Shelter availability, stress level, and digestive
|
|
|
- Cornelius Moody
- 5 years ago
- Views:
Transcription
1 First posted online on 15 November 2012 as /jeb J Exp Biol Advance Access Online the most Articles. recent version First at posted online on 15 November 2012 as doi: /jeb Access the most recent version at Shelter availability, stress level, and digestive performance in the aspic viper 4 5 Xavier Bonnet, Alain Fizesan, Catherine Louise Michel CEBC-CNRS, Villers en Bois, France Correspondance : Xavier Bonnet ; Tel ; Fax ; bonnet@cebc.cnrs.fr Submitted for consideration in Journal of Experimental Biology Keywords: corticosterone, digestive performance, ectothermy, heliotheria, reptiles, safety, shelter, snake, stress, thermoregulation Short title: Impact of lack of shelter on snakes 1 Copyright (C) Published by The Company of Biologists Ltd
2 Abstract The lack of shelter can perturb behaviors, increase stress level, and thus alter physiological performances (e.g. digestive, immune, or reproductive functions). Although intuitive, such potential impacts of lack of shelter remain poorly documented. We manipulated shelter availability, environmental and physiological variables (i.e. access to a heat source, predator attack, feeding status) in a viviparous snake. We assessed sun-basking behavior, digestive performance (i.e., digestive transit time, crude estimate of assimilation, regurgitation rate) and plasma corticosterone levels (a proxy of stress level). Shelter deprivation provoked a strong increase in sun-basking behavior and thus elevated body temperature; even in unfed individuals for which energy savings would have been otherwise beneficial. The lack of heat was detrimental digestive performance (i.e. all the metrics used to assess it). Simulated predator attacks worsened the situation and entailed a further deterioration of digestion. The combination of the lack of shelter with cool ambient temperatures markedly elevated basal corticosterone level and was associated with low digestive performance. This hormonal effect was absent when only one negative factor was involved, suggesting a threshold response. Overall, our results revealed important non-linear cascading impacts of shelter availability on stress-hormone levels, behaviors and physiological performance. These results infer shelter availability is important for laboratory studies, captive husbandry, and possibly conservation plans. 2
3 Introduction In most animals, individuals spend a considerable amount of time in refuges, and shelter availability is often essential for the persistence of populations (Wright and Shapiro 1990, Armstrong and Griffiths 2001, Souter et al. 2004, Bonnet et al. 2009, Grillet et al. 2010, Lagarde et al. 2012). Shelters provide protection against predators and buffer environmental conditions (Anderson 1986, Schwarzkopf and Alford 1996, Sih 1997; Roper et al. 2001, Berryman and Hawkins 2006). The benefits of retreat site selection have been examined in species belonging to various lineages (arachnids, fish, reptiles, mammals or amphibians for instance; Bulova 2002, Kearney 2002, Goldsbrough et al. 2004, Kotler et al. 2004, Millidine et al. 2006, Hossie and Murray 2010, 2011). However, previous investigations focused on constraining temperature conditions: hibernation, aestivation or sun-scorching periods (Seebacher and Alford 2002, Beck and Jennings 2003, Lagarde et al. 2002, 2012, Cooper and Withers 2005). Yet, many animals intensively use their refuges for other reasons than to escape severe climatic conditions: for instance to sleep, for reproduction or digestion (Siegel, 2005, Pike et al. 2010, 2011). Consequently, important behavioral and physiological consequences unrelated to overheating or dehydration should be associated with shelter availability; yet, this eco-physiological issue is poorly documented When they leave their refuge, animals are exposed to both challenging abiotic and biotic factors (Sih 1997, Kearney 2002, Langkilde et al. 2003, Berryman and Hawkins 2006). Assessing the consequences of the absence of shelter on stress level, behaviors and physiological performances is thus crucial to understand how behavioral tradeoffs are mediated. Notably, many animals retreat into their shelter between foraging episodes. Such shuttling activity offers an appropriate context to test behavioral 3
4 trade-offs. For example, individuals deprived from a shelter may be forced to undertake displacements to escape unfavorable and/or dangerous conditions with possible negative consequences on other activities. A lack of shelter may also generate physiological stress, which in turn may perturb immunity, sexual behaviors, and possibly many other fitness related processes (Rabin 1999). To examine these issues, we manipulated shelter availability, ambient temperature, feeding status, and predatory threat in a snake species. We assessed the consequences on three major traits: thermoregulation, digestive performance, and plasma corticoid levels (Möstl and Palme 2002, Bassett and Buchanan-Smith 2007). We measured corticoid hormones as they influence the process of acquisition and allocation of resources, the mobilization of body reserves, metabolism, and various behaviors (DeNardo and Sinervo 1994, Guillette et al. 1995, Romero 2004). We expected that snakes would display contrasting behavioral and physiological responses to different environmental conditions. We notably hypothesized that unfavorable conditions (e.g. lack of shelter, low ambient temperature) would provoke an elevation of plasma corticoid levels and would impede digestive performances. By contrast, favorable conditions (presence of a shelter plus high ambient temperature) should be associated with low plasma corticoid level and high digestive performance. Finally, intermediate situations (combination of favorable and unfavorable factors) should produce intermediate stress level and average digestive performance. Overall, these predictions can be expressed through one main hypothesis: environmental conditions should exert an additive effect (either positive or negative) on physiological parameters associated 4
5 with stress and resource assimilation. Further, we expected state dependent effects where individual physiological status influences response (Aubret and Bonnet 2005). Notably, recently fed snakes should exhibit different behaviors compared to unfed snakes (second hypothesis). Indeed, as digestion requires elevated body temperatures, fed snakes should exhibit a positive thermoregulation shift to select hot microhabitat more often than unfed snakes. Snakes are suitable organisms to test these hypotheses. Most species are secretive and individuals spend considerable amounts of time sheltered (Bonnet and Brischoux 2008). They usually leave their refuge to feed on large prey items, and they tend to cease moving during long digestive episodes. Therefore, foraging and digestion can be easily distinguished and monitored (by comparison these processes overlap in herbivorous animals, rendering analyses complicated). Digestion speed depends on body temperature and is often associated with prolonged phases of sun basking (at least in temperate climates) that increases predation risk (Garland and Arnold 1983, Beck 1996); consequently, there is a conflict between rapid digestion associated to sun exposure versus survival provided by the refuge (Sih 1997). The experiments presented in this study are largely based on this trade-off Material and methods Ethics Statement The authors attest their adherence to NIH standards. No snake was mistreated and the mice were euthanized following rapid cervical dislocation. Permit to collect the snakes were issued by the DIREN Poitou-Charentes (# 09/346/DEROG). All the experiments were performed under permit A and
6 Study species Previous studies provided important background (Saint Girons 1957, Bonnet et al. 1999, 2000, 2001, 2002; Bonnet 2011; Ladyman et al. 2003; Zuffi et al. 2009; Michel and Bonnet 2010). Aspic vipers feed mainly on rodents and digestion is impeded when body temperature falls below 15 C (Naulleau 1983). In the field, animals are active under ambient temperatures ranging from 10 C to more than 30 C (depending upon cloudy layer). Vipers shuttle between periods of heliotheria (i.e. sun-basking behavior to warm up the body, Bailly 1894) and periods sheltered in their refuge. This shuttling activity is a central element of their thermoregulation. The snakes involved in the experiment were captured in the vicinity of the laboratory (between spring 2003 and autumn 2008; forest of Chizé, N, W). Prior to the experiment the individuals were maintained in captivity (during at least 3 months) in standard boxes (42cm*34cm*19cm; L*l*h), with artificial grass as substratum, clean water ad libitum, a thermal gradient (20-35 C), and a rooftile as a refuge. All the snakes accepted their food regularly (dead mice approximately every two weeks). The experiments were performed during the normal activity period of the species (May August) Experimental design We performed two main experiments, and to obtain comparable data between them we used a single modular system (Figure 1). We built ten three-compartment cages fitted with artificial grass substratum (total length 150cm, 50x30x20cm per compartment) and with a small door ( 5cm) between adjacent compartments. When 6
7 the doors were open, the snakes could freely move across the compartments (experiment 1). When the doors were shut, the snakes were restricted to one compartment (experiment 2). In one compartment (A, Figure 1) we placed a 50-W halogen lamp to create a heat source (i.e. mimicking solar radiation). Roof-tiles were used as shelters. The cages were placed in a temperature-controlled room set at 16 C (14 hours of daylight: 9:00AM-21:00PM). Ambient temperatures were measured with data loggers (ibuttons Thermochron ). The temperature of the shelter and the surface body temperature of the snakes were measured from distance (20-30cm) using a 3 dots infrared laser thermometer (Raytek MX2, Fotronic Corporation; measurement diameter 19mm, range -30 C-900 C, precision ±1 C, 250ms/reading; see supplementary material). Surface and internal body temperature are highly correlated in reptiles (Lagarde et al. 2012). The front of the cages was transparent; hence, the observer and the snakes could see each other. a) Experiment 1 We first assessed the behavioral trade-off between safety and sun basking. More precisely, we focused on the impact of shelter availability and feeding status on heliotheria. Each snake was placed in a three-compartment cage, and thus in a 16 C C thermal gradient during the day (Figure 1-A). Two compartments, A and C, were fitted or not with a shelter, and we obtained four combinations: No shelter available: the snake was always visible and could select either the hot compartment (A) versus one of the two cold compartments (B or C, figure 1). 7
8 One shelter was placed in the cold compartment (C) at the opposite of the hot spot (A). The snake could select the hot-unsheltered (A), the cold-unsheltered (B), or the cold-sheltered compartment (C). Two shelters were provided: one in the hot compartment and one in the opposite cold compartment. The snake could select the hot-sheltered, the coldsheltered, or the cold-unsheltered compartment. The medium compartment (B) was never provided with a shelter to better assess selection; indeed the snakes were forced to cross it to find a heat source and/or a shelter in the compartments A and/or C. To obtain comparative data, we also tested other situations. Notably, the snakes (fed or unfed) were given the choice between hot-sheltered versus cold-unsheltered compartments, or sheltered versus unsheltered compartments without any heat source. However, these situations do not entail any major conflict (hence, they were partly out of scope of the aim of the current study). Indeed, terrestrial snakes prefer to remain sheltered rather than to stay in the open and they prefer a warm place rather than a cold one. For simplicity, these specific results, although useful to test our protocol will be briefly presented and referred as checking tests We used 32 snakes in this first experiment, balancing sex and age ratios, and encompassing a wide range of body sizes (Table 1). At least two weeks after their last meal, unfed snakes were randomly allocated to one of the four situations described above (following a random order). The snake was gently placed in the medium compartment of the cage one night before the beginning of records. It was observed every hour between 09h05 to 20h05 during two subsequent days (36h). Its position 8
9 and attitude (e.g. coiled, stretched, moving, motionless) were noted (N=12 observations per day, total number of observations per snake: N=24). Then, we performed again the same experiment with the same recently fed snakes. We offered a mouse to each snake (mouse mass represented 25±2% of snake mass, all individuals accepted the meal except during skin shedding [a relatively rare event] hence slight variations in sample sizes) and we placed the viper into the cage immediately after the prey was swallowed Overall, most of the snakes were tested either fed or unfed, and for each digestive status they experienced four different situations (4 combinations of shelter(s)). We collected a total of 4,752 focal observations. In addition, we collected 816 focal observations for the checking tests on a random subsample of 25 snakes. b) Experiment 2 In the second experiment, we examined the consequences associated with the selection of one of the three compartments available to the snakes in the first experiment. Thus, we shifted from measuring heliotheria (i.e. sun basking behavior) and sheltering behaviors to digestive performance and stress levels. During two weeks, we imposed the four combinations of heat source and shelter availability to recently fed snakes: cold-unsheltered; cold-sheltered; hot-unsheltered; hot-sheltered. The doors of the boxes were thus shut (total 30 separate compartments, Figure 1). Half of the compartments were fitted with a shelter and half were not (following a random procedure), water was provided in each compartment. The heat source was available during the diurnal phase in the 10 hot 9
10 compartments, the 20 other compartments remained cold all the time (16 C). We placed one snake per compartment during 16 consecutive days and the compartments were inspected three times per day (9h00, 13h00, 17h00). The feces were rapidly collected, weighed and dried (see below). We first tested 88 recently fed snakes (32 vipers of the first experiment plus 56 vipers, Table 1), immediately after the prey was swallowed. The snakes were weighed twice: before feeding and 16 days later Second, we analyzed the impact of repeated simulated predator attacks on recently-fed snakes. We randomly selected and tested 56 vipers (among the 88 available). We followed the same procedure as above but every day we touched each snake with a leather glove (10 times, total duration 15 seconds, the glove was attached to a 50cm rod, standard movements, same observer), and this was repeated three times per day (9h00, 13h00, 17h00). The snakes subjected to the simulated attack displayed typical defensive behaviors (e.g. hissing, striking). As above, the snakes were monitored during 16 days. Third, we assayed plasma corticoid levels in unfed vipers (N=63 vipers large enough for blood sampling) that experienced the same treatment from above, but without predatory attack simulations. These vipers were then not involved into any experiment for at least two weeks before being tested. They were not fed and not attacked to limit potential confounding effects of digestion and predatory threat of basal plasma corticosterone level. Indeed, our objective was to assess the impact of shelter and heat source availability on endocrine stress levels. All individuals were blood sampled 10 days after the beginning of the test. This duration was selected to 10
11 limit the effect of initial handling stress (i.e. placing snake into the box) and to offer to the snakes enough time to adapt to experimental condition Digestive performance Experiment 2 provided sufficient amount of time (16 days) to monitor digestion: the production of feces requires one week on average (Michel and Bonnet 2010). We considered the date for the first feces as an index of transit time; we also counted the other feces and noted their date of production. We used a crude estimate of assimilation rate: the proportion of ingested prey in relation to snake body mass variation. For example, a 20g mouse inducing a 4g increase in snake mass after digestion provides a value of 20% (see Michel and Bonnet 2010 for details). We emphasize that such calculation does not provide accurate values of digestion efficiency (e.g. other factors such as metabolism, energy content of feces, or specific body composition are involved), but is a useful index for inter-individual or inter-group comparisons. Several individuals regurgitated their prey. Snakes tend to regurgitate when threatened or exposed to unfavorable ambient temperatures (Greene 1988; Naulleau 1983). The date of regurgitation was noted, the vomited items were examined, weighed (fresh and dry mass). In case of regurgitation, to estimate the proportion of the prey digested (not assimilated) by the snake we divided the dry mass of the regurgitated prey by the estimated dry mass of the corresponding ingested mouse. This index reflects the proportion of the prey wasted versus digested (we note that actual calculation of digestive efficiency requires passage through the gut). The dry masses of vomited prey were obtained after two weeks in a 60 C autoclave. The 11
12 estimated dry mass of the ingested prey was derived from the linear correlation between fresh and dry masses measured in a range of euthanized mice (body mass ranged from 2.8g to 55.0g; N=7; r²=0.99, P<0.01; estimated dry mass = (fresh mass) ). In several cases, snakes that regurgitated did not produce any feces; generating some variations in the sample size. Importantly, the strong variations in crude estimate of digestive performance caused by different timing of regurgitation (see below) were more critical compared to any other sources of variations (e.g presence of a heat source, Michel and Bonnet 2010); therefore, we essentially focused on this key factor. Hormonal assays Basal corticosterone levels were obtained from blood samples collected within less than 5 min after handling. We obtained µl of blood per snake via intracardiac punctures (<5% of total body mass, using 30G-needles). This technique is routinely employed in snakes (Bonnet et al. 1994, 2001; Fauvel et al. 2012). The blood was immediately centrifuged (3 min*10,000g), the plasma separated and stored at -25 C until assay. Hormonal assays were performed at the Centre d Etude Biologique de Chizé Plasma concentrations of corticosterone (Mathies et al. 2001; Moore et al. 2000; Palacios et al. 2011) were determined using radioimmunoassay (RIA). The mean extraction rate of the steroids from the plasma was of 97.3±5.2%, and was always greater than 89% (N=28 tests); intra- and inter-assays coefficients of variation remained lower than 4%; the sensitivity of the assay was of 1.9 pg/tube. Crossreactions with other steroids (using the same protocol) were as follow: 12
13 androstenedione (<0.1%), compound S (7%), cortisol (0.1%), 11-deoxy-corticosterone (0.1%), progesterone (7%), testosterone (<0.1%). Plasma corticosterone levels were not correlated with snake body mass (r²=-0.005, P=0.424) Analyses During checking tests and first experiment, the vipers were observed discontinuously. In a preliminary study, we tested (and validated) this approach through continuous monitoring of twenty snakes during 12h: we placed each snake in the central compartment of the box (B, Figure 1), the doors were open, and we noted all the subsequent movements. We found no difference between continuous observations versus a single observation per hour. Indeed, the snakes crawled slowly to select a compartment, and then remained at the same place for prolonged periods. Thus the one-hour sampling over two days (N=24 records per individual) captured all the displacements between the compartments. For unknown reasons, the snakes almost never stayed in the middle compartment B: the mean score was negligible (<1 compared to a possible maximal value of 24). This means that the vipers were essentially observed either in the compartment A or in compartment B. Consequently, we simply considered the number of observations in the compartment A (i.e. heat source turned on in this compartment, except during several checking tests); and the resulting score (0-24) was expressed as the heliotheria score (i.e. sun basking score). Different observers were involved, but we did not detect any observer effect. Indeed the snakes remained motionless most of the time during observations. 13
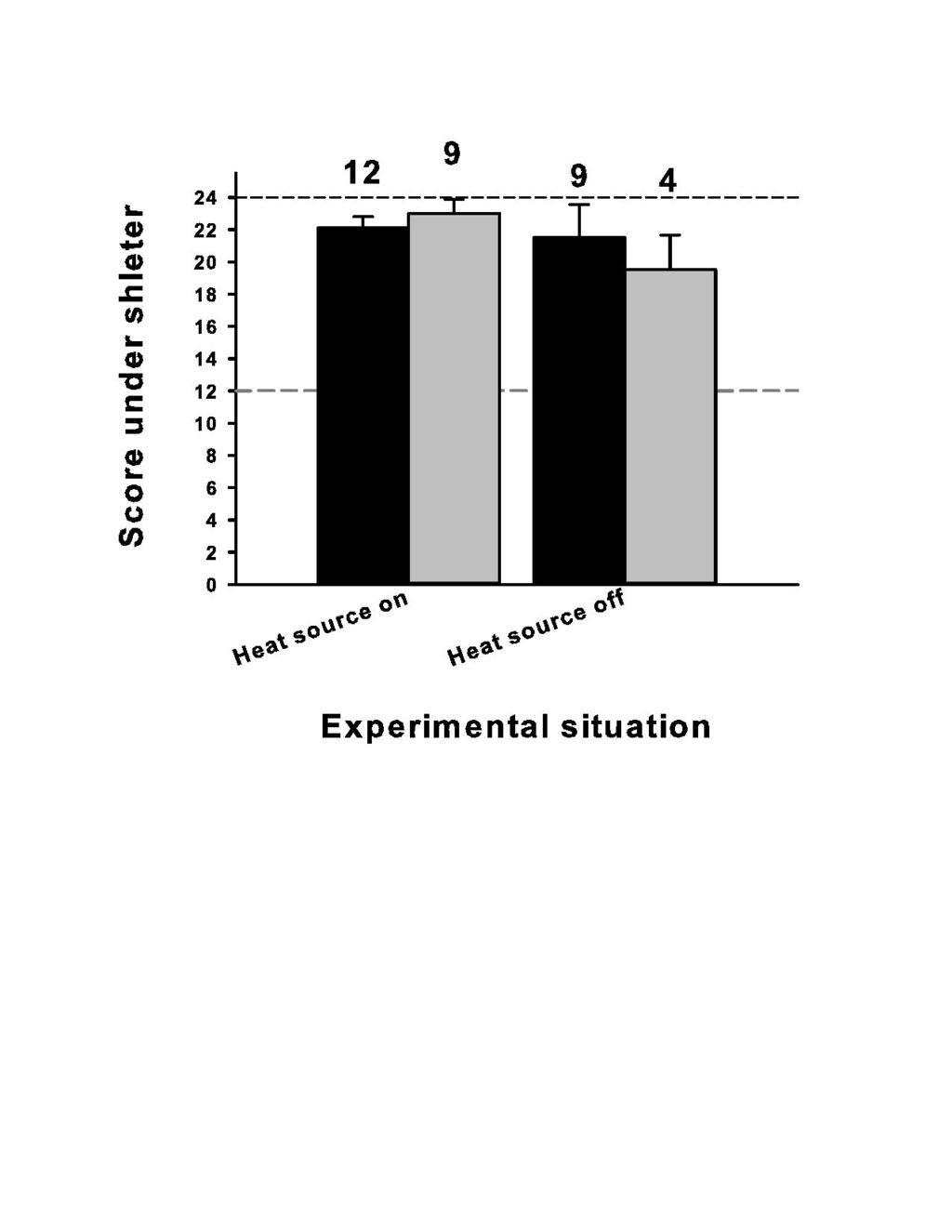
14 In all experiments, the mean characteristics of the snakes (body size, body mass, body condition, and age class) were not different between the different experimental situations (or compartments) (P>0.40 in all comparisons). Most vipers were tested repeatedly; however, at least one week elapsed between subsequent experiments, and none of the snakes presented any sign of disorder. Our experiments were not (or weakly with blood sampling) invasive and the snakes were maintained in appropriate condition. Thus, the impact of one experiment on the next remained undetectable. For conciseness, we did not present all the results (e.g. all sex effects were non-significant). This study did not aim to investigate the thermal biology of the aspic viper; thus, we simply report broad relevant descriptive thermal data. We used non-parametric (e.g. Friedman ANOVA for repeated measures on the same individual) to assess behavioral scores and contingency tables for comparisons of the occurrence of certain events among groups (e.g. regurgitation). We used parametric tests (e.g. F-test ANOVA) to compare continuous variables; the homogeneity of variance assumption was not violated. Pseudo-replicates were discarded from calculations. Statistics were performed with Stastistica 7.1 ( StatSoft Inc ). Results a) Experiment 1: microhabitat selection The checking tests showed that the snakes markedly preferred the compartment fitted with a shelter (heat source either turned on or off), all tests provided a mean value greater than 19.5 (mean = 21.9±3.9, ±SD as in subsequent results) with no difference between experimental situations (χ²=0.00, df=3, P=1.00; Figure 2). These 14
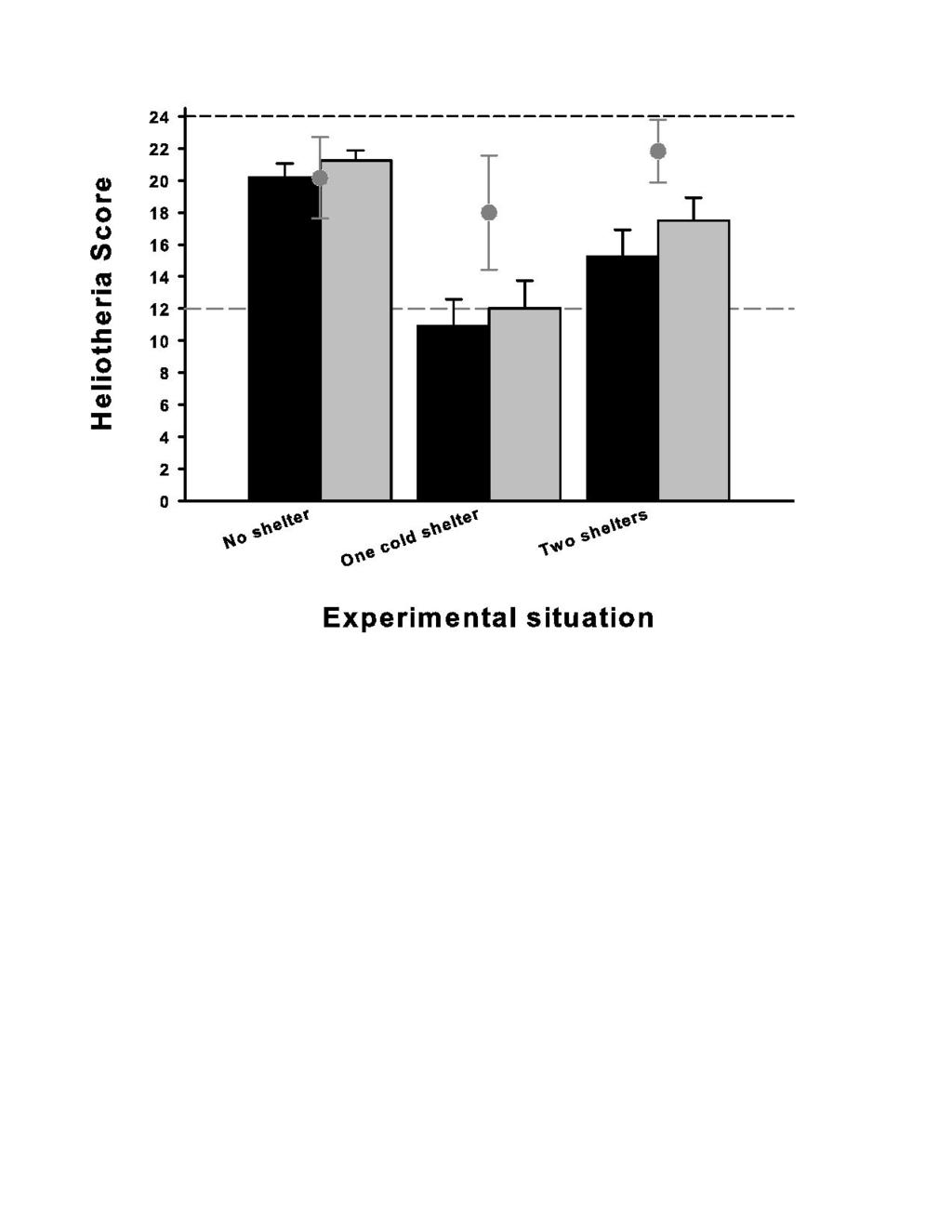
15 expected (confirmatory) results enabled us to examine the behavior of the snakes placed under more challenging situations. When no shelter was available, unfed snakes selected the hot compartment-a (i.e. heat source turned on) most of the time (Figure 3). The mean heliotheria score (20.2±3.8) was close to the maximal theoretical value (24). The addition of a shelter in the cold compartment (C) attracted an important proportion of individuals under this refuge. When two shelters were available, we observed an intermediate situation: many snakes shifted to hot with a shelter compartment-a; the others selected the cold with a shelter compartment-c. These differences were significant (Friedman ANOVA with different individual scores as the repeated dependent variables, N = 28, df = 2, χ² = 14.04, P<0.001). Snakes with recently ingested prey exhibited a similar pattern of microhabitat selection (Figure 3). Although not significant, this later result was nonetheless very close to conventional significance (same design Friedman ANOVA, N = 28, df = 2, χ² = 5.82, P=0.054). Recalculating a single score per snake irrespective of the feeding status (averaging fed and unfed scores displayed in Figure 3) supported the above patterns observed in unfed and fed snakes respectively (Friedman ANOVA, N = 32, dl = 2, χ² = 25.07, P<0.001) Further comparisons, for example between unfed versus fed vipers in one of the three experimental situations failed to detect any significant effect (P>0.30 in all Wilcoxon-tests for paired samples) suggesting that unfed and fed snakes exhibited a similar pattern of microhabitat selection (Figure 3). b) Experiment 2: shelter, heat, predator, and physiological performances 15
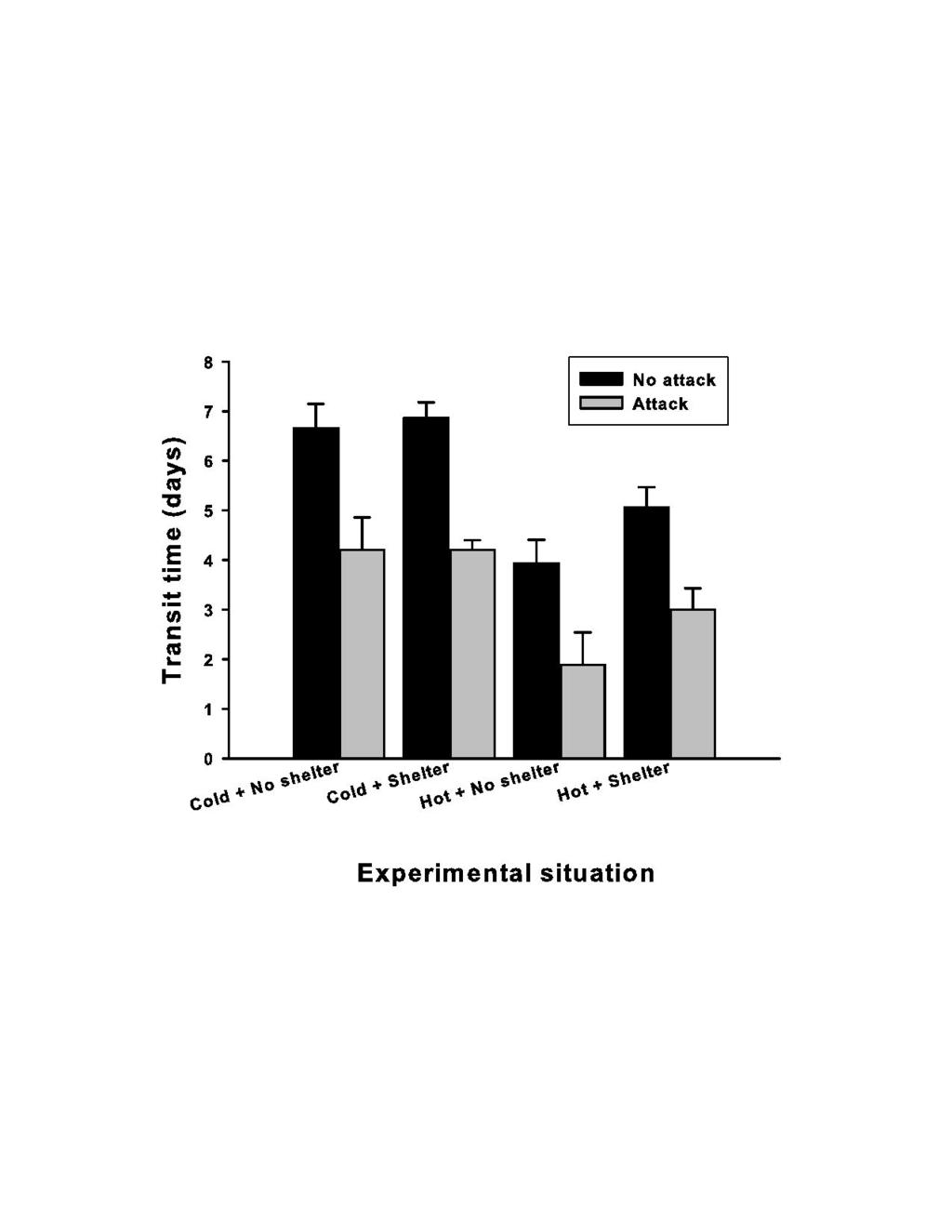
16 Ambient and body temperatures Mean ambient temperature of the cold compartments was 15.5±0.6 C (range C, N=3,450 readings) without significant variations over time; the mean body temperature of the snakes in the cold compartments was similar (15.6±0.3 C, N=212). By contrast, the ambient temperature in the hot compartments varied markedly: similar on average to the values recorded in the cold compartments during the night (15.7±0.8 C, C, N=360), but ranging over a wide gradient when the halogen lamp was turned on. The temperature measured under the tile when the heat source was turned on averaged 33.1±16.4 C ( C, N=146, logger positioned on the grass substrate in the middle of the shelter). The mean ambient temperature measured in the corner of the compartment provided with the heat source, but at the opposite of the lamp was of 17±1.8 C ( C, N=140). The surface body temperature of the snakes placed in the hot compartment varied between 14 C to 35 C and averaged 28.3±6.2 C when the thermal gradient was available (N=46 readings, restricting records from 10h00 to 18h00 to ensure that the snake had enough time for thermoregulation). Digestive performance The presence of a heat source shortened digestive transit time, but the presence of the shelter had no effect on this trait (Figure 4; ANOVA with transit time as the dependent variable, heat source and presence of a shelter as the factors; heat source, F 1, 84 =22.07, P<0.001; shelter, F 1, 84 =1.94, P=0.167; interaction, F 1, 84 =0.93, P=0.338). We found similar pattern in snakes subjected to the predator attacks with a significant effect of heat source only (Figure 4; heat source, F1, 53=5.21, P=0.027; shelter, F1, 16
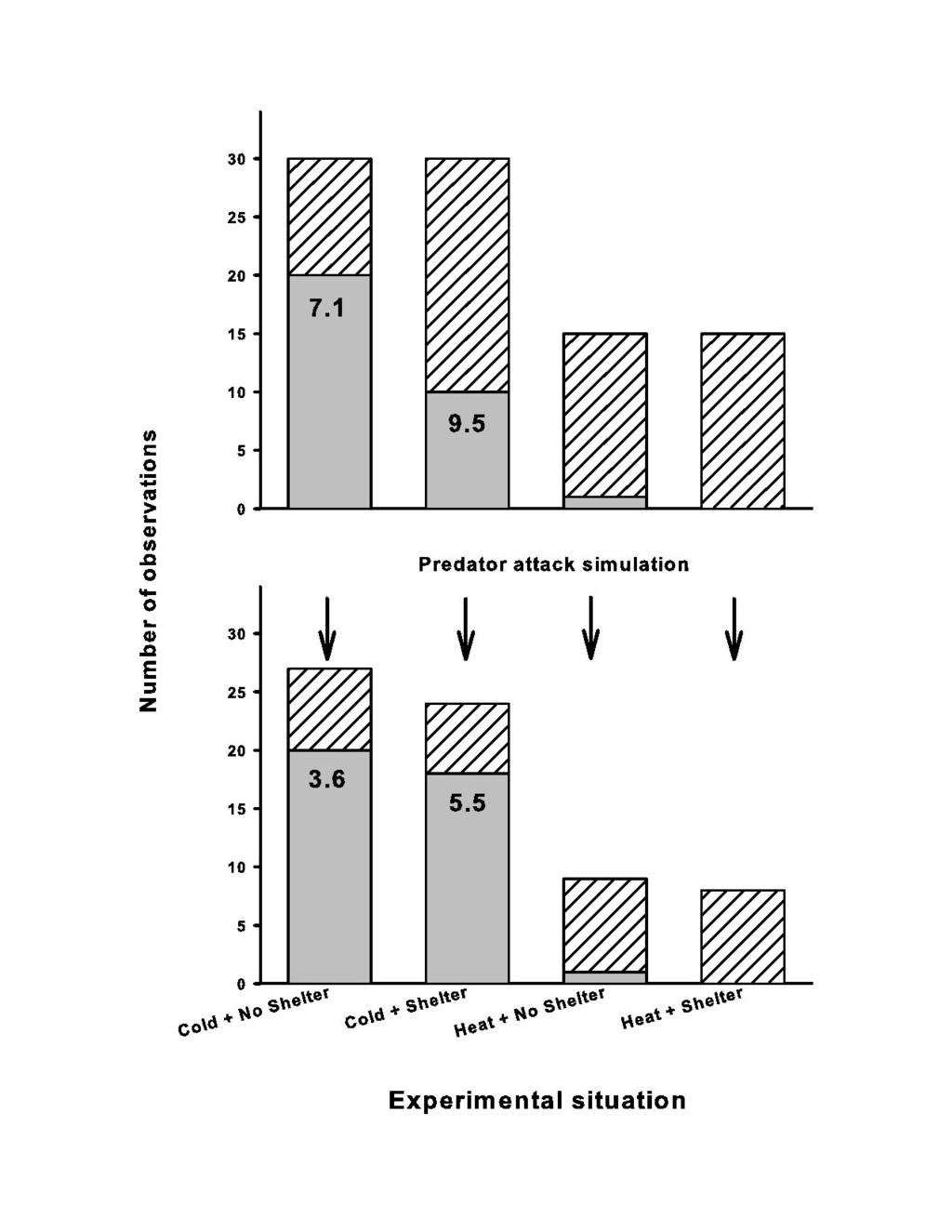
17 =1.59, P=0.213; interaction, F1, 53=0.20, P=0.660). However, the predator attacks strongly reduced transit time in all cases (Figure 4: Wilcoxon test for dependent variables, Z=4.40, P<0.001, N = 56). We found no significant effect of the experimental situation on the total number of feces (mean 2.3±1.6, range=1-7). The longer the prey remained in the stomach before regurgitation greater was the proportion of the prey digested (Figure 5). Regurgitation date had a strong effect on the total number of feces produced (r=0.50, F 1, 57 =19.43, P<0.001). The crude assimilation rate was different between the vipers that regurgitated (-8%, N = 51) versus not (20%, N = 28) (F 1, 77 = 17.36, P<0.001). Regurgitation events were not randomly distributed (Figure 6a; comparing the observed against a theoretical unbiased distribution; df = 3, χ² = 18.20, P<0.001). Regurgitation was rarely recorded (N=1) when a heat source was provided, but was frequent in the cold conditions (N=30; df = 1, Yates χ² = 17.28, P<0.001), and was twofold more frequent when the snakes were subjected to the both absence of shelter and heat source (10 versus 20; df = 1, Yates χ² = 5.4, P = 0.020). Predator attacks amplified these effects, regurgitation rate increasing from 50% to 75% (Figure 6b; df = 3, χ² = 11.37, P<0.01); again, such effect essentially affected the cold snakes with and 1 regurgitations in the cold versus hot conditions (df = 1, Yates χ² = 21.82, P<0.001). Importantly, the difference between the snakes deprived from shelter versus those provided with a shelter was cancelled out when the snakes were harassed: they all lost their prey at the same high rate (Figure 6b; df = 1, Yates χ² = 0.06, P = 0.805). Analyses of regurgitation dates supported these trends (Figure 6a, b). The snakes under the most unfavorable conditions tended to regurgitate sooner (the 17
18 mean regurgitation date was 7.7±3.8 days in non-attacked [N=30] versus 4.6±3.1 days in attacked snakes [N=38] respectively). Eleven snakes that regurgitated on two experimental situations enabled us to further test this effect on the same individuals, the predator attack simulation precipitated regurgitation (Wilcoxon-tests for paired samples Z = 2.80, N= 11, P = 0.005) Plasma corticosterone levels Vipers provided with heat source and/or shelter exhibited low basal plasma corticosterone levels (Figure 8). However, the combined absence of shelter and of heat source was associated with high plasma corticosterone levels, suggesting a chronic stress level (Figure 8; ANOVA with plasma corticosterone concentration as the dependent variable and experimental condition as the factor; interaction, F1, 59=5.15, P = 0.027). Discussion Our results show that the absence of shelter generates marked behavioral and physiological responses that appear to signal an elevation in organismal stress state of snakes. These responses are expected to have important fitness consequences. For instance, the lack of shelter provoked a strong reduction in energy intake (high regurgitation rate), notably when no heat source was available (an additional factor), or an important waste of energy (metabolism elevation due to the high body temperature in unfed individuals) when a heat source was provided. Yet, significant impairment of energy intake should be regarded as particularly detrimental to reproduction in snakes (Bonnet et al. 2002). These results support our main 18
19 hypothesis suggesting that environmental conditions should exert an additive effect on physiological parameters associated with stress and resource assimilation. We acknowledge that this conclusion was derived from different experiments rather than from a direct measure of physiological stress per se on food intake. However, the same individuals underwent different experiments, and thus provided a strong support for the suggested relationship. In vipers deprived from heat source, regurgitation rate was more pronounced when no refuge was available (Experiment 2). In many species (e.g. humans, birds) individuals vomit under stressful conditions or when threatened (Camilleri et al. 1986; Peterson et al. 1987; Siegel-Causey and Hunt 1981; Vogel 1950). Stress induced vomiting can be beneficial because it may deter predator, relaxes physical burden and increase escape capacities. However, the negative value of crude assimilation rate recorded in the snakes that regurgitated (-8%) was greater than expected as unfed (i.e. starving) snakes would display only -0.5% of body mass loss under similar condition (Michel and Bonnet 2010). This revealed a new stress induced cost, possibly due to the loss of body fluids and/or to the energy demand to initiate digestion (McCue 2006). Low ambient temperature represented a negative factor per se (Regal 1966), but it was insufficient in isolation to trigger a negative effect equivalent to the lack of shelter and heat in combination. Indeed, most cold snakes with a shelter (67%, Figure 6) successfully digested their prey and eventually gained mass. In support of this, we found a strong correlation between regurgitation date and digestion rate. We also observed a normal crude assimilation rate in the cold snakes that did not regurgitate (18% compared to 17% in Michel and Bonnet 2010). In 19
20 other words, most cold but sheltered snakes digested normally, albeit slowly. This enabled us to discard a possible regurgitation-reflex due to putrefaction of the prey and suggests that stress was a key factor. The investigations based on corticosterone provided complementary results. The most stressful condition, lack of shelter and heat source, generated chronic elevated corticosterone levels; halfway between mean normal basal and maximal levels observed during acute stress response (Figure 7). However, our results do not entirely match one of our initial predictions. We expected gradual hormonal effects in response to the accumulation of stressor agents. We rather obtained a threshold effect with a sudden massive corticosterone increase when two adverse conditions were combined (note that this effect can nonetheless be considered as additive). Perhaps the combination of cold ambient temperature with the absence of shelter exceeded the snake capacity to cope appropriately with stressful conditions resulting into a chronic stress (Bradshaw 1986; Romero 2004). Predatory threat (simulated attacks) further lowered digestive performance, especially under cool condition: snakes regurgitated at a very high rate (>60%) without difference between individuals deprived from shelter or not (Experiment 2). In addition, this factor accelerated defecation, another typical defensive behavior (Haas 1985; Myles 2002; Weiss 2006; Tillmann 2009). These results reinforce the notion that stress induced strong negative physiological consequences. At first glance, our results may suggest that shelterless snakes provided with a heat source (even individuals subjected to simulated predator attacks) were spared from stress costs. Indeed, they exhibited very low regurgitation rate and low plasma 20
21 corticosterone levels. However, appearances are deceptive. Indeed, in the Experiment-1 (snakes move freely across compartments), the absence of shelter forced almost all snakes to increase their body temperature (hence energy expenditure) through extensive heliotheria (Figure 3). This effect was marked in unfed individuals in whom a selection for low temperature to save energy was rather expected. Our second hypothesis snakes should exhibit different behaviors compared to unfed snakes was thus challenged. As predicted, the snakes with a full stomach exploited the opportunity to shorten digestion via an elevation of their body temperature (Michel and Bonnet 2010). This pattern conforms to field observations. In natural conditions, recently fed snakes tend to bask intensively under cool ambient temperatures (Beck 1996; Regal 1966). However, unfed (or nonreproductive) vipers generally remain hidden in cool places to minimize both metabolic expenditure and predation risks (Bonnet and Naulleau 1996; Bonnet 2011). Consequently, in the current study, the fact that most snakes with an empty stomach decided to remain under the heat source most of the time was surprising. At least, we expected that the fed snakes would have exhibited higher heliotheria score compared to unfed snakes, but this was not the case. The immediate consequence of such marked apparently useless heliotheria was an obligatory waste of energy as the metabolic difference between divergent body temperatures (i.e. 28 C versus 16 C) is massive in ectotherms (Andrews and Pough 1985; Ladyman et al. 2003). Why did the unfed snakes not select cool temperature to save energy? The manipulation of shelter availability suggests that such pronounced heliotheria corresponded to an anti-predator tactic. In the absence of shelter, the 21
22 snakes situated in a cold compartment exhibited low body temperature (i.e. 16 C ambient or surface body temperature); they were slow and vulnerable (Heckrotte 1967). By maintaining high body temperature they considerably improved their defensive potential as fleeing ability, striking speed, or bite efficiency are temperature dependent (Goode and Duvall 1989; Greenwald 1974; but see Whitaker et al. 2000). When a shelter was provided (either in cold or hot compartment), an important proportion of snakes was attracted, whatever the ambient temperature and the feeding status, suggesting that the snakes essentially based their decision on two conflicting options: maintaining a costly high defensive potential (high body temperature) versus using the cold refuge associated with major energy saving. The main outcome of our study is to emphasize that a crucial characteristic of the habitat, the availability of a shelter, can exert a strong influence on stress status with cascading consequences on major behaviors and energy balance. A recent experiment on fish showed that the absence of refuge not only increased the unsafe perception caused by predation threat, but also the stress level (indicated by darker skin coloration), metabolic costs, and consequently total energy budget (Millidine et al. 2006). Thus, two convergent experimental studies carried out on very different organisms (fish and snake) documented a negative impact due to the absence of shelter associated to high stress level. The widely acknowledged climatic buffering role of the refuge cannot explain our results. Indeed, in the two studies that specifically investigated this issue (performed in both aquatic and terrestrial environments), shelter availability was experimentally manipulated under controlled ambient conditions (Millidine et al. 2006; this study). In both cases, the absence of 22
23 shelter stressed the individuals independently from other environmental factors. These results suggest that the safety feeling associated with the characteristics of the habitat is a key element to understand habitat selection, thermoregulation and energy budget. If our results are confirmed by further investigations, many laboratory studies should be re-examined. Notably, the behavioral and thermoregulation results from the numerous experiments where the animals were placed into a thermal gradient without any refuge available may well be questionable. More generally, captivity and conservation studies should respectively integrate both shelter availability and quality as important elements for animal welfare and for field managements. Acknowledgments. We thank C. Thiburce for the construction of the boxes and animal care, JP Degoit, K. Ben Kaddour for valuable help during behavioral observations, C. Trouvé for assistance during hormonal assays, and the ANR-DIAME for funding. References Anderson PK (1986). Foraging range in mice and voles: the role of risk. Can J Zool 64: Andrews RM, Pough FH (1985) Metabolism of squamate reptiles: allometric and ecological relationships. Physiol Zool 58 :
24 Armstrong JD, Griffiths SW (2001) Density-dependent refuge use among overwintering wild Atlantic salmon juveniles. J Fish Biol 58: Aubret F, Bonnet X (2005) The influence of body reserves and eye opacity in tiger snakes. J Exp Z 303A: Bailly A (1894) Dictionnaire grec-français. Eds Librairie-Hachette, Paris, France. Pp Bassett L, Buchanan-Smith HM (2007) Effects of predictability on the welfare of captive animals. Appl Anim Behav Sci 102: Beck DD (1996) Effect of feeding on body temperatures of rattlesnakes: a field experiment. Physiol Zool 69: Beck DD, Jennings RD (2003) Habitat use by Gila monsters: the importance of shelters. Herpetol Monogr 17: Berryman AA, Hawkins BA (2006) The refuge as an integrating concept in ecology and evolution. Oikos 115: Blouin-Demers G, Weatherhead PJ (2001) An experimental test of the link between foraging, habitat selection and thermoregulation in black rat snakes (Elaphe obsolete obsoleta). J Anim Ecol 70: Bonnet X, Naulleau G, Mauget R (1994) The influence of body condition on 17-ß Estradiol levels in relation to vitellogenesis in female Vipera aspis (Reptilia viperidae). Gen Comp Endocrinol 93: Bonnet X, Naulleau G. (1996). Catchability in snakes: consequences on breeding frequency estimates. Can J Zool 74:
25 Bonnet X, Naulleau G, Shine R, Lourdais O (1999) What is the appropriate time scale for measuring costs of reproduction in a capital breeder such as the aspic viper? Evol Ecol 13 : Bonnet X, Naulleau G, Shine R, Lourdais O (2000) Reproductive versus ecological advantages to larger body size in female Vipera aspis. Oikos 89: Bonnet X, Naulleau G, Bradshaw D, Shine R (2001) Changes in plasma progesterone in relation to vitellogenesis and gestation in the viviparous snake, Vipera aspis Gen Comp Endocrinol 121: Bonnet X, Naulleau G, Shine R, Lourdais O (2002) Reproduction in a typical breeder: costs, currencies, and complications in the aspic viper. Ecology 83: Bonnet X, Brischoux F (2008) Thirsty sea snakes forsake their shelter during rainfall. Austral Ecol 33: Bonnet X, Brischoux F, Pearson D, Rivalan P (2009) Beach-rock as a keystone habitat for sea kraits. Environ Conserv 36: Bonnet X (2011) The evolution of semelparity. Reproductive Biology and Phylogeny of Snakes. Reproductive Biology and Phylogeny Series, Science Publishers, Inc. Pp Bradshaw SD (1986) Ecophysiology of Desert Reptiles. Academic Press, Sydney, Australia. Bulova SJ (2002) How temperature, humidity, and burrow selection affect evaporative water loss in desert tortoises. J Therm Biol 27: Camilleri M, Malagelada J-R, Kao PC, Zinsmeisterl AR (1986) Gastric and autonomic responses to stress in functional dyspepsia. Digest Dis Sci 31:
26 Cooper CE, Withers PC (2005) Physiological significance of the microclimate in night refuges of the numbat Myrmecobius fasciatus. Austral Mammal 27: DeNardo DF, Sinervo B (1994) Effects of corticosterone on activity and home-range size of free-ranging male lizards. Horm Behav 28: Fauvel T, Brischoux F, Briand MJ, Bonnet X. (2012). Do researchers impact their study populations? Assessing the effect of field procedures in a long term population monitoring of sea kraits. Amphibia Reptilia, in press Garland TJr, Arnold SJ (1983) Effects of a full stomach on locomotory performance of juvenile garter snakes (Thamnophis elegans). Copeia 1983: Goldsbrough CL, Hochuli DF, Shine R (2004) Thermal cues used for retreat-site selection by flat rock spiders confer fitness benefits. Ecology 85: Goode MJ, Duvall D (1989) Body temperature and defensive behaviour of freeranging prairie rattlesnakes, Crotalus viridis viridis. Anim Behav 38: Greene HW (1988) Antipredator mechanisms in reptiles. In: Biology of the reptilia, vol. 16 (Gans C, Huey RB, eds). New York: Alan R. Liss, Greenwald OE (1974) Thermal dependence of striking and prey capture by gopher snakes. Copeia 1974: Grillet P, Cheylan M, Thirion JM, Doré F, Bonnet X, Dauge C, Chollet S, Marchand MA (2010) Rabbit burrows or artificial refuges are a critical habitat component for the threatened lizard, Timon lepidus (Sauria, Lacertidae). Biodiv Conserv 19: Guillette Jr LJ, Cree A, Rooney AA (1995) Biology of stress: interactions with reproduction, immunology and intermediary metabolism, in: Warwick C, Frye 26
27 FL, Murphy JB (Eds.), Health and Welfare of Captive Reptiles, Chapman and Hall, London, pp Haas V (1985) Colonial and single breeding in fieldfares Turdus pilaris L.: a comparison of nesting success in early and late broods. Behav Ecol Sociobiol 16: Heckrotte C (1967) Relations of body temperature, size, and crawling speed of the common garter snake, Thamnophis sirtalis. Copeia 1967: Hossie TJ, Murray DL (2010) You can t run but you can hide: refuge use in frog tadpoles elicits density dependent predation by dragonfly larvae. Oecologia 163: Hossie TJ, Murray DL (2011) Effects of structural refuge and density on foraging behaviour and mortality of hungry tadpoles subject to predation risk. Ethology, DOI: /j x Kearney M (2002) Hot rocks and much-too-hot rocks: seasonal patterns of retreat-site selection by a nocturnal ectotherm. J Therm Biol 27: Kotler BP, Brown SJ, Bouskila A. (2004). Apprehension and time allocation in gerbils: the effects of predatory risk and energetic state. Ecology 85: Ladyman M, Bonnet X, Lourdais O, Bradshaw D, Naulleau G (2003) Gestation, thermoregulation and metabolism in a viviparous snake, Vipera aspis: evidence for fecundity-independent costs. Physiol Biochem Zool 76: Lagarde F, Bonnet X, Corbin J, Henen B, Nagy K (2002) A short spring before a long jump: the ecological challenge to the steppe tortoise (Testudo horsfieldi). Can J Zool 80:
28 Lagarde F, Louzizi T, Slimani T, El Mouden H, Ben Kaddour K, Moulherat S, Bonnet X. (2012). Spiny bushes protect tortoises from lethal overheating in arid areas of Morocco. Environ Conserv 39: Langkilde T, O'Connor D, Shine R (2003) Shelter-site use by five species of montane scincid lizards in southeastern Australia. Aust J Zool 51: Mathies T, Felix TA, Lance VA (2001) Effects of trapping and subsequent short-term confinement stress on plasma corticosterone in the brown treesnake (Boiga irregularis) on Guam. Gen Comp Endocrinol 124: McCue MD (2006) Specific dynamic action: a century of investigation. Comp Biochem Physiol 144A: Michel CL, Bonnet X (2010) Contrasted thermal regimes do not influence digestion and growth rates in a snake from temperate climate. Physiol Biochem Zool 83: Millidine KJ, Armstrong JD, Metcalfe NB (2006) Presence of shelter reduces maintenance metabolism of juvenile salmon. Funct Ecol 20: Moore IT, LeMaster MP, Mason RT (2000) Behavioural and hormonal responses to capture stress in the male red-sided garter snake, Thamnophis sirtalis parietalis. Anim Behav 59: Moore IT, Greene MJ, Mason RT (2001) Environmental and seasonal adaptation of the adrenocortical and gonadal responses to capture stress in two population of the male garter snake, Thamnophis sirtalis. J Exp Zool 289: Moore IT, Jessop TS (2003) Stress, reproduction, and adrenocortical modulation in amphibians and reptiles. Horm Behav 43:
29 Möstl E, Palme R (2002) Hormones as indicators of stress. Dom Anim Endocrinol 23: Myles TG (2002) Alarm, aggregation, and defense by Reticulitermes flavipes in response to a naturally occurring isolate of Metarhizium anisopliae. Sociobiology 40: Naulleau G (1983) The effect of temperature on digestion in Vipera aspis. J Herpetol 17: Palacios MG, Sparkman AM, Bronikowski AM (2011) Corticosterone and pace of life in two life-history ecotypes of the garter snake Thamnophis elegans. Gen Comp Endocrinol, doi: /j.ygcen Peterson SC, Johnson ND, LeGuyader JL (1987) Defensive regurgitations of allelochemicals derived from host cyanogenesis by eastern tent caterpillars. Ecology 68: Pike DA, Webb JK, Shine R (2010) Nesting in a thermally challenging environment: nest-site selection in a rock-dwelling gecko, Oedura lesueurii (Reptilia: Gekkonidae). Biol J Lin Soc 99: Pike DA, Webb JK, Andrews RM (2011) Social and thermal cues influence nest-site selection in a nocturnal gecko, Oedura lesueurii. Ethology, DOI: /j x Rabin BS (1999) Stress, immune function and health: the connection. Wiley-Liss, New York. Regal PJ (1966) Thermophilic response following feeding in certain reptiles. Copeia 1966:
30 Robert KA, Vleck C, Bronikowski AM (2009). The effects of maternal corticosterone levels on offspring behavior in fast- and slow-growth garter snakes (Thamnophis elegans). Hormones and Behavior 55: Romero ML, (2004) Physiological stress in ecology: lessons from biomedical research. Trends Ecol Evol 19: Roper TJ, Bennet NC, Conrad TL, Molteno AJ (2001) Environmental conditions in burrows of two species of African mole-rat, Georhychus capensis and Cryptomys damarensis. J Zool L 254 : Saint Girons H (1957) Le cycle sexuel chez Vipera Aspis dans l ouest de la France. Bull Biol Fr Belg 151: Schwarzkopf L, Alford RA (1996) Desiccation and shelter-site use in a tropical amphibian: comparing toads with physical models. Func Ecol 10: Seebacher F, Alford RA (2002) Shelter microhabitats determine body temperature and dehydration rates of a terrestrial amphibian (Bufo marinus). J Herpetol 36: Siegel JM (2005) Clues to the functions of mammalian sleep. Nature 437: Siegel-Causey GL, Hunt JR (1981) Colonial defense behavior in double-crested and pelagic cormorants. Auk 98: Sih A (1997) To hide or not to hide? Refuge use in a fluctuating environment. TREE 12: Souter NJ, Bull CM, Hutchinson MN (2004) Adding burrows to enhance a population of the endangered pygmy blue tongue lizard Tiliqua adelaidensis. Biol Conserv 116:
31 Tillmann JE (2009) An ethological perspective on defecation as an integral part of anti-predatory behaviour in the grey partridge (Perdix perdix L.) at night. J Ethol 27 : Vogel HH (1950) Observations on social behavior in Turkey Vultures. Auk 67: Weiss MR (2006) Defecation behaviour and ecology of insects. Annu Rev Entomol 51: Whitaker P, Ellis K, Shine R (2000) The defensive strike of the eastern brownsnake (Pseudonaja textilis, Elapidae). Funct Ecol 14: Wingfield JC (1994) Modulation of the adrenocortical response to stress in birds. In: Davey KG, Peter RE, Tobe SS (Eds.), Perspectives in Comparative Endocrinology. National Research Council of Canada, Ottawa, pp Wright D, Shapiro J (1990) Refuge availability: a key to understanding the summer disappearance of Daphnia. Freshwater Biol 24: Zuffi MAL, Gentilli A, Cecchinelli E, Pupin F, Bonnet X, Filippi E, Luiselli LM, Barbanera F, Dini F, Fasola M (2009) Geographic variation of life-history traits and reproductive patterns in Continental and Mediterranean asp vipers, Vipera aspis. Biol J Lin Soc 96:
32 716 Captions to table and figures Table 1: Mean characteristics (expressed ±1SD, number in brackets) of the snakes involved in the two experiments (E1 & E2) Figure 1: ten large boxes with three compartments (A, B and C) were used to test microhabitat selection by the vipers (top configuration = Experiment 1), and the physiological consequences associated with such selection (bottom configuration = Experiment 2). The dashed arrow means that the snake can move freely between the compartments (top), the absence of arrow means that the snake is restrained to one compartment (bottom). The circle represents a heat source (50W halogen lamp). Optimal T means that the snake has access to a wide range of ambient temperatures during the day phase (compartment A only, see text for details); cold environment was permanently set to 16 C (compartments B and C). The grey curved line represents shelters (roof tiles). The shelters were available or not. In the experiment 1, zero, one (compartment C), or two shelters (compartments A & C) were placed in the large box. In the Experiment 2 zero versus one shelter was placed in each compartment. Figure 2: Mean score expressing the time spent by the snakes in the compartment fitted with a shelter (on a scale of 0-24 observations per snake, Y-axis). A heat source placed above the shelter was either turned on, or turned off (X-axis). The snakes were unfed (black bars) versus recently fed snakes (grey bars), and they could move freely 32
33 between the three compartments (Figure 1). Means are expressed ±1SE, the numbers indicate the number of snake involved in each situation. The dashed lines indicate respectively the maximal theoretical value (dark grey) and the absence of selection (light grey). Note that several snakes were placed in more than one situation (total number of snakes = 25) Figure 3: Mean heliotheria scores (i.e. microhabitat selection) of 32 snakes placed under three experimental situations (Experiment 1, Figure 1). Mean values are expressed ±1SE. The snakes were unfed (black bars) versus recently fed (grey bars). The dashed lines indicate respectively the maximal theoretical value (dark grey) and the absence of selection (light grey). Heliotheria scores was defined as the number of observations (total 24) the snake was observed under the heat source (an halogen lamp). Figure 4: mean transit time (time elapsed between the ingestion of the prey and the production of the first feces) of the snakes placed into four different experimental situations (Experiment 2, Figure 1). The snakes were not disturbed (black bars, N=88) or harassed through simulation of predator attacks (grey bars, N=56). Figure 5: proportion of the mass of the prey regurgitated versus digested (Y-axis) in function of the number of days the prey remained in the stomach (X-axis) (r = 0.77, P < 0.001). 33
34 Figure 6: number of snakes that regurgitated their prey (grey part of the bar) relative to the number of snakes that did not (hatched part of the bar) in four experimental situations (Experiment 2). The number indicates the mean date of regurgitation. The snakes were not disturbed (top graph) or harassed through simulation of predator attacks (bottom graph) Figure 7: mean basal plasma corticosterone concentrations (±SE) obtained in 63 unfed vipers placed under four experimental situations (Experiment 2). Blood samples were taken in less than 5 min. The dashed lines indicate respectively the mean basal and the mean stress levels obtained in vipers tested for the stress response (i.e. difference between basal and stress-induced corticosterone levels obtained in vipers approximately 30min following handling stress, Wingfield 1994; N=23 vipers, unpublished). Supplemental File: The surface body temperature of the snakes was targeted and measured from distance (20-30cm) using a 3 dots infrared laser thermometer (Raytek MX2, Fotronic Corporation; measurement diameter 19mm, range -30 C-900 C, precision ±1 C, 250ms/reading). The three dots on the back of the snake indicate the area targeted by the thermometer. On small snakes, measurements were performed on coiled individual that offer a relatively large body surface. 34
35 Table 1. Mean characteristics (expressed ±1SD, number in brackets) of the snakes involved in the two experiments 1 & 2 (see text). Experiment Sex Age class SVL (cm) Mass (g) E1 Females Juveniles (N=4) 26.7 ± ± 1.9 Females Sub-adults (N=6) 43.7 ± ± 21.5 Females Adults (N=6) 59.1 ± ± 66.2 Males Juveniles (N=4) 26.6 ± ± 0.9 Males Sub-adults (N=6) 43.1 ± ± 3.2 Males Adults (N=6) 51.8 ± ± 10.5 E2 Females Juveniles (N=18) 27.1 ± ± 8.7 Females Sub-adults (N=12) 48.0 ± ± 41.0 Females Adults (N=18) 56.2 ± ± 67.5 Males Juveniles (N=16) 26.7 ± ± 10.6 Males Sub-adults (N=10) 42.4 ± ± 9.9 Males Adults (N=14) 52.0 ± ±

36 l of Experimental Biology ACCEPTED AUTHOR MAN
37
38
39
40
41
Bio4009 : Projet de recherche/research project
 Bio4009 : Projet de recherche/research project Is emergence after hibernation of the black ratsnake (Elaphe obsoleta) triggered by a thermal gradient reversal? By Isabelle Ceillier 4522350 Supervisor :
Bio4009 : Projet de recherche/research project Is emergence after hibernation of the black ratsnake (Elaphe obsoleta) triggered by a thermal gradient reversal? By Isabelle Ceillier 4522350 Supervisor :
Social and Thermal Cues Influence Nest-site Selection in a Nocturnal Gecko, Oedura lesueurii
 RESEARCH PAPER Social and Thermal Cues Influence Nest-site Selection in a Nocturnal Gecko, Oedura lesueurii David A. Pike*, Jonathan K. Webb* & Robin M. Andrews * School of Biological Sciences A08, University
RESEARCH PAPER Social and Thermal Cues Influence Nest-site Selection in a Nocturnal Gecko, Oedura lesueurii David A. Pike*, Jonathan K. Webb* & Robin M. Andrews * School of Biological Sciences A08, University
CHOOSING YOUR REPTILE LIGHTING AND HEATING
 CHOOSING YOUR REPTILE LIGHTING AND HEATING What lights do I need for my pet Bearded Dragon, Python, Gecko or other reptile, turtle or frog? Is specialised lighting and heating required for indoor reptile
CHOOSING YOUR REPTILE LIGHTING AND HEATING What lights do I need for my pet Bearded Dragon, Python, Gecko or other reptile, turtle or frog? Is specialised lighting and heating required for indoor reptile
THE concept that reptiles have preferred
 Copeia, 2000(3), pp. 841 845 Plasticity in Preferred Body Temperature of Young Snakes in Response to Temperature during Development GABRIEL BLOUIN-DEMERS, KELLEY J. KISSNER, AND PATRICK J. WEATHERHEAD
Copeia, 2000(3), pp. 841 845 Plasticity in Preferred Body Temperature of Young Snakes in Response to Temperature during Development GABRIEL BLOUIN-DEMERS, KELLEY J. KISSNER, AND PATRICK J. WEATHERHEAD
When does a reproducing female viper (Vipera aspis) decide on her litter size?
 decide on her litter size?") Copyright 2003 Wiley-Blackwell. This is the pre-peer reviewed version of an article published in the Journal of Zoology which has been published in final form at http://dx.doi.org/10.1017/s0952836902003059.
Copyright 2003 Wiley-Blackwell. This is the pre-peer reviewed version of an article published in the Journal of Zoology which has been published in final form at http://dx.doi.org/10.1017/s0952836902003059.
Reptiles and amphibian behaviour
 Reptiles and amphibian behaviour Understanding how a healthy reptile and amphibian should look and act takes a lot of observation and practice. Reptiles and amphibians have behaviour that relates to them
Reptiles and amphibian behaviour Understanding how a healthy reptile and amphibian should look and act takes a lot of observation and practice. Reptiles and amphibians have behaviour that relates to them
Impact of colour polymorphism and thermal conditions on thermoregulation, reproductive success, and development in Vipera aspis
 Impact of colour polymorphism and thermal conditions on thermoregulation, reproductive success, and development in Vipera aspis Sylvain Dubey, Johan Schürch, Joaquim Golay, Briséïs Castella, Laura Bonny,
Impact of colour polymorphism and thermal conditions on thermoregulation, reproductive success, and development in Vipera aspis Sylvain Dubey, Johan Schürch, Joaquim Golay, Briséïs Castella, Laura Bonny,
Brumation (Hibernation) in Chelonians and Snakes
 in Chelonians and Snakes") What is Brumation? Brumation (Hibernation) in Chelonians and Snakes Often referred to as hibernation, which is a mammalian process, brumation is the term used to describe the period of dormancy where cold-blooded
What is Brumation? Brumation (Hibernation) in Chelonians and Snakes Often referred to as hibernation, which is a mammalian process, brumation is the term used to describe the period of dormancy where cold-blooded
Corn Snake Care Sheet
 Corn Snake Care Sheet Temperament With the odd exception, Corn Snakes are calm, docile, placid snakes that are hardy and thrive very well in captivity. Due to their temperament Corn Snakes are a recommended
Corn Snake Care Sheet Temperament With the odd exception, Corn Snakes are calm, docile, placid snakes that are hardy and thrive very well in captivity. Due to their temperament Corn Snakes are a recommended
Author's personal copy. Available online at
 Available online at www.sciencedirect.com General and Comparative Endocrinology 155 (2008) 607 612 www.elsevier.com/locate/ygcen A combination of body condition measurements is more informative than conventional
Available online at www.sciencedirect.com General and Comparative Endocrinology 155 (2008) 607 612 www.elsevier.com/locate/ygcen A combination of body condition measurements is more informative than conventional
Effect of Ambient Temperature in Neonate Aspic Vipers: Growth, Locomotor Performance and Defensive Behaviors
 RESEARCH ARTICLE Effect of Ambient Temperature in Neonate Aspic Vipers: Growth, Locomotor Performance and Defensive Behaviors AURÉLIE AÏDAM*, CATHERINE LOUISE MICHEL, AND XAVIER BONNET CEBC CNRS, Beauvoir
RESEARCH ARTICLE Effect of Ambient Temperature in Neonate Aspic Vipers: Growth, Locomotor Performance and Defensive Behaviors AURÉLIE AÏDAM*, CATHERINE LOUISE MICHEL, AND XAVIER BONNET CEBC CNRS, Beauvoir
Objectives: Outline: Idaho Amphibians and Reptiles. Characteristics of Amphibians. Types and Numbers of Amphibians
 Natural History of Idaho Amphibians and Reptiles Wildlife Ecology, University of Idaho Fall 2005 Charles R. Peterson Herpetology Laboratory Department of Biological Sciences, Idaho Museum of Natural History
Natural History of Idaho Amphibians and Reptiles Wildlife Ecology, University of Idaho Fall 2005 Charles R. Peterson Herpetology Laboratory Department of Biological Sciences, Idaho Museum of Natural History
Journal of Thermal Biology
 Journal of Thermal Biology 35 (2010) 324 331 Contents lists available at ScienceDirect Journal of Thermal Biology journal homepage: www.elsevier.com/locate/jtherbio Thermal benefits of artificial shelters
Journal of Thermal Biology 35 (2010) 324 331 Contents lists available at ScienceDirect Journal of Thermal Biology journal homepage: www.elsevier.com/locate/jtherbio Thermal benefits of artificial shelters
Reptilian Physiology
 Reptilian Physiology Physiology, part deux The study of chemical and physical processes in the organism Aspects of the physiology can be informative for understanding organisms in their environment Thermoregulation
Reptilian Physiology Physiology, part deux The study of chemical and physical processes in the organism Aspects of the physiology can be informative for understanding organisms in their environment Thermoregulation
Costs of Anorexia During Pregnancy in a Viviparous Snake (Vipera aspis)
") JOURNAL OF EXPERIMENTAL ZOOLOGY 292:487 493 (2002) DOI 10.1002/jez.10065 Costs of Anorexia During Pregnancy in a Viviparous Snake (Vipera aspis) OLIVIER LOURDAIS, 1,2 * XAVIER BONNET, 1,3 AND PAUL DOUGHTY
JOURNAL OF EXPERIMENTAL ZOOLOGY 292:487 493 (2002) DOI 10.1002/jez.10065 Costs of Anorexia During Pregnancy in a Viviparous Snake (Vipera aspis) OLIVIER LOURDAIS, 1,2 * XAVIER BONNET, 1,3 AND PAUL DOUGHTY
SOAR Research Proposal Summer How do sand boas capture prey they can t see?
 SOAR Research Proposal Summer 2016 How do sand boas capture prey they can t see? Faculty Mentor: Dr. Frances Irish, Assistant Professor of Biological Sciences Project start date and duration: May 31, 2016
SOAR Research Proposal Summer 2016 How do sand boas capture prey they can t see? Faculty Mentor: Dr. Frances Irish, Assistant Professor of Biological Sciences Project start date and duration: May 31, 2016
When does a reproducing female viper (Vipera aspis) decide on her litter size?
 decide on her litter size?") J. Zool., Lond. (2003) 259, 123 129 C 2003 The Zoological Society of London Printed in the United Kingdom DOI:10.1017/S0952836902003059 When does a reproducing female viper (Vipera aspis) decide on her
J. Zool., Lond. (2003) 259, 123 129 C 2003 The Zoological Society of London Printed in the United Kingdom DOI:10.1017/S0952836902003059 When does a reproducing female viper (Vipera aspis) decide on her
OLIVIER LOURDAIS*, XAVIER BONNET*, RICHARD SHINE, DALE DENARDO, GUY NAULLEAU* and MICHAEL GUILLON*
 Ecology 2002 71, Capital-breeding and reproductive effort in a variable Blackwell Science Ltd environment: a longitudinal study of a viviparous snake OLIVIER LOURDAIS*, XAVIER BONNET*, RICHARD SHINE, DALE
Ecology 2002 71, Capital-breeding and reproductive effort in a variable Blackwell Science Ltd environment: a longitudinal study of a viviparous snake OLIVIER LOURDAIS*, XAVIER BONNET*, RICHARD SHINE, DALE
Influence of meal size on postprandial thermophily in cornsnakes (Elaphe guttata)
") TRANSACTIONS OF THE KANSAS ACADEMY OF SCIENCE Vol. 109, no. 3/4 p. 184-190 (2006) Influence of meal size on postprandial thermophily in cornsnakes (Elaphe guttata) LYNETT R. BONTRAGER, DAPHNE M. JONES,
TRANSACTIONS OF THE KANSAS ACADEMY OF SCIENCE Vol. 109, no. 3/4 p. 184-190 (2006) Influence of meal size on postprandial thermophily in cornsnakes (Elaphe guttata) LYNETT R. BONTRAGER, DAPHNE M. JONES,
Weaver Dunes, Minnesota
 Hatchling Orientation During Dispersal from Nests Experimental analyses of an early life stage comparing orientation and dispersal patterns of hatchlings that emerge from nests close to and far from wetlands
Hatchling Orientation During Dispersal from Nests Experimental analyses of an early life stage comparing orientation and dispersal patterns of hatchlings that emerge from nests close to and far from wetlands
8/19/2013. Topic 12: Water & Temperature. Why are water and temperature important? Why are water and temperature important?
 Topic 2: Water & Temperature Why are water and temperature important? Why are water and temperature important for herps? What are adaptations for gaining water? What are adaptations for limiting loss of
Topic 2: Water & Temperature Why are water and temperature important? Why are water and temperature important for herps? What are adaptations for gaining water? What are adaptations for limiting loss of
Mental stim ulation it s not just for dogs!! By Danielle Middleton- Beck BSc hons, PGDip CABC
 Milo, Congo African Grey by Elaine Henley Mental stim ulation it s not just for dogs!! By Danielle Middleton- Beck BSc hons, PGDip CABC Dexter, Green Iguana by Danielle Middleton-Beck Exotic pets include
Milo, Congo African Grey by Elaine Henley Mental stim ulation it s not just for dogs!! By Danielle Middleton- Beck BSc hons, PGDip CABC Dexter, Green Iguana by Danielle Middleton-Beck Exotic pets include
Northern Copperhead Updated: April 8, 2018
 Interpretation Guide Northern Copperhead Updated: April 8, 2018 Status Danger Threats Population Distribution Habitat Diet Size Longevity Social Family Units Reproduction Our Animals Scientific Name Least
Interpretation Guide Northern Copperhead Updated: April 8, 2018 Status Danger Threats Population Distribution Habitat Diet Size Longevity Social Family Units Reproduction Our Animals Scientific Name Least
The Effect of Aerial Exposure Temperature on Balanus balanoides Feeding Behavior
 The Effect of Aerial Exposure Temperature on Balanus balanoides Feeding Behavior Gracie Thompson* and Matt Goldberg Monday Afternoon Biology 334A Laboratory, Fall 2014 Abstract The impact of climate change
The Effect of Aerial Exposure Temperature on Balanus balanoides Feeding Behavior Gracie Thompson* and Matt Goldberg Monday Afternoon Biology 334A Laboratory, Fall 2014 Abstract The impact of climate change
Appendix 6.4. Reptile Survey
 Appendix 6.4 Reptile Survey University of Reading Whiteknights Campus Reptile Survey 2008 Prepared by:, Oxford July 2008 Mallams Court 18 Milton Park Abingdon Oxon OX14 4RP Tel 01235 821888 Fax 01235 820351
Appendix 6.4 Reptile Survey University of Reading Whiteknights Campus Reptile Survey 2008 Prepared by:, Oxford July 2008 Mallams Court 18 Milton Park Abingdon Oxon OX14 4RP Tel 01235 821888 Fax 01235 820351
About Reptiles A Guide for Children. Cathryn Sill Illustrated by John Sill
 About Reptiles About Reptiles A Guide for Children Cathryn Sill Illustrated by John Sill For the One who created reptiles. Genesis 1:24 Published by PEACHTREE PUBLISHERS, LTD. 1700 Chattahoochee Avenue
About Reptiles About Reptiles A Guide for Children Cathryn Sill Illustrated by John Sill For the One who created reptiles. Genesis 1:24 Published by PEACHTREE PUBLISHERS, LTD. 1700 Chattahoochee Avenue
Conservation (last three 3 lecture periods, mostly as a led discussion). We can't cover everything, but that should serve as a rough outline.
. We can't cover everything, but that should serve as a rough outline.") Comments on the rest of the semester: Subjects to be discussed: Temperature relationships. Echolocation. Conservation (last three 3 lecture periods, mostly as a led discussion). Possibly (in order of importance):
Comments on the rest of the semester: Subjects to be discussed: Temperature relationships. Echolocation. Conservation (last three 3 lecture periods, mostly as a led discussion). Possibly (in order of importance):
Station #4. All information Adapted from:http://school.discoveryeducation.com/lessonplans/activities/makeitahabitat/adaptations.html and other sites
 Adaptation Homework Station #1 GOAL: Avoid the Sun s heat and keep themselves cool. Animals spend the daylight hours hiding in burrows or behind boulders. They come out at night to hunt and forage for
Adaptation Homework Station #1 GOAL: Avoid the Sun s heat and keep themselves cool. Animals spend the daylight hours hiding in burrows or behind boulders. They come out at night to hunt and forage for
Management of bold wolves
 Policy Support Statements of the Large Carnivore Initiative for Europe (LCIE). Policy support statements are intended to provide a short indication of what the LCIE regards as being good management practice
Policy Support Statements of the Large Carnivore Initiative for Europe (LCIE). Policy support statements are intended to provide a short indication of what the LCIE regards as being good management practice
Reptiles Notes. Compiled by the Davidson College Herpetology Laboratory
 Reptiles Notes Compiled by the Davidson College Herpetology Laboratory Eastern Hognose Snake Green Tree Frog Reptiles and Amphibians Ectothermic Regulate temperature from outside sources Water temperature
Reptiles Notes Compiled by the Davidson College Herpetology Laboratory Eastern Hognose Snake Green Tree Frog Reptiles and Amphibians Ectothermic Regulate temperature from outside sources Water temperature
Short-term Water Potential Fluctuations and Eggs of the Red-eared Slider Turtle (Trachemys scripta elegans)
") Zoology and Genetics Publications Zoology and Genetics 2001 Short-term Water Potential Fluctuations and Eggs of the Red-eared Slider Turtle (Trachemys scripta elegans) John K. Tucker Illinois Natural History
Zoology and Genetics Publications Zoology and Genetics 2001 Short-term Water Potential Fluctuations and Eggs of the Red-eared Slider Turtle (Trachemys scripta elegans) John K. Tucker Illinois Natural History
Class Reptilia Testudines Squamata Crocodilia Sphenodontia
 Class Reptilia Testudines (around 300 species Tortoises and Turtles) Squamata (around 7,900 species Snakes, Lizards and amphisbaenids) Crocodilia (around 23 species Alligators, Crocodiles, Caimans and
Class Reptilia Testudines (around 300 species Tortoises and Turtles) Squamata (around 7,900 species Snakes, Lizards and amphisbaenids) Crocodilia (around 23 species Alligators, Crocodiles, Caimans and
Station 1 1. (3 points) Identification: Station 2 6. (3 points) Identification:
 Identification: Station 2 6. (3 points) Identification:") SOnerd s 2018-2019 Herpetology SSSS Test 1 SOnerd s SSSS 2018-2019 Herpetology Test Station 20 sounds found here: https://drive.google.com/drive/folders/1oqrmspti13qv_ytllk_yy_vrie42isqe?usp=sharing Station
SOnerd s 2018-2019 Herpetology SSSS Test 1 SOnerd s SSSS 2018-2019 Herpetology Test Station 20 sounds found here: https://drive.google.com/drive/folders/1oqrmspti13qv_ytllk_yy_vrie42isqe?usp=sharing Station
rodent species in Australia to the fecal odor of various predators. Rattus fuscipes (bush
 Sample paper critique #2 The article by Hayes, Nahrung and Wilson 1 investigates the response of three rodent species in Australia to the fecal odor of various predators. Rattus fuscipes (bush rat), Uromys
Sample paper critique #2 The article by Hayes, Nahrung and Wilson 1 investigates the response of three rodent species in Australia to the fecal odor of various predators. Rattus fuscipes (bush rat), Uromys
Squamates of Connecticut
 Squamates of Connecticut Reptilia Turtles are sisters to crocodiles and birds Yeah, birds are reptiles, haven t you watched Jurassic Park yet? Lizards and snakes are part of one clade called the squamates
Squamates of Connecticut Reptilia Turtles are sisters to crocodiles and birds Yeah, birds are reptiles, haven t you watched Jurassic Park yet? Lizards and snakes are part of one clade called the squamates
Climate affects embryonic development in a viviparous snake, Vipera aspis
 OIKOS 104: 551/560, 2004 Climate affects embryonic development in a viviparous snake, Vipera aspis Olivier Lourdais, Richard Shine, Xavier Bonnet, Michaël Guillon and Guy Naulleau Lourdais, O., Shine,
OIKOS 104: 551/560, 2004 Climate affects embryonic development in a viviparous snake, Vipera aspis Olivier Lourdais, Richard Shine, Xavier Bonnet, Michaël Guillon and Guy Naulleau Lourdais, O., Shine,
Habitats provide food, water, and shelter which animals need to survive.
 Adaptation Adaptations are the way living organisms cope with environmental stresses and pressures A biological adaptation is an anatomical structure, physiological process or behavioral trait of an organism
Adaptation Adaptations are the way living organisms cope with environmental stresses and pressures A biological adaptation is an anatomical structure, physiological process or behavioral trait of an organism
Status and Management of Amphibians on Montana Rangelands
 Status and Management of Amphibians on Montana Rangelands Society For Range Management Meeting February 9, 2011 - Billings, Montana Bryce A. Maxell Interim Director / Senior Zoologist Montana Natural Heritage
Status and Management of Amphibians on Montana Rangelands Society For Range Management Meeting February 9, 2011 - Billings, Montana Bryce A. Maxell Interim Director / Senior Zoologist Montana Natural Heritage
Reptile Round Up. An Educator s Guide to the Program
 Reptile Round Up An Educator s Guide to the Program GRADES: K-3 PROGRAM DESCRIPTION: This guide provided by the Oklahoma Aquarium explores reptiles and their unique characteristics. The Reptile Round Up
Reptile Round Up An Educator s Guide to the Program GRADES: K-3 PROGRAM DESCRIPTION: This guide provided by the Oklahoma Aquarium explores reptiles and their unique characteristics. The Reptile Round Up
Estimating radionuclide transfer to reptiles
 Estimating radionuclide transfer to reptiles Mike Wood University of Liverpool What are reptiles? Animals in the Class Reptilia c. 8000 species endangered (hence protected) Types of reptile Snakes Lizards
Estimating radionuclide transfer to reptiles Mike Wood University of Liverpool What are reptiles? Animals in the Class Reptilia c. 8000 species endangered (hence protected) Types of reptile Snakes Lizards
Survivorship. Demography and Populations. Avian life history patterns. Extremes of avian life history patterns
 Demography and Populations Survivorship Demography is the study of fecundity and survival Four critical variables Age of first breeding Number of young fledged each year Juvenile survival Adult survival
Demography and Populations Survivorship Demography is the study of fecundity and survival Four critical variables Age of first breeding Number of young fledged each year Juvenile survival Adult survival
Managing Uplands with Keystone Species. The Case of the Gopher tortoise (Gopherus polyphemus)
") Managing Uplands with Keystone Species The Case of the Gopher tortoise (Gopherus polyphemus) Biology Question: Why consider the gopher tortoise for conservation to begin with? Answer: The gopher tortoise
Managing Uplands with Keystone Species The Case of the Gopher tortoise (Gopherus polyphemus) Biology Question: Why consider the gopher tortoise for conservation to begin with? Answer: The gopher tortoise
Do researchers impact their study populations? Assessing the effect of field procedures in a long term population monitoring of sea kraits
 Amphibia-Reptilia 33 (2012): 365-372 Do researchers impact their study populations? Assessing the effect of field procedures in a long term population monitoring of sea kraits Thomas Fauvel 1,2,, François
Amphibia-Reptilia 33 (2012): 365-372 Do researchers impact their study populations? Assessing the effect of field procedures in a long term population monitoring of sea kraits Thomas Fauvel 1,2,, François
Broom, D.M In Proceedings of Aquavision 1999, 1-6. Stavanger: Proceedings of Aquavision. Fish welfare and the public perception of farmed fish
 Broom, D.M. 1999. In Proceedings of Aquavision 1999, 1-6. Stavanger: Proceedings of Aquavision. Pre-publication copy Fish welfare and the public perception of farmed fish D.M. Broom Department of Clinical
Broom, D.M. 1999. In Proceedings of Aquavision 1999, 1-6. Stavanger: Proceedings of Aquavision. Pre-publication copy Fish welfare and the public perception of farmed fish D.M. Broom Department of Clinical
Contrasting Response to Predator and Brood Parasite Signals in the Song Sparrow (melospiza melodia)
") Luke Campillo and Aaron Claus IBS Animal Behavior Prof. Wisenden 6/25/2009 Contrasting Response to Predator and Brood Parasite Signals in the Song Sparrow (melospiza melodia) Abstract: The Song Sparrow
Luke Campillo and Aaron Claus IBS Animal Behavior Prof. Wisenden 6/25/2009 Contrasting Response to Predator and Brood Parasite Signals in the Song Sparrow (melospiza melodia) Abstract: The Song Sparrow
Faculty Mentor, Department of Integrative Biology, Oklahoma State University
 Sex Recognition in Anole Lizards Authors: Shelby Stavins and Dr. Matthew Lovern * Abstract: Sexual selection is the process that furthers a species, and either improves the genetic variability or weakens
Sex Recognition in Anole Lizards Authors: Shelby Stavins and Dr. Matthew Lovern * Abstract: Sexual selection is the process that furthers a species, and either improves the genetic variability or weakens
The effects of diet upon pupal development and cocoon formation by the cat flea (Siphonaptera: Pulicidae)
") June, 2002 Journal of Vector Ecology 39 The effects of diet upon pupal development and cocoon formation by the cat flea (Siphonaptera: Pulicidae) W. Lawrence and L. D. Foil Department of Entomology, Louisiana
June, 2002 Journal of Vector Ecology 39 The effects of diet upon pupal development and cocoon formation by the cat flea (Siphonaptera: Pulicidae) W. Lawrence and L. D. Foil Department of Entomology, Louisiana
Supplementary Fig. 1: Comparison of chase parameters for focal pack (a-f, n=1119) and for 4 dogs from 3 other packs (g-m, n=107).
 and for 4 dogs from 3 other packs (g-m, n=107).") Supplementary Fig. 1: Comparison of chase parameters for focal pack (a-f, n=1119) and for 4 dogs from 3 other packs (g-m, n=107). (a,g) Maximum stride speed, (b,h) maximum tangential acceleration, (c,i)
Supplementary Fig. 1: Comparison of chase parameters for focal pack (a-f, n=1119) and for 4 dogs from 3 other packs (g-m, n=107). (a,g) Maximum stride speed, (b,h) maximum tangential acceleration, (c,i)
Thermal adaptation of maternal and embryonic phenotypes in a geographically widespread ectotherm
 International Congress Series 1275 (2004) 258 266 www.ics-elsevier.com Thermal adaptation of maternal and embryonic phenotypes in a geographically widespread ectotherm Michael J. Angilletta Jr. a, *, Christopher
International Congress Series 1275 (2004) 258 266 www.ics-elsevier.com Thermal adaptation of maternal and embryonic phenotypes in a geographically widespread ectotherm Michael J. Angilletta Jr. a, *, Christopher
Institutional Animal Care & Use Program - UTEP Title: Animal Enrichment Policy#: 014 Date in Effect: 27 February 2015
 IACUC Policy #014-A, Page 1 of 5 Institutional Animal Care & Use Program - UTEP Title: Animal Enrichment Policy#: 014 Date in Effect: 27 February 2015 Version #: A Rev Date: In Effect Rescinded Date Rescinded:
IACUC Policy #014-A, Page 1 of 5 Institutional Animal Care & Use Program - UTEP Title: Animal Enrichment Policy#: 014 Date in Effect: 27 February 2015 Version #: A Rev Date: In Effect Rescinded Date Rescinded:
Reptilian Requirements Created by the North Carolina Aquarium at Fort Fisher Education Section
 Essential Question: North Carolina Aquariums Education Section Reptilian Requirements Created by the North Carolina Aquarium at Fort Fisher Education Section What physical and behavioral adaptations do
Essential Question: North Carolina Aquariums Education Section Reptilian Requirements Created by the North Carolina Aquarium at Fort Fisher Education Section What physical and behavioral adaptations do
Effects of food supplementation on the physiological ecology of female Western diamond-backed rattlesnakes (Crotalus atrox)
") Oecologia (2005) DOI 10.1007/s00442-005-0056-x ECOPHYSIOLOGY Emily N. Taylor Æ Michael A. Malawy Dawn M. Browning Æ Shea V. Lemar Æ Dale F. DeNardo Effects of food supplementation on the physiological
Oecologia (2005) DOI 10.1007/s00442-005-0056-x ECOPHYSIOLOGY Emily N. Taylor Æ Michael A. Malawy Dawn M. Browning Æ Shea V. Lemar Æ Dale F. DeNardo Effects of food supplementation on the physiological
Thermal quality influences effectiveness of thermoregulation, habitat use, and behaviour in milk snakes
 Oecologia (2006) 148: 1 11 DOI 10.1007/s00442-005-0350-7 ECOPHYSIOLOGY Jeffrey R. Row Æ Gabriel Blouin-Demers Thermal quality influences effectiveness of thermoregulation, habitat use, and behaviour in
Oecologia (2006) 148: 1 11 DOI 10.1007/s00442-005-0350-7 ECOPHYSIOLOGY Jeffrey R. Row Æ Gabriel Blouin-Demers Thermal quality influences effectiveness of thermoregulation, habitat use, and behaviour in
Proceedings of the World Small Animal Veterinary Association Sydney, Australia 2007
 Proceedings of the World Small Animal Veterinary Association Sydney, Australia 2007 Hosted by: Australian Small Animal Veterinary Association (ASAVA) Australian Small Animal Veterinary Association (ASAVA)
Proceedings of the World Small Animal Veterinary Association Sydney, Australia 2007 Hosted by: Australian Small Animal Veterinary Association (ASAVA) Australian Small Animal Veterinary Association (ASAVA)
Nest-site selection in Eastern hognose snakes (Heterodon platirhinos) Casey Peet-Paré
 Casey Peet-Paré") Nest-site selection in Eastern hognose snakes (Heterodon platirhinos) by Casey Peet-Paré Thesis submitted to the Department of Biology in partial fulfillment of the requirements for the B.Sc. Honours degree,
Nest-site selection in Eastern hognose snakes (Heterodon platirhinos) by Casey Peet-Paré Thesis submitted to the Department of Biology in partial fulfillment of the requirements for the B.Sc. Honours degree,
Sprint speed capacity of two alpine skink species, Eulamprus kosciuskoi and Pseudemoia entrecasteauxii
 Sprint speed capacity of two alpine skink species, Eulamprus kosciuskoi and Pseudemoia entrecasteauxii Isabella Robinson, Bronte Sinclair, Holly Sargent, Xiaoyun Li Abstract As global average temperatures
Sprint speed capacity of two alpine skink species, Eulamprus kosciuskoi and Pseudemoia entrecasteauxii Isabella Robinson, Bronte Sinclair, Holly Sargent, Xiaoyun Li Abstract As global average temperatures
Title of Project: Distribution of the Collared Lizard, Crotophytus collaris, in the Arkansas River Valley and Ouachita Mountains
 Title of Project: Distribution of the Collared Lizard, Crotophytus collaris, in the Arkansas River Valley and Ouachita Mountains Project Summary: This project will seek to monitor the status of Collared
Title of Project: Distribution of the Collared Lizard, Crotophytus collaris, in the Arkansas River Valley and Ouachita Mountains Project Summary: This project will seek to monitor the status of Collared
08 alberts part2 7/23/03 9:10 AM Page 95 PART TWO. Behavior and Ecology
 08 alberts part2 7/23/03 9:10 AM Page 95 PART TWO Behavior and Ecology 08 alberts part2 7/23/03 9:10 AM Page 96 08 alberts part2 7/23/03 9:10 AM Page 97 Introduction Emília P. Martins Iguanas have long
08 alberts part2 7/23/03 9:10 AM Page 95 PART TWO Behavior and Ecology 08 alberts part2 7/23/03 9:10 AM Page 96 08 alberts part2 7/23/03 9:10 AM Page 97 Introduction Emília P. Martins Iguanas have long
Keywords Correspondence
 Journal of Zoology Contrasted thermal preferences translate into divergences in habitat use and realized performance in two sympatric snakes H. Lelièvre 1,2, G. Blouin-Demers 3, D. Pinaud 1, H. Lisse 1,
Journal of Zoology Contrasted thermal preferences translate into divergences in habitat use and realized performance in two sympatric snakes H. Lelièvre 1,2, G. Blouin-Demers 3, D. Pinaud 1, H. Lisse 1,
Comparative Physiology 2007 Second Midterm Exam. 1) 8 pts. 2) 14 pts. 3) 12 pts. 4) 17 pts. 5) 10 pts. 6) 8 pts. 7) 12 pts. 8) 10 pts. 9) 9 pts.
 8 pts. 2) 14 pts. 3) 12 pts. 4) 17 pts. 5) 10 pts. 6) 8 pts. 7) 12 pts. 8) 10 pts. 9) 9 pts.") Name: Comparative Physiology 2007 Second Midterm Exam 1) 8 pts 2) 14 pts 3) 12 pts 4) 17 pts 5) 10 pts 6) 8 pts 7) 12 pts 8) 10 pts 9) 9 pts Total 1. Cells I and II, shown below, are found in the gills
Name: Comparative Physiology 2007 Second Midterm Exam 1) 8 pts 2) 14 pts 3) 12 pts 4) 17 pts 5) 10 pts 6) 8 pts 7) 12 pts 8) 10 pts 9) 9 pts Total 1. Cells I and II, shown below, are found in the gills
EFFECTS OF ENVIRONMENTAL TEMPERATURE, RELATIVE HUMIDITY, FASTING AND FEEDING ON THE BODY TEMPERATURE OF LAYING HENS
 EFFECTS OF ENVIRONMENTAL TEMPERATURE, RELATIVE HUMIDITY, FASTING AND FEEDING ON THE BODY TEMPERATURE OF LAYING HENS W. K. SMITH* Summary The separate effects of air temperature, relative humidity, fasting
EFFECTS OF ENVIRONMENTAL TEMPERATURE, RELATIVE HUMIDITY, FASTING AND FEEDING ON THE BODY TEMPERATURE OF LAYING HENS W. K. SMITH* Summary The separate effects of air temperature, relative humidity, fasting
The effect of testosterone injections on aggression and begging behaviour of black headed gull chicks (Larus ridibundus)
") The effect of testosterone injections on aggression and begging behaviour of black headed gull chicks (Larus ridibundus) Abstract L.M. van Zomeren april 2009 supervised by Giuseppe Boncoraglio and Ton
The effect of testosterone injections on aggression and begging behaviour of black headed gull chicks (Larus ridibundus) Abstract L.M. van Zomeren april 2009 supervised by Giuseppe Boncoraglio and Ton
A test of the thermal coadaptation hypothesis in the common map turtle (Graptemys geographica) Elad Ben-Ezra. Supervisor: Dr. Gabriel Blouin-Demers
 Elad Ben-Ezra. Supervisor: Dr. Gabriel Blouin-Demers") A test of the thermal coadaptation hypothesis in the common map turtle (Graptemys geographica) by Elad Ben-Ezra Supervisor: Dr. Gabriel Blouin-Demers Thesis submitted to the Department of Biology in partial
A test of the thermal coadaptation hypothesis in the common map turtle (Graptemys geographica) by Elad Ben-Ezra Supervisor: Dr. Gabriel Blouin-Demers Thesis submitted to the Department of Biology in partial
When a species can t stand the heat
 When a species can t stand the heat Featured scientists: Kristine Grayson from University of Richmond, Nicola Mitchell from University of Western Australia, & Nicola Nelson from Victoria University of
When a species can t stand the heat Featured scientists: Kristine Grayson from University of Richmond, Nicola Mitchell from University of Western Australia, & Nicola Nelson from Victoria University of
JoJoKeKe s Herpetology Exam
 ~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~~*~*~*~*~*~*~*~*~*~*~*~*~*~*~ JoJoKeKe s Herpetology Exam (SSSS) 2:30 to be given at each station- B/C Station 1: 1.) What is the family & genus of the shown
~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~~*~*~*~*~*~*~*~*~*~*~*~*~*~*~ JoJoKeKe s Herpetology Exam (SSSS) 2:30 to be given at each station- B/C Station 1: 1.) What is the family & genus of the shown
Broiler Management for Birds Grown to Low Kill Weights ( lb / kg)
") Broiler Management for Birds Grown to Low Kill Weights (3.3-4.0 lb / 1.5-1.8 kg) April 2008 Michael Garden, Regional Technical Manager Turkey, Middle East & Africa, Aviagen Robin Singleton, Technical Service
Broiler Management for Birds Grown to Low Kill Weights (3.3-4.0 lb / 1.5-1.8 kg) April 2008 Michael Garden, Regional Technical Manager Turkey, Middle East & Africa, Aviagen Robin Singleton, Technical Service
Breeding White Storks( Ciconia ciconia at Chessington World of Adventures Paul Wexler
 Breeding White Storks(Ciconia ciconia) at Chessington World of Adventures Paul Wexler The White Stork belongs to the genus Ciconia of which there are seven other species incorporated predominantly throughout
Breeding White Storks(Ciconia ciconia) at Chessington World of Adventures Paul Wexler The White Stork belongs to the genus Ciconia of which there are seven other species incorporated predominantly throughout
Pre-lab homework Lab 8: Food chains in the wild.
 Pre-lab homework Lab 8: Food chains in the wild. Lab Section: Name: Put your field hat on and complete the questions below before coming to lab! The bits of information you and your classmates collect
Pre-lab homework Lab 8: Food chains in the wild. Lab Section: Name: Put your field hat on and complete the questions below before coming to lab! The bits of information you and your classmates collect
LIFE history tradeoffs are prevalent in nature because
 Copeia 2012, No. 1, 100 105 Do Sidewinder Rattlesnakes (Crotalus cerastes, Viperidae) Cease Feeding During the Breeding Season? Michael M. Webber 1, Xavier Glaudas 1, and Javier A. Rodríguez-Robles 1 Seasonal
Copeia 2012, No. 1, 100 105 Do Sidewinder Rattlesnakes (Crotalus cerastes, Viperidae) Cease Feeding During the Breeding Season? Michael M. Webber 1, Xavier Glaudas 1, and Javier A. Rodríguez-Robles 1 Seasonal
Grade Level: 3-5. Next Generation Sunshine State Standards SC.3.L.15.1 SC.4.L.16.2; SC.4.L.17.4 SC.5.L.15.1; SC.5.L.17.1
 Grade Level: 3-5 Next Generation Sunshine State Standards SC.3.L.15.1 SC.4.L.16.2; SC.4.L.17.4 SC.5.L.15.1; SC.5.L.17.1 Program Overview Discover the realm of reptiles, amazing creatures adapted to land
Grade Level: 3-5 Next Generation Sunshine State Standards SC.3.L.15.1 SC.4.L.16.2; SC.4.L.17.4 SC.5.L.15.1; SC.5.L.17.1 Program Overview Discover the realm of reptiles, amazing creatures adapted to land
WHAT ARE HERPTILES? WHICH IS WHICH? 1. Vertebrates are animals that have 2. Complete the following chart of vertebrate groups: EGGS LAID WHERE?
 WHAT ARE HERPTILES? 1. Vertebrates are animals that have 2. Complete the following chart of vertebrate groups: SKIN COVERING? GILLS OR LUNGS? EGGS LAID WHERE? ENDOTHERMIC OR ECTOTHERMIC Fish AMPHIBIANS
WHAT ARE HERPTILES? 1. Vertebrates are animals that have 2. Complete the following chart of vertebrate groups: SKIN COVERING? GILLS OR LUNGS? EGGS LAID WHERE? ENDOTHERMIC OR ECTOTHERMIC Fish AMPHIBIANS
Adjustments In Parental Care By The European Starling (Sturnus Vulgaris): The Effect Of Female Condition
: The Effect Of Female Condition") Proceedings of The National Conference on Undergraduate Research (NCUR) 2003 University of Utah, Salt Lake City, Utah March 13-15, 2003 Adjustments In Parental Care By The European Starling (Sturnus Vulgaris):
Proceedings of The National Conference on Undergraduate Research (NCUR) 2003 University of Utah, Salt Lake City, Utah March 13-15, 2003 Adjustments In Parental Care By The European Starling (Sturnus Vulgaris):
Institutional Animal Care and Use Committee Guideline. Defining Humane Endpoints and End-stage Illness BACKGROUND. Definitions.
 Institutional Animal Care and Use Committee Guideline Applies to WSU and John D. Dingell VAMC Subject: BACKGROUND Animals used in biomedical research may at times display signs of pain or distress related
Institutional Animal Care and Use Committee Guideline Applies to WSU and John D. Dingell VAMC Subject: BACKGROUND Animals used in biomedical research may at times display signs of pain or distress related
Biological Invasions and Herpetology. 4/18/13 Chris Thawley
 Biological Invasions and Herpetology 4/18/13 Chris Thawley What are some invasive species? http://news.discovery.com/animals/videos/animals-jumping-carp-attack-explained.htm What is an Invasive species?
Biological Invasions and Herpetology 4/18/13 Chris Thawley What are some invasive species? http://news.discovery.com/animals/videos/animals-jumping-carp-attack-explained.htm What is an Invasive species?
Rodent Husbandry and Care 201 Cynthia J. Brown and Thomas M. Donnelly
 EXOTIC PET MANAGEMENT FOR THE TECHNICIAN Preface Michelle S. Schulte and Agnes E. Rupley xi Rodent Husbandry and Care 201 Cynthia J. Brown and Thomas M. Donnelly This article reviews the husbandry, care
EXOTIC PET MANAGEMENT FOR THE TECHNICIAN Preface Michelle S. Schulte and Agnes E. Rupley xi Rodent Husbandry and Care 201 Cynthia J. Brown and Thomas M. Donnelly This article reviews the husbandry, care
BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS
 Nov., 1965 505 BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS Lack ( 1954; 40-41) has pointed out that in species of birds which have asynchronous hatching, brood size may be adjusted
Nov., 1965 505 BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS Lack ( 1954; 40-41) has pointed out that in species of birds which have asynchronous hatching, brood size may be adjusted
Kris Descovich How do captive wombats cope with extreme environmental seasons? This paper was presented at the 2011 National Wombat Conference
 Kris Descovich How do captive wombats cope with extreme environmental seasons? This paper was presented at the 2011 National Wombat Conference National Wombat Conference Sponsored by The Wombat Protection
Kris Descovich How do captive wombats cope with extreme environmental seasons? This paper was presented at the 2011 National Wombat Conference National Wombat Conference Sponsored by The Wombat Protection
Silverback Male Presence and Group Stability in Gorillas (Gorilla gorilla gorilla)
") Brief Report Folia Primatol 753 Received: August 16, 2002 DOI: 10.1159/0000XXXXX Accepted after revision: October 30, 2002 Silverback Male Presence and Group Stability in Gorillas (Gorilla gorilla gorilla)
Brief Report Folia Primatol 753 Received: August 16, 2002 DOI: 10.1159/0000XXXXX Accepted after revision: October 30, 2002 Silverback Male Presence and Group Stability in Gorillas (Gorilla gorilla gorilla)
5 State of the Turtles
 CHALLENGE 5 State of the Turtles In the previous Challenges, you altered several turtle properties (e.g., heading, color, etc.). These properties, called turtle variables or states, allow the turtles to
CHALLENGE 5 State of the Turtles In the previous Challenges, you altered several turtle properties (e.g., heading, color, etc.). These properties, called turtle variables or states, allow the turtles to
Like mother, like daughter: inheritance of nest-site
 Like mother, like daughter: inheritance of nest-site location in snakes Gregory P. Brown and Richard Shine* School of Biological Sciences A0, University of Sydney, NSW 00, Australia *Author for correspondence
Like mother, like daughter: inheritance of nest-site location in snakes Gregory P. Brown and Richard Shine* School of Biological Sciences A0, University of Sydney, NSW 00, Australia *Author for correspondence
Mexican Gray Wolf Reintroduction
 Mexican Gray Wolf Reintroduction New Mexico Supercomputing Challenge Final Report April 2, 2014 Team Number 24 Centennial High School Team Members: Andrew Phillips Teacher: Ms. Hagaman Project Mentor:
Mexican Gray Wolf Reintroduction New Mexico Supercomputing Challenge Final Report April 2, 2014 Team Number 24 Centennial High School Team Members: Andrew Phillips Teacher: Ms. Hagaman Project Mentor:
FIELD GUIDE TO NORTH AMERICAN MAMMALS Northern Short tailed Shrew (Blarina brevicauda)
") Northern Short tailed Shrew (Blarina brevicauda) Northern Short tailed Shrews have poisonous saliva. This enables them to kill mice and larger prey and paralyze invertebrates such as snails and store them
Northern Short tailed Shrew (Blarina brevicauda) Northern Short tailed Shrews have poisonous saliva. This enables them to kill mice and larger prey and paralyze invertebrates such as snails and store them
BEDDING GUIDE Choose the right bedding for your reptile. Ornate Uromastyx (Uromastyx ornata)
") BEDDING GUIDE Choose the right bedding for your reptile. Ornate Uromastyx (Uromastyx ornata) Preferred Acceptable Chameleons Bearded Dragons Desert Geckos Frogs/Toads Anoles Iguanas Tortoises Monitors
BEDDING GUIDE Choose the right bedding for your reptile. Ornate Uromastyx (Uromastyx ornata) Preferred Acceptable Chameleons Bearded Dragons Desert Geckos Frogs/Toads Anoles Iguanas Tortoises Monitors
Reproductive physiology and eggs
 Reproductive physiology and eggs Class Business Reading for this lecture Required. Gill: Chapter 14 1. Reproductive physiology In lecture I will only have time to go over reproductive physiology briefly,
Reproductive physiology and eggs Class Business Reading for this lecture Required. Gill: Chapter 14 1. Reproductive physiology In lecture I will only have time to go over reproductive physiology briefly,
Desert Reptiles. A forty five Desert Discovery program
 Desert Reptiles A forty five Desert Discovery program To the Teacher: Thank you for making the Desert Reptiles discovery class a part of your curriculum. During this exciting interactive educational program,
Desert Reptiles A forty five Desert Discovery program To the Teacher: Thank you for making the Desert Reptiles discovery class a part of your curriculum. During this exciting interactive educational program,
Lacerta vivipara Jacquin
 Oecologia (Berl.) 19, 165--170 (1975) 9 by Springer-Verlag 1975 Clutch Size and Reproductive Effort in the Lizard Lacerta vivipara Jacquin R. A. Avery Department of Zoology, The University, Bristol Received
Oecologia (Berl.) 19, 165--170 (1975) 9 by Springer-Verlag 1975 Clutch Size and Reproductive Effort in the Lizard Lacerta vivipara Jacquin R. A. Avery Department of Zoology, The University, Bristol Received
All about snakes. What are snakes? Are snakes just lizards without legs? If you want to know more
 Novak.lisa@gmail.com Day 83 12/29/2017 All about snakes What are snakes? Are snakes just lizards without legs? If you want to know more keep reading to find out the answers to the question. The purpose
Novak.lisa@gmail.com Day 83 12/29/2017 All about snakes What are snakes? Are snakes just lizards without legs? If you want to know more keep reading to find out the answers to the question. The purpose
How Does Photostimulation Age Alter the Interaction Between Body Size and a Bonus Feeding Program During Sexual Maturation?
 16 How Does Photostimulation Age Alter the Interaction Between Body Size and a Bonus Feeding Program During Sexual Maturation? R A Renema*, F E Robinson*, and J A Proudman** *Alberta Poultry Research Centre,
16 How Does Photostimulation Age Alter the Interaction Between Body Size and a Bonus Feeding Program During Sexual Maturation? R A Renema*, F E Robinson*, and J A Proudman** *Alberta Poultry Research Centre,
RURAL INDUSTRIES RESEARCH AND DEVELOPMENT CORPORATION FINAL REPORT. Improvement in egg shell quality at high temperatures
 RURAL INDUSTRIES RESEARCH AND DEVELOPMENT CORPORATION FINAL REPORT Project Title: Improvement in egg shell quality at high temperatures RIRDC Project No.: US-43A Research Organisation: University of Sydney
RURAL INDUSTRIES RESEARCH AND DEVELOPMENT CORPORATION FINAL REPORT Project Title: Improvement in egg shell quality at high temperatures RIRDC Project No.: US-43A Research Organisation: University of Sydney
Thermal strategies and energetics in two sympatric colubrid snakes with contrasted exposure
 J Comp Physiol B (2010) 180:415 425 DOI 10.1007/s00360-009-0423-8 ORIGINAL PAPER Thermal strategies and energetics in two sympatric colubrid snakes with contrasted exposure Hervé Lelièvre Maxime Le Hénanff
J Comp Physiol B (2010) 180:415 425 DOI 10.1007/s00360-009-0423-8 ORIGINAL PAPER Thermal strategies and energetics in two sympatric colubrid snakes with contrasted exposure Hervé Lelièvre Maxime Le Hénanff
Shearing Lambs Improves Growth Performance During Periods with Elevated Thermal Load
 Shearing Lambs Improves Growth Performance During Periods with Elevated Thermal Load Jake J. Herrig 1, Simone. M. Holt 2, and J. A. Daniel 2 Department of Animal and Range Sciences Sheep Research Report
Shearing Lambs Improves Growth Performance During Periods with Elevated Thermal Load Jake J. Herrig 1, Simone. M. Holt 2, and J. A. Daniel 2 Department of Animal and Range Sciences Sheep Research Report
Selection for Egg Mass in the Domestic Fowl. 1. Response to Selection
 Selection for Egg Mass in the Domestic Fowl. 1. Response to Selection H. L. MARKS US Department of Agriculture, Science & Education Administration, Agricultural Research, uthern Regional Poultry Breeding
Selection for Egg Mass in the Domestic Fowl. 1. Response to Selection H. L. MARKS US Department of Agriculture, Science & Education Administration, Agricultural Research, uthern Regional Poultry Breeding
Effects of Cage Stocking Density on Feeding Behaviors of Group-Housed Laying Hens
 AS 651 ASL R2018 2005 Effects of Cage Stocking Density on Feeding Behaviors of Group-Housed Laying Hens R. N. Cook Iowa State University Hongwei Xin Iowa State University, hxin@iastate.edu Recommended
AS 651 ASL R2018 2005 Effects of Cage Stocking Density on Feeding Behaviors of Group-Housed Laying Hens R. N. Cook Iowa State University Hongwei Xin Iowa State University, hxin@iastate.edu Recommended
NAME: DATE: SECTION:
 NAME: DATE: SECTION: MCAS PREP PACKET EVOLUTION AND BIODIVERSITY 1. Which of the following observations best supports the conclusion that dolphins and sharks do not have a recent common ancestor? A. Dolphins
NAME: DATE: SECTION: MCAS PREP PACKET EVOLUTION AND BIODIVERSITY 1. Which of the following observations best supports the conclusion that dolphins and sharks do not have a recent common ancestor? A. Dolphins
AGGRESSION (CATS) DIAGNOSING AND TREATING
 DIAGNOSING AND TREATING") AGGRESSION (CATS) DIAGNOSING AND TREATING Aggression is a serious and dangerous behavior problem for cat owners. There are many different types of aggression. Making a diagnosis, determining the prognosis
AGGRESSION (CATS) DIAGNOSING AND TREATING Aggression is a serious and dangerous behavior problem for cat owners. There are many different types of aggression. Making a diagnosis, determining the prognosis
Ciccaba virgata (Mottled Owl)
") Ciccaba virgata (Mottled Owl) Family: Strigidae (Typical Owls) Order: Strigiformes (Owls) Class: Aves (Birds) Fig. 1. Mottled owl, Ciccaba virgata. [http://www.owling.com/mottled13.htm, downloaded 12 November
Ciccaba virgata (Mottled Owl) Family: Strigidae (Typical Owls) Order: Strigiformes (Owls) Class: Aves (Birds) Fig. 1. Mottled owl, Ciccaba virgata. [http://www.owling.com/mottled13.htm, downloaded 12 November
RCPS7-Science-Evolution (RCPS7-Science-Evolution) 1. Which is an adaptation that makes it possible for the animal to survive in a cold climate?
 1. Which is an adaptation that makes it possible for the animal to survive in a cold climate?") Name: Date: 1. Which is an adaptation that makes it possible for the animal to survive in a cold climate? A. tail on a lizard B. scales on a fish C. stripes on a tiger D. fur on a bear 2. Use the picture
Name: Date: 1. Which is an adaptation that makes it possible for the animal to survive in a cold climate? A. tail on a lizard B. scales on a fish C. stripes on a tiger D. fur on a bear 2. Use the picture
Habitats and Field Methods. Friday May 12th 2017
 Habitats and Field Methods Friday May 12th 2017 Announcements Project consultations available today after class Project Proposal due today at 5pm Follow guidelines posted for lecture 4 Field notebooks
Habitats and Field Methods Friday May 12th 2017 Announcements Project consultations available today after class Project Proposal due today at 5pm Follow guidelines posted for lecture 4 Field notebooks
When a species can t stand the heat
 When a species can t stand the heat Featured scientists: Kristine Grayson from University of Richmond, Nicola Mitchell from University of Western Australia, & Nicola Nelson from Victoria University of
When a species can t stand the heat Featured scientists: Kristine Grayson from University of Richmond, Nicola Mitchell from University of Western Australia, & Nicola Nelson from Victoria University of
INFLUENCE OF FEED QUALITY ON THE EXPRESSION OF POST WEANING GROWTH ASBV s IN WHITE SUFFOLK LAMBS
 INFLUENCE OF FEED QUALITY ON THE EXPRESSION OF POST WEANING GROWTH ASBV s IN WHITE SUFFOLK LAMBS Introduction Murray Long ClearView Consultancy www.clearviewconsulting.com.au Findings from an on farm trial
INFLUENCE OF FEED QUALITY ON THE EXPRESSION OF POST WEANING GROWTH ASBV s IN WHITE SUFFOLK LAMBS Introduction Murray Long ClearView Consultancy www.clearviewconsulting.com.au Findings from an on farm trial