Xu et al. 1999, 2000) have been followed by a flurry of fossil discoveries preserving integumentary soft tissues in this group (e.g. Norell & Xu 2005; Xu 2006; Hu et al. 2009; Zelenitsky et al. 2012). Filamentous integumentary structures have even been found on ornithischian dinosaurs (Mayr et al. 2002; Zheng et al. 2009a; Godefroit et al. 2014), although the homology of such structures to true feathers has not been determined (Barrett et al. 2015; Mayr et al. 2016). The evolution of feathers has become a major research topic in evolutionary biology and palaeontology. A commonly cited model for feather evolution is based on an understanding from feather development in modern birds, the evo-devo approach (Prum & Brush 2002), but fossil feathers that do not match expected morphologies based on feather development (Zhang et al. 2008) show that such a model for feather evolution, while useful, is likely overly simplified and that extinct feather morphologies existed, distinct from modern feathers. Determining the morphology of a single integumentary appendage can be difficult when such structures are preserved as dense plumage around a specimen. Descriptions can also become outdated with the discovery of more specimens that provide novel or clearer morphological details. Improved understanding of fossil feather morphology provides better insight into their function with implications for the evolution of avian flight, a key adaptation. This is important as many bizarre forms of non-avian theropods appear to have evolved aerial locomotion (e.g. gliding) prior to the evolution of the modern bird body plan, such as four-winged forms like Microraptor or Anchiornis (Xu et al. 2003; Hu et al. 2009; Palmer 2014) or even those with proposed membranous wings (Xu et al. 2015). Moreover, it is possible that some non-avian theropods, like Caudipteryx or short-armed or large dromaeosaurs, might even have been secondarily flightless (Feduccia 1999; Jones et al. 2000; Paul 2002; Zheng et al. 2009b; Lü and Brusatte 2015; Mayr 2017, but see
5 Dyke and Norell 2005; Dececchi et al for counter-positions). However, it should be noted that there is a lack of consensus on secondary flightlessness in non-avian theropods as pennaceous wings have been hypothesized to have evolved prior to biomechanical functional usage (Zelenitsky et al. 2012). It has been suggested that subsequent locomotory functions of the wing might have involved pre-aerial locomotion such as flap running, wing-assisted incline running, and wingassisted leaping (Heers et al. 2014; Dececchi et al. 2016). Beyond investigating the evolution of flight, body contour feather evolution is particularly important. Contour feathers evolved prior to flight feathers as seen by taxa like Sinosauropteryx, which possess filamentous structures around the majority of its body but lack rectrices and remiges as well as skeletal adaptations for aerial locomotion (Chen et al. 1998; Currie & Chen 2001). Contour feathers also play key functional roles beyond aerodynamic streamlining such as thermoregulation, water repellence, or display (Lucas & Stettenheim 1972). Thus, understanding the evolution of contour feathers should provide insight into the function of the earliest feathers and the palaeobiology of extinct species beyond aerodynamic capability. The main goal of this study is to better understand fossil feathers in non-avian theropod dinosaurs. In providing new feather dara from a phylogenetic range of fossil dinosaurs, we aim to identify potential plesiomorphies, highlight challenges in their study, and gain greater insight into paravian feather function and evolution. MATERIAL AND METHODS High resolution photographs of several non-avian and avian dinosaur taxa with preserved integumentary structures were taken using a Nikon D800 camera and a 60 or 105 mm macro
6 Nikkor lens in a crossed polarised light configuration with a Lowell Totalight 800W or 400W tungsten bulb: Psittacosaurus (SMF R 4970), Sinosauropteryx (NIGP , NIGP ), Caudipteryx (IVPP V12344, IVPP V12430), and Confuciusornis (IVPP V13156). Anchiornis (BMNHC PH828) was photographed using the 60 mm macro Nikkor lens on a Nikon D90 under normal incandescent light at different angles. The taxa represent a phylogenetically broad sampling within Dinosauria. Psittacosaurus and Confuciusornis represent taxa whose integumentary appendages can be thought of as fairly well understood end-members of feather-like integument evolution. Psittacosaurus had long, simple, non-branching, bristle-like filaments that potentially occurred in clusters (Mayr et al. 2016), and Confuciusornis is expected to have relatively more derived feathers compared to the more basal taxa (Chiappe et al. 1999; Prum & Brush 2002; Fucheng et al. 2006, but see Feo et al. 2015), at least with regards to its wing feathers. Importantly, the specimen of Anchiornis (BMNHC PH828) possesses isolated contour feathers that have been transported away from the rest of the plumage, allowing for easier interpretation of their morphology. The isolated contour feathers on this specimen were first figured by Li et al. (2010) in their supporting online material but were not thoroughly discussed. Using this specimen and the inclusion of relatively end-member examples, we hope to better elucidate the less understood feather morphology of non-avian theropods through comparison. Interpretive drawings are included to better portray certain observations. These also allow for testing the hypothesis that overlapping patterns of the plumage are influenced by the morphology (i.e., branching pattern) of a single integumentary structure. Fig. 1A F shows some of the studied specimens and the locations where detailed observations/comparisons of feathers were made.
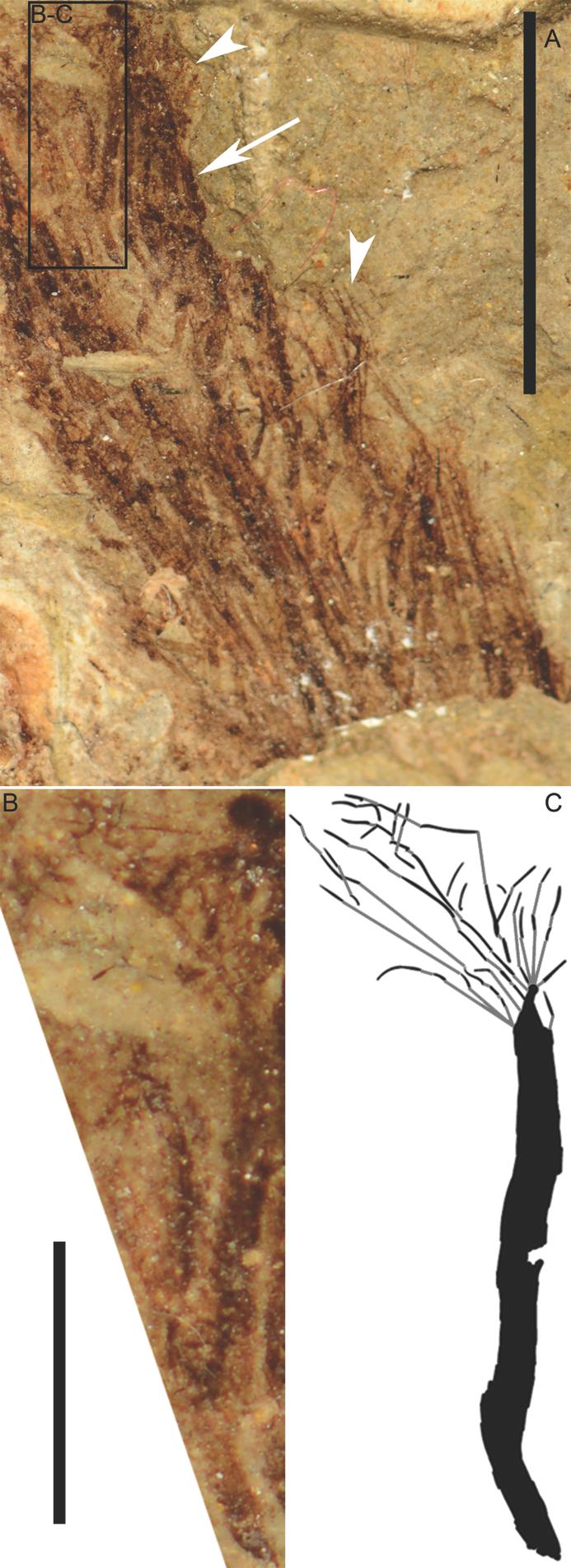
7 Contour feathers in Anchiornis and Sinosauropteryx are first discussed to highlight possible plesiomorphies among extinct contour feather morphologies and the challenges of interpreting articulated fossil plumage, followed by a discussion of potential plesiomorphies in dinosaurian wing feathers and how feather morphology in secondarily flightless taxa may provide ecological rather than evolutionary signals. The taphonomic history of these specimens, in which they are sub-aqueously buried, should not affect feather morphology as sediment works to keep tissues in place and feather barbs only clump together when removed from water, not upon submersion. Institutional abbreviations BMNHC, Beijing Museum of Natural History, Beijing, China; IVPP, Institute of Vertebrate Palaeontology and Palaeoanthropology, Beijing, China; NIGP, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Beijing, China; SMF, Forschungsinstitut Senckenberg, Frankfurt am Main, Germany RESULTS Anchiornis isolated contour feathers a novel morphotype Isolated contour feathers in the Anchiornis specimen BMNHC PH828, disarticulated and transported away from the rest of the plumage, reveal the morphology of these feathers more clearly than do those in the articulated plumage (Figs. 2A B, 3A E). Most of the isolated feathers
8 are about 1 2 cm long. They appear to show two vanes of long barbs positioned at low barb angles to a relatively short rachis in a presumably pennaceous configuration. Barbs extend significantly beyond the apical tip of the rachis, resulting in a shaggy morphology noticeably bifurcated towards its apical end in some of the isolated feathers. In those feathers that show significant bifurcation, the apical region (i.e. towards the apical tips of the barbs) are blunt or squared-off on each vane (Fig. 3A D), suggesting that barbs originating at different positions along the rachis terminate at a similar point and that the more basal barbs are longer than the distal barbs. The barbs do not always show tight association to each other and can diverge and strongly curve. This suggests an open vane lacking properly differentiated proximal and distal barbules. An open vane with flexible barbs, along with variation in the angle at which the feather is exposed in the matrix, could explain why some of the isolated contour feathers do not appear to bifurcate strongly at their apical ends, although a similar disparity in Sinornithosaurus was attributed to the presence of two contour feather morphotypes (Xu et al. 2001). One isolated feather has a trident appearance (Fig. 2A). Most likely, this feather is the disarticulated apical end of a secondary remex or major covert (these being located near the isolated contour feathers observed on the specimen). The trident appearance results from colour patterning in a feather whose morphology is consistent with the other remiges and major coverts. Sinosauropteryx non-isolated contour feathers interpreting highly primitive morphotypes With a better understanding of basal paravian Anchiornis contour feather morphology, the contour feathers of the basal coelurosaur Sinosauropteryx are re-examined. Comparing filament overlapping patterns within the articulated plumage between Sinosauropteryx and better
9 understood taxa like Psittacosaurus and Anchiornis (Fig. 4) shows how integumentary appendages with different branching patterns can lead to similar overlapping patterns within the articulated plumage. Psittacosaurus bristles have been suggested to be developmentally and/or structurally homologous at some level to bristle-like structures in other dinosaurs, Tianyulong and Beipiaosaurus (Xu et al. 1999; Zheng et al. 2009a; Mayr et al. 2016). The interpretation of Psittacosaurus bristle morphology here is based on that of Mayr et al. (2016). Non-branching Psittacosaurus bristles (Fig. 4A B) show filaments that curve in the same direction and manner. Near the skin, low angle, short filaments overlap higher angle, long filaments. Filaments originating more anteriorly along the tail can overlap more posterior filaments as they curve posteriorly. As a result, there is also an overlapping pattern further away from the skin where low angle, apical portions of anteriorly erupting bristles cross over the high angle, basal portions of posteriorly erupting bristles. Filaments of Sinosauropteryx contour feathers (Fig. 4C D) show many similar overlapping patterns, although the structures are much shorter and the plumage is denser. Like Psittacosaurus bristles, Sinosauropteryx filaments separate from each other and become more easily distinguishable apically. Theoretically, simple, singular, non-branching filaments could create such overlapping patterns that might give the illusion of branching integumentary structures, especially if they are arranged in multiple tracts on the body (Saitta et al. 2017a). However, filament orientations of contour feathers on the hindlimb and pes of Anchiornis (present in addition to remiges/major coverts on the hindlimbs) also show a similar overlapping pattern and are much closer in size to Sinosauropteryx feathers (Fig. 4E F). Anchiornis contour feather morphology was determined above by examining isolated feathers near the torso (Fig. 2),
10 a method inapplicable to known Sinosauropteryx specimens. There are additional isolated contour feathers near the pes consistent with the interpreted morphology. Therefore, the similar overlapping pattern seen in Psittacosaurus, Sinosauropteryx, and Anchiornis results from a range of integumentary appendage morphologies. Based on overlapping alone, Sinosauropteryx contour feathers might represent one of a range of plausible morphologies from simple, singular, non-branching filaments, to nonbranching filaments connected at their bases (e.g. via a calamus or shared follicle), to a morphology seen in Anchiornis with multiple barbs connected to a short rachis. Further observations are therefore needed and may benefit from additional imaging using new techniques such as the laser stimulated fluorescence (LSF) used on Psittacosaurus (Mayr et al. 2016) and other integumentbearing non-avian dinosaurs (Wang et al. 2017a; Xu et al. 2017). As in original descriptions (Chen et al. 1998; Currie & Chen 2001), this study noticed a mixture of thick and thin strands close to the body (Fig. 4C D). However: 1. Some areas show relatively thick strands away from the body. Thick strands away from the body are likely due to close-lying or overlapping thinner filaments. 2. Unlike previous descriptions (Chen et al. 1998; Currie & Chen 2001), streaks near the base are not always positioned at a higher angle (although thicker strands near the base do tend to be at high angles). Low angle strands near the body could be the apical ends of feathers that erupt from relatively lateral tracts. 3. Furthermore, finer strands often appear parallel to each other and usually curve posteriorly rather than branching off equally from larger strands in opposing directions as previously described (Currie & Chen 2001).
11 These observations might suggest that relatively simple feathers (e.g. lacking a rachis) are possible for Sinosauropteryx. One perplexing observation was of several adjacent thick streaks away from the body dorsal to the cervical vertebrae in NIGP (Fig. 5A C). They resemble modern developing feathers contained in a sheath where the apical-most barbs separate as they erupt from the sheath. Whether such a developmental pattern might suggest a relatively complex morphology in which a rachis is present is unknown. However, some of these streaks appear to curve at their basal ends, which might be less likely to occur if they were encased in a sheath (although such a sheath could have been relatively pliable). It is also unusual that several adjacent feathers would all be growing/molting in one specific region on such a large specimen. Finally, no traces of calcium phosphate are visible under normal light photography which would give strong evidence for the presence of a calcified, stiffened rachis or sheath (Pautard 1963; Vinther et al. 2016; Saitta et al. 2017b). Other fossil coelurosaurs with similar structures have been suggested to preserve developing feathers (Prum 2010). This study corroborates previous descriptions (Chen et al. 1998; Currie & Chen 2001) of regularly spaced, high angle, thick streaks close to the body (Fig. 4C D). These thick streaks suggest a morphology where multiple barbs attach at their bases but lack a rachis. Multiple filaments are present in clusters or tufts, merging downward to a basal point. The tuft is continuous to the base, suggesting that the barbs are not attached to a short rachis. If there was a short rachis, filaments would appear to emanate from a narrow strip. Instead, filaments do not branch from a point distal to the base. The thick basal region is of uniform thickness throughout, and at the distal end, it separates into smaller filaments. The Sinosauropteryx tuft (Fig. 3F G) starkly differs from isolated Anchiornis contour feathers (Fig. 3A E). Multiple filaments clumping
12 into tufts with a single basal origin also provide evidence against the prevalence of single, nonbranching filaments in the contour feathering of Sinosauropteryx. Beyond contours possible plesiomorphic traits in wing feathers Not only are Anchiornis contour feathers of interest, but their wing feathers also show unusual morphology. The remiges have fairly symmetric vanes as described by Hu et al. (2009) but some primary remiges show slight levels of curvature in the rachis (Fig. 6A B). The barbs within Anchiornis remiges and major coverts do not lie close together, especially at their tips (Fig. 6A E). As they extend away from the rachis, they separate, and this pattern is less noticeable in barbs originating very apically along the rachis. This suggests, at the very least, an open feather vane in the tips of barbs on the more basal regions of the remex and major covert. In light of the open vane of Anchiornis contour feathers, this suggests that, even in wing feathers, proximal and distal barb differentiation or functional barbicels were lacking. This interpretation is bolstered by comparison with the closed vanes of Confuciusornis remiges (Fig. 6F J). In these more derived feathers, barbs are closely positioned even as they extend away from the rachis, and adjacent barbs show similar patterns of displacement and curvature, indicating that they are zipped together via differentiated proximal and distal barbules. Caudipteryx simplified wing feathers the evolutionary significance of simplicity The wing feathers in Caudipteryx are very unusual and greatly differ from remiges of modern flying birds (Fig. 7). The remiges (or major coverts) of one specimen (IVPP V12344) have an open
13 vane where the barbs are independent and separate from each other (Fig. 7A B). The vanes are symmetric with a straight rachis. Another specimen (IVPP V12430) has feathers on its wings (presumably remiges or major coverts) with a more extreme open vane where long barbs curve flexibly and separate widely from each other to form a frond shape feather (Fig. 7C D). This specimen shows even more unusual structures near the distal end of its forelimb which might represent simplified contour feathers on the forelimb (Fig. 7E). They are disarticulated from the rest of the specimen and are long, curving, ribbon-like structures. Some potentially converge together at their base. Overall, the feathers on Caudipteryx wings are highly simplified compared to aerodynamically efficient modern flight feathers. DISCUSSION Anchiornis isolated contour feathers a novel morphotype Isolated contour feathers disarticulated from the plumage in Anchiornis allow for a much easier interpretation of their morphology. The shaggy, bifurcated morphology described here is a novel feather type (Fig. 8C) and further demonstrates how extinct feather morphologies prevent researchers from relying solely upon an evo-devo model of feather evolution. These feathers would have appeared open-vaned, and specialized barbicels like hooklets to differentiate proximal and distal barbules are possibly more derived characters than we may realize. Given that iridescence has been reported in paravians (Li et al. 2012), and iridescence is often produced in the barbules (Maia et al. 2011), it is plausible that Anchiornis did not lack barbules entirely (Li et al. 2010).
14 These contour feathers would have given at least adult Anchiornis, and possibly related non-avian theropods, a much shaggier or fuzzier appearance than typically depicted (Fig. 9), especially compared to most modern birds that have smooth, aerodynamic plumage (Fig. 8D E), keeping in mind that plumage could have varied through ontogeny. Some non-avian theropods potentially had a superficially similar plumage to modern flightless birds like kiwis, bearing in mind that morphology differed at the level of individual feathers. Another paravian, Serikornis, is preserved with dramatically long, dense contour plumage (Lefèvre et al. 2017), consistent with the shaggy appearance of paravian plumage suggested here. We hypothesize that the shaggy plumage of Anchiornis affected physiology by altering heat retention (relative to the combination of modern down and contour feathers), decreasing water repellence, and decreasing aerodynamic efficiency by increasing drag in comparison to modern feathers, which are known to effectively function in such roles (Lucas & Stettenheim 1972). These proposed functional hypotheses should be rigorously and quantitatively tested in the future. Anchiornis is a basal member of paraves and is very closely related to Aves, possibly suggesting that modern contour feathers are a feature exclusive to Avialae. Sinornithosaurus has been described with contour feather plumage consisting of two morphotypes. One morphology was described as multiple filaments connected to a central filament (Xu et al. 2001). Some of these contour feathers have become disarticulated and isolated from this specimen, and closely resemble those described here in Anchiornis. Therefore, Sinornithosaurus likely possessed the same shaggy, bifurcated contour feather morphology as Anchiornis a sensible conclusion given the fact that Sinornithosaurus is a dromaeosaur and, therefore, also a paravian. The contour feathers of the paravian Serikornis were described in a similar manner with two morphotypes present, one with bundles of filaments that are joined together proximally and remain nearly parallel as they
15 extend distally and the other with a well-defined rachis and transversely inserted barbs (Lefèvre et al. 2017). Some of the epidermal structures found associated with Yi, another taxa that has been placed within Pennaraptora (in a clade sister to Paraves), are also similar to the contour feathers observed here for Anchiornis (Xu et al. 2015, see Figure 2e within). Sinosauropteryx non-isolated contour feathers interpreting highly primitive morphotypes Given contour feather morphology in relatively derived theropods like Anchiornis, the basal coelurosaur Sinosauropteryx is also expected to show primitive contour feathers, however, a lack of isolated feather fossils makes morphological determinations challenging. Currie & Chen (2001) suggested that Sinosauropteryx feathers had central rachises and plumulaceous barbs based on the following observations: (1) a mixture of thin and thick strands close to the body; (2) thinner strands more frequent distally; (3) thick strands positioned close to the body and typically oriented at higher angles from the body than the more distal strands; (4) areas of many fine strands adjacent and parallel to each other and sometimes kinking together; and (5) finer strands tending to angle away on both sides from thicker structures. The observations made in this study largely agree with those of Currie & Chen (2001) with some differences. Regarding observations (1) and (3), this study also observed thicker filaments farther away from the body and some low angle streaks near the body. Regarding observation (5), this study observed that finer streaks tend to lie parallel and orient in common directions rather than branching of equally in different directions. Thinner strands being more frequent distally (2) could also occur in a simple, non-branching filament (e.g. Psittacosaurus) if the filaments taper in thickness apically or due to less filament overlapping away from the body resulting in more thin filaments being visible. Fine, parallel strands (4) do not
16 necessitate the presence of a rachis, although a rachis could organize strands via parallel barbs in a vane. A range of feather morphologies can produce similar overlapping patterns, contrary to the initial hypothesis that overlapping patterns are dictated by the morphology of the individual integumentary structures. This result confirms published claims about the difficulty in discerning a single feather s morphology within articulated plumage (Currie & Chen 2001) and highlights how important the disarticulated feathers on Anchiornis are to interpreting its contour feather morphology. Although this study largely supports the observations of Chen et al. (1998) and Currie & Chen (2001), it opens up the possibility that these feathers are actually morphologically simpler than originally thought (e.g. Fig. 8A B). Some observations might seem to suggest a relatively complex morphology with the presence of multiple barbs attached to a short rachis (e.g. regularly spaced, thick stains near the body where filaments converge or potential feathers erupting from a sheath). If correct, it could mean that the shaggy contour feather morphology of Anchiornis was widely distributed in coelurosaurs. However, such observations more strongly suggest a tuft of multiple barbs connected basally without a rachis. Amidst a range of plausible morphologies, we assign Sinosauropteryx contour feather morphology as, at least predominantly, a tuft of multiple barbs attached basally via a calamus or shared follicle based on the sum of the evidence of the specimens examined (Saitta et al. 2017a). Such a tuft represents an even more primitive feather morphology than seen in Anchiornis contour feathers. Tufts (i.e., stage II feathers) have been observed in Cretaceous amber (McKellar et al. 2011).
17 Overlapping integumentary appendages within articulated plumage can greatly obscure observations, meaning that the discovery of isolated contour feathers will likely be needed to conclusively assign a morphology to Sinosauropteryx feathers. Further emphasizing this point, is an indeterminate coelurosaurian tail segment preserved in amber with exceptional 3D preservation of the associated feathers (Xing et al. 2016a). The authors state, none of the observed osteological features preclude a compsognathid affinity. The feathers are weakly pennaceous and open-vaned with alternating barbs attached to a poorly-developed rachis and simple, undifferentiated barbules as well as rachidial barbules. Although evidence for close phylogenetic affinity with Sinosauropteryx is uncertain, the feathers are morphologically more complex than those predicted for Sinosauropteryx here. The second type of Sinornithosaurus contour feather morphology described is filaments joined in a basal tuft (Xu et al. 2001). This would be consistent with the morphology described here for Sinosauropteryx and would have given Sinornithosaurus an interesting combination of primitive and derived contour feather morphotypes within the plumage as well as begging the question as to whether other paravians like Anchiornis also possessed multiple contour feather morphotypes. However, the possibility that Sinornithosaurus tufts are actually misidentified feathers of the other described morphotype (i.e., the shaggy, bifurcated morphotype possessing a rachis) may need to be investigated given that they do somewhat resemble the other feathers on Sinornithosaurus. Beyond contours possible plesiomorphic traits in wing feathers
18 The symmetric wing feathers in Anchiornis suggest similar barb lengths between the leading and trailing vanes and a primitive condition whereby the barbs of the trailing vane are at low angles, which may have limited their aerodynamic efficiency (Feo et al. 2015). In addition to shaggy, open-vaned contour feathers, Anchiornis wing feathers appear to be at least partly open-vaned. A similar observation was made for the paravian Serikornis (Lefèvre et al. 2017). The wing feathers on the forelimb and hindlimb of this taxa appear to be open-vaned. Although Lefèvre et al. (2017) suggest that barbules were entirely lacking in Serikornis, this may be unlikely and represent an extreme interpretation, as discussed above where the presence of iridescence in paravians is consistent with the likely presence of barbules (Maia et al. 2011; Li et al. 2012). Evidence Lefèvre et al. (2017) cite includes a lack of visible barbs under optical and electron microscopy as well as a lack of basal expansions on the preserved barbs that indicated barbule insertions. However, such expansions are noted to be lacking in taxa believed to have barbules. Furthermore, carbonaceous preservation of keratinous structures like feathers derives from preserved pigments, so a lack of carbonaceously preserved barbules may simply indicate a lack of pigmented barbules in vivo or another taphonomic process that might eliminate fine morphological details such as weathering (Vinther 2015; Saitta et al. 2017b). The possibility of extinct, open-vaned feathers possessing barbules is in congruence with the indeterminate coelurosaurian tail preserved in amber that possesses undifferentiated barbules and is also weakly pennaceous and open-vaned (Xing et al. 2016a). Thus, Serikornis, and other paravians like Anchiornis, may have lacked functional barbicels/differentiated barbules, resulting in an open vane, rather than lacking barbules entirely. This hypothetically reduces aerodynamic capability and might further support differentiated proximal and distal barbules as relatively derived characters. Open-vaned wing
19 feathers suggest that non-avian theropods had reduced aerial locomotion ability compared to modern birds or even Mesozoic aves like Confuciusornis. Other lines of evidence have been invoked in support of this idea (e.g. Wang et al. 2011, 2017a), and gliding rather than powered flight has been proposed for four-winged non-avian theropods (e.g. Chatterjee & Templin 2007; Dyke et al. 2013, but see Dececchi et al. 2017). However, the question involves the aerodynamic functionality of the wing as a whole could open-vaned, or partially open-vaned, feathers arranged into a wing still be aerodynamically sufficient for aerial locomotion in a way that is unpredictable from a reductionist examination of individual feathers? Such a primitive vane condition may have been countered by a primitive wing arrangement in which multiple rows of major coverts extended far down the aerofoil of the wing in Anchiornis, as well as Archaeopteryx (Longrich et al. 2012, but see Nudds 2014 for a counter-position). Such extensive coverts may have helped to create a less permeable lift surface despite being composed of openvaned feathers, a feat accomplished in modern birds using tightly closed-vaned feathers but with fewer rows of such feathers in the aerofoil (i.e. less extensive coverts). This hypothesis is possibly evidenced by the fact that the major coverts in paravians like Anchiornis are very similar morphologically to the remiges both in overall size and shape, as well as in vane and barb morphology, suggesting similar functional roles. Furthermore, the presence of a propatagium in Anchiornis, and other paravians like Serikornis, may have also helped to compensate for aerial locomotion-related inefficiencies of the feathers and skeletomusculature (Wang et al. 2017a; Lefèvre et al. 2017), and the presence of hindlimb wings and tail rectrices in addition to forelimb wings might have compensated by increasing surface area for lift or stability during gliding (Xu et al. 2003; Chatterjee & Templin 2006; Alexander et al. 2010; Dyke et al. 2013; Koehl et al. 2011; Palmer 2014). Gliding capability prior to the evolution of functional barbicels and closed
20 vanes may suggest that aerodynamic functions of feathers could have predated socio-sexual display functions as the transition from filamentous feathers to closed-vane, pennaceous feathers has been suggested to have occurred through an open-vaned, pennaceous intermediate (Prum & Brush 2002), pennaceous feathers have been hypothesized to first function for display (Foth et al. 2014; Koschowitz et al. 2014), and such open-vaned, pennaceous feathers can be found in extant display feathers (Lucas & Stettenheim 1972). This question hinges upon determining whether certain, more basal non-avian theropods with pennaceous feathers represent primarily or secondarily flightless forms. The functional utility of wings containing partially open-vaned feathers can be elucidated by examining the silky trait in modern domesticated birds, such as pigeons and doves (Fig. 6K L). The silky allele is a recessive mutation and leads to aberrant, disarrayed barbule formation where barbules are brittle, easily broken, and fail to properly interlock. The resulting feathers, including remiges and rectrices, take on a more open-vaned appearance in the heterozygote resulting in reduced aerodynamic capability whereby flight is lost but low perches can be reached. Insulation is not affected but water repellence is decreased. Homozygous recessive individuals show an even more extreme morphology of aberrant barbules and open-vaned feathers and are totally incapable of aerial locomotion. Silky fowl, unlike the silky pigeon, lack barbicels entirely (Cole & Willard 1939; Miller 1956; Juhn & Bates 1960; Feng et al. 2014; Van Grouw 2016). The fact that the overall morphology of these heterozygous silky remiges closely resembles the open-vaned pattern seen in Anchiornis or Serikornis is further evidence that fullyfunctional barbicels were absent in these paravians and highlights the need to compensate for open-vaned wing feathers through other adaptations (e.g., multiple feather tiers on wing surface). Understanding silky feathers in modern birds may provide insight into the functional utility of
21 paravian feathers in aerial locomotion, thermoregulation, and water repellence as well as provide insight into the evo-devo of modern feathers given that the underlying genetics and development of the silky trait are well known (Cole & Willard 1939; Miller 1956; Juhn & Bates 1960; Feng et al. 2014; Van Grouw 2016). Open vanes are also found on the remiges of secondarily flightless ratites like ostriches and rheas as well as certain types of extant ornamental feathers (Lucas & Stettentheim 1972). Caudipteryx simplified wing feathers the evolutionary significance of simplicity Extinct feather morphologies give us insight into the macroevolution of feathers as they became adapted for modern avian flight. However, there is potential that rapidly-evolving ecological signals might confound broader macroevolutionary signals. Secondary flightlessness has been proposed for Caudipteryx (Feduccia 1999; Jones et al. 2000; Paul 2002; Mayr 2017, but see Dyke and Norell 2005; Dececchi et al for counter-positions). Caudipteryx wing feathers are highly simplified compared to those of flying birds, extant and extinct (Lucas & Stettenheim 1972). Remiges were likely open-vaned, and wing feathers appeared broad and frond shaped. Some more perplexing feathers are ribbon-like. If consensus can be built to demonstrate that some nonavian dinosaurs were secondarily flightless, then consideration must be taken when using their feather morphologies to reconstruct feather macroevolution with regards to the appearance of fully modern feathers and flight. CONCLUSIONS
22 Isolated Anchiornis contour feathers reveal a novel, extinct feather morphology, consisting of a short rachis with long, low-angle barbs in a bifurcated configuration. Functional barbicels appear to have been lacking, resulting in an open vane. Sinosauropteryx had simpler contour feather morphology than previously thought (i.e. lacking a rachis) and in comparison to Anchiornis. This study tentatively assigns a tuft morphology to Sinosauropteryx contour feathers consisting of multiple barbs attached at their bases. However, conclusive interpretations likely require the discovery of isolated Sinosauropteryx feathers. Anchiornis wing feathers appear at least partially open-vaned. These observations change the way we depict paravians (i.e. with a much 'shaggier' appearance), and hints at their palaeobiology (e.g. reduced aerodynamic capabilities). When examining feather evolution in the future, the impact of potential secondary flightlessness on feather morphology also needs to be more carefully considered if support for secondary flightlessness in non-avian dinosaurs or basal avialans can be demonstrated based on multiple lines evidence, including quantitative biomechanical evidence. Future work should examine the distribution of the novel contour feather morphotype identified herein. Finding such contour feathers in avialan or pygostylian stem birds would provide further support that they are likely plesiomorphic to modern contour feathers rather than a secondarily derived. When did truly modern contour feathers first appear? Mid-Cretaceous hatchling enantiornithines in amber (Xing et al. 2016b, 2017) provide highly detailed glimpses of neoptile plumage. A highly complete specimen has scarce body feathers and a combination of primitive and derived plumage morphotypes. Neoptile feathers on the body either resemble down feathers of modern birds, with elongate (plumulaceous) barbules, flexible barbs, and a poorly defined rachis [or share] the flattened (pennaceous) barb arrangement, and short rachis seen in modern neoptile feathers, but the barbs bear barbules that are pennaceous (Xing et al. 2017).
23 Also present are isolated bristle-like filaments (IBFs) on the crural tract and tail that are flattened structures erupting from separate follicles as well as scutellae scale filaments (SSFs) that erupt from the distal edge of the scutellae on the digits. However, this neoptile plumage may differ drastically from adult plumage. A relatively derived, yet old (Early Cretaceous), enantiornithine Cruralspennia multidona was described as having hair-like and rachis-less body feathers and peculiar crural feathers that are proximally wire-like with a short filamentous tip (PWFDTs), representing a novel feather morphotype (Wang et al. 2017b). PWFDTs are tapered, curved proximally, and narrow for about 90% of their length, ending in visibly separated, parallel barbs at their distal end. The dark, wire-like portion was interpreted by Wang et al. (2017b) as representing fusion of barbs into a single rachis-like structure. Might the contour feathers of Cruralspennia or PWFDTs instead be morphologically similar to the shaggy, bifurcated morphology described here in Anchiornis contours? Neoptile IBFs and SSFs might be evidence that PWFDTs are indeed a single, fused structure proximally, although the narrow, solid appearance of the proximal 90% of PWFDTs could represent a closed-vane variant of the Anchiornis contour feather morphology, where only the distal 10% of the PWFDT is open-vaned. Contour feathers associated with the basal enantiornithine Protopteryx show some semblance to the those described here for Anchiornis (Fucheng et al. 2006, see Figure 1b). Ultimately, truly modern contour feathers might be relatively more derived than originally thought. Acknowledgments. Many thanks to Hein Van Grouw for providing images and discussion of the silky domesticated birds, the curators and institutions that granted access to the fossil specimens, and for the helpful reviews of Gerald Mayr and Michael Pittman.
24 DATA ARCHIVING STATEMENT Data for this study are available in the Dryad Digital Repository: REFERENCES ALEXANDER, D. E., GONG, E., MARTIN, L. D., BURNHAM, D. A. and FALK, A. R Model tests of gliding with different hindwing configurations in the four-winged dromaeosaurid Microraptor gui. Proceedings of the National Academy of Sciences, 107, BARRETT, P. M., EVANS, D. C. and CAMPIONE, N. E Evolution of dinosaur epidermal structures. Biology Letters, 11, CHATTERJEE, S. and TEMPLIN, R. J Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui. Proceedings of the National Academy of Sciences, 104, CHEN, P. J., DONG, Z. M. and ZHEN, S. N An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. Nature, 391,
25 CHIAPPE, L. M., JI, S. A., JI, Q. and NORELL, M. A Anatomy and systematics of the Confuciusornithidae (Theropoda, Aves) from the late Mesozoic of northeastern China. Bulletin of the AMNH, 242. COLE, L. J. and WILLARD, F. H The inheritance of silky plumage in the domestic pigeon. Journal of Heredity, 30, CURRIE, P. J. and CHEN, P. J Anatomy of Sinosauropteryx prima from Liaoning, northeastern China. Canadian Journal of Earth Sciences, 38, DECECCHI, T. A., LARSSON, H. C. and HABIB, M. B The wings before the bird: an evaluation of flapping-based locomotory hypotheses in bird antecedents. PeerJ, 4, e2159. DYKE, G. J. and NORELL, M. A Caudipteryx as a non-avalian theropod rather than a flightless bird. Acta Palaeontologica Polonica, 50, DE KAT, R., PALMER, C., VAN DER KINDERE, J., NAISH, D. and GANAPATHISUBRAMANI, B Aerodynamic performance of the feathered dinosaur Microraptor and the evolution of feathered flight. Nature Communications, 4, FEDUCCIA, A The origin and evolution of birds. Yale University Press, New Haven, Connecticut.
26 FENG, C., GAO, Y., DORSHORST, B., SONG, C., GU, X., LI, Q., LI, J., LIU, T., RUBIN, C. J., ZHAO, Y. and WANG, Y A cis-regulatory mutation of PDSS2 causes silky-feather in chickens. PLoS genetics, 10, e FEO, T. J., FIELD, D. J. and PRUM, R. O Barb geometry of asymmetrical feathers reveals a transitional morphology in the evolution of avian flight. Proceedings of the Royal Society of London B: Biological Sciences, 282, FOTH, C., TISCHLINGER, H. and RAUHUT, O.W New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers. Nature, 511, FUCHENG, Z., ZHONGHE, Z. and DYKE, G Feathers and feather- like integumentary structures in Liaoning birds and dinosaurs. Geological Journal, 41, GODEFROIT, P., SINITSA, S.M., DHOUAILLY, D., BOLOTSKY, Y.L., SIZOV, A.V., MCNAMARA, M.E., BENTON, M.J. and SPAGNA, P A Jurassic ornithischian dinosaur from Siberia with both feathers and scales. Science, 345, HEERS, A. M., DIAL, K. P. and TOBALSKE, B. W From baby birds to feathered dinosaurs: incipient wings and the evolution of flight. Paleobiology, 40, HU, D., HOU, L., ZHANG, L. and XU, X A pre-archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature, 461,
27 JONES, T. D., FARLOW, J. O., RUBEN, J. A., HENDERSON, D. M., and HILLENIUS, W. J Cursoriality in bipedal archosaurs. Nature, 406, JUHN, M. and BATES, R.W Thyroid function in silky feathering. Journal of Experimental Zoology Part A: Ecological Genetics and Physiology, 143, KOEHL, M. A. R., EVANGELISTA, D. and YANG, K Using physical models to study the gliding performance of extinct animals. Integrative and Comparative Biology, 51, KOSCHOWITZ, M., FISCHER C. and SANDER P. M Beyond the rainbow. Science, 346, LEFÈVRE, U., CAU, A., CINCOTTA, A., HU, D., CHINSAMY, A., ESCUILLIÉ, F. and GODEFROIT, P A new Jurassic theropod from China documents a transitional step in the macrostructure of feathers. The Science of Nature, 104, 74. LI, Q., GAO, K.Q., VINTHER, J., SHAWKEY, M.D., CLARKE, J.A., D ALBA, L., MENG, Q., BRIGGS, D.E. and PRUM, R.O Plumage color patterns of an extinct dinosaur. Science, 327,
28 GAO, K. Q., MENG, Q., CLARKE, J. A., SHAWKEY, M. D., D ALBA, L., PEI, R., ELLISON, M., NORELL, M. A. and VINTHER, J Reconstruction of Microraptor and the evolution of iridescent plumage. Science, 335, LONGRICH, N. R., VINTHER, J., MENG, Q., LI, Q., RUSSELL, A. P Primitive wing feather arrangement in Archaeopteryx lithographica and Anchiornis huxleyi. Current Biology, 22, LÜ, J. and BRUSATTE, S.L A large, short-armed, winged dromaeosaurid (Dinosauria: Theropoda) from the Early Cretaceous of China and its implications for feather evolution. Scientific Reports, 5, LUCAS, A. M. and STETTENHEIM, P. R Avian Anatomy Integument. Part 1. U. S. Government Printing Office, Washington, D. C. MAIA, R., MACEDO, R. H. and SHAWKEY, M. D., Nanostructural self-assembly of iridescent feather barbules through depletion attraction of melanosomes during keratinization. Journal of the Royal Society Interface, 9, rsif MAYR, G Avian evolution: the fossil record of birds and its paleobiological significance. John Wiley & Sons, Hoboken, New Jersey.
29 PETERS, S. D., PLODOWSKI, G. and VOGEL, O Bristle-like integumentary structures at the tail of the horned dinosaur Psittacosaurus. Naturwissenschaften, 89, PITTMAN, M., SAITTA, E., KAYE, T. G. and VINTHER, J Structure and homology of Psittacosaurus tail bristles. Palaeontology, 59, MCKELLAR, R. C., CHATTERTON, B. D., WOLFE, A. P. and CURRIE, P.J A diverse assemblage of Late Cretaceous dinosaur and bird feathers from Canadian amber. Science, 333, MILLER, W. J Silky plumage in the ring neck dove. Journal of Heredity, 47, NORELL, M. and XU, X Feathered dinosaurs. Annual Review of Earth and Planetary Sciences, 33, NUDDS, R. L Reassessment of the wing feathers of Archaeopteryx lithographica suggests no robust evidence for the presence of elongated dorsal wing coverts. PloS one, 9, e PALMER, C The aerodynamics of gliding flight and its application to the arboreal flight of the Chinese feathered dinosaur Microraptor. Biological journal of the Linnean Society, 113,
30 PAUL, G. S Dinosaurs of the air: the evolution and loss of flight in dinosaurs and birds. Johns Hopkins University Press, Baltimore, Maryland. PAUTARD, F. G. E Mineralization of keratin and its comparison with the enamel matrix. Nature, 199, PRUM, R. O Moulting tail feathers in a juvenile oviraptorosaur. Nature, 468, E1. PRUM, R. O. and BRUSH, A. H The evolutionary origin and diversification of feathers. The Quarterly Review of Biology, 77, SAITTA, E. T., GELERNTER, R. and VINTHER, J. 2017a. Primitive contour feathers in paravian dinosaurs and interpreting feather plesiomorphies. Dryad Digital Repository. SAITTA, E. T., ROGERS, C., BROOKER, R. A., ABBOTT, G. D., KUMAR, S., O'REILLY, S. S., DONOHOE, P., DUTTA, S., SUMMONS, R. E. and VINTHER, J. 2017b. Low fossilization potential of keratin protein revealed by experimental taphonomy. Palaeontology, 60, VAN GROUW, H Silky pigeons. Aviculture Europe, 12, 10 p. ISSN: VINTHER, J A guide to the field of palaeo colour. BioEssays, 37,
31 NICHOLLS, R., LAUTENSCHLAGER, S., PITTMAN, M., KAYE, T.G., RAYFIELD, E., MAYR, G. and CUTHILL, I.C D camouflage in an ornithischian dinosaur. Current Biology, 26, WANG, M., O CONNOR, J. K., PAN, Y., and ZHOU, Z. 2017b. A bizarre Early Cretaceous enantiornithine bird with unique crural feathers and an ornithuromorph plough-shaped pygostyle. Nature Communications, 8, WANG, X., NUDDS, R. L. and DYKE, G. J The primary feather lengths of early birds with respect to avian wing shape evolution. Journal of Evolutionary Biology, 24, PITTMAN, M., ZHENG, X., KAYE, T. G., FALK, A. R., HARTMAN, S. A. and XU, X. 2017a. Basal paravian functional anatomy illuminated by high-detail body outline. Nature Communications, 8, XING, L., MCKELLAR, R.C., XU, X., LI, G., BAI, M., PERSONS, W.S., MIYASHITA, T., BENTON, M.J., ZHANG, J., WOLFE, A.P. and YI, Q. 2016a. A feathered dinosaur tail with primitive plumage trapped in mid-cretaceous amber. Current Biology, 26, MCKELLAR, R. C., WANG, M., BAI, M., O CONNOR, J. K., BENTON, M. J., ZHANG, J., WANG, Y., TSENG, K., LOCKLEY, M. G., LI, G., ZHANG, W. and XU, X. 2016b. Mummified precocial bird wings in mid-cretaceous Burmese amber. Nature Communications, 7,
32 O'CONNOR, J.K., MCKELLAR, R.C., CHIAPPE, L.M., TSENG, K., LI, G. and BAI, M A mid-cretaceous enantiornithine (Aves) hatchling preserved in Burmese amber with unusual plumage. Gondwana Research. XU, X Feathered dinosaurs from China and the evolution of major avian characters. Integrative Zoology, 1, TANG, Z. L. and WANG, X.L A therizinosauroid dinosaur with integumentary structures from China. Nature, 399, ZHOU, Z. and WANG, X The smallest known non-avian theropod dinosaur. Nature, 408, ZHONG-HE, Z. and PRUM, R.O Branched integumental structures in Sinornithosaurus and the origin of feathers. Nature, 410, ZHOU, Z., WANG, X., KUANG, X., ZHANG, F. and DU, X Four-winged dinosaurs from China. Nature, 421, ZHENG, X., SULLIVAN, C., WANG, X., XING, L., WANG, Y., ZHANG, X., O CONNOR, J. K., ZHANG, F. AND PAN, Y A bizarre Jurassic maniraptoran theropod with preserved evidence of membranous wings. Nature, 521,
33 CURRIE, P., PITTMAN, M., XING, L., MENG, Q., LÜ, J., HU, D. and YU, C Mosaic evolution in an asymmetrically feathered troodontid dinosaur with transitional features. Nature Communications, 8, ZELENITSKY, D. K., THERRIEN, F., ERICKSON, G. M., DEBUHR, C. L., KOBAYASHI, Y., EBERTH, D. A. and HADFIELD, F Feathered non-avian dinosaurs from North America provide insight into wing origins. Science, 338, ZHANG, F., ZHOU, Z., XU, X., WANG, X. and SULLIVAN, C A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers. Nature, 455, ZHENG, X., XU, X., YOU, H., ZHAO, Q. and DONG, Z. 2009b. A short-armed dromaeosaurid from the Jehol Group of China with implications for early dromaeosaurid evolution. Proceedings of the Royal Society of London B: Biological Sciences, 277, YOU, H. L., XU, X. and DONG, Z. M. 2009a. An Early Cretaceous heterodontosaurid dinosaur with filamentous integumentary structures. Nature, 458, FIGURES

34
. D, Caudipteryx (IVPP V12344). E, Sinosauropteryx (NIGP 127587). F, Sinosauropteryx (NIGP 127586). Psittacosaurus (Fig.")
35 Fig. 1. Specimens examined with boxes around locations of figure insets used in comparisons. A, Confuciusornis (IVPP V13156). B, Anchiornis (BMNHC PH828) where Fig. 6A E panels are of counter slab. C, Caudipteryx (IVPP V12430). D, Caudipteryx (IVPP V12344). E, Sinosauropteryx (NIGP ). F, Sinosauropteryx (NIGP ). Psittacosaurus (Fig. 4A B) only shows filamentous structures on the anterior portion of the tail figured therein. Scale bars represent: 10 cm (A F). Fig. 2. Isolated feathers near the torso of Anchiornis (BMNHC PH828). A, the feather indicated by the arrow shows typical bifurcating appearance. The feather indicated by the arrowhead shows unusual trident appearance. B, the feathers indicated by arrows show typical bifurcating appearance. See Fig. 3A E for details and drawings of these feathers. Scale bars represent: 1 cm (A B).

36 Fig. 3. Drawings of contour feathers arranged with basal ends downward. A B, Anchiornis (BMNHC PH828) bifurcated contour feathers as they appear in the rock. Black areas represent darkly pigmented regions of the fossil. Grey areas represent faintly preserved pigmented regions. A, the feather in Fig. 2A indicated by arrowhead. The darkly pigmented, non-striated block apical to the bifurcation of the feather is the result of overlapping adjacent feathers that intersect at this location. B, a feather in Fig. 2B indicated by rightmost arrow. C D, ccorresponding photographs of drawn feathers in A B, respectively, indicated by arrowhead. E, Anchiornis contour feather interpretation by Rebecca Gelernter. F, filaments representing the basal portion of one Sinosauropteryx tuft (NIGP ) within the region depicted by Fig. 4C D as it appears in the rock. The finer filaments extended much farther apically than illustrated here. G, corresponding
 tail bristles (see also Mayr et al. 2016, Figure 2).")
37 photograph of drawn feather in F indicated by arrowhead. Scale bars represent 1 cm (A, C), 0.5 cm (B, D), 0.25 cm (F G). Fig. 4. Non-avian dinosaur integumentary appendages. Photograph, A, and corresponding drawing, B, of Psittacosaurus (SMF R 4970) tail bristles (see also Mayr et al. 2016, Figure 2). Photograph, C, and corresponding drawing, D, of Sinosauropteryx (NIGP ) contour feathers dorsal to the caudal vertebrae. Thick strokes in C indicate high-angle, dark, thick streaks. Thin grey lines connect incomplete filaments. Photograph, E, and corresponding drawing, F, of Anchiornis (BMNHC PH828) contour feathers anterior to the hindlimb and pes with isolated contour feathers also present in the top-right corner of the panels. Thin grey lines connect incomplete filaments. Large grey areas indicate regions where bone is visible (B, C, F). Scale bars represent: 1 cm (A F).
38

39 Fig. 5. A, Feathers with potential developmental sheaths dorsal to the cervical vertebrae in Sinosauropteryx (NIGP ). Arrow indicates region containing several of these structures. Arrowheads indicate filaments splaying out as if erupting from sheaths. Vertebra is partly visible in the bottom-left corner. Scale bar represents 1 cm. B C, A single such feather and associated drawing, respectively. Thin grey lines connect incomplete filaments. Scale bar represents 25 mm.
40 Fig. 6. Anchiornis (BMNHC PH828 counter slab) and Confuciusornis (IVPP V13156) wing feathers compared to modern remiges. Drawing, A, corresponding to photograph, B, of Anchiornis primary remex. The feather crossing the panel from the bottom-left and extending to the top-right demonstrates rachis curvature and unzipped barbs. Thin grey lines connect incomplete barbs. Large grey areas indicate preserved rachis. C, close-up image of the basal region of the primary remex feather depicted in A B. D, barbs on Anchiornis major coverts and secondary remiges. E, drawing of major covert in D. Thin grey lines connect incomplete barbs. Large grey area indicate preserved rachis. F J, Confuciusornis remex barbs. Photographs, F & H, correspond to drawings, G & I, respectively. Large grey areas indicate preserved rachis. Arrows indicate jointly deflected barbs. K, Primary and, L, secondary remex of the Barbary Dove, Streptopelia risoria (i.e., domesticated African Collared Dove, photographs courtesy of Hein Van Grouw) that is a heterozygous for the silky trait and shows similarly open vanes to those seen in Anchiornis. Scale bars represent: 1 cm (A C, K L), 0.5 cm (D I), 0.25 cm (J).

41 Fig. 7. Feathers associated with Caudipteryx forelimbs. Photograph, A, and corresponding drawing, B, of the remiges of IVPP V Thin grey lines connect incomplete barbs. Large grey areas indicate preserved rachis. C E, feathers associated with the forelimb of IVPP V Photograph, C, and corresponding drawing, D, of feathers on the wing showing a frond shape.
42 Thin grey lines connect incomplete barbs. Large grey areas indicate preserved rachis. E, long, ribbon-like structures (arrowhead) near the distal end of the forelimb. Phalange visible at the top of the panel. Scale bars represent: 1 cm (A E). Fig. 8. Idealized diagrams of contour feathers in order of increasing complexity. A, simple, nonbranching filaments reminiscent of those in Psittacosaurus (although their homology to feathers is unsure). B, multiple non-branching filaments connected basally (e.g. via a hypothetical calamus or emerging from a single follicle). C, shaggy, bifurcated contour feather of Anchiornis with a short rachis and long barbs, hypothetically depicted with a calamus and lacking barbules (although undifferentiated barbules are possible). Sinosauropteryx may plausibly possess feathers ranging in morphology from A to C, although morphology B is most supported. D, modern, open-vaned feather without differentiated barbules. E, modern, closed-vaned feather with differentiated proximal and distal barbules.

43
Accepted Manuscript. News & Views. Primary feather vane asymmetry should not be used to predict the flight capabilities of feathered fossils
 Accepted Manuscript News & Views Primary feather vane asymmetry should not be used to predict the flight capabilities of feathered fossils Xia Wang, Robert L. Nudds, Colin Palmer, Gareth J. Dyke PII: S2095-9273(17)30453-X
Accepted Manuscript News & Views Primary feather vane asymmetry should not be used to predict the flight capabilities of feathered fossils Xia Wang, Robert L. Nudds, Colin Palmer, Gareth J. Dyke PII: S2095-9273(17)30453-X
SUPPLEMENTARY INFORMATION
 1. Preservation of STM4-1 and STM22-6 STM4-1 represents a complete skeleton with integumentary structures preserved on part and counterpart slabs. STM22-6 is a nearly complete skeleton with integumentary
1. Preservation of STM4-1 and STM22-6 STM4-1 represents a complete skeleton with integumentary structures preserved on part and counterpart slabs. STM22-6 is a nearly complete skeleton with integumentary
The Evolution of Birds & the Origin of Flight
 The Evolution of Birds & the Origin of Flight Archaeopteryx Solnhofen quarry Oldest known bird, but not ancestral to modern birds Inhabited coastal habitats where it probably glided between conifers, cycads,
The Evolution of Birds & the Origin of Flight Archaeopteryx Solnhofen quarry Oldest known bird, but not ancestral to modern birds Inhabited coastal habitats where it probably glided between conifers, cycads,
Origin and Evolution of Birds. Read: Chapters 1-3 in Gill but limited review of systematics
 Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
University of Bristol - Explore Bristol Research. Early version, also known as pre-print
 Godefroit, P., Sinitsa, S. M., Dhouailly, D., Bolotsky, Y. L., Sizov, A. V., McNamara, M. E.,... Spagna, P. (2014). Dinosaur evolution. A Jurassic ornithischian dinosaur from Siberia with both feathers
Godefroit, P., Sinitsa, S. M., Dhouailly, D., Bolotsky, Y. L., Sizov, A. V., McNamara, M. E.,... Spagna, P. (2014). Dinosaur evolution. A Jurassic ornithischian dinosaur from Siberia with both feathers
the Feather or the Bird?
 Which Came First, the Feather or the Bird? FEATHERS EVOLVED in carnivorous, bipedal dinosaurs before the origin of birds. The creatures depicted here are reconstructions of fossils found recently in northern
Which Came First, the Feather or the Bird? FEATHERS EVOLVED in carnivorous, bipedal dinosaurs before the origin of birds. The creatures depicted here are reconstructions of fossils found recently in northern
Origin and Evolution of Birds. Read: Chapters 1-3 in Gill but limited review of systematics
 Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
From Dinosaurs to Birds: Puzzles Unraveled while Evidence Building up
 From Dinosaurs to Birds: Puzzles Unraveled while Evidence Building up CHEN Pingfu 1 and SONG Jianlan 2 1 Institute of Vertebrate Paleontology and Paleoanthropology 2 BCAS Staff Reporter Rejuvenation of
From Dinosaurs to Birds: Puzzles Unraveled while Evidence Building up CHEN Pingfu 1 and SONG Jianlan 2 1 Institute of Vertebrate Paleontology and Paleoanthropology 2 BCAS Staff Reporter Rejuvenation of
A juvenile coelurosaurian theropod from China indicates arboreal habits
 Naturwissenschaften (2002) 89:394 398 DOI 10.1007/s00114-002-0353-8 SHORT COMMUNICATION Fucheng Zhang Zhonghe Zhou Xing Xu Xiaolin Wang A juvenile coelurosaurian theropod from China indicates arboreal
Naturwissenschaften (2002) 89:394 398 DOI 10.1007/s00114-002-0353-8 SHORT COMMUNICATION Fucheng Zhang Zhonghe Zhou Xing Xu Xiaolin Wang A juvenile coelurosaurian theropod from China indicates arboreal
The Origin of Birds. Technical name for birds is Aves, and avian means of or concerning birds.
 The Origin of Birds Technical name for birds is Aves, and avian means of or concerning birds. Birds have many unusual synapomorphies among modern animals: [ Synapomorphies (shared derived characters),
The Origin of Birds Technical name for birds is Aves, and avian means of or concerning birds. Birds have many unusual synapomorphies among modern animals: [ Synapomorphies (shared derived characters),
University of Bristol - Explore Bristol Research
 Mayr, G., Pittman, M., Saitta, E., Kaye, T., & Vinther, J. (2016). Structure and Homology of Psittacosaurus Tail Bristles. Palaeontology, 59(6). DOI: 10.1111/pala.12257 Peer reviewed version License (if
Mayr, G., Pittman, M., Saitta, E., Kaye, T., & Vinther, J. (2016). Structure and Homology of Psittacosaurus Tail Bristles. Palaeontology, 59(6). DOI: 10.1111/pala.12257 Peer reviewed version License (if
Barney to Big Bird: The Origin of Birds. Caudipteryx. The fuzzy raptor. Solnhofen Limestone, cont d
 Barney to Big Bird: The Origin of Birds Caudipteryx The fuzzy raptor The discovery of feathered dinosaurs in Liaoning, China, has excited the many paleontologists who suspected a direct link between dinosaurs
Barney to Big Bird: The Origin of Birds Caudipteryx The fuzzy raptor The discovery of feathered dinosaurs in Liaoning, China, has excited the many paleontologists who suspected a direct link between dinosaurs
A new maniraptoran dinosaur from China with long feathers on the metatarsus
 Naturwissenschaften (2005) 92:173 177 DOI 10.1007/s00114-004-0604-y SHORT COMMUNICATION Xing Xu Fucheng Zhang A new maniraptoran dinosaur from China with long feathers on the metatarsus Received: 18 March
Naturwissenschaften (2005) 92:173 177 DOI 10.1007/s00114-004-0604-y SHORT COMMUNICATION Xing Xu Fucheng Zhang A new maniraptoran dinosaur from China with long feathers on the metatarsus Received: 18 March
Video Assignments. Microraptor PBS The Four-winged Dinosaur Mark Davis SUNY Cortland Library Online
 Video Assignments Microraptor PBS The Four-winged Dinosaur Mark Davis SUNY Cortland Library Online Radiolab Apocalyptical http://www.youtube.com/watch?v=k52vd4wbdlw&feature=youtu.be Minute 13 through minute
Video Assignments Microraptor PBS The Four-winged Dinosaur Mark Davis SUNY Cortland Library Online Radiolab Apocalyptical http://www.youtube.com/watch?v=k52vd4wbdlw&feature=youtu.be Minute 13 through minute
Bird evolution. Primer
 R350 other to achieve their goal. There are also heroes and heroines and Ashburner identifies these as including Gerry Rubin and Suzi Lewis in particular. Heroes come in all shapes and sizes and, despite
R350 other to achieve their goal. There are also heroes and heroines and Ashburner identifies these as including Gerry Rubin and Suzi Lewis in particular. Heroes come in all shapes and sizes and, despite
Juehuaornis gen. nov.
 34 1 2015 3 GLOBAL GEOLOGY Vol. 34 No. 1 Mar. 2015 1004 5589 2015 01 0007 05 Juehuaornis gen. nov. 1 1 1 2 1. 110034 2. 110034 70% Juehuaornis zhangi gen. et sp. nov Q915. 4 A doi 10. 3969 /j. issn. 1004-5589.
34 1 2015 3 GLOBAL GEOLOGY Vol. 34 No. 1 Mar. 2015 1004 5589 2015 01 0007 05 Juehuaornis gen. nov. 1 1 1 2 1. 110034 2. 110034 70% Juehuaornis zhangi gen. et sp. nov Q915. 4 A doi 10. 3969 /j. issn. 1004-5589.
A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers
 A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers Fucheng Zhang, Zhonghe Zhou, Xing Xu, Xiaolin Wang, Corwin Sullivan Laboratory of Evolutionary Systematics of Vertebrates,
A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers Fucheng Zhang, Zhonghe Zhou, Xing Xu, Xiaolin Wang, Corwin Sullivan Laboratory of Evolutionary Systematics of Vertebrates,
An Archaeopteryx-like theropod dinosaur newly
 BCAS Vol.25 No.4 2011 Archaeopteryx: Dinosaur or Bird? By SONG Jianlan (Staff Reporter) An Archaeopteryx-like theropod dinosaur newly found from western Liaoning Province in northeastern China would make
BCAS Vol.25 No.4 2011 Archaeopteryx: Dinosaur or Bird? By SONG Jianlan (Staff Reporter) An Archaeopteryx-like theropod dinosaur newly found from western Liaoning Province in northeastern China would make
On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds
 and the origin of birds") On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds by Qiang Ji and Shu an Ji Chinese Geological Museum, Beijing Chinese Geology Volume 233 1996 pp.
On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds by Qiang Ji and Shu an Ji Chinese Geological Museum, Beijing Chinese Geology Volume 233 1996 pp.
6. The lifetime Darwinian fitness of one organism is greater than that of another organism if: A. it lives longer than the other B. it is able to outc
 1. The money in the kingdom of Florin consists of bills with the value written on the front, and pictures of members of the royal family on the back. To test the hypothesis that all of the Florinese $5
1. The money in the kingdom of Florin consists of bills with the value written on the front, and pictures of members of the royal family on the back. To test the hypothesis that all of the Florinese $5
The Fossil Record of Vertebrate Transitions
 The Fossil Record of Vertebrate Transitions The Fossil Evidence of Evolution 1. Fossils show a pattern of change through geologic time of new species appearing in the fossil record that are similar to
The Fossil Record of Vertebrate Transitions The Fossil Evidence of Evolution 1. Fossils show a pattern of change through geologic time of new species appearing in the fossil record that are similar to
S06-5 Selection for feather structure. 1 Introduction
 52(Supplement): 131 135, 2006 S06-5 Selection for feather structure Edward H. BURTT Jr. 1, Jann M. ICHIDA 2 1. Dept. of Zoology, Ohio Wesleyan University, Delaware, OH 43015, USA; ehburtt@owu.edu 2. Dept.
52(Supplement): 131 135, 2006 S06-5 Selection for feather structure Edward H. BURTT Jr. 1, Jann M. ICHIDA 2 1. Dept. of Zoology, Ohio Wesleyan University, Delaware, OH 43015, USA; ehburtt@owu.edu 2. Dept.
Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported
 Drawing of early ankle development of Alligator mississippiensis, as reported") Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported by a previous study 1. The intermedium is formed at
Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported by a previous study 1. The intermedium is formed at
Geo 302D: Age of Dinosaurs. LAB 7: Dinosaur diversity- Saurischians
 Geo 302D: Age of Dinosaurs LAB 7: Dinosaur diversity- Saurischians Last lab you were presented with a review of major ornithischian clades. You also were presented with some of the kinds of plants that
Geo 302D: Age of Dinosaurs LAB 7: Dinosaur diversity- Saurischians Last lab you were presented with a review of major ornithischian clades. You also were presented with some of the kinds of plants that
Adaptation to the Sky: Defining The Feather With Integument Fossils From Mesozoic China and Experimental Evidence From Molecular Laboratories
 JOURNAL OF EXPERIMENTAL ZOOLOGY (MOL DEV EVOL) 298B:42 56 (2003) Adaptation to the Sky: Defining The Feather With Integument Fossils From Mesozoic China and Experimental Evidence From Molecular Laboratories
JOURNAL OF EXPERIMENTAL ZOOLOGY (MOL DEV EVOL) 298B:42 56 (2003) Adaptation to the Sky: Defining The Feather With Integument Fossils From Mesozoic China and Experimental Evidence From Molecular Laboratories
Dilong Paradoxus. (DIE-long) Early Tyrannosaurid Found in the Middle Jurassic to the Early Cretaceous. Claudia Montilla
 Early Tyrannosaurid Found in the Middle Jurassic to the Early Cretaceous. Claudia Montilla") Dilong Paradoxus (DIE-long) Early Tyrannosaurid Found in the Middle Jurassic to the Early Cretaceous Claudia Montilla William Parker BRY Trilobites to T-Rex Dilong Paradoxus 2 Abstract Dilong paradoxus,
Dilong Paradoxus (DIE-long) Early Tyrannosaurid Found in the Middle Jurassic to the Early Cretaceous Claudia Montilla William Parker BRY Trilobites to T-Rex Dilong Paradoxus 2 Abstract Dilong paradoxus,
A Feathered Dinosaur Tail with Primitive Plumage Trapped in Mid-Cretaceous Amber
 Report A Feathered Dinosaur Tail with Primitive Plumage Trapped in Mid-Cretaceous Amber Highlights d The first non-avialan theropod fragments preserved in amber are described d d d Vertebral outlines,
Report A Feathered Dinosaur Tail with Primitive Plumage Trapped in Mid-Cretaceous Amber Highlights d The first non-avialan theropod fragments preserved in amber are described d d d Vertebral outlines,
Name: GEOL 104 Dinosaurs: A Natural History Video Assignment. DUE: Wed. Oct. 20
 GEOL 104 Dinosaurs: A Natural History Video Assignment DUE: Wed. Oct. 20 Documentaries represent one of the main media by which scientific information reaches the general public. For this assignment, you
GEOL 104 Dinosaurs: A Natural History Video Assignment DUE: Wed. Oct. 20 Documentaries represent one of the main media by which scientific information reaches the general public. For this assignment, you
Pre-Archaeopteryx coelurosaurian dinosaurs and their implications for understanding avian origins
 Invited Review Geology December 2010 Vol.55 No.35: 3971 3977 doi: 10.1007/s11434-010-4150-z SPECIAL TOPICS: Pre-Archaeopteryx coelurosaurian dinosaurs and their implications for understanding avian origins
Invited Review Geology December 2010 Vol.55 No.35: 3971 3977 doi: 10.1007/s11434-010-4150-z SPECIAL TOPICS: Pre-Archaeopteryx coelurosaurian dinosaurs and their implications for understanding avian origins
A new species of Confuciusornis from Lower Cretaceous of Jianchang Liaoning China
 29 2 2010 6 GLOBAL GEOLOGY Vol. 29 No. 2 Jun. 2010 1004-5589 2010 02-0183 - 05 1 2 2 2 1. 110004 2. 110034 Confuciusornis jianchangensis sp. nov. 蹠 V 蹠 Q915. 865 A doi 10. 3969 /j. issn. 1004-5589. 2010.
29 2 2010 6 GLOBAL GEOLOGY Vol. 29 No. 2 Jun. 2010 1004-5589 2010 02-0183 - 05 1 2 2 2 1. 110004 2. 110034 Confuciusornis jianchangensis sp. nov. 蹠 V 蹠 Q915. 865 A doi 10. 3969 /j. issn. 1004-5589. 2010.
Discovery of an Avialae bird from China, Shenzhouraptor sinensis gen. et sp. nov.
 Discovery of an Avialae bird from China, Shenzhouraptor sinensis gen. et sp. nov. by Qiang Ji 1, Shuan Ji 2, Hailu You 1, Jianping Zhang 3, Chongxi Yuan 3, Xinxin Ji 4, Jinglu Li 5, and Yinxian Li 5 1.
Discovery of an Avialae bird from China, Shenzhouraptor sinensis gen. et sp. nov. by Qiang Ji 1, Shuan Ji 2, Hailu You 1, Jianping Zhang 3, Chongxi Yuan 3, Xinxin Ji 4, Jinglu Li 5, and Yinxian Li 5 1.
Modern taxonomy. Building family trees 10/10/2011. Knowing a lot about lots of creatures. Tom Hartman. Systematics includes: 1.
 Modern taxonomy Building family trees Tom Hartman www.tuatara9.co.uk Classification has moved away from the simple grouping of organisms according to their similarities (phenetics) and has become the study
Modern taxonomy Building family trees Tom Hartman www.tuatara9.co.uk Classification has moved away from the simple grouping of organisms according to their similarities (phenetics) and has become the study
Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida. Evo-Devo Revisited. Development of the Tetrapod Limb
 Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida Evo-Devo Revisited Development of the Tetrapod Limb Limbs whether fins or arms/legs for only in particular regions or LIMB FIELDS. Primitively
Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida Evo-Devo Revisited Development of the Tetrapod Limb Limbs whether fins or arms/legs for only in particular regions or LIMB FIELDS. Primitively
Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui
 Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui Sankar Chatterjee* and R. Jack Templin *Department of Geosciences, Museum of Texas Tech University, P.O. Box 43191,
Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui Sankar Chatterjee* and R. Jack Templin *Department of Geosciences, Museum of Texas Tech University, P.O. Box 43191,
Title: Phylogenetic Methods and Vertebrate Phylogeny
 Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Do the traits of organisms provide evidence for evolution?
 PhyloStrat Tutorial Do the traits of organisms provide evidence for evolution? Consider two hypotheses about where Earth s organisms came from. The first hypothesis is from John Ray, an influential British
PhyloStrat Tutorial Do the traits of organisms provide evidence for evolution? Consider two hypotheses about where Earth s organisms came from. The first hypothesis is from John Ray, an influential British
ARE CURRENT CRITIQUES OF THE THEROPOD ORIGIN OF BIRDS SCIENCE? REBUTTAL TO FEDUCCIA (2002)
") Commentary The Auk 120(2):550 561, 2003 ARE CURRENT CRITIQUES OF THE THEROPOD ORIGIN OF BIRDS SCIENCE? REBUTTAL TO FEDUCCIA (2002) Department of Ecology and Evolutionary Biology, and Natural History Museum,
Commentary The Auk 120(2):550 561, 2003 ARE CURRENT CRITIQUES OF THE THEROPOD ORIGIN OF BIRDS SCIENCE? REBUTTAL TO FEDUCCIA (2002) Department of Ecology and Evolutionary Biology, and Natural History Museum,
LABORATORY EXERCISE 6: CLADISTICS I
 Biology 4415/5415 Evolution LABORATORY EXERCISE 6: CLADISTICS I Take a group of organisms. Let s use five: a lungfish, a frog, a crocodile, a flamingo, and a human. How to reconstruct their relationships?
Biology 4415/5415 Evolution LABORATORY EXERCISE 6: CLADISTICS I Take a group of organisms. Let s use five: a lungfish, a frog, a crocodile, a flamingo, and a human. How to reconstruct their relationships?
Shedding Light on the Dinosaur-Bird Connection
 Shedding Light on the Dinosaur-Bird Connection This text is provided courtesy of the American Museum of Natural History. When people think of dinosaurs, two types generally come to mind: the huge herbivores
Shedding Light on the Dinosaur-Bird Connection This text is provided courtesy of the American Museum of Natural History. When people think of dinosaurs, two types generally come to mind: the huge herbivores
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1
 Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
WHY ORNITHOLOGISTS SHOULD CARE ABOUT THE THEROPOD ORIGIN OF BIRDS
 The Auk A Quarterly Journal of Ornithology Vol. 119 No. 1 January 2002 The Auk 119(1):1 17, 2002 PERSPECTIVES IN ORNITHOLOGY WHY ORNITHOLOGISTS SHOULD CARE ABOUT THE THEROPOD ORIGIN OF BIRDS RICHARD O.
The Auk A Quarterly Journal of Ornithology Vol. 119 No. 1 January 2002 The Auk 119(1):1 17, 2002 PERSPECTIVES IN ORNITHOLOGY WHY ORNITHOLOGISTS SHOULD CARE ABOUT THE THEROPOD ORIGIN OF BIRDS RICHARD O.
The wing of Archaeopteryx as a primary thrust generator
 Page 1 of 5 The wing of Archaeopteryx as a primary thrust generator Nature 399, pp. 60-62 (1999) Macmillan Publishers Ltd. PHILLIP BURGERS* AND LUIS M. CHIAPPE * San Diego Natural History Museum, PO Box
Page 1 of 5 The wing of Archaeopteryx as a primary thrust generator Nature 399, pp. 60-62 (1999) Macmillan Publishers Ltd. PHILLIP BURGERS* AND LUIS M. CHIAPPE * San Diego Natural History Museum, PO Box
Theoretical Morphology and Development of Flight Feather Vane Asymmetry with Experimental Tests in Parrots
 RESEARCH ARTICLE Theoretical Morphology and Development of Flight Feather Vane Asymmetry with Experimental Tests in Parrots TERESA J. FEO 1,2 * AND RICHARD O. PRUM 1,2 1 Department of Ecology and Evolutionary
RESEARCH ARTICLE Theoretical Morphology and Development of Flight Feather Vane Asymmetry with Experimental Tests in Parrots TERESA J. FEO 1,2 * AND RICHARD O. PRUM 1,2 1 Department of Ecology and Evolutionary
Early diversification of birds: Evidence from a new opposite bird
 Early diversification of birds: Evidence from a new opposite bird ZHANG Fucheng 1, ZHOU Zhonghe 1, HOU Lianhai 1 & GU Gang 2 1. Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy
Early diversification of birds: Evidence from a new opposite bird ZHANG Fucheng 1, ZHOU Zhonghe 1, HOU Lianhai 1 & GU Gang 2 1. Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy
1 Describe the anatomy and function of the turtle shell. 2 Describe respiration in turtles. How does the shell affect respiration?
 GVZ 2017 Practice Questions Set 1 Test 3 1 Describe the anatomy and function of the turtle shell. 2 Describe respiration in turtles. How does the shell affect respiration? 3 According to the most recent
GVZ 2017 Practice Questions Set 1 Test 3 1 Describe the anatomy and function of the turtle shell. 2 Describe respiration in turtles. How does the shell affect respiration? 3 According to the most recent
Preservation of ovarian follicles reveals evolution of avian reproductive behaviour
 Preservation of ovarian follicles reveals evolution of avian reproductive behaviour Xiaoting Zheng, 1,2 Jingmai O Connor, 3 Fritz Huchzermeyer, 4 Xiaoli Wang, 1 Yan Wang, 1 Min Wang 3, Zhonghe Zhou 3 1
Preservation of ovarian follicles reveals evolution of avian reproductive behaviour Xiaoting Zheng, 1,2 Jingmai O Connor, 3 Fritz Huchzermeyer, 4 Xiaoli Wang, 1 Yan Wang, 1 Min Wang 3, Zhonghe Zhou 3 1
Fossilized remains of cat-sized flying reptile found in British Columbia
 Fossilized remains of cat-sized flying reptile found in British Columbia By Washington Post, adapted by Newsela staff on 09.06.16 Word Count 768 An artist's impression of the small-bodied, Late Cretaceous
Fossilized remains of cat-sized flying reptile found in British Columbia By Washington Post, adapted by Newsela staff on 09.06.16 Word Count 768 An artist's impression of the small-bodied, Late Cretaceous
What is evolution? Transitional fossils: evidence for evolution. In its broadest sense, evolution is simply the change in life through time.
 Transitional fossils: evidence for evolution http://domain- of- darwin.deviantart.com/art/no- Transitional- Fossils- 52231284 Western MA Atheists and Secular Humanists 28 May 2016 What is evolution? In
Transitional fossils: evidence for evolution http://domain- of- darwin.deviantart.com/art/no- Transitional- Fossils- 52231284 Western MA Atheists and Secular Humanists 28 May 2016 What is evolution? In
What is a bird? Endothermic, vertebrate. Beak with no teeth Bipedal. Lays egg with hard outer shell Strong, light weight skeleton Sister to Crocodiles
 Birds What is a bird? Endothermic, vertebrate High metabolic rate Four chambered heart Beak with no teeth Bipedal Arms modified as wings Lays egg with hard outer shell Strong, light weight skeleton Sister
Birds What is a bird? Endothermic, vertebrate High metabolic rate Four chambered heart Beak with no teeth Bipedal Arms modified as wings Lays egg with hard outer shell Strong, light weight skeleton Sister
LABORATORY EXERCISE 7: CLADISTICS I
 Biology 4415/5415 Evolution LABORATORY EXERCISE 7: CLADISTICS I Take a group of organisms. Let s use five: a lungfish, a frog, a crocodile, a flamingo, and a human. How to reconstruct their relationships?
Biology 4415/5415 Evolution LABORATORY EXERCISE 7: CLADISTICS I Take a group of organisms. Let s use five: a lungfish, a frog, a crocodile, a flamingo, and a human. How to reconstruct their relationships?
Early evolution of the biological bird: perspectives from new fossil discoveries in China
 DOI 10.1007/s10336-015-1222-5 REVIEW Early evolution of the biological bird: perspectives from new fossil discoveries in China Jingmai O Connor 1 Zhonghe Zhou 1 Received: 8 December 2014 / Revised: 1 April
DOI 10.1007/s10336-015-1222-5 REVIEW Early evolution of the biological bird: perspectives from new fossil discoveries in China Jingmai O Connor 1 Zhonghe Zhou 1 Received: 8 December 2014 / Revised: 1 April
Anatomy. Name Section. The Vertebrate Skeleton
 Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Introduction to Cladistic Analysis
 3.0 Copyright 2008 by Department of Integrative Biology, University of California-Berkeley Introduction to Cladistic Analysis tunicate lamprey Cladoselache trout lungfish frog four jaws swimbladder or
3.0 Copyright 2008 by Department of Integrative Biology, University of California-Berkeley Introduction to Cladistic Analysis tunicate lamprey Cladoselache trout lungfish frog four jaws swimbladder or
PERSPECTIVES IN ORNITHOLOGY ARCHAEOPTERYX 2007: QUO VADIS?
 PERSPECTIVES IN ORNITHOLOGY ARCHAEOPTERYX 2007: QUO VADIS? Author(s): Alan Feduccia, Larry D. Martin, Sam Tarsitano Source: The Auk, 124(2):373-380. Published By: The American Ornithologists' Union https://doi.org/10.1642/0004-8038(2007)124[373:pioaqv]2.0.co;2
PERSPECTIVES IN ORNITHOLOGY ARCHAEOPTERYX 2007: QUO VADIS? Author(s): Alan Feduccia, Larry D. Martin, Sam Tarsitano Source: The Auk, 124(2):373-380. Published By: The American Ornithologists' Union https://doi.org/10.1642/0004-8038(2007)124[373:pioaqv]2.0.co;2
Tetrapod Similarites The Origins of Birds
 Tetrapod Similarites The Origins of Birds Birds Reptiles Mammals Integument Feathers, scales Scales Hair Digestive Horny bill Teeth Teeth Skeletal Fusion of bones Some fusion Some fusion Reduction in number
Tetrapod Similarites The Origins of Birds Birds Reptiles Mammals Integument Feathers, scales Scales Hair Digestive Horny bill Teeth Teeth Skeletal Fusion of bones Some fusion Some fusion Reduction in number
A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin
 Chinese Science Bulletin 2008 SCIENCE IN CHINA PRESS ARTICLES Springer A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin XU Xing 1, ZHAO Qi 1, NORELL Mark 2, SULLIVAN
Chinese Science Bulletin 2008 SCIENCE IN CHINA PRESS ARTICLES Springer A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin XU Xing 1, ZHAO Qi 1, NORELL Mark 2, SULLIVAN
From Reptiles to Aves
 First Vertebrates From Reptiles to Aves Evolutions of Fish to Amphibians Evolution of Amphibians to Reptiles Evolution of Reptiles to Dinosaurs to Birds Common Ancestor of Birds and Reptiles: Thecodonts
First Vertebrates From Reptiles to Aves Evolutions of Fish to Amphibians Evolution of Amphibians to Reptiles Evolution of Reptiles to Dinosaurs to Birds Common Ancestor of Birds and Reptiles: Thecodonts
8/19/2013. Topic 5: The Origin of Amniotes. What are some stem Amniotes? What are some stem Amniotes? The Amniotic Egg. What is an Amniote?
 Topic 5: The Origin of Amniotes Where do amniotes fall out on the vertebrate phylogeny? What are some stem Amniotes? What is an Amniote? What changes were involved with the transition to dry habitats?
Topic 5: The Origin of Amniotes Where do amniotes fall out on the vertebrate phylogeny? What are some stem Amniotes? What is an Amniote? What changes were involved with the transition to dry habitats?
Melanosome evolution indicates a key physiological shift within feathered dinosaurs 2,000 1,500. Length (nm) 1,000
 1,000") doi:10.1038/nature12973 Melanosome evolution indicates a key physiological shift within feathered dinosaurs Quanguo Li 1, Julia A. Clarke 2, Ke-Qin Gao 3, Chang-Fu Zhou 4, Qingjin Meng 5, Daliang Li 6,
doi:10.1038/nature12973 Melanosome evolution indicates a key physiological shift within feathered dinosaurs Quanguo Li 1, Julia A. Clarke 2, Ke-Qin Gao 3, Chang-Fu Zhou 4, Qingjin Meng 5, Daliang Li 6,
COMPARING DNA SEQUENCES TO UNDERSTAND EVOLUTIONARY RELATIONSHIPS WITH BLAST
 Big Idea 1 Evolution INVESTIGATION 3 COMPARING DNA SEQUENCES TO UNDERSTAND EVOLUTIONARY RELATIONSHIPS WITH BLAST How can bioinformatics be used as a tool to determine evolutionary relationships and to
Big Idea 1 Evolution INVESTIGATION 3 COMPARING DNA SEQUENCES TO UNDERSTAND EVOLUTIONARY RELATIONSHIPS WITH BLAST How can bioinformatics be used as a tool to determine evolutionary relationships and to
Mendelian Genetics Using Drosophila melanogaster Biology 12, Investigation 1
 Mendelian Genetics Using Drosophila melanogaster Biology 12, Investigation 1 Learning the rules of inheritance is at the core of all biologists training. These rules allow geneticists to predict the patterns
Mendelian Genetics Using Drosophila melanogaster Biology 12, Investigation 1 Learning the rules of inheritance is at the core of all biologists training. These rules allow geneticists to predict the patterns
Cladistics (reading and making of cladograms)
") Cladistics (reading and making of cladograms) Definitions Systematics The branch of biological sciences concerned with classifying organisms Taxon (pl: taxa) Any unit of biological diversity (eg. Animalia,
Cladistics (reading and making of cladograms) Definitions Systematics The branch of biological sciences concerned with classifying organisms Taxon (pl: taxa) Any unit of biological diversity (eg. Animalia,
These small issues are easily addressed by small changes in wording, and should in no way delay publication of this first- rate paper.
 Reviewers' comments: Reviewer #1 (Remarks to the Author): This paper reports on a highly significant discovery and associated analysis that are likely to be of broad interest to the scientific community.
Reviewers' comments: Reviewer #1 (Remarks to the Author): This paper reports on a highly significant discovery and associated analysis that are likely to be of broad interest to the scientific community.
古脊椎动物学报 VERTEBRATA PALASIATICA. Corwin SULLIVAN
 第 52 卷第 1 期 2014 年 1 月 古脊椎动物学报 VERTEBRATA PALASIATICA pp. 3-30 figs. 1-9 Reinterpretation of the Early Cretaceous maniraptoran (Dinosauria: Theropoda) Zhongornis haoae as a scansoriopterygid-like non-avian,
第 52 卷第 1 期 2014 年 1 月 古脊椎动物学报 VERTEBRATA PALASIATICA pp. 3-30 figs. 1-9 Reinterpretation of the Early Cretaceous maniraptoran (Dinosauria: Theropoda) Zhongornis haoae as a scansoriopterygid-like non-avian,
Biology 1B Evolution Lecture 11 (March 19, 2010), Insights from the Fossil Record and Evo-Devo
, Insights from the Fossil Record and Evo-Devo") Biology 1B Evolution Lecture 11 (March 19, 2010), Insights from the Fossil Record and Evo-Devo Extinction Important points on extinction rates: Background rate of extinctions per million species per year:
Biology 1B Evolution Lecture 11 (March 19, 2010), Insights from the Fossil Record and Evo-Devo Extinction Important points on extinction rates: Background rate of extinctions per million species per year:
Evolution of Birds. Summary:
 Oregon State Standards OR Science 7.1, 7.2, 7.3, 7.3S.1, 7.3S.2 8.1, 8.2, 8.2L.1, 8.3, 8.3S.1, 8.3S.2 H.1, H.2, H.2L.4, H.2L.5, H.3, H.3S.1, H.3S.2, H.3S.3 Summary: Students create phylogenetic trees to
Oregon State Standards OR Science 7.1, 7.2, 7.3, 7.3S.1, 7.3S.2 8.1, 8.2, 8.2L.1, 8.3, 8.3S.1, 8.3S.2 H.1, H.2, H.2L.4, H.2L.5, H.3, H.3S.1, H.3S.2, H.3S.3 Summary: Students create phylogenetic trees to
Species: Panthera pardus Genus: Panthera Family: Felidae Order: Carnivora Class: Mammalia Phylum: Chordata
 CHAPTER 6: PHYLOGENY AND THE TREE OF LIFE AP Biology 3 PHYLOGENY AND SYSTEMATICS Phylogeny - evolutionary history of a species or group of related species Systematics - analytical approach to understanding
CHAPTER 6: PHYLOGENY AND THE TREE OF LIFE AP Biology 3 PHYLOGENY AND SYSTEMATICS Phylogeny - evolutionary history of a species or group of related species Systematics - analytical approach to understanding
Phylogeny Reconstruction
 Phylogeny Reconstruction Trees, Methods and Characters Reading: Gregory, 2008. Understanding Evolutionary Trees (Polly, 2006) Lab tomorrow Meet in Geology GY522 Bring computers if you have them (they will
Phylogeny Reconstruction Trees, Methods and Characters Reading: Gregory, 2008. Understanding Evolutionary Trees (Polly, 2006) Lab tomorrow Meet in Geology GY522 Bring computers if you have them (they will
Origins of avian flight a new perspective
 Origins of avian flight a new perspective Larry D. Martin Department of Ecology and Evolutionary Biology; Museum of Natural History and Biodiversity Research Center, University of Kansas, Lawrence, KS
Origins of avian flight a new perspective Larry D. Martin Department of Ecology and Evolutionary Biology; Museum of Natural History and Biodiversity Research Center, University of Kansas, Lawrence, KS
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes)
") Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
TAXONOMIC HIERARCHY. science of classification and naming of organisms
 TAXONOMIC HIERARCHY Taxonomy - science of classification and naming of organisms Taxonomic Level Kingdom Phylum subphylum Class subclass superorder Order Family Genus Species Example Animalae Chordata
TAXONOMIC HIERARCHY Taxonomy - science of classification and naming of organisms Taxonomic Level Kingdom Phylum subphylum Class subclass superorder Order Family Genus Species Example Animalae Chordata
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A.
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
muscles (enhancing biting strength). Possible states: none, one, or two.
. Possible states: none, one, or two.") Reconstructing Evolutionary Relationships S-1 Practice Exercise: Phylogeny of Terrestrial Vertebrates In this example we will construct a phylogenetic hypothesis of the relationships between seven taxa
Reconstructing Evolutionary Relationships S-1 Practice Exercise: Phylogeny of Terrestrial Vertebrates In this example we will construct a phylogenetic hypothesis of the relationships between seven taxa
Evolution as Fact. The figure below shows transitional fossils in the whale lineage.
 Evolution as Fact Evolution is a fact. Organisms descend from others with modification. Phylogeny, the lineage of ancestors and descendants, is the scientific term to Darwin's phrase "descent with modification."
Evolution as Fact Evolution is a fact. Organisms descend from others with modification. Phylogeny, the lineage of ancestors and descendants, is the scientific term to Darwin's phrase "descent with modification."
A Pterodactylus with Remains of Flight Membrane. by F. Broili (with 3 plates). Read at the Conference on 7th February 1925.
. Read at the Conference on 7th February 1925.") Broili, F. (1925) Ein Pterodactylus mit Resten der Flughaut. Sitzungsberichte der Bayerischen Königlichen Akademie der Wissenschaften, Mathematischen-Physicalischen Classe, 1925, 23-32. A Pterodactylus
Broili, F. (1925) Ein Pterodactylus mit Resten der Flughaut. Sitzungsberichte der Bayerischen Königlichen Akademie der Wissenschaften, Mathematischen-Physicalischen Classe, 1925, 23-32. A Pterodactylus
Crocs and Birds as Dino models Crocs and birds united with dinos by morphology Both also have parental care and vocal communication between offspring
 Chapter 16. Mesozoic Diapsids Phylogenetic relationships Earliest from late carboniferous stem diapsids Petrolacosaurus Lineage split into two: Archosauromorpha Crocs, birds, dinos, pterosaurs Lepidosauromorpha
Chapter 16. Mesozoic Diapsids Phylogenetic relationships Earliest from late carboniferous stem diapsids Petrolacosaurus Lineage split into two: Archosauromorpha Crocs, birds, dinos, pterosaurs Lepidosauromorpha
FORENSIC ORNITHOLOGY
 FORENSIC ORNITHOLOGY Carla J. Dove Division of Birds Smithsonian Institution Washington, DC 20560 INTRODUCTION Feathers are among the most uniquely designed, beautiful structures in nature and have evolved
FORENSIC ORNITHOLOGY Carla J. Dove Division of Birds Smithsonian Institution Washington, DC 20560 INTRODUCTION Feathers are among the most uniquely designed, beautiful structures in nature and have evolved
Deposited on: 16 November 2012
 Wang, X., McGowan, A.J., and Dyke, G.J. (2011) Avian wing proportions and flight styles: first step towards predicting the flight modes of Mesozoic birds. PLoS ONE, 6 (12). e28672. ISSN 1932-6203 http://eprints.gla.ac.uk/71705
Wang, X., McGowan, A.J., and Dyke, G.J. (2011) Avian wing proportions and flight styles: first step towards predicting the flight modes of Mesozoic birds. PLoS ONE, 6 (12). e28672. ISSN 1932-6203 http://eprints.gla.ac.uk/71705
Ceri Pennington VELOCIRAPTOR
 Ceri Pennington VELOCIRAPTOR The Velociraptor - meaning swift seizer - lived during the late Cretaceous period - 75-71 million years ago. They were a genus of dromaeosaurid theropod dinosaur and there
Ceri Pennington VELOCIRAPTOR The Velociraptor - meaning swift seizer - lived during the late Cretaceous period - 75-71 million years ago. They were a genus of dromaeosaurid theropod dinosaur and there
Ahypertrophied ossified sternum characterizes all living birds,
 On the absence of sternal elements in Anchiornis (Paraves) and Sapeornis (Aves) and the complex early evolution of the avian sternum Xiaoting Zheng a,b, Jingmai O Connor c,1, Xiaoli Wang a, Min Wang c,
On the absence of sternal elements in Anchiornis (Paraves) and Sapeornis (Aves) and the complex early evolution of the avian sternum Xiaoting Zheng a,b, Jingmai O Connor c,1, Xiaoli Wang a, Min Wang c,
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
286 œvo. 72 THE MOLT OF HUMMINGBIRDS
 [ Auk 286 œvo. 72 THE MOLT OF HUMMINGBIRDS BY HELMUTH O. WAGNER FEw details are available about the molts of hummingbirds. When collecting in Mexico, I was struck by characteristic variations in the sequence
[ Auk 286 œvo. 72 THE MOLT OF HUMMINGBIRDS BY HELMUTH O. WAGNER FEw details are available about the molts of hummingbirds. When collecting in Mexico, I was struck by characteristic variations in the sequence
Diapsida. BIO2135 Animal Form and Function. Page 1. Diapsida (Reptilia, Sauropsida) Amniote eggs. Amniote egg. Temporal fenestra.
 Amniote eggs. Amniote egg. Temporal fenestra.") Diapsida (Reptilia, Sauropsida) Vertebrate phylogeny Mixini Chondrichthyes Sarcopterygii Mammalia Pteromyzontida Actinopterygii Amphibia Reptilia! 1! Amniota (autapomorphies) Costal ventilation Amniote
Diapsida (Reptilia, Sauropsida) Vertebrate phylogeny Mixini Chondrichthyes Sarcopterygii Mammalia Pteromyzontida Actinopterygii Amphibia Reptilia! 1! Amniota (autapomorphies) Costal ventilation Amniote
Diapsida. BIO2135 Animal Form and Function. Page 1. Diapsida (Reptilia, Sauropsida) Amniote egg. Membranes. Vertebrate phylogeny
 Amniote egg. Membranes. Vertebrate phylogeny") Diapsida (Reptilia, Sauropsida) 1 Vertebrate phylogeny Mixini Chondrichthyes Sarcopterygii Mammalia Pteromyzontida Actinopterygii Amphibia Reptilia!! Amniota (autapomorphies) Costal ventilation Amniote
Diapsida (Reptilia, Sauropsida) 1 Vertebrate phylogeny Mixini Chondrichthyes Sarcopterygii Mammalia Pteromyzontida Actinopterygii Amphibia Reptilia!! Amniota (autapomorphies) Costal ventilation Amniote
First Flightless Pterosaur
 First Flightless Pterosaur David Peters no affiliation 9 Greenfield Court, Saint Charles, MO 63303 USA Pterosaur fossils have been discovered all over the world [1], but so far no flightless pterosaurs
First Flightless Pterosaur David Peters no affiliation 9 Greenfield Court, Saint Charles, MO 63303 USA Pterosaur fossils have been discovered all over the world [1], but so far no flightless pterosaurs
CLADISTICS Student Packet SUMMARY Phylogeny Phylogenetic trees/cladograms
 CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
Interpreting Evolutionary Trees Honors Integrated Science 4 Name Per.
 Interpreting Evolutionary Trees Honors Integrated Science 4 Name Per. Introduction Imagine a single diagram representing the evolutionary relationships between everything that has ever lived. If life evolved
Interpreting Evolutionary Trees Honors Integrated Science 4 Name Per. Introduction Imagine a single diagram representing the evolutionary relationships between everything that has ever lived. If life evolved
2. Using an appropriate illustration and words, describe the physics of flight.
 1. Besides the obvious, like feathers and wings, birds have many special features that allow them to fly. Explain how each of the characteristics are specialized to help birds fly. A. Skeletal System-
1. Besides the obvious, like feathers and wings, birds have many special features that allow them to fly. Explain how each of the characteristics are specialized to help birds fly. A. Skeletal System-
Vol. XIV, No. 1, March, The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S.
 By S.") Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Are the dinosauromorph femora from the Upper Triassic of Hayden Quarry (New Mexico) three stages in a growth series of a single taxon?
 three stages in a growth series of a single taxon?") Anais da Academia Brasileira de Ciências (2017) 89(2): 835-839 (Annals of the Brazilian Academy of Sciences) Printed version ISSN 0001-3765 / Online version ISSN 1678-2690 http://dx.doi.org/10.1590/0001-3765201720160583
Anais da Academia Brasileira de Ciências (2017) 89(2): 835-839 (Annals of the Brazilian Academy of Sciences) Printed version ISSN 0001-3765 / Online version ISSN 1678-2690 http://dx.doi.org/10.1590/0001-3765201720160583
The evolution of feathers: a major problem for Darwinism
 The evolution of feathers: a major problem for Darwinism Jerry Bergman The origin of birds has always been a major problem for Darwinism, and even today little agreement about the evolution of birds exists.
The evolution of feathers: a major problem for Darwinism Jerry Bergman The origin of birds has always been a major problem for Darwinism, and even today little agreement about the evolution of birds exists.
UNIT III A. Descent with Modification(Ch19) B. Phylogeny (Ch20) C. Evolution of Populations (Ch21) D. Origin of Species or Speciation (Ch22)
 B. Phylogeny (Ch20) C. Evolution of Populations (Ch21) D. Origin of Species or Speciation (Ch22)") UNIT III A. Descent with Modification(Ch9) B. Phylogeny (Ch2) C. Evolution of Populations (Ch2) D. Origin of Species or Speciation (Ch22) Classification in broad term simply means putting things in classes
UNIT III A. Descent with Modification(Ch9) B. Phylogeny (Ch2) C. Evolution of Populations (Ch2) D. Origin of Species or Speciation (Ch22) Classification in broad term simply means putting things in classes
May 10, SWBAT analyze and evaluate the scientific evidence provided by the fossil record.
 May 10, 2017 Aims: SWBAT analyze and evaluate the scientific evidence provided by the fossil record. Agenda 1. Do Now 2. Class Notes 3. Guided Practice 4. Independent Practice 5. Practicing our AIMS: E.3-Examining
May 10, 2017 Aims: SWBAT analyze and evaluate the scientific evidence provided by the fossil record. Agenda 1. Do Now 2. Class Notes 3. Guided Practice 4. Independent Practice 5. Practicing our AIMS: E.3-Examining
A new species of sauropod, Mamenchisaurus anyuensis sp. nov.
 A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
Bio 1B Lecture Outline (please print and bring along) Fall, 2006
 Fall, 2006") Bio 1B Lecture Outline (please print and bring along) Fall, 2006 B.D. Mishler, Dept. of Integrative Biology 2-6810, bmishler@berkeley.edu Evolution lecture #4 -- Phylogenetic Analysis (Cladistics) -- Oct.
Bio 1B Lecture Outline (please print and bring along) Fall, 2006 B.D. Mishler, Dept. of Integrative Biology 2-6810, bmishler@berkeley.edu Evolution lecture #4 -- Phylogenetic Analysis (Cladistics) -- Oct.
 Oct. 2017 ACTA GEOLOGICA SINICA (English Edition) Vol. 91 No. 5 1529 http://www.geojournals.cn/dzxben/ch/index.aspx of Yumenerpeton and that of all the other bystrowianids. On the other hand, the primitive
Oct. 2017 ACTA GEOLOGICA SINICA (English Edition) Vol. 91 No. 5 1529 http://www.geojournals.cn/dzxben/ch/index.aspx of Yumenerpeton and that of all the other bystrowianids. On the other hand, the primitive
Red Eared Slider Secrets. Although Most Red-Eared Sliders Can Live Up to Years, Most WILL NOT Survive Two Years!
 Although Most Red-Eared Sliders Can Live Up to 45-60 Years, Most WILL NOT Survive Two Years! Chris Johnson 2014 2 Red Eared Slider Secrets Although Most Red-Eared Sliders Can Live Up to 45-60 Years, Most
Although Most Red-Eared Sliders Can Live Up to 45-60 Years, Most WILL NOT Survive Two Years! Chris Johnson 2014 2 Red Eared Slider Secrets Although Most Red-Eared Sliders Can Live Up to 45-60 Years, Most
Evolution on Exhibit Hints for Teachers
 1 Evolution on Exhibit Hints for Teachers This gallery activity explores a variety of evolution themes that are well illustrated by gallery specimens and exhibits. Each activity is aligned with the NGSS
1 Evolution on Exhibit Hints for Teachers This gallery activity explores a variety of evolution themes that are well illustrated by gallery specimens and exhibits. Each activity is aligned with the NGSS
What are taxonomy, classification, and systematics?
 Topic 2: Comparative Method o Taxonomy, classification, systematics o Importance of phylogenies o A closer look at systematics o Some key concepts o Parts of a cladogram o Groups and characters o Homology
Topic 2: Comparative Method o Taxonomy, classification, systematics o Importance of phylogenies o A closer look at systematics o Some key concepts o Parts of a cladogram o Groups and characters o Homology
Supplementary Note 1. Additional osteological description
 Supplementary Note 1 Additional osteological description The text below provides additional details of Jianianhualong that were not pertinent to the salient osteological description provided in the main
Supplementary Note 1 Additional osteological description The text below provides additional details of Jianianhualong that were not pertinent to the salient osteological description provided in the main