Papers in Press. Papers in Press includes peer-reviewed, accepted manuscripts of research
|
|
|
- Annabella Dickerson
- 5 years ago
- Views:
Transcription

1 Papers in Press Papers in Press includes peer-reviewed, accepted manuscripts of research articles, reviews, and short notes to be published in Paleontological Research. They have not yet been copy edited and/or formatted in the publication style of Paleontological Research. As soon as they are printed, they will be removed from this website. Please note they can be cited using the year of online publication and the DOI, as follows: Humblet, M. and Iryu, Y. 2014: Pleistocene coral assemblages on Irabu-jima, South Ryukyu Islands, Japan. Paleontological Research, doi: /2014PR020.
2 1 doi: /2017pr024 A Turonian polycotylid plesiosaur (Reptilia; Sauropterygia) from Obira Town, Hokkaido, and its biostratigraphic and paleoecological significance TAMAKI SATO 1, TOMOYA HANAI 2, SHOJI HAYASHI 3,4, and TOMOHIRO NISHIMURA 5 1 Department of Astronomy and Earth Sciences, Tokyo Gakugei University, Koganei City, Tokyo , Japan. [tsato@u-gakugei.ac.jp] 2 Department of Earth and Planetary Science, Graduate School of Science, The University of Tokyo, Hongo, Bunkyo-ku, Tokyo Japan. [thanai@eps.s.u-tokyo.ac.jp] 3 Faculty of Biosphere-Geosphere Science, Okayama University of Science, Kita-ku Ridaicho 1-1, Okayama, , Japan. [hayashi@big.ous.ac.jp] 4 Division of Materials and Manufacturing Science, Graduate School of Engineering, Osaka University, 2-1, Yamada-Oka, Suita, Osaka , Japan. 5 Hobetsu Museum, 80-6 Hobetsu, Mukawa Town, Hokkaido Japan.
3 2 Abstract. The Polycotylidae are short-necked plesiosaurs known from the Cretaceous in various parts of the world, but only a few occurrences have been documented in Japan where elasmosaurid remains are much more common. An indeterminate polycotylid specimen from the Upper Cretaceous in Obira Town, Hokkaido, is described. Characteristics of the vertebrae and clavicular arch support its taxonomic affinity. The Turonian occurrence of the specimen indicates the continuous presence of the Polycotylidae across the Cenomanian-Turonian boundary in the northwestern Pacific. Macroscopic osteological features of the vertebrae and clavicular arch indicate an advanced stage of ossification, and there are histological characteristics suggesting slowed growth. The osteoporotic-like condition implies a high degree of aquatic adaption. Key words: Plesiosauria, Polycotylidae, Cenomanian-Turonian boundary, ontogeny, histology
4 3 Introduction The Yezo Group in Japan and Sakhalin is so far the only place in the northern Pacific where a continuous stratigraphic sequence of mid to uppermost Cretaceous is available with a reasonable number of reptilian fossils and good stratigraphic controls (Sato et al., 2012; Takashima et al., 2004; Konishi et al., 2016). Basic taxonomic studies of Yezo fauna are therefore crucial to understand the temporal and geographic distributions of various Cretaceous marine reptiles. Remains of the Plesiosauria (Reptilia: Sauropterygia) are fairly common in the Japanese Upper Cretaceous (Sato et al.., 2012), and the long-necked Elasmosauridae numerically dominate the record, represented by several incomplete but articulated skeletons (e.g., Nakaya 1989; Ogawa and Nakaya, 1998; Sato et al., 2006). In contrast, non-elasmosaurid specimens, such as those of the Polycotylidae and Pliosauroidea, are much fewer in number and represented by more fragmentary remains. However, reports of their occurrences have doubled in the last decade or so (e.g., Sato et al., 2012), suggesting their taxonomic and/or morphological diversity is higher than perceived. In this study, we describe a polycotylid specimen from the Turonian of the Yezo Group in Hokkaido. This specimen is important because 1) it includes the first reasonably complete clavicular arch of the Polycotylidae from Japan, 2) its Turonian
5 4 occurrence demonstrates the survival of Polycotylidae across the Cenomanian-Turonian boundary in northwest Pacific, and 3) it provides the first histological data of definite Polycotylidae using CT-scanning and thin-sectioning. Much of the Yezo plesiosaurian specimens were collected as isolated floats in a riverbed. Even the preserved bones tend to be incomplete due to their large size and weathering of the float, and their stratigraphic origins are often given as a range of the horizons exposed in the area. For example, Echizenya and Minoura (2005) reported the occurrence of two damaged vertebrae and the anterior tip of the interclavicle from the upper Cenomanian to lower Turonian of the Yezo Group, but the fragmentary nature of the specimen and lack of co-occurring age-indicative fossils did not allow in-depth investigation on the morphology and stratigraphic significance of their specimen. Previous studies on plesiosaurian bone histology are limited in their taxonomic, geographic, and stratigraphic coverage. Indeterminate elasmosaurids of southern high latitudes in the latest Cretaceous (Campanian Maastrichtian) are the most common material (e.g., Wiffen et al., 1995; Fostowicz Frelik and Gaździcki, 2001; Talevi and Fernández, 2014), whereas Cruickshank et al. (1996) and Street and O Keefe (2010) studied a pliosaurid and a cryptoclidid of the Jurassic, respectively. To our knowledge, there are no previous studies on the histology of definite polycotylid
6 5 specimens. Wiffen et al. (1995) examined skeletal histological features of two pliosaurid vertebrae from the Campanian-Maastrichtian of New Zealand. The classification back then was the one proposed by Brown (1981) in which the Polycotylidae was not recognized as a discrete taxon, and it is quite possible that Wiffen s pliosaurid specimens may actually represent or include polycotylid material, but it is difficult to prove it. Material and Methods The specimen described here, HMG-357 of Hobetsu Museum in Mukawa Town, Hokkaido, came from the Upper Cretaceous Yezo Group in Obira Town, Hokkaido; see Figure 1 and Remarks on stratigraphy in Systematic Paleontology for details on its geology. Sato and Nishimura (2012) and Sato et al. (2012) briefly reported/mentioned its occurrence. Two anteromost dorsal ribs, #7 and #15 (see Systematic Paleontology below) are used for our histological study. CT scanning was performed using Tesco Microfocus CT TXS320-ACTIS (0.2 mm resolution, 260kV, 200μA) at the National Museum of Nature and Science, Tokyo, Japan, and image segmentation and visualization were performed using VG Studio Max (Volume Graphics GmBH, Heidelberg,
7 6 Germany). The thin sectioning procedure generally follows Chinsamy and Raath (1992). The rib #15 was sectioned at six points, and prepared sections were numbered #15-I to #15-VI from the proximal to distal end. Thin sections were observed in transmitted light using a Leica Z6 APO microscope at Tokyo Gakugei University and Leica M125 microscope at the University of Tokyo, and photographed using Leica DFC290 HD digital sight camera and Canon EOS Kiss X6i digital single-lens camera, respectively. Because of the large size of the samples, multiple photographs were integrated into a single image showing the entire section. The polarizing microscope Olympus BX41 at the University of Tokyo was also used in observation of thin sections, using Canon EOS Kiss X6i digital camera. Original CT images were processed on the software (Photoshop Creative Cloud) to emphasize the contrast. Nomenclature and definitions of technical terms in bone histology are based on Francillon-Vieillot et al. (1990). SYSTEMATIC PALEONTOLOGY Sauropterygia Owen, 1860 Plesiosauria de Blainville, 1835
8 7 Polycotylidae Cope, 1869 Polycotylidae gen. et sp. indet. Referred specimen.--hmg-357, a partial skeleton consisting of five articulated vertebrae at the base of the neck, eight ribs, clavicular arch, and a scapular dorsal blade. Locality.--Collected as a float in the Obirashibe River in the Lake Obirashibe area, Obira Town, Hokkaido, Japan. Horizon.--Estimated to be in an upper part of the Saku Formation (Mm-o unit) of the Yezo Group, upper middle to lower upper Turonian. Remarks on the stratigraphy.--hmg-357 was collected in 1986 as a float in the Obirashibe River by Ken-ichi Chiyokawa and donated to the Hobetsu Museum. A reservoir (Lake Obirashibe) was constructed in 1992 and the locality has been submerged. The nodule containing HMG-357 was prepared sometime before our research,
9 8 but the remaining matrix consists of bioturbated mudstone and includes very fine sands and abundant wood particles. Judging from matrix, the nodule came from Mm-o stratigraphic unit, i.e., the uppermost geological unit of Saku Formation. Mm-o unit correlates to the upper part of Inoceramus hobetsensis Zone and the lower part of Inoceramus teshioensis Zone (Tanabe et al., 1977; Funaki and Hirano, 2004). In addition, the ammonoid Scalarites sp. co-occurred in the nodule; this genus is known from the Turonian to the Coniacian in the Yezo Group (Matsumoto, 1977; Toshimitsu and Hirano, 2000). Based on these evidence, the stratigraphic origin of the plesiosaur specimen is confidently estimated to be in an upper part of the Saku Formation (Mm-o unit) of the Yezo Group, representing the upper middle Turonian to the lower upper Turonian. Osteological description: HMG-357 includes five vertebrae, eight ribs, a part of the left scapula and a clavicular arch. Judging from the anatomical positions and size of individual elements, it is safely assumed that this specimen originated from the pectoral region of a single individual. The neurocentral suture is completely closed in all vertebrae, indicating this individual was an adult or old adult sensu Brown (1981; also see Discussion). An
10 9 Arabic numeral is painted on each element (but it does not match the anatomical order), and in the following description this number is employed to distinguish ribs, e.g., #15 for the rib with the painted number of HMG The articulated vertebrae (Figure 2A, B) represent the transition from the cervical to dorsal vertebrae, and the rib facets gradually shift from the centrum in cervicals to the tip of the transverse process in dorsals. The centra are amphicoelous (Figure 2C, D), and wider than tall and taller than long (Table 1), and there is no ventral notch on the ventral edge of the intervertebral facet. The foramina subcentralia are located in the shallow depressions on the ventral surface separated by a longitudinal ridge (Figure 2E). The neural spine slightly narrows dorsally in lateral view, and lacks the thickening at the anterior and posterior corners seen in the pectoral vertebrae of elasmosaurids (e.g., Sato et al., 2006). All of the eight ribs (#7, 8, 10 to 15: Figure 3) are more or less damaged, but most of them can be matched with the corresponding vertebrae with reasonable confidence based on the fit of the head and rib facet, as well as the symmetry. We consider #11 as the anteromost right rib in the preserved series because of the shortness and the presence of the anterior process. #10 is slightly longer than #11, and likely represents the next right rib, because the partly damaged distal end retains the base of
11 10 the anterior process. The articular facet is elongate and seems to match the facet on the second anteromost vertebra, although it may have been compressed slightly. #12 is longer than #10, and has an elongate articular facet which corresponds to the rib facets of the 3 rd anterior vertebra, and paired with #14 based on the characteristics of the constriction near the head, as well as the thickness of the shaft. #15 and #7 are used for the histological study and described in detail. #7 is the best preserved rib in this specimen, although the distal end and the rim of the articular facet are slightly damaged. It is about 13 cm long from the rib facet to the broken distal end, and the longitudinal diameter of the damaged proximal end is about 25 mm. The shaft is gently curved, and the articulation facet is deeply concave to fit the convex facet on the transverse process, and there are ridges and gentle grooves on the surface near the head. #15 is heavily abraded and missing the rim of the head, the dorsal surface and the distal part, but paired with #7 based on the similarity of the ridge and groove near the ventral edge, as well as the concavity and inferred size of the articular facet. It is about 75 mm long, and the longitudinal diameter of the damaged proximal end is about 20 mm. #8 and #13 represent the posteromost pair belonging to the posteromost vertebra in the preserved series. The rib head is more massive than #7, the articular facet is deeply concave with a well-defined rim, and the shaft is thicker.
12 11 In plesiosaurs, the transition from the cervical to dorsal vertebrae is gradual and they are conventionally distinguished based on the location of the rib facet relative to the neurocentral suture (e.g., Welles, 1943; Carpenter, 1999). The neurocentral suture is completely closed in HMG-357 and cannot be traced in the vertebra bearing #7 and #15 ribs, but the rib facet on the short transverse process is convex without any trace of suture, suggesting it is a dorsal vertebra. In addition, the damaged shaft of #7 indicates that it was much longer than more anterior ribs. Therefore, we interpret #7 and #15 as the first dorsal ribs of this plesiosaur. The preserved portion of the clavicular arch is missing much of the original edge of the bone (Figure 4). It is fairly thick in the anteromedial portion near the large interclavicular foramen, but the element thins posterolaterally, and it is only 1 mm thick or so at the broken edge of the interclavicle and clavicles. The three bones are fused, but the suture is partially traceable. There are longitudinal sharp ridges on the interclavicle lateral to the interclavicular foramen. This portion of the arch is slightly asymmetrical. Judging from the thickness of the anterior edge and concavity on the medial side, the preserved dorsal blade came from the left scapula (Figure 5). The distal end of the blade is damaged but indicates that the tip of the blade widens and did not simply taper.
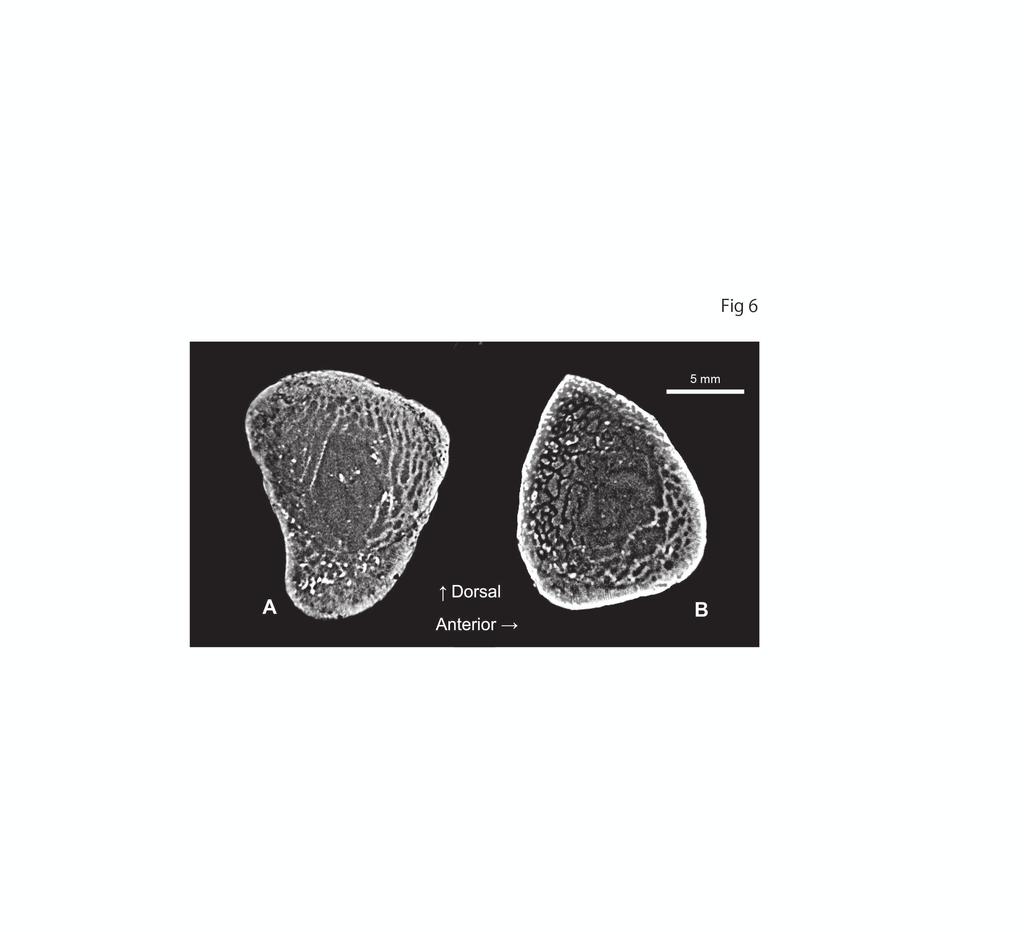
13 12 Microanatomical and histological descriptions CT images of HMG-357 rib #7. The CT value (X-ray transmissivity) of the matrix and bone did not differ considerably, and the contrast of the image was enhanced in Figure 6. The comparison with the structures observed in thin sections supports our interpretation of these images. The brightness of the bone is reversed in some places within the same image, i.e., the trabeculae appear brighter than the matrix in the anterior half of the rib but darker in the posterior half, likely reflecting the subtle difference in the relative transmissivity between the matrix and bone. The bright spots in spogiosa likely represent pyrite crystals (see microscopic description below). The inner rib structure exhibits osteoporotic-like condition at the sectioned two points (i.e., rib head and mid-shaft). The medullary cavity is surrounded by extensive spongiosa, and a thin layer of compact cortical bone forms the solid surface. Microscopic description.--the longitudinal section of a rib head (HMG-357 #15-I, Figure 7A) displays loose spongiosa, consisting of thin trabeculae (about 0.2 mm) oriented longitudinally to the articular facet. The outermost cortex at the ventral side of the rib exhibits nearly avascular primary bone with few secondary osteons. There is a
14 13 mass of tissue at the ventral corner of the articular facet, which appears homogeneous and contains numerous fiber bundles, possibly related with tendon insertions (Figure 7D; Haines and Mohuiddin, 1968; Petermann and Sander, 2013). Bands of alternating light and dark tissue are present in the fiber bundles showing possible tempo and mode of calcification of existing soft tissue (see also Main et al. 2005). In the medullary region, the trabecular tissue is mostly secondary remodeled. In the shaft sections, there is a large open medullary cavity surrounded by spongiosa with numerous erosion lacunae throughout the preserved portion (Figure 7B, C). Most parts of the cortex are heavily remodeled; especially, the ventral region is completely remodeled by dense Haversian tissue (Figure 7E). A thin layer of primary bone tissue consisting of parallel-fibered bone is present in the outermost cortex of the posterior side (Figure 7G). A part of the peripheral bone tissue comprises mostly avascular tissue including possible lines of arrested growth (LAG) (Figure 7I), which resembles an external fundamental system (EFS) (e.g., Sander et al., 2006). The outermost edge of ventral side includes numerous fiber bundles running toward periphery between secondary osteons (Figure 7D). There are numerous erosion cavities in the deeper cortex; they are irregularly shaped and larger in deeper area. Howship s lacunae are occasionally present in the wall of the erosion cavity,
15 14 indicating active resorption by osteoclasts (Figure 7F). The trabeculae are about mm thick and form a coarse network and are composed of secondary lamellar bone tissue (Figure 7H). The marrow spaces of HMG-357-#15 are filled with minerals. Dark irregular spots are observed on the trabecular surfaces and form thick black layers in places (e.g., Figure 7F). They are opaque in transmitted light, but exhibit metallic luster in reflected light, likely representing pyrite crystals commonly occurring in fossilized bones from Yezo Group (e.g., Kaim et al., 2008). DISCUSSION Justification of taxonomic identification HMG-357 is identified as a polycotylid based on the morphology of the vertebrae and clavicular arch. Preserved bones of HMG-357 are comparable in size to the previously described Yezo polycotylid specimens which represent relatively small adult individuals (e.g., Sato and Storrs, 2000; Echizennya and Minoura, 2005). The vertebrae of HMG-357 shows typical features of polycotylid pectorals, such as the short centrum with oval articular faces without a ventral notch, a pair of
16 15 foramina subcentralia separated by a ridge, large zygapophyses, and the neural spine which narrows dorsally (e.g., Williston, 1903; Adams, 1997; Frey et al., 2017). In contrast, more common elasmosaurid pectoral vertebrae are characterized by the square neural spine in which the anterior and posterior corners are thickened and the intervertebral facet bears a ventral notch, and they lack the sharp ridge on the ventral surface of the centrum separating the paired depressions hosting the foramina subcentralia (e.g., Sato et al., 2006). Morphology of the clavicular arch is quite variable among the elasmosaurids (e.g., Welles 1962), but an elongate interclavicular foramen between the sharp longitudinal ridges as in the HMG specimen has not been reported in any known elasmosaurid taxa. Much less information is available for the Cretaceous pliosauroids because of the paucity of the postcranial materials, but HMG-357 can be distinguished from the two genera with comparable specimens: Brachauchenius which lacks foramina subcentralia (e.g., Williston, 1903; Albright et al., 2006a), and Kronosaurus whose cervical and anterior dorsal centra are taller than wide (Romer and Lewis, 1959). The clavicular arch has been described only in a few pliosauroids (e.g., Peloneustes in Andrews, 1913), but we are not aware of any with an interclavicular foramen and surrounding ridge. Characteristics of the interclavicular arch of HMG-357 distinguish it from
17 16 non-polycotylids, but they are fairly common among polycotylids for which detailed comparison is available. HMG-357 shares the presence of a large interclavicular foramen with ridges on the sides with Dolichorhynchops osborni, D. bonneri, Eopolycotylus rankini, and Trinacromerum bentonianum (Williston 1903, 1908; Adams, 1997; Albright et al., 2007b; O Keefe, 2008); the taxa with the large foramen range from the Cenomanian to Santonian or Campanian. In the Campanian-Maastichtian D. herschelensis Sato 2005, however, the area for the interclavicular foramen is mostly ossified, leaving very small openings and weak lateral ridges. The interclavicle is poorly preserved or unknown in many polycotylid taxa and it is hard to assess the effect of ontogenetic variability, but the presence of well-defined foramen in the adult individual HMG-357 and other early polycotylids suggests that the closure of this foramen in D. herschelensis likely represents an autapomorphic condition. Significance of the occurrence of Turonian polycotylids in Japan Early polycotylid remains have been reported from the upper Lower Cretaceous of North America and Australia (Druckenmiller and Russell, 2009), although phylogenetic relationships within the group and referral of certain genera at higher taxonomic level conflict in recent studies (e.g., Albright et al., 2007b;
18 17 Druckenmiller and Russell, 2008; Ketchum and Benson, 2010). Diverse polycotylids are known from the Cenomanian to Campanian, mostly from the Western Interior Seaway (WIS) of North America but also from Asia and northern Africa, with the latest ones from the Campanian-Maastrichtian of the North America and the southern hemisphere (summarized in Druckenmiller and Russell, 2009; Schmeisser McKean and Gillette, 2012). The fossil record of Late Cretaceous polycotylids is geographically and stratigraphically sparse and uneven outside the WIS, and the polycotylid remains from Japanese Cretaceous are important to understand the global distribution of the group. For example, the Cenomanian occurrence of an indeterminate polycotylid (Sato and Storrs, 2000) and the Turonian age of HMG-357 indicate that polycotylids crossed the Cenomanian-Turonian (CT) boundary in the northwestern Pacific as they did in the Western Interior Seaway, indicating the survival was not a local phenomenon restricted to the latter. No other Late Cretaceous polycotylid localities in the Pacific have a stratigraphic range across the boundary. In the Yezo Group of Japan and Russian Far East, CT boundary events such as faunal turnover among shelled molluscs (ammonoids, inoceramids) and some microfossils, as well as the excursion of carbon isotope ratio, are recognized, and their
19 18 causal relationship with an oceanic anoxic event (OAE2) has been debated (e.g., Toshimitsu et al., 2003; Takashima et al., 2004; Yazykova et al., 2004; Takahashi, 2005; Kurihara et al., 2012). Meanwhile, some groups appear to have been less affected. There are no marked extinction events among radiolarians and dinoflagellate cysts (Yazykova et al., 2004; Kaneko and Hirano, 2005), and the coleoid cephalopods Longibelus matsumotoi and Conotheuthis hayakawai occur in Cenomanian and Turonian strata (Tanabe et al., 2017). Sato and Tanabe (1998) reported the occurrence of a Cenomanian polycotylid skeleton which contains many jaw apparati of ammonoids as its stomach contents. Most ammonoid species went extinct at the CT boundary just to be replaced with other new species in the Turonian (Toshimitsu et al., 2003). Although the number and quality of Yezo plesiosaur specimens are not sufficient to analyze changes at lower taxonomic levels as in these invertebrates, they do demonstrate the survival of the clade across the C/T boundary, thanks to the continuous stratigraphic sequence with fine biostratigraphic resolution. Ontogenetic stages and aquatic adaptation inferred from bone histology Ontogeny affects morphology and various other aspects of an animal, and the importance of assessing the growth stage of studied specimen(s) cannot be
20 19 over-emphasized. However, growth stages are only roughly estimated for large fossil vertebrates, because their remains are often fragmentary and/or rare, and it is hard to document an ontogenetic sequence. Here we assess the growth stage of HMG-357 based on the osteological and histological features. Unlike dinosaurs for which various criteria to distinguish ontogenetic stages have been proposed (summarized in Hone et al., 2016), there are very few previous studies for plesiosaurs. Brown (1981: 255) distinguished juvenile, adult, and old adult individuals of plesiosaurs based on osteological characters. An individual is juvenile if the neural arches and centra are not fused; it is adult if they are fused, and old adult if the individual exhibits additional characters indicative of advanced ossification. As Brown himself clearly indicated, these terms should be always used with quotation marks to indicate their special meanings which do not necessary correspond to the biological age or maturity. Unfortunately this practice is not strictly followed, and it is sometimes unclear whether the distinction of juvenile and adult is based on the criterion set by Brown or not (e.g., Wiffen et al., 1995; Liebe and Hurum, 2012). Brown s criterion has been widely used in the description of plesiosaurian specimens, probably because it is so practical; vertebrae are commonly found and easily
21 20 identified as plesiosaurian. At the same time, it is quite approximate, in particular for the distinction of adult and old adult specimens. Ontogenetic sequence of ossification remains unknown for plesiosaurs, and it is hard to judge if a particular character of ossification appears earlier or later than other features. In the first place, the sequence of the closure of neurocental sutures within the vertebral column (e.g., Brochu, 1996; Hone et al., 2016) has not been established to show if there is a consistent pattern within the Plesiosauria or subgroups, so even the distinction of juvenile and adult is challenged if the suture is partially closed within an individual. It is practically impossible to distinguish adult and old adult individuals for the fragmentary specimens in which additional characters indicative of advanced ossification cannot be confirmed. In the case of HMG-357, the closure and complete obliteration of the neurocentral suture indicates it does not represent a juvenile sensu Brown. The interclavicle and clavicles are fused, leaving partially obliterated sutures, and this may be taken as an evidence of advanced ossification at old adult level. These bones are fused but sutures are clearly visible at least in one juvenile Trinacromerum specimen (the holotype of T. willistoni Riggs 1944; T. bentonianum in Carpenter 1996), and they are at least partially visible in several polycotylid specimens in which neurocentral
22 21 sutures are closed (e.g., Sato, 2005; Albright et al., 2007b). The parallel-fibered avascular bone observed in HMG-357 (Figure 7G) can be interpreted as a sign of slow periosteal growth immediately before death. Woven-fibered tissue with plexiform or radial vascular canals have been observed in the juvenile and subadult individuals of various non-polycotylid plesiosaurs and pliosaurid (see Introduction) plesiosaurs (Wiffen et al., 1995; Fostowicz Frelik and Gaździcki, 2001; Liebe and Hurum, 2012), suggesting relatively rapid bone deposition (Huttenlocker et al., 2013). Assuming that rapid deposition of bone tissue also occurred in an early ontogenetic stage of polycotylid plesiosaurs, the peripheral bone tissue in HMG-357 may indicate slowed deposition at a late ontogenetic stage of the individual. Given that the degree of remodeling is mostly related with skeletal maturity in tetrapods such as dinosaurs, marine reptiles, and terrestrial mammals (Zug et al., 1986; Wiffen et al., 1995; Klein and Sander, 2008; Mitchell et al., 2017), the extensive remodeling of the rib may suggest that HMG-357 have reached skeletal maturity. In addition, although obscured by the extensive remodeling, the presence of the peripheral bone tissue which resembles an EFS implies an effective cessation of any significant periosteal growth of the bone in HMG-357 (e.g., Erickson, 2005; Woodward et al., 2011).
23 22 The extensive spongiosa and thin compacta in the studied HMG ribs represents an osteoporotic-like condition (Ricqlès and Buffrénil, 2001), which has been reported in highly aquatic amniotes including extinct taxa such as ichthyosaurs (Houssaye et al., 2014; Houssaye et al., 2016; Nakajima et al., 2014) and mosasaurs (Houssaye et al., 2013). Prominence of the erosion spaces including the Howship s lacunae, suggests ongoing osteoclast activity and intense resorption activity. In extant taxa, active swimmers such as cetacean and pinnipedians displays a similar microanatomical features (notably the spongy inner organization) requiring efficient swimming abilities (e.g. manoeuvrability, speed) and relying on a hydrodynamic buoyancy and body trim control (Ricqlès and Buffrénil, 2001; Houssaye, 2009). Therefore, these results on HMG-357 suggest that this polycotylid plesiosaur has acquired highly aquatic adaptation in bone microstructure. In summary, osteological features indicate that HMG-357 represents an old adult sensu Brown (1981), in which the periosteal deposition of bone tissue was probably slowed by maturity at least in this part of the skeleton. The microscopic features also exhibit an osteoporotic-like condition, suggesting that polycotylid plesiosaurs were highly aquatic animals.
24 23 Conclusions HMG-357 represents an indeterminate polycotylid plesiosaur from the Upper Cretaceous Yezo Group in Hokkaido, Japan. Its stratigraphic origin is estimated to be the upper middle to lower upper Turonian, and the Turonian occurrence demonstrates the survival of the polycotylids across the Cenomanian-Turonian boundary in northwestern Pacific. The specimen consists of five vertebrae and ribs at the base of the neck, clavicular arch, and a scapular blade, and the ribs were subjected to the first detailed histological study of the Polycotylidae. The complete closure of the neurocentral suture and partial obliteration of the clavicular sutures indicates that the individual reached an advanced stage of ossification. Parallel-fibered tissue and a possible EFS in the rib are signs of slowed growth, and osteoporotic-like condition suggests that this polycotylid was highly adapted to life in water. Acknowledgements We thank Ken-ichi Chiyokawa for donating the specimen to the Hobetsu Museum. We are grateful to the museum for the permission to use their specimen for destructive analysis, and former and current curators Tsutomu Chitoku and Kazuhiko Sakurai for
25 24 assisting various aspects of our research. Two anonymous reviewers provided constructive comments and suggestions which improved our manuscript considerably. Chisako Sakata and Makoto Manabe (National Museum of Nature and Science) are thanked for assistance in the use of the CT-scanner, and Koichiro Fujimoto, Masataka Nakata (Tokyo Gakugei University) and Takanobu Tsuihiji (University of Tokyo) for assistance in the use of laboratory equipment. We also thank Yasuhisa Nakajima and Momo Yamashita (University of Tokyo) for their helpful advice, and Janet and David Gillette of Museum of Northern Arizona for the access to comparative specimens. Financial support was provided by the Inamori Foundation and Tokyo Gakugei University (to TS), JSPS KAKENHI grants 15K05327 (to TS) and (to SH). References Adams, D. A. 1997: Trinacromerum bonneri, new species, last and fastest plesiosaur of the Western Interior Seaway. Texas Journal of Science, vol. 49, p Albright, L. B., Gillette, D. D. and Titus, A. L., 2007a: Plesiosaurs from the Upper Cretaceous (Cenomanian-Turonian) Tropic Shale of southern Utah, part 1: new records of the pliosaur Brachauchenius lucasi. Journal of Vertebrate Paleontology, vol. 27, p
26 25 Albright, L. B. III, Gillette, D. D. and Titus, A. L., 2007b: Plesiosaurs from the Upper Cretaceous (Cenomanian Turonian) Tropic Shale of southern Utah, part 2: Polycotylidae. Journal of Vertebrate Paleontology, vol. 27, p Andrews, C. W., 1913: A Catalogue of the Marine Reptiles of the Oxford Clay, Part II. British Museum (Natural History), 206p, pls 1-13, London. Blainville, H. D. de, 1835: Description de quelques espèces de reptiles de la Californie, précédé de l analyse d un système general d Erpetologie et d Amphibiologie. Nouvelles Annales du Museum National d Histoire Naturelle, Paris, vol. 4, p Brochu, C., 1996: Closure of neurocetntral sutures during crocodilian ontogeny: implications for maturity assessment in fossil archosaurs. Journal of Vertebrate Paleontology, vol. 16, p Brown, D. S., 1981: The English Upper Jurassic Plesiosauroidea. Bulletin of British Museum of Natural History (Geology), vol. 35, p Canoville, A. and Laurin, M., 2010: Evolution of humeral microanatomy and lifestyle in amniotes, and some comments on palaeobiological inferences. Biological Journal of the Linnean Society, vol. 100, p Carpenter, K. 1996: A review of short-necked plesiosaurs from the Cretaceous of the
27 26 Western Interior, North America. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, vol. 201, p Carpenter, K., 1999: Revision of North American elasmosaurs from the Cretaceous of the Western Interior. Paludicola, vol. 2, p Chinsamy, A. and Raath, M., 1992: Preparation of fossil bone for histological examination. Palaeontologia africana, vol. 29, p Cohen, K. M., Finney, S. C., Gibbard, P. L. and Fan, J.-X., 2013 (updated in 2017): The ICS International Chronostratigraphic Chart. Episodes 36: URL: Cope, E. D., Synopsis of the extinct Batrachia, Reptilia and Aves of North America, part 1. Transactions of the American Philosophical Society, New Series, vol. 14, p Cruickshank, A., Martill, D. and Noe, L., 1996: A pliosaur (Reptilia, Sauropterygia) exhibiting pachyostosis from the Middle Jurassic of England. Journal of the Geological Society, vol. 153, p Druckenmiller, P. S. and Russell, A. P., 2008: A phylogeny of Plesiosauria (Sauropterygia) and its bearing on the systematic status of Leptocleidus Andrews, Zootaxa, vol. 1863, p
28 27 Druckenmiller, P. S. and Russell, A. P., 2009: Earliest North American occurrence of Polycotylidae (Sauropterygia: Plesiosauria) from the Lower Cretaceous (Albian) Clearwater Formation, Alberta, Canada. Journal of Paleontology, vol. 83, p Echizenya, H., and Minoura, N., 2005: Late Cretaceous polycotylid plesiosaur from Mikasa District, Hokkaido. Bulletin of the Hobetsu Museum vol. 21, p (in Japanese with English abstract). Erickson, G. M., 2005: Assessing dinosaur growth patterns: a microscopic revolution. Trends in Ecology and Evolution, vol. 20, no. 12, p Fostowicz-Frelik, L. and Gaździcki, A., 2001: Anatomy and histology of plesiosaur bones from the Late Cretaceous of Seymour Island, Antarctic Peninsula. Acta Palaeontologica Polonica, vol. 60, p Francillon-Vieillot, H., de Buffrénil, V., Castanet, J., Géraudie, J. F., Meunier, J., Sire, J. Y., Zylberberg, L. and Ricqlès, A. de., 1990: Microstructure and mineralization of vertebrate skeletal tissues. In Carter, J. G. ed., Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends, vol. 1. Van Nostrand Reinhold, New York. Frey, E., Mulder, E. W. A., Stinnesbeck, W., Rivera-Sylva, H. E., Padilla-Gutiérrez, J. M.
29 28 and González-González, A. H., 2017:A new polycotylid plesiosaur with extensive soft tissue preservation from the early Late Cretaceous of northeast Mexico. Boletín de la Sociedad Geológica Mexicana, vol. 69(1), p Funaki, H. and Hirano, H., 2004: Cretaceous stratigraphy in the northeastern part of the Obira area, Hokkaido, Japan. Bulletin of the Mikasa City Museum, no. 8, p (in Japanese with English abstract) Haines, R. W. and Mohuiddin, A., 1968: Metaplastic bone. Journal of Anatomy, vol. 103, p Hayashi, S., Houssaye, A., Nakajima, Y., Chiba, K., Ando, T., Sawamura, H., Inuzuka, N., Kaneko, N. and Osaki, T., 2013: Bone inner structure suggests increasing aquatic adaptations in Desmostylia (Mammalia, Afrotheria). PloS One, vol. 8, no. 4, e Hone, D. W. E., Farke, A. A., and Wedel, M. J. 2016: Ontogeny and the fossil record: what, if anything, is an adult dinosaur? Biology Letters, vol. 12, Houssaye, A. 2009: Pachyostosis in aquatic amniotes: a review. Integrative Zoology, 4, p Houssaye, A. and Bardet, N., 2012: Rib and vertebral micro-anatomical characteristics
30 29 of hydropelvic mosasauroids. Lethaia, vol. 45, p Houssaye, A., Lindgren, J., Pellegrini, R., Lee, A. H, Germain, D. and Polcyn, J. M., 2013: Microanatomical and histological features in the long bones of mosasaurine mosasaurs (Reptilia, Squamata): implications for aquatic adaptation and growth rates. Plos One, vol. 8, no. 10, e Houssaye, A., Scheyer, T. M., Kolb, C., Fischer, V., Sander, P. M., 2014: A New Look at Ichthyosaur Long Bone Microanatomy and Histology: Implications for Their Adaptation to an Aquatic Life. PLoS ONE, vol. 9, no. 4, e Houssaye, A., Sander, P. M., Klein, N., 2016: Adaptive Patterns in Aquatic Amniote Bone Microanatomy-More Complex than Previously Thought. Integrative and Comparative Biology, vol. 56, no. 6, p Huttenlocker, A. K., Woodward, H. N. and Hall, B. K., 2013: The biology of bone. In, Padian, K. and Lamm, E. T., eds., Bone Histology of Fossil Tetrapods: Advancing Methods, Analysis, and Interpretation, p University of California Press, Berkeley. Kaim, A., Kobayashi, Y., Echizenya, H., Jenkins, R. G., and Tanabe, K., 2008: Chemosynthesis-based associations on Cretaceous plesiosaurid carcasses. Acta Palaeontologica Polonica vol. 53, p
31 30 Kaneko, M. and Hirano, H., 2005: Dinoflagellate cyst assemblages and reconstruction of primary productivity across the Cenomanian/Turonian boundary in the Obira area, northwestern Hokkaido, Japan. Bulletin of the Mikasa City Museum, no. 9, p (In Japanese with English abstract) Ketchum, H. F. and Benson, R. B., 2010: Global interrelationships of Plesiosauria (Reptilia, Sauropterygia) and the pivotal role of taxon sampling in determining the outcome of phylogenetic analyses. Biological Reviews, vol. 85, p Klein, N. and Sander, P. M., 2008: Ontogenetic stages in the long bone histology of sauropod dinosaurs. Paleobiology, vol. 34, p Konishi, T., Caldwell, M.W., Nishimura, T., Sakurai, K., and Tanoue, K A new halisaurine mosasaur (Squamata: Halisaurinae) from Japan: the first record in the western Pacific realm and the first documented insights into binocular vision in mosasaurs. Journal of Systematic Palaeontology: vol. 14, p , Kubo, T., Mitchell, M. T. and Henderson, D. M., 2012: Albertonectes vanderveldei, a new elasmosaur (Reptilia, Sauropterygia) from the Upper Cretaceous of Alberta. Journal of Vertebrate Paleontology, vol. 32, p Kurihara, K., Toshimitsu, S. and Hirano, H., 2012: Ammonoid biodiversity changes
32 31 across the Cenomanian-Turonian boundary in the Yezo Group, Hokkaido, Japan. Acta Palaeontologica Polonica, vol. 57, p Liebe, L. and Hurum, J. H., 2012: Gross internal structure and microstructure of plesiosaur limb bones from the Late Jurassic, central Spitsbergen. Norwegian Journal of Geology, vol. 92, p Main, R., Ricqlès, A., Horner, J. and Padian, K., 2005: The evolution and function of thyreophoran dinosaur scutes: implications for plate function in stegosaurs. Paleobiology, vol. 31, p Matsumoto, T., 1977: Some heteromorph ammonites from the Cretaceous of Hokkaido (Studies of the Cretaceous ammonites from Hokkaido and Saghalien-XXXI). Science Reports of the Department of Geology, Kyushu University, vol. 12, no. 3, p , pls Mitchell, J., Sander, P. M., Stein, K., 2017: Can secondary osteons be used as ontogenetic indicators in sauropods? Extending the histological ontogenetic stages into senescence. Paleobiology, vol. 43, no. 2, p Nakajima, Y., Houssaye, A. and Endo, H., 2014: Osteohistology of the Early Triassic ichthyopterygian reptile Utatsusaurus hataii: Implications for early ichthyosaur biology. Acta Palaeontologica Polonica, vol. 59, p
33 32 Nakaya, H., 1989: Upper Cretaceous elasmosaurid (Reptilia, Plesiosauria) from Hobetsu, Hokkaido, northern Japan. Transactions and Proceedings of the Palaeontological Society of Japan, New Series, vol. 154, p Ogawa, K. and Nakaya, H., 1998: Late Cretaceous Elasmosauridae fossils from Nakagawa, Hokkaido, Japan. Bulletin of the Nakagawa Museum of Natural History, vol. 1, p (In Japanese with English abstract). O Keefe, F. R., 2008: Cranial anatomy and taxonomy of Dolichorhynchops bonneri new combination, a polycotylid (Sauropterygia: Plesiosauria) from the Pierre Shale of Wyoming and South Dakota. Journal of Vertebrate Paleontology, vol. 28, p Owen, R., 1860: On the orders of fossil and recent Reptilia, and their distribution in time. Reports of the British Association for the Advancement of Science, vol. 29, p Petermann, H. and Sander, P. M., 2013: Histological evidence for muscle insertion in extant amniote femora: implications for muscle reconstruction in fossils. Journal of Anatomy, vol. 222, p Ricqlès, A. and Buffrénil, V., 2001: Bone histology, heterochronies and the return of Tetrapods to life in water: where are we? In: Mazin, J. M. and Buffrénil, V.,
34 33 eds., Secondary adaptation of tetrapods to life in water, pp München: Verlag. Riggs, E. S., 1944: A new polycotylid plesiosaur. University of Kansas Science Bulletin, vol. 30, p Romer, A. S. and Lewis, A. D., 1959: A mounted skeleton of the giant plesiosaur Kronosaurus. Breviora, vol. 112, p Sander, P. M., Mateus, O., Laven, T. and Knötschke, N. 2006: Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur. Nature, vol. 441, p Sato, T. and Tanabe, K., 1998: Cretaceous plesiosaurs ate ammonites. Nature, vol. 394, p. 629 Sato, T., 2005: A new polycotylid plesiosaur (Reptilia, Sauropterygia) from the Upper Cretaceous Bearpaw Formation in Saskatchewan, Canada. Journal of Paleontology, vol. 79, p Sato, T., and Storrs, G. W., 2000: An early polycotylid plesiosaur (Reptilia; Sauropterygia) from the Cretaceous of Hokkaido, Japan. Journal of Paleontology, vol. 74, p Sato, T. and Nishimura, T., 2012: Clavicular arch of a polycotylid plesiosaur from
35 34 Obira Town, Hokkaido. Abstracts with Programs, 161st Regular Meeting of the Palaeontological Society of Japan, Tomioka, Gunma, p. 14. (in Japanese with English title). Sato, T., Hasegawa, Y. and Manabe, M., 2006: A new elasmosaurid plesiosaur from the Upper Cretaceous of Fukushima, Japan. Palaeontology, vol. 49, p Sato, T., Wu, X.-C., Tirabasso, A. and Bloskie, P., 2011: Braincase of a polycotylid plesiosaur (Reptilia: Sauropterygia) from the Upper Cretaceous of Manitoba, Canada. Journal of Vertebrate Paleontology, vol. 31, p Sato, T., Konishi, T., Hirayama, R. and Caldwell, M. W., 2012: A review of the Upper Cretaceous marine reptiles from Japan. Cretaceous Research, vol. 37, p Schmeisser McKean R., 2012: A new species of a polycotylid plesiosaur (Reptilia: Sauropterygia) from the Lower Turonian of Utah: Extending the stratigraphic range of Dolichorhynchops. Cretaceous Research, vol. 34: Street, H. P. and O'Keefe, F. R., 2010: Evidence of pachyostosis in the cryptocleidoid plesiosaur Tatenectes laramiensis from the Sundance Formation of Wyoming. Journal of Vertebrate Paleontology, vol. 30, p Talevi, M. and Fernández, M. S., 2014: Remodeling of skeletal tissues bone and
36 35 structural specialisations in an elasmosaurid (Sauropterygia: Plesiosauroidea) from the Upper Cretaceous of Patagonia, Argentina. Historical Biology, vol. 27, p Takahashi, A., 2005: Responses of inoceramid bivalves to environmental disturbances across the Cenomanian/Turonian boundary in the Yezo forearc basin, Hokkaido, Japan. Cretaceous Research, vol. 26, p Takashima, R., Kawabe, F., Nishi, H., Moriya, K., Wani, R. and Ando, H., 2004: Geology and stratigraphy of forearc basin sediments in Hokkaido, Japan: Cretaceous environmental events on the northwest Pacific margin. Cretaceous Research, vol. 25, p Tanabe, K., Hirano, H., Matsumoto, T. and Miyata, Y., 1977: Stratigraphy of the Upper Cretaceous deposits in the Obira area, northwestern Hokkaido. Science Reports of the Department of Geology, Kyushu University, vol. 12, p (in Japanese with English abstract) Tanabe, K., Misaki, A., Hikida, Y. and Nishimura, T., 2016: New records of coleoid cephalopod jaws from the Upper Cretaceous of Hokkaido, Japan, and their paleobiogeographic and evolutionary implications. Cretaceous Research, vol. 70,
37 36 Toshimitsu, S. and Hirano, H., 2000: Database of the Cretaceous ammonoids in Japan Stratigraphic distribution and bibliography. Bulletin of the Geological Survey of Japan, vol. 51, p Toshimitsu, S., Hirano, H., Matsumoto, T. and Takahashi, K., 2003: Databse and species diversity of Japanese Cretaceous ammonoids. Journal of Asian Earth Sciences, vol. 21, p Tsushima, K., Tanaka, K., Matsuno, K. and Yamaguchi, S., 1958: Geological Sheet Map Tappu, Scale 1:50,000 and its Explanatory Text, 74p. Hokkaido Development Agency. (in Japanese with English abstract) Welles, S. P., 1943: Elasmosaurid plesiosaurs with description of new material from California and Colorado. Memoirs of the University of California, vol. 13, p Welles, S. P., 1962: A new species of elasmosaur from the Aptian of Columbia and a review of the Cretaceous Plesiosaurs. University of California, Publications in Geological Sciences, vol. 44, p Wiffen, J., Buffrénil, V. de, Ricqlès, A. de and Mazin, J., 1995: Ontogenetic evolution of bone structurein Late Cretaceous Plesiosauria from New Zealand. Geobios, vol. 28, p
38 37 Williston, S. W., 1903: North American plesiosaurs, Part 1. Field Columbian Museum Publication 73, Geological Series vol. 2, Williston, S. W., 1908: North American plesiosaurs: Trinacromerum. Journal of Geology, vol. 16, p Woodward, H. N., Horner, J. R. and Farlow, J. O., 2011: Osteohistological evidence for determinate growth in the American alligator. Journal of Herpetology, vol. 45, p Yazykova, E. A., Peryt, D., Zonova, T. D., and Kasintzova, L. I., 2004: The Cenomanian/Turonian boundary in Sakhalin, Far East Russia: Ammonites, inoceramids, foraminifera, and radiolarians. New Zealand Journal of Geology and Geophysics, vol. 47, p Zug, G. R., Wynn, A. H. and Ruckdeschel, C., 1986: Age determination of loggerhead sea turtles, Caretta caretta, by incremental growth marks in the skeleton. Smithsonian Institution Contributions to Zoology, 427, p
39 38 Figure and Table captions Figure 1. Locality map and stratigraphy of HMG-357. A, map of Hokkaido with a star indicating the fossil locality and grey areas the distribution of the Yezo Group; B, details of the now-submerged locality area: Mm-o refers to local stratigraphic units, Ng to Neogene, Pg to Paleogene; C, local stratigraphy with a star indicating HMG-357 horizon. Geological map based on Tsushima et al., Timescale follows Cohen et al. (2013). Figure 2. Photograph (A) and interpretation (B) of the left lateral view of the five vertebrae of HMG-357 in articulation, and anterior (C), posterior (D) and ventral (E) views of the second vertebra. Hatching indicates damaged surface, and the open-circle-and-dot pattern marks matrix/adhesive in this and following figures. Abbreviations: rf, rib facet; tp, transverse process. Figure 3. Photographs (A to I) and interpretation (J to R) of the ribs of HMG-357. A and J, dorsal (?) view of the 1st right rib (#11); B and K, dorso-poterior view of the 2nd right(?) rib (#10); C and L, posterior (?) view of the 3rd left
40 39 rib (#12); proximal (D and M), anterior (E and N), posterior (F and O) views of the 4th left rib (#7); posterodorsal (G and P) and anterior (H and Q) views of the 4 th right rib (#15); I and R, posterior view of the 5th left rib (#13). Figure 4. Photograph (A to C) and interpretation (D to F) of the clavicular arch of HMG-357 in anterior (A, D), ventral (B, E), and dorsal or visceral (C, F) views. Abbreviations: cl, clavicle; frag, fragmentary; icf, interclavicular foramen; icl, interclavicle; l, left; r, right. Figure 5. Photograph (A, B) and interpretation (C, D) of the dorsal blade of the right (?) scapula of HMG-357 in lateral (A, C) and medial (B, D) views. Figure 6. CT images of the proximal (A) and midshaft (B) portions of the left (?) anteromost dorsal rib (#7) of HMG-357. Figure 7. Histological thin sections of the right (?) anteromost dorsal rib (#15) of HMG-357. A, longitudinal section of the proximal portion under natural transmitted light, with square D indicating the location of the close-up image in
41 40 D of this figure; B and C, transverse sections (B is a few millimeters more distal than C) under natural transmitted light, with squares E to I indicating locations of close-up images in E to I of this figure; D to I, close-ups under polarized microscope showing fiber bundles and the bands of alternating light and dark tissue (D), Haversian tissue (E), Howship s lacunae (arrow heads, F), primary parallel-fibered bone tissue (arrow head, G), secondary lamellar bone tissue (H), and avascular bone tissue with possible lines of arrested growth (arrow head, I). Table 1. Vertebral measurements of HMG-357. Length (L), height (H), and width (W) of the centrum in mm. Subscripts d and e refer to distorted centrum and estimated length, respectively.

42

43 ccepted manuscript

44

45

46 ted manusc
47

48
49 Table 1 L H W 1 (anteromost) 42.9 e d 61.8 e 64.8
These small issues are easily addressed by small changes in wording, and should in no way delay publication of this first- rate paper.
 Reviewers' comments: Reviewer #1 (Remarks to the Author): This paper reports on a highly significant discovery and associated analysis that are likely to be of broad interest to the scientific community.
Reviewers' comments: Reviewer #1 (Remarks to the Author): This paper reports on a highly significant discovery and associated analysis that are likely to be of broad interest to the scientific community.
Papers in Press. Papers in Press includes peer-reviewed, accepted manuscripts of research
 Papers in Press Papers in Press includes peer-reviewed, accepted manuscripts of research articles, reviews, and short notes to be published in Paleontological Research. They have not yet been copy edited
Papers in Press Papers in Press includes peer-reviewed, accepted manuscripts of research articles, reviews, and short notes to be published in Paleontological Research. They have not yet been copy edited
Sauropterygia. Lepidosauromorpha
 Sauropterygia Lepidosauromorpha ***cladogram of lepids*** Pachypleurosauridae Nothosauria Pliosauroidea Plesiosauroidea Mosasauridae Thalattosauriformes? Placodontia Pistosauridae Plesiosauria Sauropterygia
Sauropterygia Lepidosauromorpha ***cladogram of lepids*** Pachypleurosauridae Nothosauria Pliosauroidea Plesiosauroidea Mosasauridae Thalattosauriformes? Placodontia Pistosauridae Plesiosauria Sauropterygia
Anatomy. Name Section. The Vertebrate Skeleton
 Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
BIBLIOGRAPHIE SUR LES SAUROPTERYGIENS
 BIBLIOGRAPHIE SUR LES SAUROPTERYGIENS Bakker, R. T. 1993. Plesiosaur extinction cycles- Events that mark the beginning, middle and end of the Cretaceous. In Caldwell, W. G. E. and Kaufman, E. G. (eds.).
BIBLIOGRAPHIE SUR LES SAUROPTERYGIENS Bakker, R. T. 1993. Plesiosaur extinction cycles- Events that mark the beginning, middle and end of the Cretaceous. In Caldwell, W. G. E. and Kaufman, E. G. (eds.).
Fossils explained 54
 Fossils explained 54 Plesiosaurs Plesiosaurs are an unusual and intriguing group of extinct aquatic reptiles (Fig. 1). They are sauropterygians, a group known from an array of semiaquatic forms during
Fossils explained 54 Plesiosaurs Plesiosaurs are an unusual and intriguing group of extinct aquatic reptiles (Fig. 1). They are sauropterygians, a group known from an array of semiaquatic forms during
A study of the morphology of Tatenectes laramiensis, a cryptocleidoid plesiosaur from the Sundance Formation (Wyoming, USA)
") Marshall University Marshall Digital Scholar Theses, Dissertations and Capstones 2009 A study of the morphology of Tatenectes laramiensis, a cryptocleidoid plesiosaur from the Sundance Formation (Wyoming,
Marshall University Marshall Digital Scholar Theses, Dissertations and Capstones 2009 A study of the morphology of Tatenectes laramiensis, a cryptocleidoid plesiosaur from the Sundance Formation (Wyoming,
Description of an Unusual Cervical Vertebral Column of a Plesiosaur from the Kiowa Shale
 Fort Hays State University FHSU Scholars Repository Master's Theses Graduate School Spring 2014 Description of an Unusual Cervical Vertebral Column of a Plesiosaur from the Kiowa Shale Ian N. Cost Fort
Fort Hays State University FHSU Scholars Repository Master's Theses Graduate School Spring 2014 Description of an Unusual Cervical Vertebral Column of a Plesiosaur from the Kiowa Shale Ian N. Cost Fort
William R. Wahl. Wyoming Dinosaur Center, 110 Carter Ranch Rd, Thermopolis, WY INTRODUCTION
 Paludicola 9(1):32-39 November 2012 by the Rochester Institute of Vertebrate Paleontology GASTRIC CONTENTS OF A PLESIOSAUR FROM THE SUNDANCE FORMATION (JURASSIC), HOT SPRINGS COUNTY, WYOMING, AND IMPLICATIONS
Paludicola 9(1):32-39 November 2012 by the Rochester Institute of Vertebrate Paleontology GASTRIC CONTENTS OF A PLESIOSAUR FROM THE SUNDANCE FORMATION (JURASSIC), HOT SPRINGS COUNTY, WYOMING, AND IMPLICATIONS
A new species of sauropod, Mamenchisaurus anyuensis sp. nov.
 A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
TRACHEMYS SCULPTA. A nearly complete articulated carapace and plastron of an Emjdd A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE,
 A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
Erycine Boids from the Early Oligocene of the South Dakota Badlands
 Georgia Journal of Science Volume 67 No. 2 Scholarly Contributions from the Membership and Others Article 6 2009 Erycine Boids from the Early Oligocene of the South Dakota Badlands Dennis Parmley J. Alan
Georgia Journal of Science Volume 67 No. 2 Scholarly Contributions from the Membership and Others Article 6 2009 Erycine Boids from the Early Oligocene of the South Dakota Badlands Dennis Parmley J. Alan
Lower Cretaceous Kwanmon Group, Northern Kyushu
 Bull. Kitakyushu Mus. Nat. Hist., 11: 87-90. March 30, 1992 A New Genus and Species of Carnivorous Dinosaur from the Lower Cretaceous Kwanmon Group, Northern Kyushu Yoshihiko Okazaki Kitakyushu Museum
Bull. Kitakyushu Mus. Nat. Hist., 11: 87-90. March 30, 1992 A New Genus and Species of Carnivorous Dinosaur from the Lower Cretaceous Kwanmon Group, Northern Kyushu Yoshihiko Okazaki Kitakyushu Museum
Introduction and objectives
 Chapter 1 Plesiosaurus! A fresh-water Plesiosaurus! cried Summerlee. That I should have lived to see such a sight! We are blessed, my dear Challenger, above all zoologists since the world began! The Lost
Chapter 1 Plesiosaurus! A fresh-water Plesiosaurus! cried Summerlee. That I should have lived to see such a sight! We are blessed, my dear Challenger, above all zoologists since the world began! The Lost
A R T I C L E S STRATIGRAPHIC DISTRIBUTION OF VERTEBRATE FOSSIL FOOTPRINTS COMPARED WITH BODY FOSSILS
 A R T I C L E S STRATIGRAPHIC DISTRIBUTION OF VERTEBRATE FOSSIL FOOTPRINTS COMPARED WITH BODY FOSSILS Leonard Brand & James Florence Department of Biology Loma Linda University WHAT THIS ARTICLE IS ABOUT
A R T I C L E S STRATIGRAPHIC DISTRIBUTION OF VERTEBRATE FOSSIL FOOTPRINTS COMPARED WITH BODY FOSSILS Leonard Brand & James Florence Department of Biology Loma Linda University WHAT THIS ARTICLE IS ABOUT
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC
 FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
Further records of plesiosaurian reptiles of Jurassic and Cretaceous age from Western Australia
 l\ccord5 of thc WC51crn AII5Im//l1/I IVl115CIIIII 19: 475:; (1998). Further records of plesiosaurian reptiles of Jurassic and Cretaceous age from Western Australia John A. Long l and Arthur R. I. Cruickshank
l\ccord5 of thc WC51crn AII5Im//l1/I IVl115CIIIII 19: 475:; (1998). Further records of plesiosaurian reptiles of Jurassic and Cretaceous age from Western Australia John A. Long l and Arthur R. I. Cruickshank
A new partial skeleton of a cryptocleidoid plesiosaur from the Upper Jurassic Sundance Formation of Wyoming
 Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 12-2010 A new partial skeleton of a cryptocleidoid plesiosaur from the Upper Jurassic Sundance Formation
Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 12-2010 A new partial skeleton of a cryptocleidoid plesiosaur from the Upper Jurassic Sundance Formation
FIELDIANA GEOLOGY NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA
 FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
Earliest record of the genus Tylosaurus (Squamata; Mosasauridae) from the Fort Hays Limestone (Lower Coniacian) of western Kansas
 from the Fort Hays Limestone (Lower Coniacian) of western Kansas") Earliest record of the genus Tylosaurus (Squamata; Mosasauridae) from the Fort Hays Limestone (Lower Coniacian) of western Kansas Author: Michael J. Everhart Source: Transactions of the Kansas Academy
Earliest record of the genus Tylosaurus (Squamata; Mosasauridae) from the Fort Hays Limestone (Lower Coniacian) of western Kansas Author: Michael J. Everhart Source: Transactions of the Kansas Academy
Bibliographie de Kenshu Shimada
 Bibliographie de Kenshu Shimada Shimada, K. 1986. [Elasmobranchs from the Early Pliocene Naarai Formation, Choshi City, Chiba Prefecture, Japan]; pp. 357-359, Twenty-ninth Japanese Students Science Prize
Bibliographie de Kenshu Shimada Shimada, K. 1986. [Elasmobranchs from the Early Pliocene Naarai Formation, Choshi City, Chiba Prefecture, Japan]; pp. 357-359, Twenty-ninth Japanese Students Science Prize
The occurrence of Elasmosaurus and Polycotylus in Russian deposits. *
 Yezregodneek Geologhee i Meeneraloghee Rossiee [Annuaire Geologique et Minéralogique de la Russie] Edited under the direction of N. Krishtofovich Vol. 14, Moscow, 1912 The occurrence of Elasmosaurus and
Yezregodneek Geologhee i Meeneraloghee Rossiee [Annuaire Geologique et Minéralogique de la Russie] Edited under the direction of N. Krishtofovich Vol. 14, Moscow, 1912 The occurrence of Elasmosaurus and
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A.
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Non-Dinosaurians of the Mesozoic
 Non-Dinosaurians of the Mesozoic Calling the Mesozoic the Age of Dinosaurs is actually not quite correct Not all reptiles of the Mesozoic were dinosaurs. Many reptiles (and other amniotes) have returned
Non-Dinosaurians of the Mesozoic Calling the Mesozoic the Age of Dinosaurs is actually not quite correct Not all reptiles of the Mesozoic were dinosaurs. Many reptiles (and other amniotes) have returned
LOWER CRETACEOUS OF SOUTH DAKOTA.
 A NEW DINOSAUR, STP^GOSAURUS MARSHl, FROM THE LOWER CRETACEOUS OF SOUTH DAKOTA. By Frederic A. Lucas, Curator, Divisioii of Coiiipnrative Anatomy, in charge, of Section of Vertebrate Fossils. The name
A NEW DINOSAUR, STP^GOSAURUS MARSHl, FROM THE LOWER CRETACEOUS OF SOUTH DAKOTA. By Frederic A. Lucas, Curator, Divisioii of Coiiipnrative Anatomy, in charge, of Section of Vertebrate Fossils. The name
Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved
 Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved This was a private report in 2003 on my thoughts on Platecarpus planifrons.
Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved This was a private report in 2003 on my thoughts on Platecarpus planifrons.
SUPPLEMENTARY INFORMATION
 In comparison to Proganochelys (Gaffney, 1990), Odontochelys semitestacea is a small turtle. The adult status of the specimen is documented not only by the generally well-ossified appendicular skeleton
In comparison to Proganochelys (Gaffney, 1990), Odontochelys semitestacea is a small turtle. The adult status of the specimen is documented not only by the generally well-ossified appendicular skeleton
Letting rotting sharks lie: further evidence for shark identity of the Zuiyo-maru carcass
 Letting rotting sharks lie: further evidence for shark identity of the Zuiyo-maru carcass Pierre Jerlström and Bev Elliott Another basking shark carcass washed up on the New Zealand coast reveals features
Letting rotting sharks lie: further evidence for shark identity of the Zuiyo-maru carcass Pierre Jerlström and Bev Elliott Another basking shark carcass washed up on the New Zealand coast reveals features
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
35. DATA REPORT: CRETACEOUS OSTRACODES FROM HOLES 865A AND 866A (MID-PACIFIC MOUNTAINS) 1. Renée Damotte 2
 1. Renée Damotte 2") Winterer, E.L., Sager, W.W., Firth, J.V., and Sinton, J.M. (Eds.), 1995 Proceedings of the Ocean Drilling Program, Scientific Results, Vol. 143 35. DATA REPORT: CRETACEOUS OSTRACODES FROM HOLES 865A AND
Winterer, E.L., Sager, W.W., Firth, J.V., and Sinton, J.M. (Eds.), 1995 Proceedings of the Ocean Drilling Program, Scientific Results, Vol. 143 35. DATA REPORT: CRETACEOUS OSTRACODES FROM HOLES 865A AND
A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China
 SUPPLEMENTARY INFORMATION A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China Ya-Ming Wang 1, Hai-Lu You 2,3 *, Tao Wang 4 1 School of Earth Sciences and Resources, China
SUPPLEMENTARY INFORMATION A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China Ya-Ming Wang 1, Hai-Lu You 2,3 *, Tao Wang 4 1 School of Earth Sciences and Resources, China
Marshall Digital Scholar. Marshall University. F. Robin O Keefe Marshall University,
 Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 2008 Cranial anatomy and taxonomy of Dolichorhynchops bonneri new combination, a polycotylid (Sauropterygia:
Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 2008 Cranial anatomy and taxonomy of Dolichorhynchops bonneri new combination, a polycotylid (Sauropterygia:
New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia
 1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
Biology Slide 1 of 50
 Biology 1 of 50 2 of 50 What Is a Reptile? What are the characteristics of reptiles? 3 of 50 What Is a Reptile? What Is a Reptile? A reptile is a vertebrate that has dry, scaly skin, lungs, and terrestrial
Biology 1 of 50 2 of 50 What Is a Reptile? What are the characteristics of reptiles? 3 of 50 What Is a Reptile? What Is a Reptile? A reptile is a vertebrate that has dry, scaly skin, lungs, and terrestrial
Mesozoic Marine Life Invertebrate Vertebrate
 Mesozoic Marine Life Invertebrate Vertebrate Cenozoic Marine Life - Invertebrates (Mollusks) Cenozoic Marine Life - Invertebrates (Arthropods) Cenozoic Marine Life - Vertebrates Marine fossils are abundant
Mesozoic Marine Life Invertebrate Vertebrate Cenozoic Marine Life - Invertebrates (Mollusks) Cenozoic Marine Life - Invertebrates (Arthropods) Cenozoic Marine Life - Vertebrates Marine fossils are abundant
Locomotion and skeletal morphology of Late Cretaceous mosasaur, Tylosaurus proriger
 Georgia Southern University Digital Commons@Georgia Southern University Honors Program Theses Student Research Papers 2017 Locomotion and skeletal morphology of Late Cretaceous mosasaur, Tylosaurus proriger
Georgia Southern University Digital Commons@Georgia Southern University Honors Program Theses Student Research Papers 2017 Locomotion and skeletal morphology of Late Cretaceous mosasaur, Tylosaurus proriger
NEW MATERIAL FROM THE TYPE SPECIMEN OF MEGALNEUSAURUS REX (REPTILIA: SAUROPTERYGIA) FROM THE JURASSIC SUNDANCE FORMATION, WYOMING
 FROM THE JURASSIC SUNDANCE FORMATION, WYOMING") Paludicola 7(4):170-180 May 2010 by the Rochester Institute of Vertebrate Paleontology NEW MATERIAL FROM THE TYPE SPECIMEN OF MEGALNEUSAURUS REX (REPTILIA: SAUROPTERYGIA) FROM THE JURASSIC SUNDANCE FORMATION,
Paludicola 7(4):170-180 May 2010 by the Rochester Institute of Vertebrate Paleontology NEW MATERIAL FROM THE TYPE SPECIMEN OF MEGALNEUSAURUS REX (REPTILIA: SAUROPTERYGIA) FROM THE JURASSIC SUNDANCE FORMATION,
A new skeleton of the cryptoclidid plesiosaur Tatenectes laramiensis reveals a novel body shape among plesiosaurs
 Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 2011 A new skeleton of the cryptoclidid plesiosaur Tatenectes laramiensis reveals a novel body shape
Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 2011 A new skeleton of the cryptoclidid plesiosaur Tatenectes laramiensis reveals a novel body shape
Animal Diversity III: Mollusca and Deuterostomes
 Animal Diversity III: Mollusca and Deuterostomes Objectives: Be able to identify specimens from the main groups of Mollusca and Echinodermata. Be able to distinguish between the bilateral symmetry on a
Animal Diversity III: Mollusca and Deuterostomes Objectives: Be able to identify specimens from the main groups of Mollusca and Echinodermata. Be able to distinguish between the bilateral symmetry on a
Red Eared Slider Secrets. Although Most Red-Eared Sliders Can Live Up to Years, Most WILL NOT Survive Two Years!
 Although Most Red-Eared Sliders Can Live Up to 45-60 Years, Most WILL NOT Survive Two Years! Chris Johnson 2014 2 Red Eared Slider Secrets Although Most Red-Eared Sliders Can Live Up to 45-60 Years, Most
Although Most Red-Eared Sliders Can Live Up to 45-60 Years, Most WILL NOT Survive Two Years! Chris Johnson 2014 2 Red Eared Slider Secrets Although Most Red-Eared Sliders Can Live Up to 45-60 Years, Most
Appendix 1. Peter Alsen
 Appendix 1 Description of a new Bajocian (Middle Jurassic) ammonite species, Cranocephalites tvaerdalensis sp.nov., from Geographical Society Ø, North-East Greenland. Peter Alsen A new Cranocephalites
Appendix 1 Description of a new Bajocian (Middle Jurassic) ammonite species, Cranocephalites tvaerdalensis sp.nov., from Geographical Society Ø, North-East Greenland. Peter Alsen A new Cranocephalites
NEW MATERIAL FROM THE TYPE SPECIMEN OF MEGAL1VEUSAURUS REX (REPTILIA: SAUROPTERYGIA) FROM THE JURASSIC SUNDANCE FORMATION, WYOMING
 FROM THE JURASSIC SUNDANCE FORMATION, WYOMING") Paludicola 7(4):170-180 May 2010 CO by the Rochester Institute of Vertebrate Paleontology NEW MATERIAL FROM THE TYPE SPECIMEN OF MEGAL1VEUSAURUS REX (REPTILIA: SAUROPTERYGIA) FROM THE JURASSIC SUNDANCE
Paludicola 7(4):170-180 May 2010 CO by the Rochester Institute of Vertebrate Paleontology NEW MATERIAL FROM THE TYPE SPECIMEN OF MEGAL1VEUSAURUS REX (REPTILIA: SAUROPTERYGIA) FROM THE JURASSIC SUNDANCE
The Geological Society of America Special Paper
 GSA_SP427_15_Meredith.qxd 8/8/07 12:16 PM Page 209 The Geological Society of America Special Paper 427 2007 The largest mosasaur (Squamata: Mosasauridae) from the Missouri River area (Late Cretaceous;
GSA_SP427_15_Meredith.qxd 8/8/07 12:16 PM Page 209 The Geological Society of America Special Paper 427 2007 The largest mosasaur (Squamata: Mosasauridae) from the Missouri River area (Late Cretaceous;
Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported
 Drawing of early ankle development of Alligator mississippiensis, as reported") Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported by a previous study 1. The intermedium is formed at
Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported by a previous study 1. The intermedium is formed at
First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia
 from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia") First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia Tsogtbaatar Chinzorig¹, ³ *, Yoshitsugu Kobayashi², Khishigjav Tsogtbaatar³,
First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia Tsogtbaatar Chinzorig¹, ³ *, Yoshitsugu Kobayashi², Khishigjav Tsogtbaatar³,
Juehuaornis gen. nov.
 34 1 2015 3 GLOBAL GEOLOGY Vol. 34 No. 1 Mar. 2015 1004 5589 2015 01 0007 05 Juehuaornis gen. nov. 1 1 1 2 1. 110034 2. 110034 70% Juehuaornis zhangi gen. et sp. nov Q915. 4 A doi 10. 3969 /j. issn. 1004-5589.
34 1 2015 3 GLOBAL GEOLOGY Vol. 34 No. 1 Mar. 2015 1004 5589 2015 01 0007 05 Juehuaornis gen. nov. 1 1 1 2 1. 110034 2. 110034 70% Juehuaornis zhangi gen. et sp. nov Q915. 4 A doi 10. 3969 /j. issn. 1004-5589.
Biology 1B Evolution Lecture 11 (March 19, 2010), Insights from the Fossil Record and Evo-Devo
, Insights from the Fossil Record and Evo-Devo") Biology 1B Evolution Lecture 11 (March 19, 2010), Insights from the Fossil Record and Evo-Devo Extinction Important points on extinction rates: Background rate of extinctions per million species per year:
Biology 1B Evolution Lecture 11 (March 19, 2010), Insights from the Fossil Record and Evo-Devo Extinction Important points on extinction rates: Background rate of extinctions per million species per year:
Title: Phylogenetic Methods and Vertebrate Phylogeny
 Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
8/19/2013. Topic 4: The Origin of Tetrapods. Topic 4: The Origin of Tetrapods. The geological time scale. The geological time scale.
 Topic 4: The Origin of Tetrapods Next two lectures will deal with: Origin of Tetrapods, transition from water to land. Origin of Amniotes, transition to dry habitats. Topic 4: The Origin of Tetrapods What
Topic 4: The Origin of Tetrapods Next two lectures will deal with: Origin of Tetrapods, transition from water to land. Origin of Amniotes, transition to dry habitats. Topic 4: The Origin of Tetrapods What
8/19/2013. Topic 5: The Origin of Amniotes. What are some stem Amniotes? What are some stem Amniotes? The Amniotic Egg. What is an Amniote?
 Topic 5: The Origin of Amniotes Where do amniotes fall out on the vertebrate phylogeny? What are some stem Amniotes? What is an Amniote? What changes were involved with the transition to dry habitats?
Topic 5: The Origin of Amniotes Where do amniotes fall out on the vertebrate phylogeny? What are some stem Amniotes? What is an Amniote? What changes were involved with the transition to dry habitats?
1/9/2013. Divisions of the Skeleton: Topic 8: Appendicular Skeleton. Appendicular Components. Appendicular Components
 /9/203 Topic 8: Appendicular Skeleton Divisions of the Skeleton: Cranial Postcranial What makes up the appendicular skeleton? What is the pattern of serial homology of the limbs? Tetrapod front limb morphology
/9/203 Topic 8: Appendicular Skeleton Divisions of the Skeleton: Cranial Postcranial What makes up the appendicular skeleton? What is the pattern of serial homology of the limbs? Tetrapod front limb morphology
GEOL 104 Dinosaurs: A Natural History Homework 6: The Cretaceous-Tertiary Extinction. DUE: Fri. Dec. 8
 GEOL 104 Dinosaurs: A Natural History Homework 6: The Cretaceous-Tertiary Extinction DUE: Fri. Dec. 8 Part I: Victims and Survivors Below is a list of various taxa. Indicate (by letter) if the taxon: A.
GEOL 104 Dinosaurs: A Natural History Homework 6: The Cretaceous-Tertiary Extinction DUE: Fri. Dec. 8 Part I: Victims and Survivors Below is a list of various taxa. Indicate (by letter) if the taxon: A.
BEHAVIORAL AND PALEOENVIRONMENTAL IMPLICATIONS OF REPTILE SWIM TRACKS FROM THE EARLY TRIASSIC OF WESTERN NORTH AMERICA
 Tracy Thomson attended the College of Eastern Utah and then received his B.Sc. in geology from the University of Utah. He is currently attending the University of California-Riverside and Dr. Mary Droser
Tracy Thomson attended the College of Eastern Utah and then received his B.Sc. in geology from the University of Utah. He is currently attending the University of California-Riverside and Dr. Mary Droser
Origin and Evolution of Birds. Read: Chapters 1-3 in Gill but limited review of systematics
 Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
complex in cusp pattern. (3) The bones of the coyote skull are thinner, crests sharper and the
 The bones of the coyote skull are thinner, crests sharper and the") DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
Revision of polycotylid plesioaur systematics (sauropterygia plesiosauria) and description of the axial osteology of a juvenile polycotylid
 and description of the axial osteology of a juvenile polycotylid") Marshall University Marshall Digital Scholar Theses, Dissertations and Capstones 2016 Revision of polycotylid plesioaur systematics (sauropterygia plesiosauria) and description of the axial osteology of
Marshall University Marshall Digital Scholar Theses, Dissertations and Capstones 2016 Revision of polycotylid plesioaur systematics (sauropterygia plesiosauria) and description of the axial osteology of
A Fossil Snake (Elaphe vulpina) From A Pliocene Ash Bed In Nebraska
 From A Pliocene Ash Bed In Nebraska") University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
Feature. Mesozoic marine reptiles from Spitsbergen and their ecosystems
 Feature Mesozoic marine reptiles from Spitsbergen and their ecosystems In the Mesozoic seas, the apex predators were reptiles. From the Arctic archipelago of Svalbard, the Spitsbergen Mesozoic Research
Feature Mesozoic marine reptiles from Spitsbergen and their ecosystems In the Mesozoic seas, the apex predators were reptiles. From the Arctic archipelago of Svalbard, the Spitsbergen Mesozoic Research
Evolution of Biodiversity
 Long term patterns Evolution of Biodiversity Chapter 7 Changes in biodiversity caused by originations and extinctions of taxa over geologic time Analyses of diversity in the fossil record requires procedures
Long term patterns Evolution of Biodiversity Chapter 7 Changes in biodiversity caused by originations and extinctions of taxa over geologic time Analyses of diversity in the fossil record requires procedures
Today there are approximately 250 species of turtles and tortoises.
 I WHAT IS A TURTLE OR TORTOISE? Over 200 million years ago chelonians with fully formed shells appeared in the fossil record. Unlike modern species, they had teeth and could not withdraw into their shells.
I WHAT IS A TURTLE OR TORTOISE? Over 200 million years ago chelonians with fully formed shells appeared in the fossil record. Unlike modern species, they had teeth and could not withdraw into their shells.
.56 m. (22 in.). COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition
. COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition") Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Vol. XIV, No. 1, March, The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S.
 By S.") Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Preliminary results on the stratigraphy and taphonomy of multiple bonebeds in the Triassic of Algarve
 Preliminary results on the stratigraphy and taphonomy of multiple bonebeds in the Triassic of Algarve Hugo Campos 1,2*, Octávio Mateus 1,2, Miguel Moreno-Azanza 1,2 1 Faculdade de Ciências e Tecnologia,
Preliminary results on the stratigraphy and taphonomy of multiple bonebeds in the Triassic of Algarve Hugo Campos 1,2*, Octávio Mateus 1,2, Miguel Moreno-Azanza 1,2 1 Faculdade de Ciências e Tecnologia,
Natural Sciences 360 Legacy of Life Lecture 3 Dr. Stuart S. Sumida. Phylogeny (and Its Rules) Biogeography
 Biogeography") Natural Sciences 360 Legacy of Life Lecture 3 Dr. Stuart S. Sumida Phylogeny (and Its Rules) Biogeography So, what is all the fuss about phylogeny? PHYLOGENETIC SYSTEMATICS allows us both define groups
Natural Sciences 360 Legacy of Life Lecture 3 Dr. Stuart S. Sumida Phylogeny (and Its Rules) Biogeography So, what is all the fuss about phylogeny? PHYLOGENETIC SYSTEMATICS allows us both define groups
d a Name Vertebrate Evolution - Exam 2 1. (12) Fill in the blanks
 Fill in the blanks") Vertebrate Evolution - Exam 2 1. (12) Fill in the blanks 100 points Name f e c d a Identify the structures (for c and e, identify the entire structure, not the individual elements. b a. b. c. d. e. f.
Vertebrate Evolution - Exam 2 1. (12) Fill in the blanks 100 points Name f e c d a Identify the structures (for c and e, identify the entire structure, not the individual elements. b a. b. c. d. e. f.
LEIDY, SHOWING THE BONES OF THE FEET 'AND LIMBS
 CQNTEUBUTIONS FBOM THE MUSEUM OF PALEONTOLOGY (Confindion of Con&&&m froin UB Muaercm of Gcologg) UNIVERSITY OF ' MICHIGAN VOL V, No. 6, pp. 6W3 (e ph.) DEAXMBER 31,1036 A SPECIMEN OF STYLEMYS NEBRASCENSIS
CQNTEUBUTIONS FBOM THE MUSEUM OF PALEONTOLOGY (Confindion of Con&&&m froin UB Muaercm of Gcologg) UNIVERSITY OF ' MICHIGAN VOL V, No. 6, pp. 6W3 (e ph.) DEAXMBER 31,1036 A SPECIMEN OF STYLEMYS NEBRASCENSIS
Master thesis by Patrick Ronander 2007
 A description and phylogenetic relationship of a polycotylid plesiosaur (Reptilia: Sauropterygia) from the Upper Cretaceous (Turonian) of Goulmima, Morocco Master thesis by Patrick Ronander 2007 Contents
A description and phylogenetic relationship of a polycotylid plesiosaur (Reptilia: Sauropterygia) from the Upper Cretaceous (Turonian) of Goulmima, Morocco Master thesis by Patrick Ronander 2007 Contents
On the cranial anatomy of the polycotylid plesiosaurs, including new material of Polycotylus latipinnis, Cope, from Alabama
 Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 2004 On the cranial anatomy of the polycotylid plesiosaurs, including new material of Polycotylus latipinnis,
Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 2004 On the cranial anatomy of the polycotylid plesiosaurs, including new material of Polycotylus latipinnis,
PRELIMINARY REPORT ON A CLUTCH OF SIX DINOSAURIAN EGGS FROM THE UPPER TRIASSIC ELLIO T FORMATION, NORTHERN ORANGE FREE STATE. J. W.
 41 Pa/aeont. afr., 22, 41-45 (1979) PRELIMINARY REPORT ON A CLUTCH OF SIX DINOSAURIAN EGGS FROM THE UPPER TRIASSIC ELLIO T FORMATION, NORTHERN ORANGE FREE STATE b y J. W. Kitching ABSTRACT A clutch of
41 Pa/aeont. afr., 22, 41-45 (1979) PRELIMINARY REPORT ON A CLUTCH OF SIX DINOSAURIAN EGGS FROM THE UPPER TRIASSIC ELLIO T FORMATION, NORTHERN ORANGE FREE STATE b y J. W. Kitching ABSTRACT A clutch of
Animal Form and Function. Amphibians. United by several distinguishing apomorphies within the Vertebrata
 Animal Form and Function Kight Amphibians Class Amphibia (amphibia = living a double life) United by several distinguishing apomorphies within the Vertebrata 1. Skin Thought Question: For whom are integumentary
Animal Form and Function Kight Amphibians Class Amphibia (amphibia = living a double life) United by several distinguishing apomorphies within the Vertebrata 1. Skin Thought Question: For whom are integumentary
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS
 AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.
REVISION OF REDONDASUCHUS (ARCHOSAURIA: AETOSAURIA) FROM THE UPPER TRIASSIC REDONDA FORMATION, NEW MEXICO, WITH DESCRIPTION OF A NEW SPECIES
 FROM THE UPPER TRIASSIC REDONDA FORMATION, NEW MEXICO, WITH DESCRIPTION OF A NEW SPECIES") Harris et al., eds., 2006, The Triassic-Jurassic Terrestrial Transition. New Mexico Museum of Natural History and Science Bulletin 37. REVISION OF REDONDASUCHUS (ARCHOSAURIA: AETOSAURIA) FROM THE UPPER
Harris et al., eds., 2006, The Triassic-Jurassic Terrestrial Transition. New Mexico Museum of Natural History and Science Bulletin 37. REVISION OF REDONDASUCHUS (ARCHOSAURIA: AETOSAURIA) FROM THE UPPER
Title. CitationJapanese Journal of Veterinary Research, 24(1-2): 37. Issue Date DOI. Doc URL. Type. File Information
: 37. Issue Date DOI. Doc URL. Type. File Information") Title DISTRIBUTION OF LYMPHATIC TISSUES IN DUCK CAECA Author(s)KITAMURA, Hirokazu; SUGIMURA, Makoto; HASHIMOTO, Yos CitationJapanese Journal of Veterinary Research, 24(1-2): 37 Issue Date 1976-05 DOI 10.14943/jjvr.24.1-2.37
Title DISTRIBUTION OF LYMPHATIC TISSUES IN DUCK CAECA Author(s)KITAMURA, Hirokazu; SUGIMURA, Makoto; HASHIMOTO, Yos CitationJapanese Journal of Veterinary Research, 24(1-2): 37 Issue Date 1976-05 DOI 10.14943/jjvr.24.1-2.37
Animal Diversity wrap-up Lecture 9 Winter 2014
 Animal Diversity wrap-up Lecture 9 Winter 2014 1 Animal phylogeny based on morphology & development Fig. 32.10 2 Animal phylogeny based on molecular data Fig. 32.11 New Clades 3 Lophotrochozoa Lophophore:
Animal Diversity wrap-up Lecture 9 Winter 2014 1 Animal phylogeny based on morphology & development Fig. 32.10 2 Animal phylogeny based on molecular data Fig. 32.11 New Clades 3 Lophotrochozoa Lophophore:
From Slime to Scales: Evolution of Reptiles. Review: Disadvantages of Being an Amphibian
 From Slime to Scales: Evolution of Reptiles Review: Disadvantages of Being an Amphibian Gelatinous eggs of amphibians cannot survive out of water, so amphibians are limited in terms of the environments
From Slime to Scales: Evolution of Reptiles Review: Disadvantages of Being an Amphibian Gelatinous eggs of amphibians cannot survive out of water, so amphibians are limited in terms of the environments
CURRICULUM VITAE SIMON SCARPETTA (July 2018)
") CURRICULUM VITAE SIMON SCARPETTA (July 2018) PhD Candidate in Paleontology Jackson School of Geosciences Email: scas100@utexas.edu RESEARCH AREAS AND INTERESTS Evolutionary biology, herpetology, paleontology,
CURRICULUM VITAE SIMON SCARPETTA (July 2018) PhD Candidate in Paleontology Jackson School of Geosciences Email: scas100@utexas.edu RESEARCH AREAS AND INTERESTS Evolutionary biology, herpetology, paleontology,
Field Trip: Harvard Museum of Natural History (HMNH)
") Field Trip: Harvard Museum of Natural History (HMNH) Objectives To observe the diversity of animals. To compare and contrast the various adaptations, body plans, etc. of the animals found at the HMNH.
Field Trip: Harvard Museum of Natural History (HMNH) Objectives To observe the diversity of animals. To compare and contrast the various adaptations, body plans, etc. of the animals found at the HMNH.
TOPOTYPES OF TYPOTHORAX COCCINARUM, A LATE TRIASSIC AETOSAUR FROM THE AMERICAN SOUTHWEST
 Lucas, S.G. and Spielmann, J.A., eds., 2007, The Global Triassic. New Mexico Museum of Natural History and Science Bulletin 41. TOPOTYPES OF TYPOTHORAX COCCINARUM, A LATE TRIASSIC AETOSAUR FROM THE AMERICAN
Lucas, S.G. and Spielmann, J.A., eds., 2007, The Global Triassic. New Mexico Museum of Natural History and Science Bulletin 41. TOPOTYPES OF TYPOTHORAX COCCINARUM, A LATE TRIASSIC AETOSAUR FROM THE AMERICAN
May 10, SWBAT analyze and evaluate the scientific evidence provided by the fossil record.
 May 10, 2017 Aims: SWBAT analyze and evaluate the scientific evidence provided by the fossil record. Agenda 1. Do Now 2. Class Notes 3. Guided Practice 4. Independent Practice 5. Practicing our AIMS: E.3-Examining
May 10, 2017 Aims: SWBAT analyze and evaluate the scientific evidence provided by the fossil record. Agenda 1. Do Now 2. Class Notes 3. Guided Practice 4. Independent Practice 5. Practicing our AIMS: E.3-Examining
Outline 17: Reptiles and Dinosaurs
 Outline 17: Reptiles and Dinosaurs Evolution of Reptiles The first reptiles appeared in the Mississippian. They evolved from amphibians, which first appeared in the Devonian. The evolutionary jump was
Outline 17: Reptiles and Dinosaurs Evolution of Reptiles The first reptiles appeared in the Mississippian. They evolved from amphibians, which first appeared in the Devonian. The evolutionary jump was
AMERICAN MUSEUM NOVITATES Published by
 AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes)
") Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
Living Dinosaurs (3-5) Animal Demonstrations
 Animal Demonstrations") Living Dinosaurs (3-5) Animal Demonstrations At a glance Students visiting the zoo will be introduced to live animals and understand their connection to a common ancestor, dinosaurs. Time requirement One
Living Dinosaurs (3-5) Animal Demonstrations At a glance Students visiting the zoo will be introduced to live animals and understand their connection to a common ancestor, dinosaurs. Time requirement One
290 SHUFELDT, Remains of Hesperornis.
 290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
click for previous page SEA TURTLES
 click for previous page SEA TURTLES FAO Sheets Fishing Area 51 TECHNICAL TERMS AND PRINCIPAL MEASUREMENTS USED head width (Straight-line distances) head prefrontal precentral carapace central (or neural)
click for previous page SEA TURTLES FAO Sheets Fishing Area 51 TECHNICAL TERMS AND PRINCIPAL MEASUREMENTS USED head width (Straight-line distances) head prefrontal precentral carapace central (or neural)
Are the dinosauromorph femora from the Upper Triassic of Hayden Quarry (New Mexico) three stages in a growth series of a single taxon?
 three stages in a growth series of a single taxon?") Anais da Academia Brasileira de Ciências (2017) 89(2): 835-839 (Annals of the Brazilian Academy of Sciences) Printed version ISSN 0001-3765 / Online version ISSN 1678-2690 http://dx.doi.org/10.1590/0001-3765201720160583
Anais da Academia Brasileira de Ciências (2017) 89(2): 835-839 (Annals of the Brazilian Academy of Sciences) Printed version ISSN 0001-3765 / Online version ISSN 1678-2690 http://dx.doi.org/10.1590/0001-3765201720160583
Ch 34: Vertebrate Objective Questions & Diagrams
 Ch 34: Vertebrate Objective Questions & Diagrams Invertebrate Chordates and the Origin of Vertebrates 1. Distinguish between the two subgroups of deuterostomes. 2. Describe the four unique characteristics
Ch 34: Vertebrate Objective Questions & Diagrams Invertebrate Chordates and the Origin of Vertebrates 1. Distinguish between the two subgroups of deuterostomes. 2. Describe the four unique characteristics
TRUE SKULL ROOF CONFIGURATION OF ICHTHYOSAURUS AND STENOPTERYGIUS AND ITS IMPLICATIONS
 Journal of Vertebrate Paleontology 25(2):338 342, June 2005 2005 by the Society of Vertebrate Paleontology TRUE SKULL ROOF CONFIGURATION OF ICHTHYOSAURUS AND STENOPTERYGIUS AND ITS IMPLICATIONS RYOSUKE
Journal of Vertebrate Paleontology 25(2):338 342, June 2005 2005 by the Society of Vertebrate Paleontology TRUE SKULL ROOF CONFIGURATION OF ICHTHYOSAURUS AND STENOPTERYGIUS AND ITS IMPLICATIONS RYOSUKE
BREVIORA LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1
, A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1") ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
ON SOME REPTILIAN REMAINS FROM THE DINOSAUR BEDS OF NYASALAND. By S. H. HAUGHTON, D.Sc., F.G.S.
 ( 67 ) ON SOME REPTILIAN REMAINS FROM THE DINOSAUR BEDS OF NYASALAND. By S. H. HAUGHTON, D.Sc., F.G.S. (Published by permission of the Hon. the Minister for Mines and Industries.) (With Plates II-V and
( 67 ) ON SOME REPTILIAN REMAINS FROM THE DINOSAUR BEDS OF NYASALAND. By S. H. HAUGHTON, D.Sc., F.G.S. (Published by permission of the Hon. the Minister for Mines and Industries.) (With Plates II-V and
Origin and Evolution of Birds. Read: Chapters 1-3 in Gill but limited review of systematics
 Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Everhart, Bite marks on an elasmosaur vertebrate palaeontology, 2, 2, (2005)
") Bite marks on an elasmosaur (Sauropterygia; Plesiosauria) paddle from the Niobrara Chalk (Upper Cretaceous) as probable evidence of feeding by the lamniform shark, Cretoxyrhina mantelli M.J. Everhart Sternberg
Bite marks on an elasmosaur (Sauropterygia; Plesiosauria) paddle from the Niobrara Chalk (Upper Cretaceous) as probable evidence of feeding by the lamniform shark, Cretoxyrhina mantelli M.J. Everhart Sternberg
Non-fiction: Sea Monsters. A new wave of fossils reveals the oceans prehistoric giants.
 Sea Monsters By Stephen Fraser A new wave of fossils reveals the oceans prehistoric giants. Way back when Tyrannosaurus rex shook the ground, another giant reptile lurked in the prehistoric oceans. A 50-foot
Sea Monsters By Stephen Fraser A new wave of fossils reveals the oceans prehistoric giants. Way back when Tyrannosaurus rex shook the ground, another giant reptile lurked in the prehistoric oceans. A 50-foot
2018 SVP Schedule of Events (subject to change) All events are held at the Albuquerque Convention Center unless otherwise noted with an **
 All events are held at the Albuquerque Convention Center unless otherwise noted with an **") 2018 SVP Schedule of Events (subject to change) All events are held at the Albuquerque Convention Center unless otherwise noted with an ** Tuesday, October 16 3:00pm 7:00pm 7:00pm 9:00pm Special Lecture
2018 SVP Schedule of Events (subject to change) All events are held at the Albuquerque Convention Center unless otherwise noted with an ** Tuesday, October 16 3:00pm 7:00pm 7:00pm 9:00pm Special Lecture
Name: GEOL 104 Dinosaurs: A Natural History Video Assignment. DUE: Mon. Oct. 29
 GEOL 104 Dinosaurs: A Natural History Video Assignment DUE: Mon. Oct. 29 Documentaries represent one of the main media by which scientific information reaches the general public. For this assignment, you
GEOL 104 Dinosaurs: A Natural History Video Assignment DUE: Mon. Oct. 29 Documentaries represent one of the main media by which scientific information reaches the general public. For this assignment, you
Current Taxonomic Status of the Plesiosaur Pantasaurus Striatus from the Upper Jurassic Sundance Formation, Wyoming
 Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 7-23 Current Taxonomic Status of the Plesiosaur Pantasaurus Striatus from the Upper Jurassic Sundance
Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 7-23 Current Taxonomic Status of the Plesiosaur Pantasaurus Striatus from the Upper Jurassic Sundance
F. Robin O'Keefe, Ph.D. Phone: Education Ph.D., University of Chicago B.S., Stanford University Professional Experience Associate Professor,
 Curriculum Vitae F. Robin O'Keefe, Ph.D. Associate Professor Department of Biological Sciences, Marshall University S267, One John Marshall Drive Huntington, WV 25755 Phone: 304 696 2427 email: okeefef@marshall.edu
Curriculum Vitae F. Robin O'Keefe, Ph.D. Associate Professor Department of Biological Sciences, Marshall University S267, One John Marshall Drive Huntington, WV 25755 Phone: 304 696 2427 email: okeefef@marshall.edu
VERTEBRATE READING. Fishes
 VERTEBRATE READING Fishes The first vertebrates to become a widespread, predominant life form on earth were fishes. Prior to this, only invertebrates, such as mollusks, worms and squid-like animals, would
VERTEBRATE READING Fishes The first vertebrates to become a widespread, predominant life form on earth were fishes. Prior to this, only invertebrates, such as mollusks, worms and squid-like animals, would
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * Dr. L.D. Boonstra. Paleontologist, South African Museum, Cape Town
 THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
ALFRED GILLETT AND FOSSILS FROM STREET
 ALFRED GILLETT AND FOSSILS FROM STREET This collection of local fossils was formerly in the Crispin Hall, Street. Most of these fossils came from Alfred Gillett (1814-1904), a retired ironmonger who lived
ALFRED GILLETT AND FOSSILS FROM STREET This collection of local fossils was formerly in the Crispin Hall, Street. Most of these fossils came from Alfred Gillett (1814-1904), a retired ironmonger who lived