Title. CitationChromosome Research, 15(6): Issue Date Doc URL. Rights. Type. File Information.
|
|
|
- Russell Jordan
- 6 years ago
- Views:
Transcription
1 Title The molecular basis of chromosome orthologies and se Nishida-Umehara, Chizuko; Tsuda, Yayoi; Ishijima, Ju Author(s) Darren K. CitationChromosome Research, 15(6): Issue Date Doc URL Rights The original publication is available at Type article (author version) File Information CR15-6.pdf Instructions for use Hokkaido University Collection of Scholarly and Aca
2 The molecular basis of chromosome orthologies and sex chromosomal differentiation in palaeognathous birds Chizuko Nishida-Umehara 1, 2, Yayoi Tsuda 2, Junko Ishijima 1, Junko Ando 1, Atushi Fujiwara 3, Yoichi Matsuda 1, 2 * & Darren K. Griffin 4 1 Laboratory of Animal Cytogenetics, Division of Genome Dynamics, Creative Research Initiative Sousei, Hokkaido University, North 10 West 8, Kita-ku, Sapporo , Japan; Tel: ; Fax: ; yoimatsu@ees.hokusai.ac.jp, 2 Laboratory of Cytogenetics, Division of Bioscience, Graduate School of Environmental Earth Science, Hokkaido University, Sapporo , Japan 3 Immunology Section, National Research Institute of Aquaculture, Fisheries Research Agency, Tamaki, Mie , Japan; 4 Department of Biosciences, University of Kent, Canterbury CT2 7NJ, UK. *Correspondence Key words: chromosome painting, gene mapping, sex chromosome, Struthioniformes, Tinamiformes 1
3 Abstract Palaeognathous birds (Struthioniformes and Tinamiformes) have morphologically conserved karyotypes and less differentiated ZW sex chromosomes. To delineate interspecific chromosome orthologies in palaeognathous birds we conducted comparative chromosome painting with chicken (Gallus gallus, GGA) chromosome 1-9 and Z chromosome paints (GGA1-9 and GGAZ) for emu, double-wattled cassowary, ostrich, greater rhea, lesser rhea and elegant crested tinamou. All six species showed the same painting patterns: each probe was hybridized to a single pair of chromosomes with the exception that the GGA4 was hybridized to the fourth largest chromosome and a single pair of microchromosomes. The GGAZ was also hybridized to the entire region of the W chromosome, indicating that extensive homology remains between the Z and W chromosomes on the molecular level. Comparative FISH mapping of four Z- and/or W-linked markers, the ACO1/IREBP, ZOV3 and CHD1 genes and the EE0.6 sequence, revealed the presence of a small deletion in the proximal region of the long arm of the W chromosome in greater rhea and lesser rhea. These results suggest that the karyotypes and sex chromosomes of palaeognathous birds are highly conserved not only morphologically, but also at the molecular level; moreover, palaeognathous birds appear to retain the ancestral lineage of avian karyotypes. 2
4 Introduction Avian karyotypes are generally characterized by the high diploid number of chromosomes, ranging from 74 to 86 in about two thirds of species. Karyotypes can be subdivided into large macrochromosomes including ZW-type sex chromosomes (7-10 pairs) and a large number of almost indistinguishable microchromosomes (Takagi & Sasaki 1974, Belterman & de Boer 1984) or into groups A-D (Masabanda et al. 2004). The karyotypic similarities and differences between bird species have been studied morphologically by conventional Giemsa staining and chromosome banding, and, for the past 10 years, molecular cytogenetically by comparative FISH mapping with chromosome-specific paints, cdna and genomic DNA clones, mostly developed in chicken (Gallus gallus) (Griffin et al. 1999, Suzuki et al. 1999, Schmid et al. 2000, Shibusawa et al. 2001, 2002). Cross-species chromosome hybridization (termed Zoo-FISH) and subsequent comparative gene mapping delineates accurately the chromosomal orthologies between distantly related species and the chromosome rearrangements that have occurred during evolution. Comparative chromosome painting in birds with chicken probes has been performed for over 30 species in at least nine orders (Shetty et al. 1999, Schmid et al. 2000, Raudsepp et al. 2002, Guttenbach et al. 2003, Kasai et al. 2003, Derjusheva et al. 2004, Shibusawa et al. 2004a, b, Itoh & Arnold 2005, de Oliveria et al. 2005, Nanda et al. 2006, reviewed in Griffin et al. 2007). These studies collectively have revealed that the avian karyotypes are highly conserved at the molecular level with rare exceptions including the Falconiformes and the Psittaciformes which display both the fragmentation of macrochromosomes and the 3
5 decrease in microchromosomal number through fusions (de Oliveira et al. 2005). Reciprocal chromosome translocations have not yet been reported. Avian species are categorized into two large clades based on the palatal form: the Palaeognathae and the Neognathae. The Palaeognathae consist of the Struthioniformes (Ratites) and the sister group Tinamiformes. This classification is also confirmed molecular phylogenetically (Sibley & Ahlquist 1990, van Tuinen & Hedges 1998, 2000, Cracraft 2001). G-banding suggests that palaeognathous birds have morphologically the typical avian karyotype (Takagi et al. 1972, de Boer 1980, Ansari et al. 1988), which is also shared by most neognathous birds (Takagi & Sasaki 1974, Belterman & de Boer 1984); there is however a remarkable difference in the sex chromosome differentiation between the two taxa. The W chromosomes of neognathous birds are highly differentiated: they are smaller than the Z chromosomes, conspicuously heterochromatin-rich and late replicating (Takagi et al. 1972, Schmid et al. 1989). In contrast, the sex chromosomes of Struthioniformes species remain the most primitive stage of avian sex chromosome differentiation: the Z and W chromosomes are extensively homomorphic and euchromatic (Takagi et al. 1972, de Boer 1980, Ansari et al. 1988). Chromosome numbers of Tinamiformes species have been reported to be around 2n=80, and they also appear to have the typical avian karyotypes (Sasaki et al. 1984, Belterman & de Boer 1990, Pigozzi & Solari 1999, 2005), whereas the W chromosomes with the large heterochromatin blocks are in the intermediate state between the euchromatic W chromosomes of the Ratites and the highly heterochomatized W 4
6 chromosomes of neognathous birds (Sasaki et al. 1980, Pigozzi & Solari 1999, 2005, Tsuda et al. 2007). In palaeognathous birds, cross-species chromosome hybridization has been performed only for emu (Dromaius novaehollandiae) (Shetty et al. 1999) and for greater rhea (Rhea americana) (Guttenbach et al. 2003), with chicken chromosome 1-9 paints demonstrating that their karyotypes have complete orthology with the chicken macrochromosomes except that the chicken chromosome 4 paint hybridizes to the fourth largest chromosome and an additional pair of microchromosomes. Chromosome painting with the chicken Z probe showed that the homology between the Z and W chromosomes is highly conserved at the molecular level in emu, however, the sex chromosomes of other Ratites and Tinamous have only been morphologically studied by conventional Giemsa staining and/or G- and C-banding (Takagi et al. 1972, Sasaki et al. 1980, Ansari et al. 1988, Belterman & de Boer 1990, Pigozzi & Solari, 1999, 2005). Comparative mapping with a few Z- and/or W-linked molecular makers has been also performed only for emu, ostrich and double-wattled cassowary (Ogawa et al. 1998, Nishida-Umehara et al. 1999). Here we have conducted comparative chromosome painting with chicken chromosome-specific DNA probes for six species belonging to two Palaeognathae orders, and delineated chromosome homologies and interchromosomal rearrangements among the palaeognathous bird species. The state of sex chromosome differentiation was also examined by comparative chromosome mapping of Z- and/or W-linked markers and morphological comparison of banding patterns between the Z and W chromosomes. Karyotypic evolution in the context of sex chromosome differentiation in 5
7 palaeognathous birds is discussed. Materials and methods Specimens Five species of the Struthioniformes and one species of the Tinamiformes were used for chromosomal analysis: emu (Dromaius novaehollandiae), double-wattled cassowary (Casuarius casuarius), ostrich (Struthio camelus), greater rhea (Rhea americana) and lesser rhea (Pterocnemia pennata) of the Struthioniformes and elegant crested tinamou (Eudromia elegans) of the Tinamiformes. Small pieces of skin tissues were taken by biopsy and used for cell culture. Cell culture and chromosome preparation The fibroblast cells prepared from the skin tissues were cultured in 199 medium (Invitrogen-GIBCO) supplemented with 18 % fetal bovine serum at 39 C in 5% CO 2 in air. After colcemid (0.025 μg/ml) treatment for 30 min, the chromosome preparation was made following a standard protocol. For karyotyping the chromosome slides were stained with 3% Giemsa solution for 10 min. Chromosome banding To examine the distribution of constitutive heterochromatin on the Z and W chromosomes, C-banding was carried out with the BSG (Barium 6
8 hydroxide/saline/giemsa) method (Sumner 1972). For morphological comparison of the Z and W chromosomes, replication G-banded chromosome preparations were made using the GBG (G-bands by BrdU using Giemsa) method as described in Nishida-Umehara et al. (1999). The replication R-banded chromosomes were prepared following Suzuki et al. (1999), and used for chromosome painting, Ag-NOR staining and chromosome mapping of DNA clones. DNA probes Chicken (Gallus gallus, GGA) chromosome-specific DNA probes of chromosome 1 9 and Z (GGA1-9 and GGAZ) were used for comparative chromosome painting (Griffin et al. 1999, Masabanda et al. 2004, Each probe was amplified by DOP-PCR following Carter et al. (1992) and hybridized to the metaphase spreads. The cosmid DNA clones of the ACO1/IREBP and ZOV3 genes, the EE0.6 sequence derived from emu (Ogawa et al. 1998) and the genomic DNA fragment of the CHD1 gene cloned from lesser rhea in this study were used as probes for comparative mapping of the Z and W chromosomes. A 4.5 kb DNA fragment of the CHD1 gene was amplified by polymerase chain reaction (PCR) with the genomic DNA of lesser rhea using the forward primer 2550(F): 5 -GTTACTGATTCGTCTACGAGA-3 (Fridolfsson & Ellegren 1999) and the reverse primer P2(R): 5 -TCTGCATCGCTAAATCCTTT-3 (Griffiths et al. 1996). For chromosome mapping of the 18S-28S ribosomal RNA genes, the 5.8-kb phr21ab and 7.3-kb phr14e3 fragments of the human ribosomal RNA gene provided by the Japanese Cancer Research Resource Bank (JCRB), Tokyo, were used. 7
9 Fluorescence in-situ hybridization (FISH) and Ag-NOR-staining FISH was performed as described in Matsuda & Chapman (1995) with slight modifications. For comparative chromosome painting, 1 microgram of chicken chromosome-specific DNA probe was labeled with biotin 16-dUTP using a nick translation kit (Roche Diagnostics), and hybridized to chromosome slides at 37 C for four days. After hybridization, the slides were incubated with fluorosceinated avidin (FITC-avidin) (Roche Diagnostics), and stained with 0.50 μg/ml propidium iodide after washing. The FISH images were captured using a cooled CCD camera (MicroMAX 782Y, Princeton Instruments) mounted on a Leica DMRA microscope, and analysed with the 550CW-QFISH application program of Leica Microsystems Imaging Solutions Ltd. (Cambridge, UK). For chromosome mapping of the ACO1/IREBP, ZOV3 and CHD1 genes, the EE0.6 sequence and the 18S-28S ribosomal RNA (rrna) genes, 0.5 μg probe was labeled by nick translation with biotin 16-dUTP or digoxigenin (DIG) 11-dUTP (Roche Diagnostics). The genomic DNA clones of ACO1/IREBP, ZOV3 and EE0.6 were hybridized to chromosomes of greater rhea, lesser rhea and elegant crested tinamou, and the DNA fragments of CHD1 and the rrna genes were hybridized to chromosomes of all six species. After hybridization overnight, the slides were washed, and then the hybridized probes labeled with biotin 16-dUTP and DIG 11-dUTP were detected with FITC-avidin and anti-dig-rhodamine (Roche Diagnostics), respectively. The FISH signals were observed under a Nikon E800 fluorescence microscope using a B-2A filter 8
10 and microphotographed using Kodak Ektachrome ASA100 films. The chromosomal distribution of the nucleolar organizer regions (NORs) was examined on the same metaphase spreads used for FISH analysis. After FISH, the slides were washed with distilled water, fixed with 3:1 methanol/glacial acetic acid for 5 min, rinsed with methanol and air-dried. Ag-NOR staining was performed as described in Howell & Black (1980). Results Karyotypes The diploid chromosome numbers of the six palaeognathous bird species were as follows: emu (2n=80), double-wattled cassowary (2n=92), ostrich (2n=80), greater rhea (2n=80), lesser rhea (2n=80) and elegant crested tinamou (2n=80), being identical with those reported by previous authors (Takagi et al. 1972, Ansari et al. 1988, Sasaki et al. 1980, 1984, Nishida-Umehara et al. 1999). Giemsa-stained preparations of the nine largest autosomes and ZW sex chromosomes of the six species are shown in Figure 1. The third largest chromosomes were acrocentric in ostrich, greater rhea, lesser rhea and elegant crested tinamou but subtelocentric in emu and double-wattled cassowary. The fourth largest chromosomes were all acrocentric except for the submetacentric chromosome 4 of elegant crested tinamou. The fifth largest autosomes were acrocentric except for the submetacentric chromosome 5 of greater rhea and lesser rhea. The Z and W chromosomes were acrocentric in emu, double-wattled cassowary, 9
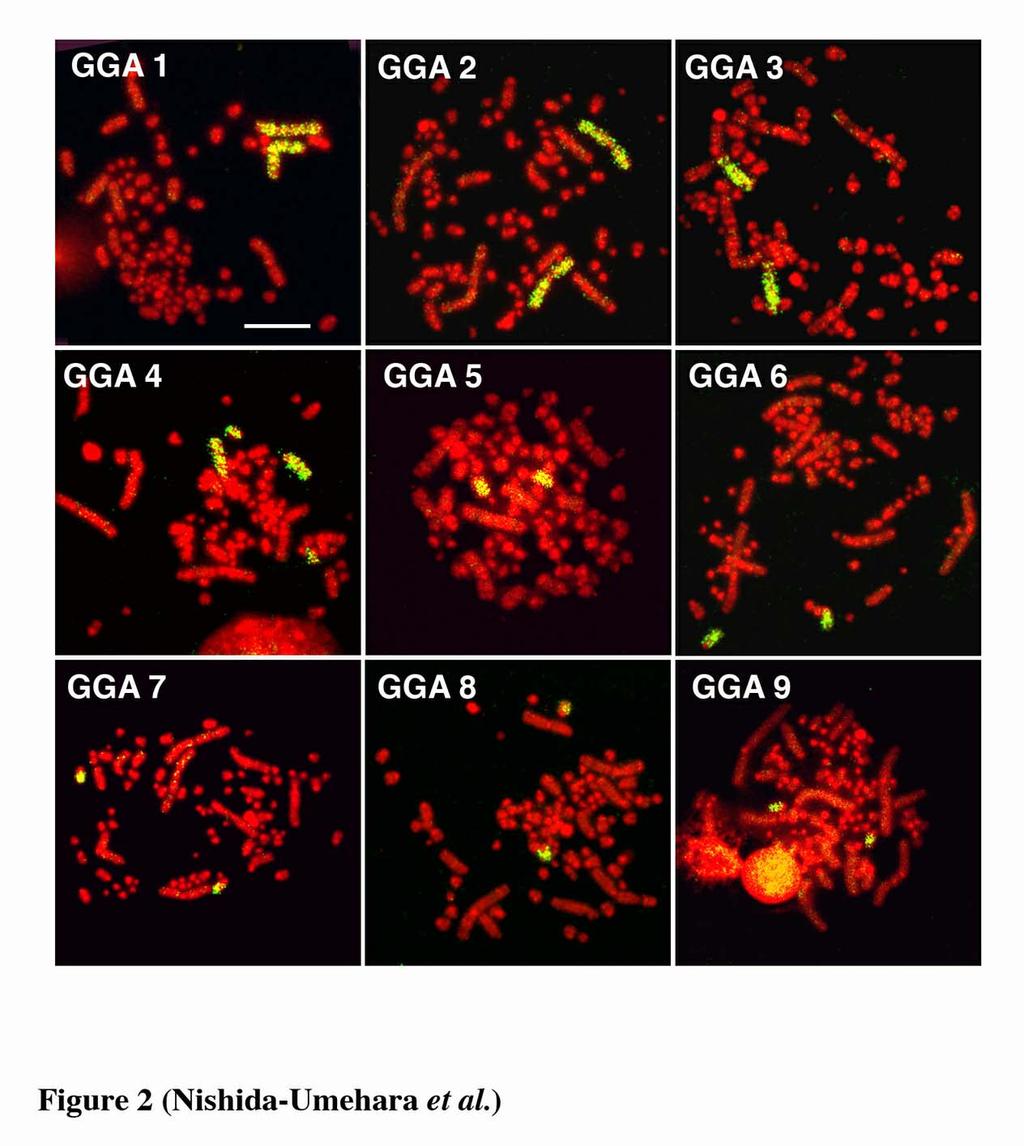
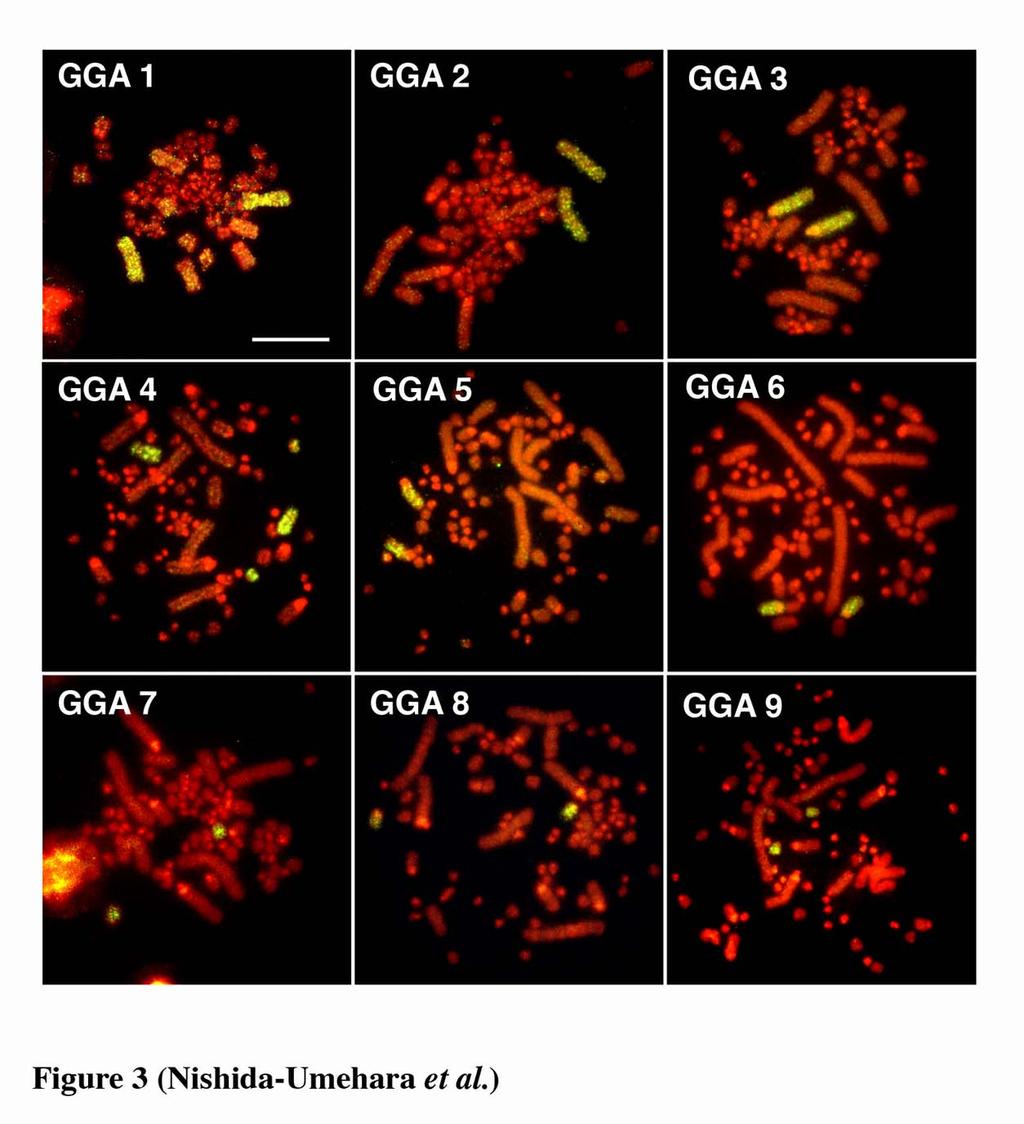
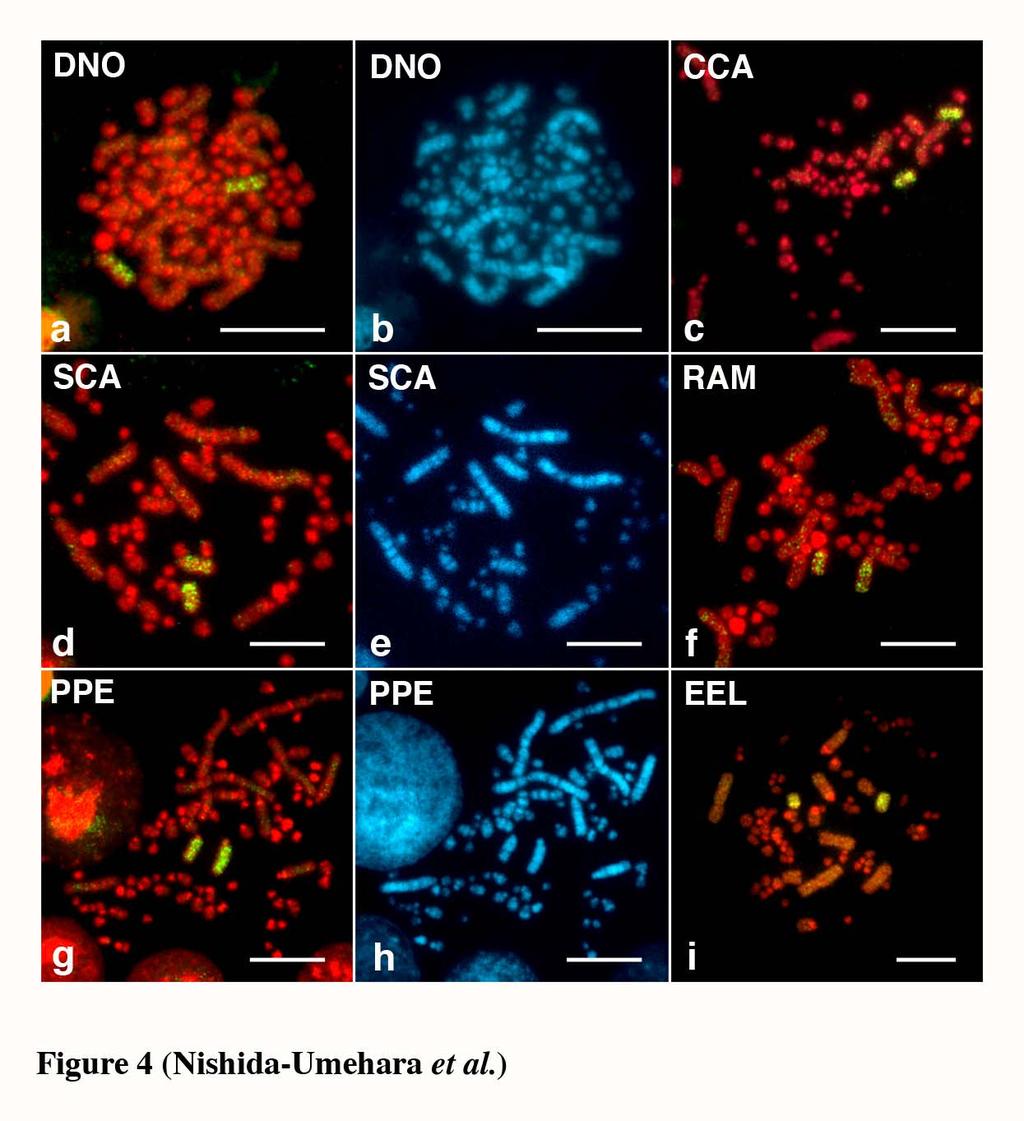
11 greater rhea, lesser rhea and elegant crested tinamou, whereas the Z and W chromosomes of ostrich were acrocentric and subtelocentric, respectively. The size differences between the Z and W chromosomes were small in the five species except elegant crested tinamou (Takagi et al. 1972, Sasaki et al. 1980, Ansari et al. 1988, Nishida-Umehara et al. 1999). Interspecific chromosomal homologies of macrochromosomes Chromosomal homologies were examined at the molecular level among the six palaeognathous bird species by comparative painting with chicken chromosome-specific DNA probes. All chicken painting probes (GGA1-9 and GGAZ) were efficiently cross-hybridized to chromosomes of all six species. The chromosome painting patterns with GGA1-9 in double-wattled cassowary and elegant crested tinamou were shown in Figures 2 and 3, respectively. Each chicken probe painted a single pair of chromosomes with the exception for GGA4 in all six species (Table 1), and chicken chromosome 1, 2, 3 and 5 corresponded to each chromosome 1, 2, 3 and 5, respectively. The GGA4 was hybridized to the fourth largest macrochromosome and additionally to a single pair of microchromosomes. The GGAZ was cross-hybridized to both the Z and W chromosomes in all six species, and the W chromosomes were also entirely painted like the Z chromosomes (Figure 4). The painting patterns of emu chromosomes with GGA1-9 and GGAZ and greater rhea chromosomes with GGA1-9 were consistent with the published data (Shetty et al. 1999, Guttenbach et al. 2003). 10
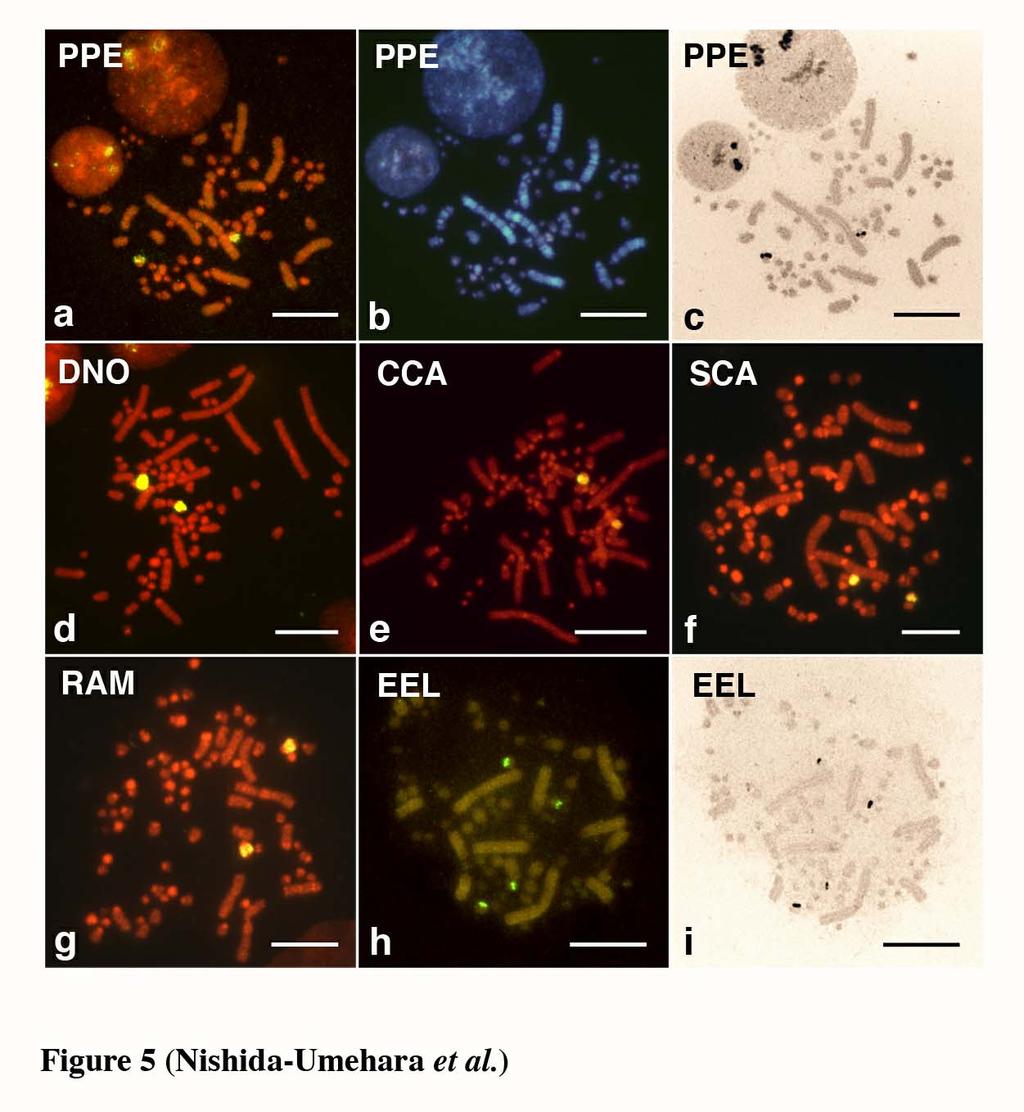
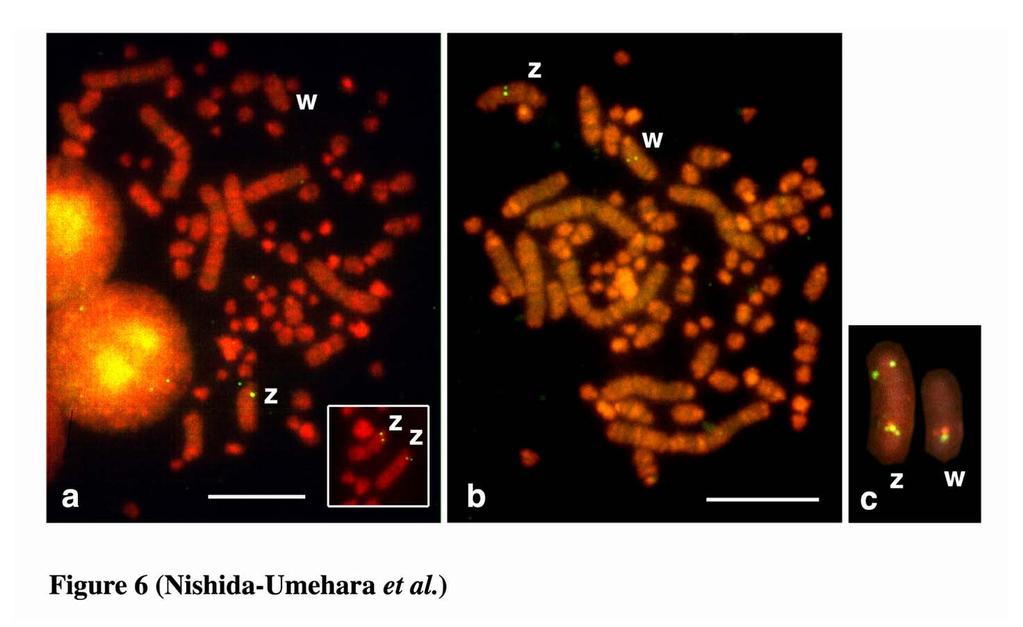
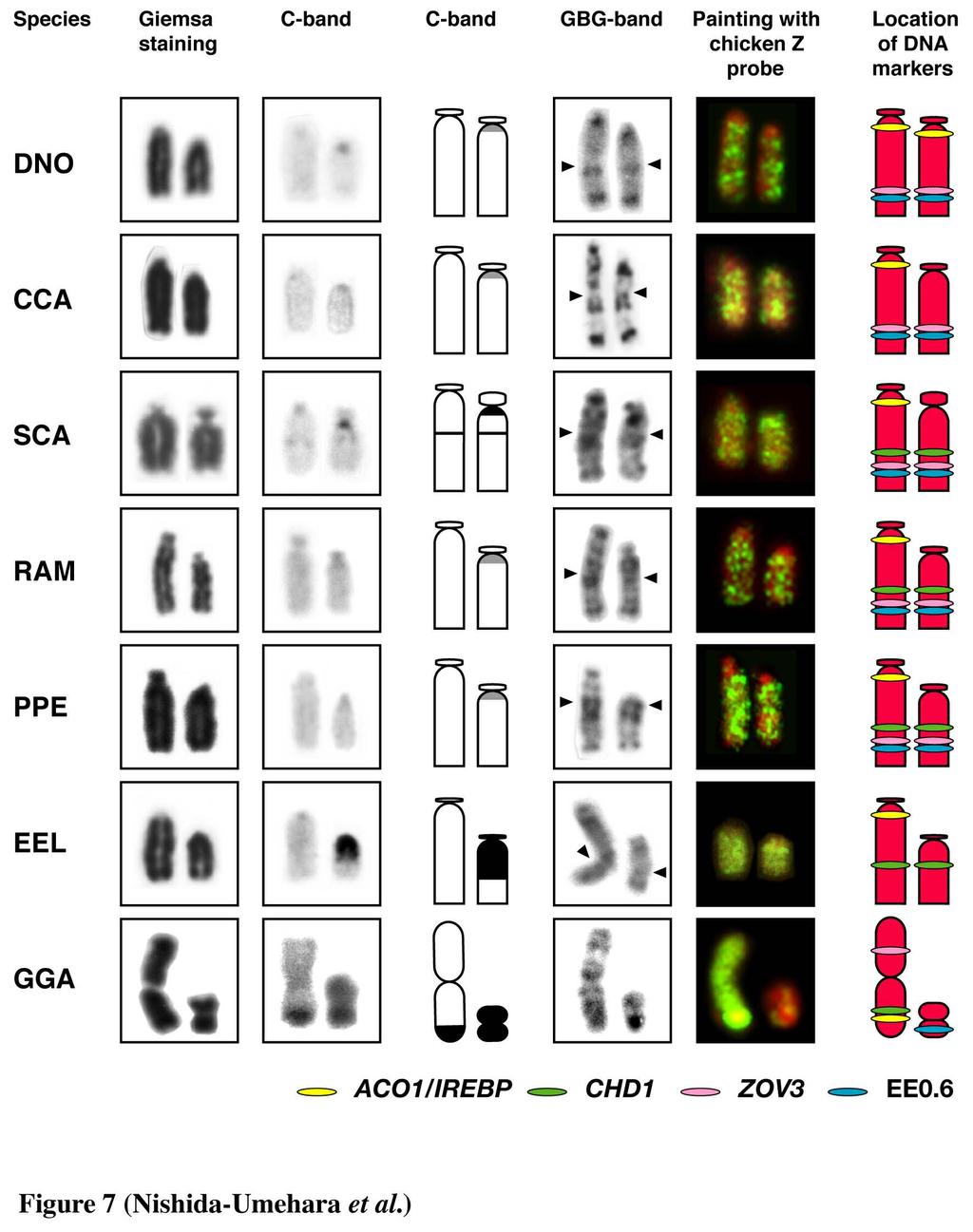
12 Chromosomal location of the 18S-28S rrna genes The 18S-28S rrna genes were localized to a single pair of indistinguishable microchromosomes in emu, double-wattled cassowary, ostrich, greater rhea and lesser rhea, and to two pairs of microchromosomes in elegant crested tinamou (Figure 5). Ag-NORs were all localized to the chromosomal sites where the hybridization signals of the 18S-28S rrna genes were located. Comparative mapping of Z- and W-linked DNA markers Comparative mapping of the ACO1/IREBP and ZOV3 genes and the EE0.6 sequence was performed for greater rhea, lesser rhea and elegant crested tinamou. The hybridization signals of the clones were all detected for lesser rhea (Figure 6) and greater rhea (data not shown) but not for elegant crested tinamou. The 4.5 kb genomic DNA fragment of the CHD1 gene isolated from lesser rhea was applied to chromosomes of all six species, but the hybridization signals were obtained only for two rhea species. The hybridization signal of ACO1/IREBP was detected near the centromere on the Z chromosome but not on the W chromosome in two rhea species (Figure 6a). The CHD1, ZOV3 and EE0.6 were localized to both the Z and W chromosomes in the two species (Figure 6b, c). ZOV3 was closely localized proximal to EE0.6, and the CHD1 was localized proximal to ZOV3 on the Z chromosomes. The locations of the genes and the order of CHD1 - ZOV3 - EE0.6 from the proximal on the Z chromosome was the same as those on the W chromosome. The locations of the four markers on the Z and W chromosomes in the two species were the same as those in ostrich reported previously 11
13 (Ogawa et al. 1998, Tsuda et al. 2007) (Figure 7). C-banded and replication G-banded patterns of the Z and W chromosomes The structural differences between the Z and W chromosomes were examined by C-banding and replication G-banding. The results are summarized in Figure 7 with the data of chromosome painting with the chicken Z probe and chromosome mapping of Z- and/or W-linked DNA markers. In ostrich, distinct C-positive bands were observed in the interstitial region of both the Z and W chromosomes and in the centromeric region of the W chromosome. In emu, double-wattled cassowary, greater rhea and lesser rhea, weak C-positive bands were observed in the centromeric regions of the W chromosomes. The GBG method produced high resolution G-bands, which were effective for morphological comparison between the Z and W chromosomes. The replication G-banded patterns in the distal half (approximately) of the Z chromosome (indicated by arrows in Figure 7) were identical between the Z and W chromosomes in five Struthioniformes species, whereas there were differences in banding patterns between the proximal half of the Z chromosomes and the corresponding regions of the W chromosomes. In elegant crested tinamou, the replication G-banded pattern was similar between the Z and W chromosomes in the distal quarter (roughly) of the Z chromosome. Two thirds of the W chromosome were composed of C-positive heterochromatin in this species, whereas no C-positive band was observed on the Z chromosome. These results strongly suggest that structural changes have occurred in the proximal regions of the W chromosomes of all six palaeognathous bird species. 12
14 Discussion The morphological comparison of macrochromosomes and ZW sex chromosomes among five Struthioniformes species and one Tinamiformes species confirmed that the karyotypes have been highly conserved among palaeognathous birds, although there are a few morphological differences between the macrochromosomes (Takagi et al. 1972, Sasaki et al. 1980, 1984, Ansari et al. 1988, Nishida-Umehara et al. 1999, Guttenbach et al. 2003, present study). The karyotypes of palaeognathous birds, which are composed of several macrochromosomes and a large number of almost indistinguishable microchromosomes, are quite similar to those of the earliest diverged Carinates, the Galliformes and the Anseriformes (Takagi & Sasaki 1974, Belterman & de Boer 1984). Cross-species chromosome hybridization with chicken chromosome paints confirmed the karyotypic orthologies between the Palaeognathae and the Galloanserae at the molecular level. The painting probes of chicken chromosome 1-9 and Z were each hybridized to a single pair of chromosomes in all six palaeognathous bird species except that the GGA4 was hybridized to the fourth largest chromosome and an additional pair of microchromosomes. This result suggests that the higher diploid chromosome number of double-wattled cassowary (2n=92) than five other palaeognathous bird species (2n=80) was attributed to the fissions of chromosomes smaller than number 9. Chromosome painting with the GGA4 probe confirmed that the submetacentric chicken chromosome 4 resulted from a centric fusion between the ancestral type of acrocentric 13
15 chromosome 4 (GGA4q) and a smaller (GGA4p) (Shetty et al. 1999, Shibusawa et al. 2004b). The short arm of GGA4 is GC-rich, preserving the feature of GC-rich bird microchromosome (McQueen et al. 1996, 1998, International Chicken Genome Sequencing Consortium 2004); Griffin and colleagues have suggested that this smaller chromosome is orthologous to turkey chromosome 9 which has been since designated ancestral chromosome 10 (International Chicken Genome Sequencing Consortium 2004; Griffin et al. 2007). We previously delineated the process of karyotypic evolution of the Galliformes by comparing the chromosome painting data of 13 Galliformes species with their molecular phylogenetic tree constructed with the mitochondrial DNA sequences, and consequently proposed that the karyotype of emu is identical with the ancestral karyotype of the Galliformes (at least for the largest chromosome pairs) (Shibusawa et al. 2004b). The 18S-28S rrna genes were localized to a pair of undistinguishable microchromosomes in all the five Struthioniformes species like chicken with the rrna genes located on chromosome 16, suggesting that the rrna genes might have been located on a single pair of microchromosomes in the ancestral avian karyotype. These data support that palaeognathous birds retain the ancestral karyotypes of the Galloanserae (Sibley & Ahlquist 1990), and that the ancient types of avian karyotypes have been independently conserved in both the lineages of the Palaeognathae and the Neognathae (including the Galloanserae and Neoaves) since they diverged about 120 million years ago (van Tuinen & Hedges 2001, Paton et al. 2002). We recently constructed a comparative cytogenetic map between chicken and Chinese soft-shelled turtle (Pelodiscus sinensis, Trionychidae) with a large number of 14
16 cdna clones (Matsuda et al. 2005). The chicken-turtle comparative map revealed that the GGA4q and GGA4p correspond to the subtelocentric chromosome 4 and a pair of microchromosomes of the turtle, respectively. The acrocentric chromosome 4 (GGA4q) and the GGA4p microchromosome are an ancient feature of the reptile-avian lineage; indeed GGA4q appears almost intact in mammals, being represented as human chromosome 4 (Chowdhary & Raudsepp 2000). Homologies between the turtle and chicken chromosomes have been highly conserved, with the six largest chromosomes being almost equivalent to one another. These results suggest that the Archosauromprha (reptiles and birds) have shared the similar ancestral types of chromosomal organization for more than 210 million years (Janke & Arnason 1997, Kumar & Hedges 1998, Hedges & Poling 1999, Kumazawa & Nishida 1999, Mannen & Li 1999). The GGAZ probe was intensely cross-hybridized to the entire W chromosomes of five Struthioniformes species, indicating that their W chromosomes have been hardly differentiated molecularly and still retain most of their genes. Cytogenetic studies on meiotic chromosome pairing also showed the presence of high homologies between the Z and W chromosomes in two rhea species (Pigozzi & Solari 1997, 1999). The comparative cytogenetic mapping of four Z- and/or W-linked molecular markers showed that the proximal regions including the ACO1/IREBP gene of the acrocentric Z chromosomes have been deleted in the W chromosomes of greater rhea and lesser rhea as previously reported in ostrich and double-wattled cassowary; however, ACO1/IREBP is still located on the emu W chromosome (Ogawa et al. 1998, Nishida-Umehara et al. 1999). In addition to the chromosome deletion, GBG-banding demonstrated that some 15
17 chromosomal rearrangements have occurred in the W chromosomal regions corresponding to the proximal half of the Z chromosomes. It seems more likely that the structural differentiation in the proximal region of the W chromosome was caused by the cessation of meiotic recombination between the Z and W chromosomes (Nishida-Umehara et al. 1999, García-Moreno & Mindell 2000, de Kloet & de Kloet 2003, Tsuda et al. 2007). In contrast to the Struthioniformes, there are large heterochromatic regions on the W chromosomes of Tinamiformes species (Sasaki et al. 1980, Pigozzi & Solari 1999, 2005). The chromosomal deletion in the proximal region of the W chromosome was much larger in elegant crested tinamou than ostrich (Tsuda et al. 2007), and the C-banding and GBG-banding also showed the presence of the large deletion of the euchromatic regions in the W chromosome of this species. The sexual dimorphism has been found for the CHD1 and spindlin (SPIN) genes of elegant crested tinamou, which are located in the proximal half of the W chromosome (de Kloet 2002, de Kloet & de Kloet 2003, Tsuda et al. 2007), suggesting that the Z and W copies of CHD1 and SPIN have been diverged by the cessation of meiotic recombination in the region containing the two genes between the Z and W chromosomes. Recently we molecularly cloned the BamHI-family of repetitive DNA sequences that consist of the large W-heterochromatin of elegant crested tinamou (Tsuda et al. 2007); however, this heterochromatin region was also intensely painted with chicken Z probe. This result suggests that a single copy or a few number of copies of the unique sequences on the Z chromosome, which are shared by both the Palaeognathae and the Neognathae, were site-specifically amplified on the W chromosome in this species. The W chromosomes 16
18 of Tinamiformes species, therefore, are considered to be at a transitional stage between the largely euchromatic W chromosomes of the Struthioniformes and the highly condensed heterochromatic W chromosomes of neognathous birds. Acknowledgements We express our appreciation to Dr. Shigeki Mizuno, Nihon University, for providing the cosmid clone of ZOV3, ACO1/IREBP and EE0.6 derived from emu, and Dr. Yasuyuki Ishibashi, Forestry and Forest Products Research Institute, for helping molecular cloning of the CHD1 DNA fragment from lesser rhea. We express our appreciation to Kanazawa Zoological Gardens, Yokohama, for providing the specimen of double-wattled cassowary, greater rhea and lesser rhea, Yokohama Zoological Gardens, Yokohama, for emu and elegant crested tinamou, Ueno Zoological Gardens, Tokyo, for emu, and Dr. Kimiyuki Tsuchiya, Tokyo University of Agriculture, for ostrich. This work was supported by Grants-in-Aid for Scientific Research (No and No ) from the Ministry of Education, Culture, Sports, Science and Technology, Japan and by a BBSRC project grant awarded to DKG. References Ansari HA, Takagi N, Sasaki M (1988) Morphological differentiation of sex chromosomes in three species of ratite birds. Cytogenet Cell Genet 47:
19 Belterman RHR, de Boer LEM (1984) A karyological study of 55 species of birds, including karyotypes of 39 species new to cytology. Genetica 65: Belterman RHR, de Boer LEM (1990) A miscellaneous collection of bird karyotypes. Genetica 83: Carter NP, Ferguson-Smith MA, Perryman MT et al. (1992) Reverse chromosome painting: a method for the rapid analysis of aberrant chromosomes in clinical cytogenetics. J Med Genet 29: Chowdhary BP, Raudsepp T (2000) HSA4 and GGA4: remarkable conservation despite 300-Myr divergence. Genomics 64: Cracraft J (2001) Avian evolution, Gondwana biogeography and the Cretaceous-Tertiary mass extinction event. Proc R Soc Lond B 268: de Boer LEM (1980) Do the chromosomes of the kiwi provide evidence for a monophyletic origin of the ratites? Nature 287: de Kloet SR (2002) Molecular sex identification of tinamous with PCR using primers derived from the spindlin gene. Mol Ecol Notes 2: de Kloet RS, de Kloet SR (2003) Evolution of the spindlin gene in birds: independent cessation of the recombination of sex chromosomes at the spindlin locus in neognathous birds and tinamous, a palaeognathous avian family. Genetica 119: de Oliveira EHC, Habermann FA, Lacerda O, Sbalqueiro IJ, Wienberg J, Müller S (2005) Chromosome reshuffling in birds of prey: the karyotype of the world's largest eagle (Harpy eagle, Harpia harpyja) compared to that of the chicken 18
20 (Gallus gallus). Chromosoma 114: Derjusheva S, Kurganova A, Habermann F, Gaginskaya E (2004) High chromosome conservation detected by comparative chromosome painting in chicken, pigeon and passerine birds. Chromosome Res 12: Fridolfsson A-K, Ellegren H (1999) A simple and universal method for molecular sexing of non-ratite birds. J Avian Biol 30: García-Moreno J, Mindell DP (2000) Rooting a phylogeny with homologous genes on opposite sex chromosomes (gametologs): a case study using avian CHD. Mol Biol Evol 17: Griffin DK, Haberman F, Masabanda J et al. (1999) Micro- and macrochromosome paints generated by flow cytometry and microdissection: tools for mapping the chicken genome. Cytogenet Cell Genet 87: Griffin DK, Robertson LBW, Tempest HG, Skinner BM (2007) The evolution of the avian genome as revealed by comparative molecular cytogenetics. Cytogenet Genome Res (in press). Griffiths R, Daan S, Dijkstra C (1996) Sex identification in birds using two CHD genes. Proc R Soc Lond B 263: Guttenbach M, Nanda I, Feichtinger W, Masabanda JS, Griffin DK, Schmid M (2003) Comparative chromosome painting of chicken autosomal paints 1-9 in nine different bird species. Cytogenet Genome Res 103: Hedges SB, Poling LL (1999) A molecular phylogeny of reptiles. Science 283:
21 Howell WM, Black DA (1980) Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method. Experientia 36: International Chicken Genome Sequencing Consortium (2004) Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 432: Itoh Y, Arnold AP (2005) Chromosomal polymorphism and comparative painting analysis in the zebra finch. Chromosome Res 13: Janke A, Arnason U (1997) The complete mitochondrial genome of Alligator mississippiensis and the separation between recent Archosauria (birds and crocodiles). Mol Biol Evol 14: Kasai F, Garcia C, Arruga MV, Ferguson-Smith MA (2003) Chromosome homology between chicken (Gallus gallus domesticus) and the red-legged partridge (Alectoris rufa); evidence of the occurrence of a neocentromere during evolution. Cytogenet Genome Res 102: Kumar S, Hedges SB (1998) A molecular timescale for vertebrate evolution. Nature 392: Kumazawa Y, Nishida M (1999) Complete mitochondrial DNA sequences of the green turtle and blue-tailed mole skink: statistical evidence for Archosaurian affinity of turtles. Mol Biol Evol 16: Mannen H, Li SS-L (1999) Molecular evidence for a clade of turtles. Mol Phyl Evol 13:
22 Masabanda JS, Burt DW, O'Brien PC, Vignal A, Fillon V, Walsh PS, Cox H, Tempest HG, Smith J, Habermann F, Schmid M, Matsuda Y, Ferguson-Smith MA, Crooijmans RP, Groenen MA, Griffin DK (2004): Molecular cytogenetic definition of the chicken genome: the first complete avian karyotype. Genetics 166: Matsuda Y, Chapman VM (1995) Application of fluorescence in situ hybridization in genome analysis of the mouse. Electrophoresis 16: Matsuda Y, Nishida-Umehara C, Tarui H et al. (2005) Highly conserved linkage homology between birds and turtles: birds and turtle chromosomes are precise counterparts of each other. Chromosome Res 13: McQueen HA, Fantes J, Cross SH, Clark VH, Archibald AL, Bird AP (1996) CpG islands of chicken are concentrated on microchromosomes. Nature Genet 12: McQueen HA, Siriaco G, Bird AP (1998) Chicken microchromosomes are hyperacetylated, early replicating, and gene rich. Genome Res 8: Nanda I, Karl E, Volobouev V, Griffin DK, Schartl M, Schmid M (2006) Extensive gross genomic rearrangements between chicken and Old World vultures (Falconiformes: Accipitridae). Cytogenet Genome Res 112: Nishida-Umehara C, Fujiwara A, Ogawa A. Mizuno S, Abe S, Yoshida MC (1999) Differentiation of Z and W chromosomes revealed by replication banding and FISH mapping of sex-chromosome-linked DNA markers in the cassowary (Aves, Ratitae). Chromosome Res 7:
23 Ogawa A, Murata K, Mizuno S (1998) The location of Z- and W-linked marker genes and sequence on the homomorphic sex chromosomes of the ostrich and the emu. Proc Natl Acad Sci USA 95: Paton T, Haddrath O, Baker AJ (2002) Complete mitochondrial DNA genome sequences show that modern birds are not descended from transitional shorebirds. Proc R Soc Lond B 269: Pigozzi MI, Solari AJ (1997) Extreme axial equalization and wide distribution of recombination nodules in the primitive ZW pair of Rhea americana (Aves, Ratitae). Chromosome Res 5: Pigozzi MI, Solari AJ (1999) The ZW pairs of two paleognath birds from two orders show transitional stages of sex chromosome differentiation. Chromosome Res 7: Pigozzi MI, Sorali AJ (2005) Meiotic recombination in the ZW pair of a tinamid bird shows a differential pattern compared with neognaths. Genome 48: Raudsepp T, Houck ML, O'Brien PC, Ferguson-Smith MA, Ryder OA, Chowdhary BP (2002) Cytogenetic analysis of California condor (Gymnogyps californianus) chromosomes: comparison with chicken (Gallus gallus) macrochromosomes. Cytogenet Cell Genet 98: Sasaki M, Nishida C, Takagi N, Hori H (1980) Sex-chromosomes of the elegant crested tinamou, Eudromia elegans (Aves: Tinamiformes: Tinamidae). Chromosome Info Serv 29: Sasaki M, Takagi N, Nishida C (1984) Current profiles of avian cytogenetics, with notes 22
24 on chromosomal diagnosis of sex in birds. the nucleus 27: Schmid M, Nanda I, Guttenbach M et al. (2000) First report on chicken genes and chromosomes Cytogenet Cell Genet 90: Schmid M, Enderle E, Schindler D, Schempp W (1989) Chromosome banding and DNA replication patterns in bird karyotypes. Cytogenet Cell Genet 52: Shetty S, Griffin DK, Graves JAM (1999) Comparative painting reveals strong chromosome homology over 80 million years of bird evolution. Chromosome Res 7: Shibusawa M, Minai S, Nishida-Umehara C, Suzuki T, Mano T, Yamada K, Namikawa T, Matsuda Y (2001) A comparative cytogenetic study of chromosome homology between chicken and Japanese quail. Cytogenet Cell Genet 95: Shibusawa M, Nishida-Umehara C, Masabanda J, Griffin DK, Isobe T, Matsuda Y (2002) Chromosome rearrangements between chicken and guinea fowl defined by comparative chromosome painting and FISH mapping of DNA clones. Cytogenet Cell Genet 98: Shibusawa M, Nishida-Umehara C, Tsudzuki M, Masabanda J, Griffin DK, Matsuda Y (2004a) A comparative karyological study of the blue-breasted quail (Coturnix chinensis, Phasianidae) and California quail (Callipepla californica, Odontophoridae). Cytogenet Genome Res 106: Shibusawa M, Nishibori M, Nishida-Umehara C, Tsudzuki M, Masabanda J, Griffin DK, Matsuda Y (2004b) Karyotypic evolution in the Galliformes: An examination of the process of karyotypic evolution by comparison of the molecular cytogenetic 23
25 findings with the molecular phylogeny. Cytogenet Genome Res 106: Sibley CG, Ahlquist JE (1990) Phylogeny and Classification of Birds: A study in molecular evolution. New Haven: Yale University Press. Sumner AT (1972) A simple technique for demonstrating centromeric heterochromatin. Exptl Cell Res 75: Suzuki T, Kurosaki T, Shimada K et al. (1999) Cytogenetic mapping of 31 functional genes on chicken chromosomes by direct R-banding FISH. Cytogenet Cell Genet 87: Takagi N, Sasaki M (1974) A phylogenetic study of bird karyotypes. Chromosoma 46: Takagi N, Itoh M, Sasaki M (1972) Chromosome studies in four species of Ratitae (Aves). Chromosoma 36: Tsuda Y, Nishida-Umehara C, Ishijima J, Yamada Y, Matsuda Y (2007) Comparison of the Z and W sex chromosomal architectures in elegant crested tinamou (Eudromia elegans) and ostrich (Struthio camelus) and the process of sex chromosome differentiation in palaeognathous birds. Chromosoma 116: van Tuinen M, Sibley CG, Hedges SB (1998) Phylogeny and biogeography of ratite birds inferred from DNA sequences of the mitochondrial ribosomal genes. Mol Biol Evol 15: van Tuinen M, Sibley CG, Hedges SB (2000) The early history of modern birds inferred from DNA sequences of nuclear and mitochondrial ribosomal genes. Mol Biol Evol 17: van Tuinen M, Hedges SB (2001) Calibration of avian molecular clock. Mol Biol Evol 24
26 18:
27 Figure legends Figure 1. Nine largest autosomal pairs and ZW sex chromosomes of six palaeognathous bird species represented by conventional Giemsa-staining. (a) emu (Dromaius novaehollandiae). (b) double-wattled cassowary (Casuarius casuarius). (c) ostrich (Struthio camelus). (d) greater rhea (Rhea americana). (e) lesser rhea (Pterocnemia pennata). (f) elegant crested tinamou (Eudromia elegans). Figure 2. Chromosome painting with chicken chromosome 1-9 probes to PI-stained R-banded metaphase spreads of double-wattled cassowary (Casuarius casuarius) female. Scale bar indicates 10 µm. Figure 3. Chromosome painting with chicken chromosome 1-9 probes to PI-stained R-banded metaphase spreads of elegant crested tinamou (Eudromia elegans) female. Scale bar indicates 10 µm. Figure 4. Chromosome painting with chicken chromosome Z probe to metaphase spreads of the ZW females of five Struthioniformes species and one Tinamiformes species. (a, b) emu (Dromaius novaehollandiae, DNO). (c) double-wattled cassowary (Casuarius casuarius, CCA). (d, e) ostrich (Struthio camelus, SCA). (f) greater rhea (Rhea americana, RAM). (g, h) lesser rhea (Pterocnemia pennata, PPE). (i) elegant crested tinamou (Eudromia elegans, EEL). (b), (e) and (h) are Hoechst-stained patterns of the PI-stained R-banded metaphase spreads shown in (a), (d) and (g), respectively, 26
28 which show the same banding patterns as G-banding. Scale bar indicates 10 µm. Figure 5. Chromosomal distribution of the 18S-28S ribosomal RNA genes and the nucleolar organizer regions (NORs) on metaphase spreads of the ZW females of five Struthioniformes species and one Tinamiformes species. (a-c) lesser rhea (Pterocnemia pennata, PPE). (d) emu (Dromaius novaehollandiae, DNO). (e) double-wattled cassowary (Casuarius casuarius, CCA). (f) ostrich (Struthio camelus, SCA). (g) greater rhea (Rhea americana, RAM). (h, i) elegant crested tinamou (Eudromia elegans, EEL). (a, d, e, f, g, h) FISH patterns of the 18S-28S ribosomal RNA genes on PI-stained R-banded metaphase spreads. (c) and (i) are Ag-NOR-stained patterns of the metaphase spreads shown in (a, b) and (h), respectively. (b) Hoechst-stained pattern of the metaphase spread shown in (a, c), which shows the same banding pattern as G-banding. Scale bar indicates 10 µm. Figure 6. FISH mapping of the ACO1/IREBP, CHD1 and ZOV3 genes and the EE0.6 sequence on PI-stained R-banded metaphase spreads of lesser rhea (Pterocnemia pennata) female. (a) FISH mapping of ACO1/IREBP gene using a cosmid clone as biotinylated probe on PI-stained metaphase spread. (b) Detection of the CHD1 gene on PI-stained metaphase spread using a 4.5 kb genomic DNA fragment as biotinylated probe. (c) Simultaneous detection of the ACO1/IREBP and ZOV3 genes and the EE0.6 sequence on the Z and W chromosomes. The FITC signals of biotin-labeled ACO1/IRFBP and EE0.6 (greenish-yellow) and the rhodamin signals of DIG-labeled 27
29 ZOV3 (red) are localized to the Z and W sex chromosomes. The order of the clones is ACO1/IREBP - ZOV3 - EE0.6 from the proximal on the Z chromosome, whereas ACO1/IREBP is deleted on the W chromosome. Scale bar indicates 10 µm. Figure 7. Summary of Giemsa-stained, C-banded and GBG-banded patterns and chromosome painting patterns with the chicken Z probe of the Z and W chromosomes, and the locations of four DNA markers on the Z and W chromosomes in emu (Dromaius novaehollandiae, DNO), double-wattled cassowary (Casuarius casuarius, CCA), ostrich (Struthio camelus, SCA), greater rhea (Rhea americana, RAM) and lesser rhea (Pterocnemia pennata, PPE) of the Struthioniformes, elegant crested tinamou (Eudromia elegans, EEL) of the Tinamiformes and chicken (Gallus gallus, GGA). The chromosomal locations of ACO1/IREBP, ZOV3 and EE0.6 on the Z and W chromosomes of emu and ostrich, and double-wattled cassowary were taken from Ogawa et al. (1998) and Nishida-Umehara et al. (1999), respectively. The chromosomal location of CHD1 in ostrich and the location of ACO1/IREBP and CHD1 in elegant crested tinamou were taken from Tsuda et al. (2007). 28
30 Table 1. Comparative chromosome painting in six Palaeognathae species with chicken chromosome paints 1-9 and Z Species 2n Chromosome chicken Gallus domesticus 78 GGA1 GGA2 GGA3 GGA4 GGA5 GGA6 GGA7 GGA8 GGA9 emu a Dromaius novaehollandiae m 5 x x x x cassowary Casuarius casuarius m 5 x x x x ostrich Struthio camelus m 5 x x x x greater rhea b Rhea americana m 5 x x x x lesser rhea Pterocnemia pennata m 5 x x x x tinamou Eudromia elegans m 5 x x x x x : the corresponding chromosome detected by chicken probe m : microchromosome a Shetty et al b The data of chromosome painting with GGA1-9 were taken from Guttenbach et al. 2003
31
32
33
34
35
36
37
Different origins of bird and reptile sex chromosomes inferred from comparative mapping of chicken Z-linked genes
 Evolution DOI: 10.1159/000103169 Different origins of bird and reptile sex chromosomes inferred from comparative mapping of chicken Z-linked genes a a, b b b c A. Kawai C. Nishida-Umehara J. Ishijima Y.
Evolution DOI: 10.1159/000103169 Different origins of bird and reptile sex chromosomes inferred from comparative mapping of chicken Z-linked genes a a, b b b c A. Kawai C. Nishida-Umehara J. Ishijima Y.
Molecular study for the sex identification in Japanese quails (Coturnix Japonica) Iran.
 Iran.") Molecular study for the sex identification in Japanese quails (Coturnix Japonica) Nasrollah Vali1 1 and Abbas Doosti 2 1 Department of Animal Sciences, Faculty of Agriculture, Islamic Azad University,
Molecular study for the sex identification in Japanese quails (Coturnix Japonica) Nasrollah Vali1 1 and Abbas Doosti 2 1 Department of Animal Sciences, Faculty of Agriculture, Islamic Azad University,
Abstract. Journal of Heredity 2013:104(6): doi: /jhered/est061
: doi: /jhered/est061") Journal of Heredity 2013:104(6):798 806 doi:10.1093/jhered/est061 The American Genetic Association 2013. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com Highly Species-Specific
Journal of Heredity 2013:104(6):798 806 doi:10.1093/jhered/est061 The American Genetic Association 2013. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com Highly Species-Specific
CHROMOSOMA 9 Springer-Verlag Behaviour of the ZW Sex Bivalent in the Snake Bothrops jararaca. Chromosoma (Berl.) 83, (1981)
 83, (1981)") Chromosoma (Berl.) 83, 289-293 (1981) CHROMOSOMA 9 Springer-Verlag 1981 Behaviour of the ZW Sex Bivalent in the Snake Bothrops jararaca Maria Luiza Be~ak* and Willy Be~ak Servigo de Gen~tica, Instituto
Chromosoma (Berl.) 83, 289-293 (1981) CHROMOSOMA 9 Springer-Verlag 1981 Behaviour of the ZW Sex Bivalent in the Snake Bothrops jararaca Maria Luiza Be~ak* and Willy Be~ak Servigo de Gen~tica, Instituto
Genetics and Molecular Biology, 34, 4, (2011) Copyright 2011, Sociedade Brasileira de Genética. Printed in Brazil
 Copyright 2011, Sociedade Brasileira de Genética. Printed in Brazil") Short Communication Genetics and Molecular Biology, 34, 4, 582-586 (2011) Copyright 2011, Sociedade Brasileira de Genética. Printed in Brazil www.sbg.org.br Chromosomal localization of the 18S-28S and
Short Communication Genetics and Molecular Biology, 34, 4, 582-586 (2011) Copyright 2011, Sociedade Brasileira de Genética. Printed in Brazil www.sbg.org.br Chromosomal localization of the 18S-28S and
Lecture 11 Wednesday, September 19, 2012
 Lecture 11 Wednesday, September 19, 2012 Phylogenetic tree (phylogeny) Darwin and classification: In the Origin, Darwin said that descent from a common ancestral species could explain why the Linnaean
Lecture 11 Wednesday, September 19, 2012 Phylogenetic tree (phylogeny) Darwin and classification: In the Origin, Darwin said that descent from a common ancestral species could explain why the Linnaean
Kornsorn Srikulnath & Yoshinobu Uno & Chizuko Nishida & Yoichi Matsuda
 Chromosome Res (2013) 21:805 819 DOI 10.1007/s10577-013-9398-0 Karyotype evolution in monitor lizards: cross-species chromosome mapping of cdna reveals highly conserved synteny and gene order in the Toxicofera
Chromosome Res (2013) 21:805 819 DOI 10.1007/s10577-013-9398-0 Karyotype evolution in monitor lizards: cross-species chromosome mapping of cdna reveals highly conserved synteny and gene order in the Toxicofera
NOR association in Canis familiaris
 NOR association in Canis familiaris M Rønne, BS Poulsen, Y Shibasaki Odense University, Institute of Medical Biology, Department of Anatomy and Cytology, Campusvej 55, DK-5230 Odense M, Denmark (Proceedings
NOR association in Canis familiaris M Rønne, BS Poulsen, Y Shibasaki Odense University, Institute of Medical Biology, Department of Anatomy and Cytology, Campusvej 55, DK-5230 Odense M, Denmark (Proceedings
6. The lifetime Darwinian fitness of one organism is greater than that of another organism if: A. it lives longer than the other B. it is able to outc
 1. The money in the kingdom of Florin consists of bills with the value written on the front, and pictures of members of the royal family on the back. To test the hypothesis that all of the Florinese $5
1. The money in the kingdom of Florin consists of bills with the value written on the front, and pictures of members of the royal family on the back. To test the hypothesis that all of the Florinese $5
Modern Evolutionary Classification. Lesson Overview. Lesson Overview Modern Evolutionary Classification
 Lesson Overview 18.2 Modern Evolutionary Classification THINK ABOUT IT Darwin s ideas about a tree of life suggested a new way to classify organisms not just based on similarities and differences, but
Lesson Overview 18.2 Modern Evolutionary Classification THINK ABOUT IT Darwin s ideas about a tree of life suggested a new way to classify organisms not just based on similarities and differences, but
9. Summary & General Discussion CHAPTER 9 SUMMARY & GENERAL DISCUSSION
 9. Summary & General Discussion CHAPTER 9 SUMMARY & GENERAL DISCUSSION 143 The Evolution of the Paleognathous Birds 144 9. Summary & General Discussion General Summary The evolutionary history of the Palaeognathae
9. Summary & General Discussion CHAPTER 9 SUMMARY & GENERAL DISCUSSION 143 The Evolution of the Paleognathous Birds 144 9. Summary & General Discussion General Summary The evolutionary history of the Palaeognathae
Inference of the Protokaryotypes of Amniotes and Tetrapods and the Evolutionary Processes of Microchromosomes from Comparative Gene Mapping
 Inference of the Protokaryotypes of Amniotes and Tetrapods and the Evolutionary Processes of Microchromosomes from Comparative Gene Mapping Yoshinobu Uno 1, Chizuko Nishida 2, Hiroshi Tarui 3 a, Satoshi
Inference of the Protokaryotypes of Amniotes and Tetrapods and the Evolutionary Processes of Microchromosomes from Comparative Gene Mapping Yoshinobu Uno 1, Chizuko Nishida 2, Hiroshi Tarui 3 a, Satoshi
Phylogeny and Biogeography of Ratite Birds Inferred from DNA Sequences of the Mitochondrial Ribosomal Genes
 Phylogeny and Biogeography of Ratite Birds Inferred from DNA Sequences of the Mitochondrial Ribosomal Genes Marcel van Tuinen,* Charles G. Sibley, and S. Blair Hedges* *Department of Biology and Institute
Phylogeny and Biogeography of Ratite Birds Inferred from DNA Sequences of the Mitochondrial Ribosomal Genes Marcel van Tuinen,* Charles G. Sibley, and S. Blair Hedges* *Department of Biology and Institute
The karyotype and C-banding pattern of domestic Greylag geese Anser anser anser. Populations (Aves: Anatidae) in Egypt
 in Egypt") PL-ISSN 0015-5497 (print), ISSN 1734-9168 (online) Folia Biologica (Kraków), vol. 62 (2014), No 1 Institute of Systematics and Evolution of Animals, PAS, Kraków, 2014 doi:10.3409/fb62_1.49 Karyotype and
PL-ISSN 0015-5497 (print), ISSN 1734-9168 (online) Folia Biologica (Kraków), vol. 62 (2014), No 1 Institute of Systematics and Evolution of Animals, PAS, Kraków, 2014 doi:10.3409/fb62_1.49 Karyotype and
Species: Panthera pardus Genus: Panthera Family: Felidae Order: Carnivora Class: Mammalia Phylum: Chordata
 CHAPTER 6: PHYLOGENY AND THE TREE OF LIFE AP Biology 3 PHYLOGENY AND SYSTEMATICS Phylogeny - evolutionary history of a species or group of related species Systematics - analytical approach to understanding
CHAPTER 6: PHYLOGENY AND THE TREE OF LIFE AP Biology 3 PHYLOGENY AND SYSTEMATICS Phylogeny - evolutionary history of a species or group of related species Systematics - analytical approach to understanding
Molecular evidence for hybridisation between the two living species of South American ratites: potential. conservation implications
 Molecular evidence for hybridisation between the two living species of South American ratites: potential conservation implications Frédéric Delsuc, Mariella Superina, Guillermo Ferraris, Marie-Ka Tilak,
Molecular evidence for hybridisation between the two living species of South American ratites: potential conservation implications Frédéric Delsuc, Mariella Superina, Guillermo Ferraris, Marie-Ka Tilak,
107. Segregation o f Karyotypes in the F2 Generation o f the Hybrids between Mauritius and Oceanian Type Black Rats with a Note on their Litter Size*'
 No. 9] Proc. Japan Acad., 5'6, Ser. B (1980) 557 107. Segregation o f Karyotypes in the F2 Generation o f the Hybrids between Mauritius and Oceanian Type Black Rats with a Note on their Litter Size*' By
No. 9] Proc. Japan Acad., 5'6, Ser. B (1980) 557 107. Segregation o f Karyotypes in the F2 Generation o f the Hybrids between Mauritius and Oceanian Type Black Rats with a Note on their Litter Size*' By
THE PALAEOGNATHOUS PTERYGOID-PALATINUM COMPLEX. A TRUE CHARACTER?
 2. The Palaeognathous Pterygoid-Palatinum Complex. A True Character? CHAPTER 2 THE PALAEOGNATHOUS PTERYGOID-PALATINUM COMPLEX. A TRUE CHARACTER? Summary Molecular analyses show that modern birds can be
2. The Palaeognathous Pterygoid-Palatinum Complex. A True Character? CHAPTER 2 THE PALAEOGNATHOUS PTERYGOID-PALATINUM COMPLEX. A TRUE CHARACTER? Summary Molecular analyses show that modern birds can be
Highly Differentiated ZW Sex Microchromosomes in the Australian Varanus Species Evolved through Rapid Amplification of Repetitive Sequences
 Highly Differentiated ZW Sex Microchromosomes in the Australian Varanus Species Evolved through Rapid Amplification of Repetitive Sequences Kazumi Matsubara 1 *, Stephen D. Sarre 1, Arthur Georges 1, Yoichi
Highly Differentiated ZW Sex Microchromosomes in the Australian Varanus Species Evolved through Rapid Amplification of Repetitive Sequences Kazumi Matsubara 1 *, Stephen D. Sarre 1, Arthur Georges 1, Yoichi
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes)
") Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
CLADISTICS Student Packet SUMMARY Phylogeny Phylogenetic trees/cladograms
 CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
Geo 302D: Age of Dinosaurs. LAB 7: Dinosaur diversity- Saurischians
 Geo 302D: Age of Dinosaurs LAB 7: Dinosaur diversity- Saurischians Last lab you were presented with a review of major ornithischian clades. You also were presented with some of the kinds of plants that
Geo 302D: Age of Dinosaurs LAB 7: Dinosaur diversity- Saurischians Last lab you were presented with a review of major ornithischian clades. You also were presented with some of the kinds of plants that
A ZZ/ZW microchromosome system in the spiny softshell turtle, Apalone spinifera, reveals an intriguing sex chromosome conservation in Trionychidae
 Chromosome Res (2013) 21:137 147 DOI 10.1007/s10577-013-9343-2 A ZZ/ZW microchromosome system in the spiny softshell turtle, Apalone spinifera, reveals an intriguing sex chromosome conservation in Trionychidae
Chromosome Res (2013) 21:137 147 DOI 10.1007/s10577-013-9343-2 A ZZ/ZW microchromosome system in the spiny softshell turtle, Apalone spinifera, reveals an intriguing sex chromosome conservation in Trionychidae
Time Lapse. A glimpse into prehistoric genomics. DK Griffin University of Kent CoGen 2018 No conflict of interest
 Time Lapse A glimpse into prehistoric genomics DK Griffin University of Kent CoGen 2018 No conflict of interest This is not a talk about time lapse imaging Karyomapping So, where do we start? Karyomapping
Time Lapse A glimpse into prehistoric genomics DK Griffin University of Kent CoGen 2018 No conflict of interest This is not a talk about time lapse imaging Karyomapping So, where do we start? Karyomapping
Research Note. A novel method for sexing day-old chicks using endoscope system
 Research Note A novel method for sexing day-old chicks using endoscope system Makoto Otsuka,,1 Osamu Miyashita,,1 Mitsuru Shibata,,1 Fujiyuki Sato,,1 and Mitsuru Naito,2,3 NARO Institute of Livestock and
Research Note A novel method for sexing day-old chicks using endoscope system Makoto Otsuka,,1 Osamu Miyashita,,1 Mitsuru Shibata,,1 Fujiyuki Sato,,1 and Mitsuru Naito,2,3 NARO Institute of Livestock and
Kazumi Matsubara *, Theresa Knopp, Stephen D Sarre, Arthur Georges and Tariq Ezaz *
 Matsubara et al. Molecular Cytogenetics 2013, 6:60 RESEARCH Open Access Karyotypic analysis and FISH mapping of microsatellite motifs reveal highly differentiated XX/XY sex chromosomes in the pink-tailed
Matsubara et al. Molecular Cytogenetics 2013, 6:60 RESEARCH Open Access Karyotypic analysis and FISH mapping of microsatellite motifs reveal highly differentiated XX/XY sex chromosomes in the pink-tailed
Kazumi Matsubara 1,2,5*, Chizuko Nishida 3, Yoichi Matsuda 2,4 and Yoshinori Kumazawa 1
 Matsubara et al. Zoological Letters (2016) 2:19 DOI 10.1186/s40851-016-0056-1 RESEARCH ARTICLE Open Access Sex chromosome evolution in snakes inferred from divergence patterns of two gametologous genes
Matsubara et al. Zoological Letters (2016) 2:19 DOI 10.1186/s40851-016-0056-1 RESEARCH ARTICLE Open Access Sex chromosome evolution in snakes inferred from divergence patterns of two gametologous genes
Name Kornsorn Srikulnath Position Lecturer (Kasetsart University) Researcher (Reptile Cytogenetics, Nagoya University)
 Researcher (Reptile Cytogenetics, Nagoya University)") Name Kornsorn Srikulnath Position Lecturer (Kasetsart University) Researcher (Reptile Cytogenetics, Nagoya University) Associate Editor of Thai Journal of Genetics Tel. +66-25625444 ext.4240 Email address:
Name Kornsorn Srikulnath Position Lecturer (Kasetsart University) Researcher (Reptile Cytogenetics, Nagoya University) Associate Editor of Thai Journal of Genetics Tel. +66-25625444 ext.4240 Email address:
Bi156 Lecture 1/13/12. Dog Genetics
 Bi156 Lecture 1/13/12 Dog Genetics The radiation of the family Canidae occurred about 100 million years ago. Dogs are most closely related to wolves, from which they diverged through domestication about
Bi156 Lecture 1/13/12 Dog Genetics The radiation of the family Canidae occurred about 100 million years ago. Dogs are most closely related to wolves, from which they diverged through domestication about
A new karyotypic formula for the genus Amphisbaena (Squamata: Amphisbaenidae)
") Phyllomedusa 9(1):75-80, 2010 2010 Departamento de Ciências Biológicas - ESALQ - USP ISSN 1519-1397 Short Communication A new karyotypic formula for the genus Amphisbaena (Squamata: Amphisbaenidae) Camila
Phyllomedusa 9(1):75-80, 2010 2010 Departamento de Ciências Biológicas - ESALQ - USP ISSN 1519-1397 Short Communication A new karyotypic formula for the genus Amphisbaena (Squamata: Amphisbaenidae) Camila
Karyotype, constitutive heterochromatin and nucleolus organizer regions in two species of Liolaemus (Squamata, Tropiduridae)
") CARYOLOGIA Vol. 56, no. 3: 269-273, 2003 Karyotype, constitutive heterochromatin and nucleolus organizer regions in two species of Liolaemus (Squamata, Tropiduridae) ALEJANDRA HERNANDO Departamento de
CARYOLOGIA Vol. 56, no. 3: 269-273, 2003 Karyotype, constitutive heterochromatin and nucleolus organizer regions in two species of Liolaemus (Squamata, Tropiduridae) ALEJANDRA HERNANDO Departamento de
History of Lineages. Chapter 11. Jamie Oaks 1. April 11, Kincaid Hall 524. c 2007 Boris Kulikov boris-kulikov.blogspot.
 History of Lineages Chapter 11 Jamie Oaks 1 1 Kincaid Hall 524 joaks1@gmail.com April 11, 2014 c 2007 Boris Kulikov boris-kulikov.blogspot.com History of Lineages J. Oaks, University of Washington 1/46
History of Lineages Chapter 11 Jamie Oaks 1 1 Kincaid Hall 524 joaks1@gmail.com April 11, 2014 c 2007 Boris Kulikov boris-kulikov.blogspot.com History of Lineages J. Oaks, University of Washington 1/46
UNIT III A. Descent with Modification(Ch19) B. Phylogeny (Ch20) C. Evolution of Populations (Ch21) D. Origin of Species or Speciation (Ch22)
 B. Phylogeny (Ch20) C. Evolution of Populations (Ch21) D. Origin of Species or Speciation (Ch22)") UNIT III A. Descent with Modification(Ch9) B. Phylogeny (Ch2) C. Evolution of Populations (Ch2) D. Origin of Species or Speciation (Ch22) Classification in broad term simply means putting things in classes
UNIT III A. Descent with Modification(Ch9) B. Phylogeny (Ch2) C. Evolution of Populations (Ch2) D. Origin of Species or Speciation (Ch22) Classification in broad term simply means putting things in classes
Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported
 Drawing of early ankle development of Alligator mississippiensis, as reported") Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported by a previous study 1. The intermedium is formed at
Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported by a previous study 1. The intermedium is formed at
Karyotypic polymorphism of the zebra finch Z chromosome
 Chromosoma (2011) 120:255 264 DOI 10.1007/s00412-010-0308-3 RESEARCH ARTICLE Karyotypic polymorphism of the zebra finch Z chromosome Yuichiro Itoh & Kathy Kampf & Christopher N. Balakrishnan & Arthur P.
Chromosoma (2011) 120:255 264 DOI 10.1007/s00412-010-0308-3 RESEARCH ARTICLE Karyotypic polymorphism of the zebra finch Z chromosome Yuichiro Itoh & Kathy Kampf & Christopher N. Balakrishnan & Arthur P.
DIVISION 056 IMPORTATION, POSSESSION, CONFINEMENT, TRANSPORTATION AND SALE OF NONNATIVE WILDLIFE
 DIVISION 056 IMPORTATION, POSSESSION, CONFINEMENT, TRANSPORTATION AND SALE OF NONNATIVE WILDLIFE 635 056 0010 Definitions For the purposes of these rules, the definitions in ORS 496.004 and OAR 635 045
DIVISION 056 IMPORTATION, POSSESSION, CONFINEMENT, TRANSPORTATION AND SALE OF NONNATIVE WILDLIFE 635 056 0010 Definitions For the purposes of these rules, the definitions in ORS 496.004 and OAR 635 045
Supporting Online Material
 Supporting Online Material Supporting Text: Rapprochement in dating the early branching of modern mammals It is important to distinguish the meaning of nodes in the tree (Fig. S1): successive branching
Supporting Online Material Supporting Text: Rapprochement in dating the early branching of modern mammals It is important to distinguish the meaning of nodes in the tree (Fig. S1): successive branching
The Karyotype of Plestiodon anthracinus (Baird, 1850) (Sauria: Scincidae): A Step Toward Solving an Enigma
 (Sauria: Scincidae): A Step Toward Solving an Enigma") 2017 2017 SOUTHEASTERN Southeastern Naturalist NATURALIST 16(3):326 330 The Karyotype of Plestiodon anthracinus (Baird, 1850) (Sauria: Scincidae): A Step Toward Solving an Enigma Laurence M. Hardy 1, *,
2017 2017 SOUTHEASTERN Southeastern Naturalist NATURALIST 16(3):326 330 The Karyotype of Plestiodon anthracinus (Baird, 1850) (Sauria: Scincidae): A Step Toward Solving an Enigma Laurence M. Hardy 1, *,
PHYSICAL MAP OF THE AUSTRALIAN CENTRAL BEARDED DRAGON. (Pogona vitticeps) AND COMPARATIVE MAPPING AMONG DRAGONS. (Squamata, Agamidae) AND AMNIOTES
 AND COMPARATIVE MAPPING AMONG DRAGONS. (Squamata, Agamidae) AND AMNIOTES") PHYSICAL MAP OF THE AUSTRALIAN CENTRAL BEARDED DRAGON (Pogona vitticeps) AND COMPARATIVE MAPPING AMONG DRAGONS (Squamata, Agamidae) AND AMNIOTES By MATTHEW JOHN YOUNG B. Environmental Science Institute
PHYSICAL MAP OF THE AUSTRALIAN CENTRAL BEARDED DRAGON (Pogona vitticeps) AND COMPARATIVE MAPPING AMONG DRAGONS (Squamata, Agamidae) AND AMNIOTES By MATTHEW JOHN YOUNG B. Environmental Science Institute
1 Describe the anatomy and function of the turtle shell. 2 Describe respiration in turtles. How does the shell affect respiration?
 GVZ 2017 Practice Questions Set 1 Test 3 1 Describe the anatomy and function of the turtle shell. 2 Describe respiration in turtles. How does the shell affect respiration? 3 According to the most recent
GVZ 2017 Practice Questions Set 1 Test 3 1 Describe the anatomy and function of the turtle shell. 2 Describe respiration in turtles. How does the shell affect respiration? 3 According to the most recent
> BACK TO CONTENTS PAGE
 Human interaction: previously pursued for their feathers; nowadays farmed for meat. In the wild they will attack if threatened (treacherous kick); passive in captive environments. If raised, they may display
Human interaction: previously pursued for their feathers; nowadays farmed for meat. In the wild they will attack if threatened (treacherous kick); passive in captive environments. If raised, they may display
Ch 1.2 Determining How Species Are Related.notebook February 06, 2018
 Name 3 "Big Ideas" from our last notebook lecture: * * * 1 WDYR? Of the following organisms, which is the closest relative of the "Snowy Owl" (Bubo scandiacus)? a) barn owl (Tyto alba) b) saw whet owl
Name 3 "Big Ideas" from our last notebook lecture: * * * 1 WDYR? Of the following organisms, which is the closest relative of the "Snowy Owl" (Bubo scandiacus)? a) barn owl (Tyto alba) b) saw whet owl
CHAPTER 6 CRANIAL KINESIS IN PALAEOGNATHOUS BIRDS. 6. Cranial Kinesis in Palaeognathous Birds
 6. Cranial Kinesis in Palaeognathous Birds CHAPTER 6 CRANIAL KINESIS IN PALAEOGNATHOUS BIRDS Summary In palaeognathous birds the morphology of the Pterygoid-Palatinum Complex (PPC) is remarkably different
6. Cranial Kinesis in Palaeognathous Birds CHAPTER 6 CRANIAL KINESIS IN PALAEOGNATHOUS BIRDS Summary In palaeognathous birds the morphology of the Pterygoid-Palatinum Complex (PPC) is remarkably different
PCR detection of Leptospira in. stray cat and
 PCR detection of Leptospira in 1 Department of Pathology, School of Veterinary Medicine, Islamic Azad University, Shahrekord Branch, Shahrekord, Iran 2 Department of Microbiology, School of Veterinary
PCR detection of Leptospira in 1 Department of Pathology, School of Veterinary Medicine, Islamic Azad University, Shahrekord Branch, Shahrekord, Iran 2 Department of Microbiology, School of Veterinary
University of Groningen
 University of Groningen No sexual differences in embryonic period in jackdaws Corvus monedula and black-headed gulls Larus ridibundus Salomons, Henri; Mueller, Wendt; Dijkstra, C; Eising, Corine; Verhulst,
University of Groningen No sexual differences in embryonic period in jackdaws Corvus monedula and black-headed gulls Larus ridibundus Salomons, Henri; Mueller, Wendt; Dijkstra, C; Eising, Corine; Verhulst,
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
May 10, SWBAT analyze and evaluate the scientific evidence provided by the fossil record.
 May 10, 2017 Aims: SWBAT analyze and evaluate the scientific evidence provided by the fossil record. Agenda 1. Do Now 2. Class Notes 3. Guided Practice 4. Independent Practice 5. Practicing our AIMS: E.3-Examining
May 10, 2017 Aims: SWBAT analyze and evaluate the scientific evidence provided by the fossil record. Agenda 1. Do Now 2. Class Notes 3. Guided Practice 4. Independent Practice 5. Practicing our AIMS: E.3-Examining
RICKETTSIA SPECIES AMONG TICKS IN AN AREA OF JAPAN ENDEMIC FOR JAPANESE SPOTTED FEVER
 RICKETTSIA SPECIES AMONG TICKS IN AN AREA OF JAPAN ENDEMIC FOR JAPANESE SPOTTED FEVER Makoto Kondo 1, Katsuhiko Ando 2, Keiichi Yamanaka 1 and Hitoshi Mizutani 1 1 Department of Dermatology, 2 Department
RICKETTSIA SPECIES AMONG TICKS IN AN AREA OF JAPAN ENDEMIC FOR JAPANESE SPOTTED FEVER Makoto Kondo 1, Katsuhiko Ando 2, Keiichi Yamanaka 1 and Hitoshi Mizutani 1 1 Department of Dermatology, 2 Department
Giant croc with T. rex teeth roamed Madagascar
 Giant croc with T. rex teeth roamed Madagascar www.scimex.org/newsfeed/giant-croc-with-t.-rex-teeth-used-to-roam-in-madagascar Embargoed until: Publicly released: PeerJ A fossil of the largest and oldest
Giant croc with T. rex teeth roamed Madagascar www.scimex.org/newsfeed/giant-croc-with-t.-rex-teeth-used-to-roam-in-madagascar Embargoed until: Publicly released: PeerJ A fossil of the largest and oldest
BioSci 110, Fall 08 Exam 2
 1. is the cell division process that results in the production of a. mitosis; 2 gametes b. meiosis; 2 gametes c. meiosis; 2 somatic (body) cells d. mitosis; 4 somatic (body) cells e. *meiosis; 4 gametes
1. is the cell division process that results in the production of a. mitosis; 2 gametes b. meiosis; 2 gametes c. meiosis; 2 somatic (body) cells d. mitosis; 4 somatic (body) cells e. *meiosis; 4 gametes
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1
 Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
Bioinformatics: Investigating Molecular/Biochemical Evidence for Evolution
 Bioinformatics: Investigating Molecular/Biochemical Evidence for Evolution Background How does an evolutionary biologist decide how closely related two different species are? The simplest way is to compare
Bioinformatics: Investigating Molecular/Biochemical Evidence for Evolution Background How does an evolutionary biologist decide how closely related two different species are? The simplest way is to compare
In the first half of the 20th century, Dr. Guido Fanconi published detailed clinical descriptions of several heritable human diseases.
 In the first half of the 20th century, Dr. Guido Fanconi published detailed clinical descriptions of several heritable human diseases. Two disease syndromes were named after him: Fanconi Anemia and Fanconi
In the first half of the 20th century, Dr. Guido Fanconi published detailed clinical descriptions of several heritable human diseases. Two disease syndromes were named after him: Fanconi Anemia and Fanconi
Evolution in dogs. Megan Elmore CS374 11/16/2010. (thanks to Dan Newburger for many slides' content)
") Evolution in dogs Megan Elmore CS374 11/16/2010 (thanks to Dan Newburger for many slides' content) Papers for today Vonholdt BM et al (2010). Genome-wide SNP and haplotype analyses reveal a rich history
Evolution in dogs Megan Elmore CS374 11/16/2010 (thanks to Dan Newburger for many slides' content) Papers for today Vonholdt BM et al (2010). Genome-wide SNP and haplotype analyses reveal a rich history
Turtles (Testudines) Abstract
 Abstract") Turtles (Testudines) H. Bradley Shaffer Department of Evolution and Ecology, University of California, Davis, CA 95616, USA (hbshaffer@ucdavis.edu) Abstract Living turtles and tortoises consist of two
Turtles (Testudines) H. Bradley Shaffer Department of Evolution and Ecology, University of California, Davis, CA 95616, USA (hbshaffer@ucdavis.edu) Abstract Living turtles and tortoises consist of two
Effectivenesss of a sexing technique on free-range day-old chick
 Effectivenesss of a sexing technique on free-range day-old chick Hervé Brice Dakpogan a*, Sahidou Salifou a, André Aboh b, Christophe Chrysostome c a Applied Biology Research Laboratory of Polytechnic
Effectivenesss of a sexing technique on free-range day-old chick Hervé Brice Dakpogan a*, Sahidou Salifou a, André Aboh b, Christophe Chrysostome c a Applied Biology Research Laboratory of Polytechnic
PHYSICAL MAP OF THE AUSTRALIAN CENTRAL BEARDED DRAGON. (Pogona vitticeps) AND COMPARATIVE MAPPING AMONG DRAGONS. (Squamata, Agamidae) AND AMNIOTES
 AND COMPARATIVE MAPPING AMONG DRAGONS. (Squamata, Agamidae) AND AMNIOTES") PHYSICAL MAP OF THE AUSTRALIAN CENTRAL BEARDED DRAGON (Pogona vitticeps) AND COMPARATIVE MAPPING AMONG DRAGONS (Squamata, Agamidae) AND AMNIOTES By MATTHEW JOHN YOUNG B. Environmental Science Institute
PHYSICAL MAP OF THE AUSTRALIAN CENTRAL BEARDED DRAGON (Pogona vitticeps) AND COMPARATIVE MAPPING AMONG DRAGONS (Squamata, Agamidae) AND AMNIOTES By MATTHEW JOHN YOUNG B. Environmental Science Institute
Differential human impact on the survival of genetically distinct avian lineages
 Bird Conservation International (1999) 9:147-154. BirdLife International 1999 Differential human impact on the survival of genetically distinct avian lineages AUSTIN L. HUGHES Summary At the present time
Bird Conservation International (1999) 9:147-154. BirdLife International 1999 Differential human impact on the survival of genetically distinct avian lineages AUSTIN L. HUGHES Summary At the present time
INQUIRY & INVESTIGATION
 INQUIRY & INVESTIGTION Phylogenies & Tree-Thinking D VID. UM SUSN OFFNER character a trait or feature that varies among a set of taxa (e.g., hair color) character-state a variant of a character that occurs
INQUIRY & INVESTIGTION Phylogenies & Tree-Thinking D VID. UM SUSN OFFNER character a trait or feature that varies among a set of taxa (e.g., hair color) character-state a variant of a character that occurs
oxfitates Mllsdum M ie'ican Group of Lizards in the Genus Sceloporusl Karyotypes and Evolution of the spinosus COLE2 BY CHARLES J.
 M ie'ican Mllsdum oxfitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N. Y. I0024 NUMBER 243I SEPTEMBER 28, 1970 Karyotypes and Evolution of the spinosus
M ie'ican Mllsdum oxfitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N. Y. I0024 NUMBER 243I SEPTEMBER 28, 1970 Karyotypes and Evolution of the spinosus
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and
 Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
Evolution of Birds. Summary:
 Oregon State Standards OR Science 7.1, 7.2, 7.3, 7.3S.1, 7.3S.2 8.1, 8.2, 8.2L.1, 8.3, 8.3S.1, 8.3S.2 H.1, H.2, H.2L.4, H.2L.5, H.3, H.3S.1, H.3S.2, H.3S.3 Summary: Students create phylogenetic trees to
Oregon State Standards OR Science 7.1, 7.2, 7.3, 7.3S.1, 7.3S.2 8.1, 8.2, 8.2L.1, 8.3, 8.3S.1, 8.3S.2 H.1, H.2, H.2L.4, H.2L.5, H.3, H.3S.1, H.3S.2, H.3S.3 Summary: Students create phylogenetic trees to
First named as a separate species of rodent in 1946, Tokudaia muenninki, also known as
 First named as a separate species of rodent in 1946, Tokudaia muenninki, also known as Muennink s spiny rat or the Okinawa spiny rat, lives in the northern region of Yanbaru Forest on Okinawa Island, Japan.
First named as a separate species of rodent in 1946, Tokudaia muenninki, also known as Muennink s spiny rat or the Okinawa spiny rat, lives in the northern region of Yanbaru Forest on Okinawa Island, Japan.
Current status of the evaluation of genetic diversity in livestock breeds
 1st Globaldiv Workshop, Bydgoszcz Current status of the evaluation of genetic diversity in livestock breeds Groeneveld LF, Lenstra JA, Eding H, Toro MA, Scherf B, Pilling D, Negrini R, Finlay EK, Jianlin
1st Globaldiv Workshop, Bydgoszcz Current status of the evaluation of genetic diversity in livestock breeds Groeneveld LF, Lenstra JA, Eding H, Toro MA, Scherf B, Pilling D, Negrini R, Finlay EK, Jianlin
Video Assignments. Microraptor PBS The Four-winged Dinosaur Mark Davis SUNY Cortland Library Online
 Video Assignments Microraptor PBS The Four-winged Dinosaur Mark Davis SUNY Cortland Library Online Radiolab Apocalyptical http://www.youtube.com/watch?v=k52vd4wbdlw&feature=youtu.be Minute 13 through minute
Video Assignments Microraptor PBS The Four-winged Dinosaur Mark Davis SUNY Cortland Library Online Radiolab Apocalyptical http://www.youtube.com/watch?v=k52vd4wbdlw&feature=youtu.be Minute 13 through minute
A search for sequence similarity between chicken (Gallus domesticus) and ostrich (Struthio camelus) microsatellite markers*
 and ostrich (Struthio camelus) microsatellite markers*") Animal Science Papers and Reports vol. 25 (2007) no. 4, 283-288 Institute of Genetics and Animal Breeding, Jastrzębiec, Poland SHORT REPORT A search for sequence similarity between chicken (Gallus domesticus)
Animal Science Papers and Reports vol. 25 (2007) no. 4, 283-288 Institute of Genetics and Animal Breeding, Jastrzębiec, Poland SHORT REPORT A search for sequence similarity between chicken (Gallus domesticus)
Bio 1B Lecture Outline (please print and bring along) Fall, 2006
 Fall, 2006") Bio 1B Lecture Outline (please print and bring along) Fall, 2006 B.D. Mishler, Dept. of Integrative Biology 2-6810, bmishler@berkeley.edu Evolution lecture #4 -- Phylogenetic Analysis (Cladistics) -- Oct.
Bio 1B Lecture Outline (please print and bring along) Fall, 2006 B.D. Mishler, Dept. of Integrative Biology 2-6810, bmishler@berkeley.edu Evolution lecture #4 -- Phylogenetic Analysis (Cladistics) -- Oct.
Fig Phylogeny & Systematics
 Fig. 26- Phylogeny & Systematics Tree of Life phylogenetic relationship for 3 clades (http://evolution.berkeley.edu Fig. 26-2 Phylogenetic tree Figure 26.3 Taxonomy Taxon Carolus Linnaeus Species: Panthera
Fig. 26- Phylogeny & Systematics Tree of Life phylogenetic relationship for 3 clades (http://evolution.berkeley.edu Fig. 26-2 Phylogenetic tree Figure 26.3 Taxonomy Taxon Carolus Linnaeus Species: Panthera
Title: Phylogenetic Methods and Vertebrate Phylogeny
 Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Complete mitochondrial DNA sequence of Chinese alligator, Alligator sinensis, and phylogeny of crocodiles
 Chinese Science Bulletin 2003 Vol. 48 No. 19 2050 2054 Complete mitochondrial DNA sequence of Chinese alligator, Alligator sinensis, and phylogeny of crocodiles WU Xiaobing 1,3, WANG Yiquan 1,2,ZHOUKaiya
Chinese Science Bulletin 2003 Vol. 48 No. 19 2050 2054 Complete mitochondrial DNA sequence of Chinese alligator, Alligator sinensis, and phylogeny of crocodiles WU Xiaobing 1,3, WANG Yiquan 1,2,ZHOUKaiya
Copy number variation, chromosome rearrangement, and their association with recombination during avian evolution
 Copy number variation, chromosome rearrangement, and their association with recombination during avian evolution Martin Völker, Niclas Backström, Benjamin M. Skinner, et al. Genome Res. 2010 20: 503-511
Copy number variation, chromosome rearrangement, and their association with recombination during avian evolution Martin Völker, Niclas Backström, Benjamin M. Skinner, et al. Genome Res. 2010 20: 503-511
Title. CitationJapanese Journal of Veterinary Research, 24(1-2): 37. Issue Date DOI. Doc URL. Type. File Information
: 37. Issue Date DOI. Doc URL. Type. File Information") Title DISTRIBUTION OF LYMPHATIC TISSUES IN DUCK CAECA Author(s)KITAMURA, Hirokazu; SUGIMURA, Makoto; HASHIMOTO, Yos CitationJapanese Journal of Veterinary Research, 24(1-2): 37 Issue Date 1976-05 DOI 10.14943/jjvr.24.1-2.37
Title DISTRIBUTION OF LYMPHATIC TISSUES IN DUCK CAECA Author(s)KITAMURA, Hirokazu; SUGIMURA, Makoto; HASHIMOTO, Yos CitationJapanese Journal of Veterinary Research, 24(1-2): 37 Issue Date 1976-05 DOI 10.14943/jjvr.24.1-2.37
Animals & Reptiles (PA) LD P KER CHIPS. *** Variations
 LD P KER CHIPS. *** Variations") Animals & Reptiles (PA) LD P KER CHIPS 1 PA-AB thru PA-CW PA-AB Beaver PA-AF Bear *** PA-AJ Dancing Bears Embossed / v:e PA-AP Buffalo Head PA-AS Buffalo Head PA-AV Old Tom *** PA-BC House Cat PA-BG House
Animals & Reptiles (PA) LD P KER CHIPS 1 PA-AB thru PA-CW PA-AB Beaver PA-AF Bear *** PA-AJ Dancing Bears Embossed / v:e PA-AP Buffalo Head PA-AS Buffalo Head PA-AV Old Tom *** PA-BC House Cat PA-BG House
Origin and Evolution of Birds. Read: Chapters 1-3 in Gill but limited review of systematics
 Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Research Article A Comparative Morphometrical Study of the Pecten Oculi in Different Avian Species
 The Scientific World Journal Volume 2013, Article ID 968652, 5 pages http://dx.doi.org/10.1155/2013/968652 Research Article A omparative Morphometrical Study of the Pecten Oculi in Different Avian Species
The Scientific World Journal Volume 2013, Article ID 968652, 5 pages http://dx.doi.org/10.1155/2013/968652 Research Article A omparative Morphometrical Study of the Pecten Oculi in Different Avian Species
Warm-Up: Fill in the Blank
 Warm-Up: Fill in the Blank 1. For natural selection to happen, there must be variation in the population. 2. The preserved remains of organisms, called provides evidence for evolution. 3. By using and
Warm-Up: Fill in the Blank 1. For natural selection to happen, there must be variation in the population. 2. The preserved remains of organisms, called provides evidence for evolution. 3. By using and
Received 16 November 1998; received in revised form and accepted for publication by M. Schmid 8 March 1999
 Chromosome Research 7: 247±254, 1999. # 1999 Kluwer Academic Publishers. Printed in the Netherlands 247 Chromosomal polymorphisms due to supernumerary chromosomes and pericentric inversions in the eyelidless
Chromosome Research 7: 247±254, 1999. # 1999 Kluwer Academic Publishers. Printed in the Netherlands 247 Chromosomal polymorphisms due to supernumerary chromosomes and pericentric inversions in the eyelidless
Caecilians (Gymnophiona)
") Caecilians (Gymnophiona) David J. Gower* and Mark Wilkinson Department of Zoology, The Natural History Museum, London SW7 5BD, UK *To whom correspondence should be addressed (d.gower@nhm. ac.uk) Abstract
Caecilians (Gymnophiona) David J. Gower* and Mark Wilkinson Department of Zoology, The Natural History Museum, London SW7 5BD, UK *To whom correspondence should be addressed (d.gower@nhm. ac.uk) Abstract
In situ and Ex situ gene conservation in Russia
 In situ and Ex situ gene conservation in Russia Osadchaya Olga, Phd, Academic Secretary Bagirov Vugar, Dr. Biol. Sci., Professor, Laboratory Head Zinovieva Natalia, Dr. Biol. Sci., Professor, Director
In situ and Ex situ gene conservation in Russia Osadchaya Olga, Phd, Academic Secretary Bagirov Vugar, Dr. Biol. Sci., Professor, Laboratory Head Zinovieva Natalia, Dr. Biol. Sci., Professor, Director
What are taxonomy, classification, and systematics?
 Topic 2: Comparative Method o Taxonomy, classification, systematics o Importance of phylogenies o A closer look at systematics o Some key concepts o Parts of a cladogram o Groups and characters o Homology
Topic 2: Comparative Method o Taxonomy, classification, systematics o Importance of phylogenies o A closer look at systematics o Some key concepts o Parts of a cladogram o Groups and characters o Homology
BreenLab - Molecular Cytogenetic Investigation of Soft Tissue Sarcoma General information and sample submission requirements
 PARTICIPANTS NEEDED FOR RESEARCH ON CANINE CANCER THE STUDY The research project Cellular Genomics- A molecular cytogenetics investigation of canine soft tissue sarcoma is part of Dr. Matthew Breen s laboratory
PARTICIPANTS NEEDED FOR RESEARCH ON CANINE CANCER THE STUDY The research project Cellular Genomics- A molecular cytogenetics investigation of canine soft tissue sarcoma is part of Dr. Matthew Breen s laboratory
Analysis of CR1 repeats in the zebra finch genome
 Analysis of CR1 repeats in the zebra finch genome George E. Liu, Yali Hou* and Twain Brown Bovine Functional Genomics Laboratory, ANRI, ARS, USDA, Beltsville, Maryland 20705, USA *Also affiliated with
Analysis of CR1 repeats in the zebra finch genome George E. Liu, Yali Hou* and Twain Brown Bovine Functional Genomics Laboratory, ANRI, ARS, USDA, Beltsville, Maryland 20705, USA *Also affiliated with
18 August Puerto Rican Crested Toad Dustin Smith, North Carolina Zoological Park
 18 www.aza.org August 2015 Puerto Rican Crested Toad Dustin Smith, North Carolina Zoological Park MANAGING SSP POPULATIONS WITH MOLECULAR GENETICS BY ALINA TUGEND Are they one species? Are they two? How
18 www.aza.org August 2015 Puerto Rican Crested Toad Dustin Smith, North Carolina Zoological Park MANAGING SSP POPULATIONS WITH MOLECULAR GENETICS BY ALINA TUGEND Are they one species? Are they two? How
sex ratio 5 5 Common Peafowl Rainbow Lorikeet
 MOST-NUMEROUS AVES IN NORTH AMERICAN ISIS INSTITUTIONS December 31, 2011 AND A COMPARISON OF POPULATIONS FROM ONE AND TEN YEARS PAST Robert Webster The Toledo Zoo Current 12-10 species Population/ sex
MOST-NUMEROUS AVES IN NORTH AMERICAN ISIS INSTITUTIONS December 31, 2011 AND A COMPARISON OF POPULATIONS FROM ONE AND TEN YEARS PAST Robert Webster The Toledo Zoo Current 12-10 species Population/ sex
Evolution as Fact. The figure below shows transitional fossils in the whale lineage.
 Evolution as Fact Evolution is a fact. Organisms descend from others with modification. Phylogeny, the lineage of ancestors and descendants, is the scientific term to Darwin's phrase "descent with modification."
Evolution as Fact Evolution is a fact. Organisms descend from others with modification. Phylogeny, the lineage of ancestors and descendants, is the scientific term to Darwin's phrase "descent with modification."
ADOPTED REGULATION OF THE BOARD OF WILDLIFE COMMISSIONERS. LCB File No. R Effective December 30, 2011
 ADOPTED REGULATION OF THE BOARD OF WILDLIFE COMMISSIONERS LCB File No. R056-11 Effective December 30, 2011 EXPLANATION Matter in italics is new; matter in brackets [omitted material] is material to be
ADOPTED REGULATION OF THE BOARD OF WILDLIFE COMMISSIONERS LCB File No. R056-11 Effective December 30, 2011 EXPLANATION Matter in italics is new; matter in brackets [omitted material] is material to be
THREE-DIMENSIONAL KINEMATICS OF SKELETAL ELEMENTS IN AVIAN PROKINETIC AND RHYNCHOKINETIC SKULLS DETERMINED BY ROENTGEN STEREOPHOTOGRAMMETRY
 The Journal of Experimental Biology 4, 1735 1744 (01) Printed in Great Britain The Company of Biologists Limited 01 JEB3132 1735 THREE-DIMENSIONAL KINEMATICS OF SKELETAL ELEMENTS IN AVIAN PROKINETIC AND
The Journal of Experimental Biology 4, 1735 1744 (01) Printed in Great Britain The Company of Biologists Limited 01 JEB3132 1735 THREE-DIMENSIONAL KINEMATICS OF SKELETAL ELEMENTS IN AVIAN PROKINETIC AND
AKC Canine Health Foundation Grant Updates: Research Currently Being Sponsored By The Vizsla Club of America Welfare Foundation
 AKC Canine Health Foundation Grant Updates: Research Currently Being Sponsored By The Vizsla Club of America Welfare Foundation GRANT PROGRESS REPORT REVIEW Grant: 00748: SNP Association Mapping for Canine
AKC Canine Health Foundation Grant Updates: Research Currently Being Sponsored By The Vizsla Club of America Welfare Foundation GRANT PROGRESS REPORT REVIEW Grant: 00748: SNP Association Mapping for Canine
8/19/2013. What is convergence? Topic 11: Convergence. What is convergence? What is convergence? What is convergence? What is convergence?
 Topic 11: Convergence What are the classic herp examples? Have they been formally studied? Emerald Tree Boas and Green Tree Pythons show a remarkable level of convergence Photos KP Bergmann, Philadelphia
Topic 11: Convergence What are the classic herp examples? Have they been formally studied? Emerald Tree Boas and Green Tree Pythons show a remarkable level of convergence Photos KP Bergmann, Philadelphia
Ratite Standards and Guidelines
 Exhibited Animals - Ratite and Australian Animal Welfare and Exhibited Animals - Ratite and December 2011 Page 1 of 18 Exhibited Animals - Ratite and Introduction Purpose The principal purpose of this
Exhibited Animals - Ratite and Australian Animal Welfare and Exhibited Animals - Ratite and December 2011 Page 1 of 18 Exhibited Animals - Ratite and Introduction Purpose The principal purpose of this
Salmonella National Poultry Improvement Plan Washington State Regulations
 Salmonella National Poultry Improvement Plan Washington State Regulations Lyndon Badcoe BVSc,, MVS, DVSc, Avian Health Veterinarian and Epidemiologist Outline Describe Pathogenesis of Salmonellosis in
Salmonella National Poultry Improvement Plan Washington State Regulations Lyndon Badcoe BVSc,, MVS, DVSc, Avian Health Veterinarian and Epidemiologist Outline Describe Pathogenesis of Salmonellosis in
Karyotype of a Ranid Frog, Platymantis pelewensis, from Belau, Micronesia, with Comments on Its Systematic Implications l
 Pacific Science (1995), vol. 49, no. 3: 296-300 1995 by University of Hawai'i Press. All rights reserved Karyotype of a Ranid Frog, Platymantis pelewensis, from Belau, Micronesia, with Comments on Its
Pacific Science (1995), vol. 49, no. 3: 296-300 1995 by University of Hawai'i Press. All rights reserved Karyotype of a Ranid Frog, Platymantis pelewensis, from Belau, Micronesia, with Comments on Its
Population/ sex ratio
 Current MOST-NUMEROUS AVES IN NORTH AMERICAN ISIS INSTITUTIONS** December 31, 2012 AND A COMPARISON OF POPULATIONS FROM ONE AND TEN YEARS PAST Robert Webster The Toledo Zoo Species * - species is represented
Current MOST-NUMEROUS AVES IN NORTH AMERICAN ISIS INSTITUTIONS** December 31, 2012 AND A COMPARISON OF POPULATIONS FROM ONE AND TEN YEARS PAST Robert Webster The Toledo Zoo Species * - species is represented
ox4tates )J ieuican%usellm Groups of Lizards in the Genus Sceloporus Karyotypes of the Five Monotypic Species BY CHARLES J. COLE
J ieuican%usellm Groups of Lizards in the Genus Sceloporus Karyotypes of the Five Monotypic Species BY CHARLES J. COLE") )J ieuican%usellm ox4tates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N. Y. I0024 NUMBER 2450 FEBRUARY II, I971 Karyotypes of the Five Monotypic Species
)J ieuican%usellm ox4tates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N. Y. I0024 NUMBER 2450 FEBRUARY II, I971 Karyotypes of the Five Monotypic Species
Rapid Growth Phase of Ovum in the Guinea Fowl
 300 Rapid Growth Phase of Ovum in the Guinea Fowl Hiroshi OGAWA1), Takehito KUWAYAMA2) and Katuhide TANAKA2) Fuji Zootechnical Station, Tokyo University of Agriculture, Fujinomiya-shi 418-01 Department
300 Rapid Growth Phase of Ovum in the Guinea Fowl Hiroshi OGAWA1), Takehito KUWAYAMA2) and Katuhide TANAKA2) Fuji Zootechnical Station, Tokyo University of Agriculture, Fujinomiya-shi 418-01 Department
d. Wrist bones. Pacific salmon life cycle. Atlantic salmon (different genus) can spawn more than once.
 can spawn more than once.") Lecture III.5b Answers to HW 1. (2 pts). Tiktaalik bridges the gap between fish and tetrapods by virtue of possessing which of the following? a. Humerus. b. Radius. c. Ulna. d. Wrist bones. 2. (2 pts)
Lecture III.5b Answers to HW 1. (2 pts). Tiktaalik bridges the gap between fish and tetrapods by virtue of possessing which of the following? a. Humerus. b. Radius. c. Ulna. d. Wrist bones. 2. (2 pts)
Animal Diversity wrap-up Lecture 9 Winter 2014
 Animal Diversity wrap-up Lecture 9 Winter 2014 1 Animal phylogeny based on morphology & development Fig. 32.10 2 Animal phylogeny based on molecular data Fig. 32.11 New Clades 3 Lophotrochozoa Lophophore:
Animal Diversity wrap-up Lecture 9 Winter 2014 1 Animal phylogeny based on morphology & development Fig. 32.10 2 Animal phylogeny based on molecular data Fig. 32.11 New Clades 3 Lophotrochozoa Lophophore:
THE CORPORATION OF THE CITY OF BRAMPTON BY-LAW. Number '_6_5_-9_2. To prohibit and regulate the keeping of animals other than dogs
 THE CORPORATION OF THE CITY OF BRAMPTON BY-LAW Number '_6_5_-9_2 To prohibit and regulate the keeping of animals other than dogs The Council of The Corporation of the city of Brampton ENACTS AS FOLLOWS:
THE CORPORATION OF THE CITY OF BRAMPTON BY-LAW Number '_6_5_-9_2 To prohibit and regulate the keeping of animals other than dogs The Council of The Corporation of the city of Brampton ENACTS AS FOLLOWS:
Lineage Classification of Canine Title Disorders Using Mitochondrial DNA 宮原, 和郎, 鈴木, 三義. Journal of Veterinary Medical Sci Citation
 ' ' Lineage Classification of Canine Title Disorders Using Mitochondrial DNA TAKAHASI, Shoko, MIYAHARA, Kazuro Author(s) Hirosi, ISHIGURO, Naotaka, SUZUKI 宮原, 和郎, 鈴木, 三義 Journal of Veterinary Medical Sci
' ' Lineage Classification of Canine Title Disorders Using Mitochondrial DNA TAKAHASI, Shoko, MIYAHARA, Kazuro Author(s) Hirosi, ISHIGURO, Naotaka, SUZUKI 宮原, 和郎, 鈴木, 三義 Journal of Veterinary Medical Sci
Three tiers of genome evolution in reptiles
 494 Three tiers of genome evolution in reptiles Chris L. Organ, 1 Ricardo Godínez Moreno and Scott V. Edwards Department of Organismic and Evolutionary Biology, Museum of Comparative Zoology, Harvard University,
494 Three tiers of genome evolution in reptiles Chris L. Organ, 1 Ricardo Godínez Moreno and Scott V. Edwards Department of Organismic and Evolutionary Biology, Museum of Comparative Zoology, Harvard University,