THE CENTRAL CONNEXIONS OF DORSAL SPIN [l NERVE ROOTS AND THE ASCENDING TRACT IN THE SPINAL CORD OF LACERTA VIRIDIS
|
|
|
- Camilla Rogers
- 5 years ago
- Views:
Transcription

1 J. Anat., Lond. (1962), 96, 2, pp With 2 plates and 2 text-figures Printed in Great Britain THE CENTRAL CONNEXIONS OF DORSAL SPIN [l NERVE ROOTS AND THE ASCENDING TRACT IN THE SPINAL CORD OF LACERTA VIRIDIS By F. GOLDBY AND L. R. ROBINSON* Department of Anatomy, St Mary's Hospital Medical School, Paddington, London W. 2 Until recently our knowledge of the detailed structure of the reptilian nervous system has been based almost entirely on observations made on serial sections of normal material prepared by routine neuro-histological methods. Most of these observations were reported in the older literature and were ably summarised by Kappers, Huber & Crosby, in In spite of the well-known limitations of the methods a considerable body of knowledge has been built up. Its interpretation, however, in terms of connexions between different parts of the nervous system, has depended in many cases on a presumed homology with tracts or collections of nerve cells similarly situated in the mammalian nervous system, where they have been subjected to more rigorous investigation by experimental methods. Recently, several papers have appeared in which the results of the differential staining of degenerating fibres or cells have been described in the reptilian brain. The degeneration has been caused by lesions so placed that it should demonstrate the connexions of particular nerves or parts of the brain, e.g. Armstrong (1950) for optic connexions, Gamble (1952) for olfactory connexions, and Powell & Kruger (1960) for striato-thalamic connexions. So far this work has been limited to the foreand mid-brain, where it has demonstrated many fundamental differences between reptilian and mammalian anatomy which could not have been demonstrated conclusively in preparations of normal material. The present investigation extends observations of this kind to lower levels of the nervous system and its main purpose is to determine the central connexions of the dorsal roots of spinal nerves and the course and destination of long ascending tracts in the spinal cord. MATERIALS AND METHODS The animals were all specimens of Lacerta viridis, between 8 and 10 cm. in length, excluding the tail. Operations were performed with aseptic precautions under ether anaesthesia. The spinal cord was exposed by the removal of the laminae (together with the dura, which was adherent to the bone) from two or three vertebrae. In the root section experiments three successive dorsal spinal roots on the same side were cut as far from the cord as possible but central to the ganglia; slight direct damage to the spinal cord, or damage due to traction on the roots, was not always avoided, but was checked microscopically and allowed for in assessing the results. Three animals, in which the spinal cord and dorsal roots were exposed but no lesions * On leave from the University of Otago, New Zealand. 11 Anat. 96

2 154 F. Goldby and L. R. Robinson made, were used as control specimens. The spinal cord lesions were all attempted hemisections either of the right or left side or of the dorsal half of the cord, or complete transverse sections, and were made by cutting. In nearly all cases the hemisections were found to be more extensive than expected and to have divided between i and i of the cord. The animals were kept in cages maintained at 250 to 320 C., temperatures at which degeneration is known to progress at a rate comparable with that in mammals (Gamble, Goldby & Smith, 1957). No specimen showed any obvious signs of infection but there was some operative mortality. Of thirty-three hemisections and spinal root sections, two died immediately post-operatively and five before the 11th day; all the others appeared healthy and were killed at the time which had been arranged. In one case early death seemed due to an intercurrent infection which led to abscess formation in the liver and probably had no relation to the operation. Occasional deaths from a similar cause occurred in the stock animals. The mortality from complete transactions was much heavier. This operation was attempted on only three animals, two of which died within the first 4 days and were useless for histological purposes. The survivor was killed on the 8th day, when it still appeared healthy. It was found that axonal degeneration could be demonstrated in paraffin sections by the Holmes silver impregnation technique (Holmes, 1947) as modified by Powell, Guillery & Cowan (1957). Clearer results were obtained with a modification of the technique of Nauta & Gygax (1951) introduced by Guillery, Shirra & Webster (1961) and applicable to paraffin sections. The small size of the blocks made it possible to work with complete series of sections in all cases. All specimens were fixed by perfusion after washing out with 065 % saline. The spinal cord and brain were then exposed and the specimens stored in the fixative. For the Holmes technique either 10% neutral formalin or 70 % alcohol containing 2 % of acetic acid was used. With the formalin fixed specimens there was considerable background staining, particularly in degenerated areas, and normal fibres were frequently distorted and irregular in calibre. With acid alcohol the distortion of normal fibres and the background staining were both much less. For the modified Nauta technique neutral formalin was the fixative in all cases. Although the results varied, this did not appear to depend on the length of time in the fixative. Entirely satisfactory impregnations were obtained from specimens which had been stored in fixative for no more than a few days and from others which had been stored for as long as 6 months. The results obtained by this method form the principal basis of the findings to be reported; in addition a number of series stained by toluidine blue were available and were used to study the normal structure of the spinal cord and brain stem. The following experimental specimens were used for the work to be reported: I. Spinal root lesions (all unilateral) Specimen Roots cut Days Method no. survival 23 27th-29th 23 Modified Nauta 25 25th-27th 22 Modified Nauta 38 7th-9th 35 Holmes and modified Nauta 34 8th-lOth 57 Modified Nauta

3 Dorsal spinal nerve roots and spinal cord of Lacerta viridis 155 II. Hemisections of spinal cord (right or left) Specimen Segmental level Days Method no. survival 7 Between 10th and 11th 21 Holmes and modified Nauta 10 Between 13th and 14th 15 Holmes and modified Nauta 22 27th 24 Holmes and modified Nauta 36 Between 11th and 12th 73 Modified Nauta 42 28th 25 Modified Nauta 45 Between 10th and 11th 28 Modified Nauta III. Section of dorsal half of spinal cord Specimen Segmental level Days Method no. survival 18 Between 9th and 10th 21 Holmes and modified Nauta 19 Between 14th and 15th 38 Holmes and modified Nauta Only one specimen with a complete transaction of the spinal cord gave satisfactory histological results. This was a transaction between the 12th and 13th segments, and the animal was killed on the 8th day. Sections were prepared by both the Holmes and the modified Nauta method. It should be noted that the nerve roots or segments of the spinal cord are numbered cranio-caudally and no attempt is made to specify cervical, dorsal, lumbar or sacral regions. The enlargement of the cord opposite the forelimb extends from about the 5th to the 10th and opposite the hindlimb from about the 25th to the 30th segment. The nature and criteria of degeneration In Holmes preparations, after a survival period of 21 days, degenerating coarse fibres (4-8g in diameter) showed very obvious irregularities in calibre, vacuolation and fragmentation. In transverse sections many spaces were seen from which such fibres had disappeared. Fine fibres (21s or less in diameter) disappeared more rapidly leaving behind a little debris which was only faintly argentophilic. The disappearance of fine fibres could be recognized with confidence only when most of a compact group were affected together (PI. 1, fig. 2), but at earlier stages (15 days after operation) fusiform swellings and fragmentation were seen in individual fibres (PI. 1, fig. 7). In general, the rather pale impregnation of the Holmes preparations made the identification of degenerative changes difficult except in very coarse fibres, so that after the first few experiments the method was used chiefly for studying normal structure or to confirm results obtained by the modified Nauta technique. Preterminal or terminal degeneration was not recognized in the Holmes preparations. The modified Nauta technique showed degeneration debris as intensely argentophilic granules of very irregular shape and size distributed along the course of fibres (e.g. PI. 1, fig. 9). Adjacent normal fibres usually stained a golden brown colour, but in a few cases their staining was suppressed entirely. It was common, however, to find at least some normal fibres (usually fine) densely impregnated and showing minor irregularities of calibre, a feature also seen in control specimens. It followed that argentophilia and irregularity of calibre alone could not be accepted as criteria 11-2

4 156 F. Goldby and L. R. Robinson of degeneration. It was necessary also to see fragmentation with irregular granules arranged in a linear fashion. Usually this was possible only in longitudinal sections, although occasionally, when the degeneration was massive and the staining of normal fibres more completely suppressed than in most of our specimens, transverse sections gave convincing results, very similar to those obtained in a good Marchi preparation (PI. 1, fig. 3). Because the stained fibres had a tubular appearance in some of our preparations and stained debris was still visible after 73 days, it seemed possible that it was the myelin sheath debris rather than that of the axon which we were examining in these specimens. The earliest degeneration seen in the central nervous system was in an 8-day specimen. Although in this specimen the spinal cord had been completely transacted, only occasional fragmented fibres were present, and these did not show the intense argentophilia seen after longer periods. A survival period between 20 and 40 days was optimal under the conditions of our experiments. In the 73-day specimen the degeneration debris was less abundant and more widely scattered as if active absorption were in progress. We noticed, however, that after spinal root section all fibres and debris disappeared from the nerve root close to its entry in the spinal cord at an early stage (21 days) so that in this situation (outside the central nervous system) the degeneration process was much more rapid. What was interpreted as 'pre-terminal' degeneration was seen as the fragmentation of fine fibres into very small argentophilic granules, usually linearly arranged and often close to perikarya, in the grey matter. Where this had occurred the surrounding neuropil was generally paler than in control specimens, e.g. PI. 1, figs. 5, 6. In all the posterior root sections the degeneration (except for some pre-terminal changes in the ventral horns) was entirely unilateral and comparison with the opposite side in the same section provided a very satisfactory control, e.g. PI. 1, fig. 10. The spinal cord lesions were never confined strictly to one side and always led to some bilateral degeneration. Such specimens were controlled by comparison with sections from the same region of the cord in a normal animal. In three cases the latter were mounted for processing on the same slide with sections from the experimental animal, by far the most satisfactory form of control for bilateral effects. With long fibre tracts, consistency of results in several blocks from widely separated parts of the cord gave added confidence to the interpretation in many cases. It must be remembered, however, that interpretation depends always on the assumption that the degeneration observed is the result of the lesion. It is known that in mammals the mere exposure of the central nervous system by removal of the dura after laminectomy (Peele & Windle, 1946) or craniotomy (Harris, 1960) can cause degeneration which may be severe. This may also occur in reptiles, for in a lizard with a simple exposure of the spinal cord in the 12th and 13th segments which survived for 27 days, almost complete disorganization of the spinal cord was found at the site of exposure. The result was like that of an almost complete division of the spinal cord. This was an isolated case however. In two similarly treated specimens only occasional degenerated fibres could be found; they were limited to the segments close to the exposure, and showed no tendency to concentration in one region or tract rather than another. Gross disorganization of the spinal cord was seen in none of the experimental specimens with dorsal root lesions, and the slight

5 Dorsal spina nerve roots and spinal cord of Lacerta viridis 157 and non-specific degeneration which may have been present as a result of exposure only could not have accounted for the findings or have led to any serious difficulty in interpretation. It was necessary only to ignore occasional and isolated fibres showing degeneration near the lesion, particularly when they could not be found consistently in particular regions or traced in more than one block of tissue. There remains only the possibility of ischaemic degeneration which could follow the interruption of radicular arteries. No evidence which would suggest such an occurrence was seen in any of our specimens, unless a small extravasation of blood, in the ventral horn of lizard 34 was due to this cause. In the specimens with spinal cord lesions any additional damage caused by exposure or by interference with the blood supply was of little importance. The lesions were always checked histologically in serial sections and allowance was made for the fact that an intended hemisection had usually damaged considerably more than half the spinal cord. In one case, however (lizard 22), more extensive degeneration was found throughout the central nervous system then in any other specimen with a comparable lesion and some of this may have been due to a generalized pathological condition rather than to the lesion directly. In two others (lizard 23 and 34) apparent degeneration was seen in the habenular commissure of the forebrain which it seemed impossible to attribute to the experimental lesion (spinal root section). The significance of occasional observations of this kind which cannot be confirmed in all specimens must remain obscure. The principal interpretations which we report are based only on observations which could be made consistently in all relevant and well impregnated specimens; in a few cases where the basis of interpretation was less secure, this has been pointed out. The dorsal roots and spinal cord in normal material Kappers et al. (1936) reviewed the results obtained by the earlier workers and little has since been added to our knowledge of the nerve roots and spinal cord in reptiles. It is unnecessary therefore to do more than draw attention to some features which are characteristic of Lacerta viridis or particularly relevant to the present investigation. The axons which form the dorsal roots are of varying calibre; many are fine (in the Holmes preparations 1iu or less in diameter) although the most conspicuous component consists of coarser axons, about 2-3# in diameter. For each segment they are arranged in a single flattened bundle, which runs dorsally in close apposition with the lateral column of the spinal cord, and, for the last part of its course, deep to the pia mater (PI. 1, fig. 4). It is probably for this reason that it is difficult to cut the roots without causing some direct damage to fibres in the lateral columns. No clear cut segregation of fine from coarse fibres could be detected as the root enters the spinal cord. Many fibres, both fine and coarse, can be followed into the posterior columns. Fibres which are predominantly fine enter a marginal tract, superficial to the dorsal horn, which resembles Lissauer's tract in mammals, and a few (both coarse and fine) enter the lateral columns as has been described in turtles, lizards and snakes (see Kappers et al. 1936, pp ). The fibres which enter the lateral columns appear to run close to the grey matter towards the ventral horn. In the dorsal columns

6 158 F. Goldby and L. R. Robinson fibres bifurcate into ascending and descending branches from which collaterals pass to the grey matter but beyond this no certain information about the termination of dorsal root fibres could be obtained from the normal material. Coarse efferent fibres which arise from the large cells of von Lenhossdk in the medial part of the ventral horn have been described in the cervical dorsal roots of L. murals (Beccari, 1913, 1914). They may represent the spinal roots of the accessory nerve of mammals. What appeared to be similar fibres were seen in a 2nd cervical dorsal root in L. viridis but their origin and destination were not investigated experimentally. The roots which are attached to the limb plexuses are obviously larger than in other regions, but no fibre counts were made. In reptiles generally, and this is certainly true of L. viridis, the distinction between grey and white matter is less definite than in mammals, although the same main subdivisions (apart from a lateral horn) can be recognized. Large dendrites extend from the grey matter into the anterior and lateral columns, dividing them into fasciculi and ending in a superficial or marginal dendritic plexus where the cells of the marginal (Hofmann's) nuclei are situated (see Terni, 1926). The dorsal columns are well developed at all levels and share in the general enlargements of the cord opposite the limb plexuses. Measurements of cross-sectional area showed no significant increase in the more cranial segments and failed to provide evidence for the frontal accummulation of fibres which is said to be 'clearly evident in reptiles' (Kappers et al. 1936). This is not to say, however, that a frontal accummulation of long ascending fibres does not occur in L. viridis. The gross size of the dorsal column is to a large extent dependent on the abundance or otherwise of descending collaterals and intersegmental Ii br.,s and variations in the number of such fibres at different levels might mask the effects of additions from each segment of a few long ascending fibres. Most of the axons of the dorsal columns are fine, particularly those on the surface and adjacent to the dorsomedian septum (P1. 1, fig. 1). A proportion of coarser axons is present in the deeper parts of the columns. The proportion of coarser axons increases in the lateral and becomes very large in the ventral columns where the coarsest of all (6-8/i in diameter) are found. In some sections this results in the white matter of the ventral half of the cord having a strikingly different appearance from the dorsal half. The deeper part of the ventral columns, immediately ventral to the central canal, consists of particularly large fibres which are cut off from the rest of the columns by an accessory grey commissure to form two conspicuous bundles. They can be traced into continuity with the medial longitudinal bundles of the brain stem and were studied very thoroughly by Terni (1922) in a lizard (Gongylus ocellatus) and later by Leghissa (1954) in a series of reptiles. Terni thought they were largely ascending tracts, formed by axons which decussate below the central canal. Leghissa described a descending component arising in large part from the brain-stem, and Stefanelli (1941) considered they were concerned with movements of the trunk. The further division of the white matter into tracts is largely hypothetical and chiefly based on observations made on normal material from the brain-stem and the junctional region between the medulla oblongata and spinal cord. Only the supposed
 are also present and a relay could occur into a medial lemniscus which has been identified by some authors but has not been traced further than")
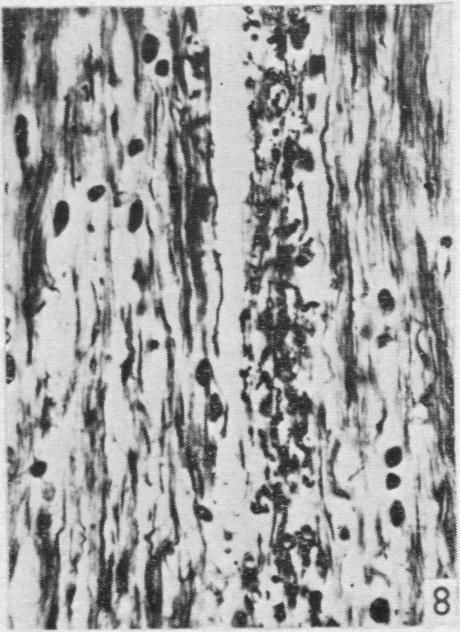
7 Dorsal spinal nerve roots and spinal cord of Lacerta viridis 159 ascending tracts will be considered here. The dorsal columns are said to contain cuneate and gracile fasciculi. Rudimentary dorsal column nuclei (Zeehandelaar, 1920) are also present and a relay could occur into a medial lemniscus which has been identified by some authors but has not been traced further than the mid-brain (Kappers et al. 1936). In L. viridis the dorsal column nuclei are poorly developed. They are represented by some small multipolar cells scattered in the deeper part of the dorsal columns in the upper cervical segments, which can be traced into the medulla. In Nissl preparations they show no definite arrangement in nuclear groups. In fibre preparations dendrites from these cells stream into the dorsal columns in two regions, medially close to the dorso-median septum and again laterally, producing an appearance very like that of the cuneate and gracile nuclei in mammals. Larsell (1926, 1932) and others have described large ventral and smaller dorsal spino-cerebellar tracts the presence of which Larsell (1932) tried to confirm (but without much success) by using the Marchi technique after spinal cord lesions in the cervical region. A secondary sensory tract (spino-tectal or spino-mesencephalic) from the lateral columns has been described (Edinger, 1908), and it is certainly true that fibres from the lateral columns can be traced into the reticular formation of the brain stem and possibly as high as the midbrain. There is no evidence to show whether these are ascending or descending, as indeed is the case with most of the tracts described in the reptilian spinal cord. Apart from Larsell's (1932) and Ingvar's (1918) disappointing attempts with the Marchi technique, we have found no descriptions in the literature of studies of degenerating fibres after experimental lesions in the reptilian spinal cord. Degeneration following dorsal spinal nerve root section Section of three successive dorsal roots connected either to the fore- or hindlimb plexus caused little functional disturbance. The limbs behaved normally and in a perfectly coordinated manner in walking, but the affected limb was not withdrawn when stimulated and the claws did not grip as effectively as in the normal limbs. The degeneration can be illustrated by two specimens (lizards 23 and 34). In lizard 23 blocks were examined at the level of the lesion (27th, 28th and 29th segments), cranial to the lesion at the 4th and 5th segments and in the brain stem, but not caudal to the lesion. All sections were stained by the modified Nauta method. Section of the roots was found to be accompanied by superficial damage to the lateral column on the same side, there being some loss of tissue and cellular reaction in this situation. In the dorsal columns degeneration was confined to the side of the lesion. Close to the lesion it was scattered throughout the column, affecting both fine and coarse fibres, but a fair number of apparently normal fibres were interspersed among those which were fragmented and intensely argentophilic. In higher segments (4th and 5th) the degenerated fibres were concentrated in a crescentic area adjacent to the dorsomedian septum (PI. 1, fig. 8) and extending on to the dorsal surface of the cord about half way to the line of entry of the dorsal nerve roots. At this level degeneration was confined to fine fibres. Similar degeneration was found at the caudal end of the medulla and could be traced for a short distance cranially. It became diffuse in a region lateral to the lower extremity of the 4th ventricle and

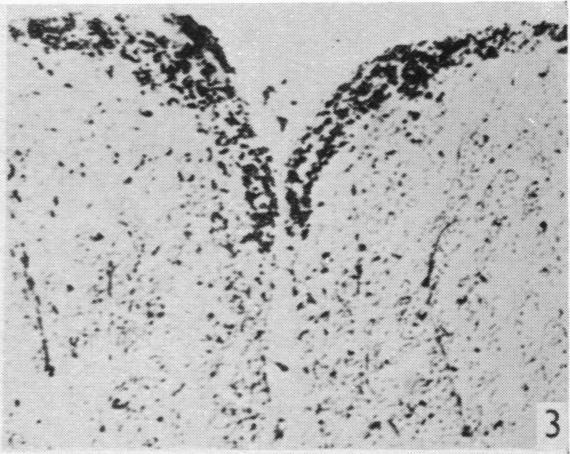
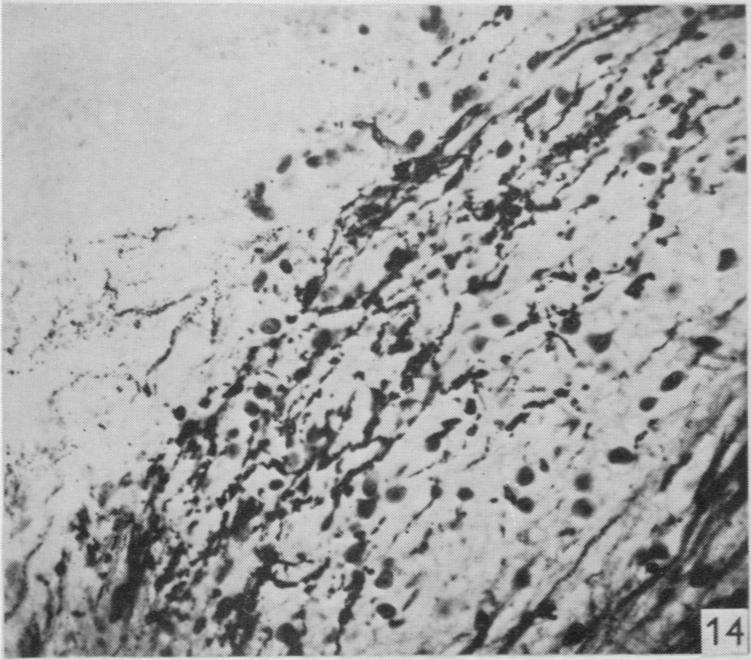
8 160 F. Goldby and L. R. Robinson could not be followed further (Text-fig. 1 and PI. 2, fig. 14). The scattering of debris suggested degeneration in terminal or preterminal fibres and since few perikarya were present in this situation any connexions established were presumably axodendritic. The fact that the degenerated tract occupies a far more restricted region of the dorsal column in the more cranial segments suggests that more ascending fibres are added from the roots cranial to those divided. Degeneration was also seen in the superficial part of the lateral column close to the dorsal horn, and could be followed into the brain stem. It is almost certain that this was due to the direct damage to the lateral column already mentioned, and not to the root section. In another specimen (lizard 25) where lateral column Vill Fig. 1 Fig. 2 Text-fig. 1. A reconstruction of the dorsal aspect of the medulla in lizard 23 made by superimposing the outlines of horizontal sections. The position of the degenerated fibres is shown in stipple. The section which is cross-hatched is the one illustrated in PI. 2, fig. 14. Text-fig. 2. A reconstruction ofthe dorsal aspect ofthe medulla in lizard 34 made by superimposing the outlines of horizontal sections. The position of the degenerated fibres is shown in stipple. The section which is cross-hatched is the one illustrated in PI. 2, fig. 15. damage was less extensive, it was less marked and not recognizable in the brain stem. In two further specimens (lizards 83 and 34) in which roots attached to the forelimb enlargement had been cut, but without any apparent damage to the spinal cord, only occasional degenerated fibres were found in the superficial part of the lateral column or brain stem; on the other hand, lateral column degeneration was always abundant after spinal cord lesions. It is reasonable to conclude that this degeneration is not related to the root section and its nature and extent will be described later.

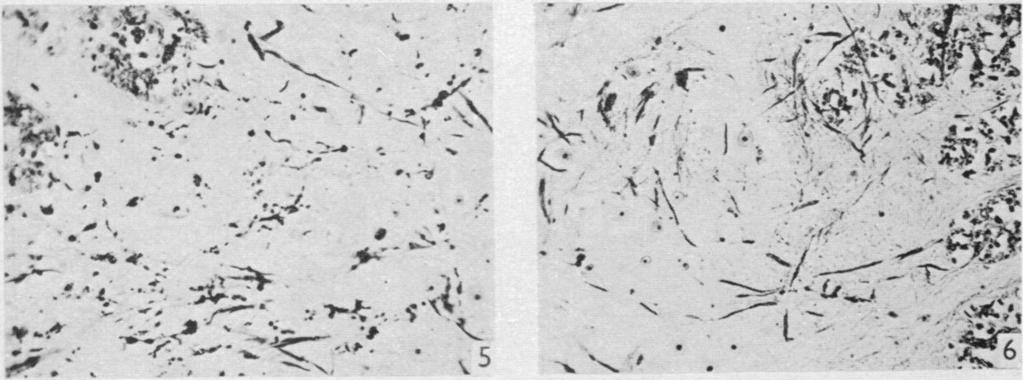
9 Dorsal spinal nerve roots and spinal cord of Lacerta viridis 161 The only additional degeneration seen in lizard 23 was in segments close to the lesion. From the dorsal column fine degenerated fibres, possibly collaterals, could be followed into the base of the dorsal horn of grey matter, and what appeared to be terminal or pre-terminal degeneration was concentrated towards the lateral side of the horn where the fibres seemed to have a predominantly longitudinal orientation. This degeneration was limited to the side of the lesion. Degeneration was also seen around large cells in the ventral horn; this was bilateral, but more marked on the side of the lesion, where some of the adjacent lateral column fibres were also degenerate. The ventral horn degeneration was particularly well shown in a second specimen, lizard 33 (PI. 1, figs. 5, 6). The interpretation of terminal degeneration in the grey matter near the lesion is made difficult by the existence of lateral column damage. It was present, however, in all specimens with root sections and there is a strong probability that it represents degeneration in the terminations of root fibres or their collaterals. Degeneration of this kind was not present in the 4th and 5th segments, a long way cranial to the lesion. In lizard 34, the roots divided were connected to the brachial plexus. Immediately cranial to the lesion (7th segment) degenerated fibres were widespread throughout the ipsilateral dorsal column except in the crescentic region occupied by the tract described in lizard 23, where only fine normal fibres were present. There was no difficulty in tracing the degeneration to a region of the medulla which lay lateral to the region where the degeneration in lizard 23 appeared to end, and extended a little further cranially (Text-fig. 2; PI. 2, fig. 15). Again few perikarya were found at the site of termination so that any connexions were presumably axodendritic. It will be noticed in the reconstructions (Text-figs. 1 and 2) that the two degenerated tracts described in lizards 23 and 34 respectively appear to occupy the same position in the dorsal columns. This results from the fact that both are flattened bundles which lie obliquely, superficial and deep respectively, so that in orthogonal projection they overlap almost completely. The distribution of degeneration in the grey matter of the spinal cord was very similar to that found in the previous specimen (lizard 23). It is illustrated in PI. 1, fig. 10, in collaterals from the dorsal columns and in PI. 1, fig. 11, as preterminal degeneration in the grey matter at the root of the dorsal horn. It was present in sections 4 segments cranial and 5 segments caudal to the lesion, and could be traced into the ventral horns from a few degenerate fibres deep in the lateral columns. In the superficial part of the lateral column only an occasional degenerate fibre could be seen, and, as already stated, there was no obvious direct damage to the spinal cord in this situation. It is possible, however, that the degeneration in the grey matter was increased by the presence of a small extravasation of blood which appeared to have occurred ante mortem near the base of the ventral horn in the 9th segment, but there is little doubt that most if not all was due to preterminal degeneration in the fibres of the dorsal roots which had been cut. In lizard 34, segment 15, five segments caudal to the lesion, was also examined. It showed a few degenerate fibres deep in the dorsal column on the side of the lesion, probably descending collaterals from ascending axons. There was also very slight evidence of degeneration in some of the deeper fibres of the lateral column on the side of the lesion and in collateral branches to the ventral horn. Traces of terminal
 gave similar results and do not require separate description.")
10 162 F. Goldby and L. R. Robinson degeneration in both ventral horns and in the root of the ipsilateral posterior horn were also seen. Two further specimens (lizards 33 and 25) gave similar results and do not require separate description. Degeneration following spinal cord lesions Most of the spinal cord lesions were between the 11th and 15th segments, i.e. just caudal to the enlargement of the spinal cord opposite the forelimb. Hemisection at this level had no apparent effect on movements of progression in most specimens. In one, where subsequent examination showed continuity of nerve fibres only in the lateral column on one side, and in two, where only the superficial parts of both anterior columns remained intact, no regular movements of the hind-limbs occurred in walking. Uncoordinated movements were sometimes seen however. In the two animals with hemisections in the enlargement opposite the hindlimbs, the only obvious change was relative inactivity of the limb on the side of the lesion. After complete transaction of the spinal cord between the 12th and 13th segments no hind-limb movements were observed up to the 8th day when the animal was killed. The degeneration which resulted from theselesions is well illustrated in two specimens, lizards 7 and 18. In lizard 7 about three-quarters of the transverse extent of the cord was destroyed and fibres could be traced in continuity across the lesion only in the superficial parts of the opposite lateral and anterior columns. Horizontal and transverse sections were examined immediately above the lesion at the 9th segment (Holmes) and further cranially at the 7th (modified Nauta -PI. 2, fig. 12), 6th (Holmes) and 2nd segments (modified Nauta). A horizontal series of sections of the brain stem (Holmes) was also available. All spinal cord sections above the lesion showed degeneration in the dorsal columns similar to that seen after root section in lizard 23 (P1. 1, figs. 2, 3). It was traced to the level of the foramen magnum, but not with certainty to its termination in the medulla oblongata where only Holmes preparations were available. It has already been pointed out that the impregnation by this method is too light for the identification of fine scattered degenerating fibres. The lateral and deeper parts of the dorsal columns showed little or no change, so that any ascending fibres in this situation must have entered the spinal cord cranial to the 10th segment. Other ascending fibres were degenerated in the lateral columns, and were demonstrated most convincingly in the block from the 7th segment (modified Nauta preparations), i.e. three segments cranial to the lesion, (P1. 2, figs. 12, 13). Degeneration was concentrated in the superficial fibres and was most abundant near the dorsal horn. It was very much more marked on the side where the lateral column had been completely transacted but was also present on the other. In the 2nd segment (also a modified Nauta preparation), this degeneration was still recognizable but there was considerably less difference between the two sides. In Holmes preparations from intermediate segments (5th and 6th) the only evidence of degeneration which could be recognized in the lateral columns was vacuolation and fragmentation in some of the coarser fibres, although the loss of the more compact bundles in the posterior columns was quite obvious. Similar sections from the 9th

11 Dorsal spinal nerve roots and spinal cord of Lacerta viridis 163 segment showed many fragmenting fibres in the lateral columns. While no doubt some of these were the ascending fibres recognized further cranially, others may have been short intersegmental fibres and the possibility of direct traumatic effeets cannot be ruled out so close to the lesion. For reasons already given the Holmes preparations available were not satisfactory for tracing the lateral column degeneration to its termination in the brain stem. In the anterior columns a few fragmented fibres were found in segments adjacent to the lesion, but no satisfactory evidence was found in this or any other specimen for the presence of long ascending tracts in this part of the spinal cord. Observations caudal to the lesion are chiefly relevant to the descending pathways which will not be discussed in this paper. They provided a convenient control for the ascending degeneration in the dorsal columns, however, the position of which was occupied by completely normal fibres at all levels examined (12th, 14th and 20th segments). A little degeneration in the central parts of the dorsal columns, still present at the 14th, but not at the 20th segment, probably represented descending collaterals of dorsal root fibres or intersegmental fibres of propriospinal origin. Descending degeneration was very marked in the anterior columns, which, apart from an occasional degenerated fibre, were normal cranial to the lesion, and was also present in the lateral columns. The latter did not therefore provide a satisfactory control for the ascending degeneration found in this situation cranial to the lesion. It appears that there is considerable intermingling of ascending and descending fibres in the lateral columns. Degeneration in the grey matter was present for at least eight segments cranial and caudal to the lesion. It was very similar to the degeneration seen after root section but more abundant and bilateral in distribution, no doubt due to the involvement of intersegmental fibres of propriospinal origin as well as root fibres. In many sections what appeared to be degenerating collaterals could be seen entering the grey matter from the dorsal and the lateral columns. Lizard 18, in which the dorsal two-thirds, of the spinal cord was transacted sparing only the anterior columns, confirmed the findings in lizard 7 in relation to the dorsal columns and also demonstrated the ascending fibres of the lateral columns; collateral branches to the ventral horn were particularly well shown (PI. 1, fig. 9). The lesion was one segment higher than in lizard 7 and some scattered degeneration was present in the lateral and deeper parts of the dorsal columns. The 9th is the most caudal segment to receive a large dorsal root from the region of the forelimb plexus and it is clear that it is only when this or higher segments are affected that ascending degeneration in this part of the dorsal column occurs. In lizard 18 modified Nauta preparations from the brain-stem were available. These confirmed the site of termination of the dorsal column degeneration and also made it possible to trace the lateral column degeneration to its termination. The latter was found in a fairly compact flattened bundle on the ventro-lateral surface of the medulla, from which degenerated fibres passed radially inwards to end among cells of the reticular formation. Such fibres were particularly numerous about the level of exit of the abducens nerve, and could represent terminating fibres or collateral branches. The degeneration extended cranially ventral to the roots of the auditory and trigeminal nerves and then curved dorsally, cranial to the latter, to
.")
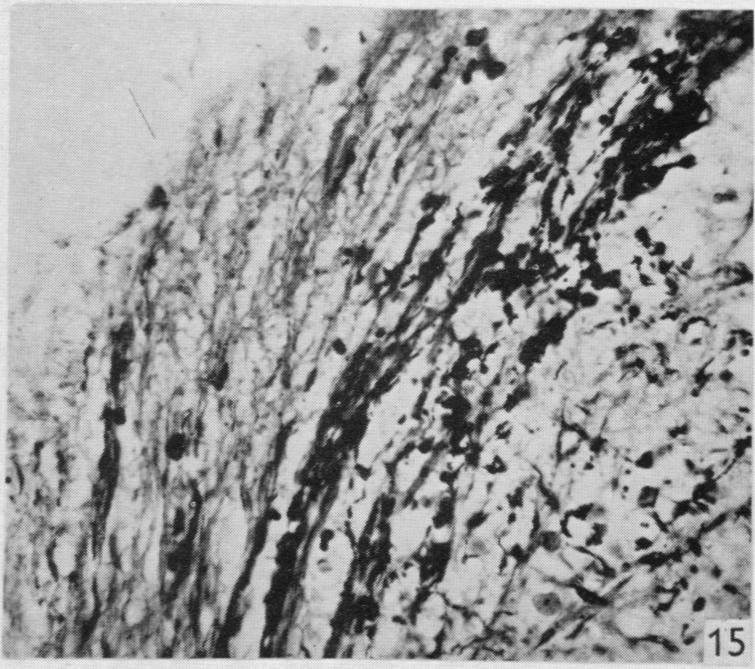
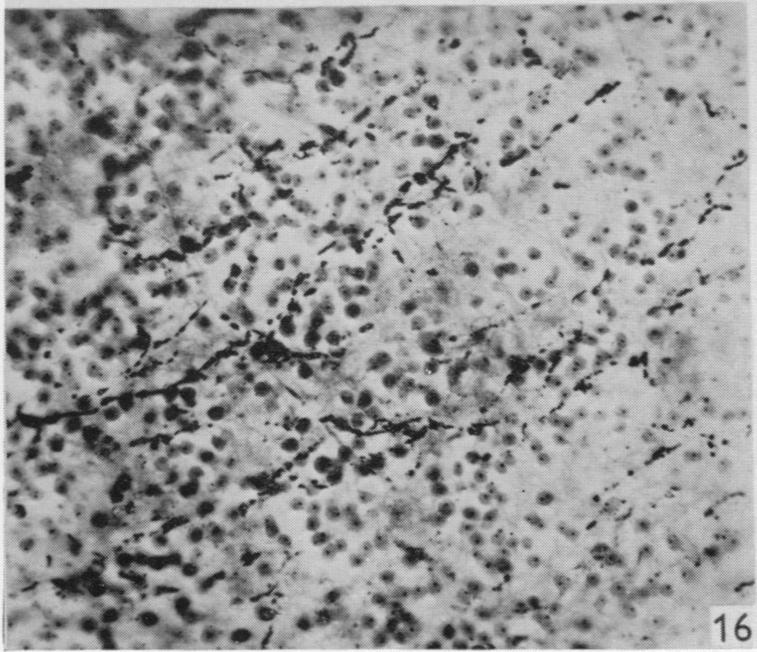
12 164 F. Goldby and L. R. Robinson turn medially between the tectum of the midbrain and the cerebellum. From this situation most of the degenerated fibres entered the cerebellum where they could be traced among the granule cells (see P1. 2, figs. 16 and 17). The cerebellar degeneration was bilateral and many degenerated fibres were seen in the cerebellar commissure which lies adjacent to the decussation of the trochlear nerves. There was also degeneration of a more diffuse character in certain regions of the midbrain but it could not be traced with confidence into continuity with the fibre system just described. The regions in question are in the central grey matter around the caudal part of the aqueduct and also lateral to the posterior corpora quadrigemina in relation to a group of cells which seems to correspond in position with the nucleus isthmi of other reptiles (see Huber & Crosby, 1926). The latter is also very close to the part of the brain stem where Huber & Crosby (1926) identified ascending lemniscus fibres in the alligator. A few degenerated fibres entered the tectum close to the root of the trochlear nerve. These findings, including those in the brain-stem, were confirmed in another animal (lizard 19) with a hemisection of the same type but between the 14th and 15th segments. No degeneration in the lateral and deeper parts of the dorsal columns was seen in this specimen. A hemisection in the 28th segment (lizard 42), which had divided one lateral column only, gave similar results, but failed to demonstrate degeneration in the cerebellum although it was present in the cerebellar commissure. In this case the superficial degeneration in the medulla was unilateral; in the midbrain it was not seen in the region of the nucleus isthmi, but was present in the central grey matter. Another hemisection in the hindlimb enlargement (lizard 22) showed degeneration in the lateral columns which could be traced to the cerebellum and midbrain, but the findings of this specimen are suspect because degenerated fibres were widespread throughout the-medulla and also in the anterior columns of the spinal cord. Such widespread degeneration was seen in no other specimen and may have been partly due to some general pathological condition and not directly to the experimental lesion. The remaining two specimens were hemisections just caudal to the forelimb enlargement and were generally confirmatory of the results already described. CONCLUSIONS AND DISCUSSION The conclusions which can be based on an investigation of this kind vary in the degree of certainty with which they are established. So far as dorsal spinal nerve roots are concerned, it can be said that all appropriate specimens have demonstrated that a proportion of their fibres enter the dorsal columns to form a long ascending tract which reaches the medulla. Within this tract the fibres from the more caudal segments lie dorsomedially to those from more cranial segments. The fibres are fine, but most and perhaps all possess a myelin sheath. The fibres arising caudally end in the dorsal and caudal part of the medulla near the ventricle, while the fibres from more cranial segments end lateral and slightly rostral to the others. A similar somatotopic localization in the posterior column nuclei of Alligator has recently been described by Kruger & Witkovsky (1961) using electrical recording techniques. In Lacerta, the endings are almost certainly axodendritic contacts in the neuropil of this situation, although this particular point has not been established by direct

13 Dorsal spinal nerve roots and spinal cord of Lacerta viridis 165 observation. It is probable that most of these ascending fibres are derived from roots associated with the limb plexuses, because the amount of degeneration at the cervical level in specimens with a lesion just caudal to the forelimb plexus did not differ very much from that in animals with lesions in the region of the hindlimb plexus. From the material available it is not possible to say whether any significant contribution is provided from segments in the tail or whether a median nucleus of Bischoff (Kappers et al. 1936, p. 264) is present in Lacerta. The fibres from segments caudal to the forelimb plexus occupy a region in the cervical spinal cord which corresponds almost exactly with that occupied by the fasciculus gracilis in mammals; the tract in the lizard may be rather smaller in proportion, but the material is not suitable for precise quantitative assessments. Fibres entering from the roots of the forelimb plexus form a tract which is equally comparable with a fasciculus cuneatus, and again the material gives the impression that the proportion of fibres which reach as far as the medulla may be smaller than in mammals. The dorsal columns also contain descending fibres, some passing at least 5 segments below a lesion. These are probably collaterals from ascending fibres, but the material used does not allow a definite distinction to be made between collateral branches and the main stem of a fibre. Clearly the constitution of the dorsal columns in the lizard and in mammals is very similar. The principal differences lie in the poor development and definition of the dorsal column nuclei, the probability that terminations in the medulla are all axodendritic, and the possibility that the columns contain a larger proportion of intersegmental fibres and descending collaterals. It is possible also that a smaller proportion of the ascending root fibres reach brain-stem level for in a mammal (the cat) it has been stated that not more than about 25 % do so (Glees & Soler, 1951). The terminal connexions of dorsal root fibres in the spinal cord are established with slightly less certainty because of the lateral column damage which often accompanied section of the nerve roots. The results were consistent in several specimens, however, and are as reliable as most of those obtained by degeneration methods. They indicate that a major site for the termination of dorsal root fibres lies in the lateral part of the grey matter at the base of the dorsal horn presumably on internuncial neurones, and that these terminations are derived from fibres (or their collaterals) which have passed into the dorsal columns, and are entirely ipsilateral. Terminating fibres are found bilaterally in the anterior horns, more abundantly on the side of the lesion, which they reach through the deeper parts of the lateral columns. This represents the reptilian 'reflexo-motor' pathway for mono-synaptic reflexes. Functional contacts may also be made with the dendritic arborizations which extend into the white matter, but we have no direct evidence of this. The intra-spinal connexions are widespread, extending for at least 5 segments above and below the point of entry of a dorsal root. Again the similarity with the mammalian condition is evident, the principle difference being the presence of the reflexo-motor pathway in the lateral columns. The presence of ascending tracts in the lateral columns is also established by the material examined, but the evidence for the precise sites of termination is less

14 166 F. Goldby and L. R. Robinson satisfactory than in the case of the posterior column tracts. Although scattered to some extent throughout the lateral columns they tend to be concentrated near the dorsal horn and superficially. In this respect they resemble ascending fibres in the lateral columns of mammals. The cells of origin have not been identified. By analogy with mammals one would expect to find them in or near the dorsal horn of the opposite side, but direct evidence is lacking. At least a proportion of these fibres reach the brain stem where some end or give collaterals to the reticular formation of the hindbrain. Others can be traced to the cerebellum and, from the course they follow, are comparable with the ventral or indirect spino-cerebellar fibres ofmammals. Endings in the caudal part of the midbrain tectum, the central grey matter around the aqueduct and in the nucleus isthmi are also probable, although the evidence is not entirely satisfactory. Degenerated fibres were not seen in the midbrain in every specimen in which the lateral columns had been divided. It is true that such degeneration was not seen in any of the control specimens, but it must be remembered that in the experimental animals, owing to its scattered nature, we were unable to trace it with confidence into continuity with the degenerated ascending tracts in the hindbrain. The occasional observation of what appeared to be degenerated fibres in the habenular commissure also throws doubt on the significance of degeneration seen in the midbrain. It is difficult to believe that fibres in the habenular commissure could have been affected directly by any of the lesions and if apparent degeneration in this situation was due to some other cause, no strong reasons can be given for interpreting the midbrain degeneration differently. However, the absence of midbrain degeneration from the control specimens and its presence in most of the experimental animals with spinal lesions provides at least prima face evidence for spino-mesencephalic connexions from the lateral columns. No evidence for spinothalamic connexions was obtained, and perhaps should not be expected. In the more primitive mammals very few fibres from the lateral columns reach the thalamus (Mehler,1957). One may conclude that spino-reticular, spino-cerebellar and probably spinomesencephalic fibres form a definite component of the lateral columns in a reptile, and that they occupy very much the same situation as in mammals. Spino-thalamic fibres are probably not present and no evidence for a dorsal spino-cerebellar tract, entering the cerebellum through an inferior peduncle, was found. No spinal cord lesion in the present series of experiments was more cranial than the 9th segment however. Larsell (1932) thought that a small dorsal spino-cerebellar tract might be present and if so it could arise between the 1st and the 9th spinal segments and derive impulses solely from the forelimb or neck. An origin for spino-thalamic fibres solely from this part of the spinal cord is perhaps less likely. We have also failed to demonstrate ascending fibres in the anterior columns of the spinal cord, unless the degeneration seen in lizard 22 can be taken as evidence for their presence. We have already given reasons why the widespread degeneration seen in this specimen, much of which was found in no other animal with a similar lesion, is of very doubtful significance. It is worth pointing out, however, that Terni (1922) has described ascending fibres in the medial longitudinal fasciculi of the anterior columns, but apart from the one doubtful specimen just mentioned, we have found no confirmatory evidence. It is clear that so far as the present investigation goes the marked differences from

15 Dorsal spinal nerve roots and spinal cord of Lacerta viridis 167 mammals in the forebrain structure of reptiles, which have been made clear by recent work, are not reflected in the ascending pathways of the spinal cord. Dorsal nerve root connexions, and the ascending components of the dorsal and lateral columns are essentially similar to the corresponding features in mammals, and the differences are mostly of a minor character. This is perhaps to be expected. In vertebrates generally the main afferent pathways appear to have been established at an early evolutionary stage. It is in the mechanisms for integration in the forebrain and brain stem, and in those concerned with the elaboration of complex patterns of behaviour, that the principal new developments in the more advanced vertebrates such as mammals have occurred. At the level of the spinal cord, such developments are more likely to be reflected in elaboration and specialization of the descending or motor pathways than in those that are ascending or sensory. SUMMARY The degeneration resulting from lesions in dorsal spinal root fibres and in ascending tracts of the spinal cord in Lacerta viridis has been studied, using the Holmes silver impregnation method and a modified Nauta technique. The nature and criteria of degeneration as revealed by these methods are reviewed. Some features of the normal dorsal roots and spinal cord are described. Severance of three adjacent dorsal roots at the levels of the limb plexuses produced ipsilateral degeneration in dorsal column fibres ascending to the dorsal part of the medulla in fasciculi corresponding closely with the cuneate (forelimb) and gracile (hindlimb) fasciculi of mammals. Intraspinal connexions in the ventral horns and in the roots of the dorsal horns were also demonstrated. Hemisections of the cord resulted in additional degeneration in the superficial part of the lateral column. This could be traced to the cerebellum and, less certainly, to the midbrain. Numerous collaterals were given off to the reticular formation in the medulla. It is concluded that the arrangement of dorsal spinal root connexions and of ascending tracts in the dorsal and lateral columns of the spinal cord is very similar to that found in mammals. No evidence for the presence of long ascending tracts in the anterior columns of the spinal cord was found. The authors wish to thank Mr P. Attwood for technical assistance, and Mr R. J. Fant for preparing the photographs in this study. They also thank Dr R. W. Guillery for the loan of his manuscript describing the modified Nauta technique which was used here. REFERENCES ARMSTRONG, J. A. (1950). An experimental study of the visual pathways in a reptile (Lacerta vivipara). J. Anat., Lond., 84, BECCARI, N. (1913). Sulla spettanza delle fibre del Lenhossek al sistema del nervo accessorio e contribute alla morfologia di questo nervo. (Osservazioni in Lacerta muralss. Arch. ital. Anat. Embriol. 11, BECCARI, N. (1914). II IX, X, XI e XII pajo di nervi cranici e i nervi cervicali negli embrioni di Lacerta muralis. (Contribuzioni allo studio del significato morfologico dei nervi della testa.) Arch. ital. Anat. Embriol. 13, 1-78.
.")
16 168 F. Goldby and L. R. Robinson EDINGER, L. (1908). Vorlesungen fiber den Bau der nervosen Zentralorgane des Menschen und der Tiere. Bd. 2, 7te Aufl. Leipzig: F. C. W. Vogel. GAMBLE, H. J. (1952). An experimental study of the secondary olfactory connexions in Lacerta viridis. J. Anat., Lond., 86, GAMBLE, H. J., GOLDBY, F. & SMITH, G. M. R. (1957). Effect of temperature on the degeneration of nerve fibres. Nature, Lond., 179, 527. GLEES. P. & SOLER, J. (1951). Fibre content of the posterior column and synaptic connections of nucleus gracilis. Z. Zellforsch. 36, GUILLERY, R. XV., SHIRRA, B. & WVEBSTER, K. E. (1961). Differential impregnation of degenerating nerve fibres in paraffin-embedded material. Stain. tech 36, HARRIS, AV. G. (1960). Fibre degeneration in the cerebral cortex of the cat and rabbit following experimental craniotomy. J. Anat., Lond., 94, HOLMIES, AV. (1947). The peripheral nerve biopsy, p In Recent Advances in Clinical Pathology. London: J. and A. Churchill. HUBER, G. C. & CROSBY, E. C. (1926). On thalamic and tectal nuclei and fiber paths in the brain of the American alligator. J. comp. Neurol. 40, INGVAR, S. (1918). Zur Phylo- und Ontogenese des Kleinhirns. Folia neuro-biol., Lpz., 11, KAPPERS, C. U. A., HUBER, G. C. & CROSBY, E. C. (1936). The Comparative Anatomy of the Nervous System of Vertebrates, Including Man. New York: The Macmillan Co. KRUGER, L. & XVITROVSKY, P. (1961). A functional analysis of neurons in the dorsal column nuclei and spinal nucleus of the trigeminal in the reptile (Alligator mississippiensis). (In the Press.) LARSELL, 0. (1926). The cerebellum of reptiles: lizards and snake. J. comp. Neurol. 41, LARSELL, 0. (1932). The cerebellum of reptiles: chelonians and alligator. J. comp. Neurol. 56, LEGHISSA, S. (1954). Richerche anatomo-comparative sul sistema longitudinale mediate nelle serie dei vertebrate. Comment. pontif. Acad. Sci. 16, MIEHLER, NV. R. (1957). The mammalian 'pain tract' in phylogeny. Anat. Rec. 127, 332. NAUTA, W. J. H. & GYGAX, P. A. (1951). Silver impregnation of degenerating axon terminals in the central nervous system: (1) Technic. (2) Chemical notes. Stain. Tech 26, PEELE, T. L. & XVINDLE, WV. F. (1946). Reaction of the spinal cord to laminectomy. Anat. Rec. 94, 488. POWELL, T. P. S., GUILLERY, R. WX. & COWAN, W. M. (1957). A quantitative study of the fornixmamillo-thalamic system. J. Anat., Lond., 91, POWELL, T. P. S. & KRUGER, L. (1960). The thalamic projection upon the telencephalon in Lacerta viridis. J. Anat., Lond., 94, STEFANELLI, A. (1941). Ricerche comparative sui centri tegmentali dei Rettili in rapporto alla loro locomozione. Arch. zool. (ital.), Napoli, 29, TERNI, T. (1922). Ricerche istologiche sul midollo spinale dei rettili con particolare riguardo a componenti spinali del fascicolo longitudinale mediate (Osservazioni in Gonglyus ocellatus Wagl). Arch. ital. Anat. Embriol. 18, Suppl., TERNI, T. (1926). Sui nuclei marginali del midollo spinale dei Sauropsidi. Arch. ital. Anat. Embriol. 23, ZEEBANDELAAR, I. (1920). Ontogenese und Phylogenese der Hinterstrangkerne in Verband mit der Sensibilitat. Folia neuro-biol., Lpz., 12,



17 Journal of Anatorny9 Vol. 96, Part 2 Plate 1 Vi't}.4.+ X 2<WX'''','.~~~~~~~'d GOLDBY AND ROBINON DORSAL SPINA NERO E ROOTS AN SPINAL CORD OFLACERTA Vl~l DI t*~~~~~~~~~~~~~~(fcn.18


18 Journal of Anatomy., Vol. 96, Part 2 Plate 2 12~~~~~~~~~1 GOLDBY AND ROBINSON-DORSAL SPINAL NERVE ROOTS AND SPINAL CORD OF LACERTA VJRIDIS

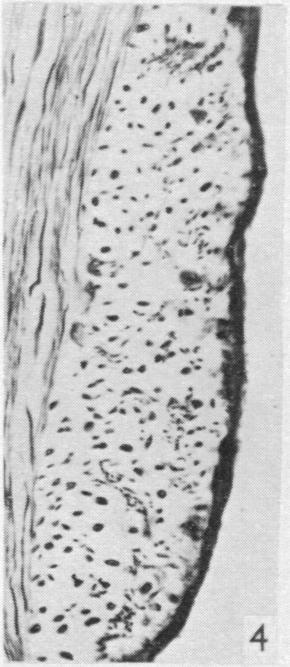
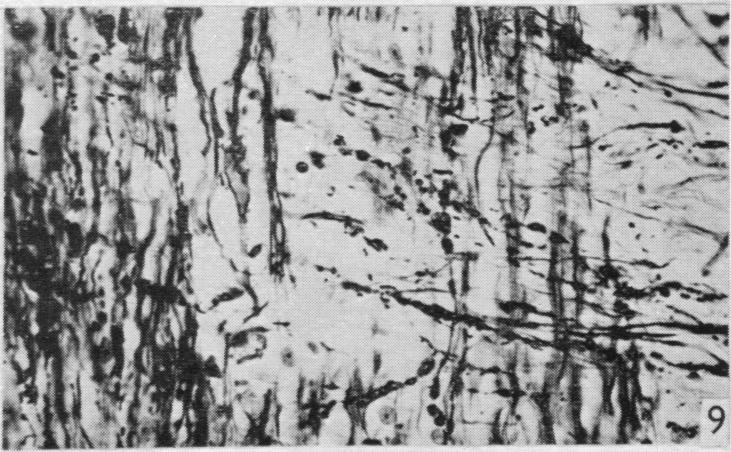
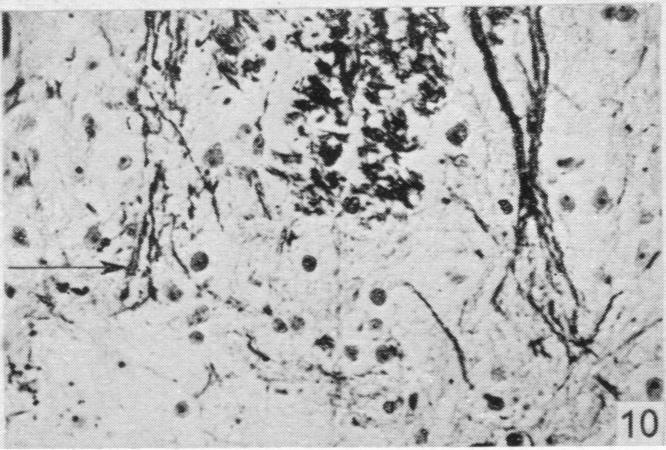
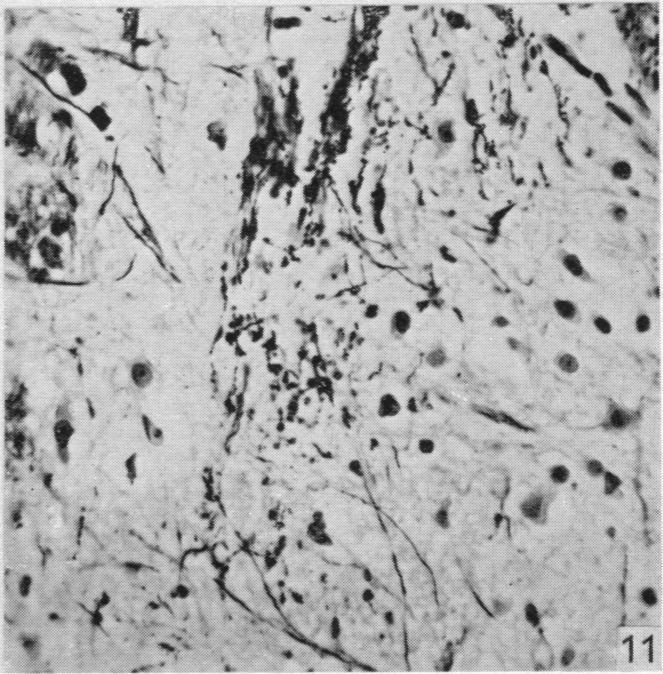
19 Dorsal spinal nerve roots and spinal cord of Lacerta viridis 169 EXPLANATION OF PLATES The regions of the spinal cord of L. viridis which are illustrated photographically in Pls 1 and 2 are indicated in the figure below. This is a diagrammatic outline of a transverse section of the spinal cord inwhichthe rectangles indicate the positions oftransverse andthelinesthe positions of horizontal sections in relation to the cord as a whole. The numbers refer to the figures in the plates. Figs. 1, 2, 4 and 7 are of Holmes preparations; all the others are of modified Nauta preparations. In all the figures which illustrate horizontal sections the cranio-caudal axis of the cord or medulla lies vertically. 8 6~~~~~~~~~~~~~- PLATE 1 Fig. 1. Transverse section throughthe dorsal columns in the 13th spinal segment ofa normal animal. Note the closely packed fine fibres situated superficially on each side of the dorso-median sulcus; compare with fig. 2. x 165. Fig. 2. Lizard 7 (cord lesion at 10th segment). Transverse section through the dorsal columns in the 5th spinal segment, showing bilateral loss of superficial fibres; compare with Fig. 1. (The black material at the surface is India ink used in marking the block.) x 165. Fig. 3. Lizard 7 (cord lesion at 10th segment). Transverse section through the dorsal columns in the 7th spinal segment showing bilateral degeneration in the superficial fibres. The dense impregnation of the degeneration debris in this situation is in striking contrast with the surrounding areas. Compare this Nauta preparation with Fig. 2, which shows the same areas in a Holmes preparation. (See also PI. 2, fig. 12, which is a low power view of the same section.) x165. Fig. 4. Slightly oblique horizontal section through the spinal cord of a normal lizard to show a dorsal spinal root (cut transversely) lying beneath the pia mater and immediately adjacent to the fibres of the lateral column (cut longitudinally). In the dorsal root, the very fine fibres tend to form small groups scattered among the coarser fibres. x 330. Figs. 5, 6. Lizard 33 (7th, 8th and 9th dorsal roots cut). These two photographs are of the anterior horns in the same section from the 8th spinal segment. There is marked degeneration in the anterior horn on the side of the cut spinal roots (fig. 5) and very little in the other (fig. 6). Note the difference in the impregnation of the neuropil on the two sides. x 280. Fig. 7. Lizard 16 (cord lesion at 14th segment). Horizontal section through the dorsal columns at the junction of the spinal cord and medulla 16 days after division of the dorsal columns. Fusiform swellings are present in axons near the midline (indicated by arrows). x 310. Fig. 8. Lizard 23 (27th, 28th and 29th dorsal roots cut). Horizontal section through the dorsal columns in the 4th spinal segment showing unilateral degeneration adjacent to the median plane. x Anat. 96

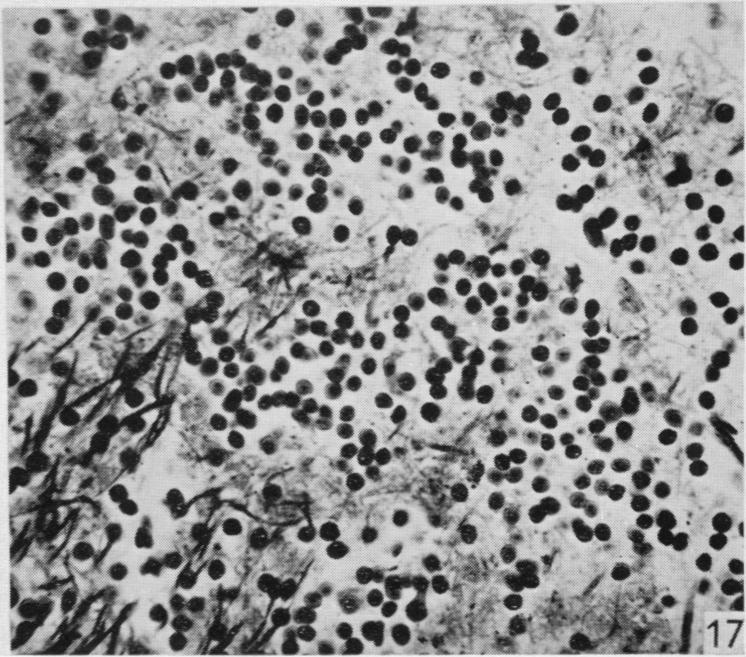
20 170 F. Goldby and L. R. Robinson Fig. 9. Lizard 18 (cord lesion at 9th segment). Horizontal section through the lateral column adjacent to one ventral horn at the 4th segment. Degeneration can be seen in the lateral column on the left of the photograph. There are conspicuous degenerating collaterals passing at right angles to the lateral column into the ventral horn on the right. x 310. Fig. 10. Lizard 34 (8th, 9th and 10th dorsal roots cut). Transverse section at the 4th spinal segment showing grey matter dorsal to the central canal. Degenerating collaterals from the dorsal column are seen on the left. The corresponding fibres on the right are normal. (See also fig. 11.) x 280. Fig. 11. Lizard 34 (8th, 9th and 10th dorsal roots cut). Transverse section at the base of one dorsal horn at the 7th segment to show degeneration in the grey matter. On the left a few fasciculi of the lateral column can be seen and at the top degenerating collaterals passing into the grey matter from the dorsal column. (See also Fig. 10.) x 310. PLATE 2 Fig. 12. Lizard 7 (cord lesion at 10th segment). Transverse section at the 7th spinal segment showing the distribution of degeneration debris. Note the concentrations of degenerating fibres in the dorsal columns (see also Plate 1, fig. 3) and on the surface in one lateral column. There is also scattered degeneration in the rest of the white matter. x 45. Fig. 18. Lizard 7 (cord lesion at 10th segment). Horizontal section through the dorsal part of the lateral column shown in transverse section on the right side of Fig. 12. The entering 7th dorsal root fibres can be seen running tranversely from right to left. Degeneration is conspicuous in the superficial part of the lateral column on the right. The dorsal horn grey matter is on the left. x 310. Fig. 14. Lizard 23 (27th, 28th and 29th dorsal roots cut). Horizontal section through the dorsal part of the medulla near the midline at the caudal end of the lwth ventricle (which is shown at the top left corner of the photograph). This section was one of those (shaded) used in preparing the reconstruction (Text-fig. 1) and shows the degenerating fasciculus gracilis near its apparent termination. x 280. Fig. 15. Lizard 34 (8th, 9th and 10th dorsal roots cut). Horizontal section through the dorsal part of the medulla near the midline at the caudal end of the IVth ventricle (which is shown at the top left corner of the photograph). This section was one of those (shaded) used in preparing the reconstruction (Text-fig. 2) and shows the degenerating fasciculus cuneatus near its apparent termination. The area occupied by the fasciculus gracilis lies medial to it on the left (compare fig. 14). x 280 Fig. 16. Lizard 45 (cord lesion at 10th segment). Horizontal section through the cerebellum showing degenerating fibres in the granular layer following hemisection of the spinal cord. Compare this appearance with that of the normal cerebellum shown in Fig. 17. x 310 Fig. 17. Horizontal section of the cerebellum of a normal lizard taken at a level corresponding approximately to that shown in Fig. 16. x 310.
VARIATION IN MONIEZIA EXPANSA RUDOLPHI
 VARIATION IN MONIEZIA EXPANSA RUDOLPHI STEPHEN R. WILLIAMS, Miami University, Oxford, Ohio In making a number of preparations of proglottids for class study at the stage when sex organs are mature and
VARIATION IN MONIEZIA EXPANSA RUDOLPHI STEPHEN R. WILLIAMS, Miami University, Oxford, Ohio In making a number of preparations of proglottids for class study at the stage when sex organs are mature and
Weekly Schedule of Neuroscience (2017/2018) Week 1
 Week 1") Week 1 28/1/2018 29/1/2018 30/1/2018 31/1/2018 1/2/2018 8:00 9:00 Health politics & 9:00 10:00 Introductory lecture Gross morphology of the brain Gross morphology of spinal cord Health politics & Blood
Week 1 28/1/2018 29/1/2018 30/1/2018 31/1/2018 1/2/2018 8:00 9:00 Health politics & 9:00 10:00 Introductory lecture Gross morphology of the brain Gross morphology of spinal cord Health politics & Blood
Weekly Schedule of Neuroscience (2018/2019) Week 1
 Week 1") Week 1 27/1/2019 28/1/2019 29/1/2019 30/1/2019 31/1/2019 9:00 10:00 Introductory lecture Gross morphology of the brain Gross morphology of spinal cord Health politics & Blood supply of the CNS Language
Week 1 27/1/2019 28/1/2019 29/1/2019 30/1/2019 31/1/2019 9:00 10:00 Introductory lecture Gross morphology of the brain Gross morphology of spinal cord Health politics & Blood supply of the CNS Language
UTILITY OF THE NEUROLOGICAL EXAMINATION IN RATS
 ACTA NEUROBIOL. ELW. 1980, 40 : 999-3 Short communication UTILITY OF THE NEUROLOGICAL EXAMINATION IN RATS David E. TUPPER and Robert B. WALLACE Laboratory of Developmental Psychobiology, University of
ACTA NEUROBIOL. ELW. 1980, 40 : 999-3 Short communication UTILITY OF THE NEUROLOGICAL EXAMINATION IN RATS David E. TUPPER and Robert B. WALLACE Laboratory of Developmental Psychobiology, University of
Anatomy. Name Section. The Vertebrate Skeleton
 Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Class Date. After you read this section, you should be able to answer these questions:
 CHAPTER 14 4 Vertebrates SECTION Introduction to Animals BEFORE YOU READ After you read this section, you should be able to answer these questions: How are vertebrates different from invertebrates? How
CHAPTER 14 4 Vertebrates SECTION Introduction to Animals BEFORE YOU READ After you read this section, you should be able to answer these questions: How are vertebrates different from invertebrates? How
Title: Phylogenetic Methods and Vertebrate Phylogeny
 Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Vertebrates. Vertebrate Characteristics. 444 Chapter 14
 4 Vertebrates Key Concept All vertebrates have a backbone, which supports other specialized body structures and functions. What You Will Learn Vertebrates have an endoskeleton that provides support and
4 Vertebrates Key Concept All vertebrates have a backbone, which supports other specialized body structures and functions. What You Will Learn Vertebrates have an endoskeleton that provides support and
DEVELOPMENT OF THE HEAD AND NECK PLACODES
 DEVELOPMENT OF THE HEAD AND NECK Placodes and the development of organs of special sense L. Moss-Salentijn PLACODES Localized thickened areas of specialized ectoderm, lateral to the neural crest, at the
DEVELOPMENT OF THE HEAD AND NECK Placodes and the development of organs of special sense L. Moss-Salentijn PLACODES Localized thickened areas of specialized ectoderm, lateral to the neural crest, at the
texp. Biol. (196a), 39,
, 39,") texp. Biol. (196a), 39, 239-242 ith 1 plate Printed in Great Britain INNERVATION OF LOCOMOTOR MOVEMENTS BY THE LUMBOSACRAL CORD IN BIRDS AND MAMMALS BY J. TEN CATE Physiological Laboratory, University
texp. Biol. (196a), 39, 239-242 ith 1 plate Printed in Great Britain INNERVATION OF LOCOMOTOR MOVEMENTS BY THE LUMBOSACRAL CORD IN BIRDS AND MAMMALS BY J. TEN CATE Physiological Laboratory, University
Frog Dissection Information Manuel
 Frog Dissection Information Manuel Anatomical Terms: Used to explain directions and orientation of a organism Directions or Positions: Anterior (cranial)- toward the head Posterior (caudal)- towards the
Frog Dissection Information Manuel Anatomical Terms: Used to explain directions and orientation of a organism Directions or Positions: Anterior (cranial)- toward the head Posterior (caudal)- towards the
NOTE XVII. Dr. A.A.W. Hubrecht. which should he in accordance with. of my predecessors. alive or in excellent. further
 further either EUROPEAN NEMERTEANS. 93 NOTE XVII. New Species of European Nemerteans. First Appendix to Note XLIV, Vol. I BY Dr. A.A.W. Hubrecht In the above-mentioned note, published six months ago, several
further either EUROPEAN NEMERTEANS. 93 NOTE XVII. New Species of European Nemerteans. First Appendix to Note XLIV, Vol. I BY Dr. A.A.W. Hubrecht In the above-mentioned note, published six months ago, several
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
examination, the slight resistance encountered being sufficient By J. HERBERT PARSONS.
 PROCEEDI NGS OF THE PHYSIOLOGICAL May 10, 1902. SOCIETY, A method of measuring a visual illusion. By HORACE DARWIN and W. H. R. RIVERS. The instrument we show is designed for the quantitative study of
PROCEEDI NGS OF THE PHYSIOLOGICAL May 10, 1902. SOCIETY, A method of measuring a visual illusion. By HORACE DARWIN and W. H. R. RIVERS. The instrument we show is designed for the quantitative study of
The Laminar and Size Distribution of Commissural Efferent Neurons in the Cat Visual Cortex*
 Arch. histol. jap., Vol. 42, No. 2 (1979) p. 119-128 The Laminar and Size Distribution of Commissural Efferent Neurons in the Cat Visual Cortex* Kazuhiko SHOUMURA Department of Anatomy (Prof. S. DEURA),
Arch. histol. jap., Vol. 42, No. 2 (1979) p. 119-128 The Laminar and Size Distribution of Commissural Efferent Neurons in the Cat Visual Cortex* Kazuhiko SHOUMURA Department of Anatomy (Prof. S. DEURA),
Vertebrates. skull ribs vertebral column
 Vertebrates skull ribs vertebral column endoskeleton in cells working together tissues tissues working together organs working together organs systems Blood carries oxygen to the cells carries nutrients
Vertebrates skull ribs vertebral column endoskeleton in cells working together tissues tissues working together organs working together organs systems Blood carries oxygen to the cells carries nutrients
Mobile Slaughter Unit
 Mobile Slaughter Unit Name of the business/responsible entity USDA Facility Number: 00000 Standard Operating Procedures (SOP) Signature Page Slaughter: beef, swine, goat, and lamb (list all species you
Mobile Slaughter Unit Name of the business/responsible entity USDA Facility Number: 00000 Standard Operating Procedures (SOP) Signature Page Slaughter: beef, swine, goat, and lamb (list all species you
INHERITANCE OF BODY WEIGHT IN DOMESTIC FOWL. Single Comb White Leghorn breeds of fowl and in their hybrids.
 440 GENETICS: N. F. WATERS PROC. N. A. S. and genetical behavior of this form is not incompatible with the segmental interchange theory of circle formation in Oenothera. Summary.-It is impossible for the
440 GENETICS: N. F. WATERS PROC. N. A. S. and genetical behavior of this form is not incompatible with the segmental interchange theory of circle formation in Oenothera. Summary.-It is impossible for the
Introduction and methods will follow the same guidelines as for the draft
 Locomotion Paper Guidelines Entire paper will be 5-7 double spaced pages (12 pt font, Times New Roman, 1 inch margins) without figures (but I still want you to include them, they just don t count towards
Locomotion Paper Guidelines Entire paper will be 5-7 double spaced pages (12 pt font, Times New Roman, 1 inch margins) without figures (but I still want you to include them, they just don t count towards
Flatworms Flatworms Platyhelminthes dorsoventrally free-living planarian parasitic fluke tapeworm label three body layers ectoderm mesoderm
 Flatworms Flatworms are in the phylum Platyhelminthes. Flatworms are flattened dorsoventrally (top to bottom). The group includes the freshwater, free-living planarian and the parasitic fluke and tapeworm.
Flatworms Flatworms are in the phylum Platyhelminthes. Flatworms are flattened dorsoventrally (top to bottom). The group includes the freshwater, free-living planarian and the parasitic fluke and tapeworm.
abnormal lateral geniculate body. His anatomical study suggested that chiasm instead of remaining uncrossed. They thus reach the wrong hemispheres,
 J. Physiol. (1971), 218, pp. 33-62 33 With 1 plate and 9 text-figures Printed in Great Britain ABERRANT VISUAL PROJECTIONS IN THE SIAMESE CAT BY D. H. HUBEL AND T. N. WIESEL From the Department of Neurobiology,
J. Physiol. (1971), 218, pp. 33-62 33 With 1 plate and 9 text-figures Printed in Great Britain ABERRANT VISUAL PROJECTIONS IN THE SIAMESE CAT BY D. H. HUBEL AND T. N. WIESEL From the Department of Neurobiology,
Alimentary System 解剖學科徐淑媛
 Alimentary System 解剖學科徐淑媛 本堂重點 1. Structures derived from primitive guts 2. Specific events Alimentary System endoderm of primordial gut epithelium & glands of digestive tract ectoderm of stomodeum epithelium
Alimentary System 解剖學科徐淑媛 本堂重點 1. Structures derived from primitive guts 2. Specific events Alimentary System endoderm of primordial gut epithelium & glands of digestive tract ectoderm of stomodeum epithelium
THE JOURNAL OF COMPARATIVE NEUROLOGY 233: (1985)
") THE JOURNAL OF COMPARATIVE NEUROLOGY 233:190-212 (1985) Termination Patterns of Individual XI and Y-Cell Axons in the Visual Cortex of the Cat: Projections to Area 18, to the 17/18 Border Region, and to
THE JOURNAL OF COMPARATIVE NEUROLOGY 233:190-212 (1985) Termination Patterns of Individual XI and Y-Cell Axons in the Visual Cortex of the Cat: Projections to Area 18, to the 17/18 Border Region, and to
Distribution of Thalamic Projection Neurons to the Wulst in the Japanese Quail (Coturnix coturnix japonica)
") Distribution of Thalamic Projection Neurons to the Wulst in the Japanese Quail (Coturnix coturnix japonica) Michi YAMADA and Shoei SUGITA Department of Bioproductive Science, Faculty of Agriculture, Utsunomiya
Distribution of Thalamic Projection Neurons to the Wulst in the Japanese Quail (Coturnix coturnix japonica) Michi YAMADA and Shoei SUGITA Department of Bioproductive Science, Faculty of Agriculture, Utsunomiya
Table of Contents. About the Author. Preface. Acknowledgments. Part One: Performing the Feline Physical Examination
 Table of Contents About the Author Preface Acknowledgments Part One: Performing the Feline Physical Examination 1Setting the Stage: Feline-Friendly Practice 1.1 Challenges Faced in Feline Practice 1.2
Table of Contents About the Author Preface Acknowledgments Part One: Performing the Feline Physical Examination 1Setting the Stage: Feline-Friendly Practice 1.1 Challenges Faced in Feline Practice 1.2
cyst&' appeared to be of two kinds-one smaller and Smnith "is inclined to regard these epithelial cell parasites as
 COCCIDIA IN SUBEPITHELIAL INFECTIONS OF THE INTESTINES OF BIRDS PHILIP B. HADLEY From the Agricultural Experiment Station of the Rhode Island State College' Received for publication, July 10, 1916 In an
COCCIDIA IN SUBEPITHELIAL INFECTIONS OF THE INTESTINES OF BIRDS PHILIP B. HADLEY From the Agricultural Experiment Station of the Rhode Island State College' Received for publication, July 10, 1916 In an
A comparison of placental tissue in the skinks Eulamprus tympanum and E. quoyii. Yates, Lauren A.
 A comparison of placental tissue in the skinks Eulamprus tympanum and E. quoyii Yates, Lauren A. Abstract: The species Eulamprus tympanum and Eulamprus quoyii are viviparous skinks that are said to have
A comparison of placental tissue in the skinks Eulamprus tympanum and E. quoyii Yates, Lauren A. Abstract: The species Eulamprus tympanum and Eulamprus quoyii are viviparous skinks that are said to have
The Worms / Chapter 34 and Partial 35 (pgs )
") Name: The Worms / Chapter 34 and Partial 35 (pgs.712-716) 1-6. Worms are not the simple organisms most people think of at first sight. List three specific features that support the idea that worms are
Name: The Worms / Chapter 34 and Partial 35 (pgs.712-716) 1-6. Worms are not the simple organisms most people think of at first sight. List three specific features that support the idea that worms are
AMERICAN MUSEUM NOVITATES Published by
 AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
The ascending tectofugal visual system in amniotes: New insights
 Brain Research Bulletin 66 (2005) 290 296 The ascending tectofugal visual system in amniotes: New insights Salvador Guirado,1,M a. Ángeles Real 1, José Carlos Dávila Department of Cell Biology, Genetics
Brain Research Bulletin 66 (2005) 290 296 The ascending tectofugal visual system in amniotes: New insights Salvador Guirado,1,M a. Ángeles Real 1, José Carlos Dávila Department of Cell Biology, Genetics
Title. Author(s)YAMASHITA, Jiro; OHBAYASHI, Masashi; KONNO, Seiji. CitationJapanese Journal of Veterinary Research, 4(3): Issue Date
YAMASHITA, Jiro; OHBAYASHI, Masashi; KONNO, Seiji. CitationJapanese Journal of Veterinary Research, 4(3): Issue Date") Title STUDIES ON ECHINOCOCCOSIS : III. ON EXPERIMENTAL INF DEVELOPMENT OF ECHINOCOCCUS GRANULOSUS (BATSCH, 1786 Author(s)YAMASHITA, Jiro; OHBAYASHI, Masashi; KONNO, Seiji CitationJapanese Journal of Veterinary
Title STUDIES ON ECHINOCOCCOSIS : III. ON EXPERIMENTAL INF DEVELOPMENT OF ECHINOCOCCUS GRANULOSUS (BATSCH, 1786 Author(s)YAMASHITA, Jiro; OHBAYASHI, Masashi; KONNO, Seiji CitationJapanese Journal of Veterinary
LABORATORY EXERCISE 6: CLADISTICS I
 Biology 4415/5415 Evolution LABORATORY EXERCISE 6: CLADISTICS I Take a group of organisms. Let s use five: a lungfish, a frog, a crocodile, a flamingo, and a human. How to reconstruct their relationships?
Biology 4415/5415 Evolution LABORATORY EXERCISE 6: CLADISTICS I Take a group of organisms. Let s use five: a lungfish, a frog, a crocodile, a flamingo, and a human. How to reconstruct their relationships?
COMPARATIVE VERTEBRATE HISTOLOGY ZOO 4756c Syllabus for Fall 2018
 COMPARATIVE VERTEBRATE HISTOLOGY ZOO 4756c Syllabus for Fall 2018 Instructor: Frank T. Logiudice Office: Biology Building, Room 202c Office Phone Number: (407) - 823-2495 Email Address: Frank.Logiudice@ucf.edu
COMPARATIVE VERTEBRATE HISTOLOGY ZOO 4756c Syllabus for Fall 2018 Instructor: Frank T. Logiudice Office: Biology Building, Room 202c Office Phone Number: (407) - 823-2495 Email Address: Frank.Logiudice@ucf.edu
HISTOPATHOLOGY. Introduction:
 Introduction: HISTOPATHOLOGY Goats and sheep are the major domestic animal species in India. Much of the economy of the country has been depend upon the domestication of these animals. Especially economy
Introduction: HISTOPATHOLOGY Goats and sheep are the major domestic animal species in India. Much of the economy of the country has been depend upon the domestication of these animals. Especially economy
Williston, and as there are many fairly good specimens in the American
 56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
An introduction to ear cytology in small animal patients
 Vet Times The website for the veterinary profession https://www.vettimes.co.uk An introduction to ear cytology in small animal patients Author : Ariane Neuber Categories : RVNs Date : November 1, 2009
Vet Times The website for the veterinary profession https://www.vettimes.co.uk An introduction to ear cytology in small animal patients Author : Ariane Neuber Categories : RVNs Date : November 1, 2009
VERTEBRATE READING. Fishes
 VERTEBRATE READING Fishes The first vertebrates to become a widespread, predominant life form on earth were fishes. Prior to this, only invertebrates, such as mollusks, worms and squid-like animals, would
VERTEBRATE READING Fishes The first vertebrates to become a widespread, predominant life form on earth were fishes. Prior to this, only invertebrates, such as mollusks, worms and squid-like animals, would
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS
 AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.
Dairy Cattle Assessment protocol
 Dairy Cattle Assessment protocol Guidance on sampling: Individual measures 1a. Mobility individual scoring 2. Body condition 3. Cleanliness 4. Hair loss, Lesions 5. Swellings Assessed on 20 cows from the
Dairy Cattle Assessment protocol Guidance on sampling: Individual measures 1a. Mobility individual scoring 2. Body condition 3. Cleanliness 4. Hair loss, Lesions 5. Swellings Assessed on 20 cows from the
Animal Form and Function. Amphibians. United by several distinguishing apomorphies within the Vertebrata
 Animal Form and Function Kight Amphibians Class Amphibia (amphibia = living a double life) United by several distinguishing apomorphies within the Vertebrata 1. Skin Thought Question: For whom are integumentary
Animal Form and Function Kight Amphibians Class Amphibia (amphibia = living a double life) United by several distinguishing apomorphies within the Vertebrata 1. Skin Thought Question: For whom are integumentary
What is the evidence for evolution?
 What is the evidence for evolution? 1. Geographic Distribution 2. Fossil Evidence & Transitional Species 3. Comparative Anatomy 1. Homologous Structures 2. Analogous Structures 3. Vestigial Structures
What is the evidence for evolution? 1. Geographic Distribution 2. Fossil Evidence & Transitional Species 3. Comparative Anatomy 1. Homologous Structures 2. Analogous Structures 3. Vestigial Structures
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC
 FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
FROG DISSECTION. a. Why is there a difference in size proportion between the hind and fore limbs?
 FROG DISSECTION External Anatomy 1. The division of a frog s body includes the head, trunk and limbs. Examine the front and hind limbs of the frog. The hind limbs are the long, more muscular limbs of the
FROG DISSECTION External Anatomy 1. The division of a frog s body includes the head, trunk and limbs. Examine the front and hind limbs of the frog. The hind limbs are the long, more muscular limbs of the
Phylum Platyhelminthes Flatworms
 Phylum Platyhelminthes Flatworms The Acoelomates The acoelomates are animals that lack a coelom. Acoelomates lack a body cavity, and instead the space between the body wall and the digestive tract is filled
Phylum Platyhelminthes Flatworms The Acoelomates The acoelomates are animals that lack a coelom. Acoelomates lack a body cavity, and instead the space between the body wall and the digestive tract is filled
Biology. Slide 1of 50. End Show. Copyright Pearson Prentice Hall
 Biology 1of 50 2of 50 Phylogeny of Chordates Nonvertebrate chordates Jawless fishes Sharks & their relatives Bony fishes Reptiles Amphibians Birds Mammals Invertebrate ancestor 3of 50 A vertebrate dry,
Biology 1of 50 2of 50 Phylogeny of Chordates Nonvertebrate chordates Jawless fishes Sharks & their relatives Bony fishes Reptiles Amphibians Birds Mammals Invertebrate ancestor 3of 50 A vertebrate dry,
ON THE FPERYLOSIS OF THE BLACK-THROATED DIVER.
 ON THE FPERYLOSIS OF THE BLACK-THROATED DIVER. BY W. P. PYCRAFT. IT is surely a matter for regret that so little interest has been taken in that side of ornithology which concerns structural characters,
ON THE FPERYLOSIS OF THE BLACK-THROATED DIVER. BY W. P. PYCRAFT. IT is surely a matter for regret that so little interest has been taken in that side of ornithology which concerns structural characters,
( 142 ) NOTES ON THE GREAT NORTHERN DIVER.
 NOTES ON THE GREAT NORTHERN DIVER.") ( 142 ) NOTES ON THE GREAT NORTHERN DIVER. BY ERIC B. DUNXOP. THE Great Northern Diver (Gavia immer) is best known in the British Isles as a winter-visitor, though in the Orkneys I have frequently seen
( 142 ) NOTES ON THE GREAT NORTHERN DIVER. BY ERIC B. DUNXOP. THE Great Northern Diver (Gavia immer) is best known in the British Isles as a winter-visitor, though in the Orkneys I have frequently seen
experimental studies of many workers (Hetherington, 1941; Hetherington & nucleus, resulted in obesity. The confusion introduced by the notion
 143 J. Physiol. (I955) I27, I43-152 HYPOHALAMIC CONROL OF FOOD INAKE IN CAS AND MONKEYS BY B. K. ANAND, S. DUA AND KAE SHOENBERG From the Department of Physiology, Lady Hardinge Medical College, New Delhi,
143 J. Physiol. (I955) I27, I43-152 HYPOHALAMIC CONROL OF FOOD INAKE IN CAS AND MONKEYS BY B. K. ANAND, S. DUA AND KAE SHOENBERG From the Department of Physiology, Lady Hardinge Medical College, New Delhi,
Anat. Labor. of Prof. H. SETO, Tohoku University, On the Sensory Terminations Formed along the Ductus
 Anat. Labor. of Prof. H. SETO, Tohoku University, Sendai. On the Sensory Terminations Formed along the Ductus Pancreaticus in Cat. The existence of PACINIan bodies in the pancreas of mammals, especially
Anat. Labor. of Prof. H. SETO, Tohoku University, Sendai. On the Sensory Terminations Formed along the Ductus Pancreaticus in Cat. The existence of PACINIan bodies in the pancreas of mammals, especially
F.L. Andr6s. Rua Tristao Vaz No Esq., 1400 Lisboa, Portugal
 Supranumerary Barrels Develop in the Somatosensory Cortex of Mice, After the Implantation of the Vibrissal Follicle Parts Containing Large Numbers of Receptors F.L. Andr6s Rua Tristao Vaz No. 37 1 Esq.,
Supranumerary Barrels Develop in the Somatosensory Cortex of Mice, After the Implantation of the Vibrissal Follicle Parts Containing Large Numbers of Receptors F.L. Andr6s Rua Tristao Vaz No. 37 1 Esq.,
SELECTION FOR AN INVARIANT CHARACTER, VIBRISSA NUMBER, IN THE HOUSE MOUSE. IV. PROBIT ANALYSIS
 SELECTION FOR AN INVARIANT CHARACTER, VIBRISSA NUMBER, IN THE HOUSE MOUSE. IV. PROBIT ANALYSIS BERENICE KINDRED Division of Animal Genetics, C.S.I.R.O., University of Sydney, Australia Received November
SELECTION FOR AN INVARIANT CHARACTER, VIBRISSA NUMBER, IN THE HOUSE MOUSE. IV. PROBIT ANALYSIS BERENICE KINDRED Division of Animal Genetics, C.S.I.R.O., University of Sydney, Australia Received November
OXYGEN POISONING IN COLD BLOODED ANIMALS, By JAMES M. FAULKNER, M.D., AND CARL A. L. BINGER, M.D. (Received for publication, January 3, 1927.
 Published Online: 1 May, 1927 Supp Info: http://doi.org/10.1084/jem.45.5.865 Downloaded from jem.rupress.org on September 21, 2018 OXYGEN POISONING IN COLD BLOODED ANIMALS, By JAMES M. FAULKNER, M.D.,
Published Online: 1 May, 1927 Supp Info: http://doi.org/10.1084/jem.45.5.865 Downloaded from jem.rupress.org on September 21, 2018 OXYGEN POISONING IN COLD BLOODED ANIMALS, By JAMES M. FAULKNER, M.D.,
Comparative Zoology Portfolio Project Assignment
 Comparative Zoology Portfolio Project Assignment Using your knowledge from the in class activities, your notes, you Integrated Science text, or the internet, you will look at the major trends in the evolution
Comparative Zoology Portfolio Project Assignment Using your knowledge from the in class activities, your notes, you Integrated Science text, or the internet, you will look at the major trends in the evolution
LABORATORY EXERCISE 7: CLADISTICS I
 Biology 4415/5415 Evolution LABORATORY EXERCISE 7: CLADISTICS I Take a group of organisms. Let s use five: a lungfish, a frog, a crocodile, a flamingo, and a human. How to reconstruct their relationships?
Biology 4415/5415 Evolution LABORATORY EXERCISE 7: CLADISTICS I Take a group of organisms. Let s use five: a lungfish, a frog, a crocodile, a flamingo, and a human. How to reconstruct their relationships?
Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida. Evo-Devo Revisited. Development of the Tetrapod Limb
 Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida Evo-Devo Revisited Development of the Tetrapod Limb Limbs whether fins or arms/legs for only in particular regions or LIMB FIELDS. Primitively
Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida Evo-Devo Revisited Development of the Tetrapod Limb Limbs whether fins or arms/legs for only in particular regions or LIMB FIELDS. Primitively
BREATHING WHICH IS NOT RESPIRATION
 BREATHING WHICH IS NOT RESPIRATION Breathing vs. Respiration All animals respire. A lot of people think respiration means breathing- this is not true! Breathing is the physical process of inhaling oxygen
BREATHING WHICH IS NOT RESPIRATION Breathing vs. Respiration All animals respire. A lot of people think respiration means breathing- this is not true! Breathing is the physical process of inhaling oxygen
Active sensing. Ehud Ahissar
 Active sensing Ehud Ahissar 1 Active sensing Passive vs active sensing (touch) Comparison across senses Basic coding principles -------- Perceptual loops Sensation-targeted motor control Proprioception
Active sensing Ehud Ahissar 1 Active sensing Passive vs active sensing (touch) Comparison across senses Basic coding principles -------- Perceptual loops Sensation-targeted motor control Proprioception
A FAMILY OF SPOTTED Q. I. SIMPSON AND W. E. CASTLE1
 A FAMILY OF SPOTTED NEGROES Q. I. SIMPSON AND W. E. CASTLE1 IT is the purpose of this note to put on record an interesting variation in human skin color which made its appearance as a mutation or sport
A FAMILY OF SPOTTED NEGROES Q. I. SIMPSON AND W. E. CASTLE1 IT is the purpose of this note to put on record an interesting variation in human skin color which made its appearance as a mutation or sport
NOTE XXXVIII. Three new species of the genus Helota DESCRIBED BY. C. Ritsema+Cz. is very. friend René Oberthür who received. Biet.
 Subshining; HELOTA MARIAE. 249 NOTE XXXVIII. Three new species of the genus Helota DESCRIBED BY C. Ritsema+Cz. The first of these species is very interesting as it belongs to the same section as the recently
Subshining; HELOTA MARIAE. 249 NOTE XXXVIII. Three new species of the genus Helota DESCRIBED BY C. Ritsema+Cz. The first of these species is very interesting as it belongs to the same section as the recently
$? 479 THE FUNCTION OF M. DEPRESSOR CAUDAE AND M. CAUDOFEMORALIS IN PIGEONS
 Oct.1 $? 479 THE FUNCTION OF M. DEPRESSOR CAUDAE AND M. CAUDOFEMORALIS IN PIGEONS BY HARVEY I. FISHER THE usual method of determining the function of a muscle is by gross dissection and study of attachments.
Oct.1 $? 479 THE FUNCTION OF M. DEPRESSOR CAUDAE AND M. CAUDOFEMORALIS IN PIGEONS BY HARVEY I. FISHER THE usual method of determining the function of a muscle is by gross dissection and study of attachments.
What s Your Diagnosis?
 What s Your Diagnosis? Signalment: Maine Coone (8 month old, female intact) Presenting complaint: Lethargy, inappetence, serosanguinous vaginal discharge History: Lives with 11 other Maine Coone cats (males
What s Your Diagnosis? Signalment: Maine Coone (8 month old, female intact) Presenting complaint: Lethargy, inappetence, serosanguinous vaginal discharge History: Lives with 11 other Maine Coone cats (males
A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae)
") Genus Vol. 14 (3): 413-418 Wroc³aw, 15 X 2003 A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae) JAROS AW KANIA Zoological Institute, University of Wroc³aw, Sienkiewicza
Genus Vol. 14 (3): 413-418 Wroc³aw, 15 X 2003 A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae) JAROS AW KANIA Zoological Institute, University of Wroc³aw, Sienkiewicza
Variations in the White Pigment of the Eye in Gammaru8 cheureuxisexton, with a Description of a New Genetic Type, the" Clotted eye.
 [ 691 ] Variations in the White Pigment of the Eye in Gammaru8 cheureuxisexton, with a Description of a New Genetic Type, the" Clotted eye." By E. W. Sexton and A. R. Clark. From the Plymouth Laboratory.
[ 691 ] Variations in the White Pigment of the Eye in Gammaru8 cheureuxisexton, with a Description of a New Genetic Type, the" Clotted eye." By E. W. Sexton and A. R. Clark. From the Plymouth Laboratory.
Alternatives in Veterinary Anatomy Training
 Training Computer Software The items in this category are numerous. The following are some good examples. Comparative Anatomy: Mammals, Birds and Fish This computer software covers an introduction to:
Training Computer Software The items in this category are numerous. The following are some good examples. Comparative Anatomy: Mammals, Birds and Fish This computer software covers an introduction to:
Fischthal and Kuntz (1964) reported the
 reported the") Zoological Studies 41(3): 283-287 (2002) Meristocotyle provitellaria sp. nov. (Digenea: Meristocotylidae) from Varanus salvator in China Wei Liu 1, Qing-Kui Li 2, Hsiu-Hui Shih 3 and Zhao-Zhi Qiu 1, *
Zoological Studies 41(3): 283-287 (2002) Meristocotyle provitellaria sp. nov. (Digenea: Meristocotylidae) from Varanus salvator in China Wei Liu 1, Qing-Kui Li 2, Hsiu-Hui Shih 3 and Zhao-Zhi Qiu 1, *
COMPONENTS OF EMOTIONAL EXPRESSION: AN EXPERIMENTAL STUDY*
 VOCAL AND RESPIRATORY COMPOUNDS OF EMOTIONXL EXPRESSION 241 ASSOCIATED FACIAL, VOCAL AND RESPIRATORY COMPONENTS OF EMOTIONAL EXPRESSION: AN EXPERIMENTAL STUDY* BY H. W. MAGOUN, D. ATLAS, E. H. INGERSOLL
VOCAL AND RESPIRATORY COMPOUNDS OF EMOTIONXL EXPRESSION 241 ASSOCIATED FACIAL, VOCAL AND RESPIRATORY COMPONENTS OF EMOTIONAL EXPRESSION: AN EXPERIMENTAL STUDY* BY H. W. MAGOUN, D. ATLAS, E. H. INGERSOLL
A quantitative study of hair growth using mouse and rat vibrissal follicles
 /. Embryol. exp. Morph. Vol. 72, pp. 209-224, 1982 209 Printed in Great Britain Company of Biologists Limited 1982 A quantitative study of hair growth using mouse and rat vibrissal follicles I. Dermal
/. Embryol. exp. Morph. Vol. 72, pp. 209-224, 1982 209 Printed in Great Britain Company of Biologists Limited 1982 A quantitative study of hair growth using mouse and rat vibrissal follicles I. Dermal
Pre-natal construction of neural circuits (the highways are genetically specified):
:") Modification of Brain Circuits as a Result of Experience Chapter 24, Purves et al. 4 th Ed. Pre-natal construction of neural circuits (the highways are genetically specified): (1/6/2010) Mona Buhusi Postnatal
Modification of Brain Circuits as a Result of Experience Chapter 24, Purves et al. 4 th Ed. Pre-natal construction of neural circuits (the highways are genetically specified): (1/6/2010) Mona Buhusi Postnatal
LABORATORY SYLLABUS. Emphasis: Detailed and organismic understanding of the morphology of the shark and cat as models of a fish and a tetrapod.
 Syllabus page 1 LABORATORY SYLLABUS Comparative Anatomy, 4 credit hours Actual Lab Sessions: Tu/Th 10:40am-1:30pm Independent studies: Mon/Wed/Fri 9:00am-4:30pm; Tu/Th 1:40pm-4:30pm Laboratory manual:
Syllabus page 1 LABORATORY SYLLABUS Comparative Anatomy, 4 credit hours Actual Lab Sessions: Tu/Th 10:40am-1:30pm Independent studies: Mon/Wed/Fri 9:00am-4:30pm; Tu/Th 1:40pm-4:30pm Laboratory manual:
RECESSIVE BUDGIES: A BEGINNERS INTRODUCTION TO RECESSIVES IN BUDGERIGARS.
 RECESSIVE BUDGIES: A BEGINNERS INTRODUCTION TO RECESSIVES IN BUDGERIGARS. Published on the AWEBSA webpage with the kind permission of the author: Robert Manvell. Please visit his page and view photos of
RECESSIVE BUDGIES: A BEGINNERS INTRODUCTION TO RECESSIVES IN BUDGERIGARS. Published on the AWEBSA webpage with the kind permission of the author: Robert Manvell. Please visit his page and view photos of
May 4-6, 2004 University of Arkansas
 May 4-6, 2004 University of Arkansas BSE Update Meat Industry Perspective Randall Huffman, Ph.D. V.P. Scientific Affairs American Meat Institute Foundation Tuesday, December 23 USDA Announcement Overview
May 4-6, 2004 University of Arkansas BSE Update Meat Industry Perspective Randall Huffman, Ph.D. V.P. Scientific Affairs American Meat Institute Foundation Tuesday, December 23 USDA Announcement Overview
HISTOPHYSIOLOGICAL STUDIES ON THE HYPOPHYSIO- MAMMARY AXIS IN SHEEP (Ovis aries) - MAMMOTROPHS
 - MAMMOTROPHS") International Journal of Science, Environment and Technology, Vol. 5, No 3, 2016, 912 917 ISSN 2278-3687 (O) 2277-663X (P) HISTOPHYSIOLOGICAL STUDIES ON THE HYPOPHYSIO- MAMMARY AXIS IN SHEEP (Ovis aries)
International Journal of Science, Environment and Technology, Vol. 5, No 3, 2016, 912 917 ISSN 2278-3687 (O) 2277-663X (P) HISTOPHYSIOLOGICAL STUDIES ON THE HYPOPHYSIO- MAMMARY AXIS IN SHEEP (Ovis aries)
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes)
") Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
A Lymphosarcoma in an Atlantic Salmon (Salmo salar)
") A Lymphosarcoma in an Atlantic Salmon (Salmo salar) Authors: Paul R. Bowser, Marilyn J. Wolfe, and Timothy Wallbridge Source: Journal of Wildlife Diseases, 23(4) : 698-701 Published By: Wildlife Disease
A Lymphosarcoma in an Atlantic Salmon (Salmo salar) Authors: Paul R. Bowser, Marilyn J. Wolfe, and Timothy Wallbridge Source: Journal of Wildlife Diseases, 23(4) : 698-701 Published By: Wildlife Disease
THE EFFECT OF DEAFFERENTATION UPON THE LOCOMOTORY ACTIVITY OF AMPHIBIAN LIMBS
 227 THE EFFECT OF DEAFFERENTATION UPON THE LOCOMOTORY ACTIVITY OF AMPHIBIAN LIMBS BY J. GRAY AND H. W. LISSMANN Zoological Laboratory, Cambridge (Received i December 1939) (With One Plate and One Text-figure)
227 THE EFFECT OF DEAFFERENTATION UPON THE LOCOMOTORY ACTIVITY OF AMPHIBIAN LIMBS BY J. GRAY AND H. W. LISSMANN Zoological Laboratory, Cambridge (Received i December 1939) (With One Plate and One Text-figure)
Vol. XIV, No. 1, March, The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S.
 By S.") Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
CAT DISSECTION A LABORATORY GUIDE
 8546d_fm_i-iv 6/26/02 3:51 PM Page 3 mac62 mac62:1253_ge: CAT DISSECTION A LABORATORY GUIDE CONNIE ALLEN VALERIE HARPER Edison Community College John Wiley & Sons, Inc. 8546d_fm_i-iv 6/26/02 12:17 PM Page
8546d_fm_i-iv 6/26/02 3:51 PM Page 3 mac62 mac62:1253_ge: CAT DISSECTION A LABORATORY GUIDE CONNIE ALLEN VALERIE HARPER Edison Community College John Wiley & Sons, Inc. 8546d_fm_i-iv 6/26/02 12:17 PM Page
Progressive ataxia due to central demyelination in Rottweiler dogs
 Veterinary Quarterly ISSN: 0165-2176 (Print) 1875-5941 (Online) Journal homepage: https://www.tandfonline.com/loi/tveq20 Progressive ataxia due to central demyelination in Rottweiler dogs W. Wouda & J.
Veterinary Quarterly ISSN: 0165-2176 (Print) 1875-5941 (Online) Journal homepage: https://www.tandfonline.com/loi/tveq20 Progressive ataxia due to central demyelination in Rottweiler dogs W. Wouda & J.
The Brain and Senses. Birds perceive the world differently than humans. Avian intelligence. Novel feeding behaviors
 The Brain and Senses Birds perceive the world differently than humans Color and IR vision are highly developed Hearing is superior, owls track prey in total darkness Birds navigate using abilities to sense:
The Brain and Senses Birds perceive the world differently than humans Color and IR vision are highly developed Hearing is superior, owls track prey in total darkness Birds navigate using abilities to sense:
THE EFFECT OF MUTILATION ON THE TAPEWORM TAENIA TAENIAEFORMIS
 THE EFFECT OF MUTILATION ON THE TAPEWORM TAENIA TAENIAEFORMIS JOE N. MILLER AND WM. P. BUNNER The reader is undoubtedly aware of work which has been done by Child (1910) and others in mutilating certain
THE EFFECT OF MUTILATION ON THE TAPEWORM TAENIA TAENIAEFORMIS JOE N. MILLER AND WM. P. BUNNER The reader is undoubtedly aware of work which has been done by Child (1910) and others in mutilating certain
Polecats & Ferrets. How to tell them apart
 Polecats & Ferrets How to tell them apart Introduction The polecat (Mustela putorius) is expanding its range in Britain, and in many areas across Britain, ferrets (Mustela furo) occur either as individuals
Polecats & Ferrets How to tell them apart Introduction The polecat (Mustela putorius) is expanding its range in Britain, and in many areas across Britain, ferrets (Mustela furo) occur either as individuals
Liver and Gallbladder Morphology of the juvenile Nile crocodile, Crocodylus niloticus (Laurenti, 1768)
") Liver and Gallbladder Morphology of the juvenile Nile crocodile, Crocodylus niloticus (Laurenti, 1768) by ERNA VAN WILPE Submitted in partial fulfilment of the requirements for the degree MSc DEPARTMENT
Liver and Gallbladder Morphology of the juvenile Nile crocodile, Crocodylus niloticus (Laurenti, 1768) by ERNA VAN WILPE Submitted in partial fulfilment of the requirements for the degree MSc DEPARTMENT
.56 m. (22 in.). COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition
. COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition") Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
examnined when three weeks old. Not one of the eyes showed
 A NOTE ON THE DOG'S TAPETUM IN EARLY LIFE* BY C. H. USHER ABERDEEN THIS note is written for the purpose of pointing out that the tapetum of the dog is not recognizable ophthalmoscopically for several weeks
A NOTE ON THE DOG'S TAPETUM IN EARLY LIFE* BY C. H. USHER ABERDEEN THIS note is written for the purpose of pointing out that the tapetum of the dog is not recognizable ophthalmoscopically for several weeks
Invertebrates. Brain. Brain 12/2/2017. The Invertebrate Brain. The Invertebrate Brain. Invertebrate brain general layouts some specific functions
 Brain Invertebrate brain general layouts some specific functions Vertebrate brain general layout cortical fields evolutionary theory Brain Brain size Invertebrates 1) No brain (only nerve net) jellyfish,
Brain Invertebrate brain general layouts some specific functions Vertebrate brain general layout cortical fields evolutionary theory Brain Brain size Invertebrates 1) No brain (only nerve net) jellyfish,
Reptile Identification Guide
 Care & preservation of Surrey s native amphibians and reptiles Reptile Identification Guide This identification guide is intended to act as an aid for SARG surveyors. Adder, Vipera berus A short, stocky
Care & preservation of Surrey s native amphibians and reptiles Reptile Identification Guide This identification guide is intended to act as an aid for SARG surveyors. Adder, Vipera berus A short, stocky
Ectoparasites Myobia musculi Radfordia affinis Radfordia ensifera
 Ectoparasites Fleas, ticks, and lice are uncommon in modern laboratory facilities, but may be seen on wild or feral rodents. Most ectoparasite infestations seen in rats and mice used for research are various
Ectoparasites Fleas, ticks, and lice are uncommon in modern laboratory facilities, but may be seen on wild or feral rodents. Most ectoparasite infestations seen in rats and mice used for research are various
ROSENTHAL(1) was the first to point out that the effect of section of
 was the first to point out that the effect of section of") THE EFFECT OF SECTION OF THE VAGI ON THE RESPIRATION OF THE CAT. BY J. TREVAN AND E. BOOCK. (From the Wellcome Physiological Research Laboratories.) ROSENTHAL(1) was the first to point out that the effect
THE EFFECT OF SECTION OF THE VAGI ON THE RESPIRATION OF THE CAT. BY J. TREVAN AND E. BOOCK. (From the Wellcome Physiological Research Laboratories.) ROSENTHAL(1) was the first to point out that the effect
New Species of Black Coral (Cnidaria: Antipatharia) from the Northern Gulf of Mexico
 from the Northern Gulf of Mexico") Northeast Gulf Science Volume 12 Number 2 Number 2 Article 2 10-1992 New Species of Black Coral (Cnidaria: Antipatharia) from the Northern Gulf of Mexico Dennis M. Opresko Oak Ridge National Laboratory
Northeast Gulf Science Volume 12 Number 2 Number 2 Article 2 10-1992 New Species of Black Coral (Cnidaria: Antipatharia) from the Northern Gulf of Mexico Dennis M. Opresko Oak Ridge National Laboratory
Key words: Mouse motor cortex, intracortical microstimulation, motor representation,.corticomotor asymmetry.
 Neuroscience and Behavioral Physiology, Vol. 28, No. 1, 1998 FUNCTIONAL MAPPING OF THE MOTOR CORTEX OF THE WHITE MOUSE BY A MICROSTIMULATION METHOD I. V. Pronichev and D. N. Lenkov Studies on 33 anesthetized
Neuroscience and Behavioral Physiology, Vol. 28, No. 1, 1998 FUNCTIONAL MAPPING OF THE MOTOR CORTEX OF THE WHITE MOUSE BY A MICROSTIMULATION METHOD I. V. Pronichev and D. N. Lenkov Studies on 33 anesthetized
OBSERVATIONS ON SWALLOWS AND HOUSE- MARTINS AT THE NEST. BY
 (140) OBSERVATIONS ON SWALLOWS AND HOUSE- MARTINS AT THE NEST. BY R. E. MOREAU AND W. M. MOREAU. RECENT studies of the parental care by African Hinindinidae and Swifts have suggested that, in addition
(140) OBSERVATIONS ON SWALLOWS AND HOUSE- MARTINS AT THE NEST. BY R. E. MOREAU AND W. M. MOREAU. RECENT studies of the parental care by African Hinindinidae and Swifts have suggested that, in addition
Course Curriculum for Master Degree in Poultry Diseases/Veterinary Medicine
 Course Curriculum for Master Degree in Poultry Diseases/Veterinary Medicine The Master Degree in Poultry Diseases /Veterinary Medicine, is awarded by the Faculty of Graduate Studies at Jordan University
Course Curriculum for Master Degree in Poultry Diseases/Veterinary Medicine The Master Degree in Poultry Diseases /Veterinary Medicine, is awarded by the Faculty of Graduate Studies at Jordan University
OBSERVATIONS ON THE QUALITATIVE AND QUANTITATIVE STRUCTURAL CHARACTERISTICS OF THE REPTILIAN KIDNEYS.
 OBSERVATIONS ON THE QUALITATIVE AND QUANTITATIVE STRUCTURAL CHARACTERISTICS OF THE REPTILIAN KIDNEYS. ~B~SI"Y OF Nmlll,.tpj,Tb 1.11.,,)' A Thesis submitted to the university of Nairobi in partial fulfillment
OBSERVATIONS ON THE QUALITATIVE AND QUANTITATIVE STRUCTURAL CHARACTERISTICS OF THE REPTILIAN KIDNEYS. ~B~SI"Y OF Nmlll,.tpj,Tb 1.11.,,)' A Thesis submitted to the university of Nairobi in partial fulfillment
Page # Diversity of Arthropoda Crustacea Morphology. Diversity of Arthropoda. Diversity of Arthropoda. Diversity of Arthropoda. Arthropods, from last
 Arthropods, from last time Crustacea are the dominant marine arthropods Crustacea are the dominant marine arthropods any terrestrial crustaceans? Should we call them shellfish? sowbugs 2 3 Crustacea Morphology
Arthropods, from last time Crustacea are the dominant marine arthropods Crustacea are the dominant marine arthropods any terrestrial crustaceans? Should we call them shellfish? sowbugs 2 3 Crustacea Morphology
Flight patterns of the European bustards
 Flight patterns of the European bustards By Vhilip J. Stead THE BUSTARDS, as a family, are terrestial birds and spend the major part of their time on the ground, but both the Great Bustard Otis tarda and
Flight patterns of the European bustards By Vhilip J. Stead THE BUSTARDS, as a family, are terrestial birds and spend the major part of their time on the ground, but both the Great Bustard Otis tarda and
Australian and New Zealand College of Veterinary Scientists. Membership Examination. Veterinary Radiology (Small Animal) Paper 1
 Paper 1") Australian and New Zealand College of Veterinary Scientists Membership Examination June 2016 Veterinary Radiology (Small Animal) Paper 1 Perusal time: Fifteen (15) minutes Time allowed: Two (2) hours after
Australian and New Zealand College of Veterinary Scientists Membership Examination June 2016 Veterinary Radiology (Small Animal) Paper 1 Perusal time: Fifteen (15) minutes Time allowed: Two (2) hours after
specific innervation of the muscle, so that when the nerves of a fast and of a
 Quart. J. exp. Phy8iol. (1967) 52, 293-304 THE DIFFERENTIATION OF CONDUCTION VELOCITIES OF SLOW TWITCH AND FAST TWITCH MUSCLE MOTOR INNERVATIONS IN KITTENS AND CATS. By R. M. A. P. RIDGE.* From the Physiology
Quart. J. exp. Phy8iol. (1967) 52, 293-304 THE DIFFERENTIATION OF CONDUCTION VELOCITIES OF SLOW TWITCH AND FAST TWITCH MUSCLE MOTOR INNERVATIONS IN KITTENS AND CATS. By R. M. A. P. RIDGE.* From the Physiology
Do the traits of organisms provide evidence for evolution?
 PhyloStrat Tutorial Do the traits of organisms provide evidence for evolution? Consider two hypotheses about where Earth s organisms came from. The first hypothesis is from John Ray, an influential British
PhyloStrat Tutorial Do the traits of organisms provide evidence for evolution? Consider two hypotheses about where Earth s organisms came from. The first hypothesis is from John Ray, an influential British
BSE Update Meat Industry Perspective. Randall Huffman, Ph.D. V.P. Scientific Affairs American Meat Institute Foundation
 BSE Update Meat Industry Perspective Randall Huffman, Ph.D. V.P. Scientific Affairs American Meat Institute Foundation Tuesday, December 23 USDA Announcement Overview BSE and how it spreads Control measures
BSE Update Meat Industry Perspective Randall Huffman, Ph.D. V.P. Scientific Affairs American Meat Institute Foundation Tuesday, December 23 USDA Announcement Overview BSE and how it spreads Control measures
posterior part of the second segment may show a few white hairs
 April, 1911.] New Species of Diptera of the Genus Erax. 307 NEW SPECIES OF DIPTERA OF THE GENUS ERAX. JAMES S. HINE. The various species of Asilinae known by the generic name Erax have been considered
April, 1911.] New Species of Diptera of the Genus Erax. 307 NEW SPECIES OF DIPTERA OF THE GENUS ERAX. JAMES S. HINE. The various species of Asilinae known by the generic name Erax have been considered