AMicroscaleHumanLiverPlatform that Supports the Hepatic Stages of Plasmodium falciparum and vivax
|
|
|
- Jonathan Bishop
- 6 years ago
- Views:
Transcription
1 Resource AMicroscaleHumanLiverPlatform that Supports the Hepatic Stages of Plasmodium falciparum and vivax Sandra March, 1,4 Shengyong Ng, 2 Soundarapandian Velmurugan, 5 Ani Galstian, 1,4 Jing Shan, 1 David J. Logan, 4 Anne E. Carpenter, 4 David Thomas, 4 B. Kim Lee Sim, 5 Maria M. Mota, 6 Stephen L. Hoffman, 5 and Sangeeta N. Bhatia 1,3,4,7, * 1 Health Sciences and Technology, Institute for Medical Engineering and Science 2 Department of Biological Engineering 3 Howard Hughes Medical Institute, Koch Institute, and Electrical Engineering and Computer Science Massachusetts Institute of Technology, Cambridge, MA 02139, USA 4 Broad Institute, Cambridge, MA 02142, USA 5 Sanaria Inc., Rockville, MD 20850, USA 6 Unidade de Malária, Instituto de Medicina Molecular, Universidade de Lisboa, Lisboa, Portugal 7 Department of Medicine, Brigham and Women s Hospital, Boston, MA 02115, USA *Correspondence: sbhatia@mit.edu SUMMARY The Plasmodium liver stage is an attractive target for the development of antimalarial drugs and vaccines, as it provides an opportunity to interrupt the life cycle of the parasite at a critical early stage. However, targeting the liver stage has been difficult. Undoubtedly, a major barrier has been the lack of robust, reliable, and reproducible in vitro liver-stage cultures. Here, we establish the liver stages for both Plasmodium falciparum and Plasmodium vivax in a microscale human liver platform composed of cryopreserved, micropatterned human primary hepatocytes surrounded by supportive stromal cells. Using this system, we have successfully recapitulated the full liver stage of P. falciparum, including the release of infected merozoites and infection of overlaid erythrocytes, as well as the establishment of small forms in late liver stages of P. vivax. Finally, we validate the potential of this platform as a tool for mediumthroughput antimalarial drug screening and vaccine development. INTRODUCTION Despite major advances in the prevention and treatment of malaria, this disease continues to be a major global health problem in human populations, with 250 million cases and nearly 1 million deaths every year (World Health Organization, 2010). Malaria is transmitted by Plasmodium sporozoites after they are injected by an infected mosquito. Uninucleate sporozoites travel to and invade liver hepatocytes, where they mature and multiply to form liver-stage schizonts. These schizonts eventually release pathogenic merozoites into the blood, where they invade erythrocytes and lead to the major clinical symptoms, signs, and pathology of the disease. Human malaria is primarily caused by four species of Plasmodium parasites. Plasmodium falciparum (P. falciparum) is the most virulent and causes the vast majority of deaths. Plasmodium vivax (P. vivax) is less deadly but highly disabling. Notably, some P. vivax sporozoites develop into dormant hypnozoites, which remain in the liver and serve as a P. vivax reservoir that gives rise to a chronic, relapsing infection and causes significant added clinical and financial burden (Price et al., 2007). Currently, there is a renewed interest and focus on global malaria eradication, and it is now widely recognized that existing tools are insufficient to meet this goal (Alonso et al., 2011a, 2011b). In particular, the clinical options that target the liver stages of the parasite life cycle are inadequate. There is only one licensed drug that eliminates hypnozoites, only a few drugs that target liver-stage parasites, and no licensed malaria vaccines. This problem has been exacerbated by the emergence of drug resistance and the inability to treat some populations with primaquine, the only currently approved drug with antihypnozoite activity (Wells et al., 2010). The Plasmodium liver stage is an attractive therapeutic target for the development of both antimalarial drugs and vaccines, as it provides an opportunity to interrupt the life cycle of the parasite at a critical early stage. Therefore, screening platforms that model the in vivo Plasmodium liver stage could be used to advance the pipeline for antimalarial drug development and to validate promising liver-stage vaccine candidates (Epstein et al., 2011; Plowe et al., 2009). Studies of rodent Plasmodium pathogens (P. berghei and P. yoelii) have provided important insights, due in part to the capacity to conduct both in vitro and in vivo assays (Hoffman et al., 1989; Rodrigues et al., 2008; Silvie et al., 2007). Nonetheless, there are essential differences between the rodent and human parasites, such as their antigenic variation and mechanisms of host cell invasion (Carlton et al., 2002; McCutchan et al., 1985). To date, our understanding of the liver stage of human malaria, mainly P. falciparum and P. vivax, is based in large part on the infection of human hepatoma cell lines (Chattopadhyay et al., 2010; Epstein et al., 2011; Hollingdale et al., 1983; 104 Cell Host & Microbe 14, , July 17, 2013 ª2013 Elsevier Inc.
2 Karnasuta et al., 1995; Sattabongkot et al., 2006). These cell lines, however, display abnormal proliferation, aberrant signaling, dysregulated gene expression, altered host responses to infection, and inadequate CYP450 and drug metabolism activity and thus do not accurately recapitulate human hepatocyte biology. Furthermore, in situ observation of pathogen development in liver cell lines is typically obscured after 6 days in culture, due to the continued proliferation of infected cells (Yokoo et al., 2004). Primary cultured human hepatocytes that exhibit more physiologic human liver functions have been studied previously, albeit less frequently than cell lines, and can support the development of the liver forms of P. falciparum and P. vivax (Mazier et al., 1984, 1985; Rodrigues et al., 2008; van Schaijk et al., 2008; Yalaoui et al., 2008). Nonetheless, in the 25 years since these findings were first published, primary hepatocyte systems are rarely employed and difficult to translate to screening platforms due to limited cell availability and challenges in maintaining their functional phenotype over extended periods of time in vitro (Bhatia et al., 1999). Two recent advances may help overcome the deficiencies of existing in vitro liver models. First, several groups have developed methods and novel culture platforms that support the maintenance of primary human hepatocytes (Bhatia et al., 1999; Guguen-Guillouzo and Guillouzo, 2010; LeCluyse et al., 2012). In particular, our group has developed a microliver platform, which leverages bioengineering techniques to organize primary human hepatocytes among supportive stromal cells (Khetani and Bhatia, 2008). Hepatocytes in these micropatterned cocultures (MPCCs) exhibit human-specific drug metabolism, retain drug responsiveness and hepatic energy metabolism, secrete of liver-specific proteins, polarize, and do not proliferate. Importantly, the hepatocytes in MPCCs maintain a functional phenotype for up to 4 6 weeks and are compatible with medium-throughput drug screening methods and automated data collection. This platform has been found to be more predictive than existing in vitro liver models for generating and identifying human drug metabolites and drug-induced liver toxicity and supports the persistent replication of hepatitis C virus (Jones et al., 2010; Ploss et al., 2010; Wang et al., 2010). Furthermore, the availability of large cryopreserved lots of human primary hepatocytes means that donor-dependent interexperimental variability can be minimized. Nonetheless, reproducible access to sporozoites is also critical to achieve a practical system. Cryopreservation of large batches of aseptic sporozoites has also now been established for both P. vivax (Chattopadhyay et al., 2010) and P. falciparum (Epstein et al., 2011) sporozoites. In this resource report, we demonstrate the feasibility of integrating cryopreserved human hepatocytes in MPCCs with cryopreserved P. falciparum and P. vivax sporozoites to form an in vitro platform that supports the liver stages of human malaria infection. This platform offers the potential for automation based on several factors, including preselection of cryopreserved human hepatocyte and sporozoite batches to standardize infection rate, a machine-learning algorithm that enables an automated imaging-based readout of immunofluorescent staining, and the capacity to generate a positive Z factor in response to drug exposure. The 96-well, medium-throughput format requires fewer reagents (drugs, sporozoites) than larger footprint in vitro assays, or than most in vivo assays, including humanized mouse models (Vaughan et al., 2012). Collectively, our data document the development and characterization of a highly-reproducible, medium-throughput microscale human liver platform that may aid in the development of safe and efficacious antimalarial drugs and liver-stage vaccines. RESULTS Functional Characterization of Micropatterned Cocultures from Cryopreserved Primary Human Hepatocytes In order to establish an MPCC in vitro culture of primary human hepatocytes (Figure 1A) suitable for Plasmodium infection, we screened cryopreserved hepatocyes from several individual patient donors to identify those that met the following criteria: (1) selective adhesion to collagen type I; (2) maintenance of a functional hepatocyte phenotype for up to 3 weeks as assessed by albumin expression, urea production, and CYP450 activity; and (3) expression of the previously identified Plasmodium host entry factor, CD81 (Silvie et al., 2003). We identified eight donor sources of cryopreserved human hepatocytes that were plateable on collagen I and displayed typical hepatocyte morphology, including the presence of bile canaliculi (Figure S1 available online). Seven of these sample sets demonstrated functional capacity in culture, as quantified by their production of albumin and urea, and also exhibited CYP450 activity for up to 3 weeks (Figure 1B). We next quantified the expression of the host entry factor CD81 on hepatocytes from each of the seven donors at the day of infection. Four of these samples (donors 2, 3, 7, and 8) expressed high levels of CD81 by immunofluorescence (IF) (Figure 1D). Infection of Primary Human Hepatocyte MPCCs with P. falciparum To test whether primary hepatocytes stabilized by culturing in MPCCs can be infected with P. falciparum, we exposed MPCCs to cryopreserved P. falciparum sporozoites (NF54) (Figures 2A and 2B). We confirmed productive infection in all seven donors by staining for HSP70 expression. However, a wide range of infection rates were observed between lots (Figure 1C, left panel). Specifically, at day 3 postinfection, the number of HSP70-positive, P. falciparum-infected hepatocytes was much higher in samples from donors 7 and 8 relative to the other tested lots. Hepatocytes from donors 7 and 8 were selected for further characterization of the model. We also examined the susceptibility of the same donor samples to productive infection by two different rodent species of Plasmodium. As shown in Figure 1C (middle panel), donors 7 and 8 also exhibit the highest levels of P. yoelii infection. However, this correlation was not observed in P. berghei (Figure 1C, right panel). Overall, infection with P. berghei was higher compared with P. falciparum and P. yoelii, with donor 1 showing slightly higher infection with P. berghei than the other donors. Interestingly, high CD81 expression was necessary, but not sufficient, to support robust infection by P. falciparum (Figures 1C and 1D). We next evaluated three different batches of cryopreserved P. falciparum sporozoites. As seen in Figure 1F, this experiment confirmed productive infection using sporozoites from three cryopreserved batches. However, infection efficiencies varied across batches. Cell Host & Microbe 14, , July 17, 2013 ª2013 Elsevier Inc. 105
 Morphology of primary human hepatocytes in")
3 Figure 1. Functional Characterization of Cryopreserved Human Hepatocytes in Micropatterned Cocultures and Cryopreserved Plasmodium falciparum Sporozoites (A) Morphology of primary human hepatocytes in micropatterned cocultures (left; hepatocytes, red and fibroblasts, green). Representative coculture of hepatocytes with (middle) and without (right) fibroblasts 18 days postseeding. (B) Albumin secretion, urea synthesis, and CYP450 activity in MPCCs of different donors. Red dashed lines indicate the average level observed in 6-day hepatocyte monocultures (SD = 0.9, 5.4, and 0.06 for the left, middle, and right panels, respectively). (C) P. falciparum, P. yoelii, and P. berghei infection across donors. (legend continued on next page) 106 Cell Host & Microbe 14, , July 17, 2013 ª2013 Elsevier Inc.
4 Based on higher infection levels, sporozoites from batch 2 were selected for further characterization of the model. Importantly, we observed higher infection rates when hepatocytes were cultured in the MPCC format as opposed to standard, unpatterned (randomly distributed) monocultures (Figure 1E, Hep Random). MPCCs remained susceptible to P. falciparum infection for many weeks after they were patterned; however, infection rates are optimal at day 2 after patterning (data not shown). Recapitulation of the Entire P. falciparum Liver Stage in MPCCs Having established that P. falciparum can infect MPCCs, we next sought to establish whether the entire liver stage could be reproduced in vitro. The viability of the cryopreserved P. falciparum sporozoites was evaluated by assessing their gliding motility (Figure 2C) and by their capacity to traverse cells using a cellwounding assay (Mota et al., 2001). On average, the addition of 37,000 motile cryopreserved P. falciparum sporozoites to 10,000 patterned hepatocytes resulted in 3% rhodaminepositive traversed cells (Figure 2D). To quantify the ability of the P. falciparum sporozoites to invade hepatocytes, P. falciparum-treated MPCCs were fixed 3 hr postinfection (Rénia et al., 1988). On average, 10% of hepatocytes contained intracellular cryopreserved P. falciparum sporozoites (Figure 2E). The human hepatocyte infection rate, based on the percentage of cells containing HSP70-expressing parasites at day 3 postinfection, was 0.2% (Figure 2J). Notably, at this same time point, fresh sporozoites achieved infection rates that were 7- to 13-fold higher (Figure 5A). Representative images of the HSP70-expressing parasites at day 3 and day 5 are shown in Figures 2F and 2G. The maturation of parasites derived from cryopreserved P. falciparum was assessed using three parameters: (1) immunostaining of the infected cultures with antibodies against two proteins first expressed during late liver stages, PfEBA-175 (P. falciparum erythrocyte-binding antigen, 175 kda) and PfMSP-1 (merozoite surface protein 1), which indicate full development of the respective schizonts inside the hepatocytes into merozoites; (2) the size of HSP70-expressing schizonts relative to previous reports of P. falciparum schizont size during liverstage development; and (3) the progression rate, calculated as the percentage of schizont-bearing cells at day 6 relative to day 3. At day 5 postinfection, cryopreserved sporozoites express both PfEBA-175 and PfMSP-1 (Figure 3C). The parasites we observed were similar in size to what has been reported in other in vitro settings (10 15 mm at day 5) (Mazier et al., 1985; van Schaijk et al., 2008) but smaller than those reported in vivo (Shortt et al., 1951; Shortt and Garnham, 1948; Vaughan et al., 2012) (Figures 2L and 3B). Importantly, we demonstrated that the rate at which parasites progressed to the schizont stage between day 3 and day 6 postinfection was higher in infected MPCCs (33%) relative to the commonly used hepatoma line HC04 (14%) (Figure 2K). Finally, to demonstrate that hepatic schizonts derived from cryopreserved P. falciparum sporozoites could achieve full maturity and release infective merozoites, red blood cells (RBCs) were added to the hepatocyte cultures at day 6 postinfection. After 10 days, Giemsa staining revealed infection of erythrocytes by P. falciparum merozoites (Figures 2H and 2I). Assessing Progression of Attenuated P. falciparum for Vaccine Applications The ability of the MPCCs to recapitulate the entire liver stage of P. falciparum in vitro highlights the potential to use this platform to study the biology of P. falciparum-infected hepatocytes. For example, this capability should enable the assessment of candidate pre-erythrocyte malarial vaccines that are based on liveattenuated parasites (Annoura et al., 2012; Epstein et al., 2011; Mueller et al., 2005; van Schaijk et al., 2008). To illustrate the potential use of MPCCs in vaccine development, we compared the infection capacity of a pre-erythrocytic malaria vaccine candidate comprised of cryopreserved, liveattenuated P. falciparum sporozoites to that of cryopreserved nonattenuated (wild-type) P. falciparum sporozoites. Entry of the attenuated parasites was evaluated relative to the function of nonattenuated parasites. As seen in Figure 3A, entry by both groups of parasites was similar. Late liver-stage development was evaluated at day 5 postinfection. Immunofluorescence staining with EBA-175 and MSP-1 antibodies detected mature schizonts only in MPCCs infected with the nonattenuated sporozoites, whereas MPCCs infected with the live-attenuated sporozoites were positive only for the early liver-stage antigen, LSA-1 (Figure 3C). As expected, schizonts that were established by the nonattenuated sporozoites were larger than the immature forms established by the attenuated sporozoites where the hepaticstage development is arrested. Figure 3B shows a scatter plot of the range of exoerythrocytic form (EEF) sizes generated by each group. Development of a Semiautomated, Medium-Throughput Platform for Antimalarial Applications We next explored the utility of the MPCCs as a potential antimalarial drug screening platform. The hepatocyte serves as both the site of antimalarial drug metabolism (or bioactivation) and the host for the parasite. Thus, phenotypic stability of the hepatocyte and a full drug metabolism repertoire has the potential to capture a full range of drug responses such as efficacy, drug-drug interaction, and toxicity (Ploss et al., 2010). MPCCs were established in a 96-well format, infected with P. falciparum, and treated with the canonical malaria drug, primaquine. The impact of the drug was evaluated based on its ability to reduce parasite infection relative to control cultures in a multiday dose. As seen in Figures 4A and 4B, the half maximal inhibitory concentration (IC 50 ) for primaquine was approximately 1 mm for P. falciparum during 3 days in culture. The IC 50 for P. falciparum in the MPCC was lower compared to the IC 50 (D) Representative CD81 immunofluorescence staining at day 4 postseeding (left). Heatmap indicates relative CD81 expression per donor as measured by IF (right; n.d., not detected). (E) P. falciparum infection in hepatocyte monocultures, micropattern (MP), or randomly distributed (Random) relative to infection in MPCCs. 10,000 hepatocytes were plated in each case. (F) Levels of infection by three sporozoite batches in a single hepatocyte donor. Error bars represent SD. See also Figure S1. Cell Host & Microbe 14, , July 17, 2013 ª2013 Elsevier Inc. 107
 Typical morphology of primary human hepatocytes in MPCCs (hepatocytes, red; fibroblasts, green). (C) Representative image of P. falciparum sporozoites gliding.")
5 Figure 2. Liver-Stage Recapitulation in Primary Human Hepatocyte MPCCs (A) Schematic of P. falciparum infection assay. (B) Typical morphology of primary human hepatocytes in MPCCs (hepatocytes, red; fibroblasts, green). (C) Representative image of P. falciparum sporozoites gliding. CSP immunostaining was used to visualize trails. Quantification based on the average fraction of sporozoites that perform at least one circle. (legend continued on next page) 108 Cell Host & Microbe 14, , July 17, 2013 ª2013 Elsevier Inc.
 Number of infected hepatocytes observed after wild-type (nonattenuated) and attenuated cryopreserved")
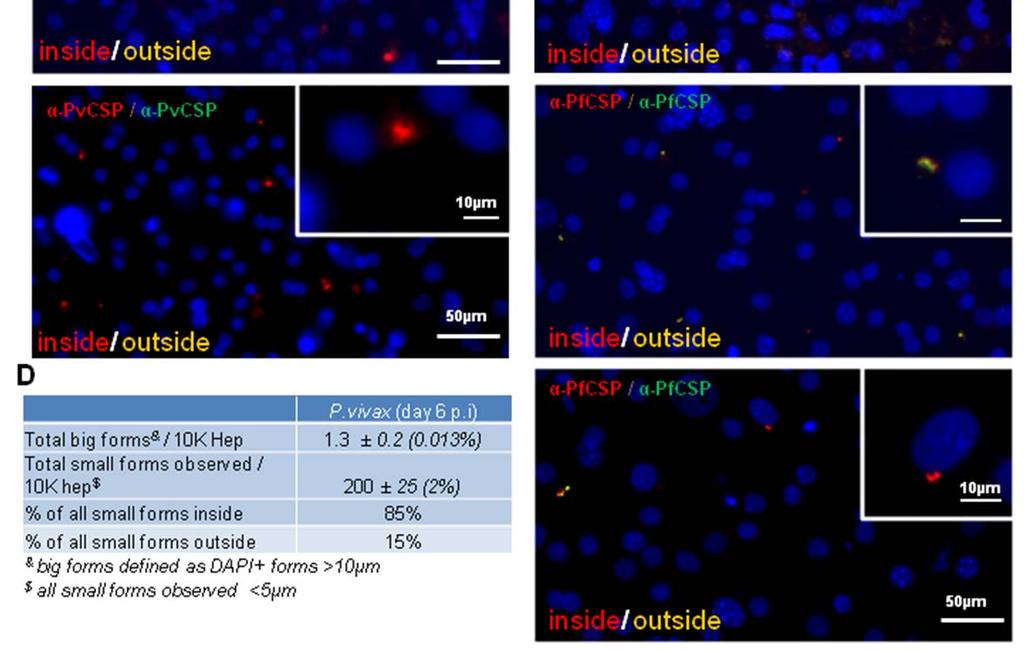
6 Figure 3. Comparison of Live-Attenuated versus Wild-Type Parasites for Candidate Vaccine Evaluation (A) Number of infected hepatocytes observed after wild-type (nonattenuated) and attenuated cryopreserved sporozoite infection. (B) Size distribution of wild-type and attenuated parasites in MPCCs after 5 days of culture. (C) Representative images of parasites at day 5 postinfection. Wild-type parasites are identified by anti-msp-1 and anti-eba-175 staining. Attenuated parasites are identified by anti-lsa-1 staining. Nuclei are visualized with DAPI (blue). Scale bar = 10 mm. Error bars represent SEM. obtained using unpatterned, monocultures of the hepatoma cell line HC04 (Figures 4A and 4B). Primaquine is known to require bioactivation in the liver in order to produce active daughter metabolites that mediate drug action (Pybus et al., 2012). Thus, the clearance of the parent compound can be used as a proxy for the level of bioactivation in culture. We monitored depletion of primaquine in MPCCs, patterned monocultures of primary hepatocytes only (Hep MP), and the HC04 cell line over 2 days by high-performance liquid chromatography and found that MPCCs most efficiently cleared primaquine over this time frame (Figure 4C). We next compared the expression level of 83 human-specific drug metabolism genes in order to assess whether the observed macroscopic differences in parent compound depletion correlates with the drug metabolism reportoire of these model systems. In general, the bulk of hepatocyte drug metabolism genes were expressed at higher levels in the MPCCs (Figure 4E). Of the three major enzymes involved with primaquine metabolism, two of the three were expressed at higher levels in MPCC than the other systems. In particular, monoamine oxidase A (MAO-A), which is thought to account for over 75% of primaquine metabolism, was expressed at a level 4-fold higher than hepatoma cell lines, suggesting that the platform may offer a more predictive system with which to assay candidate drug performance, including studies of mechanisms of action. Next, to further adapt the microliver platform for mediumthroughput screens, we characterized several critical parameters. As seen in Figures 5A and 5B, we found that day-to-day variability of hepatocyte infection using the same batch of cryopreserved sporozoites could be minimized (coefficient of variation [CV] = 10%) when compared to the range of infection rates obtained using three independent batches of fresh sporozoites (CV = 42%). Next, a positive Z factor (Z 0 > 0), which indicates confidence in separating two normally distributed populations, was obtained when primaquine-treated wells (10 mm) were compared to control cultures (Figure 5C). In addition, the reproducibility of a phenotypic assay was assessed by establishing consistent dose-dependent inhibitory effects of two different drugs at three different concentrations in two independent experiments (atovaquone and primaquine) (Figure 5D). Furthermore, the magnitude of the inhibitory impact of primaquine was consistent across two independent human hepatocyte donors (Figure S2). Finally, we established and validated an automated image analysis readout using this platform, following the infection of MPCCs with two different doses of sporozoites. As shown in Figures 5E and S3, the number of parasites detected by automated image acquisition and analysis closely matches the counts obtained using conventional manual fluorescence microscopy. Infection of MPCCs with Plasmodium vivax Plasmodium vivax differs from P. falciparum in several important ways. A key feature of P. vivax that underlies its persistence in the population is that the liver acts as a reservoir for dormant hypnozoites or small forms (Cogswell, 1992; Krotoski et al., 1982). These hypnozoites can reactivate after weeks, months, or even years, depending on the strain of P. vivax (Dao et al., 2007; Durante Mangoni et al., 2003; Garnham et al., 1975). However, due to a lack of model systems available to investigate this elusive organism, the biology of the dormant form of P. vivax is underexplored. Based on our ability to establish the liver stage of P. falciparum in MPCCs and maintain the hepatocyte phenotype for 4 6 weeks and the observation that some strains of P. vivax can reactivate over a similar timescale, we next explored the (D) Cell traversal ability of P. falciparum sporozoites as visualized by dextran-positive staining of primary human hepatocytes. (E) Representative double immunofluorescence stain (anti-pfcsp, both before and after cell permeablization) of P. falciparum-infected MPCCs. Extracellular and intracellular sporozoites are labeled with yellow and red, respectively. Nuclei are visible with blue DAPI stain. (F and G) Representative images of P. falciparum in human primary hepatocytes at days 3 (F) and 5 (G) postinfection. Parasites are identified by anti-pfhsp70 staining (red). (H and I) Infection of human RBCs by merozoites released from infected liver-stage culture. Representative images of Giemsa-stained RBCs in the ring stage (H) and the trophozoite stage (I). (J) Infection rates using MPCC primary human hepatocytes or HC04 hepatoma cells, calculated based on the plated number of sporozoites or hepatocytes. (K) Progression rate from day 3 to day 6 in MPCC and HC04 calculated as infection rate of day 6 O infection rate of day (based on hepatocytes and sporozoites). (L) Schizont size distribution at days 3, 4.5, 6, and 7. Scale bar = 5 mm (F I), 10 mm (C and E), or 100 mm (D). Error bars represent SEM. Cell Host & Microbe 14, , July 17, 2013 ª2013 Elsevier Inc. 109
 IC 50 of primaquine in MPCCs versus HC04 (p = 0.0002 by one-way ANOVA; ***p < 0.001 by Tukey s multiple comparison test).")
7 Figure 4. Utility of Medium-Throughput Human Hepatocyte Platform to Identify Lead Compounds (A) Primaquine treatment of MPCCs or HC04 infected with fresh or cryopreserved sporozoites. (B) IC 50 of primaquine in MPCCs versus HC04 (p = by one-way ANOVA; ***p < by Tukey s multiple comparison test). (C) Primaquine metabolism by HC04, MPCCs, and patterned monocultures of primary human hepatocytes (Hep MP) quantified by liquid chromatographytandem mass spectrometry (LC-MS/MS). (D) Relative expression of three putative metabolism genes implicated in primaquine metabolism. (E) Heatmap displays of LMA-Luminex analysis for 83 human-specific drug metabolism genes. Columns represent triplicate loadings of RNA extracted from HEPG2, HC04, MPCC, and Hep MP. Gene expression relative to average of control gene transferrin, and heatmaps are row normalized. Error bars represent SEM. See also Figure S Cell Host & Microbe 14, , July 17, 2013 ª2013 Elsevier Inc.
 Interexperimental variability measured by the coefficient of variation (CV) and infection rate using fresh and cryopreserved sporozoites from three different")
8 Figure 5. Adapting the Format to Drug Screening (A) Interexperimental variability measured by the coefficient of variation (CV) and infection rate using fresh and cryopreserved sporozoites from three different batches. (B) Interexperimental variability measured by the coefficient of variation (CV) and infection rate using cryopreserved sporozoites from the same batch. (C) Heatmap indicating levels of infection (green, highest EEF numbers; red, lowest EEF numbers) observed in seven representative control or primaquine-treated wells. Comparison yields positive Z factor. (D) P. falciparum infection with primaquine or atovaquone in two independent experiments performed on different days. (E) P. falciparum infection in MPCCs following two doses of fresh sporozoites determined by manual counts (indicated by M) or image analysis automation (indicated by A). Error bars represent SEM. See also Figure S3. feasibility of establishing the liver stages of P. vivax in MPCCs over time. As for the P. falciparum experiments, the viability of the cryopreserved P. vivax sporozoites was evaluated by assessing their gliding motility prior to each experiment (Figure 6A). We explored several strains of P. vivax including Chesson, a strain known to efficiently form dormant forms and reactivate at shorter timescales (Hollingdale et al., 1986; Krotoski et al., 1986). Cultures were infected and then fixed at various time points and stained for circumsporozoite protein (CSP). Using immunofluorescence microscopy, we analyzed the size distribution and localization of the P. vivax forms over a 3-week period. We readily observed P. vivax liver forms in MPCC, including mature, liver-stage schizonts larger than 20 mm found at day 6 postinfection (Figures 6C, 6B, S4A, and S4B). In addition, a population of smaller P. vivax CSP (PvCSP)-positive forms (<5 mm) was detected within MPCCs at all time points, particularly identifiable at 6 and 21 days postinfection (Figures 6A, 6B, and S4A S4C). The infection efficiency, based on the percentage of hepatocytes containing large and small PvCSPpositive forms at day 6 postinfection, was 0.013% and 2%, respectively (Figure S4D). Similar numbers of small forms were observed up to day 21 (data not shown). Importantly, 85% of the small forms observed were intracellular, based on immunostaining performed before and after permeabilization (Figure S4C). In contrast, cultures infected with P. falciparum contained very few small forms after 15 days in culture, and those were predominantly extracellular (Figure S4E). These P. vivax small forms may represent the dormant hypnozoite stage of the parasite life cycle, which is responsible for clinical relapses in P. vivax malaria patients; however, further characterization will be required to substantiate this hypothesis. Both small and large forms were observed when other strains of fresh and frozen P. vivax sporozoites were examined (Figure S4A and data not shown). Finally, we demonstrated that the MPCC system can support maturation of hepatic P. vivax schizonts, based on the detection of the late-stage antigen PvMSP-1 at day 12 postinfection (Figure S4B). DISCUSSION In this report, we describe an in vitro cell-based platform that recapitulates the human liver stage of P. falciparum and P. vivax infection. Although some attempts to infect cryopreserved human primary hepatocytes have been described in the past (Meis et al., 1985; Silvie et al., 2004), this source of hepatocytes has not been routinely adopted by the field to date. The advantages of screening individual donor-derived, cryopreserved hepatocytes is paralleled by the successful production of purified, vialed, cryopreserved sporozoites (Chattopadhyay et al., 2010; Epstein et al., 2011; Hoffman et al., 2010). These cryopreserved resources overcome the donor-to-donor variability seen in primary cultured human hepatocyes as well as infectivity rate variability introduced by different batches of mosquitoes or sporozoites. In addition, the use of cryopreserved components in this platform allows for a reliable source of reagents for use in longitudinal experiments, including screening and subsequent validation. Furthermore, since cell culture of patterned hepatocytes using the MPCC platform can maintain individual patient samples for 4 and 6 weeks (Khetani and Bhatia, 2008), it is feasible to perform long-term monitoring of both liver- and blood-stage Plasmodium infections, analyze genetic changes acquired inside Anopheles mosquitoes, assess the safety of attenuated sporozoite vaccine candidates, and characterize P. vivax hypnozoites in vitro using this system. Notably, development of vaccines and drugs against this stage of P. vivax has been identified as a critically important goal for research to eradicate malaria (Alonso et al., 2011a, 2011b). Cell Host & Microbe 14, , July 17, 2013 ª2013 Elsevier Inc. 111
 Size distribution of P. vivax parasites in MPCCs over time. Red, all observed parasites bigger than 5 mm; black, 20 representative forms smaller than 5 mm. See also Figure S4.")
9 Figure 6. Infection with Plasmodium vivax (A) Representative image of PvCSP-stained P. vivax (Chesson) sporozoites gliding. (B) Representative images of PvCSP-positive parasites over time. Scale bar = 10 mm. (C) Size distribution of P. vivax parasites in MPCCs over time. Red, all observed parasites bigger than 5 mm; black, 20 representative forms smaller than 5 mm. See also Figure S4. Recent publications have highlighted that existing candidate vaccines continue to underperform in clinical trials, and significant blood breakthrough of presumably attenuated parasites formulations has been observed (Annoura et al., 2012). In our experience with disease modeling of drug-induced liver injury and hepatitis C infection, establishing platforms that better reflect host biology is an important first step to determining where existing model systems were lacking (e.g., P450 activity and interferon signaling, respectively). In this case, we have already observed three advantages over in vitro hepatoma cultures: (1) sporozoites appear to progress through the parasite life cycle more efficiently in MPCCs relative to infected hepatoma cells, offering the potential to improve studies of drugs and sporozoite attenuation strategies that act in the second half of the liver life cycle; (2) MPCCs are able to predict differences in infection rates of sporozoites in vivo that result from cryopreservation, likely reflecting the presence of critical host factors that are altered in hepatoma cells (Albuquerque et al., 2009; Epiphanio et al., 2008; Prudêncio et al., 2008; Rodrigues et al., 2008; Silvie et al., 2003); and (3) MPCCs fabricated from different human hepatocyte donors enable direct comparison of host factors that impact entry of different sporozoite species and strains (e.g., CD81 in P. yoeli, P. berghei, and P. falciparum), whereas hepatoma cells are typically limited to one or very few donor genotypes. We demonstrated that maintenance of hepatocyte function and expression of the host entry factor CD81 (Silvie et al., 2003) were necessary, but not sufficient, to obtain adequate levels of infection by P. falciparum. This finding suggests the existence of molecular differences among donors that determine their permissiveness to Plasmodium infection. Furthermore, most antimalarial drug development leads identified in RBC-based high-throughput screens do not require metabolic activation. Thus, in such cases, screening via the MPCC format might yield the same, higher, or lower IC 50 predictions, should the candidate compound be cleared rapidly or bioactivated via metabolic pathways (Gamo et al., 2010; Guiguemde et al., 2010). The infection rates reported in this study using cryopreserved sporozoites are comparable or higher than those previously documented with fresh sporozoites, and they are an additional 10-fold higher when fresh sporozoites are used in this platform (Mazier et al., 1984, 1985; Sattabongkot et al., 2006; van Schaijk et al., 2008). The infection rates of hepatocytes in MPCCs are similar in comparison to the infection rates recently reported in HC04 cells, which range from 0.4% to 0.06% (Epstein et al., 2011; Sattabongkot et al., 2006). However, in our system, the progression rate from one stage of the life cycle to the other was much higher than in HC04, offering the potential for studying later stages of the liver life cycle more efficiently. Nonetheless, our MPCC infection rates remain low relative to those recorded in in vivo settings (Shortt et al., 1951; Shortt and Garnham, 1948). Experiments done in mice with P. yoelii (Conteh et al., 2010) and nonhuman primates with P. knowlesi (Jiang et al., 2009) have demonstrated that intravenous inoculation of only a few noncryopreserved sporozoites (10 sporozoites [SPZ]) can lead to a productive malaria infection that results in detectable parasitemia in the blood stage. Varying hypotheses have been put forward to 112 Cell Host & Microbe 14, , July 17, 2013 ª2013 Elsevier Inc.
10 explain the discrepancy between model systems. For example, our in vitro systems do not provide the host hepatocytes with potentially necessary, physiologically relevant cellular components such as Kupffer cells or sinusoidal endothelial cells. Further, the MPCC platform conformational cues may be important for EEF growth (e.g., two-dimensional versus three-dimensional). These hypotheses will be explored further. In conclusion, the ability to support the liver stages of P. falciparum and P. vivax parasites in a medium-throughput format offers promise to improve our fundamental understanding of the liver stages of human malaria as well as accelerate the development of drugs and vaccines to aid in its eradication. EXPERIMENTAL PROCEDURES Micropatterned Cocultures 12 mm coverslips that were placed into tissue culture polystyrene 24-well plates or glass-bottomed 96-well plates were coated homogenously with rat tail type I collagen (50 mg/ml) and subjected to soft lithographic techniques (Ploss et al., 2010) to pattern the collagen into microdomains (islands of 500 mm) that mediate selective hepatocyte adhesion. To create MPCCs, cryopreserved primary human hepatocytes were pelleted by centrifugation at g for 6 min at 4 C, assessed for viability using trypan blue exclusion (typically 70% 90%), and then seeded on collagenmicropatterned plates. The cells were washed with medium 2 3 hr later and replaced with human hepatocyte culture medium. 3T3-J2 murine embryonic fibroblasts were seeded (40,000 cells per well of a 24-well plate and 10,000 cells per well of a 96-well plate) in fibroblast medium 3 days later. Fibroblast medium was replaced with human hepatocyte medium 24 hr after fibroblast seeding and subsequently replaced daily (Khetani and Bhatia, 2008). P. falciparum and P. vivax Sporozoites Mosquitoes were fed on Pf- and Pv-infected blood as previously described (Chattopadhyay et al., 2010; Epstein et al., 2011). Briefly, P. falciparum and P. vivax sporozoites were extracted from infected mosquitoes by dissection of their salivary glands and passing the glands back and forth through a 26G needle fitted to a 1 ml syringe. Following extraction, sporozoites were purified from mosquito salivary gland material contamination and cryopreserved in liquid nitrogen vapor phase (LNVP) (Epstein et al., 2011; Hoffman et al., 2010). Live-attenuated P. falciparum sporozoites were attenuated by exposure of PfSPZ-infected mosquitoes to 150 Gy (Epstein et al., 2011; Hoffman et al., 2010). Infection of MPCCs with P. falciparum and P. vivax Typically, infection of MPCCs is conducted 2 days after hepatocyte seeding, but it can also be initiated after longer culture periods. Cryovials containing P. falciparum or P. vivax were warmed for 30 s at 37 C, 200 ml of human hepatocyte medium was added, and the cryovials were centrifuged for 2 min at 14,000 3 g. Next, 200 ml of the supernatant was aspirated, and the sporozoite pellet was resuspended and diluted accordingly. Each well was infected with a ratio of 3:1 (infective sporozoites:hepatocytes). After incubation at 37 C and 5% CO 2 for 3 hr, the wells were washed once, and fresh medium was added. Medium was replaced daily. Samples were fixed on days 3 and 5 postinfection with P. falciparum, and days 3, 6, 12, and 21 postinfection with P. vivax. Cell Wounding and Membrane Repair Assay Sporozoite migration through cells can be quantified by the detection of sporozoite-wounded hepatocytes using a cell-impermeant fluorescent tracer macromolecule as previously described (Mota et al., 2001). Briefly, MPCCs were infected with P. falciparum in the presence of 1 mg/ml of 10,000 Da tetramethylrhodamine-dextran (lysine-fixable) (Sigma). At 3 hr postinfection, MPCCs were washed thrice with PBS, fixed with 1% paraformaldehyde at room temperature for 20 min, and mounted on glass slides. Migration of sporozoites through cells is quantified by the number of dextran-positive hepatocytes per island. Double-Staining Assay for Sporozoite Entry At 3 hr postinfection, primary human hepatocytes or MPCCs were fixed and stained using a double-staining protocol as previously described (Rénia et al., 1988). Briefly, to label extracellular sporozoites, the samples were fixed with 4% paraformaldehyde for 10 min at room temperature, blocked with 2% BSA in PBS, incubated with a primary mouse anti-pfcsp (1:100, Sanaria) (Nardin et al., 1982), washed thrice in PBS, and incubated with a secondary goat anti-mouse Alexa Fluor 488 conjugate. This was followed by permeabilization with 20 C methanol for 10 min at 4 C, incubation with the same primary mouse anti-pfcsp, washing thrice with PBS, and incubation with a secondary goat anti-mouse Alexa Fluor 594 conjugate. This second step labels both intracellular and extracellular sporozoites. In case of MPCCs, the samples were counterstained with Hoechst and mounted on glass slides as described above. The number of invaded sporozoites (only green) in primary human hepatocytes was counted using Acumen Explorer. Gliding Assay Motility of cryopreserved sporozoites was determined in each batch to define the number of infective sporozoites. Sporozoite gliding was evaluated with 20,000 sporozoites for 40 min in complete Dulbecco s modified Eagle s medium (DMEM), at 37 C on glass coverslips covered with anti-csp monoclonal antibody (clone 2A10 for P. falciparum and 210 clone NSV3 for P. vivax). Sporozoites were subsequently fixed in 4% paraformaldehyde (PFA) for 10 min and stained with anti-csp. The percentage of sporozoites associated with CSP trails was visualized by fluorescence microscopy. Quantification was performed by counting the average percentage of sporozoites that perform at least one circle. Primaquine Treatment of P. falciparum EEFs in MPCCs Infected MPCCs were incubated with media containing primaquine diphosphate (Sigma) ranging from mm. Fresh primaquine-containing medium was added daily until the samples were fixed at days 3 and 5 postinfection. RNA Isolation and LMA-Luminex Analysis Total RNA from three wells per condition was purified using TRIzol (Invitrogen) and RNeasy Mini kit (QIAGEN) and pooled for analysis. LMA-Luminex procedures and probes are previously described (Chen et al., 2011). Briefly, data for triplicate loadings, expressed in mean fluorescent intensity of at least 100 beads per sample, were scaled to the human transferrin gene and row normalized for heatmap representation using GenePattern open-source software (Broad Institute). Image Automation Images of 96-well plates were acquired using high-content screening microscopes (ImageXpress Micro XL, Molecular Device) and then analyzed by Cell- Profiler and CellProfiler Analyst (Broad Institute) (Carpenter et al., 2006; Jones et al., 2008). Parasites were visualized through immunofluorescent staining of the HSP70 protein and can be distinguished from imaging artifacts by their proximity to a hepatocyte nucleus (within 30 pixels) and lack of autofluorescence (no signal in unlabeled channels). We developed an automated image analysis pipeline to identify every infection in every image and measure hundreds of features (e.g., shape, area, texture) of each parasite. These features are then used to train machine learning algorithms to identify and count the number of parasites in each image using CellProfiler Analyst. SUPPLEMENTAL INFORMATION Supplemental Information includes Supplemental Experimental Procedures and four figures and can be found with this article online at /j.chom ACKNOWLEDGMENTS We thank the Malaria Research and Reference Reagent Resource Center for access to the polyclonal rabbit antiserum against purified recombinant Cell Host & Microbe 14, , July 17, 2013 ª2013 Elsevier Inc. 113
11 P. vivax (Sal-1) MSP-1 (MRA-16, deposited by J.H. Adams); NYU for the monoclonal antibody 2A10; NIAID, NIH for R217; and Dr. R. Wirtz (Centers for Disease Control and Prevention) and Dr. F. Zavala (Johns Hopkins University) for PvCSP and HSP70 monoclonal antibodies, respectively. We are grateful to the Sanaria Manufacturing Team for the production of PfSPZ and PvSPZ. We thank R. Schwartz for confocal microscopy help; S. Suresh and M. Diez for aid in establishing RBC cocultures; A. Rodriguez (NYU), D. Wirth, and E. Lund (HSPH) for providing mosquitoes infected with P. yoelii and P. berghei; J. Prachumsri (Mahidol Vivax Research Center) and J. Adams (University of South Florida) for providing fresh P. vivax; T. Golub (Broad Institute) for advice with the Luminex-based characterization of drug-metabolism transcripts; H. Green (Harvard University) for providing J2-3T3 fibroblasts; and H. Fleming for manuscript editing. PvSPZ production was supported by a grant from Medicines for Malaria Venture, and PfSPZ production was supported by a Phase II NIAID, NIH Small Business Innovative Research Grant (2R44A ) awarded to S.L.H. Software improvements were supported by an NIH grant (R01GM089652) to A.E.C. This work was supported by the Bill & Melinda Gates Foundation (51066). S.N. is supported by an Agency for Science, Technology and Research (A * STAR, Singapore) NSS. S.N.B. is an HHMI Investigator. The authors wish to dedicate this paper to the memory of Officer Sean Collier for his caring service to the MIT community and for his sacrifice. Received: January 28, 2012 Revised: January 15, 2013 Accepted: June 5, 2013 Published: July 17, 2013 REFERENCES Albuquerque, S.S., Carret, C., Grosso, A.R., Tarun, A.S., Peng, X., Kappe, S.H., Prudêncio, M., and Mota, M.M. (2009). Host cell transcriptional profiling during malaria liver stage infection reveals a coordinated and sequential set of biological events. BMC Genomics 10, 270. Alonso, P.L., Ballou, R., Brown, G., Chitnis, C., Loucq, C., Moorthy, V., Saul, A., and Wirth, D.; malera Consultative Group on Vaccines. (2011a). A research agenda for malaria eradication: vaccines. PLoS Med. 8, e Alonso, P.L., Djimde, A., Kremsner, P., Magill, A., Milman, J., Najera, J., Plowe, C.V., Rabinovich, R., Wells, T., and Yeung, S.; malera Consultative Group on Drugs. (2011b). A research agenda for malaria eradication: drugs. PLoS Med. 8, e Annoura, T., Ploemen, I.H., van Schaijk, B.C., Sajid, M., Vos, M.W., van Gemert, G.J., Chevalley-Maurel, S., Franke-Fayard, B.M., Hermsen, C.C., Gego, A., et al. (2012). Assessing the adequacy of attenuation of genetically modified malaria parasite vaccine candidates. Vaccine 30, Bhatia, S.N., Balis, U.J., Yarmush, M.L., and Toner, M. (1999). Effect of cell-cell interactions in preservation of cellular phenotype: cocultivation of hepatocytes and nonparenchymal cells. FASEB J. 13, Carlton, J.M., Angiuoli, S.V., Suh, B.B., Kooij, T.W., Pertea, M., Silva, J.C., Ermolaeva, M.D., Allen, J.E., Selengut, J.D., Koo, H.L., et al. (2002). Genome sequence and comparative analysis of the model rodent malaria parasite Plasmodium yoelii yoelii. Nature 419, Carpenter, A.E., Jones, T.R., Lamprecht, M.R., Clarke, C., Kang, I.H., Friman, O., Guertin, D.A., Chang, J.H., Lindquist, R.A., Moffat, J., et al. (2006). CellProfiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biol. 7, R100. Chattopadhyay, R., Velmurugan, S., Chakiath, C., Andrews Donkor, L., Milhous, W., Barnwell, J.W., Collins, W.E., and Hoffman, S.L. (2010). Establishment of an in vitro assay for assessing the effects of drugs on the liver stages of Plasmodium vivax malaria. PLoS ONE 5, e Chen, A.A., Thomas, D.K., Ong, L.L., Schwartz, R.E., Golub, T.R., and Bhatia, S.N. (2011). Humanized mice with ectopic artificial liver tissues. Proc. Natl. Acad. Sci. USA 108, Cogswell, F.B. (1992). The hypnozoite and relapse in primate malaria. Clin. Microbiol. Rev. 5, Conteh, S., Chattopadhyay, R., Anderson, C., and Hoffman, S.L. (2010). Plasmodium yoelii-infected A. stephensi inefficiently transmit malaria compared to intravenous route. PLoS ONE 5, e8947. Dao, N.V., Cuong, B.T., Ngoa, N.D., Thuy, T.T., The, N.D., Duy, D.N., Dai, B., Thanh, N.X., Chavchich, M., Rieckmann, K.H., and Edstein, M.D. (2007). Vivax malaria: preliminary observations following a shorter course of treatment with artesunate plus primaquine. Trans. R. Soc. Trop. Med. Hyg. 101, Durante Mangoni, E., Severini, C., Menegon, M., Romi, R., Ruggiero, G., and Majori, G. (2003). Case report: An unusual late relapse of Plasmodium vivax malaria. Am. J. Trop. Med. Hyg. 68, Epiphanio, S., Mikolajczak, S.A., Gonçalves, L.A., Pamplona, A., Portugal, S., Albuquerque, S., Goldberg, M., Rebelo, S., Anderson, D.G., Akinc, A., et al. (2008). Heme oxygenase-1 is an anti-inflammatory host factor that promotes murine plasmodium liver infection. Cell Host Microbe 3, Epstein, J.E., Tewari, K., Lyke, K.E., Sim, B.K., Billingsley, P.F., Laurens, M.B., Gunasekera, A., Chakravarty, S., James, E.R., Sedegah, M., et al. (2011). Live attenuated malaria vaccine designed to protect through hepatic CD8 + T cell immunity. Science 334, Gamo, F.J., Sanz, L.M., Vidal, J., de Cozar, C., Alvarez, E., Lavandera, J.L., Vanderwall, D.E., Green, D.V., Kumar, V., Hasan, S., et al. (2010). Thousands of chemical starting points for antimalarial lead identification. Nature 465, Garnham, P.C., Bray, R.S., Bruce-Chwatt, L.J., Draper, C.C., Killick-Kendrick, R., Sergiev, P.G., Tiburskaja, N.A., Shute, P.G., and Maryon, M. (1975). A strain of Plasmodium vivax characterized by prolonged incubation: morphological and biological characteristics. Bull. World Health Organ. 52, Guguen-Guillouzo, C., and Guillouzo, A. (2010). General review on in vitro hepatocyte models and their applications. Methods Mol. Biol. 640, Guiguemde, W.A., Shelat, A.A., Bouck, D., Duffy, S., Crowther, G.J., Davis, P.H., Smithson, D.C., Connelly, M., Clark, J., Zhu, F., et al. (2010). Chemical genetics of Plasmodium falciparum. Nature 465, Hoffman, S.L., Isenbarger, D., Long, G.W., Sedegah, M., Szarfman, A., Waters, L., Hollingdale, M.R., van der Meide, P.H., Finbloom, D.S., and Ballou, W.R. (1989). Sporozoite vaccine induces genetically restricted T cell elimination of malaria from hepatocytes. Science 244, Hoffman, S.L., Billingsley, P.F., James, E., Richman, A., Loyevsky, M., Li, T., Chakravarty, S., Gunasekera, A., Chattopadhyay, R., Li, M., et al. (2010). Development of a metabolically active, non-replicating sporozoite vaccine to prevent Plasmodium falciparum malaria. Hum. Vaccin. 6, Hollingdale, M.R., Leland, P., and Schwartz, A.L. (1983). In vitro cultivation of the exoerythrocytic stage of Plasmodium berghei in a hepatoma cell line. Am. J. Trop. Med. Hyg. 32, Hollingdale, M.R., Collins, W.E., and Campbell, C.C. (1986). In vitro culture of exoerythrocytic parasites of the North Korean strain of Plasmodium vivax in hepatoma cells. Am. J. Trop. Med. Hyg. 35, Jiang, G., Shi, M., Conteh, S., Richie, N., Banania, G., Geneshan, H., Valencia, A., Singh, P., Aguiar, J., Limbach, K., et al. (2009). Sterile protection against Plasmodium knowlesi in rhesus monkeys from a malaria vaccine: comparison of heterologous prime boost strategies. PLoS ONE 4, e6559. Jones, T.R., Kang, I.H., Wheeler, D.B., Lindquist, R.A., Papallo, A., Sabatini, D.M., Golland, P., and Carpenter, A.E. (2008). CellProfiler Analyst: data exploration and analysis software for complex image-based screens. BMC Bioinformatics 9, 482. Jones, C.T., Catanese, M.T., Law, L.M., Khetani, S.R., Syder, A.J., Ploss, A., Oh, T.S., Schoggins, J.W., MacDonald, M.R., Bhatia, S.N., and Rice, C.M. (2010). Real-time imaging of hepatitis C virus infection using a fluorescent cell-based reporter system. Nat. Biotechnol. 28, Karnasuta, C., Pavanand, K., Chantakulkij, S., Luttiwongsakorn, N., Rassamesoraj, M., Laohathai, K., Webster, H.K., and Watt, G. (1995). Complete development of the liver stage of Plasmodium falciparum in a human hepatoma cell line. Am. J. Trop. Med. Hyg. 53, Khetani, S.R., and Bhatia, S.N. (2008). Microscale culture of human liver cells for drug development. Nat. Biotechnol. 26, Cell Host & Microbe 14, , July 17, 2013 ª2013 Elsevier Inc.
12 Krotoski, W.A., Collins, W.E., Bray, R.S., Garnham, P.C., Cogswell, F.B., Gwadz, R.W., Killick-Kendrick, R., Wolf, R., Sinden, R., Koontz, L.C., and Stanfill, P.S. (1982). Demonstration of hypnozoites in sporozoite-transmitted Plasmodium vivax infection. Am. J. Trop. Med. Hyg. 31, Krotoski, W.A., Garnham, P.C., Cogswell, F.B., Collins, W.E., Bray, R.S., Gwasz, R.W., Killick-Kendrick, R., Wolf, R.H., Sinden, R., Hollingdale, M., et al. (1986). Observations on early and late post-sporozoite tissue stages in primate malaria. IV. Pre-erythrocytic schizonts and/or hypnozoites of Chesson and North Korean strains of Plasmodium vivax in the chimpanzee. Am. J. Trop. Med. Hyg. 35, LeCluyse, E.L., Witek, R.P., Andersen, M.E., and Powers, M.J. (2012). Organotypic liver culture models: meeting current challenges in toxicity testing. Crit. Rev. Toxicol. 42, Mazier, D., Landau, I., Druilhe, P., Miltgen, F., Guguen-Guillouzo, C., Baccam, D., Baxter, J., Chigot, J.P., and Gentilini, M. (1984). Cultivation of the liver forms of Plasmodium vivax in human hepatocytes. Nature 307, Mazier, D., Beaudoin, R.L., Mellouk, S., Druilhe, P., Texier, B., Trosper, J., Miltgen, F., Landau, I., Paul, C., Brandicourt, O., et al. (1985). Complete development of hepatic stages of Plasmodium falciparum in vitro. Science 227, McCutchan, T.F., Lal, A.A., de la Cruz, V.F., Miller, L.H., Maloy, W.L., Charoenvit, Y., Beaudoin, R.L., Guerry, P., Wistar, R., Jr., Hoffman, S.L., et al. (1985). Sequence of the immunodominant epitope for the surface protein on sporozoites of Plasmodium vivax. Science 230, Meis, J.F., Rijntjes, P.J., Verhave, J.P., Ponnudurai, T., Hollingdale, M.R., and Yap, S.H. (1985). Infection of cryopreserved adult human hepatocytes with Plasmodium falciparum sporozoites. Cell Biol. Int. Rep. 9, 976. Mota, M.M., Pradel, G., Vanderberg, J.P., Hafalla, J.C., Frevert, U., Nussenzweig, R.S., Nussenzweig, V., and Rodríguez, A. (2001). Migration of Plasmodium sporozoites through cells before infection. Science 291, Mueller, A.K., Labaied, M., Kappe, S.H., and Matuschewski, K. (2005). Genetically modified Plasmodium parasites as a protective experimental malaria vaccine. Nature 433, Nardin, E.H., Nussenzweig, V., Nussenzweig, R.S., Collins, W.E., Harinasuta, K.T., Tapchaisri, P., and Chomcharn, Y. (1982). Circumsporozoite proteins of human malaria parasites Plasmodium falciparum and Plasmodium vivax. J. Exp. Med. 156, Ploss, A., Khetani, S.R., Jones, C.T., Syder, A.J., Trehan, K., Gaysinskaya, V.A., Mu, K., Ritola, K., Rice, C.M., and Bhatia, S.N. (2010). Persistent hepatitis C virus infection in microscale primary human hepatocyte cultures. Proc. Natl. Acad. Sci. USA 107, Plowe, C.V., Alonso, P., and Hoffman, S.L. (2009). The potential role of vaccines in the elimination of falciparum malaria and the eventual eradication of malaria. J. Infect. Dis. 200, Price, R.N., Tjitra, E., Guerra, C.A., Yeung, S., White, N.J., and Anstey, N.M. (2007). Vivax malaria: neglected and not benign. Am. J. Trop. Med. Hyg. 77 (6, Suppl), Prudêncio, M., Rodrigues, C.D., Hannus, M., Martin, C., Real, E., Gonçalves, L.A., Carret, C., Dorkin, R., Röhl, I., Jahn-Hoffmann, K., et al. (2008). Kinome-wide RNAi screen implicates at least 5 host hepatocyte kinases in Plasmodium sporozoite infection. PLoS Pathog. 4, e Pybus, B.S., Sousa, J.C., Jin, X., Ferguson, J.A., Christian, R.E., Barnhart, R., Vuong, C., Sciotti, R.J., Reichard, G.A., Kozar, M.P., et al. (2012). CYP450 phenotyping and accurate mass identification of metabolites of the 8-aminoquinoline, anti-malarial drug primaquine. Malar. J. 11, 259. Rénia, L., Miltgen, F., Charoenvit, Y., Ponnudurai, T., Verhave, J.P., Collins, W.E., and Mazier, D. (1988). Malaria sporozoite penetration. A new approach by double staining. J. Immunol. Methods 112, Rodrigues, C.D., Hannus, M., Prudêncio, M., Martin, C., Gonçalves, L.A., Portugal, S., Epiphanio, S., Akinc, A., Hadwiger, P., Jahn-Hofmann, K., et al. (2008). Host scavenger receptor SR-BI plays a dual role in the establishment of malaria parasite liver infection. Cell Host Microbe 4, Sattabongkot, J., Yimamnuaychoke, N., Leelaudomlipi, S., Rasameesoraj, M., Jenwithisuk, R., Coleman, R.E., Udomsangpetch, R., Cui, L., and Brewer, T.G. (2006). Establishment of a human hepatocyte line that supports in vitro development of the exo-erythrocytic stages of the malaria parasites Plasmodium falciparum and P. vivax. Am. J. Trop. Med. Hyg. 74, Shortt, H.E., and Garnham, P.C. (1948). The pre-erythrocytic development of Plasmodium cynomolgi and Plasmodium vivax. Trans. R. Soc. Trop. Med. Hyg. 41, Shortt, H.E., Fairley, N.H., Covell, G., Shute, P.G., and Garnham, P.C. (1951). The pre-erythrocytic stage of Plasmodium falciparum. Trans. R. Soc. Trop. Med. Hyg. 44, Silvie, O., Rubinstein, E., Franetich, J.F., Prenant, M., Belnoue, E., Rénia, L., Hannoun, L., Eling, W., Levy, S., Boucheix, C., and Mazier, D. (2003). Hepatocyte CD81 is required for Plasmodium falciparum and Plasmodium yoelii sporozoite infectivity. Nat. Med. 9, Silvie, O., Franetich, J.F., Charrin, S., Mueller, M.S., Siau, A., Bodescot, M., Rubinstein, E., Hannoun, L., Charoenvit, Y., Kocken, C.H., et al. (2004). A role for apical membrane antigen 1 during invasion of hepatocytes by Plasmodium falciparum sporozoites. J. Biol. Chem. 279, Silvie, O., Franetich, J.F., Boucheix, C., Rubinstein, E., and Mazier, D. (2007). Alternative invasion pathways for Plasmodium berghei sporozoites. Int. J. Parasitol. 37, van Schaijk, B.C., Janse, C.J., van Gemert, G.J., van Dijk, M.R., Gego, A., Franetich, J.F., van de Vegte-Bolmer, M., Yalaoui, S., Silvie, O., Hoffman, S.L., et al. (2008). Gene disruption of Plasmodium falciparum p52 results in attenuation of malaria liver stage development in cultured primary human hepatocytes. PLoS ONE 3, e3549. Vaughan, A.M., Mikolajczak, S.A., Wilson, E.M., Grompe, M., Kaushansky, A., Camargo, N., Bial, J., Ploss, A., and Kappe, S.H. (2012). Complete Plasmodium falciparum liver-stage development in liver-chimeric mice. J. Clin. Invest. 122, Wang, W.W., Khetani, S.R., Krzyzewski, S., Duignan, D.B., and Obach, R.S. (2010). Assessment of a micropatterned hepatocyte coculture system to generate major human excretory and circulating drug metabolites. Drug Metab. Dispos. 38, Wells, T.N., Burrows, J.N., and Baird, J.K. (2010). Targeting the hypnozoite reservoir of Plasmodium vivax: the hidden obstacle to malaria elimination. Trends Parasitol. 26, World Health Organization. (2010). World Malaria Report 2010 (Geneva: World Health Organization). Yalaoui, S., Huby, T., Franetich, J.F., Gego, A., Rametti, A., Moreau, M., Collet, X., Siau, A., van Gemert, G.J., Sauerwein, R.W., et al. (2008). Scavenger receptor BI boosts hepatocyte permissiveness to Plasmodium infection. Cell Host Microbe 4, Yokoo, H., Kondo, T., Fujii, K., Yamada, T., Todo, S., and Hirohashi, S. (2004). Proteomic signature corresponding to alpha fetoprotein expression in liver cancer cells. Hepatology 40, Cell Host & Microbe 14, , July 17, 2013 ª2013 Elsevier Inc. 115
13 , Volume 14 Supplemental Information A Microscale Human Liver Platform that Supports the Hepatic Stages of Plasmodium falciparum and vivax Sandra March, Shengyong Ng, Soundarapandian Velmurugan, Ani Galstian, Jing Shan, David J. Logan, Anne E. Carpenter, David Thomas, B. Kim Lee Sim, Maria M. Mota, Stephen L. Hoffman, and Sangeeta N. Bhatia 1

. Figure S2.")
14 Figure S1. Cryopreserved Human Hepatocytes Morphology, Related to Figure 1 Morphology of cryopreserved human hepatocytes from different donors in micropatterned cocultures (day 18 post-seeding). Figure S2. Inhibitory Effect of Primaquine, Related to Figure 4 Inhibitory effect of primaquine (PQ) using two different hepatocyte donors. 2


15 Figure S3. Image Automation Workflow, Related to Figure 5 Images of 96 well plates are acquired using high content screening microscopes and analyzed by Cell Profiler (Broad Institute). Parasites are visualized through immunofluorescent staining of the HSP70 protein. Images of potential parasites are manually classified using Cell Profiler, which then learns the algorithm to score all images. 3

16 4
Infecting Anopheles stephensi With Rodent Malaria Parasites Alida Coppi & Photini Sinnis
 Infecting Anopheles stephensi With Rodent Malaria Parasites Alida Coppi & Photini Sinnis A. Reagents: 1. DMEM or RPMI DMEM (4.5g/L glucose) RPMI 1640 Cellgro #MT-10-017-CM Cellgro #MT-10-040-CM 2. Giemsa
Infecting Anopheles stephensi With Rodent Malaria Parasites Alida Coppi & Photini Sinnis A. Reagents: 1. DMEM or RPMI DMEM (4.5g/L glucose) RPMI 1640 Cellgro #MT-10-017-CM Cellgro #MT-10-040-CM 2. Giemsa
Gliding Motility Assay for P. berghei Sporozoites
 Gliding Motility Assay for P. berghei Sporozoites Important Notes: 1. For all dilutions (including antibodies and sporozoites), always make slightly more than needed. For instance, if you need 200 µl sporozoites
Gliding Motility Assay for P. berghei Sporozoites Important Notes: 1. For all dilutions (including antibodies and sporozoites), always make slightly more than needed. For instance, if you need 200 µl sporozoites
alaria Parasite Bank Collection sites of P. falciparum isolates PARASITE BIOLOGY
 M alaria Parasite Bank established in 1992 is a supporting unit for research activities on different aspects of malaria. The main objective of establishing this facility is to strengthen researches at
M alaria Parasite Bank established in 1992 is a supporting unit for research activities on different aspects of malaria. The main objective of establishing this facility is to strengthen researches at
A. Effect upon human culture 1. Control of malaria has contributed to world=s population explosion 2. Africans brought to U.S.
 VI. Malaria A. Effect upon human culture 1. Control of malaria has contributed to world=s population explosion 2. Africans brought to U.S. because they were resistant to malaria & other diseases 3. Many
VI. Malaria A. Effect upon human culture 1. Control of malaria has contributed to world=s population explosion 2. Africans brought to U.S. because they were resistant to malaria & other diseases 3. Many
Quantitative Dynamics of Plasmodium yoelii Sporozoite Transmission by Infected Anopheline Mosquitoes
 INFECTION AND IMMUNITY, July 2005, p. 4363 4369 Vol. 73, No. 7 0019-9567/05/$08.00 0 doi:10.1128/iai.73.7.4363 4369.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Quantitative
INFECTION AND IMMUNITY, July 2005, p. 4363 4369 Vol. 73, No. 7 0019-9567/05/$08.00 0 doi:10.1128/iai.73.7.4363 4369.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Quantitative
Parasitology Departement Medical Faculty of USU
 Malaria Mechanism of infection Parasitology Departement Medical Faculty of USU Introduction Malaria parasites Phylum Order Suborder Family Genus Species : : Apicomplexa : Eucoccidiida : Haemosporida :
Malaria Mechanism of infection Parasitology Departement Medical Faculty of USU Introduction Malaria parasites Phylum Order Suborder Family Genus Species : : Apicomplexa : Eucoccidiida : Haemosporida :
PLASMODIUM MODULE 39.1 INTRODUCTION OBJECTIVES 39.2 MALARIAL PARASITE. Notes
 Plasmodium MODULE 39 PLASMODIUM 39.1 INTRODUCTION Malaria is characterized by intermittent fever associated with chills and rigors in the patient. There may be enlargement of the liver and spleen in the
Plasmodium MODULE 39 PLASMODIUM 39.1 INTRODUCTION Malaria is characterized by intermittent fever associated with chills and rigors in the patient. There may be enlargement of the liver and spleen in the
Novel ELISA method as exploratory tool to assess immunity induced by radiated attenuated sporozoites to decipher protective immunity
 DOI 10.1186/s12936-017-2129-9 Malaria Journal METHODOLOGY Open Access Novel ELISA method as exploratory tool to assess immunity induced by radiated attenuated sporozoites to decipher protective immunity
DOI 10.1186/s12936-017-2129-9 Malaria Journal METHODOLOGY Open Access Novel ELISA method as exploratory tool to assess immunity induced by radiated attenuated sporozoites to decipher protective immunity
Malaria. This sheet is from both sections recording and includes all slides and diagrams.
 Malaria This sheet is from both sections recording and includes all slides and diagrams. Malaria is caused by protozoa family called plasmodium (Genus) mainly affect blood system specially RBCs and each
Malaria This sheet is from both sections recording and includes all slides and diagrams. Malaria is caused by protozoa family called plasmodium (Genus) mainly affect blood system specially RBCs and each
Identification of an AP2-family Protein That Is Critical for Malaria Liver Stage Development
 Identification of an AP2-family Protein That Is Critical for Malaria Liver Stage Development Shiroh Iwanaga, Izumi Kaneko, Tomomi Kato, Masao Yuda* Department of Medical Zoology, Mie University School
Identification of an AP2-family Protein That Is Critical for Malaria Liver Stage Development Shiroh Iwanaga, Izumi Kaneko, Tomomi Kato, Masao Yuda* Department of Medical Zoology, Mie University School
PRINCIPAL INVESTIGATOR: Dr. Jetsumon (Sattabongkot) Prachumsri
 Prachumsri") AD (Leave blank) Award Number: W81XWH-07-2-0090 TITLE: Proteomic Study of Human Malaria Parasite Plasmodium Vivax Liver Stages for Development of Vaccines and Drugs PRINCIPAL INVESTIGATOR: Dr. Jetsumon
AD (Leave blank) Award Number: W81XWH-07-2-0090 TITLE: Proteomic Study of Human Malaria Parasite Plasmodium Vivax Liver Stages for Development of Vaccines and Drugs PRINCIPAL INVESTIGATOR: Dr. Jetsumon
INVESTIGATING THE MOTILITY OF PLASMODIUM
 INVESTIGATING THE MOTILITY OF PLASMODIUM by Natasha Vartak A thesis submitted to Johns Hopkins University in conformity with the requirements for the degree of Master of Science Baltimore, Maryland April,
INVESTIGATING THE MOTILITY OF PLASMODIUM by Natasha Vartak A thesis submitted to Johns Hopkins University in conformity with the requirements for the degree of Master of Science Baltimore, Maryland April,
Blood protozoan: Plasmodium
 Blood protozoan: Plasmodium Dr. Hala Al Daghistani The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans: four species are associated The Plasmodium spp.
Blood protozoan: Plasmodium Dr. Hala Al Daghistani The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans: four species are associated The Plasmodium spp.
Blood protozoan: Plasmodium
 Blood protozoan: Plasmodium The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans:four species are associated The Plasmodium spp. life cycle can be divided
Blood protozoan: Plasmodium The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans:four species are associated The Plasmodium spp. life cycle can be divided
Arrested oocyst maturation in Plasmodium parasites. lacking type II NADH:ubiquinone dehydrogenase
 Supplemental Information for: Arrested oocyst maturation in Plasmodium parasites lacking type II NADH:ubiquinone dehydrogenase Katja E. Boysen and Kai Matuschewski Contents: - Supplemental Movies 1 and
Supplemental Information for: Arrested oocyst maturation in Plasmodium parasites lacking type II NADH:ubiquinone dehydrogenase Katja E. Boysen and Kai Matuschewski Contents: - Supplemental Movies 1 and
A:Malaria (Plasmodium species) Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign
 Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign") A:Malaria (Plasmodium species) Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign tertian malaria P. ovale: causes benign tertian malaria
A:Malaria (Plasmodium species) Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign tertian malaria P. ovale: causes benign tertian malaria
Chimeric Plasmodium falciparum parasites expressing Plasmodium vivax circumsporozoite protein fail to produce salivary gland sporozoites
 https://doi.org/10.1186/s12936-018-2431-1 Malaria Journal RESEARCH Open Access Chimeric Plasmodium falciparum parasites expressing Plasmodium vivax circumsporozoite protein fail to produce salivary gland
https://doi.org/10.1186/s12936-018-2431-1 Malaria Journal RESEARCH Open Access Chimeric Plasmodium falciparum parasites expressing Plasmodium vivax circumsporozoite protein fail to produce salivary gland
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12234 Supplementary Figure 1. Embryonic naked mole-rat fibroblasts do not undergo ECI. Embryonic naked mole-rat fibroblasts ( EF) were isolated from eight mid-gestation embryos. All the
doi:10.1038/nature12234 Supplementary Figure 1. Embryonic naked mole-rat fibroblasts do not undergo ECI. Embryonic naked mole-rat fibroblasts ( EF) were isolated from eight mid-gestation embryos. All the
BIO Parasitology Spring 2009
 BIO 475 - Parasitology Spring 2009 Stephen M. Shuster Northern Arizona University http://www4.nau.edu/isopod Lecture 10 Malaria-Life Cycle a. Micro and macrogametocytes in mosquito stomach. b. Ookinete
BIO 475 - Parasitology Spring 2009 Stephen M. Shuster Northern Arizona University http://www4.nau.edu/isopod Lecture 10 Malaria-Life Cycle a. Micro and macrogametocytes in mosquito stomach. b. Ookinete
Malaria parasites: virulence and transmission as a basis for intervention strategies
 Malaria parasites: virulence and transmission as a basis for intervention strategies Matthias Marti Department of Immunology and Infectious Diseases Harvard School of Public Health The global malaria burden
Malaria parasites: virulence and transmission as a basis for intervention strategies Matthias Marti Department of Immunology and Infectious Diseases Harvard School of Public Health The global malaria burden
Epigenetic regulation of Plasmodium falciparum clonally. variant gene expression during development in An. gambiae
 Epigenetic regulation of Plasmodium falciparum clonally variant gene expression during development in An. gambiae Elena Gómez-Díaz, Rakiswendé S. Yerbanga, Thierry Lefèvre, Anna Cohuet, M. Jordan Rowley,
Epigenetic regulation of Plasmodium falciparum clonally variant gene expression during development in An. gambiae Elena Gómez-Díaz, Rakiswendé S. Yerbanga, Thierry Lefèvre, Anna Cohuet, M. Jordan Rowley,
A Cysteine Protease Inhibitor of Plasmodium berghei Is Essential for Exo-erythrocytic Development
 A Cysteine Protease Inhibitor of Plasmodium berghei Is Essential for Exo-erythrocytic Development Christine Lehmann 1, Anna Heitmann 1, Satish Mishra 2, Paul-Christian Burda 3, Mirko Singer 4, Monica Prado
A Cysteine Protease Inhibitor of Plasmodium berghei Is Essential for Exo-erythrocytic Development Christine Lehmann 1, Anna Heitmann 1, Satish Mishra 2, Paul-Christian Burda 3, Mirko Singer 4, Monica Prado
Understanding Epidemics Section 3: Malaria & Modelling
 Understanding Epidemics Section 3: Malaria & Modelling PART B: Biology Contents: Vector and parasite Biology of the malaria parasite Biology of the anopheles mosquito life cycle Vector and parasite Malaria
Understanding Epidemics Section 3: Malaria & Modelling PART B: Biology Contents: Vector and parasite Biology of the malaria parasite Biology of the anopheles mosquito life cycle Vector and parasite Malaria
Plasmodium 18S rrna of intravenously administered sporozoites does not persist in peripheral blood
 https://doi.org/10.1186/s12936-018-2422-2 Malaria Journal RESEARCH Open Access Plasmodium 18S rrna of intravenously administered sporozoites does not persist in peripheral blood Sean C. Murphy 1,2*, Andrew
https://doi.org/10.1186/s12936-018-2422-2 Malaria Journal RESEARCH Open Access Plasmodium 18S rrna of intravenously administered sporozoites does not persist in peripheral blood Sean C. Murphy 1,2*, Andrew
GUIDE TO INFECTION CONTROL IN THE HOSPITAL. Antibiotic Resistance
 GUIDE TO INFECTION CONTROL IN THE HOSPITAL CHAPTER 4: Antibiotic Resistance Author M.P. Stevens, MD, MPH S. Mehtar, MD R.P. Wenzel, MD, MSc Chapter Editor Michelle Doll, MD, MPH Topic Outline Key Issues
GUIDE TO INFECTION CONTROL IN THE HOSPITAL CHAPTER 4: Antibiotic Resistance Author M.P. Stevens, MD, MPH S. Mehtar, MD R.P. Wenzel, MD, MSc Chapter Editor Michelle Doll, MD, MPH Topic Outline Key Issues
Malaria parasites of rodents of the Congo (Brazzaville) :
 :") Annales de Parasitologie (Paris), 1976, t. 51, n 6, pp. 637 à 646 Malaria parasites of rodents of the Congo (Brazzaville) : Plasmodium cbabaudi adami subsp. nov. and Plasmodium vinckei lentum Landau, Michel,
Annales de Parasitologie (Paris), 1976, t. 51, n 6, pp. 637 à 646 Malaria parasites of rodents of the Congo (Brazzaville) : Plasmodium cbabaudi adami subsp. nov. and Plasmodium vinckei lentum Landau, Michel,
Malaria Parasite Pre-Erythrocytic Stage Infection: Gliding and Hiding
 Malaria Parasite Pre-Erythrocytic Stage Infection: Gliding and Hiding Ashley M. Vaughan, 1 Ahmed S.I. Aly, 1 and Stefan H.I. Kappe 1,2, * 1 Seattle Biomedical Research Institute, Seattle, WA 98109, USA
Malaria Parasite Pre-Erythrocytic Stage Infection: Gliding and Hiding Ashley M. Vaughan, 1 Ahmed S.I. Aly, 1 and Stefan H.I. Kappe 1,2, * 1 Seattle Biomedical Research Institute, Seattle, WA 98109, USA
Sera from 2,500 animals from three different groups were analysed:
 FIELD TRIAL OF A BRUCELLOSIS COMPETITIVE ENZYME LINKED IMMUNOABSORBENT ASSAY (ELISA) L.E. SAMARTINO, R.J. GREGORET, G. SIGAL INTA-CICV Instituto Patobiología Area Bacteriología, Buenos Aires, Argentina
FIELD TRIAL OF A BRUCELLOSIS COMPETITIVE ENZYME LINKED IMMUNOABSORBENT ASSAY (ELISA) L.E. SAMARTINO, R.J. GREGORET, G. SIGAL INTA-CICV Instituto Patobiología Area Bacteriología, Buenos Aires, Argentina
PDF hosted at the Radboud Repository of the Radboud University Nijmegen
 PDF hosted at the Radboud Repository of the Radboud University Nijmegen The following full text is a publisher's version. For additional information about this publication click this link. http://hdl.handle.net/2066/70169
PDF hosted at the Radboud Repository of the Radboud University Nijmegen The following full text is a publisher's version. For additional information about this publication click this link. http://hdl.handle.net/2066/70169
Boosting Bacterial Metabolism to Combat Antibiotic Resistance
 Boosting Bacterial Metabolism to Combat Antibiotic Resistance The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published
Boosting Bacterial Metabolism to Combat Antibiotic Resistance The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published
COMMITTEE FOR MEDICINAL PRODUCTS FOR VETERINARY USE
 European Medicines Agency Veterinary Medicines and Inspections EMEA/CVMP/211249/2005-FINAL July 2005 COMMITTEE FOR MEDICINAL PRODUCTS FOR VETERINARY USE DIHYDROSTREPTOMYCIN (Extrapolation to all ruminants)
European Medicines Agency Veterinary Medicines and Inspections EMEA/CVMP/211249/2005-FINAL July 2005 COMMITTEE FOR MEDICINAL PRODUCTS FOR VETERINARY USE DIHYDROSTREPTOMYCIN (Extrapolation to all ruminants)
Plasmodium yoelii Sporozoites with Simultaneous Deletion of P52 and P36 Are Completely Attenuated and Confer Sterile Immunity against Infection
 INFECTION AND IMMUNITY, Aug. 2007, p. 3758 3768 Vol. 75, No. 8 0019-9567/07/$08.00 0 doi:10.1128/iai.00225-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Plasmodium yoelii Sporozoites
INFECTION AND IMMUNITY, Aug. 2007, p. 3758 3768 Vol. 75, No. 8 0019-9567/07/$08.00 0 doi:10.1128/iai.00225-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Plasmodium yoelii Sporozoites
Fluoroquinolones ELISA KIT
 Fluoroquinolones ELISA KIT Cat. No.:DEIA6883 Pkg.Size:96T Intended use The Fluoroquinolones ELISA KIT is an immunoassay for the detection of Fluoroquinolones in contaminated samples including water, fish
Fluoroquinolones ELISA KIT Cat. No.:DEIA6883 Pkg.Size:96T Intended use The Fluoroquinolones ELISA KIT is an immunoassay for the detection of Fluoroquinolones in contaminated samples including water, fish
The silent path to thousands of merozoites: the Plasmodium liver stage
 The silent path to thousands of merozoites: the Plasmodium liver stage Miguel Prudêncio*, Ana Rodriguez and Maria M. Mota* Abstract Plasmodium sporozoites are deposited in the skin of their vertebrate
The silent path to thousands of merozoites: the Plasmodium liver stage Miguel Prudêncio*, Ana Rodriguez and Maria M. Mota* Abstract Plasmodium sporozoites are deposited in the skin of their vertebrate
Evaluation of the hair growth and retention activity of two solutions on human hair explants
 activity of two solutions on human hair explants Study Directed by Dr E. Lati of Laboratoire Bio-EC, Centre de Recherches Biologiques et d Experimentations Cutanees, on behalf of Pangaea Laboratories Ltd.
activity of two solutions on human hair explants Study Directed by Dr E. Lati of Laboratoire Bio-EC, Centre de Recherches Biologiques et d Experimentations Cutanees, on behalf of Pangaea Laboratories Ltd.
Malaria in the Mosquito Dr. Peter Billingsley
 Malaria in the Mosquito Senior Director Quality Systems and Entomology Research Sanaria Inc. Rockville MD. 1 Malaria: one of the world s foremost killers Every year 1 million children die of malaria 250
Malaria in the Mosquito Senior Director Quality Systems and Entomology Research Sanaria Inc. Rockville MD. 1 Malaria: one of the world s foremost killers Every year 1 million children die of malaria 250
Visit ABLE on the Web at:
 This article reprinted from: Lessem, P. B. 2008. The antibiotic resistance phenomenon: Use of minimal inhibitory concentration (MIC) determination for inquiry based experimentation. Pages 357-362, in Tested
This article reprinted from: Lessem, P. B. 2008. The antibiotic resistance phenomenon: Use of minimal inhibitory concentration (MIC) determination for inquiry based experimentation. Pages 357-362, in Tested
The Hypnozoite and Relapse in Primate Malaria
 CLINICAL MICROBIOLOGY REVIEWS, Jan. 1992, p. 26-35 Vol. 5, No. 1 0893-8512/92/010026-10$02.00/0 Copyright 1992, American Society for Microbiology The Hypnozoite and Relapse in Primate Malaria FRANK B.
CLINICAL MICROBIOLOGY REVIEWS, Jan. 1992, p. 26-35 Vol. 5, No. 1 0893-8512/92/010026-10$02.00/0 Copyright 1992, American Society for Microbiology The Hypnozoite and Relapse in Primate Malaria FRANK B.
CelTOS, a novel malarial protein that mediates transmission to mosquito and vertebrate hosts
 Blackwell Publishing LtdOxford, UKMMIMolecular Microbiology0950-382X 2005 The Authors; Journal compilation 2005 Blackwell Publishing Ltd? 200559513691379Original ArticleA protein that mediates malarial
Blackwell Publishing LtdOxford, UKMMIMolecular Microbiology0950-382X 2005 The Authors; Journal compilation 2005 Blackwell Publishing Ltd? 200559513691379Original ArticleA protein that mediates malarial
A rapid and scalable density gradient purification method for Plasmodium sporozoites
 Kennedy et al. Malaria Journal 2012, 11:421 METHODOLOGY Open Access A rapid and scalable density gradient purification method for Plasmodium sporozoites Mark Kennedy 1, Matthew E Fishbaugher 1, Ashley
Kennedy et al. Malaria Journal 2012, 11:421 METHODOLOGY Open Access A rapid and scalable density gradient purification method for Plasmodium sporozoites Mark Kennedy 1, Matthew E Fishbaugher 1, Ashley
EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS WORK-PROGRAMME PROPOSAL Version 2 VISAVET. Universidad Complutense de Madrid
 FOR BOVINE TUBERCULOSIS WORK-PROGRAMME PROPOSAL Version 2 VISAVET. Universidad Complutense de Madrid") EUROPEAN COMMISSION HEALTH & CONSUMERS DIRECTORATE-GENERAL Directorate D Animal Health and Welfare Unit D1- Animal health and Standing Committees EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS
EUROPEAN COMMISSION HEALTH & CONSUMERS DIRECTORATE-GENERAL Directorate D Animal Health and Welfare Unit D1- Animal health and Standing Committees EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
PREPARED FOR: U.S. Army Medical Research and Materiel Command Fort Detrick, Maryland X Approved for public release; distribution unlimited
 Award Number: W8XWH--- TITLE: Defining the Role of Autophagy Kinase ULK Signaling in Therapeutic Response of Tuberous Sclerosis Complex to Inhibitors PRINCIPAL INVESTIGATOR: Reuben J. Shaw, Ph.D. CONTRACTING
Award Number: W8XWH--- TITLE: Defining the Role of Autophagy Kinase ULK Signaling in Therapeutic Response of Tuberous Sclerosis Complex to Inhibitors PRINCIPAL INVESTIGATOR: Reuben J. Shaw, Ph.D. CONTRACTING
Applied-for scope of designation and notification of a Conformity Assessment Body Regulation (EU) 2017/746 (IVDR)
 2017/746 (IVDR)") Ref. Ares(2018)2576484-17/05/2018 NBOG s Best Practice Guide applicable for MDR IVDR NBOG F 2017-4 This document has been endorsed by the Medical Device Coordination Group (MDCG) established by Article
Ref. Ares(2018)2576484-17/05/2018 NBOG s Best Practice Guide applicable for MDR IVDR NBOG F 2017-4 This document has been endorsed by the Medical Device Coordination Group (MDCG) established by Article
Development and improvement of diagnostics to improve use of antibiotics and alternatives to antibiotics
 Priority Topic B Diagnostics Development and improvement of diagnostics to improve use of antibiotics and alternatives to antibiotics The overarching goal of this priority topic is to stimulate the design,
Priority Topic B Diagnostics Development and improvement of diagnostics to improve use of antibiotics and alternatives to antibiotics The overarching goal of this priority topic is to stimulate the design,
VETERINARY SCIENCE CURRICULUM. Unit 1: Safety and Sanitation
 Chariho Regional School District - Science Curriculum September, 2016 VETERINARY SCIENCE CURRICULUM Unit 1: Safety and Sanitation Students will gain an understanding of the types of hazards common in veterinary
Chariho Regional School District - Science Curriculum September, 2016 VETERINARY SCIENCE CURRICULUM Unit 1: Safety and Sanitation Students will gain an understanding of the types of hazards common in veterinary
Research with Animals
 Research with Animals Matthew Olugbenga Oyeyemi momattyemi@gmail.com +2348038059952 Research with Animals 1 Objectives Describe situations when animals may be research subjects Identify laws and regulations
Research with Animals Matthew Olugbenga Oyeyemi momattyemi@gmail.com +2348038059952 Research with Animals 1 Objectives Describe situations when animals may be research subjects Identify laws and regulations
Developmental Biology of Sporozoite-Host. Malaria: Implications for Vaccine Design. Javier E. Garcia, Alvaro Puentes and Manuel E.
 Developmental Biology of Sporozoite-Host Interactions in Plasmodium falciparum Malaria: Implications for Vaccine Design Javier E. Garcia, Alvaro Puentes and Manuel E. Patarroyo Clin. Microbiol. Rev. 2006,
Developmental Biology of Sporozoite-Host Interactions in Plasmodium falciparum Malaria: Implications for Vaccine Design Javier E. Garcia, Alvaro Puentes and Manuel E. Patarroyo Clin. Microbiol. Rev. 2006,
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and
 Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
Informing Public Policy on Agricultural Use of Antimicrobials in the United States: Strategies Developed by an NGO
 Informing Public Policy on Agricultural Use of Antimicrobials in the United States: Strategies Developed by an NGO Stephen J. DeVincent, DVM, MA Director, Ecology Program Alliance for the Prudent Use of
Informing Public Policy on Agricultural Use of Antimicrobials in the United States: Strategies Developed by an NGO Stephen J. DeVincent, DVM, MA Director, Ecology Program Alliance for the Prudent Use of
Formation of Proximal and Anterior Limb Skeleton Requires Early Function of Irx3 and Irx5 and Is Negatively Regulated by Shh Signaling
 Developmental Cell, Volume 29 Supplemental Information Formation of Proximal and Anterior Limb Skeleton Requires Early Function of Irx3 and Irx5 and Is Negatively Regulated by Shh Signaling Danyi Li, Rui
Developmental Cell, Volume 29 Supplemental Information Formation of Proximal and Anterior Limb Skeleton Requires Early Function of Irx3 and Irx5 and Is Negatively Regulated by Shh Signaling Danyi Li, Rui
Diurnal variation in microfilaremia in cats experimentally infected with larvae of
 Hayasaki et al., Page 1 Short Communication Diurnal variation in microfilaremia in cats experimentally infected with larvae of Dirofilaria immitis M. Hayasaki a,*, J. Okajima b, K.H. Song a, K. Shiramizu
Hayasaki et al., Page 1 Short Communication Diurnal variation in microfilaremia in cats experimentally infected with larvae of Dirofilaria immitis M. Hayasaki a,*, J. Okajima b, K.H. Song a, K. Shiramizu
SAFETY PHARMACOLOGY: CARDIOVASCULAR TELEMETRY. Aileen Milne PhD, Manager, Safety Pharmacology
 SAFETY PHARMACOLOGY: CARDIOVASCULAR TELEMETRY Aileen Milne PhD, Manager, Safety Pharmacology SAFETY PHARMACOLOGY SERVICES OVERVIEW Full Range of S7A and S7B studies herg assay Respiratory function plethysmography(rat/mouse)
SAFETY PHARMACOLOGY: CARDIOVASCULAR TELEMETRY Aileen Milne PhD, Manager, Safety Pharmacology SAFETY PHARMACOLOGY SERVICES OVERVIEW Full Range of S7A and S7B studies herg assay Respiratory function plethysmography(rat/mouse)
Comparative Evaluation of Online and Paper & Pencil Forms for the Iowa Assessments ITP Research Series
 Comparative Evaluation of Online and Paper & Pencil Forms for the Iowa Assessments ITP Research Series Catherine J. Welch Stephen B. Dunbar Heather Rickels Keyu Chen ITP Research Series 2014.2 A Comparative
Comparative Evaluation of Online and Paper & Pencil Forms for the Iowa Assessments ITP Research Series Catherine J. Welch Stephen B. Dunbar Heather Rickels Keyu Chen ITP Research Series 2014.2 A Comparative
EVOLUTIONARY GENETICS (Genome 453) Midterm Exam Name KEY
 Midterm Exam Name KEY") PLEASE: Put your name on every page and SHOW YOUR WORK. Also, lots of space is provided, but you do not have to fill it all! Note that the details of these problems are fictional, for exam purposes only.
PLEASE: Put your name on every page and SHOW YOUR WORK. Also, lots of space is provided, but you do not have to fill it all! Note that the details of these problems are fictional, for exam purposes only.
COMMITTEE FOR VETERINARY MEDICINAL PRODUCTS
 The European Agency for the Evaluation of Medicinal Products Veterinary Medicines and Information Technology EMEA/MRL/728/00-FINAL April 2000 COMMITTEE FOR VETERINARY MEDICINAL PRODUCTS STREPTOMYCIN AND
The European Agency for the Evaluation of Medicinal Products Veterinary Medicines and Information Technology EMEA/MRL/728/00-FINAL April 2000 COMMITTEE FOR VETERINARY MEDICINAL PRODUCTS STREPTOMYCIN AND
Quantification of Chloramphenicol in Chicken Using Xevo TQD with RADAR Technology
 Quantification of Chloramphenicol in Chicken Using Xevo TQD with RADAR Technology Dimple Shah, Marian Twohig, and Jennifer A. Burgess Waters Corporation, Milford, MA, U.S.A. A P P L I C AT ION B E N E
Quantification of Chloramphenicol in Chicken Using Xevo TQD with RADAR Technology Dimple Shah, Marian Twohig, and Jennifer A. Burgess Waters Corporation, Milford, MA, U.S.A. A P P L I C AT ION B E N E
Antimicrobial Selection to Combat Resistance
 Antimicrobial Selection to Combat Resistance (Dead Bugs Don t Mutate!) Shelley C Rankin PhD Associate Professor CE Microbiology Head of Diagnostic Services & Chief of Clinical Microbiology Ryan Veterinary
Antimicrobial Selection to Combat Resistance (Dead Bugs Don t Mutate!) Shelley C Rankin PhD Associate Professor CE Microbiology Head of Diagnostic Services & Chief of Clinical Microbiology Ryan Veterinary
ENVIRACOR J-5 aids in the control of clinical signs associated with Escherichia coli (E. coli) mastitis
 mastitis") GDR11136 ENVIRACOR J-5 aids in the control of clinical signs associated with Escherichia coli (E. coli) mastitis February 2012 Summary The challenge data presented in this technical bulletin was completed
GDR11136 ENVIRACOR J-5 aids in the control of clinical signs associated with Escherichia coli (E. coli) mastitis February 2012 Summary The challenge data presented in this technical bulletin was completed
Enzootic Bovine Leukosis: Milk Screening and Verification ELISA: VF-P02210 & VF-P02220
 Enzootic Bovine Leukosis: Milk Screening and Verification ELISA: VF-P02210 & VF-P02220 Introduction Enzootic Bovine Leukosis is a transmissible disease caused by the Enzootic Bovine Leukosis Virus (BLV)
Enzootic Bovine Leukosis: Milk Screening and Verification ELISA: VF-P02210 & VF-P02220 Introduction Enzootic Bovine Leukosis is a transmissible disease caused by the Enzootic Bovine Leukosis Virus (BLV)
STATISTICAL REPORT. Preliminary Analysis of the Second Collaborative Study of the Hard Surface Carrier Test
 STATISTICAL REPORT To: From: Subject: Diane Boesenberg, Reckitt Benckiser Emily Mitchell, Product Science Branch, Antimicrobials Division/Office of Pesticide Programs/US EPA Martin Hamilton, Statistician
STATISTICAL REPORT To: From: Subject: Diane Boesenberg, Reckitt Benckiser Emily Mitchell, Product Science Branch, Antimicrobials Division/Office of Pesticide Programs/US EPA Martin Hamilton, Statistician
Dual Antibiotic Delivery from Chitosan Sponges Prevents In Vivo Polymicrobial Biofilm Infections
 Dual Antibiotic Delivery from Chitosan Sponges Prevents In Vivo Polymicrobial Biofilm Infections Ashley Parker, MS 1, James Smith, MS 1, Karen Beenken, PhD 2, Jessica Amber Jennings, PhD 3, Mark Smeltzer,
Dual Antibiotic Delivery from Chitosan Sponges Prevents In Vivo Polymicrobial Biofilm Infections Ashley Parker, MS 1, James Smith, MS 1, Karen Beenken, PhD 2, Jessica Amber Jennings, PhD 3, Mark Smeltzer,
23 Plasmodium coatneyi Eyles, Fong, Warren, Guinn, Sandosham, and Wharton, 1962
 23 Plasmodium coatneyi Eyles, Fong, Warren, Guinn, Sandosham, and Wharton, 1962 IN the course of studies on simian malaria begun by the late Dr. Don Eyles in Malaya, he and his co-workers isolated a new
23 Plasmodium coatneyi Eyles, Fong, Warren, Guinn, Sandosham, and Wharton, 1962 IN the course of studies on simian malaria begun by the late Dr. Don Eyles in Malaya, he and his co-workers isolated a new
THE TRANSMISSION EFFICIENCY OF PLASMODIUM YOELII INFECTED MOSQUITOES
 THE TRANSMISSION EFFICIENCY OF PLASMODIUM YOELII INFECTED MOSQUITOES by Maya A. Aleshnick A thesis submitted to Johns Hopkins University in conformity with the requirements for the degree of Master of
THE TRANSMISSION EFFICIENCY OF PLASMODIUM YOELII INFECTED MOSQUITOES by Maya A. Aleshnick A thesis submitted to Johns Hopkins University in conformity with the requirements for the degree of Master of
Application of Peristaltic Filling for Flexibility and Accuracy
 E03 - Aseptic Processing Technology 2008 Case Study: Application of Peristaltic Filling for Flexibility and Accuracy by Ted Kemnitz Automated Machine Technologies, Inc. AMT (919) 361 0121 Ted.Kemnitz@AMTLiquidFilling.com
E03 - Aseptic Processing Technology 2008 Case Study: Application of Peristaltic Filling for Flexibility and Accuracy by Ted Kemnitz Automated Machine Technologies, Inc. AMT (919) 361 0121 Ted.Kemnitz@AMTLiquidFilling.com
A Unique Approach to Managing the Problem of Antibiotic Resistance
 A Unique Approach to Managing the Problem of Antibiotic Resistance By: Heather Storteboom and Sung-Chul Kim Department of Civil and Environmental Engineering Colorado State University A Quick Review The
A Unique Approach to Managing the Problem of Antibiotic Resistance By: Heather Storteboom and Sung-Chul Kim Department of Civil and Environmental Engineering Colorado State University A Quick Review The
The OIE Relevant Standards and Guidelines for Vaccines
 The OIE Relevant Standards and Guidelines for Vaccines GALVMED/OIE STAKEHOLDER WORKSHOP ON THE HARMONISATION OF THE REGISTRATION OF VETERINARY MEDICINAL PRODUCTS, JOHANNESBURG, SOUTH AFRICA 9-11 MAY 2017
The OIE Relevant Standards and Guidelines for Vaccines GALVMED/OIE STAKEHOLDER WORKSHOP ON THE HARMONISATION OF THE REGISTRATION OF VETERINARY MEDICINAL PRODUCTS, JOHANNESBURG, SOUTH AFRICA 9-11 MAY 2017
Marissa Vignali*, Cate Speake* and Patrick E Duffy*
 Minireview Malaria sporozoite proteome leaves a trail Marissa Vignali*, Cate Speake* and Patrick E Duffy* Addresses: *Malaria Program, Seattle Biomedical Research Institute, Seattle, Washington 98109,
Minireview Malaria sporozoite proteome leaves a trail Marissa Vignali*, Cate Speake* and Patrick E Duffy* Addresses: *Malaria Program, Seattle Biomedical Research Institute, Seattle, Washington 98109,
Dynamic Drug Combination Response on Pathogenic Mutations of Staphylococcus aureus
 2011 International Conference on Biomedical Engineering and Technology IPCBEE vol.11 (2011) (2011) IACSIT Press, Singapore Dynamic Drug Combination Response on Pathogenic Mutations of Staphylococcus aureus
2011 International Conference on Biomedical Engineering and Technology IPCBEE vol.11 (2011) (2011) IACSIT Press, Singapore Dynamic Drug Combination Response on Pathogenic Mutations of Staphylococcus aureus
GUIDELINES ON CHOOSING THE CORRECT ERADICATION TECHNIQUE
 GUIDELINES ON CHOOSING THE CORRECT ERADICATION TECHNIQUE PURPOSE... 2 1. RODENTS... 2 1.1 METHOD PROS AND CONS... 3 1.1. COMPARISON BETWEEN BROUDIFACOUM AND DIPHACINONE... 4 1.2. DISCUSSION ON OTHER POSSIBLE
GUIDELINES ON CHOOSING THE CORRECT ERADICATION TECHNIQUE PURPOSE... 2 1. RODENTS... 2 1.1 METHOD PROS AND CONS... 3 1.1. COMPARISON BETWEEN BROUDIFACOUM AND DIPHACINONE... 4 1.2. DISCUSSION ON OTHER POSSIBLE
Differential Somatic Cell Count with the Fossomatic 7 DC - a novel parameter
 Differential Somatic Cell Count with the Fossomatic 7 DC - a novel parameter By: Dr. Daniel Schwarz, Cattle Disease Specialist, FOSS, Denmark Dedicated Analytical Solutions Somatic cell count (SCC) represents
Differential Somatic Cell Count with the Fossomatic 7 DC - a novel parameter By: Dr. Daniel Schwarz, Cattle Disease Specialist, FOSS, Denmark Dedicated Analytical Solutions Somatic cell count (SCC) represents
No-leaching. No-resistance. No-toxicity. >99.999% Introducing BIOGUARD. Best-in-class dressings for your infection control program
 Introducing BIOGUARD No-leaching. >99.999% No-resistance. No-toxicity. Just cost-efficient, broad-spectrum, rapid effectiveness you can rely on. Best-in-class dressings for your infection control program
Introducing BIOGUARD No-leaching. >99.999% No-resistance. No-toxicity. Just cost-efficient, broad-spectrum, rapid effectiveness you can rely on. Best-in-class dressings for your infection control program
Reproductive Vaccination- Deciphering the MLV impact on fertility
 Reproductive Vaccination- Deciphering the MLV impact on fertility Safety Decision Efficacy Prebreeding Vaccination of Cattle should Provide fetal & abortive protection (BVD and BoHV-1) Not impede reproduction
Reproductive Vaccination- Deciphering the MLV impact on fertility Safety Decision Efficacy Prebreeding Vaccination of Cattle should Provide fetal & abortive protection (BVD and BoHV-1) Not impede reproduction
Rapid molecular testing to detect Staphylococcus aureus in positive blood cultures improves patient management. Martin McHugh Clinical Scientist
 Rapid molecular testing to detect Staphylococcus aureus in positive blood cultures improves patient management Martin McHugh Clinical Scientist 1 Staphylococcal Bacteraemia SAB is an important burden on
Rapid molecular testing to detect Staphylococcus aureus in positive blood cultures improves patient management Martin McHugh Clinical Scientist 1 Staphylococcal Bacteraemia SAB is an important burden on
Consequences of Antimicrobial Resistant Bacteria. Antimicrobial Resistance. Molecular Genetics of Antimicrobial Resistance. Topics to be Covered
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Impact of Antimicrobial Resistance on Human Health. Robert Cunney HSE HCAI/AMR Programme and Temple Street Children s University Hospital
 Impact of Antimicrobial Resistance on Human Health Robert Cunney HSE HCAI/AMR Programme and Temple Street Children s University Hospital AMR in Foodchain Conference, UCD, Dec 2014 Sir Patrick Dun s Hospital
Impact of Antimicrobial Resistance on Human Health Robert Cunney HSE HCAI/AMR Programme and Temple Street Children s University Hospital AMR in Foodchain Conference, UCD, Dec 2014 Sir Patrick Dun s Hospital
MID 23. Antimicrobial Resistance. Consequences of Antimicrobial Resistant Bacteria. Molecular Genetics of Antimicrobial Resistance
 Antimicrobial Resistance Molecular Genetics of Antimicrobial Resistance Micro evolutionary change - point mutations Beta-lactamase mutation extends spectrum of the enzyme rpob gene (RNA polymerase) mutation
Antimicrobial Resistance Molecular Genetics of Antimicrobial Resistance Micro evolutionary change - point mutations Beta-lactamase mutation extends spectrum of the enzyme rpob gene (RNA polymerase) mutation
Selection for Egg Mass in the Domestic Fowl. 1. Response to Selection
 Selection for Egg Mass in the Domestic Fowl. 1. Response to Selection H. L. MARKS US Department of Agriculture, Science & Education Administration, Agricultural Research, uthern Regional Poultry Breeding
Selection for Egg Mass in the Domestic Fowl. 1. Response to Selection H. L. MARKS US Department of Agriculture, Science & Education Administration, Agricultural Research, uthern Regional Poultry Breeding
Antimicrobial Resistance
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Acquisition of Foreign DNA
 Antimicrobial Resistance Acquisition of Foreign DNA Levy, Scientific American Horizontal gene transfer is common, even between Gram positive and negative bacteria Plasmid - transfer of single or multiple
Antimicrobial Resistance Acquisition of Foreign DNA Levy, Scientific American Horizontal gene transfer is common, even between Gram positive and negative bacteria Plasmid - transfer of single or multiple
CIRCUMSPOROZOITE PROTEINS OF HUMAN MALARIA PARASITES PLASMODIUM FALCIPARUM AND PLASMODIUM VIVA,F*
 CIRCUMSPOROZOITE PROTEINS OF HUMAN MALARIA PARASITES PLASMODIUM FALCIPARUM AND PLASMODIUM VIVA,F* BY ELIZABETH H. NARDIN, VICTOR NUSSENZWEIG, RUTH S. NUSSENZWEIG, WILLIAM E. COLLINS, K. TRANAKCHIT HARINASUTA,
CIRCUMSPOROZOITE PROTEINS OF HUMAN MALARIA PARASITES PLASMODIUM FALCIPARUM AND PLASMODIUM VIVA,F* BY ELIZABETH H. NARDIN, VICTOR NUSSENZWEIG, RUTH S. NUSSENZWEIG, WILLIAM E. COLLINS, K. TRANAKCHIT HARINASUTA,
Evolution in Action: Graphing and Statistics
 Evolution in Action: Graphing and Statistics OVERVIEW This activity serves as a supplement to the film The Origin of Species: The Beak of the Finch and provides students with the opportunity to develop
Evolution in Action: Graphing and Statistics OVERVIEW This activity serves as a supplement to the film The Origin of Species: The Beak of the Finch and provides students with the opportunity to develop
INFECTIOUS HEPATITIS, PARVOVIRUS & DISTEMPER
 Canine VacciCheck INFECTIOUS HEPATITIS, PARVOVIRUS & DISTEMPER IgG ANTIBODY TEST KIT INSTRUCTION MANUAL Sufficient for 12/120 assays 13 JUL 2015 Biogal Galed Laboratories Acs. Ltd., tel: 972-4-9898605.
Canine VacciCheck INFECTIOUS HEPATITIS, PARVOVIRUS & DISTEMPER IgG ANTIBODY TEST KIT INSTRUCTION MANUAL Sufficient for 12/120 assays 13 JUL 2015 Biogal Galed Laboratories Acs. Ltd., tel: 972-4-9898605.
Drug Discovery: Supporting development of new drugs to treat global parasitic diseases
 Drug Discovery: Supporting development of new drugs to treat global parasitic diseases UC Santa Cruz Bio 117 Feb. 23, 2016 Judy Sakanari Center for Parasitic Diseases UC San Francisco Parasitic Diseases,
Drug Discovery: Supporting development of new drugs to treat global parasitic diseases UC Santa Cruz Bio 117 Feb. 23, 2016 Judy Sakanari Center for Parasitic Diseases UC San Francisco Parasitic Diseases,
POST SCREENING METHODS FOR THE DETECTION OF BETA-LACTAM RESIDUES IN PIGS.
 POST SCREENING METHODS FOR THE DETECTION OF BETA-LACTAM RESIDUES IN PIGS. Lorraine Lynas, Deborah Currie and John D.G. McEvoy. Department of Agriculture and Rural Development for Northern Ireland, Veterinary
POST SCREENING METHODS FOR THE DETECTION OF BETA-LACTAM RESIDUES IN PIGS. Lorraine Lynas, Deborah Currie and John D.G. McEvoy. Department of Agriculture and Rural Development for Northern Ireland, Veterinary
Pain management: making the most of the latest options
 Vet Times The website for the veterinary profession https://www.vettimes.co.uk Pain management: making the most of the latest options Author : James Westgate Categories : Business, Business planning Date
Vet Times The website for the veterinary profession https://www.vettimes.co.uk Pain management: making the most of the latest options Author : James Westgate Categories : Business, Business planning Date
Effects of Cage Stocking Density on Feeding Behaviors of Group-Housed Laying Hens
 AS 651 ASL R2018 2005 Effects of Cage Stocking Density on Feeding Behaviors of Group-Housed Laying Hens R. N. Cook Iowa State University Hongwei Xin Iowa State University, hxin@iastate.edu Recommended
AS 651 ASL R2018 2005 Effects of Cage Stocking Density on Feeding Behaviors of Group-Housed Laying Hens R. N. Cook Iowa State University Hongwei Xin Iowa State University, hxin@iastate.edu Recommended
Developmentally Regulated!nfectivity of Malaria Sporozoites for Mosquito Salivary Glands and the Vertebrate Host
 Developmentally Regulated!nfectivity of Malaria Sporozoites for Mosquito Salivary Glands and the Vertebrate Host By Musa G. Touray, Alon Warburg, Andre Laughinghouse, Antoniana U. Krettli,* and Louis H.
Developmentally Regulated!nfectivity of Malaria Sporozoites for Mosquito Salivary Glands and the Vertebrate Host By Musa G. Touray, Alon Warburg, Andre Laughinghouse, Antoniana U. Krettli,* and Louis H.
Human-Animal Interactions in the Turkey Industry
 Human-Animal Interactions in the Turkey Industry Dr. Naomi A. Botheras 1, Ms. Jessica A. Pempek 2, Mr. Drew K. Enigk 2 1 PI, 222E Animal Sciences Building, 2029 Fyffe Court, Columbus, OH 43210 (614) 292-3776;
Human-Animal Interactions in the Turkey Industry Dr. Naomi A. Botheras 1, Ms. Jessica A. Pempek 2, Mr. Drew K. Enigk 2 1 PI, 222E Animal Sciences Building, 2029 Fyffe Court, Columbus, OH 43210 (614) 292-3776;
Detection of inducible clindamycin resistance among clinical isolates of Staphylococcus aureus in a tertiary care hospital
 ISSN: 2319-7706 Volume 3 Number 9 (2014) pp. 689-694 http://www.ijcmas.com Original Research Article Detection of inducible clindamycin resistance among clinical isolates of Staphylococcus aureus in a
ISSN: 2319-7706 Volume 3 Number 9 (2014) pp. 689-694 http://www.ijcmas.com Original Research Article Detection of inducible clindamycin resistance among clinical isolates of Staphylococcus aureus in a
An LC-MS/MS method to determine antibiotic residues in distillers grains
 An LC-MS/MS method to determine antibiotic residues in distillers grains Hemakanthi de Alwis FDA Center for Veterinary Medicine Office of Research 07-31-2018 Distillers grain (DG) q DG is a major co-product
An LC-MS/MS method to determine antibiotic residues in distillers grains Hemakanthi de Alwis FDA Center for Veterinary Medicine Office of Research 07-31-2018 Distillers grain (DG) q DG is a major co-product
The Friends of Nachusa Grasslands 2016 Scientific Research Project Grant Report Due June 30, 2017
 The Friends of Nachusa Grasslands 2016 Scientific Research Project Grant Report Due June 30, 2017 Name: Laura Adamovicz Address: 2001 S Lincoln Ave, Urbana, IL 61802 Phone: 217-333-8056 2016 grant amount:
The Friends of Nachusa Grasslands 2016 Scientific Research Project Grant Report Due June 30, 2017 Name: Laura Adamovicz Address: 2001 S Lincoln Ave, Urbana, IL 61802 Phone: 217-333-8056 2016 grant amount:
Tick-Borne Disease Research Program
 Tick-Borne Disease Research Program Strategic Plan INTRODUCTION The Congressionally Directed Medical Research Programs (CDMRP) represents a unique partnership among the U.S. Congress, the military, and
Tick-Borne Disease Research Program Strategic Plan INTRODUCTION The Congressionally Directed Medical Research Programs (CDMRP) represents a unique partnership among the U.S. Congress, the military, and
VETERINARY MEDICINAL PRODUCTS CONTROLLING VARROA JACOBSONI AND ACARAPIS WOODI PARASITOSIS IN BEES
 VETERINARY MEDICINAL PRODUCTS CONTROLLING VARROA JACOBSONI AND ACARAPIS WOODI PARASITOSIS IN BEES Guideline Title Veterinary Medicinal Products controlling Varroa jacobsoni and Acarapis woodi parasitosis
VETERINARY MEDICINAL PRODUCTS CONTROLLING VARROA JACOBSONI AND ACARAPIS WOODI PARASITOSIS IN BEES Guideline Title Veterinary Medicinal Products controlling Varroa jacobsoni and Acarapis woodi parasitosis
WHY IS THIS IMPORTANT?
 CHAPTER 20 ANTIBIOTIC RESISTANCE WHY IS THIS IMPORTANT? The most important problem associated with infectious disease today is the rapid development of resistance to antibiotics It will force us to change
CHAPTER 20 ANTIBIOTIC RESISTANCE WHY IS THIS IMPORTANT? The most important problem associated with infectious disease today is the rapid development of resistance to antibiotics It will force us to change
FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT
![FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT](/thumbs/93/113164093.jpg "FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT") FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT INSTRUCTION MANUAL Sufficient for 12/120 assays 22 APR 2018 Biogal Galed Laboratories Acs Ltd. tel: 972-4-9898605. fax: 972-4-9898690 e-mail:info@biogal.co.il
FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT INSTRUCTION MANUAL Sufficient for 12/120 assays 22 APR 2018 Biogal Galed Laboratories Acs Ltd. tel: 972-4-9898605. fax: 972-4-9898690 e-mail:info@biogal.co.il
American Association of Feline Practitioners American Animal Hospital Association
 American Association of Feline Practitioners American Animal Hospital Association Basic Guidelines of Judicious Therapeutic Use of Antimicrobials August 1, 2006 Introduction The Basic Guidelines to Judicious
American Association of Feline Practitioners American Animal Hospital Association Basic Guidelines of Judicious Therapeutic Use of Antimicrobials August 1, 2006 Introduction The Basic Guidelines to Judicious
Plasmodium sporozoites acquire virulence and. immunogenicity during mosquito hemocoel transit
 IAI Accepts, published online ahead of print on 30 December 2013 Infect. Immun. doi:10.1128/iai.00758-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 Plasmodium sporozoites
IAI Accepts, published online ahead of print on 30 December 2013 Infect. Immun. doi:10.1128/iai.00758-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 Plasmodium sporozoites
ETX0282, a Novel Oral Agent Against Multidrug-Resistant Enterobacteriaceae
 ETX0282, a Novel Oral Agent Against Multidrug-Resistant Enterobacteriaceae Thomas Durand-Réville 02 June 2017 - ASM Microbe 2017 (Session #113) Disclosures Thomas Durand-Réville: Full-time Employee; Self;
ETX0282, a Novel Oral Agent Against Multidrug-Resistant Enterobacteriaceae Thomas Durand-Réville 02 June 2017 - ASM Microbe 2017 (Session #113) Disclosures Thomas Durand-Réville: Full-time Employee; Self;
Sporozoae: Plasmodium.
 Sporozoae: Plasmodium. Coccidian. Asexual division in Man (Schizogony), sexual division in the mosquito (sporogony). Plasmodium vivax > P. falciparum > P. malariae > P. ovale as a cause of malaria. P.
Sporozoae: Plasmodium. Coccidian. Asexual division in Man (Schizogony), sexual division in the mosquito (sporogony). Plasmodium vivax > P. falciparum > P. malariae > P. ovale as a cause of malaria. P.