Department of Life Sciences, Ecology Section, University of Alcalá, Madrid, Spain;
|
|
|
- Elijah Daniel
- 5 years ago
- Views:
Transcription
1 1 Red tails are effective decoys for avian predators 2 3 Belén Fresnillo a, Josabel Belliure a, José Javier Cuervo b a Department of Life Sciences, Ecology Section, University of Alcalá, Madrid, Spain; b Department of Evolutionary Ecology, Museo Nacional de Ciencias Naturales (CSIC), Madrid, Spain Corresponding author: Belén Fresnillo Department of Life Sciences, Ecology Section, University of Alcalá, University Campus, Alcalá de Henares, Madrid, Spain belen.fresnillo@edu.uah.es Running title: Red tails are effective decoys for avian predators Key words: Acanthodactylus erythrurus, attack diversion, predator-prey interaction, evolution of signals Total words: Introduction: 692, Material and Methods: 1 967, Results: 644, Discussion: Cited references: 61 Figures: 3 Tables: 2 1
2 26 Abstract The decoy or deflection hypothesis, which states that conspicuous colouration is present in non-vital parts of the body to divert attacks from head and trunk, thus increasing survival probability, is a possible explanation for the presence of such colouration in juveniles of non-aposematic species. To test this hypothesis we made plasticine and plaster lizard models of two colour morphs, red or dark-and-light striped tails, based on the colouration of spiny-footed lizard (Acanthodactylus erythrurus) hatchlings, which naturally show a dark-and-light striped dorsal pattern and red tail. Lizard models were placed in the field and also presented to captive common kestrels (Falco tinnunculus), a common avian lizard predator. The number of attacks and the body part attacked (tail or rest-of-body) were recorded, as well as the latency to attack. Our results suggest that models of both colour morphs were recognized as prey and attacked at a similar rate, but in the field, red-tailed models were detected, and thus attacked, sooner than stripedtailed. Despite this increase in detection rate by predators, red-tailed models effectively diverted attacks to the tail from the more vulnerable body parts, thus supporting the decoy hypothesis. Greater fitness benefits of attack diversion to the tail compared to the costs of increased detection rate by predators would explain the evolution and maintenance of red tail colouration in lizards. 2
3 45 Introduction Understanding the function of conspicuous colouration in prey has attracted much interest because it may also increase detection by predators (Arnold 1984; Endler 1980; Haskell 1996). Conspicuous colouration may be a consequence of sexual selection in adults (Darwin 1871; Kemp and Rutowski 2011; Stuart-Fox et al. 2003) because it may improve mating success. Because conspicuous colouration is sometimes also present in juveniles (Booth 1990), and sexual selection cannot be driving colour evolution in such cases, at least three alternative hypotheses have been proposed to explain its evolution. The inter-age-class signalling hypothesis proposes that conspicuous colouration in juveniles evolves to reduce aggression from conspecific adults (Clark and Hall 1970; Ochi and Awata 2009; VanderWerf and Freed 2003). Conspicuous colouration in juveniles may also advertise to predators prey toxicity or unpalatability (aposematism hypothesis), or may mimic an aposematic prey model (Booth 1990; Kraus and Allison 2009). Lastly, conspicuous colouration of non-vital body parts may divert attacks from vital parts (decoy or deflection hypothesis; Bateman et al. 2014; Johnson et al. 2008; Kodandaramaiah et al. 2013; Telemeco et al. 2011; Watson et al. 2012). This function as a lure can be reinforced by stereotyped movements of the conspicuously coloured body part (Telemeco et al. 2011). The decoy hypothesis is more plausible in species where conspicuous colouration is present in expendable body parts. This is taken to the extreme in reptiles, where juveniles and/or adults of many lizard species show conspicuous colouration in the tail (Castilla et al. 1999; Clark and Hall 1970; Hawlena et al. 2006; Watson et al. 2012), which can be easily detached from the rest of the body, thus facilitating the escape from predators, although its loss might eventually lead to increased risk of 3
4 mortality (Arnold 1998; Fox and McCoy 2000; Wilson 1992; review in Bateman and Fleming 2009). Although conspicuous colouration very often increases detection by predators (Arnold 1984; Bateman et al. 2014; Endler 1980; Haskell 1996; but see Castilla et al. 1999; Watson et al. 2012), this signal may still be selected if the benefits of escaping an attack exceed the costs of increased attack rate (Cooper and Vitt 1991). The first studies addressing the decoy hypothesis in lizards were conducted in captivity (Clark and Hall 1970; Cooper and Vitt 1985; Vitt and Cooper 1986), but the use of plasticine (Mochida 2011; Valkonen et al. 2011; Watson et al. 2012) or plaster (Baylis et al. 2012; Ruiz-Rodríguez et al. 2013) models has allowed experiments in the field with different vertebrate species, obtaining information on predator attacks either by video recording or by inferring the attacks by the marks left on the models. The efficiency of blue and green lizard tails in diverting attacks has already been tested (Castilla et al. 1999; Cooper and Vitt 1985; Watson et al. 2012), but there are a number of species from several lizard families (e.g., Scincidae, Gymnophthalmidae, Lacertidae) that have red tails. Avian predators are known to discriminate colours (Bowmaker 2008), and different colours might cause different responses in bird predators, either because of differences in conspicuousness (e.g., Schaefer et al. 2006) or because of a sensory bias for certain colour (e.g., Møller and Erritzøe 2010). Consequently, results obtained when testing the decoy hypothesis in blue and green tails might not apply to red tails. The effectiveness of red tails as decoys has not yet been addressed. In the present study, our aim was to answer two specific questions: (1) do red tails make lizards more detectable to avian predators? and (2) are red tails effective decoys for diverting bird attacks from vital body parts? To answer these questions we used plasticine and plaster models based on the morphology of spiny-footed lizards (Acanthodactylus erythrurus, Schinz 1833), a species in which hatchlings have red tails 4
5 and dark-and-light striped bodies. We checked whether birds attacked red-tailed models (striped body and red tail) more and/or sooner than those with striped tails (striped body and tail), and whether red-tailed models received more attacks on the tail than stripedtailed. Models were placed in the field and also presented to captive common kestrels (Falco tinnunculus, Linnaeus 1758), one of the main avian predators of A. erythrurus (Martín and López 1990) Materials and methods Study species A. erythrurus is a medium-sized lizard (snout-vent length (SVL) and total length up to around 80 and 230 mm, respectively, although size can vary among the populations (Carretero and Llorente 1993; Seva Román 1982)) present in southern and central Iberian Peninsula and northern Africa (Belliure 2006). In populations in central Spain, new-born lizards appear in mid-august and are active until November. At the beginning of April the following spring, these lizards become active again, and they do not reach sexual maturity until their second spring (Castilla et al. 1992; Pollo and Pérez-Mellado 1990). Therefore, all individuals can be classified in three age classes, hereafter referred to as hatchlings (from hatching until the first winter), juveniles (from first to second winter) and adults (from the second winter onwards). Colouration in this species undergoes ontogenetic changes. Hatchlings show a dorsal pattern of strongly contrasting dark and light bands that run the length of their bodies. This pattern starts reticulating in juveniles, and becomes dark and light patches 5
6 in adults (Seva Román 1982). The ventrolateral parts of hatchling and juvenile tails are red (Carretero and Llorente 1993; Seva Román 1982), and this colour is clearly visible from above in hatchlings and small juveniles (personal observation). The rear part of the hind limbs of juveniles is also red. Juvenile males lose their red colouring when they are around one year old, whereas juvenile females retain it through adulthood until they become gravid (Cuervo and Belliure 2013; Seva Román 1982) Lizard models We made some lizard models with plasticine and some with plaster, because both types of models have been widely used in similar studies in various taxa (Castilla et al. 1999; Castilla and Labra 1998; Mochida 2011; Ruiz-Rodríguez et al. 2013; Valkonen et al. 2011; Vervust et al. 2007; Watson et al. 2012). Plaster models were made using plastic lizard toys which we covered with a thin layer of plaster (Aquaplast Standard, Beissier S.A., Guipúzcoa, Spain). These models were larger than real hatchlings (plastic lizard toys: mean SVL ± SD = 56.6 ± 7.9 mm, mean total length ± SD = ± 31.4 mm, N = 10; hatchlings: mean SVL ± SD = 38.3 ± 2.6 mm, mean total length ± SD = ± 10.7 mm, N = 23; t-student test, t , P in the two tests), but both juveniles and adults are common prey of avian predators (Belliure 2006), so we think these models could still be easily recognized as prey. Plasticine models were made by hand from black oil-based modelling clay (Plastilina Jovi, Barcelona, Spain) and attempted to mimic the size of real hatchlings (random sample of models: mean SVL ± SD = 38.4 ± 2.8 mm, mean total length ± SD = ± 7.9 mm, N = 15; comparison with hatchlings: t-student test, 0.00 t , P in the two tests). 6
7 Model colouring was based on real A. erythrurus hatchlings. We used non-toxic paints (Satin Paint, La Pajarita, Manises, Spain) to create two colour morphs that differed in tail colouration: red-tailed models and striped-tailed models (Fig. 1). In both colour morphs, the back of the body consisted of a contrasting pattern of dark and light bands (Fig. 1). Neither the paints used in the experiment nor real lizard dorsal colouration reflect UV light (B. Fresnillo, J. Belliure and J. J. Cuervo, unpublished data), so differences in colour discrimination by birds and humans was probably not an issue in this study. In both plaster and plasticine models, it was possible to identify the type of predator by the marks left on the model based on previous studies (Baylis et al. 2012; Brodie 1993; Valkonen et al. 2011; see Fig. 2). As plaster is harder than plasticine, we are aware that some avian attacks might not have left any mark on the plaster models. We only recorded avian attacks in this study because birds are mainly visually-directed predators, whereas mammals and reptiles also rely on chemical cues (Greene 1988). Materials used to make the models are not odourless and certainly do not smell like lizards, which might have affected the predatory behaviour of mammals and reptiles Field study We distributed lizard models when hatchlings were active (August-November) in two different years (2010 and 2011) and in two localities in central Spain where A. erythrurus is common. One study area was located in Chapinería, in south-western Region of Madrid (N40º22 ; W4º13 ), a Mediterranean oak forest with a meadow structure, where oaks (Quercus ilex L.) and lavender (Lavandula stoechas Lam.) 7
8 dominated vegetation patches surrounded by open areas. The other study area was located in Aranjuez, in southern Region of Madrid (N40º1 ; W3º33 ), a steppe dominated by esparto grass (Stipa tenacissima L.) where trees were scarce and the space surrounding plants was mainly bare soil. The study areas were 67 km apart in a straight line, and were selected because vegetation cover differed dramatically (many fewer trees in Aranjuez), which may have influenced prey detectability by visually-directed predators (Cuadrado et al. 2001; Vásquez et al. 2002). Moreover, both study areas might also have different predator communities. The study was replicated in two different habitats because the effect of model colouration on avian predation rate might depend on the habitat (a stronger effect was expected in more open habitats or where the community of avian predators was more abundant). In both localities and years, models were placed in open areas near bushes (as if they were basking lizards), at least 100 m apart from one another, and alternating both colour morphs. When both plaster and plasticine models were present in the same study area, locations of the two model types were randomized. In 2010, we worked only in Chapinería and only with plaster models. We distributed an equal number of models of both colour morphs and inspected them every 4-5 days to record the number of models attacked by birds and the body parts attacked (tail and/or rest-of-body). A total of ten models were always present in the field, as a new model was added in a new location when any model was removed due to an avian attack or after four visits without avian attacks. This makes a maximum of 20 days of exposition in the field for these models. In 2011, we worked in both localities, using plasticine and plaster models in Chapinería but only plasticine models in Aranjuez. Both study areas were visited every 3 days, and models were left in each area, always evenly distributed between both colour 8
9 morphs. The protocol for recording attacks and replacing models was the same as in 2010, so models were exposed to predators a maximum of 12 days in both localities. A total of 189 lizard models were placed in the field, but 85 of them (two plaster and 83 plasticine models) disappeared. We considered these models as attacked (in both body parts) if there were bird tracks (Fig. 2g) on the ground (11 models), or as disappeared if there were no clear bird tracks (74 models). Lizard models considered as disappeared were excluded from statistical analyses. Most plasticine models that disappeared were very probably eaten by ants Captivity study F. tinnunculus is a bird of prey in the Falconidae family that is widely distributed across Eurasia and Africa (BirdLife International 2014). It commonly preys on lizards, at least in some areas (Carrillo and González-Dávila 2009; Carrillo et al. 1994; Vanzyl 1994), and is a typical predator of A. erythrurus in Spain (Martín and López 1990), what makes this species a good model organism for studying predatory interactions with our lizard models. For this study, we used captive F. tinnunculus from the Santa Faz Wildlife Rehabilitation Centre (Alicante, Spain) from August to October in three consecutive years (2011, 2012 and 2013). Only birds that were able to fly were involved in the experiment, although some of them had sustained wing injuries. We conducted a total of 31 trials in which an F. tinnunculus (which had not been fed for 24 hours before the trial) was presented with a plasticine lizard model that was either red-tailed (N = 15) or striped-tailed (N = 16). Each bird was used only once. Trials were conducted in 9
10 individual cages (4.80 x 1.65 x 3.20 m). The lizard model was placed in the centre of the concrete cage floor at the same time that the bird was placed on a perch about 3 m from the lizard model and 1.3 m off the floor. Models were checked for attacks from outside the cage two or three times per day. The observer only entered the cage when an attack on the model was suspected. The trial ended when the model was attacked or after four days without attacks. If the model was attacked, the body part attacked, i.e., with marks (tail or rest-of-body), and the latency to attack in days were recorded Statistical analyses In the field study, Logistic Regression Models (GLZs) with binomial error distribution were used to test for any effect of model colour morph (red or striped tail) on (1) the probability of being attacked and (2) the probability of being attacked on each body part (tail or rest-of-body). In the first analysis, all the models not counted as disappeared (N = 115) were included, although the effect of model colour morph was qualitatively identical if models disappearing without the presence of bird tracks were considered attacked and included in the analysis (N = 189). For the second analysis, we included only lizard models that had been attacked by birds on a single body part (N = 38), because when there were marks on both parts, it was impossible to know which part was attacked first. To test for the possible effect of model colour morph on the latency to attack, a Generalized Linear Model (GLM) with Poisson error distribution was used. Only models attacked by birds (N = 63) were included in this analysis. All analyses in the field study simultaneously tested for differences between study areas (Chapinería and Aranjuez) and types of models (plaster and plasticine), and also checked for any 10
11 interaction between these two factors and model colour morph. Year was excluded from our statistical analyses because it was strongly associated with the type of model in all data subsets used here (Fisher's exact test; P < in the three tests), but when year was included instead of type of model, results were qualitatively identical (see Results). In the captivity study, GLZs with binomial error distribution were used to test for the possible effects of model colour morph (red or striped tail) on (1) the probability of being attacked, and (2) the probability of being attacked on each body part (tail or rest-of-body). All models (N = 31) were used in the first analysis, whereas in the second, only lizard models that had been attacked on a single body part were included (N = 16). To test for the possible effect of model colour morph on the latency to attack, a GLM with Poisson error distribution was used. Only attacked models (N = 22) were included in this analysis. All analyses in the captivity study simultaneously tested for differences in years (2011, 2012 and 2013) and checked the interaction between model colour morph and year. In both studies, a backward stepwise procedure was used in all analyses, retaining only terms associated with P values below 0.10 in the final statistical models. If Akaike s information criterion (Burnham and Anderson 2002) was used to select final models, results were qualitatively identical in all analyses (results not shown for brevity). All statistical analyses were carried out using R (R Development Core Team 2013) and lme4 package (Bates et al. 2012). All tests were two-tailed and the significance level was set to Results
12 269 Field study Excluding models that were counted as disappeared, we recorded data for 115 models, 31 plaster (14 with red tails and 17 with striped tails) and 84 plasticine (48 with red tails and 36 with striped tails). A total of 63 models were attacked by birds. Of the attacked models, 38 were attacked on one body part, whereas the rest (25 models) were attacked in both body parts (tail and rest-of-body). Taking into account the total number of models placed in the field (not including models replacing those attacked by non-avian predators), 39.16% of them were counted as disappeared, 33.33% were attacked by birds, and 27.51% were not attacked by birds. The probability of being attacked by birds was not significantly different for red and striped-tailed lizard models (Likelihood test; χ 2 = 0.15, N = 115, P = 0.697; Table 2), types of models (plaster or plasticine), or study areas (Chapinería or Aranjuez), as none of these factors was retained in the final model. However, red-tailed models were attacked sooner than striped-tailed, and plasticine models were attacked sooner than plaster (Table 1, Fig. 3a). The interaction between model colour morph and type of model was statistically significant (Table 1), implying that the difference between colour morphs depended on the type of model. In separate analyses of plaster and plasticine models, we found that red-tailed plaster models were attacked sooner than striped-tailed (Likelihood test; χ 2 = 36.51, N = 19, P < 0.001), but for plasticine models, the difference was not statistically significant (Likelihood test; χ 2 = 2.44, N = 44, P = 0.118; Fig. 3a). When year was included in this analysis instead of type of model, redtailed models were also attacked sooner than striped-tailed (Likelihood test; χ 2 = 24.82, N = 63, P < 0.001). Models were attacked sooner in 2011 than in 2010 (Likelihood test; χ 2 = , N = 63, P < 0.001), and the statistically significant interaction between 12
13 model colour morph and year (Likelihood test; χ 2 = 8.81, N = 63, P = 0.003) implied that the difference between colour morphs depended on the year. We analysed data for both years separately and found that red-tailed models were attacked sooner than striped-tailed in 2010 (Likelihood test; χ 2 = 31.75, N = 16, P < 0.001), but the difference was not statistically significant in 2011 (Likelihood test; χ 2 = 1.88, N = 47, P = 0.170). The proportion of attacks to each body part (tail or rest-of-body) was different for red and striped-tailed models, as the probability of being attacked on the tail was higher for red-tailed models (Likelihood test; χ 2 = 3.91, N = 38, P = 0.048; Table 2). Neither type of model nor study area had a significant effect on the part of the body that was attacked, as none of these factors or their interactions was retained in the final model Captivity study A total of 31 models were presented to the birds, and 22 of them were attacked. Of the attacked models, 16 were attacked on one body part, whereas the rest (6 models) were attacked on both body parts. The probability of being attacked by a captive F. tinnunculus was not significantly different for red or striped-tailed lizard models (Likelihood test; χ 2 = 0.08, N = 31, P = 0.779; Table 2), and did not differ significantly among years either, as no factor was retained in the final model. Nor did we find significant differences in the latency to attack between model colour morphs (Likelihood test; χ 2 = 0.17, N = 22, P = 0.680; Fig. 3b), or among years, as no factor was retained in the final model. The part of the body (tail or rest-of-body) that was 13
14 attacked in red and striped-tailed lizard models did not differ significantly either (Likelihood test; χ 2 = 2.08, N = 16, P = 0.150; Table 2) Discussion Our first goal was to find out whether red tails increase lizard detection rate by avian predators. The field study showed that red-tailed models were attacked sooner than striped-tailed, suggesting that the red colouring did indeed make models more easily detected by avian predators. Previous studies checking the decoy hypothesis with green (Castilla et al. 1999) and blue (Watson et al. 2012) tails concluded that they were not significantly more detected by predators than brown or black, respectively. However, recent studies found that lizard models with blue tails were attacked sooner and more often than models with brown tails (Bateman et al. 2014). Blue and green are relatively common colours among lizards (Arnold 1984; Pianka and Vitt 2006), possibly because they are inconspicuous at long distances against a background of vegetation, although they might serve as a lure at closer distances (Arnold 1984, Bateman et al. 2014). Red, however, is very conspicuous against a background of vegetation (e.g., Schaefer et al. 2006), which may explain why red-tailed models in our study were more easily detected, although other mechanisms such as a possible pre-existing sensory bias in avian predators towards red objects (e.g., Møller and Erritzøe 2010) could also explain the results. Determining the precise mechanism underlying the behaviour observed (avian predators attacking red-tailed models sooner) is beyond the scope of this study, but our results suggest that red colouration attracts bird attention and probably increase predatory pressure on red-tailed lizards. A significant interaction between colour morph 14
15 and type of model was also found, but, unfortunately, our experimental design does not allow us to elucidate whether latency to attack differed between types of models or between years. In free-ranging lizards, dorsal colour pattern is very important for crypsis (Calsbeek and Cox 2012, Lancaster et al. 2007), and we assumed that striped-tailed models matched background colour better than red-tailed. Therefore, a higher rate of attacks on red-tailed models was expected. Our results did not confirm this expectation, as models of both colour morphs were subject to a similar number of attacks in the field study. The fact that red-tailed models were attacked sooner but not more often than striped-tailed is counterintuitive, but the long periods between consecutive model inspections (3-5 days) might have contributed to the mismatch between timing and number of attacks. The similar attack rate for both colour morphs suggests that both were recognized as prey by avian predators. The captivity study showed similar attack rates and latency to attack for lizard models of both colour morphs, but this was expected due to the experimental setup (models of both colour morphs were probably very easily detected in relatively small cages with bare concrete floor). Our second aim was to check whether red tails are effective decoys for diverting bird attacks from vital body parts. The field study corroborated this hypothesis, as redtailed models received a larger proportion of attacks to the tail than striped-tailed models. Birds usually attack the head and body of their prey (Greene 1988; Shepard 2007; Vervust et al. 2011), so red tails seem to cause a change in avian predatory behaviour, thus increasing lizard probability of surviving an attack, particularly when the tail can easily be detached from the rest of the body (even though loss of the tail may imply later costs; see the Introduction). The benefits of having red tails might then outweigh the costs of increased detection by predators (Cooper and Vitt 1991), what 15
16 might have driven the evolution and maintenance of this trait in A. erythrurus. In the captivity study, a similar pattern of a larger proportion of attacks on the tail was observed for red-tailed models, but the difference between both colour morphs was not statistically significant, maybe in part because of the small sample size. The present study suggests that red tails in lizards are effective decoys for avian predators, a function that has also been suggested for green and blue tails (Bateman et al. 2014; Castilla et al. 1999; Watson et al. 2012). The tail colour developed in different lizard species does not seem to be strongly phylogenetically constrained, as the same colour may be present in species of different families (e.g., red tails are found in Morethia ruficauda, Scincidae family, Vanzosaura rubricauda, Gymnophthalmidae family, or Acanthodactylus erythrurus, Lacertidae family), whereas different colours may be present in species of the same genus (e.g., Acanthodactylus erythrurus and A. schreiberi juveniles have red tails, and A.boskianus and A. beershebensis juveniles have blue tails). Therefore, the specific tail colouration of each species might be more influenced by environmental variables (e.g., contrast with background (Håstad et al. 2005; Rosenblum 2006)), other lizard traits (e.g., contrast with the rest of the body (Arnold 1984)), or even the predator s visual characteristics (e.g., different avian taxa can discriminate wavelengths at slightly different spectral ranges (Håstad et al. 2005)). If conspicuously coloured lizard tails are an effective decoy for avian predators, we wonder why some lizard species lose this anti-predatory mechanism during the ontogenetic process (Booth 1990). A. erythrurus lizards, for example, lose the red colouration when they reach sexual maturity in the case of males, or when they become gravid in the case of females (Cuervo and Belliure 2013; Seva Román 1982). This ontogenetic colour change might be due to differences in behaviour between age classes in A. erythrurus (B. Fresnillo, J. Belliure, and J. J. Cuervo, unpublished data) and other 16
17 lizard species (Hawlena 2009; Hawlena et al. 2006), as hatchlings and juveniles are usually more active than adults and, therefore, more vulnerable to predators (Jackson et al. 1976). For age classes with high activity rate, the development of an anti-predatory mechanism such as a lure to divert attacks to expendable body parts may be advantageous. However, adults might increase their survival rates by developing more cryptic colouration (Arnold 1984; Vitt and Cooper 1986). In conclusion, this study suggests that lizards with conspicuous red tails are more easily detected by avian predators, but also that red tails effectively divert avian predator attacks from vital body parts to the expendable tail. If the benefits of diverting attacks to the tail exceed the costs of being more detectable by predators, red tails have a positive net effect on survival and, thus, on fitness, then favouring the evolution and maintenance of this conspicuous colouration. Therefore, our results support the decoy or deflection hypothesis as an explanation for conspicuous red colouration in juvenile lizards Acknowledgments We thank C. Esteban, J. Calatayud, M. Cruz, M. Almarcha and C. Zaragoza for their help in the field study, and the staff of the Santa Faz Wildlife Rehabilitation Centre (Alicante, Spain) for their help in the captivity study. Deborah Fuldauer revised English language usage. These studies were funded by the Spanish Ministry of Education and Science and the European Regional Development Fund (Grant CGL /BOS). 17
18 416 References Arnold EN (1984) Evolutionary aspects of tail shedding in lizards and their relatives. Journal of Natural History 18: Arnold EN (1988) Caudal autotomy as a defense. In: Gans C, Huey RB (eds) Biology of the Reptilia. Vol. 16. Ecology B: Defense and Life History. Alan R. Liss, Inc., New York, USA, pp Bateman PW, Fleming PA (2009) To cut a long tail short: a review of lizard caudal autotomy studies carried out over the last 20 years. Journal of Zoology 277:1-14 Bateman PW, Fleming PA, Rolek B (2014) Bite me: blue tails as a risky-decoy defense tactic for lizards. Current Zoology 60: Bates D, Maechler M, Bolker B (2012) lme4: Linear mixed-effects models using S4 classes, R package version edn. Baylis SM, Cassey P, Hauber ME (2012) Capsaicin as a deterrent against introduced mammalian nest predators. The Wilson Journal of Ornithology 124: Belliure J (2006) Lagartija colirroja Acanthodactylus erythrurus. In: Carrascal LM, Salvador A (eds) Enciclopedia Virtual de los Vertebrados Españoles. Museo Nacional de Ciencias Naturales, Madrid BirdLife International (2014) Species factsheet: Falco tinnunculus. Available from (accessed January 2014) Booth CL (1990) Evolutionary significance of ontogenic colour-cange in animals. Biological Journal of the Linnean Society 40:
19 Bowmaker JK (2008) Evolution of vertebrate visual pigments. Vision Research 48: Brodie ED III (1993) Differential avoidance of coral snake banded patterns by freeranging avian predators in Costa Rica. Evolution 47: Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer-Verlag New York, Inc., New York Calsbeek R, Cox RM (2012) An experimental test of the role of predators in the maintenance of a genetically based polymorphism. Journal of Evolutionary Biology 25: Carretero MA, Llorente GA (1993) Morphometry in a community of Mediterranean lacertid lizards, and its ecological relationships. Historia Animalium 2:77-99 Carrillo J, González-Dávila E (2009) Latitudinal variation in breeding parameters of the common kestrel Falco tinnunculus. Ardeola 56: Carrillo J, Hernández EC, Nogales M, Delgado G, García R, Ramos T (1994) Geographic variation in the spring diet of Falco tinnunculus L. on the islands of Fuerteventura and El Hierro (Canary Islands). Bonner Zoologische Beiträge 45:39-48 Castilla AM, Labra A (1998) Predation and spatial distribution of the lizard Podarcis hispanica atrata: an experimental approach. Acta Oecologica 19: Castilla AM, Barbadillo LJ, Bauwens D (1992) Annual variation in reproductive traits in the lizard Acanthodactylus erythrurus. Canadian Journal of Zoology 70:
20 Castilla AM, Gosá A, Galán P, Pérez-Mellado V (1999) Green tails in lizards of the genus Podarcis: do they influence the intensity of predation? Herpetologica 55: Clark DRJ, Hall RJ (1970) Function of the blue tail coloration of the five-lined skink Eumeces fasciatus. Herpetologica 26: Cooper WE Jr, Vitt LJ (1985) Blue tails and autotomy: enhancement of predation avoidance in juvenile skinks. Zeitschrift für Tierpsychologie 70: Cooper WE Jr, Vitt LJ (1991) Influence of detectability and ability to escape on natural selection of conspicuous autotomous defenses. Canadian Journal of Zoology 69: Cuadrado M, Martín J, López P (2001) Camouflage and escape decisions in the common chameleon Chamaeleo chamaeleon. Biological Journal of the Linnean Society 72: Cuervo JJ, Belliure J (2013) Exploring the function of red colouration in female spinyfooted lizards (Acanthodactylus erythrurus): patterns of seasonal colour change. Amphibia-Reptilia 34: Darwin C (1871) The descent of man, and selection in relation to sex. D. Appleton and Company, New York Endler JA (1980) Natural selection on color patterns in Poecilia reticulata. Evolution 34:76-91 Fox SF, McCoy JK (2000) The effects of tail loss on survival, growth, reproduction, and sex ratio of offspring in the lizard Uta stansburiana in the field. Oecologia 122:
21 Greene H (1988) Antipredator mechanisms in reptiles. In: Gans C, Huey RB (eds) Biology of the Reptilia. Vol. 16. Ecology B: Defense and Life History. Alan R. Liss, Inc., New York, USA, pp Haskell DG (1996) Do bright colors at nests incur a cost due to predation? Evolutionary Ecology 10: Hawlena D (2009) Colorful tails fade when lizards adopt less risky behaviors. Behavioral Ecology and Sociobiology 64: Hawlena D, Boochnik R, Abramsky Z, Bouskila A (2006) Blue tail and striped body: why do lizards change their infant costume when growing up? Behavioral Ecology 17: Håstad O, Victorsson J, Ödeen A (2005) Differences in color vision make passerines less conspicuous in the eyes of their predators. Proceedings of the National Academy of Sciences of the United States of America 102: Jackson JF, Ingram W III, Campbell HW (1976) The dorsal pigmentation pattern of snakes as an antipredator strategy: a multivariate approach. The American Naturalist 110: Johnson J, Burt DB, DeWitt TJ (2008) Form, function, and fitness: pathways to survival. Evolution 62: Kemp DJ, Rutowski RL (2011) The role of coloration in mate choice and sexual interactions in butterflies. Advances in the Study of Behavior 43:55-92 Kodandaramaiah U, Lindenfors P, Tullberg BS (2013) Deflective and intimidating eyespots: a comparative study of eyespot size and position in butterflies. Ecology and Evolution 3:
22 Kraus F, Allison A (2009) A remarkable ontogenetic change in color pattern in a new species of Oreophryne (Anura: Microhylidae) from Papua New Guinea. Copeia 2009: Lancaster LT, McAdam AG, Wingfield JC, Sinervo BR (2007) Adaptive social and maternal induction of antipredator dorsal patterns in a lizard with alternative social strategies. Ecology Letters 10: Martín J, López P (1990) Amphibians and reptiles as prey of birds in Southwestern Europe. Smithsonian Herpetological Information Service 82:1-43 Mochida K (2011) Combination of local selection pressures drives diversity in aposematic signals. Evolutionary Ecology 25: Møller AP, Erritzøe J (2010) Why birds eat colourful grit: colour preferences revealed by the colour of gizzard stones. Journal of Evolutionary Biology 23: Ochi H, Awata S (2009) Resembling the juvenile colour of host cichlid facilitates access of the guest cichlid to host territory. Behaviour 146: Pianka ER, Vitt LJ (2006) Lizards: Windows to the Evolution of Diversity. University of California Press, Berkeley, California Pollo CJ, Pérez-Mellado V (1990) Biología reproductora de tres especies mediterráneas de Lacertidae. Mediterránea, Serie de Estudios Biológicos 12: R Development Core Team (2013) R: a language and environment for statistical computing. Version R Foundation for Statistical Computing, Vienna, Austria. Rosenblum EB (2006) Convergent evolution and divergent selection: lizards at the White Sands ecotone. The American Naturalist 167:
23 Ruiz-Rodríguez M, Avilés JM, Cuervo JJ, Parejo D, Ruano F, Zamora-Muñoz C, Sergio F, López-Jiménez L, Tanferna A, Martín-Vivaldi M (2013) Does avian conspicuous colouration increase or reduce predation risk? Oecologia 173:83-93 Schaefer HM, Levey DJ, Schaefer V, Avery ML (2006) The role of chromatic and achromatic signals for fruit detection by birds. Behavioral Ecology 17: Seva Román E (1982) Taxocenosis de lacértidos en un arenal costero alicantino. Ph.D. Thesis, Universidad de Alicante, Alicante Shepard DB (2007) Habitat but not body shape affects predator attack frequency on lizard models in the Brazilian Cerrado. Herpetologica 63: Stuart-Fox DM, Moussalli A, Marshall NJ, Owens IPF (2003) Conspicuous males suffer higher predation risk: visual modelling and experimental evidence from lizards. Animal Behaviour 66: Telemeco RS, Baird TA, Shine R (2011) Tail waving in a lizard (Bassiana duperreyi) functions to deflect attacks rather than as a pursuit-deterrent signal. Animal Behaviour 82: Valkonen J, Niskanen M, Björklund M, Mappes J (2011) Disruption or aposematism? Significance of dorsal zigzag pattern of European vipers. Evolutionary Ecology 25: VanderWerf EA, Freed LA (2003) 'Elepaio subadult plumages reduce aggression through graded status-signaling, not mimicry. Journal of Field Ornithology 74: Vanzyl AJ (1994) Comparison of the diet of the common kestrel Falco tinnunculus in South Africa and Europe. Bird Study 41:
24 Vásquez RA, Ebensperger LA, Bozinovic F (2002) The influence of habitat on travel speed, intermittent locomotion, and vigilance in a diurnal rodent. Behavioral Ecology 13: Vervust B, Grbac I, Van Damme R (2007) Differences in morphology, performance and behaviour between recently diverged populations of Podarcis sicula mirror differences in predation pressure. Oikos 116: Vervust B, Van Loy H, Van Damme R (2011) Seeing through the lizard s trick: do avian predators avoid autotomous tails? Central European Journal of Biology 6: Vitt LJ, Cooper WE Jr (1986) Tail loss, tail color, and predator escape in Eumeces (Lacertilia: Scincidae): age-specific differences in costs and benefits. Canadian Journal of Zoology 64: Watson CM, Roelke CE, Pasichnyk PN, Cox CL (2012) The fitness consequences of the autotomous blue tail in lizards: an empirical test of predator response using clay models. Zoology 115: Wilson BS (1992) Tail injuries increase the risk of mortality in free-living lizards (Uta stansburiana). Oecologia 92:
25 Table 1. Generalized linear model with Poisson error distribution analysing the effects of type of model (plaster or plasticine) and model colour morph (red or striped tail) on the latency to attack. The interaction between type of model and colour morph is also shown. Study area and interactions including this factor were not retained in the final model after a backward stepwise procedure (see Statistical analyses for details). Only models attacked by birds were included in the analysis (N = 63) d.f. χ 2 P Type of model < Colour morph < Type of model x Colour morph
26 Table 2. Percentage of lizard models that were attacked by birds and percentage of attacks on the tail for red- and striped-tailed models in the field and captivity studies. Percentage of attacks on the tail was calculated considering only models attacked by birds on a single body part (tail or rest-of-body) Type of study N Red-tailed models Striped-tailed models Percentage of models attacked Field Captivity Percentage of attacks on the tail Field Captivity
27 601 Figure legends Fig. 1 Types of models used in the study: (a) red-tailed plaster model (tail completely red), (b) striped-tailed plaster model (dark and light banded tail pattern), (c) red-tailed plasticine model (red tail with a light band on the back), and (d) striped-tailed plasticine model (dark and light banded tail pattern) Fig. 2 Types of marks left on the models by predators: (a) V-shaped mark left by a bird on a plasticine model, (b) U-shaped mark left by a bird on a plasticine model, (c) puncture mark left by a bird on a plaster model, (d) V-shaped mark with tooth marks left by lizards on a plasticine model, (e) tooth marks left by rodents on a plasticine model, (f) tooth marks left by rodents on a plaster model, and (g) tracks left on the ground after an avian attack when the model disappeared Fig. 3 Mean (± SE) latency to attack for red and striped-tailed models in (a) the field study by type of model (plaster or plasticine) (N = 38) and (b) the captivity study (N = 16). Only models attacked by birds are included. 27



28 Fig. 1 28



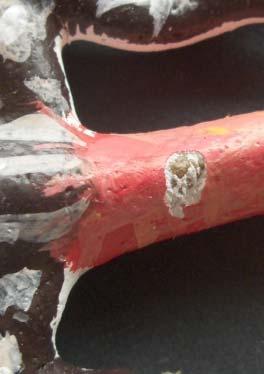


29 Fig. 2 29
30 Fig. 3 (a) Days until attack Striped tail Plaster models Plasticine models Red tail Model colour design (b) 3.0 Days until attack Striped tail Red tail Model colour design 30
Red tails are effective decoys for avian predators
 See discussions, stats, and author profiles for this publication at: http://www.researchgate.net/publication/269104580 Red tails are effective decoys for avian predators ARTICLE in EVOLUTIONARY ECOLOGY
See discussions, stats, and author profiles for this publication at: http://www.researchgate.net/publication/269104580 Red tails are effective decoys for avian predators ARTICLE in EVOLUTIONARY ECOLOGY
Introduction. Lizards: very diverse colour patterns intra- and interspecific differences in colour
 Jessica Vroonen Introduction Lizards: very diverse colour patterns intra- and interspecific differences in colour Introduction Lizards intra- and interspecific differences in colour Introduction Lizards
Jessica Vroonen Introduction Lizards: very diverse colour patterns intra- and interspecific differences in colour Introduction Lizards intra- and interspecific differences in colour Introduction Lizards
MA41 Colour variability and the ecological use of colour in the chameleons and geckos of Mahamavo
 MA41 Colour variability and the ecological use of colour in the chameleons and geckos of Mahamavo Colour and the ability to change colour are some of the most striking features of lizards. Unlike birds
MA41 Colour variability and the ecological use of colour in the chameleons and geckos of Mahamavo Colour and the ability to change colour are some of the most striking features of lizards. Unlike birds
Ciccaba virgata (Mottled Owl)
") Ciccaba virgata (Mottled Owl) Family: Strigidae (Typical Owls) Order: Strigiformes (Owls) Class: Aves (Birds) Fig. 1. Mottled owl, Ciccaba virgata. [http://www.owling.com/mottled13.htm, downloaded 12 November
Ciccaba virgata (Mottled Owl) Family: Strigidae (Typical Owls) Order: Strigiformes (Owls) Class: Aves (Birds) Fig. 1. Mottled owl, Ciccaba virgata. [http://www.owling.com/mottled13.htm, downloaded 12 November
Animal Behaviour 78 (2009) Contents lists available at ScienceDirect. Animal Behaviour. journal homepage:
 Contents lists available at ScienceDirect. Animal Behaviour. journal homepage:") Animal Behaviour 78 (2009) 1011 1018 Contents lists available at ScienceDirect Animal Behaviour journal homepage: www.elsevier.com/locate/anbehav When to run from an ambush predator: balancing crypsis
Animal Behaviour 78 (2009) 1011 1018 Contents lists available at ScienceDirect Animal Behaviour journal homepage: www.elsevier.com/locate/anbehav When to run from an ambush predator: balancing crypsis
Plestiodon (=Eumeces) fasciatus Family Scincidae
 fasciatus Family Scincidae") Plestiodon (=Eumeces) fasciatus Family Scincidae Living specimens: - Five distinct longitudinal light lines on dorsum - Juveniles have bright blue tail - Head of male reddish during breeding season - Old
Plestiodon (=Eumeces) fasciatus Family Scincidae Living specimens: - Five distinct longitudinal light lines on dorsum - Juveniles have bright blue tail - Head of male reddish during breeding season - Old
NAME: DATE: SECTION:
 NAME: DATE: SECTION: MCAS PREP PACKET EVOLUTION AND BIODIVERSITY 1. Which of the following observations best supports the conclusion that dolphins and sharks do not have a recent common ancestor? A. Dolphins
NAME: DATE: SECTION: MCAS PREP PACKET EVOLUTION AND BIODIVERSITY 1. Which of the following observations best supports the conclusion that dolphins and sharks do not have a recent common ancestor? A. Dolphins
J.K. McCoy CURRICULUM VITAE. J. Kelly McCoy. Department of Biology Angelo State University San Angelo, TX
 CURRICULUM VITAE J. Kelly McCoy Department of Biology Angelo State University San Angelo, TX 76909 325-486-6646 Kelly.McCoy@angelo.edu Education: B.S. 1990 Zoology Oklahoma State University Ph.D. 1995
CURRICULUM VITAE J. Kelly McCoy Department of Biology Angelo State University San Angelo, TX 76909 325-486-6646 Kelly.McCoy@angelo.edu Education: B.S. 1990 Zoology Oklahoma State University Ph.D. 1995
Analysis of Sampling Technique Used to Investigate Matching of Dorsal Coloration of Pacific Tree Frogs Hyla regilla with Substrate Color
 Analysis of Sampling Technique Used to Investigate Matching of Dorsal Coloration of Pacific Tree Frogs Hyla regilla with Substrate Color Madeleine van der Heyden, Kimberly Debriansky, and Randall Clarke
Analysis of Sampling Technique Used to Investigate Matching of Dorsal Coloration of Pacific Tree Frogs Hyla regilla with Substrate Color Madeleine van der Heyden, Kimberly Debriansky, and Randall Clarke
Class Reptilia Testudines Squamata Crocodilia Sphenodontia
 Class Reptilia Testudines (around 300 species Tortoises and Turtles) Squamata (around 7,900 species Snakes, Lizards and amphisbaenids) Crocodilia (around 23 species Alligators, Crocodiles, Caimans and
Class Reptilia Testudines (around 300 species Tortoises and Turtles) Squamata (around 7,900 species Snakes, Lizards and amphisbaenids) Crocodilia (around 23 species Alligators, Crocodiles, Caimans and
SOAR Research Proposal Summer How do sand boas capture prey they can t see?
 SOAR Research Proposal Summer 2016 How do sand boas capture prey they can t see? Faculty Mentor: Dr. Frances Irish, Assistant Professor of Biological Sciences Project start date and duration: May 31, 2016
SOAR Research Proposal Summer 2016 How do sand boas capture prey they can t see? Faculty Mentor: Dr. Frances Irish, Assistant Professor of Biological Sciences Project start date and duration: May 31, 2016
1.5 C: Role of the Environment in Evolution Quiz
 1. Numbers of reported cases of bedbug infestations have been increasing over the past ten years in the United States. In an attempt to combat the infestations, people began using pesticides to kill the
1. Numbers of reported cases of bedbug infestations have been increasing over the past ten years in the United States. In an attempt to combat the infestations, people began using pesticides to kill the
Impact of colour polymorphism and thermal conditions on thermoregulation, reproductive success, and development in Vipera aspis
 Impact of colour polymorphism and thermal conditions on thermoregulation, reproductive success, and development in Vipera aspis Sylvain Dubey, Johan Schürch, Joaquim Golay, Briséïs Castella, Laura Bonny,
Impact of colour polymorphism and thermal conditions on thermoregulation, reproductive success, and development in Vipera aspis Sylvain Dubey, Johan Schürch, Joaquim Golay, Briséïs Castella, Laura Bonny,
NOTES ON THE ECOLOGY AND NATURAL HISTORY OF TWO SPECIES OF EGERNIA (SCINCIDAE) IN WESTERN AUSTRALIA
 IN WESTERN AUSTRALIA") NOTES ON THE ECOLOGY AND NATURAL HISTORY OF TWO SPECIES OF EGERNIA (SCINCIDAE) IN WESTERN AUSTRALIA By ERIC R. PIANKA Integrative Biology University of Texas at Austin Austin, Texas 78712 USA Email: erp@austin.utexas.edu
NOTES ON THE ECOLOGY AND NATURAL HISTORY OF TWO SPECIES OF EGERNIA (SCINCIDAE) IN WESTERN AUSTRALIA By ERIC R. PIANKA Integrative Biology University of Texas at Austin Austin, Texas 78712 USA Email: erp@austin.utexas.edu
8/19/2013. Who eats herps? Topic 20: Predators. Who eats herps? Who eats herps? Who eats herps? Who eats herps?
 Topic 20: Predators Variation in predators across taxa Variation in predators through ontogeny How do herps avoid being eaten? Introduction to the diversity of anti-predator defenses Many animals Depends
Topic 20: Predators Variation in predators across taxa Variation in predators through ontogeny How do herps avoid being eaten? Introduction to the diversity of anti-predator defenses Many animals Depends
Suggest two features you can see in the pictures that could be used to classify these organisms (2)
") Q. (a) Organisms can be classified using features that can be seen. Organisms A, B, C, D and E below all belong to a large group called the arthropods. (i) Suggest two features you can see in the pictures
Q. (a) Organisms can be classified using features that can be seen. Organisms A, B, C, D and E below all belong to a large group called the arthropods. (i) Suggest two features you can see in the pictures
Today there are approximately 250 species of turtles and tortoises.
 I WHAT IS A TURTLE OR TORTOISE? Over 200 million years ago chelonians with fully formed shells appeared in the fossil record. Unlike modern species, they had teeth and could not withdraw into their shells.
I WHAT IS A TURTLE OR TORTOISE? Over 200 million years ago chelonians with fully formed shells appeared in the fossil record. Unlike modern species, they had teeth and could not withdraw into their shells.
6. The lifetime Darwinian fitness of one organism is greater than that of another organism if: A. it lives longer than the other B. it is able to outc
 1. The money in the kingdom of Florin consists of bills with the value written on the front, and pictures of members of the royal family on the back. To test the hypothesis that all of the Florinese $5
1. The money in the kingdom of Florin consists of bills with the value written on the front, and pictures of members of the royal family on the back. To test the hypothesis that all of the Florinese $5
Lizard malaria: cost to vertebrate host's reproductive success
 Parasilology (1983), 87, 1-6 1 With 2 figures in the text Lizard malaria: cost to vertebrate host's reproductive success J. J. SCHALL Department of Zoology, University of Vermont, Burlington, Vermont 05405,
Parasilology (1983), 87, 1-6 1 With 2 figures in the text Lizard malaria: cost to vertebrate host's reproductive success J. J. SCHALL Department of Zoology, University of Vermont, Burlington, Vermont 05405,
Colorful tails fade when lizards adopt less risky behaviors
 Behav Ecol Sociobiol (2009) 64:205 213 DOI 10.1007/s00265-009-0837-z ORIGINAL PAPER Colorful tails fade when lizards adopt less risky behaviors Dror Hawlena Received: 23 April 2009 /Revised: 30 July 2009
Behav Ecol Sociobiol (2009) 64:205 213 DOI 10.1007/s00265-009-0837-z ORIGINAL PAPER Colorful tails fade when lizards adopt less risky behaviors Dror Hawlena Received: 23 April 2009 /Revised: 30 July 2009
The role of visual cues in learning escape behaviour in the little brown skink (Scincella lateralis)
") Behaviour 151 (2014) 2015 2028 brill.com/beh The role of visual cues in learning escape behaviour in the little brown skink (Scincella lateralis) Mark A. Paulissen Department of Natural Sciences, Northeastern
Behaviour 151 (2014) 2015 2028 brill.com/beh The role of visual cues in learning escape behaviour in the little brown skink (Scincella lateralis) Mark A. Paulissen Department of Natural Sciences, Northeastern
Reptiles and amphibian behaviour
 Reptiles and amphibian behaviour Understanding how a healthy reptile and amphibian should look and act takes a lot of observation and practice. Reptiles and amphibians have behaviour that relates to them
Reptiles and amphibian behaviour Understanding how a healthy reptile and amphibian should look and act takes a lot of observation and practice. Reptiles and amphibians have behaviour that relates to them
Squamates of Connecticut
 Squamates of Connecticut Reptilia Turtles are sisters to crocodiles and birds Yeah, birds are reptiles, haven t you watched Jurassic Park yet? Lizards and snakes are part of one clade called the squamates
Squamates of Connecticut Reptilia Turtles are sisters to crocodiles and birds Yeah, birds are reptiles, haven t you watched Jurassic Park yet? Lizards and snakes are part of one clade called the squamates
Animal Adaptations. Structure and Function
 Name period date assigned date due date returned 1. What is a variation 2. What is an adaptation omplete the chart with the examples from the power point. List adaptations that help animals do the following:
Name period date assigned date due date returned 1. What is a variation 2. What is an adaptation omplete the chart with the examples from the power point. List adaptations that help animals do the following:
Station 1 1. (3 points) Identification: Station 2 6. (3 points) Identification:
 Identification: Station 2 6. (3 points) Identification:") SOnerd s 2018-2019 Herpetology SSSS Test 1 SOnerd s SSSS 2018-2019 Herpetology Test Station 20 sounds found here: https://drive.google.com/drive/folders/1oqrmspti13qv_ytllk_yy_vrie42isqe?usp=sharing Station
SOnerd s 2018-2019 Herpetology SSSS Test 1 SOnerd s SSSS 2018-2019 Herpetology Test Station 20 sounds found here: https://drive.google.com/drive/folders/1oqrmspti13qv_ytllk_yy_vrie42isqe?usp=sharing Station
Breeding White Storks( Ciconia ciconia at Chessington World of Adventures Paul Wexler
 Breeding White Storks(Ciconia ciconia) at Chessington World of Adventures Paul Wexler The White Stork belongs to the genus Ciconia of which there are seven other species incorporated predominantly throughout
Breeding White Storks(Ciconia ciconia) at Chessington World of Adventures Paul Wexler The White Stork belongs to the genus Ciconia of which there are seven other species incorporated predominantly throughout
Migration. Migration = a form of dispersal which involves movement away from and subsequent return to the same location, typically on an annual basis.
 Migration Migration = a form of dispersal which involves movement away from and subsequent return to the same location, typically on an annual basis. To migrate long distance animals must navigate through
Migration Migration = a form of dispersal which involves movement away from and subsequent return to the same location, typically on an annual basis. To migrate long distance animals must navigate through
Effect of Tail Loss on Sprint Speed and Growth in Newborn Skinks, Niveoscincus metallicus
 Effect of Tail Loss on Sprint Speed and Growth in Newborn Skinks, Niveoscincus metallicus Author(s) :David G. Chapple, Colin J. McCoull, Roy Swain Source: Journal of Herpetology, 38(1):137-140. 2004. Published
Effect of Tail Loss on Sprint Speed and Growth in Newborn Skinks, Niveoscincus metallicus Author(s) :David G. Chapple, Colin J. McCoull, Roy Swain Source: Journal of Herpetology, 38(1):137-140. 2004. Published
Mate protection in pre-nesting Canada Geese Branta canadensis
 Mate protection in pre-nesting Canada Geese Branta canadensis I. P. JOHNSON and R. M. SIBLY Fourteen individually marked pairs o f Canada Geese were observedfrom January to April on their feeding grounds
Mate protection in pre-nesting Canada Geese Branta canadensis I. P. JOHNSON and R. M. SIBLY Fourteen individually marked pairs o f Canada Geese were observedfrom January to April on their feeding grounds
A Population Analysis of the Common Wall Lizard Podarcis muralis in Southwestern France
 - 513 - Studies in Herpetology, Rocek Z. (ed.) pp. 513-518 Prague 1986 A Population Analysis of the Common Wall Lizard Podarcis muralis in Southwestern France R. BARBAULT and Y. P. MOU Laboratoire d'ecologie
- 513 - Studies in Herpetology, Rocek Z. (ed.) pp. 513-518 Prague 1986 A Population Analysis of the Common Wall Lizard Podarcis muralis in Southwestern France R. BARBAULT and Y. P. MOU Laboratoire d'ecologie
4B: The Pheasant Case: Handout. Case Three Ring-Necked Pheasants. Case materials: Case assignment
 4B: The Pheasant Case: Handout Case Three Ring-Necked Pheasants As you can see, the male ring-necked pheasant is brightly colored. The white ring at the base of the red and green head stand out against
4B: The Pheasant Case: Handout Case Three Ring-Necked Pheasants As you can see, the male ring-necked pheasant is brightly colored. The white ring at the base of the red and green head stand out against
08 alberts part2 7/23/03 9:10 AM Page 95 PART TWO. Behavior and Ecology
 08 alberts part2 7/23/03 9:10 AM Page 95 PART TWO Behavior and Ecology 08 alberts part2 7/23/03 9:10 AM Page 96 08 alberts part2 7/23/03 9:10 AM Page 97 Introduction Emília P. Martins Iguanas have long
08 alberts part2 7/23/03 9:10 AM Page 95 PART TWO Behavior and Ecology 08 alberts part2 7/23/03 9:10 AM Page 96 08 alberts part2 7/23/03 9:10 AM Page 97 Introduction Emília P. Martins Iguanas have long
The Origin of Species: Lizards in an Evolutionary Tree
 The Origin of Species: Lizards in an Evolutionary Tree NAME DATE This handout supplements the short film The Origin of Species: Lizards in an Evolutionary Tree. 1. Puerto Rico, Cuba, Jamaica, and Hispaniola
The Origin of Species: Lizards in an Evolutionary Tree NAME DATE This handout supplements the short film The Origin of Species: Lizards in an Evolutionary Tree. 1. Puerto Rico, Cuba, Jamaica, and Hispaniola
Juvenile Collared Lizards Adjust Tail Display Frequency in Response to Variable Predatory Threat
 Ethology Juvenile Collared Lizards Adjust Tail Display Frequency in Response to Variable Predatory Threat Joshua R. York* & Troy A. Baird * Department of Biology, University of Oklahoma, Norman, OK, USA
Ethology Juvenile Collared Lizards Adjust Tail Display Frequency in Response to Variable Predatory Threat Joshua R. York* & Troy A. Baird * Department of Biology, University of Oklahoma, Norman, OK, USA
Societas Europaea Herpetologica
 of 6 th Ordinary General Meeting of Societas Europaea Herpetologica 19-23 August 1991 Budapest, the the Hungary edited by Z. Korsos & I. Kiss Hungarian Natural History Museum Budapest 1992 Korsos, z.-&kis&,
of 6 th Ordinary General Meeting of Societas Europaea Herpetologica 19-23 August 1991 Budapest, the the Hungary edited by Z. Korsos & I. Kiss Hungarian Natural History Museum Budapest 1992 Korsos, z.-&kis&,
CAMBRIDGE, MASS. 26 MARCH 2010 NUMBER 519 CRUISE FORAGING OF INVASIVE CHAMELEON (CHAMAELEO JACKSONII XANTHOLOPHUS) IN HAWAI I
 IN HAWAI I") US ISSN 0006-9698 CAMBRIDGE, MASS. 26 MARCH 2010 NUMBER 519 CRUISE FORAGING OF INVASIVE CHAMELEON (CHAMAELEO JACKSONII XANTHOLOPHUS) IN HAWAI I TRAVIS J. HAGEY, 1 JONATHAN B. LOSOS, 2 AND LUKE J. HARMON
US ISSN 0006-9698 CAMBRIDGE, MASS. 26 MARCH 2010 NUMBER 519 CRUISE FORAGING OF INVASIVE CHAMELEON (CHAMAELEO JACKSONII XANTHOLOPHUS) IN HAWAI I TRAVIS J. HAGEY, 1 JONATHAN B. LOSOS, 2 AND LUKE J. HARMON
Contrasting Response to Predator and Brood Parasite Signals in the Song Sparrow (melospiza melodia)
") Luke Campillo and Aaron Claus IBS Animal Behavior Prof. Wisenden 6/25/2009 Contrasting Response to Predator and Brood Parasite Signals in the Song Sparrow (melospiza melodia) Abstract: The Song Sparrow
Luke Campillo and Aaron Claus IBS Animal Behavior Prof. Wisenden 6/25/2009 Contrasting Response to Predator and Brood Parasite Signals in the Song Sparrow (melospiza melodia) Abstract: The Song Sparrow
Biodiversity and Extinction. Lecture 9
 Biodiversity and Extinction Lecture 9 This lecture will help you understand: The scope of Earth s biodiversity Levels and patterns of biodiversity Mass extinction vs background extinction Attributes of
Biodiversity and Extinction Lecture 9 This lecture will help you understand: The scope of Earth s biodiversity Levels and patterns of biodiversity Mass extinction vs background extinction Attributes of
Parental care in the long-tailed skink, Mabuya longicaudata, on a tropical Asian island
 ANIMAL BEHAVIOUR, 2006, 72, 791e795 doi:10.1016/j.anbehav.2005.12.011 Parental care in the long-tailed skink, Mabuya longicaudata, on a tropical Asian island WEN-SAN HUANG Department of Ecology and Evolutionary
ANIMAL BEHAVIOUR, 2006, 72, 791e795 doi:10.1016/j.anbehav.2005.12.011 Parental care in the long-tailed skink, Mabuya longicaudata, on a tropical Asian island WEN-SAN HUANG Department of Ecology and Evolutionary
SKELETONS: Museum of Osteology Tooth and Eye Dentification Teacher Resource
 SKELETONS: Museum of Osteology Tooth and Eye Dentification Teacher Resource Grade Levels: 3 rd 5 th Grade 3 rd Grade: SC.3.N.1.1 - Raise questions about the natural world, investigate them individually
SKELETONS: Museum of Osteology Tooth and Eye Dentification Teacher Resource Grade Levels: 3 rd 5 th Grade 3 rd Grade: SC.3.N.1.1 - Raise questions about the natural world, investigate them individually
Weaver Dunes, Minnesota
 Hatchling Orientation During Dispersal from Nests Experimental analyses of an early life stage comparing orientation and dispersal patterns of hatchlings that emerge from nests close to and far from wetlands
Hatchling Orientation During Dispersal from Nests Experimental analyses of an early life stage comparing orientation and dispersal patterns of hatchlings that emerge from nests close to and far from wetlands
What causes lizards brains to change size?
 December 2017 What causes lizards brains to change size? GET OFF MY LAND Authors: Susan Crow, Meghan Pawlowski, Manyowa Meki, Lara LaDage, Timothy Roth II, Cynthia Downs, Barry Sinervo and Vladimir Pravosudov
December 2017 What causes lizards brains to change size? GET OFF MY LAND Authors: Susan Crow, Meghan Pawlowski, Manyowa Meki, Lara LaDage, Timothy Roth II, Cynthia Downs, Barry Sinervo and Vladimir Pravosudov
Adaptations 4. Adaptations 1 Adaptations 2
 Adaptations 1 Adaptations 2 Describe Charles Darwin s Theory of Natural Selection. Charles Darwin studied many new species and their adaptations. On which group of islands did he complete most of his research?
Adaptations 1 Adaptations 2 Describe Charles Darwin s Theory of Natural Selection. Charles Darwin studied many new species and their adaptations. On which group of islands did he complete most of his research?
Seasonal Shifts in Reproductive Investment of Female Northern Grass Lizards ( Takydromus septentrionalis
 Seasonal Shifts in Reproductive Investment of Female Northern Grass Lizards (Takydromus septentrionalis) from a Field Population on Beiji Island, China Author(s): Wei-Guo Du and Lu Shou Source: Journal
Seasonal Shifts in Reproductive Investment of Female Northern Grass Lizards (Takydromus septentrionalis) from a Field Population on Beiji Island, China Author(s): Wei-Guo Du and Lu Shou Source: Journal
Chapter 16: Evolution Lizard Evolution Virtual Lab Honors Biology. Name: Block: Introduction
 Chapter 16: Evolution Lizard Evolution Virtual Lab Honors Biology Name: Block: Introduction Charles Darwin proposed that over many generations some members of a population could adapt to a changing environment
Chapter 16: Evolution Lizard Evolution Virtual Lab Honors Biology Name: Block: Introduction Charles Darwin proposed that over many generations some members of a population could adapt to a changing environment
Scientific name: Common name: Class: Order: Suborder: Family: Etymology: Feeding behaviour: Description:
 Scientific name: Chamaeleo chamaeleon Common name: Mediterranean or Common chameleon Class: Reptilia Order: Squamata Suborder: Sauria Family: Chamaeleontidae Etymology: The name chameleon (also chamaeleon)
Scientific name: Chamaeleo chamaeleon Common name: Mediterranean or Common chameleon Class: Reptilia Order: Squamata Suborder: Sauria Family: Chamaeleontidae Etymology: The name chameleon (also chamaeleon)
*Using the 2018 List. Use the image below to answer question 6.
 Herpetology Test 1. Hearts in all herps other than consists of atria and one ventricle somewhat divided by a septum. (2 pts) a. snakes; two b. crocodiles; two c. turtles; three d. frogs; four 2. The food
Herpetology Test 1. Hearts in all herps other than consists of atria and one ventricle somewhat divided by a septum. (2 pts) a. snakes; two b. crocodiles; two c. turtles; three d. frogs; four 2. The food
Variation in predator species abundance can cause variable selection pressure on warning signaling prey
 Variation in predator species abundance can cause variable selection pressure on warning signaling prey Janne K. Valkonen 1, Ossi Nokelainen 1, Martti Niskanen 1, Janne Kilpimaa 1, Mats Björklund 2 & Johanna
Variation in predator species abundance can cause variable selection pressure on warning signaling prey Janne K. Valkonen 1, Ossi Nokelainen 1, Martti Niskanen 1, Janne Kilpimaa 1, Mats Björklund 2 & Johanna
This is the book of David, the son of Abraham and the father of Jesus Christ Matthew 1.1. Declaration
 ! ii This is the book of David, the son of Abraham and the father of Jesus Christ Matthew 1.1 Declaration This thesis is my original work and no part has been previously submitted for a degree. Chapters
! ii This is the book of David, the son of Abraham and the father of Jesus Christ Matthew 1.1 Declaration This thesis is my original work and no part has been previously submitted for a degree. Chapters
ANIMAL BEHAVIOR. Laboratory: a Manual to Accompany Biology. Saunders College Publishing: Philadelphia.
 PRESENTED BY KEN Yasukawa at the 2007 ABS Annual Meeting Education Workshop Burlington VT ANIMAL BEHAVIOR Humans have always been interested in animals and how they behave because animals are a source
PRESENTED BY KEN Yasukawa at the 2007 ABS Annual Meeting Education Workshop Burlington VT ANIMAL BEHAVIOR Humans have always been interested in animals and how they behave because animals are a source
Survivorship. Demography and Populations. Avian life history patterns. Extremes of avian life history patterns
 Demography and Populations Survivorship Demography is the study of fecundity and survival Four critical variables Age of first breeding Number of young fledged each year Juvenile survival Adult survival
Demography and Populations Survivorship Demography is the study of fecundity and survival Four critical variables Age of first breeding Number of young fledged each year Juvenile survival Adult survival
Kori Bustard Husbandry. Sara Hallager, Biologist, Smithsonian National Zoological Park
 Kori Bustard Husbandry Sara Hallager, Biologist, Smithsonian National Zoological Park Ardeotis kori 2 subspecies [?] Africa s largest flying bird Captive males: 12-19kg Seasonal weight gain up to 4kg Captive
Kori Bustard Husbandry Sara Hallager, Biologist, Smithsonian National Zoological Park Ardeotis kori 2 subspecies [?] Africa s largest flying bird Captive males: 12-19kg Seasonal weight gain up to 4kg Captive
Movement correlates of lizards dorsal pigmentation patterns
 Functional Ecology 2016 doi: 10.1111/1365-2435.12700 Movement correlates of lizards dorsal pigmentation patterns Topaz Halperin 1,2, Liran Carmel 3 and Dror Hawlena*,1,2 1 Risk-Management Ecology Lab,
Functional Ecology 2016 doi: 10.1111/1365-2435.12700 Movement correlates of lizards dorsal pigmentation patterns Topaz Halperin 1,2, Liran Carmel 3 and Dror Hawlena*,1,2 1 Risk-Management Ecology Lab,
Escape Behaviors and Flight Initiation Distance in the Common Water Snake Nerodia sipedon
 Journal of Herpetology, Vol. 42, No. 3, pp. 493 500, 2008 Copyright 2008 Society for the Study of Amphibians and Reptiles Escape Behaviors and Flight Initiation Distance in the Common Water Snake Nerodia
Journal of Herpetology, Vol. 42, No. 3, pp. 493 500, 2008 Copyright 2008 Society for the Study of Amphibians and Reptiles Escape Behaviors and Flight Initiation Distance in the Common Water Snake Nerodia
Genetics. Labrador Retrievers as a Model System to Study Inheritance of Hair Color. Contents of this Section
 Genetics Labrador Retrievers as a Model System to Study Inheritance of Hair Color Contents of this Section Unlike humans, who usually have only one child at a time, and rarely manage more than a dozen
Genetics Labrador Retrievers as a Model System to Study Inheritance of Hair Color Contents of this Section Unlike humans, who usually have only one child at a time, and rarely manage more than a dozen
Is it better to be bigger? Featured scientists: Aaron Reedy and Robert Cox from the University of Virginia Co-written by Matt Kustra
 Is it better to be bigger? Featured scientists: Aaron Reedy and Robert Cox from the University of Virginia Co-written by Matt Kustra Research Background: When Charles Darwin talked about the struggle for
Is it better to be bigger? Featured scientists: Aaron Reedy and Robert Cox from the University of Virginia Co-written by Matt Kustra Research Background: When Charles Darwin talked about the struggle for
Unit 7: Adaptation STUDY GUIDE Name: SCORE:
 Unit 7: Adaptation STUDY GUIDE Name: SCORE: 1. Which is an adaptation that makes it possible for the animal to survive in a cold climate? A. tail on a lizard B. scales on a fish C. stripes on a tiger D.
Unit 7: Adaptation STUDY GUIDE Name: SCORE: 1. Which is an adaptation that makes it possible for the animal to survive in a cold climate? A. tail on a lizard B. scales on a fish C. stripes on a tiger D.
rodent species in Australia to the fecal odor of various predators. Rattus fuscipes (bush
 Sample paper critique #2 The article by Hayes, Nahrung and Wilson 1 investigates the response of three rodent species in Australia to the fecal odor of various predators. Rattus fuscipes (bush rat), Uromys
Sample paper critique #2 The article by Hayes, Nahrung and Wilson 1 investigates the response of three rodent species in Australia to the fecal odor of various predators. Rattus fuscipes (bush rat), Uromys
AN APPLIED CASE STUDY of the complexity of ecological systems and process: Why has Lyme disease become an epidemic in the northeastern U.S.
 AN APPLIED CASE STUDY of the complexity of ecological systems and process: Why has Lyme disease become an epidemic in the northeastern U.S. over the last few decades? What causes Lyme disease? 1 Frequency
AN APPLIED CASE STUDY of the complexity of ecological systems and process: Why has Lyme disease become an epidemic in the northeastern U.S. over the last few decades? What causes Lyme disease? 1 Frequency
University of Canberra. This thesis is available in print format from the University of Canberra Library.
 University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
Animal Adaptations. EQ: How do animals adapt to survive?
 Animal Adaptations EQ: How do animals adapt to survive? What is adaptation? An adaptation is any special characteristic or skill that helps an animal to survive in its habitat. Examples: The shape of a
Animal Adaptations EQ: How do animals adapt to survive? What is adaptation? An adaptation is any special characteristic or skill that helps an animal to survive in its habitat. Examples: The shape of a
Animal Biodiversity. Teacher Resources - High School (Cycle 1) Biology Redpath Museum
 Biology Redpath Museum") Animal Biodiversity Teacher Resources - High School (Cycle 1) Biology Redpath Museum Ecology What defines a habitat? 1. Geographic Location The location of a habitat is determined by its latitude and its
Animal Biodiversity Teacher Resources - High School (Cycle 1) Biology Redpath Museum Ecology What defines a habitat? 1. Geographic Location The location of a habitat is determined by its latitude and its
Animal Defense against Predators. Ms. Levasseur Biology
 Animal Defense against Predators Ms. Levasseur Biology Animal Defense Against Predators Throughout millions of years of evolution, animals have evolved numerous ways of defending themselves against predators.
Animal Defense against Predators Ms. Levasseur Biology Animal Defense Against Predators Throughout millions of years of evolution, animals have evolved numerous ways of defending themselves against predators.
1 This question is about the evolution, genetics, behaviour and physiology of cats.
 1 This question is about the evolution, genetics, behaviour and physiology of cats. Fig. 1.1 (on the insert) shows a Scottish wildcat, Felis sylvestris. Modern domestic cats evolved from a wild ancestor
1 This question is about the evolution, genetics, behaviour and physiology of cats. Fig. 1.1 (on the insert) shows a Scottish wildcat, Felis sylvestris. Modern domestic cats evolved from a wild ancestor
North Carolina Aquariums Education Section. You Make the Crawl. Created by the NC Aquarium at Fort Fisher Education Section
 Essential Question: You Make the Crawl Created by the NC Aquarium at Fort Fisher Education Section How do scientists identify which sea turtle species has crawled up on a beach? Lesson Overview: Students
Essential Question: You Make the Crawl Created by the NC Aquarium at Fort Fisher Education Section How do scientists identify which sea turtle species has crawled up on a beach? Lesson Overview: Students
I LOVE MY DRAGONS! Dragons of Sydney Harbour Factsheet Kids Version
 I LOVE MY DRAGONS! Dragons of Sydney Harbour Factsheet Kids Version Did you know that we have dragons living right at our doorstep around the beautiful Sydney harbour? That s right! These beasts sure have
I LOVE MY DRAGONS! Dragons of Sydney Harbour Factsheet Kids Version Did you know that we have dragons living right at our doorstep around the beautiful Sydney harbour? That s right! These beasts sure have
DOES EXPERIMENTALLY INDUCED CONSPICUOUS COLORATION INCREASE RISK OF PREDATION AND CONSPECIFIC AGGRESSION IN FIRST-YEAR COLLARED LIZARD MALES?
 Herpetologica, 65(1), 2009, 31 38 E 2009 by The Herpetologists League, Inc. DOES EXPERIMENTALLY INDUCED CONSPICUOUS COLORATION INCREASE RISK OF PREDATION AND CONSPECIFIC AGGRESSION IN FIRST-YEAR COLLARED
Herpetologica, 65(1), 2009, 31 38 E 2009 by The Herpetologists League, Inc. DOES EXPERIMENTALLY INDUCED CONSPICUOUS COLORATION INCREASE RISK OF PREDATION AND CONSPECIFIC AGGRESSION IN FIRST-YEAR COLLARED
The large-scale environment and the rabbit's genetic diversity as factors to bear in mind in Iberian lynx Conservation
 PDF The large-scale environment and the rabbit's genetic diversity as factors to bear in mind in Iberian lynx Conservation A small-scale study using computer models stresses the need to, when it comes
PDF The large-scale environment and the rabbit's genetic diversity as factors to bear in mind in Iberian lynx Conservation A small-scale study using computer models stresses the need to, when it comes
Everyday Mysteries: Why most male birds are more colorful than females
 Everyday Mysteries: Why most male birds are more colorful than females By Scientific American, adapted by Newsela staff on 02.06.17 Word Count 779 Mandarin ducks, a male (left) and a female, at WWT Martin
Everyday Mysteries: Why most male birds are more colorful than females By Scientific American, adapted by Newsela staff on 02.06.17 Word Count 779 Mandarin ducks, a male (left) and a female, at WWT Martin
COMPARING BODY CONDITION ESTIMATES OF ZOO BROTHER S ISLAND TUATARA (SPHENODON GUNTHERI) TO THAT OF THE WILD, A CLINICAL CASE
 TO THAT OF THE WILD, A CLINICAL CASE") COMPARING BODY CONDITION ESTIMATES OF ZOO BROTHER S ISLAND TUATARA (SPHENODON GUNTHERI) TO THAT OF THE WILD, A CLINICAL CASE Kyle S. Thompson, BS,¹, ²* Michael L. Schlegel, PhD, PAS² ¹Oklahoma State University,
COMPARING BODY CONDITION ESTIMATES OF ZOO BROTHER S ISLAND TUATARA (SPHENODON GUNTHERI) TO THAT OF THE WILD, A CLINICAL CASE Kyle S. Thompson, BS,¹, ²* Michael L. Schlegel, PhD, PAS² ¹Oklahoma State University,
Maturity and Other Reproductive Traits of the Kanahebi Lizard Takydromus tachydromoides (Sauria, Lacertidae) in Mito
 in Mito") Japanese Journal of Herpetology 9 (2): 46-53. 1981. Maturity and Other Reproductive Traits of the Kanahebi Lizard Takydromus tachydromoides (Sauria, Lacertidae) in Mito Sen TAKENAKA SUMMARY: Reproduction
Japanese Journal of Herpetology 9 (2): 46-53. 1981. Maturity and Other Reproductive Traits of the Kanahebi Lizard Takydromus tachydromoides (Sauria, Lacertidae) in Mito Sen TAKENAKA SUMMARY: Reproduction
A different kind of ecological modelling: the use of clay model organisms to explore predator prey interactions in vertebrates
 Journal of Zoology. Print ISSN 0952-8369 REVIEW A different kind of ecological modelling: the use of clay model organisms to explore predator prey interactions in vertebrates P. W. Bateman 1, P. A. Fleming
Journal of Zoology. Print ISSN 0952-8369 REVIEW A different kind of ecological modelling: the use of clay model organisms to explore predator prey interactions in vertebrates P. W. Bateman 1, P. A. Fleming
HEALTH AND BODY CONDITION OF RABBIT DOES ON COMMERCIAL FARMS
 Pathology and Hygiene HEALTH AND BODY CONDITION OF RABBIT DOES ON COMMERCIAL FARMS Rosell J.M. 1 *, De La Fuente L.F. 2 1 Nanta SA Ronda de Poniente 9, 28760 Tres Cantos (Madrid), Spain 2 Departamento
Pathology and Hygiene HEALTH AND BODY CONDITION OF RABBIT DOES ON COMMERCIAL FARMS Rosell J.M. 1 *, De La Fuente L.F. 2 1 Nanta SA Ronda de Poniente 9, 28760 Tres Cantos (Madrid), Spain 2 Departamento
LAB. NATURAL SELECTION
 Period Date LAB. NATURAL SELECTION This game was invented by G. Ledyard Stebbins, a pioneer in the evolution of plants. The purpose of the game is to illustrate the basic principles and some of the general
Period Date LAB. NATURAL SELECTION This game was invented by G. Ledyard Stebbins, a pioneer in the evolution of plants. The purpose of the game is to illustrate the basic principles and some of the general
The Origin of Species: Lizards in an Evolutionary Tree
 The Origin of Species: Lizards in an Evolutionary Tree Cara Larracas, Stacy Lopez, Takara Yaegashi Period 4 Background Information Throughout the Caribbean Islands there is a species of anole lizards that
The Origin of Species: Lizards in an Evolutionary Tree Cara Larracas, Stacy Lopez, Takara Yaegashi Period 4 Background Information Throughout the Caribbean Islands there is a species of anole lizards that
ACTIVITY #6: TODAY S PICNIC SPECIALS ARE
 TOPIC What types of food does the turtle eat? ACTIVITY #6: TODAY S PICNIC SPECIALS ARE BACKGROUND INFORMATION For further information, refer to Turtles of Ontario Fact Sheets (pages 10-26) and Unit Five:
TOPIC What types of food does the turtle eat? ACTIVITY #6: TODAY S PICNIC SPECIALS ARE BACKGROUND INFORMATION For further information, refer to Turtles of Ontario Fact Sheets (pages 10-26) and Unit Five:
Comparing Life Cycles
 Image from Wikimedia Commons Pre-Visit Activity Grade Two Comparing Life Cycles Specific Learning Outcomes 2-1-01: Use appropriate vocabulary related to the investigations of growth and changes in animals.
Image from Wikimedia Commons Pre-Visit Activity Grade Two Comparing Life Cycles Specific Learning Outcomes 2-1-01: Use appropriate vocabulary related to the investigations of growth and changes in animals.
Intraspecific relationships extra questions and answers (Extension material for Level 3 Biology Study Guide, ISBN , page 153)
") i Intraspecific relationships extra questions and answers (Extension material for Level 3 Biology Study Guide, ISBN 978-1-927194-58-4, page 153) Activity 9: Intraspecific relationships extra questions
i Intraspecific relationships extra questions and answers (Extension material for Level 3 Biology Study Guide, ISBN 978-1-927194-58-4, page 153) Activity 9: Intraspecific relationships extra questions
Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve,
 at Sungei Buloh Wetland Reserve,") Author Title Institute Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve, Singapore Thesis (Ph.D.) National
Author Title Institute Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve, Singapore Thesis (Ph.D.) National
MATERNAL NEST-SITE CHOICE AND OFFSPRING FITNESS IN A TROPICAL SNAKE (TROPIDONOPHIS MAIRII, COLUBRIDAE)
") Ecology, 85(6), 2004, pp. 1627 1634 2004 by the Ecological Society of America MATERNAL NEST-SITE CHOICE AND OFFSPRING FITNESS IN A TROPICAL SNAKE (TROPIDONOPHIS MAIRII, COLUBRIDAE) G. P. BROWN AND R. SHINE
Ecology, 85(6), 2004, pp. 1627 1634 2004 by the Ecological Society of America MATERNAL NEST-SITE CHOICE AND OFFSPRING FITNESS IN A TROPICAL SNAKE (TROPIDONOPHIS MAIRII, COLUBRIDAE) G. P. BROWN AND R. SHINE
Adaptation. Survival of the Fittest
 Adaptation Survival of the Fittest It s all about traits Acquired Traits Happen After Birth Scars Pierced Ears Learning a Skill Changing Appearance It s all about traits Inherited Traits Programmed at
Adaptation Survival of the Fittest It s all about traits Acquired Traits Happen After Birth Scars Pierced Ears Learning a Skill Changing Appearance It s all about traits Inherited Traits Programmed at
Mimicry and Defense. Protective Strategies 3/24/2015. Professor Donald McFarlane. Camouflage ( Cryptic coloration ) Diverse Coloration
 Diverse Coloration") Professor Donald McFarlane Mimicry and Defense Protective Strategies Camouflage ( Cryptic coloration ) Diverse Coloration Diversion Structures Startle Structures 2 1 Camouflage ( Cryptic coloration ) Minimize
Professor Donald McFarlane Mimicry and Defense Protective Strategies Camouflage ( Cryptic coloration ) Diverse Coloration Diversion Structures Startle Structures 2 1 Camouflage ( Cryptic coloration ) Minimize
Trunk Contents. Crane Flight Feathers (3)
") Trunk Contents Learning occurs not only with the mind, but also with the eyes, the hands the whole child (or adult!). Items contained in the trunk are meant to be examined, handled, and shared with your
Trunk Contents Learning occurs not only with the mind, but also with the eyes, the hands the whole child (or adult!). Items contained in the trunk are meant to be examined, handled, and shared with your
Mimicry Scientific Background
 Mimicry Scientific Background The adult Monarch butterfly advertises the fact that it is bitter tasting and toxic to its predators by its bright orange, black, and white wing color pattern. This pattern
Mimicry Scientific Background The adult Monarch butterfly advertises the fact that it is bitter tasting and toxic to its predators by its bright orange, black, and white wing color pattern. This pattern
Typical Snakes Part # 1
 Advanced Snakes & Reptiles 1 Module # 4 Component # 5 Family Colubridae This is the most represented family in the course area and has the more commonly encountered species. All of these snakes only have
Advanced Snakes & Reptiles 1 Module # 4 Component # 5 Family Colubridae This is the most represented family in the course area and has the more commonly encountered species. All of these snakes only have
Effects of Diet, Migration, and Breeding on Clay Lick Use by Parrots in Southeastern Peru.
 Effects of Diet, Migration, and Breeding on Clay Lick Use by Parrots in Southeastern Peru. Donald Brightsmith, Ph.D. Duke University, Department of Biology, Durham NC, USA Rainforest Expeditions, Peru
Effects of Diet, Migration, and Breeding on Clay Lick Use by Parrots in Southeastern Peru. Donald Brightsmith, Ph.D. Duke University, Department of Biology, Durham NC, USA Rainforest Expeditions, Peru
PSY 2364 Animal Communication. Elk (Cervus canadensis) Extra credit assignment. Sad Underwing (Catocala maestosa) 10/11/2017
 Extra credit assignment. Sad Underwing (Catocala maestosa) 10/11/2017") PSY 2364 Animal Communication Elk (Cervus canadensis) Kingdom: Phylum: Class: Order: Family: Genus: Species: Animalia Chordata Mammalia Artiodactyla Cervidae Cervus canadensis Extra credit assignment Sad
PSY 2364 Animal Communication Elk (Cervus canadensis) Kingdom: Phylum: Class: Order: Family: Genus: Species: Animalia Chordata Mammalia Artiodactyla Cervidae Cervus canadensis Extra credit assignment Sad
Parthenogenesis in Varanus ornatus, the Ornate Nile Monitor.
 Parthenogenesis in Varanus ornatus, the Ornate Nile Monitor. Parthenogenesis in varanids has been reported in two other species of monitor, the Komodo dragon, Varanus komodiensis (Watts et al) and the
Parthenogenesis in Varanus ornatus, the Ornate Nile Monitor. Parthenogenesis in varanids has been reported in two other species of monitor, the Komodo dragon, Varanus komodiensis (Watts et al) and the
Evolution. Evolution is change in organisms over time. Evolution does not have a goal; it is often shaped by natural selection (see below).
.") Evolution Evolution is change in organisms over time. Evolution does not have a goal; it is often shaped by natural selection (see below). Species an interbreeding population of organisms that can produce
Evolution Evolution is change in organisms over time. Evolution does not have a goal; it is often shaped by natural selection (see below). Species an interbreeding population of organisms that can produce
Cnemidophorus lemniscatus (Rainbow Whiptail)
") Cnemidophorus lemniscatus (Rainbow Whiptail) Family: Teiidae (Tegus and Whiptails) Order: Squamata (Lizards and Snakes) Class: Reptilia (Reptiles) Fig. 1. Rainbow whiptail, Cnemidophorus lemniscatus. [https://www.flickr.com/photos/vhobus/6717385289/,
Cnemidophorus lemniscatus (Rainbow Whiptail) Family: Teiidae (Tegus and Whiptails) Order: Squamata (Lizards and Snakes) Class: Reptilia (Reptiles) Fig. 1. Rainbow whiptail, Cnemidophorus lemniscatus. [https://www.flickr.com/photos/vhobus/6717385289/,
d. Wrist bones. Pacific salmon life cycle. Atlantic salmon (different genus) can spawn more than once.
 can spawn more than once.") Lecture III.5b Answers to HW 1. (2 pts). Tiktaalik bridges the gap between fish and tetrapods by virtue of possessing which of the following? a. Humerus. b. Radius. c. Ulna. d. Wrist bones. 2. (2 pts)
Lecture III.5b Answers to HW 1. (2 pts). Tiktaalik bridges the gap between fish and tetrapods by virtue of possessing which of the following? a. Humerus. b. Radius. c. Ulna. d. Wrist bones. 2. (2 pts)
Species must be adapted to their habitat.
 Species must be adapted to their habitat. Species must protect themselves from the heat, cold, move around, feed themselves, communicate and reproduce. These are the types of adaptations which we will
Species must be adapted to their habitat. Species must protect themselves from the heat, cold, move around, feed themselves, communicate and reproduce. These are the types of adaptations which we will
ARTIFICIAL EGG-LAYING SITES FOR LIZARDS: A CONSERVATION STRATEGY
 0006-3207(94)00060-3 Biological Conservation 12 (1995) 387-391 Elsevier Science Limited Printed in Great Britain 0006-3207/95/$09.50+.00 ARTIFICIAL EGG-LAYING SITES FOR LIZARDS: A CONSERVATION STRATEGY
0006-3207(94)00060-3 Biological Conservation 12 (1995) 387-391 Elsevier Science Limited Printed in Great Britain 0006-3207/95/$09.50+.00 ARTIFICIAL EGG-LAYING SITES FOR LIZARDS: A CONSERVATION STRATEGY
Reptile Identification Guide
 Care & preservation of Surrey s native amphibians and reptiles Reptile Identification Guide This identification guide is intended to act as an aid for SARG surveyors. Adder, Vipera berus A short, stocky
Care & preservation of Surrey s native amphibians and reptiles Reptile Identification Guide This identification guide is intended to act as an aid for SARG surveyors. Adder, Vipera berus A short, stocky
A.13 BLAINVILLE S HORNED LIZARD (PHRYNOSOMA BLAINVILLII)
") A. BLAINVILLE S HORNED LIZARD (PHRYNOSOMA BLAINVILLII) A.. Legal and Other Status Blainville s horned lizard is designated as a Department of Fish and Game (DFG) Species of Concern. A.. Species Distribution
A. BLAINVILLE S HORNED LIZARD (PHRYNOSOMA BLAINVILLII) A.. Legal and Other Status Blainville s horned lizard is designated as a Department of Fish and Game (DFG) Species of Concern. A.. Species Distribution
CLADISTICS Student Packet SUMMARY Phylogeny Phylogenetic trees/cladograms
 CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
EFFECTS OF FOOD SUPPLEMENTATION AND HABITAT SELECTION ON TIMING OF LESSER KESTREL BREEDING
 Notes Ecology, 83(3), 2002, pp. 873 877 2002 by the Ecological Society of America EFFECTS OF FOOD SUPPLEMENTATION AND HABITAT SELECTION ON TIMING OF LESSER KESTREL BREEDING JOSÉ MIGUEL APARICIO 1 AND RAÚL
Notes Ecology, 83(3), 2002, pp. 873 877 2002 by the Ecological Society of America EFFECTS OF FOOD SUPPLEMENTATION AND HABITAT SELECTION ON TIMING OF LESSER KESTREL BREEDING JOSÉ MIGUEL APARICIO 1 AND RAÚL
Egg laying site preferences in Pterostichus melanarius Illiger (Coleoptera: Carabidae)
") Egg laying site preferences in Pterostichus melanarius Illiger (Coleoptera: Carabidae) H. Tréfás & J.C. van Lenteren Laboratory of Entomology, Wageningen University and Research Centre, Binnenhaven 7,
Egg laying site preferences in Pterostichus melanarius Illiger (Coleoptera: Carabidae) H. Tréfás & J.C. van Lenteren Laboratory of Entomology, Wageningen University and Research Centre, Binnenhaven 7,
Do the traits of organisms provide evidence for evolution?
 PhyloStrat Tutorial Do the traits of organisms provide evidence for evolution? Consider two hypotheses about where Earth s organisms came from. The first hypothesis is from John Ray, an influential British
PhyloStrat Tutorial Do the traits of organisms provide evidence for evolution? Consider two hypotheses about where Earth s organisms came from. The first hypothesis is from John Ray, an influential British
AnOn. Behav., 1971, 19,
 AnOn. Behav., 1971, 19, 575-582 SHIFTS OF 'ATTENTION' IN CHICKS DURING FEEDING BY MARIAN DAWKINS Department of Zoology, University of Oxford Abstract. Feeding in 'runs' of and grains suggested the possibility
AnOn. Behav., 1971, 19, 575-582 SHIFTS OF 'ATTENTION' IN CHICKS DURING FEEDING BY MARIAN DAWKINS Department of Zoology, University of Oxford Abstract. Feeding in 'runs' of and grains suggested the possibility