Light and electron microscopic study of the pathology and merogony of Goussia gadi (Apicomplexa: Coccidia) in the swimbladder wall
|
|
|
- Aubrie Osborne
- 5 years ago
- Views:
Transcription
 in the swimbladder wall of haddock Melanogrammus aeglefinus C. M. Morrisonl, J.")


1 Vol. 17: DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org., Published November 18 I Light and electron microscopic study of the pathology and merogony of Goussia gadi (Apicomplexa: Coccidia) in the swimbladder wall of haddock Melanogrammus aeglefinus C. M. Morrisonl, J. P. Leger2, C. A. Morrison' 'Department of Fisheries and Oceans, Biological Sciences Branch, Halifax, Nova Scotia, Canada B3J 2S7 'Anatomy Dept, Dalhousie University, Halifax. Nova Scotia, Canada B4H 4H7 ABSTRACT. Asexual and sexual stages in the life cycle of Goussia (Plag~llaJ gadi (Fiebiger, 1913), pardsites of haddock Melanograrnmus aeglefinus, occur in loose connective hssue between the lobes of the gas gland and in the s\vimbladrier wall. The asexual stages are usually found in the deeper layers, and the sexual stages towards the lumen of the swimbladder. There are many leukocytes in the lumens and walls of the blood vessels of the loose connective tissue, and phagocytes containing degeoerating parasites and trophozoites are seen in the surrounding connective tissue. Various stages of the parasite and disintegrating host cells arc released into the lumen of the swimbladder, forming a yellowish exudate. Mt*rozoltos form by sc\.eral generations ol endogenesis. The first generation, in which only 2 daughter meronts are formed (endodyogeny), may occur in the sporozoite after it has been released trom the sporocyst, when it is still within the oocyst membrane; in a free sporozoite, in a trophozoite which may be free ot in a phagocyte. This may be repeated, so that a group of elongate, intertwined merozoiles is formed. A second generation of rosettes of radiating meronts is formed by endopolygeny. usually in vacuoles of host cells which are reduced to a thin rim of vacuolated cytoplasm. KEY WORDS: Coccid~a Goussia gadi Haddock. Melanogrammus aeglefinus. Merogony. Swimbladder INTRODUCTION Goussia (Plagula) gadi (Fiebiger, 1913) is a coccidian parasite in gadoid fish from the North Atlantic and the Baltic Sea. It was first described in pollock Pollachius virens (Linnaeus, 1758) by M. Auerbach and in cod Gadus morhua Linnaeus, l758 by J. Mueller (Fiebiger 1913), who described yellow matter consisting mainly of spores having a capsule consisting of 2 halves. G. gad] is also frequently found in the wall of the swimbladder of haddock Melanogran~mus aeglefinus (Linnaeus, 1758) (Dykova & Lom 1983, Upton et al. 1984). It has been recorded in haddock, cod and a single fourbeard rockling Enchelyopus cimbrius (Linnaeus, 1766) in Canadian waters (Odense & Logan 1976, Morrison et al. 1986). Its prevalence vanes seasonally and geographically, but has reached 84.1 % in haddock on Browns Bank (NW Atlantic) (Scott 1981). In the swimbladder Goussia gadi encounters high oxygen concentrations and a well-vascularised substrate high in nutnents, which probably enables it to multiply rapidly (Fiebiger 1913). The swimbladder of gadoids is physoclistous, and the spores accumulate as the disease progresses, filling most of the swimbladder with yellow matter. Mueller described such infected fish as emaciated, although Fiebiger found that this was not always the case (Fiebiger 1913). It has been suggested that the infection would interfere with the regulation of buoyancy, so that feeding and reproduction would be difficult (Odense & Logan 1976). Only the anterior end of lightly infected swimbladders was filled with yellow matter, and a layer extended posteriorly along th.e ventral wall, becoming progressively thinner (Fiebiger 1913). Besides spores, droplets of lipid material, cholesterol platelets and fatty acid 'margarine needles' were found. O Inter-Research 199?
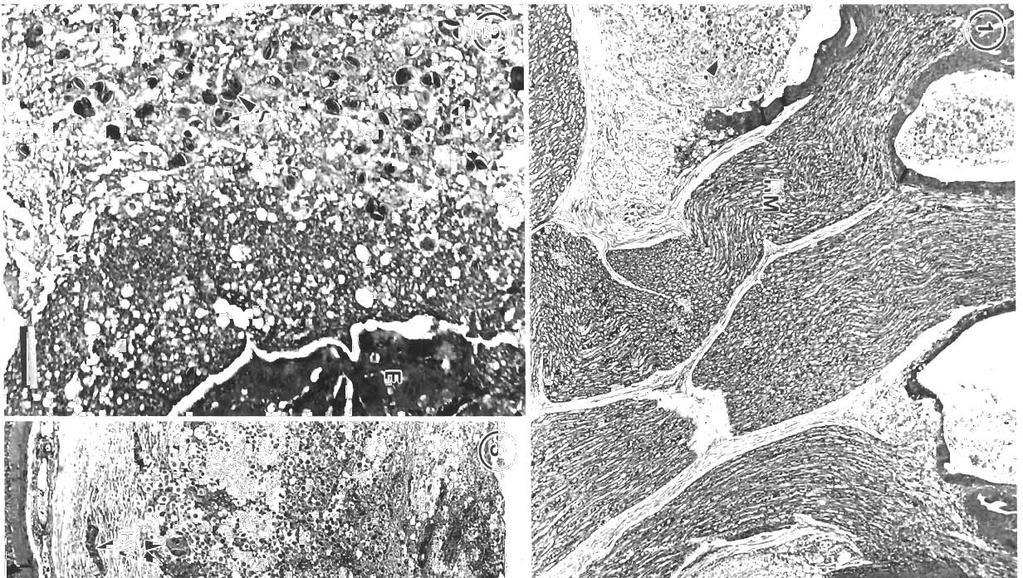
2 114 DIS. aquat. Org Other coccidians found In swimbladders are Gocissia caseosa, in the grenadier Macrourus berglax Lacepede (Lom & Dykova 19821, and Goussia cichlidarum, found in various cichlid fish, especially Sarotherodon gahlaeus (Landsberg & Paperna 1985). The only developmental stages of G. caseosa found were sporulating oocysts. These were observed mainly in the gas gland of the swimbladder, but also in the mesenteric blood vessels, gall bladder and intestinal contents. The oocysts, sporocysts and sporozoites are larger than those of G. gadi, no open sporocysts were found within the stvimbladder, and no developmental stages were found in the swimbladder wall. G. cichlidarum differs from G. gadi and G. caseosa in size, and also in that it develops within host cells which are displaced to the surface of the epithelia1 lining of the swimbladder (Paperna et al. 1986). The light microscopic appearance of all stages of Goussia gadi has been described in detail and drawn (Fiebiger 1923), but the only stage studied using electron microscopy was the thick-walled sporocyst (Odense & Logan 1976). By experimentally infecting cultured fish, Steinhagen (1991) found that the asexual stages of Goussia carpel11 were only present for a short time, and thus would be difficult to find in wild, marine fish. However, the complete life cycle of G. gadi takes place in the wall of the swimbladder. The asexual stages are in the loose connective tissue of the wall, and according to Shulman & Shtein (1962) their development is not intracellular, but takes place in interstitial spaces. Maturation of the spores occurs towards the internal surface of the swimbladder, and sporozoites are released to infect new parts of the wall (Fiebiger 1913, Shulman & Shtein 1962). This situation provides a good opportunity to study all the developmental stages, although without the advantage of a timed series as could be obtained with experimentally infected fish. Here we present light micrographs of the developmental stages of G. gadi, previously shown only as drawings (Fiebiger 1913), and electron micrographs of these same stages. A variety of terms have been used in the literature for different asexual stages, but we refer to any process that resu.lts in the formation of merozoites as merogony, as suggested by Chobotar & Scholtyseck (1982). MATERIALS AND METHODS Haddock werc examined for infection by opening the body cdvity a.nd slitting open the ventral surface of the swimbladder. Goussia gadi infections were diagnosed by the presence of a yellowish waxy exudate in the swimbladder Eight infected swimbladders were sampled on cruises in July 1990 on Emerald, La Have, Georges and Baccaro Banks, (NW Atlantic) and seven in September 1990 on Emerald Bank. Nine fish were female, six were male; they varied in length from 44 to 61 cm. Infected swimbladders from the July cruise were fixed whole in 1 % glutaraldehyde and 4 %B formaldehyde in phosphate buffer (1G4F; McDowell 1978). On return to the laboratory a longitudinal strip was removed from the anter~or to the posterior part of the ventral swimbladder wall, and cut transversely into several pieces wh~ch were processed for paraffin and JB4 resin embedding for light microscopy. Paraffin sections were stained with Harris haematoxylin and eosin (Humason 1979), whereas JB4 resin sections were stained with methylene blue/basic fuchsin (Dougherty 1981), or a 1:50 dilution of 1 % toluidine blue in 1 %, sodium borate. Samples were also taken for transmission electron microscopy (TEM). The outer tough fibrous layer was removed, then the remaining tissues were cut into cubes not larger than lmm, post-fixed in 1.5 % osmium tetroxide, dehydrated in acetone and embedded in TAAB resin. Both JB4 and TAAB resins were obtained from Marivac, Inc., Halifax, NS, Canada. Sections (0.5 pm) were stained in toluidine blue, and ultrathin gold and silver sections were stained in 25 '4 uranyl acetate in methanol (Stempack & Ward 1964) and lead citrate (Reynolds 1963). Fixation of whole swimbladders was not always adequate for TEM, so small samples of swimbladder were removed from infected haddock at sea during the September cruise, and immediately cut into small cubes in 1G4F fixative. These were then processed for TEM in the same manner as the J'uly samples. RESULTS Haddock swimbladders contained variable amounts of yellow material. Only one specimen, from Georges Bank, with small amounts of exudate with a wax-like consistency, proved to be uninfected. Coccidia were found in all other specimens, which contained significant accumulations of exudate, rna~nly around the gas gland. In histological sections there are few coccidia in the epithelium or rete mirabile of the gas gland, but disintegrating host tissue and asexual and sexual stages of th.e parasite are found in the loose connective tissue between the lobes of the gland, and in the swimbladder lumen (Pigs. 1 & 2). Many parasites are present in the loose connective tissue below the tunica interna of the scvimbladder posterior to the gas gland, but not in the outer densc, fibrous layer (Fig. 3). Most meronts and gamonts are found deep in the loose connective tissue, towards the outer f~brous wall of the swim.bladder, in, large vacuoles giving a 'honey-omb'
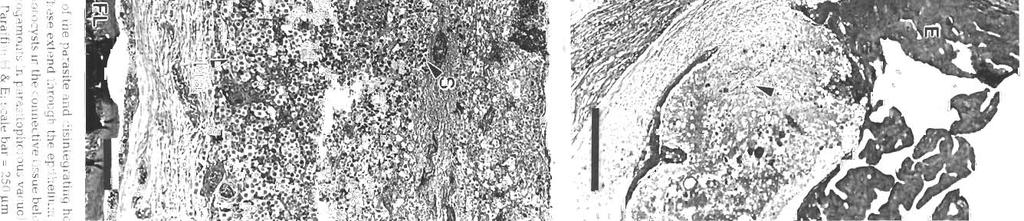
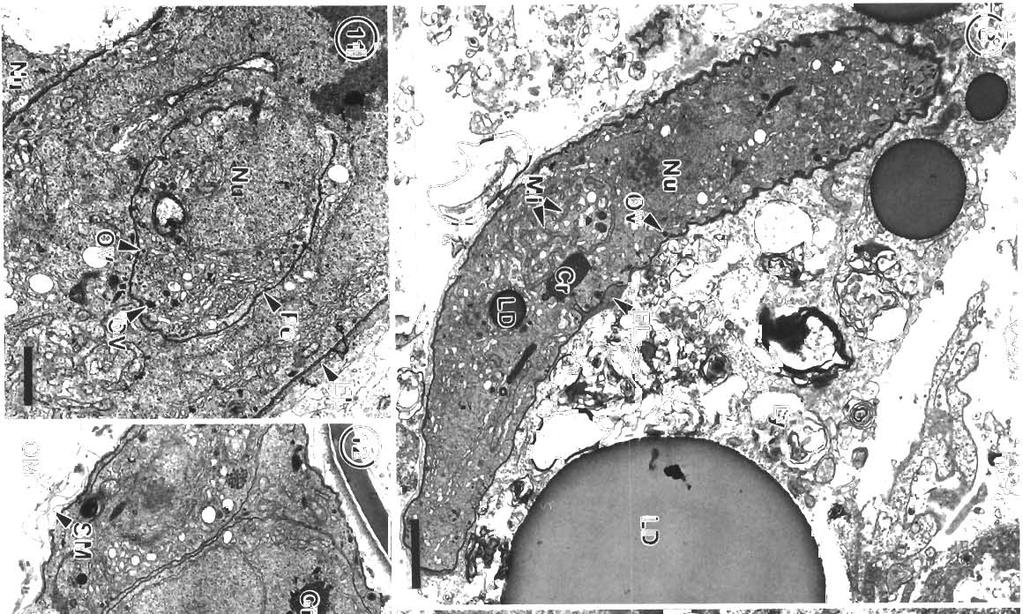
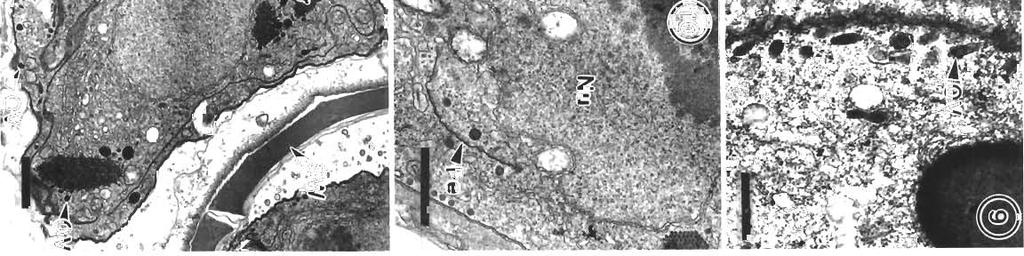
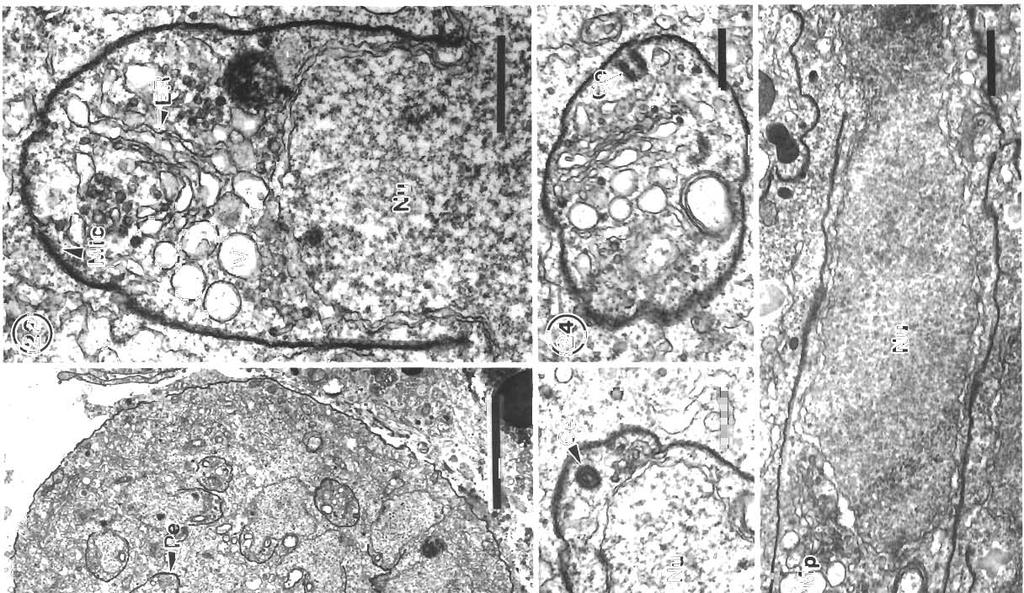
3 Morr~son et al.: Pdthology and merogony of Goussia gad; 115 appearance to the tissue. Most sporocysts and degenerating host cells are found towards the lumen of the swimbladder There is mixing of the stages, since the sporocysts release sporozoites which reinfect the swimbladder. Blood vessel walls in the loose connective tissue have a foamy appearance, cciused by many leukocytes in the endothelium and between the fibroblasts surrounding each blood vessel, and edema between the fibroblasts (Figs. 3, 4 & 5). There are phagocytes in the connective tissue around the blood vessels, some of which contain disintegrating parasites (Fig. 4). Amoeboid leukocytes containing electron-dense granules of varying sizes are seen in the endothelium and the connective tissue surrounding the blood vessels (Figs. 5 & 6). There are also groups of granulocytes containing many large granules up to about 580 nm in diameter, which might surround a parasite (Fig.?), although ingested parasites were not seen. The loose connective tissue layer of the swimbladder wall of haddock, like that of cod (Morrison 1987), contains a loose network of fibrils derived from fibroblasts (Morrison 1993), but few cells. Sporozoite-shaped trophozoites with only remnants of or no apical complex occur in phagocytes (Fig. 8). The latter are probably macrophages, because they contain many lysosomes as well as large lipid droplets. Many phagocytes have highly vacuolated cytoplasm containing myelin figures. The trophozoites are surrounded by a membrane, and in some specimens there is a thin layer of electron-dense material containing small vesicles between the membrane and the trophozoite. In some the layer is electron-transparent, and may form a large parasitophorous vacuole. The trophozoites contain a nucl.eus and a group of mitochondria, as well as lipid droplets and often a crystalloid body. Beneath the pellicle are small electron-dense vesicles, especially at each end of the parasite, which may be in contact with the pellicle (Fig. 9). Similar trophozoites are found free between the host cells. In some trophozoites there is a double-membraned pellicle around a lobe extending from the nucleus (Fig. 10). This is an early stage in the formation of a first generation merozoite. In other trophozoites the merozoite is more elongate, and organelles are present within the pellicle (Fig. 11). Two first generation merozoites are present in some trophozoites (Figs. 12 & 13). These contain all the nuclear material from the trophozoite, whose cyto- plasm apparently disappears by the end of this division, known as endodyogeny. Endodyogeny can occur in trophozoites which are still within the oocyst membrane (Fig 12), although usually this process occurs in free trophozoites, or in trophozoites within host cells. Elongate merozoites curving around each other are found (Fig. 13). Two merozoites may be seen next to each other in the host cytoplasm, but division can take place more than once (Fig. 14). Groups of elongate convoluted merozoites attached to a residual body are seen, some within host cells, others free (Fig. 15). The merozoites contain peripheral osmiophilic vesicles and crystalloids. They are surrounded by the trilayered pellicle typical of coccidia (Chobotar & Scholtyseck 1982), but the residual body is surrounded only by a unit membrane (Fig. 16). Rounded uninucleate merozoites are found in vacuoles in the host cell cytoplasm, which is highly vacuolated, and often reduced to a thin layer (Fig. 17). These become either multinucleate meronts or microgamonts, or macrogamonts (Fig. 18) Early multinucleate lneronts and microgamonts cannot be distinguished from each other (Fig. 19). Th.eir cytoplasm contains mitochondria, endoplasmic reticulum and peripheral amylopectln granules (Fig 20). Some multinucleate meronts have the early stage of the second generation merozoite, a pellicle encircling a lobe of each nucleus (Fig. 21). Subpellicular microtubules are present, and there are cisternae of endoplasmic reticulum and vesicles above the nucleus (Fig. 22). There may be a micropore (Fig. 23), and a centriole in the apical cytoplasm (Figs. 23 & 24). Some meronts contain elongate second generation merozoites (Fig. 25); others are filled with radiating merozoites continuous with the resldual body of the meront, which contains lipid droplets (Fig. 26). Rosettes of merozoites attached to the residual body of the mother meront are found, in which no cytoplasm remains between the merozoites (Figs. 27 & 28). The average size of 36 mature merozoites was 3.0 lm wide (range 1.9 to 5.0 pm) and 16.1 pm long (range 11.3 to 21.3 pm). The elongate nucleus is 2 to 3 pm in diameter, and contains a prominent nucleolus. A small apical complex is present. This consists of a conoid and a few organelles which appear to be secretory. The latter are club-shaped and electron-dense, and open via a duct at the pellicle (Figs. 29 & 30). There are also peripheral amylopectin granules and mitochondria, and a Golgi Abbrev~ations used in figures. AG = amylopectin granule; BV = blood vessel; C = conoid; Ce = centriole; CO = collagen; Cr = crystallold body; CT = connective tissue; E = epithelium; En = endothelial cell; Er = erythrocyte; EL = electron-dense layer; ER = endoplasmic reticulum; F = fibroblast; FL = fibrous layer; G = granule; GB = Golgi body; HC = host cell cytoplasm; L = leukocyte; Ly = lysosome; LD = lipid droplet; Ma = macrogamont; Me = merozoites; Mem = membrane; Mer = rneront; Mi = mitochondrion; Mic = microtubule; Mp = micropore; N = neutrophil; Nu = nucleus; OM = oocyst membrane; OV = osmiophilic vesicle; P = parasite; Pe = pellicle; Ph = phagocyte; PV = parasitophorous vacuole; RB = residual body; RM = rete mirabile; S = sporocyst; SM = sporoblast or sporocyst membrane; SO = secretory organelle; SW = sporocyst wall; V = vesicle


4
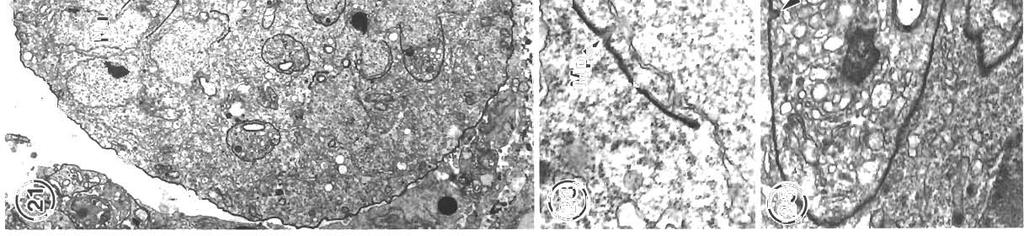
![Mornson et a1 Pathology and merogony of Gouss~agad] 117 Figs. 4 to?](/docs-images/87/95587414/images/5-0.jpg ". Goussia gadi parasitizing Melanogrammus aeglefinus. Fig. 4.")

 contalning granules of varying kocyte (neutrophil?) in the endothelium.")
5 Mornson et a1 Pathology and merogony of Gouss~agad] 117 Figs. 4 to?. Goussia gadi parasitizing Melanogrammus aeglefinus. Fig. 4. Leukocytes and erythrocytes in a blood vessel, and leukocytes between the fibroblasts external to the endothelium. One phagocyte contains a large lipld droplet and a disintegrating parasite. Fibroblasts are separated by spaces, some of which contain collagen. TEM; scale bar = 2 pm Fig. 5. Amoeboid leuleukocyte (neutrophil?) contalning granules of varying kocyte (neutrophil?) in the endothelium. TEM; scale bar = 2 pm sizes. TEM; scale bar = 1 pm. Fig. 7. Leukocyte (eosinophil?) containing large granules, surround~nga sporozoite. TEM, scale bar = l pm
6


. In some merozoites the conoid is protruded (Fig.")
.")
 state that Goussia gad1 develops in the gas gland but, as shown in this study, the gland itself is not")
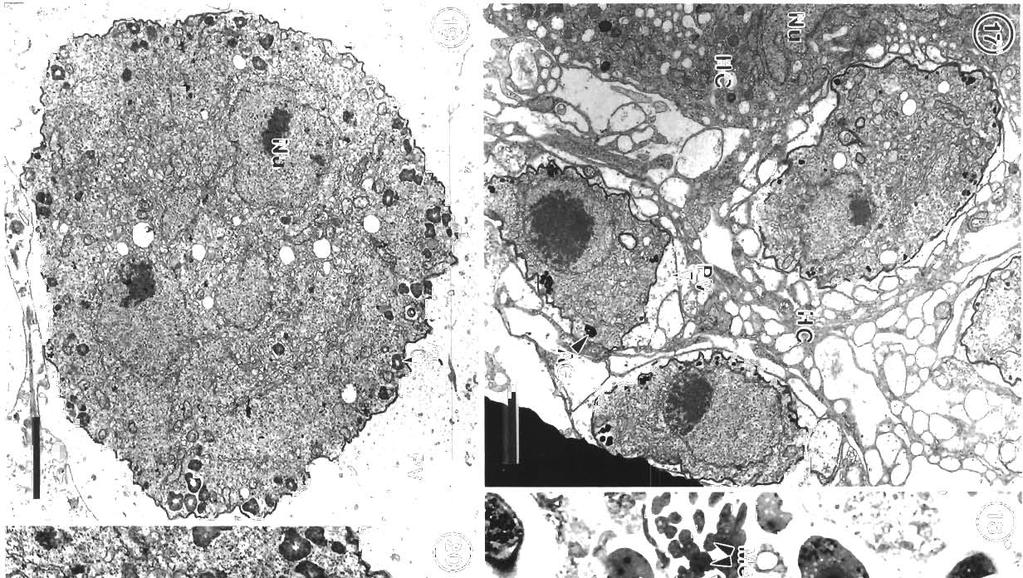
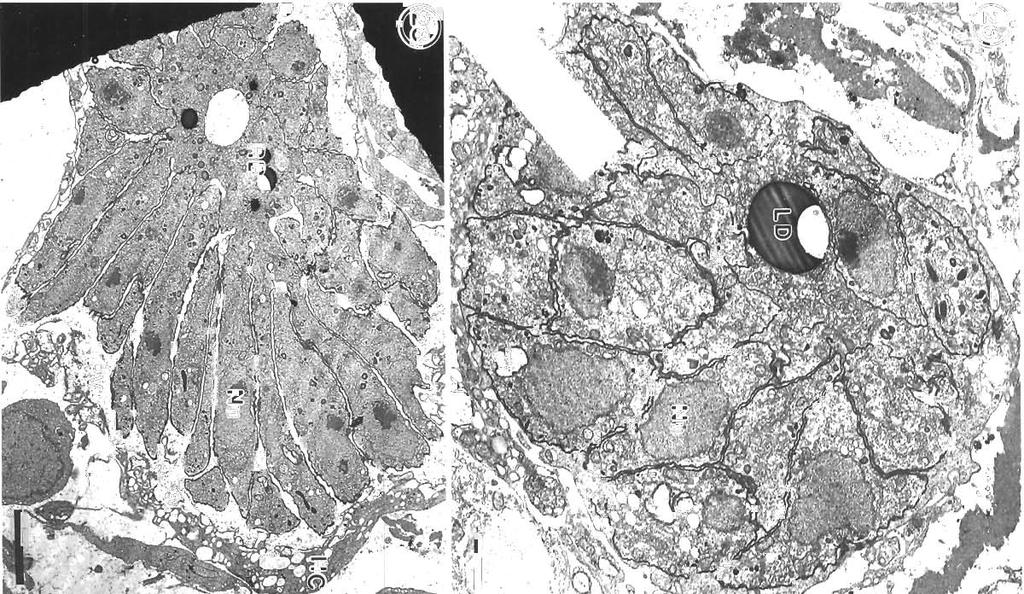
7 Morrison et al.: Pathology and merogony of Gouss~a gad1 119 Figs. 13 to 16. Goussia gadi parasitizing Melanogramrnus aeglefinus. Fig. 13. TWO convoluted merozoites in a free trophozoite. TEM; scale bar = 2 pm. Fig. 14. Two adjoining meronts each contain 2 merozoites. An electron-dense layer separates the meronts from the host cell cytoplasm containing lysosomes. TEM; scale bar = 1 pm. Fig. 15. Group of merozoites with central residual body. TEM, scale bar = 2 pm Fig. 16. Same group of merozoites at a higher magnification. Lipid droplets and amylopectin granules are present in the residual body, and merozo~tes containing crystalloid bodies and osmiophilic vesicles are surrounded by a pellicle. TEM; scale bar = l pm apparatus anterior to the nucleus (Fig. 29). In some merozoites the conoid is protruded (Fig. 30), and small crystalloid formations occur in the cytoplasm (Fig. 31). Merozoites are surrounded by a pellicle, which is not present around the residual body (Fig. 32). Some large meronts appear to be dividing in two (Fig. 33), and there are vacuoles containing 2 rosettes of merozoites (Fig. 34). DISCUSSION Landsberg & Paperna (1985) state that Goussia gad1 develops in the gas gland but, as shown in this study, the gland itself is not affected, and the parasites develop only in loose connective tissue surrounding the rete mirabile of the gland. The finding by Fiebiger (1913) that there were no coccidia in the epithelia1 cells

8


9


10



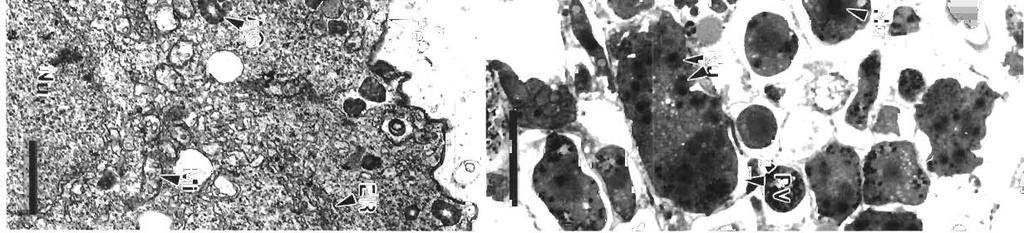
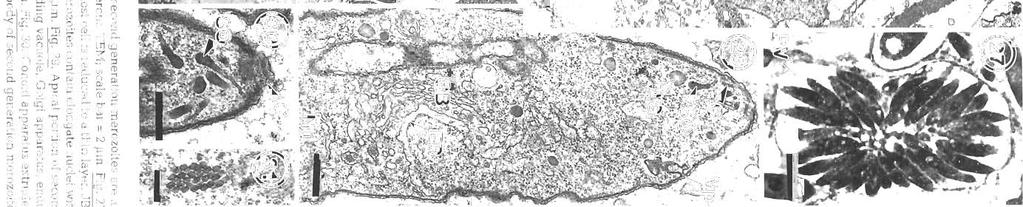
11 Morr~son et a1 Pathology dnd merogony ot Coussld gddi 123 F~gs 32 to 34 Go~rssia gadi parasitiz~ng Melanog~ammus deglefinus. Fig. 32. The residual body of the meront containing hpld droplets 15 surrounded by a single membrane, second generation merozoites are surrounded by a pellicle. =M; scale bar 1 pm Fig 33. Multlnucleate meront is partially divided into two by a membrane. TEM; scale bar = 2 5 pm. Fig 34. Two rosettes ol second generation merozoltes In a host cell vacuole. JB4, tolu~dlne blue; scale bar : 10 pm of the swimbladder is corroborated by the present study. The sporozoite-shaped trophozoltes usually found in phagocytes but also found free show no signs of degenerat~on, and appear to be able to enlarge and develop in the phagocyte. They may be derived from sporozoites or merozoites. Presumably those undergoing endodyogeny are der~ved from sporozoites, since similar stages were also fou.nd within the oocyst membrane Fiebiger (1913) described mult~nucleate sporozoites within the oocyst membrane, although he shows drawings only of blnucleate stages, as found in this study The osmiophilic vesicles at the ends of the trophozoite may be the remnants of the apical complex, and seem to be the means by which the parasite gains nutrients and disposes of waste products by pmocytosis. According to Chobotar & Scholtyseck (19821, coccidian sporozoites have often been reported In macrophages, and can repeatedly invade and escape from them. However, developmental stages such as the endodyogeny found In G, gad1 ale rarely found in other coccidlans in phagocytes The electron-dense layer around some trophozoites in phagocytes may be an early stage in the formation of the more obvious electron-lucent parasltophorous vacuole. The phagocytes apparently disintegrate and are reduced to membranous sacs, like the host cells containing the developing stages of G caseosa and G. cichlldarum (Landsberg & Paperna 1985, Paperna et a1 1986). It [vas difficult to ~dentlfy the leukocytes described In the present study, because specific stains for blood cell types were not carried out. and the ultrastructure of the leukocytes of cod has not been studied. The literature on tish granulocytes is contradictory (Ellis 1977). In the present study the amoeboid granulocytes containing numerous granules of varylng sizes are probably neutrophils (heterophils), the most commonly reported granulocyte in the peripheral blood of fish, although the granules are not elongate and striated as descnbed in some species (Hawkins et al. 1981). The leukocytes contaming numerous large, dense granules are probably eosinophils, and the granules are the same size as those reported In eoslnophlls by Hawkins et a1 (1981) Little is known about the function of these cells, although neutrophils migrate to inflammatory sites (Ellis 1977) Slnce there is an extensive host reaction to Gouss~a gadi in the swimblddder, it is likely that
.")
12 124 Dis. aquat. Org. 17; , 1993 the host may sometimes survive the infection. There is usually little host response to coccidian infections in fish, but extensive leukocyte infiltration was also found In response to infect~on by Calyptospora filnduli (Hawkins et al. 1981). The monocytes contained residual bodies, but no lipid droplets as reported in G. gad1 infections. In the latter, lipid probably accun~ulates due to the ingestion of stages of G. gadi containing lipid droplets. Fiebiger (1913) and Shulman & Shtein (1962) report that the stages of Goussia gad1 usually develop free in the connective tissue, but in the present account they were often found to be intracellular. It is not known whether phagocytes, probably macrophages, are the normal site of infection by sporozoitcs of G. gadi, or whether this only occurs in more advanced infections, such as those we studied, where numerous phagocytes had invaded the loose connective tissue layer of the swimbladder wall via the bloodstream. The merozoites resulting from endodyogeny could infect phagocytes or another type of host cell, such as a fibroblast. However, the original host cell type is difficult to determine since the cytoplasm around developing gamonts and multinucleate meronts, if still present, is reduced to a thin layer and is highly vacuolated; often only a membrane remains. However, no cc11 junctions were found between the host cells, indicdting that they were originally motile phagoc).tes. Moreover, no other host cell type is present in large numbers in the loose connective tissue of the swimbladder wa.11 (Morrison 1987). Nothing is known about the transmission of Goussia gadi to other hosts, although the possibilities are discussed by Fiebiger (1913). Like many species of Goussia, the life-cycle of G. gad1 takes place in an extra-intestinal site where oocysts have no means of discharge from the living host (Overstreet et al. 1984). Fiebiger suggests that infected fish could be ingested bp larger fish, but that the most likely mode of transmission is by death of the host, millions of spores being liberated after the abdominal and swimbladder walls disintegrate. Sporozoites may be released from the spores after ingestion by the piscine host, penetrate the intestinal epithelium, enter the bloodstream and be carried to the swimbladder. Since sporozoites easily enter phagocytes, and trophozoites of G, gadi are found in phagocytes in the connective tissue of the swimbladder, they could be transported in the bloodstream in this manner Presumably sporozoites could also be transported from the swimbladder to the intestine, and be released with the faeces to infect other hosts. Sporocysts have been reported in blood vessels near the swimbladder (Odense & Logan 1976), and so could presumably enter or exit the swimbladder by this route, although it is difficult to imagine how they could cross the endothelium lining the blood vessel. An intermediate host is not essential for Goussia gadi, since it is able to complete its cycle within the swimbladder, but one could be involved in transmission to another host. In G. carpelli (Steinhagen & Korting 1990) direct transmission by faecal contamindtion can occur, but reinfection of carp was only possible with a tubificid intermediate host. Overstreet (1981) has suggested that haddock are more likely to be infected by ingestion of a secondary host such as a crustacean containing G. gadi than by ingestion of an infected gadoid or its remains, since they are smallmouthed bottom feeders; but according to Scott & Scott (1988) they also eat fish such as elvers, herring, argentines, sdtld lance, capelin and silver hake. G. gadi was not found in 34 silver hake examined by Odense & Logan (1976). In Goussia gadi there appear to be 2 generations 01 merozoites, dividing by 2 different types of endogenesis. The firs1 type, endodyogeny, in which 2 daughter organisms form, using up the mother cell (Chobotar & Scholtyseck 1982), has rarely been described in piscine coccidians (Landsberg & Paperna 1987, Paperna 1991, Kim & Paperna 1992, Lukes 1992). Since there can be more than one cycle of this type of division, a group of first generation merozoites could be formed as shown in Figs. 15 & 16. First generation merozoites then apparently form multinucleate meronts which divide by endopolygeny, giving rise to a rosette which may contain more than 40 second generation merozoites (Fiebiger 1913). These differ from the groups of first generation rnerozoites in being more numerous and less convoluted, and in possessing numerous amylopectin granules and a small apical complex. The secretory organelles associated with this complex are open at the surface of the merozoite and are shaped like rhoptries, but they do not open through the conoid, and are not obviously paired. It has been suggested that micronernes may be an early stage in the formation of rhoptries, and may form a structural unit with them (Chobotar & Scholtyseck 1982), so the secretory organelles may be the forerunners of this system. The second generation merozoites were found to be longer in the present study than in the study by Fiebiger (1913), who reported that they were 8.0 X 2.5 pm. Endopolygeny has been described in several piscine coccidia (Paterson & Desser 1981, Steinhagen 1991), but Goussia gadi differs from these coccidia in that no vesicles are formed around the developing merozoltes, and more than one rosette may be found in a common vacuole, as reported by Fiebiger (1913). The radiating form of merogony found in G. gad1 would suggest development by exogenesis, but no early stages of exogenesis, in which the apical complexes of the merozoites develop next to the limiting membrane of the meront, were found (Paterson &
13 Morrison et al.: Pathology and merogony of Goussia gadi 125 Desser 1981, Jastrzqbski 1989). Although the merozoites originate internally in associat.ion with nuclei during endogenesis, in later development they assume a peripheral location and grow outward at the surface, so that the appearance is similar to that of exogenesis (Chobotar & Scholtyseck 1982). Acknowledgements. We thank Drs G. McCelland and D. Scarratt for their careful reviews of the manuscript, and helpful comments. LITERATURE CITED Chobotar, B., Scholtyseck, E. (1982). Ultrastructure. In: Long, P. L (ed.) The biology of the coccidia. Edward Arnold, London, p Desser. S. S. (1981). The challenge of fish coccidia. J. Protozool. 28: Dougherty, W. J. (1981). Preparation and staining of semi-thin sections of tissues embedded in water-soluble methacrylate for light microscopy. In: Clark, G. (ed.) Staining procedures, 4th edn. Williams and Wilkins Co., Baltimore, p Dykova, I., Lom, J. (1983) Fish coccidia: an annotated list of described species. Folia parasitol (Praha) 30: Ellis, A. E. (1977). The leucocytes of fish: a review. J. Fish Biol. 11: Fiebiger. J. (1913). Studien iiber die Schwimmblasencoccidien der Gadusarten (Eirneria gadi n.sp.). Arch. Protistenkd. 31: Hawkins, W. E., Solangi, M. A., Overstreet, R M. (1981). Ultrastructural effects of the coccidium, Ejmel-ia funduli Duszynski, Solangi and Overstreet, 1979 on the liver of killifishes. J. Fish Dis. 4: Humason. G. L. (1979). Animal tissue techniques, 4th edn. W. H. Freeman, San Francisco Jastrzebski. M. (1989). Ultrastructural study of the development of Goussia aculeati, a coccidium parasitizing the three-spined stickleback Gasterosteus aculeatus. Dis. aquat. Org. 6: Kim, S. H.. Paperna, 1. (1992). Fine structure of epicytoplasnlic stages of Eirneria vanasi from the gut of cichlid fish. Dis. aquat. Org. 12: Landsberg. J. H., Paperna. I. (1985). Goussja cichlidarurn n.sp. (Barrouxiidae, Apicomplexa), a coccidian parasite in the swimbladder of cichlid fish. 2. Parasitenkd. 71: Landsberg, J. H., Paperna, I. (1987). Intestinal infections by Eirneria (S.L.) vanasi n.sp. (Eimeriidae, Apicomplexa, Protozoa) in cichlid fish. Annls Parasit. hum. comp. 62: Lom, J., Dykova, I. (1982). Some marine fish coccidia of the genera Eirneria Schneider, Epieirneria Dykova and Lom and Goussia LabbB. J. Fish Dis. 5: Lukes, J. (1992). Life cycle of Goussia pannonica (Molnar, 1989) (Apicomplexa, Elmeriorina), an extracytoplasmic coccidium from the White Bream Blicca bjoerkna. J. Protozool. 39: McDowell, E. M. (1978). Fixation and processing. In: Trump, B F., Jones, R. T (eds ) Diagnostic electron microscopy. 1. John Wiley and Sons, New York Morrison, C. M. (1987). Histology of the Atlantic cod, Gadus morhua: an atlas. Part 1. Digestive tract and associated organs. Can. Spec. Publ. Fish. Aquat. Sci. 98 Morrison, C. M. (1993). Histology of the Atlantic cod, Gadus morhua: an atlas. Part 4. Eleutheroembryo and larva. Can. Spec. Publ. Fish Aqual. Sci. 119 Morrison, C. M., McClelland, G., Cornick, J., Marcogliese, D (1986). Parasites and diseases of some marine finfish off Nova Scotia. Can. Tech. Rep. Fish. Aquat. Sci Odense, P. H., Logan, V. H. (1976). Prevalence and morphology of Eimeria gad1 (Fiebiger. 1913) in the haddock. J. Protozool. 23: Overstreet, R. M. (1981). Species of Eimeria in nonepithelial sites. J. Protozool Overstreet, R. M., Hawkins, W. E., Fournie, J W. (1984). The coccidian genus Calyptospora n. g. and family Calyptosporidae n. fam. (Apicomplexa), with members infecting primarily fishes. J. Protozool. 3: Paperna, 1. (1991). Fine structure of Eirneria (S. I.) vanasi merogony stages in the intestinal mucosa of cichlid fishes. DIS. aquat. Org Paperna, I., Landsberg, J. H., Feinstein, N. (1986). Ultrastructure of the macrogamont of Goussja clchljdarurn Landsberg and Paperna, 1985, a coccidian parasite in the swimbladder of cichlid fish. Annls Parasit. hum. comp. 61: Paterson, W. B., Desser, S. S. (1981). An ultrastructural study of Eimeria iroquoina Molnar and Fernando, 1974 in experimentally ~nfected fathead minnows (Pimephales promelas, Cypnnidde). 3. Merogony. J. Protozool Reynolds, E. S. (1963). The use of lead citrate at a high ph as an electron-opaque stain in electron mlcroscopy. J Cell Biol. 17: Scott, J. S. (1981). Alimentary tract parasites of haddock (Melanogrammus aeglefinus L.) on the Scotian Shelf. Can. J Zool. 59: Scott, W. B., Scott, M G (1988). Atlantic flshes of Canada Can. Bull. Fish. Aquat. Sci. 219 Shulman, S. S., Shtein, G. A. (1962). Phylum Protozoa. Unicellular animals. In: Bykhovskii, B. E. (ed.) Key to parasites of freshwater fish of the U.S.S.R. Acad. Sci. S.S.S.R., Moscow, Leningrad. (Transl. and republ. by Israel Prog. Sci. Transl., Jerusalem, p ) Steinhagen, D. (1991) Ultrastructural observations on merogonic and gamogonic stages of Goussia cdrpell~ (Apicomplexa, Coccldia) in experimentally infected common carp Cyprinus carpio. Eur. J. Protistol. 27: Steinhagen, D., Korting. W. (1990). The role of tubificid oligochaetes in the transmission of Goussia carpelli. J. Parasitol. 76: Stempack, J. G., Ward, R. T. (1964). An improved staining method for electron microscopy. J. Cell Blol. 22: Upton, S. J., Reduker. D. W., Current, W. L., Duszynski, D. W. (1984). Taxonomy of North American fish Eirneriidae NOAA Tech. Rep. NMFS 11, Washington. DC Responsible Subject Editor: W. Korting, Hannover, Germany Manuscript first received: March 1, 1993 Revised version accepted: July 5, 1993
DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org.
 Vol. 10: 121-125. 1991 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. Published April 4 Ultrastructural observations on sporozoite stages of piscine Coccidia: Goussia carpelli and G. subepithelialis from
Vol. 10: 121-125. 1991 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. Published April 4 Ultrastructural observations on sporozoite stages of piscine Coccidia: Goussia carpelli and G. subepithelialis from
Key words: Coccidia, Choleoeimeria rochalimai, fine structure, gall bladder epithelium, Hemidactylus mabouia, Brazil
 FOLIA PARASITOLOGICA 47: 91-96, 2000 Ultrastructural study of meronts and gamonts of Choleoeimeria rochalimai (Apicomplexa: Eimeriidae) developing in the gall bladder of the gecko Hemidactylus mabouia
FOLIA PARASITOLOGICA 47: 91-96, 2000 Ultrastructural study of meronts and gamonts of Choleoeimeria rochalimai (Apicomplexa: Eimeriidae) developing in the gall bladder of the gecko Hemidactylus mabouia
Oocyst formation in the coccidian parasite Goussia carpelli
 Vol. 10: 203-209, 1991 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. Published May 8 Oocyst formation in the coccidian parasite Goussia carpelli 'Institute of Parasitology, Czech Academy of Sciences,
Vol. 10: 203-209, 1991 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. Published May 8 Oocyst formation in the coccidian parasite Goussia carpelli 'Institute of Parasitology, Czech Academy of Sciences,
Fine structure of Eimeria (S. l.) vanasi merogony stages in the intestinal mucosa of cichlid fishes
 vanasi merogony stages in the intestinal mucosa of cichlid fishes") Vol. 10: 195-201. 1991 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. Published May 8 Fine structure of Eimeria (S. l.) vanasi merogony stages in the intestinal mucosa of cichlid fishes Ilan Paperna Department
Vol. 10: 195-201. 1991 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. Published May 8 Fine structure of Eimeria (S. l.) vanasi merogony stages in the intestinal mucosa of cichlid fishes Ilan Paperna Department
The Fine Structure of the Endogenous Stages of Isospora hemidactyli Carini, 1936 in the Gecko Hemidactylus mabouia from North Brazil
 Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 95(1): 43-47, Jan./Feb. 2000 The Fine Structure of the Endogenous Stages of Isospora hemidactyli Carini, 1936 in the Gecko Hemidactylus mabouia from North Brazil
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 95(1): 43-47, Jan./Feb. 2000 The Fine Structure of the Endogenous Stages of Isospora hemidactyli Carini, 1936 in the Gecko Hemidactylus mabouia from North Brazil
Ultrastructure of Endogenous Stages of Eimeria ninakohlyakimovae Yakimoff & Rastegaieff, 1930 Emend. Levine, 1961 in Experimentally Infected Goat
 Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 92(4): 533-538, Jul./Aug. 1997 Ultrastructure of Endogenous Stages of Eimeria ninakohlyakimovae Yakimoff & Rastegaieff, 1930 Emend. Levine, 1961 in Experimentally
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 92(4): 533-538, Jul./Aug. 1997 Ultrastructure of Endogenous Stages of Eimeria ninakohlyakimovae Yakimoff & Rastegaieff, 1930 Emend. Levine, 1961 in Experimentally
Phylum:Apicomplexa Class:Sporozoa
 Phylum:Apicomplexa Class:Sporozoa The most characteristic features of sporozoa are 1-unique appearance of most protozoa makes it possible for knowledge able person to identifiy them to level of genus and
Phylum:Apicomplexa Class:Sporozoa The most characteristic features of sporozoa are 1-unique appearance of most protozoa makes it possible for knowledge able person to identifiy them to level of genus and
Protozoa. Apicomplexa Sarcomastigophora Ciliophora. Gregarinea Coccidia Piroplasma
 Protozoa Apicomplexa Sarcomastigophora Ciliophora Gregarinea Coccidia Piroplasma Coccidia characterized by thick-walled oocysts excreted in feces In Humans Cryptosporidium Isospora Cyclospora Sarcocystis
Protozoa Apicomplexa Sarcomastigophora Ciliophora Gregarinea Coccidia Piroplasma Coccidia characterized by thick-walled oocysts excreted in feces In Humans Cryptosporidium Isospora Cyclospora Sarcocystis
Key words: Plasmodium, Kentropyx calcarata, Brazil, merogony, gametocytes, ultrastructure
 FOLIA PARASITOLOGICA 49: 2-8, 2002 Fine structure of erythrocytic stages of a Plasmodium tropiduri-like malaria parasite found in the lizard Kentropyx calcarata (Teiidae) from north Brazil Ilan Paperna
FOLIA PARASITOLOGICA 49: 2-8, 2002 Fine structure of erythrocytic stages of a Plasmodium tropiduri-like malaria parasite found in the lizard Kentropyx calcarata (Teiidae) from north Brazil Ilan Paperna
HISTOPATHOLOGY. Introduction:
 Introduction: HISTOPATHOLOGY Goats and sheep are the major domestic animal species in India. Much of the economy of the country has been depend upon the domestication of these animals. Especially economy
Introduction: HISTOPATHOLOGY Goats and sheep are the major domestic animal species in India. Much of the economy of the country has been depend upon the domestication of these animals. Especially economy
Revajová, Viera, Loószová, Adrian. The Journal of Protozoology Resea Citation RightsNational Research Center for Prot
 ' ' Morphological study of partridge Title development in the foreign host - (Gallus gallus) Revajová, Viera, Loószová, Adrian Author(s) Maria, Zibrín, Martin, Herich, Ro Mikulas The Journal of Protozoology
' ' Morphological study of partridge Title development in the foreign host - (Gallus gallus) Revajová, Viera, Loószová, Adrian Author(s) Maria, Zibrín, Martin, Herich, Ro Mikulas The Journal of Protozoology
HISTOLOGY OF MAMMARY GLAND DURING LACTATING AND NON-LACTATING PHASES OF MADRAS RED SHEEP WITH SPECIAL REFERENCE TO INVOLUTION
 International Journal of Science, Environment and Technology, Vol. 5, No 3, 2016, 991 996 ISSN 2278-3687 (O) 2277-663X (P) HISTOLOGY OF MAMMARY GLAND DURING LACTATING AND NON-LACTATING PHASES OF MADRAS
International Journal of Science, Environment and Technology, Vol. 5, No 3, 2016, 991 996 ISSN 2278-3687 (O) 2277-663X (P) HISTOLOGY OF MAMMARY GLAND DURING LACTATING AND NON-LACTATING PHASES OF MADRAS
Biology of toxoplasmosis
 1 Biology of toxoplasmosis E. Petersen 1 and J. P. Dubey 2 1 Statens Seruminstitut, Copenhagen, Denmark 2 U.S. Department of Agriculture, Beltsville, USA History Toxoplasma gondii is a coccidium, with
1 Biology of toxoplasmosis E. Petersen 1 and J. P. Dubey 2 1 Statens Seruminstitut, Copenhagen, Denmark 2 U.S. Department of Agriculture, Beltsville, USA History Toxoplasma gondii is a coccidium, with
Ëtude ultrastructurale de la mérogonie de Schellackia cf. agamae (Lankesterellidae, Apicomplexa) chez le Lézard Agama stellio.
 chez le Lézard Agama stellio.") Masson, Paris, 1987 Ann. Parasitol. Hum. Comp. 1987, 62, n 5, pp. 380-386. ULTRASTRUCTURAL STUDIES ON THE MEROGONY OF SCHELLACKIA CF. AGAMAE (LANKESTERELLIDAE, APICOMPLEXA) FROM THE STARRED LIZARD AGAMA
Masson, Paris, 1987 Ann. Parasitol. Hum. Comp. 1987, 62, n 5, pp. 380-386. ULTRASTRUCTURAL STUDIES ON THE MEROGONY OF SCHELLACKIA CF. AGAMAE (LANKESTERELLIDAE, APICOMPLEXA) FROM THE STARRED LIZARD AGAMA
Sarcocystis heydorni, n. sp. (Apicomplexa: Protozoa) with cattle (Bos taurus) and human
 with cattle (Bos taurus) and human") 1 Sarcocystis heydorni, n. sp. (Apicomplexa: Protozoa) with cattle (Bos taurus) and human (Homo sapiens) cycle Jitender P. Dubey 1, Erna van Wilpe 2, Rafael Calero-Bernal 1, Shiv Kumar Verma 1, Ronald
1 Sarcocystis heydorni, n. sp. (Apicomplexa: Protozoa) with cattle (Bos taurus) and human (Homo sapiens) cycle Jitender P. Dubey 1, Erna van Wilpe 2, Rafael Calero-Bernal 1, Shiv Kumar Verma 1, Ronald
The specimens of Ameiva ameiva (Linn) were
 were") Article available at http://www.parasite-journal.org or http://dx.doi.org/10.1051/parasite/1999064359 FINE STRUCTURE OF THE EPICYTOPLASMIC EIMERID COCCIDIUM ACROEIMERIA PINTOI LAINSON & PAPERNA, 1999,
Article available at http://www.parasite-journal.org or http://dx.doi.org/10.1051/parasite/1999064359 FINE STRUCTURE OF THE EPICYTOPLASMIC EIMERID COCCIDIUM ACROEIMERIA PINTOI LAINSON & PAPERNA, 1999,
Extra-intestinal localization of Goussia sp. (Apicomplexa) oocysts in Rana dalmatina (Anura: Ranidae), and the fate of infection after metamorphosis
 oocysts in Rana dalmatina (Anura: Ranidae), and the fate of infection after metamorphosis") DISEASES OF AQUATIC ORGANISMS Vol. 70: 237 241, 2006 Published June 23 Dis Aquat Org Extra-intestinal localization of Goussia sp. (Apicomplexa) oocysts in Rana dalmatina (Anura: Ranidae), and the fate
DISEASES OF AQUATIC ORGANISMS Vol. 70: 237 241, 2006 Published June 23 Dis Aquat Org Extra-intestinal localization of Goussia sp. (Apicomplexa) oocysts in Rana dalmatina (Anura: Ranidae), and the fate
PLASMODIUM MODULE 39.1 INTRODUCTION OBJECTIVES 39.2 MALARIAL PARASITE. Notes
 Plasmodium MODULE 39 PLASMODIUM 39.1 INTRODUCTION Malaria is characterized by intermittent fever associated with chills and rigors in the patient. There may be enlargement of the liver and spleen in the
Plasmodium MODULE 39 PLASMODIUM 39.1 INTRODUCTION Malaria is characterized by intermittent fever associated with chills and rigors in the patient. There may be enlargement of the liver and spleen in the
Apicomplexans Apicomplexa Intro
 Apicomplexans Apicomplexa Intro Cryptosporidium Apicomplexan Select Characteristics Gliding motility Apical Complex organelle for invasion of host cell Life cycle alternates b/w sexual and asexual phases
Apicomplexans Apicomplexa Intro Cryptosporidium Apicomplexan Select Characteristics Gliding motility Apical Complex organelle for invasion of host cell Life cycle alternates b/w sexual and asexual phases
Ahead of print online version
 Folia Parasitologica 60 [3]: 232 236, 2013 ISSN 0015-5683 (print), ISSN 1803-6465 (online) Institute of Parasitology, Biology Centre ASCR http://folia.paru.cas.cz/ A new species of Choleoeimeria (Apicomplexa:
Folia Parasitologica 60 [3]: 232 236, 2013 ISSN 0015-5683 (print), ISSN 1803-6465 (online) Institute of Parasitology, Biology Centre ASCR http://folia.paru.cas.cz/ A new species of Choleoeimeria (Apicomplexa:
A:Malaria (Plasmodium species) Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign
 Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign") A:Malaria (Plasmodium species) Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign tertian malaria P. ovale: causes benign tertian malaria
A:Malaria (Plasmodium species) Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign tertian malaria P. ovale: causes benign tertian malaria
DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org.
 Vol. 7: 149-153, 1989 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. Published October 26 Developmental cycle of chelonian haemogregarines in leeches with extra-intestinal multiple sporozoite oocysts and
Vol. 7: 149-153, 1989 DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. Published October 26 Developmental cycle of chelonian haemogregarines in leeches with extra-intestinal multiple sporozoite oocysts and
Coccidia. Nimit Morakote, Ph.D.
 Coccidia Nimit Morakote, Ph.D. 1 Learning objectives After class, students will be able to: Describe morphology, life cycle, signs and symptoms, prevention and control, laboratory diagnosis and treatment
Coccidia Nimit Morakote, Ph.D. 1 Learning objectives After class, students will be able to: Describe morphology, life cycle, signs and symptoms, prevention and control, laboratory diagnosis and treatment
Protozoan Parasites of Veterinary importance 2017
 Protozoan Parasites of Veterinary importance 2017 VPM-122 Laboratory 4 Spencer J. Greenwood PhD, DVM Dept. of Biomedical Sciences Room 2332N AVC North Annex sgreenwood@upei.ca Office phone # 566-6002 To
Protozoan Parasites of Veterinary importance 2017 VPM-122 Laboratory 4 Spencer J. Greenwood PhD, DVM Dept. of Biomedical Sciences Room 2332N AVC North Annex sgreenwood@upei.ca Office phone # 566-6002 To
Liver and Gallbladder Morphology of the juvenile Nile crocodile, Crocodylus niloticus (Laurenti, 1768)
") Liver and Gallbladder Morphology of the juvenile Nile crocodile, Crocodylus niloticus (Laurenti, 1768) by ERNA VAN WILPE Submitted in partial fulfilment of the requirements for the degree MSc DEPARTMENT
Liver and Gallbladder Morphology of the juvenile Nile crocodile, Crocodylus niloticus (Laurenti, 1768) by ERNA VAN WILPE Submitted in partial fulfilment of the requirements for the degree MSc DEPARTMENT
' Published September 29
 Vol. 7: 1-12, 1989 l DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. ' Published September 29 Nodular and epicellular coccidiosis in the intestine of cyprinid fishes Kalman Molnar Veterinary Medical Research
Vol. 7: 1-12, 1989 l DISEASES OF AQUATIC ORGANISMS Dis. aquat. Org. ' Published September 29 Nodular and epicellular coccidiosis in the intestine of cyprinid fishes Kalman Molnar Veterinary Medical Research
A comparison of placental tissue in the skinks Eulamprus tympanum and E. quoyii. Yates, Lauren A.
 A comparison of placental tissue in the skinks Eulamprus tympanum and E. quoyii Yates, Lauren A. Abstract: The species Eulamprus tympanum and Eulamprus quoyii are viviparous skinks that are said to have
A comparison of placental tissue in the skinks Eulamprus tympanum and E. quoyii Yates, Lauren A. Abstract: The species Eulamprus tympanum and Eulamprus quoyii are viviparous skinks that are said to have
COCCIDIOSIS OF SANDHILL CRANES (GRUS CANADENSIS) WINTERING IN NEW MEXICO
 WINTERING IN NEW MEXICO") journal of Wtldltfe hemes, 22(1). 1986. pp 25-35 0 Wildlife Disease Association 1986 COCCIDIOSIS OF SANDHILL CRANES (GRUS CANADENSIS) WINTERING IN NEW MEXICO Brent B. Parker and Donald W. Duszynski Department
journal of Wtldltfe hemes, 22(1). 1986. pp 25-35 0 Wildlife Disease Association 1986 COCCIDIOSIS OF SANDHILL CRANES (GRUS CANADENSIS) WINTERING IN NEW MEXICO Brent B. Parker and Donald W. Duszynski Department
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11046 Supplementary Figure 1: Images of PB-positive cells in the subepidermal region (a-i) Representative images of PB positive cells in the subepidermis of the upper beak of the pigeon.
doi:10.1038/nature11046 Supplementary Figure 1: Images of PB-positive cells in the subepidermal region (a-i) Representative images of PB positive cells in the subepidermis of the upper beak of the pigeon.
A Scanning Electron Microscopic Study of Eggshell Surface Topography of Leidynema portentosae and L. appendiculatum (Nematoda: Oxyuroidea)
") The Ohio State University Knowledge Bank kb.osu.edu Ohio Journal of Science (Ohio Academy of Science) Ohio Journal of Science: Volume 88, Issue 5 (December, 1988) 1988-12 A Scanning Electron Microscopic
The Ohio State University Knowledge Bank kb.osu.edu Ohio Journal of Science (Ohio Academy of Science) Ohio Journal of Science: Volume 88, Issue 5 (December, 1988) 1988-12 A Scanning Electron Microscopic
DISEASES OF AQUATIC ORGANISMS Vol. 62: , 2004 Published November 23 Dis Aquat Org
 DISEASES OF AQUATIC ORGANISMS Vol. 62: 133 145, 2004 Published November 23 Dis Aquat Org Cryptosporidium scophthalmi n. sp. (Apicomplexa: Cryptosporidiidae) from cultured turbot Scophthalmus maximus. Light
DISEASES OF AQUATIC ORGANISMS Vol. 62: 133 145, 2004 Published November 23 Dis Aquat Org Cryptosporidium scophthalmi n. sp. (Apicomplexa: Cryptosporidiidae) from cultured turbot Scophthalmus maximus. Light
SCANNING electron - microscopy has
 Characteristics of the Absorptive Surface of the Small Intestine of the Chicken from 1 Day to 14 Weeks of Age 1 R. C. BAYER, C. B. CHAWAN, F. H. BIRD AND S. D. MUSGRAVE Department of Animal and Veterinary
Characteristics of the Absorptive Surface of the Small Intestine of the Chicken from 1 Day to 14 Weeks of Age 1 R. C. BAYER, C. B. CHAWAN, F. H. BIRD AND S. D. MUSGRAVE Department of Animal and Veterinary
,,, THE MORPHOLOGY AND MORPHOMETRY OF THE PECTEN OCULI IN DIURNAL AND NOCTURNAL BIRDS: A
 ,,, THE MORPHOLOGY AND MORPHOMETRY OF THE PECTEN OCULI IN DIURNAL AND NOCTURNAL BIRDS: A COMPARATIVE STUDY" BY llijama, S.G., B. V. M. (NBI), Department of Veteri nary Anatomy, University of I\Jairobi.
,,, THE MORPHOLOGY AND MORPHOMETRY OF THE PECTEN OCULI IN DIURNAL AND NOCTURNAL BIRDS: A COMPARATIVE STUDY" BY llijama, S.G., B. V. M. (NBI), Department of Veteri nary Anatomy, University of I\Jairobi.
EFFICACY OF SOME ANTICOCCIDIAL DRUGS FOR TREATING COCCIDIAL ENTERITIS OF THE COMMON CARP CAUSED BY GOUSSIA CARPELLI (APICOMPLEXA: EIMERIIDAE)
") Acta Veterinaria Hungarica 55 (1), pp. 67 76 (2007) DOI: 10.1556/AVet.55.2007.1.7 EFFICACY OF SOME ANTICOCCIDIAL DRUGS FOR TREATING COCCIDIAL ENTERITIS OF THE COMMON CARP CAUSED BY GOUSSIA CARPELLI (APICOMPLEXA:
Acta Veterinaria Hungarica 55 (1), pp. 67 76 (2007) DOI: 10.1556/AVet.55.2007.1.7 EFFICACY OF SOME ANTICOCCIDIAL DRUGS FOR TREATING COCCIDIAL ENTERITIS OF THE COMMON CARP CAUSED BY GOUSSIA CARPELLI (APICOMPLEXA:
Mesosomes are a definite event in antibiotic-treated Staphylococcus aureus ATCC 25923
 Tropical Biomedicine 24(1): 105 109 (2007) Mesosomes are a definite event in antibiotic-treated Staphylococcus aureus ATCC 25923 Santhana Raj, L. 1*, Hing, H.L. 2, Baharudin Omar 2, Teh Hamidah, Z. 1,
Tropical Biomedicine 24(1): 105 109 (2007) Mesosomes are a definite event in antibiotic-treated Staphylococcus aureus ATCC 25923 Santhana Raj, L. 1*, Hing, H.L. 2, Baharudin Omar 2, Teh Hamidah, Z. 1,
Apicomplexa of Intestinal Pathology
 LECTURES #4, #5 & #6: APICOMPLEXA 1 Apicomplexa of Intestinal Pathology Cryptosporidium, Eimeria, Cystoisospora General Characteristics of Apicomplexa A. Morphology by stage Zoite o Tear-shaped (cylindrical
LECTURES #4, #5 & #6: APICOMPLEXA 1 Apicomplexa of Intestinal Pathology Cryptosporidium, Eimeria, Cystoisospora General Characteristics of Apicomplexa A. Morphology by stage Zoite o Tear-shaped (cylindrical
A Lymphosarcoma in an Atlantic Salmon (Salmo salar)
") A Lymphosarcoma in an Atlantic Salmon (Salmo salar) Authors: Paul R. Bowser, Marilyn J. Wolfe, and Timothy Wallbridge Source: Journal of Wildlife Diseases, 23(4) : 698-701 Published By: Wildlife Disease
A Lymphosarcoma in an Atlantic Salmon (Salmo salar) Authors: Paul R. Bowser, Marilyn J. Wolfe, and Timothy Wallbridge Source: Journal of Wildlife Diseases, 23(4) : 698-701 Published By: Wildlife Disease
A. Effect upon human culture 1. Control of malaria has contributed to world=s population explosion 2. Africans brought to U.S.
 VI. Malaria A. Effect upon human culture 1. Control of malaria has contributed to world=s population explosion 2. Africans brought to U.S. because they were resistant to malaria & other diseases 3. Many
VI. Malaria A. Effect upon human culture 1. Control of malaria has contributed to world=s population explosion 2. Africans brought to U.S. because they were resistant to malaria & other diseases 3. Many
Lacerta viridis. Functional anatomy of the lungs of the green lizard, (Accepted 18 February 1977)
") J. Anat. (1978), 125, 2, pp. 421-431 421 With 9 figures Printed in Great Britain Functional anatomy of the lungs of the green lizard, Lacerta viridis C. MEBAN Department of Anatomy, The Queen's University
J. Anat. (1978), 125, 2, pp. 421-431 421 With 9 figures Printed in Great Britain Functional anatomy of the lungs of the green lizard, Lacerta viridis C. MEBAN Department of Anatomy, The Queen's University
Hepatozoon-Like Parasite (Schizonts) in the Myocardium of the Domestic Cat
 in the Myocardium of the Domestic Cat") Vet. Path. 10: 185-190 (1973) Hepatozoon-Like Parasite (Schizonts) in the Myocardium of the Domestic Cat U. KLOPFER, T.A. NOBEL and F. NEUMANN Department of Pathology, Kimron Veterinary Institute, affiliated
Vet. Path. 10: 185-190 (1973) Hepatozoon-Like Parasite (Schizonts) in the Myocardium of the Domestic Cat U. KLOPFER, T.A. NOBEL and F. NEUMANN Department of Pathology, Kimron Veterinary Institute, affiliated
Redescription of Sarcocystis fusiformis sarcocysts from the water buffalo (Bubalus bubalis)
") Redescription of Sarcocystis fusiformis sarcocysts from the water buffalo (Bubalus bubalis) 1 J. P. DUBEY 1 *, M. HILALI 2,E.VANWILPE 3,S.K.VERMA 1,R.CALERO-BERNAL 1 and A. ABDEL-WAHAB 2 1 U. S. Department
Redescription of Sarcocystis fusiformis sarcocysts from the water buffalo (Bubalus bubalis) 1 J. P. DUBEY 1 *, M. HILALI 2,E.VANWILPE 3,S.K.VERMA 1,R.CALERO-BERNAL 1 and A. ABDEL-WAHAB 2 1 U. S. Department
Coccidiosis in macropods and other species
 Coccidiosis in macropods and other species Author: Derek Spielman Wildlife Assistance and Information Foundation; Sydney School of Veterinary Science, the University of Sydney Abstract This presentation
Coccidiosis in macropods and other species Author: Derek Spielman Wildlife Assistance and Information Foundation; Sydney School of Veterinary Science, the University of Sydney Abstract This presentation
Dermatitis in a dog associated with an unidentified Toxoplasma gondii-like parasite
 Veterinary Parasitology 116 (2003) 51 59 Short communication Dermatitis in a dog associated with an unidentified Toxoplasma gondii-like parasite J.P. Dubey a,, A.L. Pimenta b, L.C.S. Abboud b, R.R. Ravasani
Veterinary Parasitology 116 (2003) 51 59 Short communication Dermatitis in a dog associated with an unidentified Toxoplasma gondii-like parasite J.P. Dubey a,, A.L. Pimenta b, L.C.S. Abboud b, R.R. Ravasani
BIO Parasitology Spring 2009
 BIO 475 - Parasitology Spring 2009 Stephen M. Shuster Northern Arizona University http://www4.nau.edu/isopod Lecture 10 Malaria-Life Cycle a. Micro and macrogametocytes in mosquito stomach. b. Ookinete
BIO 475 - Parasitology Spring 2009 Stephen M. Shuster Northern Arizona University http://www4.nau.edu/isopod Lecture 10 Malaria-Life Cycle a. Micro and macrogametocytes in mosquito stomach. b. Ookinete
Taxonomy of North American Fish Eimeriidae
 11 NOAA Technical Report NMFS 11 Taxonomy of North American Fish Eimeriidae Steve J. Upton, David W. Reduker, William L. Current, and Donald W. Duszynski August 1984 U.S. DEPARTMENT OF COMMERCE National
11 NOAA Technical Report NMFS 11 Taxonomy of North American Fish Eimeriidae Steve J. Upton, David W. Reduker, William L. Current, and Donald W. Duszynski August 1984 U.S. DEPARTMENT OF COMMERCE National
Malaria parasites of rodents of the Congo (Brazzaville) :
 :") Annales de Parasitologie (Paris), 1976, t. 51, n 6, pp. 637 à 646 Malaria parasites of rodents of the Congo (Brazzaville) : Plasmodium cbabaudi adami subsp. nov. and Plasmodium vinckei lentum Landau, Michel,
Annales de Parasitologie (Paris), 1976, t. 51, n 6, pp. 637 à 646 Malaria parasites of rodents of the Congo (Brazzaville) : Plasmodium cbabaudi adami subsp. nov. and Plasmodium vinckei lentum Landau, Michel,
Parasitology Amoebas. Sarcodina. Mastigophora
 Parasitology Amoebas Sarcodina Entamoeba hisolytica (histo = tissue, lytica = lyse or break) (pathogenic form) o Trophozoite is the feeding form o Life Cycle: personfeces cyst with 4 nuclei with thicker
Parasitology Amoebas Sarcodina Entamoeba hisolytica (histo = tissue, lytica = lyse or break) (pathogenic form) o Trophozoite is the feeding form o Life Cycle: personfeces cyst with 4 nuclei with thicker
cyst&' appeared to be of two kinds-one smaller and Smnith "is inclined to regard these epithelial cell parasites as
 COCCIDIA IN SUBEPITHELIAL INFECTIONS OF THE INTESTINES OF BIRDS PHILIP B. HADLEY From the Agricultural Experiment Station of the Rhode Island State College' Received for publication, July 10, 1916 In an
COCCIDIA IN SUBEPITHELIAL INFECTIONS OF THE INTESTINES OF BIRDS PHILIP B. HADLEY From the Agricultural Experiment Station of the Rhode Island State College' Received for publication, July 10, 1916 In an
TARENTANNULARI INFECTING THE GECKO TARENTOLA ANNULARIS. Department of Zoology, Faculty of Science, University of Ain Shams, Cairo, Egypt - - -
 Qatar Univ. Sci. J. (1995), 15 (2) : 379-387 THE ULTRASTRUCTURE OF SOME STAGES OF HAEMOGREGARINA TARENTANNULARI INFECTING THE GECKO TARENTOLA ANNULARIS BY Nadia F. Ramadan, Shadia H. Mohammed and Samia
Qatar Univ. Sci. J. (1995), 15 (2) : 379-387 THE ULTRASTRUCTURE OF SOME STAGES OF HAEMOGREGARINA TARENTANNULARI INFECTING THE GECKO TARENTOLA ANNULARIS BY Nadia F. Ramadan, Shadia H. Mohammed and Samia
Sam R. Telford, Jr The Florida Museum of Natural History, University of Florida, Gainesville, Fl32611, USA
 Systematic Parasitology 23: 203-208, 1992. 0 1992 Kluwer Academic Publishers. Printed in the Netherlands. An eimeriid species (Apicomplexa: Eimeriidae) that parasitises the gallbladder and bile-duct of
Systematic Parasitology 23: 203-208, 1992. 0 1992 Kluwer Academic Publishers. Printed in the Netherlands. An eimeriid species (Apicomplexa: Eimeriidae) that parasitises the gallbladder and bile-duct of
HYDATID CYST DISEASE
 HYDATID CYST DISEASE Hydatid disease, also called hydatidosis or echinococcosis, is a cystforming disease resulting from an infection with the metacestode, or larval form, of parasitic dog tapeworms from
HYDATID CYST DISEASE Hydatid disease, also called hydatidosis or echinococcosis, is a cystforming disease resulting from an infection with the metacestode, or larval form, of parasitic dog tapeworms from
Observations on Eimeria species of Dasyprocta leporina (Linnaeus, 1758) (Rodentia: Dasyproctidae) from the state of Pará, North Brazil
 (Rodentia: Dasyproctidae) from the state of Pará, North Brazil") Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99: 000-000, 2004 1 Observations on Eimeria species of Dasyprocta leporina (Linnaeus, 1758) (Rodentia: Dasyproctidae) from the state of Pará, North Brazil Ralph
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99: 000-000, 2004 1 Observations on Eimeria species of Dasyprocta leporina (Linnaeus, 1758) (Rodentia: Dasyproctidae) from the state of Pará, North Brazil Ralph
Flatworms Flatworms Platyhelminthes dorsoventrally free-living planarian parasitic fluke tapeworm label three body layers ectoderm mesoderm
 Flatworms Flatworms are in the phylum Platyhelminthes. Flatworms are flattened dorsoventrally (top to bottom). The group includes the freshwater, free-living planarian and the parasitic fluke and tapeworm.
Flatworms Flatworms are in the phylum Platyhelminthes. Flatworms are flattened dorsoventrally (top to bottom). The group includes the freshwater, free-living planarian and the parasitic fluke and tapeworm.
Progressive Retinal Atrophy in the Abyssinian Cat
 Progressive Retinal Atrophy in the Abyssinian Cat Electron Microscopy Kristina Narfstr6m*t and Sven Erik Nilsson* Seven adult Abyssinian cats at different stages of a recessively inherited retinal degenerative
Progressive Retinal Atrophy in the Abyssinian Cat Electron Microscopy Kristina Narfstr6m*t and Sven Erik Nilsson* Seven adult Abyssinian cats at different stages of a recessively inherited retinal degenerative
Diagnosis, treatment and control: dealing with coccidiosis in cattle
 Vet Times The website for the veterinary profession https://www.vettimes.co.uk Diagnosis, treatment and control: dealing with coccidiosis in cattle Author : Adam Martin Categories : Vets Date : January
Vet Times The website for the veterinary profession https://www.vettimes.co.uk Diagnosis, treatment and control: dealing with coccidiosis in cattle Author : Adam Martin Categories : Vets Date : January
Article available at or
 Article available at http://www.parasite-journal.org or http://dx.doi.org/10.1051/parasite/1998051017 PSEUDOKLOSSIA SEMILUNA N. SP. (APICOMPLEXA: AGGREGATIDAE): A COCCIDIAN PARASITE OF THE KIDNEY OF BLUE
Article available at http://www.parasite-journal.org or http://dx.doi.org/10.1051/parasite/1998051017 PSEUDOKLOSSIA SEMILUNA N. SP. (APICOMPLEXA: AGGREGATIDAE): A COCCIDIAN PARASITE OF THE KIDNEY OF BLUE
Joerg Kinne, Mansoor Ali*, Ulrich Wernery, and J. P. Dubey
 J. Parasitol., 88(3), 2002, pp. 548 552 American Society of Parasitologists 2002 CLINICAL LARGE INTESTINAL COCCIDIOSIS IN CAMELS (CAMELUS DROMEDARIUS) IN THE UNITED ARAB EMIRATES: DESCRIPTION OF LESIONS,
J. Parasitol., 88(3), 2002, pp. 548 552 American Society of Parasitologists 2002 CLINICAL LARGE INTESTINAL COCCIDIOSIS IN CAMELS (CAMELUS DROMEDARIUS) IN THE UNITED ARAB EMIRATES: DESCRIPTION OF LESIONS,
BEAK AND FEATHER DYSTROPHY IN WILD SULPHUR-CRESTED COCKATOOS (CACATUA GALERITA)
") BEAK AND FEATHER DYSTROPHY IN WILD SULPHUR-CRESTED COCKATOOS (CACATUA GALERITA) Author(s): Steven McOrist, Douglas G. Black, David A. Pass, Peter C. Scott, and John Marshall Source: Journal of Wildlife
BEAK AND FEATHER DYSTROPHY IN WILD SULPHUR-CRESTED COCKATOOS (CACATUA GALERITA) Author(s): Steven McOrist, Douglas G. Black, David A. Pass, Peter C. Scott, and John Marshall Source: Journal of Wildlife
Effect of Sodium Hypochlorite on the Oocyst Wall of Eimeria tenella as Shown by Electron Microscopy1
 32 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY This alteration appeared similar to that observed by light microscopy (Figs. 5, 6). Literature Cited Dixon, K. E. 1966. The physiology of excystment of the
32 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY This alteration appeared similar to that observed by light microscopy (Figs. 5, 6). Literature Cited Dixon, K. E. 1966. The physiology of excystment of the
Phylum Platyhelminthes Flatworms
 Phylum Platyhelminthes Flatworms The Acoelomates The acoelomates are animals that lack a coelom. Acoelomates lack a body cavity, and instead the space between the body wall and the digestive tract is filled
Phylum Platyhelminthes Flatworms The Acoelomates The acoelomates are animals that lack a coelom. Acoelomates lack a body cavity, and instead the space between the body wall and the digestive tract is filled
Parasitenkunde. (Odocoileus virginianus ) Ultrastructure of Sarcocystis sp. from the Muscle of a White-Tailed Deer
 Ultrastructure of Sarcocystis sp. from the Muscle of a White-Tailed Deer") Z Parasitenkd (1982) 68 : 33-38 Zeitschrift for Parasitenkunde Parasitology Research 9 Springer-Verlag 1982 Ultrastructure of Sarcocystis sp. from the Muscle of a White-Tailed Deer (Odocoileus virginianus
Z Parasitenkd (1982) 68 : 33-38 Zeitschrift for Parasitenkunde Parasitology Research 9 Springer-Verlag 1982 Ultrastructure of Sarcocystis sp. from the Muscle of a White-Tailed Deer (Odocoileus virginianus
LABORATORY. The Protozoa. At the Bench
 LABORATORY Laboratory 8, Page 1 8 The Protozoa Introduction: The protozoa are unicellular animals that are classified on the basis of the organelles used for locomotion (flagella, pseudopodia, cilia or
LABORATORY Laboratory 8, Page 1 8 The Protozoa Introduction: The protozoa are unicellular animals that are classified on the basis of the organelles used for locomotion (flagella, pseudopodia, cilia or
A NEW SPECIES OF GENUS EIMERIA (APICOMPLEXA: EUCOCCIDIORIDA) FROM GOAT.
 FROM GOAT.") A NEW SPECIES OF GENUS EIMERIA (APICOMPLEXA: EUCOCCIDIORIDA) FROM GOAT. B.V. More 1, H.A.Kamble. 2 S.V. Nikam 3, 1 Department of Zoology, Ramkrishna Paramhansa Mahavidyalaya, Osmanabad. (M.S.) India. 2
A NEW SPECIES OF GENUS EIMERIA (APICOMPLEXA: EUCOCCIDIORIDA) FROM GOAT. B.V. More 1, H.A.Kamble. 2 S.V. Nikam 3, 1 Department of Zoology, Ramkrishna Paramhansa Mahavidyalaya, Osmanabad. (M.S.) India. 2
1) Most common, infectious, pathogenic animal (zoonotic) parasite of humans; estimated that 13% of humans are infected
 Most common, infectious, pathogenic animal (zoonotic) parasite of humans; estimated that 13% of humans are infected") XX Phylum Apicomplexa (Chapter 8) 2005 A. Characteristics 1. All are parasitic 2. APICAL COMPLEX a. Group of organelles used to invade host cells b. Visible only with electron microscopy Picture Slide
XX Phylum Apicomplexa (Chapter 8) 2005 A. Characteristics 1. All are parasitic 2. APICAL COMPLEX a. Group of organelles used to invade host cells b. Visible only with electron microscopy Picture Slide
Malaria. This sheet is from both sections recording and includes all slides and diagrams.
 Malaria This sheet is from both sections recording and includes all slides and diagrams. Malaria is caused by protozoa family called plasmodium (Genus) mainly affect blood system specially RBCs and each
Malaria This sheet is from both sections recording and includes all slides and diagrams. Malaria is caused by protozoa family called plasmodium (Genus) mainly affect blood system specially RBCs and each
Blood protozoan: Plasmodium
 Blood protozoan: Plasmodium The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans:four species are associated The Plasmodium spp. life cycle can be divided
Blood protozoan: Plasmodium The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans:four species are associated The Plasmodium spp. life cycle can be divided
Systemic Apicomplexans. Toxoplasma
 Systemic Apicomplexans Toxoplasma Protozoan Groups Historically, protozoa have been grouped by mode of motility. Flagellates Hemoflagellates Trypanosoma cruzi Leishmania infantum Mucoflagellates Tritrichomonas
Systemic Apicomplexans Toxoplasma Protozoan Groups Historically, protozoa have been grouped by mode of motility. Flagellates Hemoflagellates Trypanosoma cruzi Leishmania infantum Mucoflagellates Tritrichomonas
BLOOD PARASITES MORPHOTYPES OF ROCK LIZARDS OF ARMENIA
 PROCEEDINGS OF THE YEREVAN STATE UNIVERSITY C h e m i s t r y a n d B i o l o g y 2015, 2, p. 45 49 B i o l o g y BLOOD PARASITES MORPHOTYPES OF ROCK LIZARDS OF ARMENIA T. K. HARUTYUNYAN, F. D. DANIELYAN,
PROCEEDINGS OF THE YEREVAN STATE UNIVERSITY C h e m i s t r y a n d B i o l o g y 2015, 2, p. 45 49 B i o l o g y BLOOD PARASITES MORPHOTYPES OF ROCK LIZARDS OF ARMENIA T. K. HARUTYUNYAN, F. D. DANIELYAN,
EXOSTOSIS OF THE MANDIBLE OF THE CHICKEN
 EXOSTOSIS OF THE MANDIBLE OF THE CHICKEN COMPLICATING EDEMA OF THE WATTLES GEORGE MILTON SMITH, M.D.1 (AnutomioaZ Laboratory, Yale School of Medicine, New Haven, Connecticut) During the past year opportunity
EXOSTOSIS OF THE MANDIBLE OF THE CHICKEN COMPLICATING EDEMA OF THE WATTLES GEORGE MILTON SMITH, M.D.1 (AnutomioaZ Laboratory, Yale School of Medicine, New Haven, Connecticut) During the past year opportunity
Giardia and Apicomplexa. G. A. Lozano UNBC
 Giardia and Apicomplexa G. A. Lozano UNBC NINE Protozoan diseases/parasites Ciliphora, Ichthyophthirius, Ick Sarcomastigophora, Giardia, giardiasis Apicomplexa: Eimeria, Toxoplasma, Sarcocystis, Cryptosporidium.
Giardia and Apicomplexa G. A. Lozano UNBC NINE Protozoan diseases/parasites Ciliphora, Ichthyophthirius, Ick Sarcomastigophora, Giardia, giardiasis Apicomplexa: Eimeria, Toxoplasma, Sarcocystis, Cryptosporidium.
Proteocephalus filicollis (Rud. 1810) in the Netherlands
 in the Netherlands") Proteocephalus filicollis (Rud. 1810) in the Netherlands by J.J. Willemse AND A.L.M. Veltman Zoological Laboratory, University of Amsterdam INTRODUCTION in another glass dish containing about 50 specimens
Proteocephalus filicollis (Rud. 1810) in the Netherlands by J.J. Willemse AND A.L.M. Veltman Zoological Laboratory, University of Amsterdam INTRODUCTION in another glass dish containing about 50 specimens
Blood protozoan: Plasmodium
 Blood protozoan: Plasmodium Dr. Hala Al Daghistani The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans: four species are associated The Plasmodium spp.
Blood protozoan: Plasmodium Dr. Hala Al Daghistani The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans: four species are associated The Plasmodium spp.
Biology of Isospora spp. from Humans, Nonhuman Primates, and Domestic Animals
 CLINICAL MICROBIOLOGY REVIEWS, Jan. 1997, p. 19 34 Vol. 10, No. 1 0893-8512/97/$04.00 0 Copyright 1997, American Society for Microbiology Biology of Isospora spp. from Humans, Nonhuman Primates, and Domestic
CLINICAL MICROBIOLOGY REVIEWS, Jan. 1997, p. 19 34 Vol. 10, No. 1 0893-8512/97/$04.00 0 Copyright 1997, American Society for Microbiology Biology of Isospora spp. from Humans, Nonhuman Primates, and Domestic
Hepatic Coccidiosis of the Domestic Rabbit Oryctolagus cuniculus domesticus L. in Saudi Arabia
 World Journal of Zoology 3 (1): -35, 2008 ISSN 1817-98 IDOSI Publications, 2008 Hepatic Coccidiosis of the Domestic Rabbit Oryctolagus cuniculus domesticus L. in Saudi Arabia Ebtesam M. Al-Mathal Department
World Journal of Zoology 3 (1): -35, 2008 ISSN 1817-98 IDOSI Publications, 2008 Hepatic Coccidiosis of the Domestic Rabbit Oryctolagus cuniculus domesticus L. in Saudi Arabia Ebtesam M. Al-Mathal Department
Ultrastructure of Sarcocystis bertrami sarcocysts from a naturally infected donkey (Equus
 Ultrastructure of Sarcocystis bertrami sarcocysts from a naturally infected donkey (Equus asinus) from Egypt J. P. DUBEY 1,*, E. VAN WILPE 2, S. K. VERMA 1, M. HILALI 3, 1 U. S. Department of Agriculture,
Ultrastructure of Sarcocystis bertrami sarcocysts from a naturally infected donkey (Equus asinus) from Egypt J. P. DUBEY 1,*, E. VAN WILPE 2, S. K. VERMA 1, M. HILALI 3, 1 U. S. Department of Agriculture,
Vertebrates. Vertebrate Characteristics. 444 Chapter 14
 4 Vertebrates Key Concept All vertebrates have a backbone, which supports other specialized body structures and functions. What You Will Learn Vertebrates have an endoskeleton that provides support and
4 Vertebrates Key Concept All vertebrates have a backbone, which supports other specialized body structures and functions. What You Will Learn Vertebrates have an endoskeleton that provides support and
Frog Dissection Information Manuel
 Frog Dissection Information Manuel Anatomical Terms: Used to explain directions and orientation of a organism Directions or Positions: Anterior (cranial)- toward the head Posterior (caudal)- towards the
Frog Dissection Information Manuel Anatomical Terms: Used to explain directions and orientation of a organism Directions or Positions: Anterior (cranial)- toward the head Posterior (caudal)- towards the
EXOTIC CLINICAL PATHOLOGY
 Brittney Exarhos, LVT, RVT Toledo Zoo and Aquarium 2700 Broadway St. Toledo OH 43609 EXOTIC CLINICAL PATHOLOGY Veterinary technicians in a zoo setting often spend a lot of time in the lab. They must have
Brittney Exarhos, LVT, RVT Toledo Zoo and Aquarium 2700 Broadway St. Toledo OH 43609 EXOTIC CLINICAL PATHOLOGY Veterinary technicians in a zoo setting often spend a lot of time in the lab. They must have
"Comments on the nature and methods of collection of fish coccidia. " - Molnár, K. - Parasit. Hung. _
 Parasit. Hung 10. 1977. Comments on the Nature and Methods of Collection of Fish Coccidia Dr. Kálmán MOLNÁR Research Institute for Veterinary Science, Hungarian Academy of Sciences, Budapest "Comments
Parasit. Hung 10. 1977. Comments on the Nature and Methods of Collection of Fish Coccidia Dr. Kálmán MOLNÁR Research Institute for Veterinary Science, Hungarian Academy of Sciences, Budapest "Comments
AN ULTRASTRUCTURAL STUDY OF THE DEVELOPMENT OF BABESIA. E. F. BLOUIN and LYNN VAN RENSBURG, Veterinary Research Institute, Onderstepoort OliO
 OnderstepoortJ. vet. Res., 55, 93-100(1988) AN ULTRASTRUCTURAL STUDY OF THE DEVELOPMENT OF BABESIA OCCULTANS INTHESALIVARYGLANDSOF ADULT HYALOMMA MARGINATUM RUFIPES E. F. BLOUIN and LYNN VAN RENSBURG,
OnderstepoortJ. vet. Res., 55, 93-100(1988) AN ULTRASTRUCTURAL STUDY OF THE DEVELOPMENT OF BABESIA OCCULTANS INTHESALIVARYGLANDSOF ADULT HYALOMMA MARGINATUM RUFIPES E. F. BLOUIN and LYNN VAN RENSBURG,
Title. CitationJapanese Journal of Veterinary Research, 24(1-2): 37. Issue Date DOI. Doc URL. Type. File Information
: 37. Issue Date DOI. Doc URL. Type. File Information") Title DISTRIBUTION OF LYMPHATIC TISSUES IN DUCK CAECA Author(s)KITAMURA, Hirokazu; SUGIMURA, Makoto; HASHIMOTO, Yos CitationJapanese Journal of Veterinary Research, 24(1-2): 37 Issue Date 1976-05 DOI 10.14943/jjvr.24.1-2.37
Title DISTRIBUTION OF LYMPHATIC TISSUES IN DUCK CAECA Author(s)KITAMURA, Hirokazu; SUGIMURA, Makoto; HASHIMOTO, Yos CitationJapanese Journal of Veterinary Research, 24(1-2): 37 Issue Date 1976-05 DOI 10.14943/jjvr.24.1-2.37
A Study of Coccidiosis in Livestock in the Island of Dominica. Joshua Santelises. Study Abroad Texas A&M University. Dr.
 A Study of Coccidiosis in Livestock in the Island of Dominica Joshua Santelises Study Abroad 2012 Texas A&M University Dr. Thomas Lacher Dr. Jim Woolley Abstract The following experiment was done to investigate
A Study of Coccidiosis in Livestock in the Island of Dominica Joshua Santelises Study Abroad 2012 Texas A&M University Dr. Thomas Lacher Dr. Jim Woolley Abstract The following experiment was done to investigate
A NEW TYPE OF BRYOZOAN GIZZARD, WITH REMARKS ON THE GENUS BUSKIA.
 A NEW TYPE OF BRYOZOAN GIZZARD, WITH REMARKS ON THE GENUS BUSKIA. RAYMOND C. OSBURN AND RUTH M. VETH Department of Zoology and Entomology, Ohio State University A certain few of the Ctenostome Bryozoa
A NEW TYPE OF BRYOZOAN GIZZARD, WITH REMARKS ON THE GENUS BUSKIA. RAYMOND C. OSBURN AND RUTH M. VETH Department of Zoology and Entomology, Ohio State University A certain few of the Ctenostome Bryozoa
Ectoparasites Myobia musculi Radfordia affinis Radfordia ensifera
 Ectoparasites Fleas, ticks, and lice are uncommon in modern laboratory facilities, but may be seen on wild or feral rodents. Most ectoparasite infestations seen in rats and mice used for research are various
Ectoparasites Fleas, ticks, and lice are uncommon in modern laboratory facilities, but may be seen on wild or feral rodents. Most ectoparasite infestations seen in rats and mice used for research are various
Toxoplasmosis in Atlantic Bottle-Nosed Dolphins
 Journal of Wildlife Diseases, 26(3), 1990, pp. 377-382 Toxoplasmosis in Atlantic Bottle-Nosed Dolphins (Tursiops truncatus) W. Inskeep II, C. H. Gardiner, R. K. Harris, J. P. Dubey,2 and R. T. Goldston,3
Journal of Wildlife Diseases, 26(3), 1990, pp. 377-382 Toxoplasmosis in Atlantic Bottle-Nosed Dolphins (Tursiops truncatus) W. Inskeep II, C. H. Gardiner, R. K. Harris, J. P. Dubey,2 and R. T. Goldston,3
Heartworm Disease in Dogs
 Kingsbrook Animal Hospital 5322 New Design Road, Frederick, MD, 21703 Phone: (301) 631-6900 Website: KingsbrookVet.com What causes heartworm disease? Heartworm Disease in Dogs Heartworm disease or dirofilariasis
Kingsbrook Animal Hospital 5322 New Design Road, Frederick, MD, 21703 Phone: (301) 631-6900 Website: KingsbrookVet.com What causes heartworm disease? Heartworm Disease in Dogs Heartworm disease or dirofilariasis
Report of Water Mite Larvae in the Esophagus and Stomach Walls of Mountain Whitefish in British Columbia
 Proc. Helminthol. Soc. Wash. 50(2), 1983, pp. 325-329 Report of Water Mite Larvae in the Esophagus and Stomach Walls of Mountain Whitefish in British Columbia HILDA LEI CHING AND Lois PARKER Envirocon
Proc. Helminthol. Soc. Wash. 50(2), 1983, pp. 325-329 Report of Water Mite Larvae in the Esophagus and Stomach Walls of Mountain Whitefish in British Columbia HILDA LEI CHING AND Lois PARKER Envirocon
THE STRUCTURE OF ECHINOCOCCAL CYSTS AND HISTOPATHOLOGICAL CHANGES IN LIVER
 THE STRUCTURE OF ECHINOCOCCAL CYSTS AND HISTOPATHOLOGICAL CHANGES IN LIVER Michal Juszynski Helena Palenga, Danuta Cielecka PhD Department of General Biology and Parasitology Medical University of Warsaw
THE STRUCTURE OF ECHINOCOCCAL CYSTS AND HISTOPATHOLOGICAL CHANGES IN LIVER Michal Juszynski Helena Palenga, Danuta Cielecka PhD Department of General Biology and Parasitology Medical University of Warsaw
Cryptosporidium spp. Oocysts
 Sampling and Source Tracking of Cryptosporidium spp. Oocysts June 28, 2005 Kristen L. Jellison, Ph.D. Department of Civil & Environmental Engineering Lehigh University Bethlehem, Pennsylvania Ultimate
Sampling and Source Tracking of Cryptosporidium spp. Oocysts June 28, 2005 Kristen L. Jellison, Ph.D. Department of Civil & Environmental Engineering Lehigh University Bethlehem, Pennsylvania Ultimate
Malaria in the Mosquito Dr. Peter Billingsley
 Malaria in the Mosquito Senior Director Quality Systems and Entomology Research Sanaria Inc. Rockville MD. 1 Malaria: one of the world s foremost killers Every year 1 million children die of malaria 250
Malaria in the Mosquito Senior Director Quality Systems and Entomology Research Sanaria Inc. Rockville MD. 1 Malaria: one of the world s foremost killers Every year 1 million children die of malaria 250
VERTEBRATE READING. Fishes
 VERTEBRATE READING Fishes The first vertebrates to become a widespread, predominant life form on earth were fishes. Prior to this, only invertebrates, such as mollusks, worms and squid-like animals, would
VERTEBRATE READING Fishes The first vertebrates to become a widespread, predominant life form on earth were fishes. Prior to this, only invertebrates, such as mollusks, worms and squid-like animals, would
Exotic Hematology Lab Leigh-Ann Horne, LVT, CWR Wildlife Center of Virginia
 Exotic Hematology Lab Leigh-Ann Horne, LVT, CWR Wildlife Center of Virginia lhorne@wildlifecenter.org Anne Lynch, LVT Cedarcrest Animal Clinic amllvt9@gmail.com Introduction While the general set-up for
Exotic Hematology Lab Leigh-Ann Horne, LVT, CWR Wildlife Center of Virginia lhorne@wildlifecenter.org Anne Lynch, LVT Cedarcrest Animal Clinic amllvt9@gmail.com Introduction While the general set-up for
Schistosoma mansoni, S. japonicum, S. haematobium
 Schistosoma mansoni, S. japonicum, S. haematobium The Organisms More than 200 million people are infected worldwide with Schistosoma species. The adult worms are long and slender (males are 6 12 mm in
Schistosoma mansoni, S. japonicum, S. haematobium The Organisms More than 200 million people are infected worldwide with Schistosoma species. The adult worms are long and slender (males are 6 12 mm in
This is the smallest tapeworm that can affect human being but it s not really proper human tapeworm (the human is not the primary host).
.") Echinococcus Granulosus Small Tapeworm (1 cm), Cestode. This is the smallest tapeworm that can affect human being but it s not really proper human tapeworm (the human is not the primary host). The primary
Echinococcus Granulosus Small Tapeworm (1 cm), Cestode. This is the smallest tapeworm that can affect human being but it s not really proper human tapeworm (the human is not the primary host). The primary
Canine and Feline Distemper. Description. The following chart indicates the animals which are susceptible to infection by canine and feline distemp
 Canine and Feline Distemper Description Canine and feline distemper are diseases affecting many wild and domestic carnivo The following chart indicates the animals which are susceptible to infection by
Canine and Feline Distemper Description Canine and feline distemper are diseases affecting many wild and domestic carnivo The following chart indicates the animals which are susceptible to infection by
Morphological characterization of Cryptosporidium parvum life-cycle stages in an in vitro model system
 Morphological characterization of Cryptosporidium parvum life-cycle stages in an in vitro model system 13 H. BOROWSKI 1,R.C.A.THOMPSON 1 *, T. ARMSTRONG 1 and P. L. CLODE 2 1 WHO Collaborating Centre for
Morphological characterization of Cryptosporidium parvum life-cycle stages in an in vitro model system 13 H. BOROWSKI 1,R.C.A.THOMPSON 1 *, T. ARMSTRONG 1 and P. L. CLODE 2 1 WHO Collaborating Centre for
Efficacies of fenbendazole and albendazole in the treatment of commercial turkeys artificially infected with Ascaridia dissimilis
 Efficacies of fenbendazole and albendazole in the treatment of commercial turkeys artificially infected with Ascaridia dissimilis Jessica Perkins, Thomas Yazwinski, Chris Tucker Abstract The goal of this
Efficacies of fenbendazole and albendazole in the treatment of commercial turkeys artificially infected with Ascaridia dissimilis Jessica Perkins, Thomas Yazwinski, Chris Tucker Abstract The goal of this
SEMESTER ONE 2007 INFECTION and IMMUNITY GRADUATE ENTRY PROGRAMME PARASITOLOGY PRACTICAL 9 Dr TW Jones NEMATODES
 SEMESTER ONE 2007 INFECTION and IMMUNITY GRADUATE ENTRY PROGRAMME PARASITOLOGY PRACTICAL 9 Dr TW Jones NEMATODES Objectives After this class I expect you to be able to: 1. Describe and recognise the range
SEMESTER ONE 2007 INFECTION and IMMUNITY GRADUATE ENTRY PROGRAMME PARASITOLOGY PRACTICAL 9 Dr TW Jones NEMATODES Objectives After this class I expect you to be able to: 1. Describe and recognise the range
Article available at or
 Article available at http://www.parasite-journal.org or http://dx.doi.org/10.1051/parasite/1995023307 LIGHT AND ELECTRON MICROSCOPE STUDY OF A LANKESTERELLA PETITI N. SP., (APICOMPLEXA : LANKESTERELLIDAE)
Article available at http://www.parasite-journal.org or http://dx.doi.org/10.1051/parasite/1995023307 LIGHT AND ELECTRON MICROSCOPE STUDY OF A LANKESTERELLA PETITI N. SP., (APICOMPLEXA : LANKESTERELLIDAE)
Electron Microscopic Observations on Ciliated Epithelium of Tracheal Organ Cultures Infected with Bordetella bronchiseptica
 Microbiol. Immunol. Vol. 33 (2), 111-121, 1989 Electron Microscopic Observations on Ciliated Epithelium of Tracheal Organ Cultures Infected with Bordetella bronchiseptica Kachiko SEKIYA,*,1 Yutaka FUTAESAKU,2
Microbiol. Immunol. Vol. 33 (2), 111-121, 1989 Electron Microscopic Observations on Ciliated Epithelium of Tracheal Organ Cultures Infected with Bordetella bronchiseptica Kachiko SEKIYA,*,1 Yutaka FUTAESAKU,2