Reassessment of the oldest British turtle: Protochelys from the Middle Jurassic Stonesfield Slate of Stonesfield, Oxfordshire, UK
|
|
|
- Irma Newton
- 5 years ago
- Views:
Transcription
1 Reassessment of the oldest British turtle: Protochelys from the Middle Jurassic Stonesfield Slate of Stonesfield, Oxfordshire, UK Jérémy ANQUETIN The Natural History Museum, Department of Palaeontology, Cromwell road, London SW7 5BD (United Kingdom) Julien CLAUDE Université de Montpellier 2, Institut des Sciences de l Évolution de Montpellier, UMR 5554 CNRS, F Montpellier cedex 5 (France) claude@isem.univ-montp2.fr Anquetin J. & Claude J Reassessment of the oldest British turtle: Protochelys from the Middle Jurassic Stonesfield Slate of Stonesfield, Oxfordshire, UK. Geodiversitas 30 (2) : Key Words Reptilia, Testudinata, Protochelys, Bathonian, Middle Jurassic, Stonesfield Slate, England, epidermal scale, taphonomy. ABSTRACT Protochelys Lydekker, 1889 from the Stonesfield Slate (middle Bathonian) is the oldest British turtle and the only record to date of fossil epidermal shell scales preserved isolated from underlying bone. Although known since the 1840s, these remains have never been properly described, figured or compared with other taxa. Here, we provide a thorough reassessment of the available material with a discussion of the exceptional preservation of isolated scales. We conclude that: 1) no satisfactory diagnosis of this taxon can be proposed and Protochelys blakii (Mackie, 1863) has to be considered nomen dubium; 2) the carapace of the Stonesfield turtle has a plesiomorphic morphology (vertebral scales twice as wide as long; fifth vertebral scale as wide anteriorly as it is posteriorly; pleural scales longer than wide) shared with numerous basal turtles. The fossilisation of turtle epidermal scales is extremely rare (only two other examples are known). The Stonesfield material is unique in that the scales are isolated, without underlying bone. A review of the literature shows that isolation of shell scales occurs as a result of two processes: shedding of old scale layers during growth or post mortem disarticulation. We favour the disarticulation hypothesis because complete scales are thicker and more likely to preserve the well-developed ornamentation shown by the Stonesfield scales. Publications Scientifiques du Muséum national d Histoire naturelle, Paris
2 Anquetin J. & Claude J. Mots Clés Reptilia, Testudinata, Protochelys, Bathonien, Jurassique moyen, Stonesfield Slate, Angleterre, écaille épidermique, taphonomie. RÉSUMÉ Réévaluation de la plus ancienne tortue britannique : Protochelys du Jurassique moyen du Stonesfield Slate de Stonesfield, Oxfordshire, Royaume-Uni. Protochelys Lydekker, 1889 du Stonesfield Slate (Bathonien moyen) est la plus ancienne tortue britannique et le seul exemple connu à l heure actuelle de fossiles d écailles épidermiques de carapace de tortue préservées à l état isolé. Bien que connus depuis les années 1840, ces restes n ont jamais été proprement décrits, figurés ou comparés avec les autres taxons. Ici, nous proposons une réévaluation complète du matériel disponible ainsi qu une discussion de l exceptionnelle préservation d écailles isolées. Nous concluons que : 1) aucune diagnose satisfaisante de ce taxon ne peut être proposée et Protochelys blakii (Mackie, 1863) doit être considéré nomen dubium ; 2) la dossière de la tortue de Stonesfield présente une morphologie plésiomorphe (écailles vertébrales deux fois plus larges que longues ; cinquième écaille vertébrale aussi large antérieurement que postérieurement ; écailles pleurales plus longues que larges) partagée avec de nombreuses tortues basales. La fossilisation d écailles épidermiques de tortues est extrêmement rare (seuls deux autres exemples sont connus). Le matériel de Stonesfield est unique du fait que les écailles sont isolées, détachées des os sous-jacents. Une étude de la littérature montre que l isolation des écailles de la carapace peut résulter de deux processus : la mue des anciennes couches de l écaille ou la désarticulation post mortem. Nous favorisons l hypothèse de la désarticulation car les écailles entières sont plus épaisses et plus à même de préserver l ornementation détaillée présente sur les écailles de Stonesfield. Introduction The Stonesfield Slate is well known as the type locality of the first scientifically described dinosaur, Megalosaurus (Buckland 1824). This is also one of the richest Middle Jurassic terrestrial reptile localities in Great Britain (Evans & Milner 1994; Benton & Spencer 1995). Although most of the taxa from Stonesfield have been well studied, the fact that the Stonesfield Slate has yielded the oldest known British turtle is often overlooked. Owen (1842: 160) was the first to notice impressions of turtle epidermal scales from the Stonesfield Slate. Blake (1863) confirmed the chelonian nature of these impressions and provided a short general description based on specimens in the British Museum (now housed in the Natural History Museum, London), without indicating which ones he had in hand. In the same issue of The Geologist, Mackie (1863) described an isolated coracoid from this locality that he attributed to a new species, Chelys(?) blakii Mackie, Later, Phillips (1871: 182) proposed the new species Testudo stricklandi Phillips, 1871 for epidermal scales from the Stonesfield Slate housed at the Oxford University Museum. In his Catalogue of Fossil Reptilia and Amphibia, Lydekker (1889: 220) coined the new genus name Protochelys for T. stricklandi and provisionally referred the isolated coracoid described by Mackie (1863) to this form (although he did not change P. stricklandi to P. blakii as he should have according to the Principle of Priority; ICZN 1999). Few authors mention this material in the 20th century. Romer (1956, 1966) and Bergounioux (1955) list Protochelys, which they tentatively assign to the Pleurosternidae, but do not discuss it. Evans & Milner (1994) include Protochelys in their account of the Middle Jurassic microvertebrate assemblages from the British Isles without reassessing the material. 332
3 Reassessment of the Middle Jurassic turtle from Stonesfield Benton & Spencer (1995) also mention the turtle from Stonesfield, but cast doubt on the chelonian affinities of the scale impressions. Since Lydekker (1889), the turtle material from the Stonesfield Slate has been completely overlooked. Moreover, this material has never been properly described or compared, despite the fact that turtles are extremely rare in Middle Jurassic deposits. The purpose of this paper is to reassess the available material of Protochelys in order to clarify the taxonomic status and possible affinities of this turtle. The remains are herein described and compared for the first time and a tentative reconstruction of the carapace is proposed. Moreover, the Stonesfield scales are an example of exceptional fossilisation and, consequently, are worthy of a detailed taphonomic discussion. Geological settings The Stonesfield Slate was extracted from a localised series of mines and quarries that lie within 1 km of the village of Stonesfield, Oxfordshire, England (Boneham & Wyatt 1993: fig. 1). The slates were exploited as roofing stones (tiles) between the 17th and the early 20th centuries (Aston 1974). The Stonesfield Slate consists of fine, calcareous sandstones and siltstones that are locally interbedded with thin and fissile laminae of ooliths (Boneham & Wyatt 1993). The Stonesfield Slate was previously considered to be a member of the Taynton Limestone Formation or the underlying Sharp s Hill Formation. More recently, Boneham & Wyatt (1993) demonstrated that the Stonesfield Slate represents a sporadic, recurrent lithofacies of the Taynton Limestone Formation. Based on the ammonite fauna, both the Taynton Limestone Formation and the Stonesfield Slate are referred to the Procerites progracilis Biozone, which corresponds to the lower part of the middle Bathonian (Torrens 1980; Boneham & Wyatt 1993). Turtle remains are also known from other British Bathonian localities, especially Kirtlington and Cladach a Ghlinne (Gillham 1994; Evans et al. 2006; Anquetin 2007; Scheyer & Anquetin 2008), but these localities are late Bathonian in age (Evans & Milner 1994). The Stonesfield Slate has yielded a mixed assemblage of marine and terrestrial taxa: marine invertebrates (ammonites, belemnites, bivalves, gastropods, crustaceans, etc.), terrestrial plants, insects, fish, marine and terrestrial reptiles, and mammals (Evans & Milner 1994; Benton & Spencer 1995). This association suggests a deposit in a shallow inshore marine environment. Stonesfield quickly became famous with the discovery of fossil mammals and reptiles. Mammals are represented by two small jaws, of two different species, discovered in 1812 which are still the oldest unquestionable crown-group mammals (Rowe 1999). Reptiles include marine crocodiles (steneosaurids), plesiosaurs, ichthyosaurs, pterosaurs (rhamphorhynchoids), dinosaurs (including the famous Megalosaurus), and turtles (Evans & Milner 1994; Benton & Spencer 1995). Fossil vertebrates occur in the three different lithofacies of the Stonesfield Slate (sandstones, siltstones and oolith laminae), without particular differences in faunal composition. The bone preservation is generally good but the material is disarticulated and was probably transported a short distance (Benton & Spencer 1995). However, the presence of wellpreserved fragile elements like turtle epidermal scales (see below) and terrestrial plants suggests that transport was gentle. Material Apart from two isolated bones (a coracoid and a plastron fragment), all of the Stonesfield turtle specimens consist of unassociated, isolated carapacial scales (no plastral scale has been identified). The Stonesfield scales are unique in being the first isolated fossil turtle scales known to date (see Discussion). Interestingly, these scales are not imprints, as described by 19th century authors, but thin fossilised layers picked out by iron staining or other mineralizations, in the same way as the majority of terrestrial plant remains from the same locality (Cleal & Rees 2003). Palaeobotanists use the term impression or compression-impression to designate this type of preservation (Shute & Cleal 1987; Cleal & Rees 2003). At Stonesfield, turtle scales are often represented by an association 333
 Protochelys blakii (Mackie, 18")
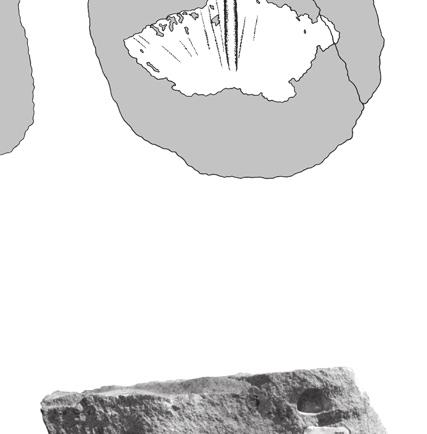
4 Anquetin J. & Claude J. A B Systematic palaeontology Testudinata Klein, 1760 (sensu Joyce et al. 2004) Protochelys blakii (Mackie, 1863) nomen dubium C Fig. 1. Specimens from the Stonesfield Slate (UK) misidentified as chelonian: A, BMNH R896, specimen identified by Lydekker (1889: 222) as a scapulo-precoracoid, but more probably an archosaur cervical rib; B, OUMNH J29907, fish scale labelled as a turtle scale; C, OUMNH unnumbered, specimen (two slabs) labelled as a turtle bony plate which consists of a splinter of compact bone maybe from a crocodilian osteoderm. Scale bars: 10 mm. (two slabs) of the fossil scale itself (representing the original morphology of the scale; i.e. growth rings are thin grooves) and an external mould (representing a mould of the external surface of the scale; i.e. growth rings are low ridges). The nomenclature for shell elements follows Zangerl (1969). Turtle shell scales usually possess growth rings (or growth annuli), though these are not always well pronounced. Generally, these rings indicate that growth was not equal in all directions. For carapacial scales mediolateral growth is faster laterally (or medially for marginals), whereas anteroposterior growth is faster anteriorly. This heterogeneous growth is common among turtles and allows isolated scales to be orientated. We used this characteristic to orientate the Stonesfield carapacial scales: the embryonic scale is at the posteromedial corner of the scale, except for marginals for which it is generally at the posterolateral corner of the scale. Abbreviations BMNH The Natural History Museum, London; OUMNH oxford University Museum of Natural History, Oxford. Chelys(?) blakii Mackie, 1863: 41, fig. 1. Testudo stricklandi Phillips, 1871: 182, diagram 41. Protochelys stricklandi Lydekker 1889: 220. Holotype. BMNH 37979; an isolated, slightly crushed right coracoid, figured in Mackie (1863) [holotype by monotypy]. Referred specimens. BMNH 37218, complete vertebral (two slabs); BMNH 37218a, complete vertebral; BMNH R247, half vertebral (two slabs); BM NH R247a, half vertebral; BMNH 37218b, almost complete vertebral (due to a misreading of the original labelling, the external mould has been erroneously numbered BMNH 39198b after Lydekker [1889]; the correct number is used herein); BMNH R247b, complete pleural (two slabs); BMNH 39198, external mould of a pleural; BMNH 39198a, external mould of a small incomplete scale; BMNH R5320, isolated fragment of plastron; OUMNH J40407, complete vertebral; OUMNH J37067, complete small vertebral; OUMNH J J77376, complete vertebral (two slabs, both figured in Phillips [1871]); OUMNH J77377, external mould of a small vertebral; OUMNH J77378, external mould of a half vertebral. Horizon and age. Stonesfield Slate, Taynton Limestone Formation, Stonesfield, Oxfordshire, England. The Stonesfield Slate is attributed to the lower part of the middle Bathonian (Torrens 1980; Boneham & Wyatt 1993). Remarks Due to the nature of the specimens, no satisfactory diagnostic characters can be identified. Comparisons with other taxa are also limited (see below). At least as far as the scales are concerned, the consistency of the morphology and preservation suggests that they only represent one taxon. There is no support for the separation of two taxa (one based on the coracoid, the other on the scales) and this would not improve the current taxonomic situation. So, it is sensible to treat all the remains as one species, whose affinities are unknown. 334

5 Reassessment of the Middle Jurassic turtle from Stonesfield Lydekker (1889: 222) referred to Protochelys the specimen BMNH R896 (Fig. 1A), which he interpreted as a chelonian scapulo-precoracoid. In fact, this specimen is not chelonian. It shows pneumatic features and may be interpreted as a possible archosaur cervical rib. The collections of the OUMNH also contain several other specimens misidentified as chelonian: OUMNH J29907 (a fish scale; Fig. 1B) and an unnumbered specimen that consists of a splinter of compact bone (maybe from the base of a crocodilian osteoderm; Fig. 1C). Description Coracoid BMNH 37979, as identified by Carter Blake in Mackie (1863), is a right coracoid presented in dorsal view (Fig. 2A). The bone is elongate and slightly crushed. The proximal head is notably enlarged laterally to form the articular glenoid. Medially, the upper part of the sutural surface with the scapula is clearly visible. Posteriorly to the proximal head, the main body of the bone is thin and was probably cylindrical before crushing. Distally, the coracoid expands into a dorsoventrally flattened blade that is characteristic of many turtles. Its posterior margin is broken. The bone may have been concave dorsally but this could be the result of deformation. Plastron BMNH R5320 is the only turtle shell bone from the Stonesfield Slate. This is a plastron fragment (hyo- or hypoplastron) probably of the bridge area (Fig. 2B). Three spiny projections are present like those that can be found in embryonic or juvenile individuals or in turtles that do not have a fully ossified shell at adult size. The size of the specimen (40 mm in maximum length) indicates that it does not belong to a hatchling turtle, but it may have belonged to a young individual. First vertebral scale BMNH 37218b (Fig. 3A, B), OUMNH J J77376 (Fig. 3C, D; see also Phillips 1871: 182, fig , 11) and OUMNH J77377 (Fig. 3E) are interpreted as first vertebrals because of their symmetrical, pentagonal shape and their concave A B Fig. 2. Turtle bones from the Stonesfield Slate (UK), middle Bathonian: A, BMNH 37979, right coracoid in dorsal view (proximal to the right) figured in Mackie (1863); B, BMNH R5320, plastron fragment (hyo- or hypoplastron) probably from the bridge area. Scale bars: 10 mm. posterior margin. The bilateral symmetry is underlined by a strong medial keel that is stronger anteriorly and ends abruptly just before reaching the posterior margin of the scale. These scales are almost 335
6 Anquetin J. & Claude J. twice as wide as they are long. It is likely that the second vertebral overlapped the first, as suggested by the presence of a smooth triangular area on the posterior part of the first vertebral and by the disappearance of the medial keel just anterior to this area (Fig. 3C, D). The anterior margin of the first vertebral is convex and longer than the posterior margin. The long lateral margin faces posterolaterally and contacts the first pleural. On BMNH 37218b, growth rings are poorly preserved in some areas of the scale. A few anteriorly radiating ridges are present on the anteromedial part of the scale. On OUMNH J J77376, growth rings and radiating ridges are well preserved. OUMNH J77377 is poorly preserved, but some anteriorly radiating ridges are still visible. Second vertebral scale BMNH (Fig. 3F, G), OUMNH J77378 (Fig. 3H) and OUMNH J37067 (Fig. 3I) are interpreted as second vertebrals because of their symmetrical, hexagonal shape and their anterior margin shorter than the posterior margin (generally, vertebrals 2-4 are hexagonal in outline and differ from each other in the relative development of their anterior and posterior margins). These scales have a straight or slightly concave anterior margin and straight posterior margin. They are twice as wide as long (OUMNH J77378 is incomplete) and have a medial keel. The anterolateral margin contacts the posteromedial margin of the first pleural and faces anterolaterally. The posterolateral margin contacts the anteromedial margin of the second pleural and extends parallel to the anteroposterior axis of the scale. The anterolateral margin is slightly shorter than the posterolateral one. The medial keel protrudes anteriorly from the anterior margin of the scale. The posterior margin presents a medial emargination that probably corresponds to the anterior protrusion of the medial keel of the third vertebral (see below). On BMNH 37218, growth rings are only slightly apparent medially but are still well defined laterally. Some rings are deeply marked but, between those, thinner rings are often present. The spacing between major growth rings is irregular. A few poorly defined anteriorly radiating ridges are apparent laterally. On OUMNH J77378, growth rings are better preserved, no intercalated thin rings are present, and the spacing between growth rings is relatively regular. An anteriorly radiating pattern covers the whole surface of the scale. On OUMNH J37067, the presence of the medial ridge is uncertain and the majority of growth rings are badly marked (this specimen is poorly preserved). A few anteriorly radiating ridges are present on the anteromedial and posterolateral portions of this specimen. Third vertebral scale BMNH 37218a (Fig. 3J) and BMNH R247a (Fig. 3K) are interpreted as third vertebrals because of their symmetrical, hexagonal shape and their anterior and posterior margins equal in length. Both specimens exhibit a medial keel that is wider and more pronounced anteriorly. The scale is twice as wide as long with sub-straight anterior and posterior margins. The medial keel protrudes anteriorly from the anterior margin of the scale, whereas a slight medial emargination of the posterior margin probably corresponds to the anterior protrusion of the fourth vertebral. Both lateral margins are oblique with respect to the anteroposterior axis: the anterolateral margin contacts the posteromedial margin of the second pleural, whereas the posterolateral margin contacts the anteromedial margin of the third pleural. The two lateral margins are equal in length. The surface ornamentation of BMNH 37218a is poorly preserved, but growth rings Fig. 3. Chelonian first to fourth vertebral scales, Stonesfield Slate (UK), middle Bathonian: A-E, first vertebral; F-I, second vertebral; J, K, third vertebral; L, M, fourth vertebral; A, B, BMNH 37218b, almost complete first vertebral (B is the external mould); C, D, OUMNH J J77376, complete first vertebral (D is the external mould OUMNH J77376) figured by Phillips (1871); E, OUMNH J77377, badly preserved external mould of a first vertebral; F, G, BMNH 37218, complete second vertebral (G is the external mould); H, OUMNH J77378, external mould of the right side of a second vertebral; I, OUMNH J37067, badly preserved small second vertebral; J, BMNH 37218a, complete third vertebral; K, BMNH R247a, left half of a third vertebral; L, M, BMNH R247, left side of a fourth vertebral (M is the external mould). Scale bars: 10 mm. 336





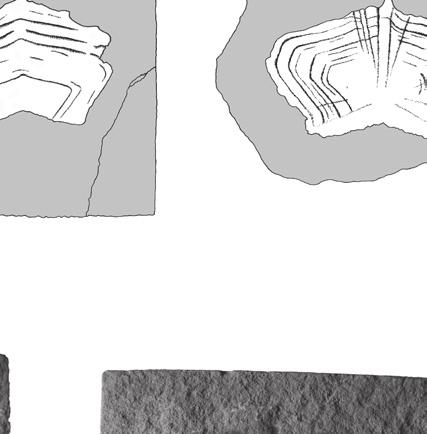









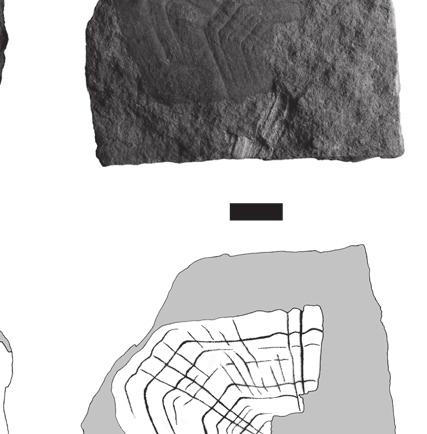
7 Reassessment of the Middle Jurassic turtle from Stonesfield A B F J C E D G H K L I M 337
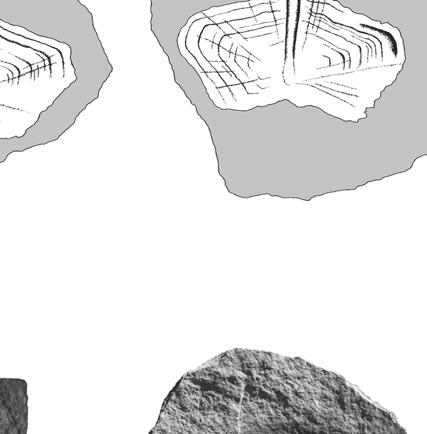
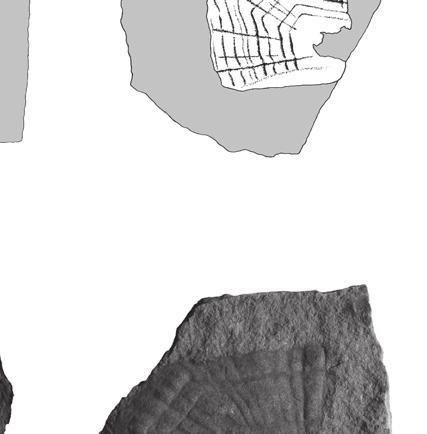
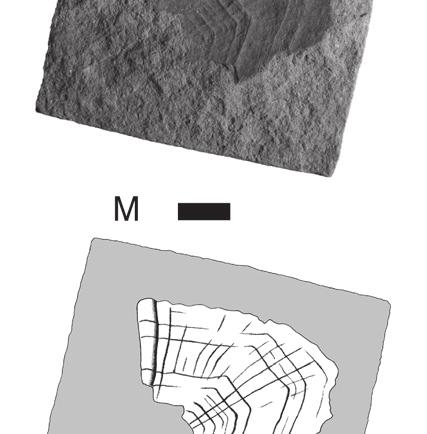
8 Anquetin J. & Claude J. are still well preserved on BMNH R247a. On the latter, thinner (less marked) rings are intercalated between the rather regularly spaced major rings. No radiating pattern is observable, but it should be noted that these specimens are less well preserved than the previously described scales. Fourth vertebral scale BMNH R247 (Fig. 3L, M) is interpreted as a fourth vertebral because of its (original) symmetrical, hexagonal shape and its anterior margin wider than the posterior margin. The right side of the scale and the former emplacement of the embryonic scale are missing. The anterior part of the medial keel is visible on the broken side of the scale. The anterior margin is slightly convex, whereas the posterior margin is slightly concave. Both lateral margins are oblique with respect to the anteroposterior axis: the anterolateral margin contacts the posteromedial margin of the third pleural, whereas the posterolateral margin contacts the anteromedial margin of the fourth pleural. The anterolateral margin is notably shorter than the posterolateral margin. The original scale was approximately twice as wide as long. Due to the preservation, it is unclear whether or not the medial keel protrudes anteriorly, but since a slight medial emargination is present on the posterior margin of the third vertebral (see above) it is probable that it did. Growth rings are relatively well defined and thinner rings are often intercalated between major rings. The spacing between major growth rings is irregular. A pattern of anteriorly radiating ridges is present. Fifth vertebral scale OUMNH J40407 (Fig. 4A) and BMNH 39198a (Fig. 4B) are interpreted as fifth vertebrals because of their symmetrical, pentagonal shape and their convex posterior margin. OUMNH J40407 has a well-developed medial keel that is wider anteriorly than posteriorly. On BMNH 39198a, an inflexion present on each growth ring near to the broken side of the scale indicates the presence of a keel although this one is not apparent on the specimen (compare with OUMNH J40407). For both specimens, both the anterior and posterior margins are convex, which differs from the condition in other vertebrals. The anterior margin contacts the posterior margin of the fourth vertebral. The long and arched posterior part of the scale in fact presents two separate margins that correspond to the contact with the two last marginals (probably eleventh and twelfth) at the posterior end of the carapace. The short lateral margin faces anterolaterally and corresponds to the contact with the posteromedial margin of the fourth pleural. The scale is wider than long, but proportionally less wide than vertebrals 2-4. Growth rings are well preserved on OUMNH J40407, although only slightly defined in some areas. The spacing between major growth rings is irregular and a few anteriorly radiating ridges are present anteromedially. By comparison, only major growth rings are preserved on BMNH 39198a and no trace of a radiating pattern is present. Pleural scales BMNH R247b (Fig. 4C, D) is interpreted as a right third pleural because of its non-symmetrical, squarish shape, its straight anterior and posterior margins, its oblique medial margins, and its short posteromedial margin. The scale is slightly longer than wide. The anterior margin is as long as the posterior. On the arched lateral margin, three distinct contacts corresponding to three different marginals can be observed. Growth rings are well preserved, especially on the external mould. Major rings are irregularly spaced and up to three or four less pronounced rings can be seen between some of them. There is no trace of radiating ridges on the scale. BMNH (Fig. 4E 1 ) is interpreted as a first or fourth pleural because of its non-symmetrical and irregular pentagonal shape. The original outline of the scale, revealed by growth rings, allows two possible interpretations of this specimen: a right fourth pleural (Fig. 4E 2 ) or a left first pleural (Fig. 4E 3 ). The following observations weaken the second hypothesis: first pleurals are usually more elongate; growth rate should have been greater anteriorly; and the posterior margin that contacts the second pleural should be sub-perpendicular to the anteroposterior axis of the scale. Consequently, we favour an interpretation of BMNH as a right fourth pleural. The scale is longer than wide. 338
, middle Bathonian: A, B, fifth vertebral; C-E, pleural scales; A, OUMNH J40407, complete fifth vertebral; B, BMNH 39198a, external")
9 Reassessment of the Middle Jurassic turtle from Stonesfield A B C D E 1 E 2 E 3 P3 V4 V1 M M M M M M V5 V2 P2 Fig. 4. Chelonian fifth vertebral and pleural scales, Stonesfield Slate (UK), middle Bathonian: A, B, fifth vertebral; C-E, pleural scales; A, OUMNH J40407, complete fifth vertebral; B, BMNH 39198a, external mould of the right side of a fifth vertebral; C, D, BMNH R247b, complete right third pleural (D is the external mould); E, BMNH 39198, external mould of an almost complete scale (E 1, non oriented) which can be interpreted either as a right fourth pleural (E 2, preferred hypothesis) or as a left first pleural (E 3 ) (the specimen is an external mould and needs to be reversed to retrieve the normal orientation). Abbreviations: M, marginal scale; P, pleural scale; V, vertebral scale. Scale bars: 10 mm (Figures E 2 and E 3 are not to scale). The anteromedial margin (that contacts the posterolateral margin of the fourth vertebral) is damaged and consequently the anterior margin (that contacts the posterior margin of the third pleural) appears shorter than it was originally. The short posteromedial margin corresponds to the contact with the lateral margin of the fifth vertebral. The lateral margin is divided into three parts corresponding to the contact with three different marginals. Growth rings are relatively well preserved. Major rings are irregularly spaced. Few less pronounced rings are present but difficult to distinguish. There is no radiating pattern on the scale. Comparison It is difficult to compare the turtle scales from the Stonesfield Slate with other taxa because no directly comparable material (i.e. fossil epidermal scales) is known. When describing bony shells, systematists can only access information on the general outlines of scales and mainly use their relative sizes for diagnostic purposes. At Stonesfield, although they match each other fairly well (Fig. 5A), the scales are isolated and obviously pertain to different individuals: they show different growth rates, different sizes and there is no indication that all of the scales come from the same stratigraphical horizon. So that the relative sizes of the Stonesfield scales cannot be assessed. However, the detailed description provided herein enables some comparisons to be made. A tentative reconstruction of the carapace of the Stonesfield turtle is provided in Figure 5B. An important characteristic of this turtle is that vertebrals are wider than long, whereas pleurals are slightly 339
10 Anquetin J. & Claude J. longer than wide. This is a rather primitive scheme commonly found in stem turtles and basal crowngroup turtles. More derived turtles tend to have vertebrals that are longer than wide and pleurals that are wider than long. Among turtles with wide vertebrals and narrow pleurals, the Stonesfield turtle more closely resembles the stem turtles Kayentachelys aprix Gaffney, Hutchison, Jenkins & Meeker, 1987 (Gaffney et al. 1987), Indochelys spatulata Datta, Manna, Ghosh & Das, 2000 (Datta et al. 2000), Heckerochelys romani Sukhanov, 2006 (Sukhanov 2006) and Kallokibotion bajazidi Nopcsa, 1923 (Gaffney & Meylan 1992), the panpleurodire Notoemys laticentralis Cattoi & Freiberg, 1961 (Fernandez & Fuente 1994; Lapparent de Broin et al. 2007), and the pleurosternid Dinochelys whitei Gaffney, 1979 (Gaffney 1979; Brinkman et al. 2000). The aforementioned species share the following features with the Stonesfield turtle: large vertebral 1 with a convex anterior margin that is at least as wide as the posterior margin; vertebral 2 with a posterolateral margin more or less parallel to the anteroposterior axis of the shell; vertebral 4 with a significantly short anterolateral margin; vertebral 5 wider than long with an anterior margin as wide as the posterior margin. The Stonesfield turtle differs from each of these species by several of its characters. Vertebrals 2-4 of Kayentachelys and Indochelys are wider than those of the Stonesfield turtle, whereas they are narrower in Kallokibotion. In Kayentachelys, vertebral 1 is at least three times wider than long and has three distinct margins anteriorly (one for each marginal 1 and one for the cervical scale). Vertebral 1 of Dinochelys and Notoemys is also wider than that of the Stonesfield turtle. In Kallokibotion, vertebral 1 is narrower and more elongated than that of the Stonesfield turtle and its lateral margin is parallel to the anteroposterior axis of the shell. In Notoemys, the lateral margin of vertebral 1 are also parallel to the anteroposterior axis of the shell and all intervertebral sulci are straight. Dinochelys also has straight intervertebral sulci. Heckerochelys differs from the Stonesfield turtle in having a longer vertebral 2, vertebral 3 with strongly concave posterior margin and vertebral 4 with only one lateral margin. Concerning the scale ornamentation, none of the aforementioned species is known to have one, although it is possible that scale ornamentation does not always leave a trace on the bone plates of the shell. One can notice that the anterior margin of vertebrals 2-5 of Kayentachelys presents a welldefined, short medial protrusion similar to that of the Stonesfield turtle. Other turtles known to have a scale ornamentation of anteriorly radiating ridges on the vertebrals (e.g., Proganochelys, Platychelys, Euryaspis, Desmemys, some plesiochelyids and chengyuchelyids) do not match the morphology of the Stonesfield turtle. Romer (1956, 1966) and Bergounioux (1955) referred the turtle from Stonesfield to the Pleurosternidae. Pleurosternon bullockii (Owen, 1842), Glyptops plicatulus (Cope, 1877) and Glyptops typocardium (Seeley, 1869) (Gaffney 1979; Milner 2004) all have vertebrals that are reduced in width and pleurals wider than long. Among pleurosternids, only the Late Jurassic Dinochelys whitei has a scale pattern somewhat similar to that of the Stonesfield turtle. However, as it is also true for other species ranging from Early Jurassic stem turtles to basal panpleurodires (see above), one can only assume that this scale pattern is plesiomorphic. In other words, a referral to the pleurosternids is not better supported than a referral to the stem Testudines or the Panpleurodira in the current state of knowledge. Taphonomy The preservation of turtle epidermal scales is rather uncommon in the fossil record. We are aware of only two other cases: a shell fragment from the Pleistocene of NE Thailand (J. Claude pers. obs.) and a shell of Neurankylus from the early Paleocene of New Mexico (Sullivan et al. 1988). In the later case, the scales are preserved on approximately 85% of the carapace, are rich in iron and manganese, and are apparently preserved with the original colour pattern (Sullivan et al. 1988: fig. 3). In both cases, the scales are preserved as a thin layer on the surface of the bony shell, but the Stonesfield specimens are the only known example of isolated scales in the fossil record. Isolated epidermal scales can result from two processes: shedding during growth or post mortem disarticulation. To our knowledge, no detailed studies of the post-hatching development of shell scales have 340
11 Reassessment of the Middle Jurassic turtle from Stonesfield been published. In a recent review on the use of growth rings to estimate turtle age, Wilson et al. (2003) discuss this lack of literature on epidermal scale growth. Two patterns are seen in extant turtles (Wilson et al. 2003: fig. 1). In some turtles (e.g., Terrapene carolina (Linnaeus, 1758)), scales do not shed and old layers are retained on the external surface of new ones. In such cases, growth rings correspond to the superposition of successively larger and younger scales. Such scales become thicker with age, although old layers are often progressively worn by abrasion. In other turtles (e.g., Trachemys, Orlitia, Chrysemys), the old scale is shed while the new one grows beneath it. Although the mechanism is not yet understood, the old layers leave an impression, corresponding to the growth ring, on the new scale. Shed keratinous scale layers are thin, translucent and flexible, and are consequently unlikely to fossilise. Moreover, we were unable to find any reference mentioning the persistence of shed scale layers in the environment, which might suggest that they are quickly destroyed after shedding. The isolated nature of Stonesfield scales probably results from disarticulation rather than shedding: shed scales are thinner and more fragile than complete scales, and are therefore less likely to fossilise. Moreover, shed scales would probably not display such marked growth rings. Indeed, extant shedding turtles tend to have smooth epidermal scales with poorly developed rings. Turtle taphonomy is poorly studied and the few available studies deal primarily with bones, which is little help in case of the Stonesfield material. Frustratingly, studies on extant species often fail to mention epidermal scales at all (see Brand et al for a more detailed review). However, among these neontological studies, Bourn & Coe (1979) reported the disarticulation sequence of Geochelone gigantea (Schweigger, 1812) (the Aldabra tortoise) and stated that, at some point in the sequence, scales detach themselves from each other and from the bony layer before falling. Dodd (1995) documented the disarticulation patterns of 80 turtle carcasses, representing three families (emydids, testudinids and trionychids), in a terrestrial sandhill habitat in North Florida. As noted by Bourn & Coe (1979), Dodd (1995) A B Fig. 5. Reconstruction of the Stonesfield turtle, middle Bathonian: A, reconstruction based on available specimens (note that these specimens are not from the same individuals nor at the same scale); B, proposed reconstruction of the carapace of the Stonesfield turtle. The morphology of the first and second pleural (dashed lines) can be deduced from that of neighbouring scales. Marginals are unknown at Stonesfield and therefore are not represented on the present reconstruction. 341
12 Anquetin J. & Claude J. also observed that the keratinous scales of emydids and testudinids eventually detached themselves individually from the shell (trionychids have no scales but a leathery skin covering the shell). He noted that the vertebrals and pleurals generally detach simultaneously, or the former closely followed by the latter. Marginals are often the first group to detach, although they persist longer than the vertebrals and pleurals in the testudinid Gopherus polyphemus (Daudin, 1802). Dodd (1995: 383) observed that disarticulated scales may remain close to the carcass for an extended period. Brand et al. (2003) carried out an experimental study to compare the disarticulation processes for the common slider turtle (Trachemys scripta (Schoepff, 1792)) in different environments. They observed that separation between scales and shell bones occurred at the same time as shell disarticulation. Unfortunately, the detached elements were removed immediately from the experimental area so that there was no estimation of the time that disarticulated scales could remain in the environment. Thus, taphonomic studies on turtles have not so far explained the fate of scales once detached or shed. For example, it is unknown how long keratinous scales can remain in the environment, how they are affected by transport or what conditions may be conducive to their preservation. The Stonesfield specimens are the only isolated fossil turtle scales known to date, though it is possible that others exist and have been misinterpreted or remain unrecognised and undescribed. The preservation of isolated keratinous elements is unusual in the fossil record and probably requires particular burial environments and transport conditions. In addition, there are currently no detailed taphonomical studies of the Stonesfield biota in general that could explained the preservation of these scales, although similarities of preservation with the plant remains from the same locality are striking (Cleal & Rees 2003). Conclusions A reassessment of the available material from the Middle Jurassic Stonesfield Slate shows that the turtle previously known as Protochelys Lydekker, 1889 cannot be properly diagnosed from other taxa nor placed within a phylogenetic framework. Protochelys blakii (Mackie, 1863) is consequently considered nomen dubium, until more evidences are found. This situation is a direct consequence of the nature of the preserved specimens. The Stonesfield turtle presents a plesiomorphic scale pattern with vertebrals that are twice as wide as long, pleurals that are reduced in width and a fifth vertebral with an anterior margin as wide as its posterior margin. These features are commonly found in stem turtles and some basal crown-group turtles. Any phylogenetic assignment of this turtle is then difficult to achieve, although the combination of these primitive features suggests a basal phylogenetic position. Although the Stonesfield turtle scales have a limited systematic value, they are important in terms of taphonomy. These remains are the only known example of isolated fossil turtle scales. The Stonesfield scales probably result from disarticulation processes as such scales are thicker, more resistant and more likely to preserve well-developed ornamentation (growth rings and radiating ridges) than shed scale layers. Now that fossil turtle scales have been described and figured extensively for the first time, it will be easier for future workers to recognise them. In the same time, this study also enlightens the fact that taphonomic literature generally overlooks the fate of shed scale layers and disarticulated scales. This should be taken into consideration by future taphonomic studies on turtles. Acknowledgements The authors thank Sandra Chapman and Angela Milner (BMNH), and Paul Jeffery (OUMNH) for access to collections in their care. The help from Andrew Ross (BMNH) and Lionel Cavin (Muséum d Histoire naturelle, Geneva) at the beginning of the study was deeply appreciated. Our gratitude also goes to Paul Barrett (BMNH), Guillaume Billet and Peggy Vincent (both at the Muséum national d Histoire naturelle, Paris) for their comments on earlier versions of the manuscript. Igor Danilov (Russian Academy of Sciences, Saint Petersburg) and an anonymous reviewer provided helpful comments. This is publication ISEM of J.C. 342
13 Reassessment of the Middle Jurassic turtle from Stonesfield References Anquetin J A new (stem?) turtle from the Middle Jurassic of Skye, Scotland. Journal of Vertebrate Paleontology 27 (3): 41A. Aston M Stonesfield Slate. Oxfordshire County Council, Department of Museum Services, Publication 5, 86 p. Benton M. J. & Spencer P. S Fossil Reptiles of Great Britain. Chapman and Hall, London, 386 p. Bergounioux F. M Testudinata, in Piveteau J. (ed.), Traité de paléontologie. Volume 5. Masson et Cie, Paris: Blake C. C On chelonian scutes from the Stonesfield Slate. The Geologist 6: Boneham B. F. W. & Wyatt R. J The stratigraphical position of the Middle Jurassic (Bathonian) Stonesfield Slate of Stonesfield, Oxfordshire, UK. Proceedings of the Geologists Association 104 (2): Bourn D. & Coe M. J Features of tortoise mortality and decomposition on Aldabra. Philosophical Transactions of the Royal Society of London, series B, Biological Science 286: Brand L. R., Hussey M. & Taylor J Taphonomy of freshwater turtles: decay and disarticulation in controlled experiments. Journal of Taphonomy 1 (4): Brinkman D. B., Stadtman K. & Smith D New material of Dinochelys whitei Gaffney, 1979, from the Dry Mesa Quarry (Morrison Formation, Jurassic) of Colorado. Journal of Vertebrate Paleontology 20 (2): Buckland W Notice on the Megalosaurus, or great fossil lizard of Stonesfield. Transactions of the Geological Society of London 1 (2): Cleal C. J. & Rees P. M The Middle Jurassic flora from Stonesfield, Oxfordshire, UK. Palaeontology 46 (4): Datta P. M., Manna P., Ghosh S. C. & Das D. P The first Jurassic turtle from India. Palaeontology 43 (1): Dodd C. K. Jr Disarticulation of turtle shells in North-central Florida: how long does a shell remain in the woods? American Midland Naturalist 134: Evans S. E. & Milner A. R Middle Jurassic microvertebrate assemblages from the British Isles, in Fraser N. C. & Sues H.-D. (eds), In the Shadow of the Dinosaurs Early Mesozoic Tetrapods. Cambridge University Press, Cambridge: Evans S. E., Barrett P. M., Hilton J., Butler R. J., Jones M. E. H., Liang M.-M., Parish J. C., Rayfield E. J., Sigogneau-Russell D. & Underwood C. J The Middle Jurassic vertebrate Assemblage of Skye, Scotland, in Barrett P. M. & Evans S. E. (eds), Ninth International Symposium on Mesozoic Terrestrial Ecosystems and Biota, Abstracts and Proceedings. The Natural History Museum, London: Fernandez M. S. & Fuente M. S. de la Redescription and phylogenetic position of Notoemys: the oldest Gondwanian pleurodiran turtle. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 193 (1): Gaffney E. S The Jurassic turtles of North America. Bulletin of the American Museum of Natural History 162 (3): Gaffney E. S. & Meylan P. A The Transylvanian turtle, Kallokibotion, a primitive cryptodire of Cretaceous age. American Museum Novitates 3040: Gaffney E. S., Hutchison J. H., Jenkins F. A. & Meeker L. J Modern turtle origins: the oldest known cryptodire. Science 237: Gillham C A fossil turtle (Reptilia: Chelonia) from the Middle Jurassic of Oxfordshire, England. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 10: ICZN International Code of Zoological Nomenclature. Fourth edition. International Trust for Zoological Nomenclature, London, 306 p. Joyce W. G., Parham J. F. & Gauthier J. A Developing a protocol for the conversion of rankbased taxon names to phylogenetically defined clade names, as exemplified by turtles. Journal of Paleontology 78 (5): Lapparent de Broin F. de, Fuente M. S. de la & Fernandez M. S Notoemys laticentralis (Chelonii, Pleurodira), Late Jurassic of Argentina: new examination of the anatomical structures and comparisons. Revue de Paléobiologie 26 (2): Lydekker R Catalogue of the Fossil Reptilia and Amphibia in the British Museum (Natural History) Part III. The Order Chelonia. Trustees of the British Museum, London, 239 p. Mackie S. J Turtles in the Stonesfield Slate. The Geologist 6: Milner A. R The turtles of the Purbeck Limestone Group of Dorset, Southern England. Palaeontology 47 (6): Owen R Report on British fossil reptiles, part 2. Report for the British Association for the Advancement of Science 11 (Plymouth 1841): Phillips J Geology of Oxford and the Valley of the Thames. Clarendon Press, Oxford, 523 p. Romer A. S Osteology of the Reptiles. University of Chicago Press, Chicago, 772 p. Romer A. S Vertebrate Paleontology, 3rd edition. University of Chicago Press, Chicago, 468 p. Rowe T At the roots of the mammalian family tree. Nature 398: Scheyer T. M. & Anquetin J Bone histology of the Middle Jurassic turtle shell remains from 343
14 Anquetin J. & Claude J. Kirtlington, Oxfordshire, England. Lethaia 41 (1): Shute C. H. & Cleal C. J Palaeobotany in museums. Geological Curator 4: Sukhanov V. B An archaic turtle, Heckerochelys romani gen. et sp. nov., from the Middle Jurassic of Moscow region, Russia, in Danilov I. G. & Parham J. F. (eds), Fossil Turtle Research. Volume 1. Russian Journal of Herpetology 13 (suppl.): Sullivan R. M., Lucas S. G., Hunt A. P. & Fritts T. H Color pattern on the selmacryptodiran turtle Neurankylus from the early Paleocene (Puercan) of the San Juan Basin, New Mexico. Contributions in Science, Natural History Museum of Los Angeles County 401: 1-9. Torrens H. S Bathonian correlation chart, in Cope J. C. W. (ed.), A Correlation of Jurassic Rocks in the British Isles, Part 2. Geological Society, London, special report 15: Wilson D. S., Tracy C. R. & Tracy C. R Estimating age of turtles from growth rings: a critical evaluation of the technique. Herpetologica 59 (2): Zangerl R The turtle shell, in Gans C., Bellairs A. & Parsons T. S. (eds), Biology of the Reptilia. Volume 1. Academic Press, London; New York: Submitted on 19 July 2007; accepted on 13 March
TRACHEMYS SCULPTA. A nearly complete articulated carapace and plastron of an Emjdd A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE,
 A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A.
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
SUPPLEMENTARY INFORMATION
 In comparison to Proganochelys (Gaffney, 1990), Odontochelys semitestacea is a small turtle. The adult status of the specimen is documented not only by the generally well-ossified appendicular skeleton
In comparison to Proganochelys (Gaffney, 1990), Odontochelys semitestacea is a small turtle. The adult status of the specimen is documented not only by the generally well-ossified appendicular skeleton
REVISION OF REDONDASUCHUS (ARCHOSAURIA: AETOSAURIA) FROM THE UPPER TRIASSIC REDONDA FORMATION, NEW MEXICO, WITH DESCRIPTION OF A NEW SPECIES
 FROM THE UPPER TRIASSIC REDONDA FORMATION, NEW MEXICO, WITH DESCRIPTION OF A NEW SPECIES") Harris et al., eds., 2006, The Triassic-Jurassic Terrestrial Transition. New Mexico Museum of Natural History and Science Bulletin 37. REVISION OF REDONDASUCHUS (ARCHOSAURIA: AETOSAURIA) FROM THE UPPER
Harris et al., eds., 2006, The Triassic-Jurassic Terrestrial Transition. New Mexico Museum of Natural History and Science Bulletin 37. REVISION OF REDONDASUCHUS (ARCHOSAURIA: AETOSAURIA) FROM THE UPPER
Title: Phylogenetic Methods and Vertebrate Phylogeny
 Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Lower Cretaceous Kwanmon Group, Northern Kyushu
 Bull. Kitakyushu Mus. Nat. Hist., 11: 87-90. March 30, 1992 A New Genus and Species of Carnivorous Dinosaur from the Lower Cretaceous Kwanmon Group, Northern Kyushu Yoshihiko Okazaki Kitakyushu Museum
Bull. Kitakyushu Mus. Nat. Hist., 11: 87-90. March 30, 1992 A New Genus and Species of Carnivorous Dinosaur from the Lower Cretaceous Kwanmon Group, Northern Kyushu Yoshihiko Okazaki Kitakyushu Museum
Vol. XIV, No. 1, March, The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S.
 By S.") Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
A new species of sauropod, Mamenchisaurus anyuensis sp. nov.
 A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
Origin and Evolution of Birds. Read: Chapters 1-3 in Gill but limited review of systematics
 Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
FOSSIL TURTLE RESEARCH
 FOSSIL TURTLE RESEARCH VOLUME 1 Proceedings of the Symposium on Turtle Origins, Evolution and Systematics August 18 20, 2003, St. Petersburg, Russia Edited by Igor G. Danilov and James F. Parham St. Petersburg,
FOSSIL TURTLE RESEARCH VOLUME 1 Proceedings of the Symposium on Turtle Origins, Evolution and Systematics August 18 20, 2003, St. Petersburg, Russia Edited by Igor G. Danilov and James F. Parham St. Petersburg,
A R T I C L E S STRATIGRAPHIC DISTRIBUTION OF VERTEBRATE FOSSIL FOOTPRINTS COMPARED WITH BODY FOSSILS
 A R T I C L E S STRATIGRAPHIC DISTRIBUTION OF VERTEBRATE FOSSIL FOOTPRINTS COMPARED WITH BODY FOSSILS Leonard Brand & James Florence Department of Biology Loma Linda University WHAT THIS ARTICLE IS ABOUT
A R T I C L E S STRATIGRAPHIC DISTRIBUTION OF VERTEBRATE FOSSIL FOOTPRINTS COMPARED WITH BODY FOSSILS Leonard Brand & James Florence Department of Biology Loma Linda University WHAT THIS ARTICLE IS ABOUT
LOWER CRETACEOUS AGE FROM VICTORIA, AUSTRALIA
 Memoirs of the National Museum of Victoria 9 April 1969 https://doi.org/10.24199/j.mmv.1969.29.02 A FOSSIL CHELONIAN OF PROBABLE LOWER CRETACEOUS AGE FROM VICTORIA, AUSTRALIA By J. W. Warren Department
Memoirs of the National Museum of Victoria 9 April 1969 https://doi.org/10.24199/j.mmv.1969.29.02 A FOSSIL CHELONIAN OF PROBABLE LOWER CRETACEOUS AGE FROM VICTORIA, AUSTRALIA By J. W. Warren Department
LOWER CRETACEOUS OF SOUTH DAKOTA.
 A NEW DINOSAUR, STP^GOSAURUS MARSHl, FROM THE LOWER CRETACEOUS OF SOUTH DAKOTA. By Frederic A. Lucas, Curator, Divisioii of Coiiipnrative Anatomy, in charge, of Section of Vertebrate Fossils. The name
A NEW DINOSAUR, STP^GOSAURUS MARSHl, FROM THE LOWER CRETACEOUS OF SOUTH DAKOTA. By Frederic A. Lucas, Curator, Divisioii of Coiiipnrative Anatomy, in charge, of Section of Vertebrate Fossils. The name
290 SHUFELDT, Remains of Hesperornis.
 290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
v:ii-ixi, 'i':;iisimvi'\>!i-:: "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi L I E) R.ARY OF THE VERSITY U N I or ILLINOIS REMO
 R.ARY OF THE VERSITY U N I or ILLINOIS REMO") "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
"^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
RATE OF SCUTE ANNULI DEPOSITION OF EASTERN BOX TURTLES (TERRAPENE CAROLINA CAROLINA) HELD IN CAPTIVITY AND IN THEIR NATURAL HABITAT
 HELD IN CAPTIVITY AND IN THEIR NATURAL HABITAT") Ana Maria Caputo December 4, 2007 RATE OF SCUTE ANNULI DEPOSITION OF EASTERN BOX TURTLES (TERRAPENE CAROLINA CAROLINA) HELD IN CAPTIVITY AND IN THEIR NATURAL HABITAT Eastern box turtles (terappene Carolina
Ana Maria Caputo December 4, 2007 RATE OF SCUTE ANNULI DEPOSITION OF EASTERN BOX TURTLES (TERRAPENE CAROLINA CAROLINA) HELD IN CAPTIVITY AND IN THEIR NATURAL HABITAT Eastern box turtles (terappene Carolina
35. DATA REPORT: CRETACEOUS OSTRACODES FROM HOLES 865A AND 866A (MID-PACIFIC MOUNTAINS) 1. Renée Damotte 2
 1. Renée Damotte 2") Winterer, E.L., Sager, W.W., Firth, J.V., and Sinton, J.M. (Eds.), 1995 Proceedings of the Ocean Drilling Program, Scientific Results, Vol. 143 35. DATA REPORT: CRETACEOUS OSTRACODES FROM HOLES 865A AND
Winterer, E.L., Sager, W.W., Firth, J.V., and Sinton, J.M. (Eds.), 1995 Proceedings of the Ocean Drilling Program, Scientific Results, Vol. 143 35. DATA REPORT: CRETACEOUS OSTRACODES FROM HOLES 865A AND
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
FIELDIANA GEOLOGY NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA
 FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
Origin and Evolution of Birds. Read: Chapters 1-3 in Gill but limited review of systematics
 Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
(Proc. Acad. Nat. Sci. Phila., p. I62) for the reception of his earlier. Chisternon. Article JX.-ON TWO INTERESTING GENERA OF EOCENE
 for the reception of his earlier. Chisternon. Article JX.-ON TWO INTERESTING GENERA OF EOCENE") 56.81,3(ii81 :78.7) Article JX.-ON TWO INTERESTING GENERA OF EOCENE TURTLES, CHISTERNON LEIDY AND ANOSTEIRA LEIDY. By OLIVER P. HAY. The genus Chisternon was proposed in I872 by Dr. Joseph Leidy (Proc.
56.81,3(ii81 :78.7) Article JX.-ON TWO INTERESTING GENERA OF EOCENE TURTLES, CHISTERNON LEIDY AND ANOSTEIRA LEIDY. By OLIVER P. HAY. The genus Chisternon was proposed in I872 by Dr. Joseph Leidy (Proc.
Anatomy. Name Section. The Vertebrate Skeleton
 Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Preliminary results on the stratigraphy and taphonomy of multiple bonebeds in the Triassic of Algarve
 Preliminary results on the stratigraphy and taphonomy of multiple bonebeds in the Triassic of Algarve Hugo Campos 1,2*, Octávio Mateus 1,2, Miguel Moreno-Azanza 1,2 1 Faculdade de Ciências e Tecnologia,
Preliminary results on the stratigraphy and taphonomy of multiple bonebeds in the Triassic of Algarve Hugo Campos 1,2*, Octávio Mateus 1,2, Miguel Moreno-Azanza 1,2 1 Faculdade de Ciências e Tecnologia,
THE TURTLES OF THE PURBECK LIMESTONE GROUP OF DORSET, SOUTHERN ENGLAND
 THE TURTLES OF THE PURBECK LIMESTONE GROUP OF DORSET, SOUTHERN ENGLAND by ANDREW R. MILNER ABSTRACT. The turtles from the Purbeck Limestone are revised and it is concluded that there are four shell-based
THE TURTLES OF THE PURBECK LIMESTONE GROUP OF DORSET, SOUTHERN ENGLAND by ANDREW R. MILNER ABSTRACT. The turtles from the Purbeck Limestone are revised and it is concluded that there are four shell-based
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE
 FROM THE") A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported
 Drawing of early ankle development of Alligator mississippiensis, as reported") Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported by a previous study 1. The intermedium is formed at
Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported by a previous study 1. The intermedium is formed at
Mesozoic Marine Life Invertebrate Vertebrate
 Mesozoic Marine Life Invertebrate Vertebrate Cenozoic Marine Life - Invertebrates (Mollusks) Cenozoic Marine Life - Invertebrates (Arthropods) Cenozoic Marine Life - Vertebrates Marine fossils are abundant
Mesozoic Marine Life Invertebrate Vertebrate Cenozoic Marine Life - Invertebrates (Mollusks) Cenozoic Marine Life - Invertebrates (Arthropods) Cenozoic Marine Life - Vertebrates Marine fossils are abundant
Today there are approximately 250 species of turtles and tortoises.
 I WHAT IS A TURTLE OR TORTOISE? Over 200 million years ago chelonians with fully formed shells appeared in the fossil record. Unlike modern species, they had teeth and could not withdraw into their shells.
I WHAT IS A TURTLE OR TORTOISE? Over 200 million years ago chelonians with fully formed shells appeared in the fossil record. Unlike modern species, they had teeth and could not withdraw into their shells.
New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia
 1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
New Late Jurassic turtle skulls from Switzerland and the diversity of the Plesiochelyidae
 New Late Jurassic turtle skulls from Switzerland and the diversity of the Plesiochelyidae Jérémy Anquetin Section d archéologie et paléontologie Office de la culture République et Canton du Jura Porrentruy,
New Late Jurassic turtle skulls from Switzerland and the diversity of the Plesiochelyidae Jérémy Anquetin Section d archéologie et paléontologie Office de la culture République et Canton du Jura Porrentruy,
Introduction. Edwin A. Cadena, Carlos A. Jaramillo, and Jonathan I. Bloch
 Chapter 8 New Material of the Platychelyid Turtle Notoemys zapatocaensis from the Early Cretaceous of Colombia; Implications for Understanding Pleurodira Evolution Edwin A. Cadena, Carlos A. Jaramillo,
Chapter 8 New Material of the Platychelyid Turtle Notoemys zapatocaensis from the Early Cretaceous of Colombia; Implications for Understanding Pleurodira Evolution Edwin A. Cadena, Carlos A. Jaramillo,
Red Eared Slider Secrets. Although Most Red-Eared Sliders Can Live Up to Years, Most WILL NOT Survive Two Years!
 Although Most Red-Eared Sliders Can Live Up to 45-60 Years, Most WILL NOT Survive Two Years! Chris Johnson 2014 2 Red Eared Slider Secrets Although Most Red-Eared Sliders Can Live Up to 45-60 Years, Most
Although Most Red-Eared Sliders Can Live Up to 45-60 Years, Most WILL NOT Survive Two Years! Chris Johnson 2014 2 Red Eared Slider Secrets Although Most Red-Eared Sliders Can Live Up to 45-60 Years, Most
The Fossil Record of the Diamond-backed Terrapin, Malaclemys terrapin (Testudines: Emydidae)
") Journal of Herpetology, Vol. 46, No. 3, 351 355, 2012 Copyright 2012 Society for the Study of Amphibians and Reptiles The Fossil Record of the Diamond-backed Terrapin, Malaclemys terrapin (Testudines:
Journal of Herpetology, Vol. 46, No. 3, 351 355, 2012 Copyright 2012 Society for the Study of Amphibians and Reptiles The Fossil Record of the Diamond-backed Terrapin, Malaclemys terrapin (Testudines:
UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA
 FROM JAVA, INDONESIA") NOTES AND NEWS UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA BY NGUYEN NGOC-HO i) Faculty of Science, University of Saigon, Vietnam Among material recently collected
NOTES AND NEWS UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA BY NGUYEN NGOC-HO i) Faculty of Science, University of Saigon, Vietnam Among material recently collected
BREVIORA LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1
, A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1") ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
Video Assignments. Microraptor PBS The Four-winged Dinosaur Mark Davis SUNY Cortland Library Online
 Video Assignments Microraptor PBS The Four-winged Dinosaur Mark Davis SUNY Cortland Library Online Radiolab Apocalyptical http://www.youtube.com/watch?v=k52vd4wbdlw&feature=youtu.be Minute 13 through minute
Video Assignments Microraptor PBS The Four-winged Dinosaur Mark Davis SUNY Cortland Library Online Radiolab Apocalyptical http://www.youtube.com/watch?v=k52vd4wbdlw&feature=youtu.be Minute 13 through minute
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL
 Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
LEIDY, SHOWING THE BONES OF THE FEET 'AND LIMBS
 CQNTEUBUTIONS FBOM THE MUSEUM OF PALEONTOLOGY (Confindion of Con&&&m froin UB Muaercm of Gcologg) UNIVERSITY OF ' MICHIGAN VOL V, No. 6, pp. 6W3 (e ph.) DEAXMBER 31,1036 A SPECIMEN OF STYLEMYS NEBRASCENSIS
CQNTEUBUTIONS FBOM THE MUSEUM OF PALEONTOLOGY (Confindion of Con&&&m froin UB Muaercm of Gcologg) UNIVERSITY OF ' MICHIGAN VOL V, No. 6, pp. 6W3 (e ph.) DEAXMBER 31,1036 A SPECIMEN OF STYLEMYS NEBRASCENSIS
ON SOME REPTILIAN REMAINS FROM THE DINOSAUR BEDS OF NYASALAND. By S. H. HAUGHTON, D.Sc., F.G.S.
 ( 67 ) ON SOME REPTILIAN REMAINS FROM THE DINOSAUR BEDS OF NYASALAND. By S. H. HAUGHTON, D.Sc., F.G.S. (Published by permission of the Hon. the Minister for Mines and Industries.) (With Plates II-V and
( 67 ) ON SOME REPTILIAN REMAINS FROM THE DINOSAUR BEDS OF NYASALAND. By S. H. HAUGHTON, D.Sc., F.G.S. (Published by permission of the Hon. the Minister for Mines and Industries.) (With Plates II-V and
SOME NEW AMERICAN PYCNODONT FISHES.
 SOME NEW AMERICAN PYCNODONT FISHES. By James Williams Gidley, Assistant Curator of Fossil Mammals, United States National Museum. In the United States National Museum are several specimens representing
SOME NEW AMERICAN PYCNODONT FISHES. By James Williams Gidley, Assistant Curator of Fossil Mammals, United States National Museum. In the United States National Museum are several specimens representing
The family Gnaphosidae is a large family
 Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
d a Name Vertebrate Evolution - Exam 2 1. (12) Fill in the blanks
 Fill in the blanks") Vertebrate Evolution - Exam 2 1. (12) Fill in the blanks 100 points Name f e c d a Identify the structures (for c and e, identify the entire structure, not the individual elements. b a. b. c. d. e. f.
Vertebrate Evolution - Exam 2 1. (12) Fill in the blanks 100 points Name f e c d a Identify the structures (for c and e, identify the entire structure, not the individual elements. b a. b. c. d. e. f.
Erycine Boids from the Early Oligocene of the South Dakota Badlands
 Georgia Journal of Science Volume 67 No. 2 Scholarly Contributions from the Membership and Others Article 6 2009 Erycine Boids from the Early Oligocene of the South Dakota Badlands Dennis Parmley J. Alan
Georgia Journal of Science Volume 67 No. 2 Scholarly Contributions from the Membership and Others Article 6 2009 Erycine Boids from the Early Oligocene of the South Dakota Badlands Dennis Parmley J. Alan
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1
 Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
Biology Slide 1 of 50
 Biology 1 of 50 2 of 50 What Is a Reptile? What are the characteristics of reptiles? 3 of 50 What Is a Reptile? What Is a Reptile? A reptile is a vertebrate that has dry, scaly skin, lungs, and terrestrial
Biology 1 of 50 2 of 50 What Is a Reptile? What are the characteristics of reptiles? 3 of 50 What Is a Reptile? What Is a Reptile? A reptile is a vertebrate that has dry, scaly skin, lungs, and terrestrial
Animal Diversity III: Mollusca and Deuterostomes
 Animal Diversity III: Mollusca and Deuterostomes Objectives: Be able to identify specimens from the main groups of Mollusca and Echinodermata. Be able to distinguish between the bilateral symmetry on a
Animal Diversity III: Mollusca and Deuterostomes Objectives: Be able to identify specimens from the main groups of Mollusca and Echinodermata. Be able to distinguish between the bilateral symmetry on a
PRELIMINARY REPORT ON A CLUTCH OF SIX DINOSAURIAN EGGS FROM THE UPPER TRIASSIC ELLIO T FORMATION, NORTHERN ORANGE FREE STATE. J. W.
 41 Pa/aeont. afr., 22, 41-45 (1979) PRELIMINARY REPORT ON A CLUTCH OF SIX DINOSAURIAN EGGS FROM THE UPPER TRIASSIC ELLIO T FORMATION, NORTHERN ORANGE FREE STATE b y J. W. Kitching ABSTRACT A clutch of
41 Pa/aeont. afr., 22, 41-45 (1979) PRELIMINARY REPORT ON A CLUTCH OF SIX DINOSAURIAN EGGS FROM THE UPPER TRIASSIC ELLIO T FORMATION, NORTHERN ORANGE FREE STATE b y J. W. Kitching ABSTRACT A clutch of
AMERICAN MUSEUM NOVITATES Published by
 AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia
 from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia") First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia Tsogtbaatar Chinzorig¹, ³ *, Yoshitsugu Kobayashi², Khishigjav Tsogtbaatar³,
First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia Tsogtbaatar Chinzorig¹, ³ *, Yoshitsugu Kobayashi², Khishigjav Tsogtbaatar³,
From Slime to Scales: Evolution of Reptiles. Review: Disadvantages of Being an Amphibian
 From Slime to Scales: Evolution of Reptiles Review: Disadvantages of Being an Amphibian Gelatinous eggs of amphibians cannot survive out of water, so amphibians are limited in terms of the environments
From Slime to Scales: Evolution of Reptiles Review: Disadvantages of Being an Amphibian Gelatinous eggs of amphibians cannot survive out of water, so amphibians are limited in terms of the environments
click for previous page SEA TURTLES
 click for previous page SEA TURTLES FAO Sheets Fishing Area 51 TECHNICAL TERMS AND PRINCIPAL MEASUREMENTS USED head width (Straight-line distances) head prefrontal precentral carapace central (or neural)
click for previous page SEA TURTLES FAO Sheets Fishing Area 51 TECHNICAL TERMS AND PRINCIPAL MEASUREMENTS USED head width (Straight-line distances) head prefrontal precentral carapace central (or neural)
New Mexico Geological Society
 New Mexico Geological Society Downloaded from: http://nmgs.nmt.edu/publications/guidebooks/56 Vertebrate fauna of the Upper Triassic Mesa Montosa Member (Petrified Forest Formation, Chinle Group), Chama
New Mexico Geological Society Downloaded from: http://nmgs.nmt.edu/publications/guidebooks/56 Vertebrate fauna of the Upper Triassic Mesa Montosa Member (Petrified Forest Formation, Chinle Group), Chama
A skull without mandihle, from the Hunterian Collection (no.
 4 MR. G. A. BOULENGER ON CHELONIAN REMAINS. [Jan. 6, 2. On some Chelonian Remains preserved in the Museum of the Eojal College of Surgeons. By G. A. Boulenger. [Eeceived December 8, 1890.] In the course
4 MR. G. A. BOULENGER ON CHELONIAN REMAINS. [Jan. 6, 2. On some Chelonian Remains preserved in the Museum of the Eojal College of Surgeons. By G. A. Boulenger. [Eeceived December 8, 1890.] In the course
Juehuaornis gen. nov.
 34 1 2015 3 GLOBAL GEOLOGY Vol. 34 No. 1 Mar. 2015 1004 5589 2015 01 0007 05 Juehuaornis gen. nov. 1 1 1 2 1. 110034 2. 110034 70% Juehuaornis zhangi gen. et sp. nov Q915. 4 A doi 10. 3969 /j. issn. 1004-5589.
34 1 2015 3 GLOBAL GEOLOGY Vol. 34 No. 1 Mar. 2015 1004 5589 2015 01 0007 05 Juehuaornis gen. nov. 1 1 1 2 1. 110034 2. 110034 70% Juehuaornis zhangi gen. et sp. nov Q915. 4 A doi 10. 3969 /j. issn. 1004-5589.
A new species of Calamagras Cope, 1873 (Serpentes, Boidae, Erycinae) from the early Eocene of Kirghizia
 from the early Eocene of Kirghizia") A new species of Calamagras Cope, 1873 (Serpentes, Boidae, Erycinae) from the early Eocene of Kirghizia Igor G. DANILOV Zoological Institute of the Russian Academy of Sciences, Universitetskaya nab. 1,
A new species of Calamagras Cope, 1873 (Serpentes, Boidae, Erycinae) from the early Eocene of Kirghizia Igor G. DANILOV Zoological Institute of the Russian Academy of Sciences, Universitetskaya nab. 1,
8/19/2013. Topic 4: The Origin of Tetrapods. Topic 4: The Origin of Tetrapods. The geological time scale. The geological time scale.
 Topic 4: The Origin of Tetrapods Next two lectures will deal with: Origin of Tetrapods, transition from water to land. Origin of Amniotes, transition to dry habitats. Topic 4: The Origin of Tetrapods What
Topic 4: The Origin of Tetrapods Next two lectures will deal with: Origin of Tetrapods, transition from water to land. Origin of Amniotes, transition to dry habitats. Topic 4: The Origin of Tetrapods What
Cladistics (reading and making of cladograms)
") Cladistics (reading and making of cladograms) Definitions Systematics The branch of biological sciences concerned with classifying organisms Taxon (pl: taxa) Any unit of biological diversity (eg. Animalia,
Cladistics (reading and making of cladograms) Definitions Systematics The branch of biological sciences concerned with classifying organisms Taxon (pl: taxa) Any unit of biological diversity (eg. Animalia,
NEW CRETACEOUS AND CENOZOIC FOSSIL TURTLES FROM COLOMBIA AND PANAMA; SYSTEMATIC PALEONTOLOGY, PHYLOGENETICAL AND PALEOBIOGEOGRAPHICAL IMPLICATIONS
 NEW CRETACEOUS AND CENOZOIC FOSSIL TURTLES FROM COLOMBIA AND PANAMA; SYSTEMATIC PALEONTOLOGY, PHYLOGENETICAL AND PALEOBIOGEOGRAPHICAL IMPLICATIONS By EDWIN ALBERTO CADENA RUEDA A THESIS PRESENTED TO THE
NEW CRETACEOUS AND CENOZOIC FOSSIL TURTLES FROM COLOMBIA AND PANAMA; SYSTEMATIC PALEONTOLOGY, PHYLOGENETICAL AND PALEOBIOGEOGRAPHICAL IMPLICATIONS By EDWIN ALBERTO CADENA RUEDA A THESIS PRESENTED TO THE
8/19/2013. Topic 5: The Origin of Amniotes. What are some stem Amniotes? What are some stem Amniotes? The Amniotic Egg. What is an Amniote?
 Topic 5: The Origin of Amniotes Where do amniotes fall out on the vertebrate phylogeny? What are some stem Amniotes? What is an Amniote? What changes were involved with the transition to dry habitats?
Topic 5: The Origin of Amniotes Where do amniotes fall out on the vertebrate phylogeny? What are some stem Amniotes? What is an Amniote? What changes were involved with the transition to dry habitats?
Bio 1B Lecture Outline (please print and bring along) Fall, 2006
 Fall, 2006") Bio 1B Lecture Outline (please print and bring along) Fall, 2006 B.D. Mishler, Dept. of Integrative Biology 2-6810, bmishler@berkeley.edu Evolution lecture #4 -- Phylogenetic Analysis (Cladistics) -- Oct.
Bio 1B Lecture Outline (please print and bring along) Fall, 2006 B.D. Mishler, Dept. of Integrative Biology 2-6810, bmishler@berkeley.edu Evolution lecture #4 -- Phylogenetic Analysis (Cladistics) -- Oct.
Living Dinosaurs (3-5) Animal Demonstrations
 Animal Demonstrations") Living Dinosaurs (3-5) Animal Demonstrations At a glance Students visiting the zoo will be introduced to live animals and understand their connection to a common ancestor, dinosaurs. Time requirement One
Living Dinosaurs (3-5) Animal Demonstrations At a glance Students visiting the zoo will be introduced to live animals and understand their connection to a common ancestor, dinosaurs. Time requirement One
TWO NEW SPECIES OF WATER MITES FROM OHIO 1-2
 TWO NEW SPECIES OF WATER MITES FROM OHIO 1-2 DAVID R. COOK Wayne State University, Detroit, Michigan ABSTRACT Two new species of Hydracarina, Tiphys weaveri (Acarina: Pionidae) and Axonopsis ohioensis
TWO NEW SPECIES OF WATER MITES FROM OHIO 1-2 DAVID R. COOK Wayne State University, Detroit, Michigan ABSTRACT Two new species of Hydracarina, Tiphys weaveri (Acarina: Pionidae) and Axonopsis ohioensis
A basal eucryptodiran turtle Sinemys efremovi (= Wuguia efremovi) from the Early Cretaceous of China
 from the Early Cretaceous of China") A basal eucryptodiran turtle Sinemys efremovi (= Wuguia efremovi) from the Early Cretaceous of China IGOR G. DANILOV and VLADIMIR B. SUKHANOV Danilov, I.G. and Sukhanov,V.B. 2006. A basal eucryptodiran
A basal eucryptodiran turtle Sinemys efremovi (= Wuguia efremovi) from the Early Cretaceous of China IGOR G. DANILOV and VLADIMIR B. SUKHANOV Danilov, I.G. and Sukhanov,V.B. 2006. A basal eucryptodiran
Amniote Relationships. Reptilian Ancestor. Reptilia. Mesosuarus freshwater dwelling reptile
 Amniote Relationships mammals Synapsida turtles lizards,? Anapsida snakes, birds, crocs Diapsida Reptilia Amniota Reptilian Ancestor Mesosuarus freshwater dwelling reptile Reptilia General characteristics
Amniote Relationships mammals Synapsida turtles lizards,? Anapsida snakes, birds, crocs Diapsida Reptilia Amniota Reptilian Ancestor Mesosuarus freshwater dwelling reptile Reptilia General characteristics
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS
 AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.
UNIVERSITY OF MICHIGAN MONTANA
 CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY UNIVERSITY OF MICHIGAN VOL. VIII, No. 4, pp. 43-58 (1 PI., 4 figs.) M~Y 31, 1950 A NEW TESTUDO FROM MADISON COUNTY, MONTANA BY THOMAS M. OELRICH UNIVERSITY
CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY UNIVERSITY OF MICHIGAN VOL. VIII, No. 4, pp. 43-58 (1 PI., 4 figs.) M~Y 31, 1950 A NEW TESTUDO FROM MADISON COUNTY, MONTANA BY THOMAS M. OELRICH UNIVERSITY
The Lower Jaws of Baenid Turtles
 AMERICAN MUSEUM Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N.Y. 10024 Number 2749, pp. 1-10, figs. 1-4, table 1 September 27, 1982 The Lower
AMERICAN MUSEUM Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N.Y. 10024 Number 2749, pp. 1-10, figs. 1-4, table 1 September 27, 1982 The Lower
 Oct. 2017 ACTA GEOLOGICA SINICA (English Edition) Vol. 91 No. 5 1529 http://www.geojournals.cn/dzxben/ch/index.aspx of Yumenerpeton and that of all the other bystrowianids. On the other hand, the primitive
Oct. 2017 ACTA GEOLOGICA SINICA (English Edition) Vol. 91 No. 5 1529 http://www.geojournals.cn/dzxben/ch/index.aspx of Yumenerpeton and that of all the other bystrowianids. On the other hand, the primitive
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * Dr. L.D. Boonstra. Paleontologist, South African Museum, Cape Town
 THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
Reptiles. Ectothermic vertebrates Very successful Have scales and toenails Amniotes (lay eggs with yolk on land) Made up of 4 orders:
 Made up of 4 orders:") Reptiles of Florida Reptiles Ectothermic vertebrates Very successful Have scales and toenails Amniotes (lay eggs with yolk on land) Made up of 4 orders: Crocodylia (alligators & crocodiles) Squamata (amphisbaenids
Reptiles of Florida Reptiles Ectothermic vertebrates Very successful Have scales and toenails Amniotes (lay eggs with yolk on land) Made up of 4 orders: Crocodylia (alligators & crocodiles) Squamata (amphisbaenids
A NEW LINDHOLMEMYDID GENUS (TESTUDINES: LINDHOLMEMYDIDAE) FROM THE MID-CRETACEOUS OF UZBEKISTAN
 FROM THE MID-CRETACEOUS OF UZBEKISTAN") Russian Journal of Herpetology Vol. 6, No. 1, 1999, pp. 63 71 A NEW LINDHOLMEMYDID GENUS (TESTUDINES: LINDHOLMEMYDIDAE) FROM THE MID-CRETACEOUS OF UZBEKISTAN Igor G. Danilov 1 Submitted December 15, 1998.
Russian Journal of Herpetology Vol. 6, No. 1, 1999, pp. 63 71 A NEW LINDHOLMEMYDID GENUS (TESTUDINES: LINDHOLMEMYDIDAE) FROM THE MID-CRETACEOUS OF UZBEKISTAN Igor G. Danilov 1 Submitted December 15, 1998.
These small issues are easily addressed by small changes in wording, and should in no way delay publication of this first- rate paper.
 Reviewers' comments: Reviewer #1 (Remarks to the Author): This paper reports on a highly significant discovery and associated analysis that are likely to be of broad interest to the scientific community.
Reviewers' comments: Reviewer #1 (Remarks to the Author): This paper reports on a highly significant discovery and associated analysis that are likely to be of broad interest to the scientific community.
.56 m. (22 in.). COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition
. COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition") Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
BEHAVIORAL AND PALEOENVIRONMENTAL IMPLICATIONS OF REPTILE SWIM TRACKS FROM THE EARLY TRIASSIC OF WESTERN NORTH AMERICA
 Tracy Thomson attended the College of Eastern Utah and then received his B.Sc. in geology from the University of Utah. He is currently attending the University of California-Riverside and Dr. Mary Droser
Tracy Thomson attended the College of Eastern Utah and then received his B.Sc. in geology from the University of Utah. He is currently attending the University of California-Riverside and Dr. Mary Droser
Dinosaurs and Dinosaur National Monument
 Page 1 of 6 Dinosaurs and Dinosaur National Monument The Douglass Quarry History of Earl's Excavation... Geology of the Quarry Rock Formations and Ages... Dinosaur National Monument protects a large deposit
Page 1 of 6 Dinosaurs and Dinosaur National Monument The Douglass Quarry History of Earl's Excavation... Geology of the Quarry Rock Formations and Ages... Dinosaur National Monument protects a large deposit
PEABODY MUSEUM OF NATURAL HISTORY, YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 85 September 21, 1964 A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA STANLEY J. RIEL
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 85 September 21, 1964 A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA STANLEY J. RIEL
In North America 1. the Triassic is represented by the thick Newark Group along the east coast, 2. by widespread red-bed and fluvial sediments in the
 The Triassic System The name Triassic derives from the three parts into which the Triassic is divided on the European platform: 3. Keuper (highest) 2. Muschelkalk 1. Bunter (lowest) In North America 1.
The Triassic System The name Triassic derives from the three parts into which the Triassic is divided on the European platform: 3. Keuper (highest) 2. Muschelkalk 1. Bunter (lowest) In North America 1.
NAUSHONIA PAN AMEN SIS, NEW SPECIES (DECAPODA: THALASSINIDEA: LAOMEDIIDAE) FROM THE PACIFIC COAST OF PANAMA, WITH NOTES ON THE GENUS
 FROM THE PACIFIC COAST OF PANAMA, WITH NOTES ON THE GENUS") 5 October 1982 PROC. BIOL. SOC. WASH. 95(3), 1982, pp. 478-483 NAUSHONIA PAN AMEN SIS, NEW SPECIES (DECAPODA: THALASSINIDEA: LAOMEDIIDAE) FROM THE PACIFIC COAST OF PANAMA, WITH NOTES ON THE GENUS Joel
5 October 1982 PROC. BIOL. SOC. WASH. 95(3), 1982, pp. 478-483 NAUSHONIA PAN AMEN SIS, NEW SPECIES (DECAPODA: THALASSINIDEA: LAOMEDIIDAE) FROM THE PACIFIC COAST OF PANAMA, WITH NOTES ON THE GENUS Joel
complex in cusp pattern. (3) The bones of the coyote skull are thinner, crests sharper and the
 The bones of the coyote skull are thinner, crests sharper and the") DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
A NEW PLIOCENE FOSSIL CRAB OF THE GENUS (Trichopeltarion) FROM NEW ZEALAND
 FROM NEW ZEALAND") De/i & I f f n 8 t 0 * of Orustac^ A NEW PLIOCENE FOSSIL CRAB OF THE GENUS (Trichopeltarion) FROM NEW ZEALAND by R. K. DELL Dominion Museum, Wellington, New Zealand ABSTRACT A new Pliocene species of Trichopeltarion
De/i & I f f n 8 t 0 * of Orustac^ A NEW PLIOCENE FOSSIL CRAB OF THE GENUS (Trichopeltarion) FROM NEW ZEALAND by R. K. DELL Dominion Museum, Wellington, New Zealand ABSTRACT A new Pliocene species of Trichopeltarion
Diurus, Pascoe. sp. 1). declivity of the elytra, but distinguished. Length (the rostrum and tails 26 included) mm. Deep. exception
. declivity of the elytra, but distinguished. Length (the rostrum and tails 26 included) mm. Deep. exception") 210 DIURUS ERYTIIROPUS. NOTE XXVI. Three new species of the Brenthid genus Diurus, Pascoe DESCRIBED BY C. Ritsema+Cz. 1. Diurus erythropus, n. sp. 1). Allied to D. furcillatus Gylh. ²) by the short head,
210 DIURUS ERYTIIROPUS. NOTE XXVI. Three new species of the Brenthid genus Diurus, Pascoe DESCRIBED BY C. Ritsema+Cz. 1. Diurus erythropus, n. sp. 1). Allied to D. furcillatus Gylh. ²) by the short head,
INDIVIDUAL IDENTIFICATION OF GREEN TURTLE (CHELONIA MYDAS) HATCHLINGS
 HATCHLINGS") INDIVIDUAL IDENTIFICATION OF GREEN TURTLE (CHELONIA MYDAS) HATCHLINGS Ellen Ariel, Loïse Corbrion, Laura Leleu and Jennifer Brand Report No. 15/55 Page i INDIVIDUAL IDENTIFICATION OF GREEN TURTLE (CHELONIA
INDIVIDUAL IDENTIFICATION OF GREEN TURTLE (CHELONIA MYDAS) HATCHLINGS Ellen Ariel, Loïse Corbrion, Laura Leleu and Jennifer Brand Report No. 15/55 Page i INDIVIDUAL IDENTIFICATION OF GREEN TURTLE (CHELONIA
Three new species of Microctenochira SPAETH from Brazil and Panama (Coleoptera: Chrysomelidae: Cassidinae)
") Genus Vol. 10 (1): 109-116 Wroc³aw, 31 III 1999 Three new species of Microctenochira SPAETH from Brazil and Panama (Coleoptera: Chrysomelidae: Cassidinae) JOLANTA ŒWIÊTOJAÑSKA and LECH BOROWIEC Zoological
Genus Vol. 10 (1): 109-116 Wroc³aw, 31 III 1999 Three new species of Microctenochira SPAETH from Brazil and Panama (Coleoptera: Chrysomelidae: Cassidinae) JOLANTA ŒWIÊTOJAÑSKA and LECH BOROWIEC Zoological
A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev Doklady Akademii Nauk, SSSR 87:
 translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
ONLINE APPENDIX 1. Morphological phylogenetic characters scored in this paper. See Poe (2004) for
 for") ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
Class Reptilia Testudines Squamata Crocodilia Sphenodontia
 Class Reptilia Testudines (around 300 species Tortoises and Turtles) Squamata (around 7,900 species Snakes, Lizards and amphisbaenids) Crocodilia (around 23 species Alligators, Crocodiles, Caimans and
Class Reptilia Testudines (around 300 species Tortoises and Turtles) Squamata (around 7,900 species Snakes, Lizards and amphisbaenids) Crocodilia (around 23 species Alligators, Crocodiles, Caimans and
New Records of Cladocera (Crustacea) for Trinidad, West Indies
 for Trinidad, West Indies") New Records of Cladocera (Crustacea) for Trinidad, West Indies Azad Mohammed Mohammed, A. 2004. A New Records of Cladocera (Crustacea) for Trinidad, West Indies. Living World, Journal of The Trinidad and
New Records of Cladocera (Crustacea) for Trinidad, West Indies Azad Mohammed Mohammed, A. 2004. A New Records of Cladocera (Crustacea) for Trinidad, West Indies. Living World, Journal of The Trinidad and
Do the traits of organisms provide evidence for evolution?
 PhyloStrat Tutorial Do the traits of organisms provide evidence for evolution? Consider two hypotheses about where Earth s organisms came from. The first hypothesis is from John Ray, an influential British
PhyloStrat Tutorial Do the traits of organisms provide evidence for evolution? Consider two hypotheses about where Earth s organisms came from. The first hypothesis is from John Ray, an influential British
A NEW SALTICID SPIDER FROM VICTORIA By R. A. Dunn
 Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae)
") Genus Vol. 14 (3): 413-418 Wroc³aw, 15 X 2003 A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae) JAROS AW KANIA Zoological Institute, University of Wroc³aw, Sienkiewicza
Genus Vol. 14 (3): 413-418 Wroc³aw, 15 X 2003 A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae) JAROS AW KANIA Zoological Institute, University of Wroc³aw, Sienkiewicza
TOPOTYPES OF TYPOTHORAX COCCINARUM, A LATE TRIASSIC AETOSAUR FROM THE AMERICAN SOUTHWEST
 Lucas, S.G. and Spielmann, J.A., eds., 2007, The Global Triassic. New Mexico Museum of Natural History and Science Bulletin 41. TOPOTYPES OF TYPOTHORAX COCCINARUM, A LATE TRIASSIC AETOSAUR FROM THE AMERICAN
Lucas, S.G. and Spielmann, J.A., eds., 2007, The Global Triassic. New Mexico Museum of Natural History and Science Bulletin 41. TOPOTYPES OF TYPOTHORAX COCCINARUM, A LATE TRIASSIC AETOSAUR FROM THE AMERICAN
The Triassic Transition
 The Triassic Transition The Age of Reptiles Begins As the Paleozoic drew to a close through the Carboniferous and Permian several important processes were at work. Assembly of Pangea Evolutionary radiation
The Triassic Transition The Age of Reptiles Begins As the Paleozoic drew to a close through the Carboniferous and Permian several important processes were at work. Assembly of Pangea Evolutionary radiation
Are the dinosauromorph femora from the Upper Triassic of Hayden Quarry (New Mexico) three stages in a growth series of a single taxon?
 three stages in a growth series of a single taxon?") Anais da Academia Brasileira de Ciências (2017) 89(2): 835-839 (Annals of the Brazilian Academy of Sciences) Printed version ISSN 0001-3765 / Online version ISSN 1678-2690 http://dx.doi.org/10.1590/0001-3765201720160583
Anais da Academia Brasileira de Ciências (2017) 89(2): 835-839 (Annals of the Brazilian Academy of Sciences) Printed version ISSN 0001-3765 / Online version ISSN 1678-2690 http://dx.doi.org/10.1590/0001-3765201720160583
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO
 A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes)
") Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
ALFRED GILLETT AND FOSSILS FROM STREET
 ALFRED GILLETT AND FOSSILS FROM STREET This collection of local fossils was formerly in the Crispin Hall, Street. Most of these fossils came from Alfred Gillett (1814-1904), a retired ironmonger who lived
ALFRED GILLETT AND FOSSILS FROM STREET This collection of local fossils was formerly in the Crispin Hall, Street. Most of these fossils came from Alfred Gillett (1814-1904), a retired ironmonger who lived
A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China
 SUPPLEMENTARY INFORMATION A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China Ya-Ming Wang 1, Hai-Lu You 2,3 *, Tao Wang 4 1 School of Earth Sciences and Resources, China
SUPPLEMENTARY INFORMATION A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China Ya-Ming Wang 1, Hai-Lu You 2,3 *, Tao Wang 4 1 School of Earth Sciences and Resources, China
Barney to Big Bird: The Origin of Birds. Caudipteryx. The fuzzy raptor. Solnhofen Limestone, cont d
 Barney to Big Bird: The Origin of Birds Caudipteryx The fuzzy raptor The discovery of feathered dinosaurs in Liaoning, China, has excited the many paleontologists who suspected a direct link between dinosaurs
Barney to Big Bird: The Origin of Birds Caudipteryx The fuzzy raptor The discovery of feathered dinosaurs in Liaoning, China, has excited the many paleontologists who suspected a direct link between dinosaurs
d. Wrist bones. Pacific salmon life cycle. Atlantic salmon (different genus) can spawn more than once.
 can spawn more than once.") Lecture III.5b Answers to HW 1. (2 pts). Tiktaalik bridges the gap between fish and tetrapods by virtue of possessing which of the following? a. Humerus. b. Radius. c. Ulna. d. Wrist bones. 2. (2 pts)
Lecture III.5b Answers to HW 1. (2 pts). Tiktaalik bridges the gap between fish and tetrapods by virtue of possessing which of the following? a. Humerus. b. Radius. c. Ulna. d. Wrist bones. 2. (2 pts)
Biology. Slide 1of 50. End Show. Copyright Pearson Prentice Hall
 Biology 1of 50 2of 50 Phylogeny of Chordates Nonvertebrate chordates Jawless fishes Sharks & their relatives Bony fishes Reptiles Amphibians Birds Mammals Invertebrate ancestor 3of 50 A vertebrate dry,
Biology 1of 50 2of 50 Phylogeny of Chordates Nonvertebrate chordates Jawless fishes Sharks & their relatives Bony fishes Reptiles Amphibians Birds Mammals Invertebrate ancestor 3of 50 A vertebrate dry,