Fine autoradiographical study on scale morphogenesis in the regenerating tail of lizards
|
|
|
- Prosper Waters
- 5 years ago
- Views:
Transcription

1 Histol Histopath (1 994) 9: Histology and Histopathology Fine autoradiographical study on scale morphogenesis in the regenerating tail of lizards L. Alibardi Depariment of Histology and Embryology, University of Sydney, Sydney, Australia Summary. Regenerating scales in lizards originate as pockets in the epidermis instead of epidermal elevations as during embryo development. The morphogenesis of scales in the regenerating tail of the lizards. Anolis and Lamphropholis was studied after peritoneal injection of 3~-thymidine. The tracer was localized in the forming epidermis after progressive post-injection times, by means of autoradiograph on plastic sections. After 4-5 hours post-injection of Y H-thymidine, the radioactivity was localized in the basal layer. After 2 to 4 days postinjection labelled cells were seen in the basal and intermediate spinosus layers but not in the uppermost keratinizing layers. Labelled cells were seen in the differentiating cornifying layers (pre-b and pre-a) 6-8 days post-injection. At days post-injection almost no radioactivity was seen in the basal layer or in the living part of the epidermis. A few labelled cells were present in the dense keratinizing layers of the sloughing wound and interscale lacunar layers. This study shows that scale formation and morphogenesis in the regenerating tail is brought about by a localized cell proliferation along the regenerating epidermis. In the forming scales the percentage of labelled cells in the distal side (future dorsal part of the new scale) is much higher than in the proximal side (the future ventral side of the scale), so that overlapped scales are generated. Key words: Lizard, Scale formation, Regenerating tail, Electron microscouv. Autoradiograuhv lntroduction During tail regeneration in lizards the new epidermis progressively differentiates new scales which resemble, with different degrees from species to species, the original ones (Quattrini, 1952, 1954). In most lizard species, the regenerated scales are different from the original ones both in macroscopical and microscopical - Offprint requests to: Dr. Alibardi Lorenzo. Departrnent of Biology, University of Padova, via Trieste 75, Padova, ltaly appearance (Quattrini, 1952; Werner, 1967). Despite this difference, sensory organs can also be reformed in the regenerated tail skin (Hughes and New, 1959; Maderson. 1971). Scale morphogenesis during regeneration of the tail in lizards presents some interesting problems. In fact, regenerating scales begin as epidermal invaginations instead of externa1 elevations as in normal development (Maderson, 1965, 1985; Dhouailly and Maderson, 1984). This modality of scale morphogenesis shows some analogies with the initial stages of hair development in mammals, due to the epidermal migration into the dermis (Bryant and Bellairs, 1967). The knowledge of scale morphogenesis during regeneration appears particularly interesting since squamate epidermis contains an outer layer of cornified cells containing B-keratin, resembling bird keratin, and an inner cornified layer made up by a-keratin, which resembles mammal keratin (Alexander and Parakkal, 1969; Alexander, 1970; Maderson et al., 1972; Maderson, 1985). The vertical sequence of B and a keratinized layers in the normal squamate skin has been divided into six arbitrary stages (Maderson, 1965, 1985; Landmann, 1979). Though the general development of the regenerating scales of lizards is known (Hughes and New, 1959; Shah and Chakko, 1967; Cox, 1969; Liu and Maneelly, 1969), many important details on the fine cytology, keratin types, morphogenesis, timing and cell migration during the scaling process are missing. The present research describes al1 these processes using the fine resolution of semithin plastic sections and autoradiography. This study shows that only the very early stages of scale formation resemble hair morphogenesis in mammals and that the two processes are completely different. In fact, the following differential pattern of cell division, cell shaping, differentiation and the spatial aggregation of cells in the proximal side (the future ventral side of the scale) and the distal side (the future well-keratinized dorsal side of the scale) of the

2 Autoradiographical study of regenerating lizard scales invaginations, produces asymmetric and overlapping scales. Materials and methods Adult individuals of the American iguanid lizard Anolis carolinensis and of the Australian scincid lizard Lamphopholis delicata and L. Gouchenoti were studied. The animals were kept in the laboratory, the former species at "C and the latter at "C, and fed with insect larvae. Some animals were peritoneally injected with a single dose of 3~-thymidine (10-15 pc/gbw; 3~-thymidine Amersham, sa 29 Cilmmole or Cilmmole) and tissues sampled after 4-5 hours post-injection as reported later. In other specimens, the original tail was removed by induced autotomy, and from the tail stump a new tail was left to regenerate until about days later, when the length of the regenerating tail was ranging from about 3 to 6 mm. At this stage the animals received a single intraperitoneal injection of 3~-thymidine as previously reported. Regenerating tails were fixed after 4-5 hours, 2-4, 6-8, and 20 days post-injection. The cool fixative (0-4 "C) consisted of a 2.5% solution of glutaraldehyde and 0.5% acrolein in reptile Ringer solution at ph After about 8 hours in cold fixation the tissues were immersed in 1% Oso4 for about 1-2 hours, dehydrated in alcohol and embedded in epon or spurr resins. After sectioning with LKB or Reichert ultramicrotomes, semithin sections containing regenerating scales were collected on clean slides. Some thin sections, to be studied with the electron microscope, were collected with a wire loop and deposited over slides previously coated with collodium. The sections were coated and prepared for electron microscopic autoradiography (Weakly, 1981). Semithin sections were coated in a darkroom with Ilford K5 nuclear emulsion, while thin sections (for electron microscopy) were coated with Ilford L4 nuclear emulsion. Exposure time ranged between 5-7 weeks for semithin sections and 3-5 months for thin sections. After the exposure time, the slides were developed with Kodak D 19 and fixed in Agfa fixer. From the L4-coated slides, thin sections were recuperated by stripping and floating the supporting collodium membrane on distilled water. The sections were picked up with copper grids, lightly stained with uranyl acetate and lead citrate and observed with a Jeol CXI 00 electron microscope operating at Kv. The autoradiographical quantification was done by counting the labelled cells in the different regions of the apical prescaling epidermis and in the forming scales of the regenerating tail (Fig. 1). The percentage of labelled cells (%L) was calculated over counted cells. The cells were counted only in the basal germinative layer of the epidermis (Germinative %L, Fig. la) or in al1 the layers of the epidermis (Global %L, Fig. lb). The %L of the distal and proximal sides of the regenerating scales, was obtained by adding the numbers of unlabelled and labelled cells of at least 3-7 consequent scales in each animal (in order to reach at least counted cells). Results Four-five hours post-injection The epidermis located by the apex of the regenerating tail, both in Anolis and Lampropholis, appeared pluristratified (7-10 cell layers) with the external layers more or less keratinized (Figs. 1, 2). The cells in the external and intermediate layers were more or less flattened and were unlabelled. Labelled and dividing cells were only present in the germinative or the basal layer of the new epidermis, as was confirmed with the electron microscope (Figs. 2-4). As reported in Fig. la,b, the highest %L was found in the basal layer of the epidermis behind the tip of the regenerating tail (mean of Germinative %L 42.8; mean of Global %L 11.8). The cells of the basal layer in the apical prescaling epidermis were in contact with an irregularly differentiated basement membrane and were often organized in a pseudo-stratified manner. In this layer most of mitosis also occurred. The tip of the regenerating tail (apical cup, Fig. la,b) showed scarce or no basal labelled cells. Lateral to the epidermal tip, in the crescent epidermis (Fig. la,b), labelled cells were more numerous (mean of Germinative %L 15.9; mean of Global %L 6.6). In these distal thinner epidermis (5-7 layers of epidermal cells), there was no sign of scale formation ti mm from the tail tip. Under the apical epidermis, mesenchimo-fibroblasts and many small clotted or lacunar capillaries were observed. Extravasated blood cells were commonly seen in the loose connective tissue underlying the apical epidermis, 1-2 mm from the tail tip. The formation of scales in a regenerating tail 21 days old began at about mm from the tail tip, and varied according to the length of the new tail at that stage (3-6 mm) and also from animal to animal. While the labelled cells in the apical non-scaled epidermis were located more or less uniformly along the basal germinative layer (beside the apical cup, Fig. l), in the prescaling epidermis it was observed that labelled cells were grouped to form alternated areas with unlabelled epidermal cells (Figs. 5, 6). These labelled spots of epithelial cells foreshadowed the formation of scales. In fact, in more proximal regions of the regenerating tail, the epidermis gradually folded and sank into the dermis forming invaginations or epidermal papillae. These latter progressively deepened and lengthened in a disto-proximal direction (Fig. Ic). Most of the labelled cells were concentrated in these invaginations (mean of Germinative %L 31.0; mean of Global %L 12.8) which gave a wave-like outline to the basement membrane separating the epidermis from the
3 Fig. 1. Schematic drawings of scale formation in the regenerating lizard tail. P, prescaling apical epidermis. 1, intermediate scale anlage region. S, proximal scaling epidermis. a, distal side made up of cells. A, distal side made up of cells. W, distal side made up of cells. e, distal side made up of cells. V, distal side made up of cells. Fig. la indicates the %L in the basal layer (black cuwes), (as mean f standard error; numbers in parentheses indicate the number of samples). L, Lampropholis. A, Anolis. Fig. 1 b indicates the %L referring to the whole epidermis layers (striped regions). Fig. Ic shows the general sequence of scale morphogenesis from the tail tip backward (rectangle). Legends: w, wound epidermis. L, lacunar tissue. p, proximal side. d, distal side. H, hinge region. do, dorsal side. v, ventral side. k, keratinized layer. The arrow points to the tail tip.

4 122 Autoradiographical study of regenerating lizard scales
 near the basal layer of the apical epidermis.")
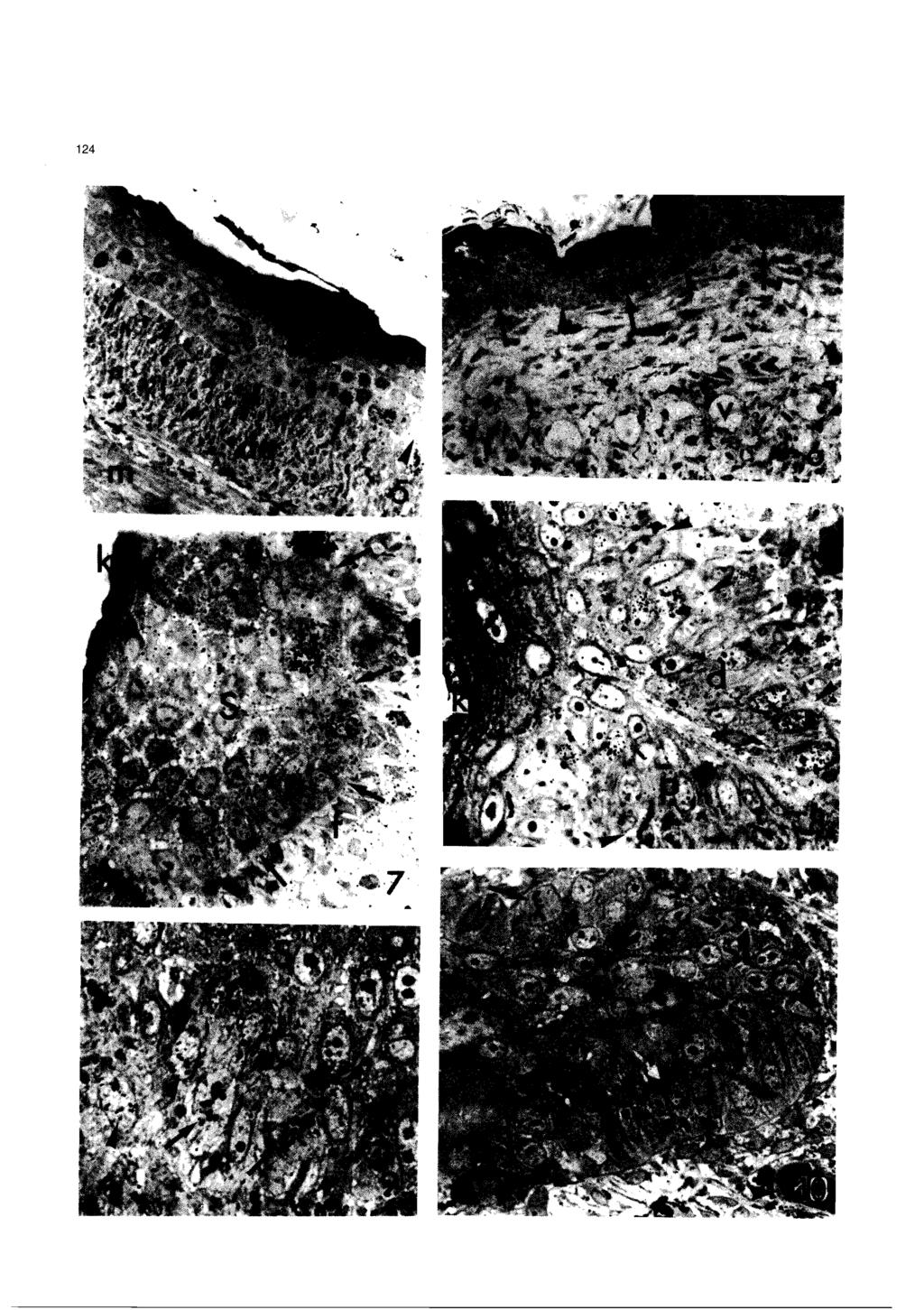
5 Autoradiographical study of regenerating lizard scales Fig. 2. Lampropholis. Pluristratified epidermis (e) of the apical regenerating tail. In the basal layer two labelled cells are seen (arrows). de, dermis; K, outer lightly keratinized layer. x 700 Fig. 3. Anolis. Electron microscopic view of two labelled cells (arrows) near the basal layer of the apical epidermis. Arrowheads on the incomplete basement membrane. S, cells of the stratum spinosum; de, dermal fibroblast. x 3,100 Fig. 4. Anolis. Close-up of labelled vacuolated epidermal cells, showing desmosomes (arrows). A few tonofilaments are seen in the cytoplasm. x 11,500 underlying dermis (Figs. 1, 6-8). Mitoses were observed with relatively high frequency in these epidermal papillae and many dermal fibroblasts were oriented more or less perpendicularly to the irregular basement membrane (Figs. 5, 9-1 1). Also, these fibroblasts often appeared more concentrated under the invaginated epidermal papillae than in the interpapillar regions where blood vessels were numerous (Fig. 10). The process of invagination and deepening of the epidermal papillae continued in a progressive fashion in a disto-proximal direction in the following days (Fig. 8). From the bottom part of the invagination (forming a hinge region), two columns (or sides) of epithelial cells reached the epithelial surface. The distal side was oriented by the tail tip while the proximal side was oriented by the animal body (Fig. lc). When the two sides were made up by cells they had a simil- number of labelled cells and dividing cells (Figs. 7, 8, lo), but with more cells, the labelled and dividing cells were more frequent in the distal than in the proximal side. Later the invagination appeared to progressively lose the initial symmetry and tilted backward (Figs. 12, 13). Also, in this columnar cell stage, the cells of the distal side appeared cuboidal or polygonal and larger than the smaller flat cells of the proximal side; this, contrast appeared particularly evident in the bottom part of the invagination (the presumptive hinge region of the forming scale, Figs. 14, 15). During the following stages of deepening of the epidermal invaginations, the proximal side became flatter and flatter, the cells progressively stretched and few of them appeared labelled. On the contrary, the distal side of the developing scale kept for most of the stages of development a higher %L and number of dividing cells than the proximal side; as a result the distal side lengthened more than the proximal side and the epidermal impocketings tilted further backward. The Germinative %L ranged between 7.0 and 23.5, depending on the degree of scale development (%L was higher in younger scales during rapid scale lengthening than in later stages, see representative values in Fig. la,b). Labelled cells were often seen close to the hinge region and in the apical region of the forming scale. The distal side showed the complete sequence of differentiating layers of the normal epidermis during the shedding cycle (Maderson, 1985; Landmann, 1986), with a hypertrophic germinative layer, and intermediate and outer keratinized a and B layers (Fig. 15). In the distal side of the forming scale the number of keratinizing strata was higher moving from the hinge region up toward the scale tip. The germinative layer contained mostly clear, and some darker cells of thinner diameter. In the proximal side, clear flat cells were seen in the stratified narrow layers over the basal layer. These paler stretched cells were in contact with the more stratified layers produced by the distal side (Figs. 14, 15). One or more blood vessels, clothed or of synusoidal aspect, were always seen on both sides of the scale. Furthermore, melanocytes were essentially localized under the epidermis of the distal side of the developing scale and were rarely seen under the proximal side b Fig. 5. Anolis. Group of labelled cells (arrows) in the pre-scaling epidermis. Arrowheads point to areas without labelled cells. de, dermis with fusiform fibroblasts; m, regenerating muscles. x 320 Fig. 6. Anolis. Cross-sectioned apical prescaling epidermis. Labelled cells (arrows) tend to concentrate in spots along the epidermis. Arrowheads point to non-labelled areas. v, cross sectioned dermal blood plexus. x 320 Fig. 7. Anolis. Beginning of invaginating epidermis (arrows) where some labelled cells are visible. Note the row of fibroblasts (f) just below the basal layer. S, spinosus cells; K, keratinized thin layer. x 850 Fig. 8. Lamphropolis. Epidermal invagination showing labelled cells in both proximal (p) and distal (d) sides. Arrowheads outline the basement membrane; K, superficial keratinizing wound layer. x 800 Fig. 9. Lamphropolis. Basal layer of apical non scaled epidermis showing mitotic cells (arrows). Two weakly-labelled cells are also seen (arrowheads). S, spinosus layers. x 1,100 Fig. 10. Lamphropolis. Two mitotic cells (arrows) in the proximal side (p) and in the distal side (d) of a deepening epidermal papilla. The two sides are roughly similar at this stage. The arrowhead points to a blood capillafy. x 800
6

7 Autoradiographical study of regenerating lizard scales 125
8 A utoradiographical study of regenerating lizard scales Fig. 11. Lamphropolis. Electron microscopic view of mitotic cell in the distal side of a forming scale. A few tonofibrils are seen in the cytoplasm of basal epidermis. Arrowheads point to the basement membrane. x 4,900 Fig. 12. Anolis. Two deepening epidermal papillae in the scaling region (the distal direction is uppeiward). Labelled cells (arrowheads) are essentially localized in the distal side (d). In the shorter proximal side (p) no labelled cells are seen. de, dermis; arrows point to a blood vessel. x 320 Fig. 13. Lamphropolis. Sequence of advanced proximo-dista1 scale formation by epidermal papillae (e) downgrowih migration into the dermis (de). K, keratinized wound epidermis. H, forming hinge region. P, proximal side. d, distal side. x 300 Fig. 14. Lamphropolis. Hinge region (H) of deepening scale at stage 2-3. Note most labelled cells (arrowheads) in the distal side (d) and only one in the proximal side (p). From the hinge region rightward the number of flat cell layers increases. On the proximal side flat pale cells are often seen (arrows). 6, interscale blood vessel. x 800 Fig. 15. Lamphropolis. Hinge region (H) of more developed scale at stage 3-4. Most of the labelled cells are in the distal side (d). o, possible Oberhautchen layer, B, pre-b layer; arrows on pale flat cells from the proximal side (p). x 830. (Fig. 16). Two-four days post-injection Both in non-scaled apical epidermis as well as in the scaling epidermis, labelled cells were found in the basal and in the intermediate (spinosus) layers of the epidermis above (Figs ). Due to cell division and upward migration, the intensity of the labelled nuclei was diminished in the basal layer with respect to 4-5 hours post-injection. Labelled cells of the spinosus layers were often more labelled than the basal cells. However, no labelled cells were seen in the forming flat keratinizing layer (presumptive or mature-b layer) of the regenerating scales. This was seen both in the distal and proximal sides of the forming scale at any stage of their development, in early as well as in the more developed scales (Fig. 2 1). Also, in the apical epidermis, near the regenerating tail tip, the basal cells appeared both in the basal and spinosus layer (Fig. 19). In the initial epidermis invaginations (forming papillae), the labelling was also present in the spinosus layer of the epidermal papillae. Six-eight days post-injection Labelled cells were commonly seen both in the apical epidermis and in the forming scales. Rare labelled cells were seen in the germinative layer. Though some labelled cells were still located in the layer just above the germinative layer (close to the hinge region which represents the youngest part of the forming scale), most of the labelled cells were seen in the intermediate and upper pre-keratinized (pre-b) layers. In the apical epidermis, where no scales were yet seen, labelled cells were often seen to contain less trace granules than in earlier stages. These cells were located in the intermediate layers, 3-4 cell layers above the germinative layer, and even in the upper flat prekeratinized cell layer (Figs ). Labelled cells were also seen in the pre-keratinizing layers of the developing distal side of the new scale (Fig. 24, pre-(3, located 3-4 layers above the germinative, during stage 4 according to Maderson, 1985). Labelled cells were less frequently seen in the thin layer just over the germinative and in pre-keratinizing layers by the tip Fig. 16. Anolis. Forming scale two days post-injection. Stratified labelled cells (arrows) are seen throughout the distal side (d). No labelled cells are in the flat pre-r layer (B) nor in the proximal side (p) of the scale. Note the melanophores (arrows) mostly under the distal side of the scale. B, blood vessel. S, spinosus lacunar layer of the outer interscaling epidermis. x 400 Fig. 17. Anolis. Prescaling apical epidermis two days post-injection. Five strongly labelled cells (arrowheads) are located in the intermediate (spinosus) layer, just above the germinative layer, this latter still showing fewer labelled cells (arrows). K, keratinized ~~wound~~ epidermis. de, dermis. x 800 Fig. 18. Anolis. High concentration of labelled cells throughout the living part of the distal side (d) in a forming scale two days post-injection. Most labelled cells are in the intermediate layer (arrowheads) more than in the basal layer (ba). Only one labelled cell is seen in the proximal side (p) and no labelled cells are in pre-b and the putative Oberhautchen (o) layer. S, spinosus lacunar cells of the outer interscaling epidermis. x 800 Fig. 19. Anolis. Electron microscopic view of a labelled cell (arrowheads) in the intermediate spinosus layer two days post-injection. Stretched intercellular spaes (i) among epidermal cells are joined by desmosomes (arrows). t, tonofilaments; M, melanosome arrms. x 13,400 Fig. 20. Anolis. Labelled cells (arrowheads) in the spinosus layer of pre-scaling epidermis aíter four days post-injection. No labelled cells are in the uppermost keratinized layer (K). Arrows point to the basal epidermal layer. x 760 Fig. 21. Lamphropolis. Proximo-dista1 sequence (upward) of the splitting lines (arrowheads) from the hinge region (H) of forming scales into new scales. The distal side (d) of the epidermal papillae becomes the dorsal outer part of the new scale (S). The proximal side (p) forms the ventral inner part of the scale (v). x 200

9

10
 are in the spinosus layer, 4-5 cell layers above the basal layer (arrowheads). m, epiderrnal rnelanocyte. K, upper keratinized wound epidermis. x 830 Fig. 23. Lamphropolis.")
11 A utoradiographical study of regenera ting lizard scales Fig. 22. Lamphropolis. Prescaling apical epidermis six days post-injection (the distal direction is downward). Two labelled cells (arrows) are in the spinosus layer, 4-5 cell layers above the basal layer (arrowheads). m, epiderrnal rnelanocyte. K, upper keratinized wound epidermis. x 830 Fig. 23. Lamphropolis. Prescaling wound epidermis at six days post-injection (the distal direction is leftwards). Two streched labelled cells (arrows) are seen under the keratinized layer, at 6-7 cell layers frorn the basal stratum. S, spinosus cells; m, intra-epiderrnal rnelanocyte. x 850 Fig. 24. Lamphropolis. Hinge region (H), with epidermis of the distal side (d) at stage 4, six days post-injection. Labelled cells (arrows) are obse~ed in the forrning pre-b-layer (B) and in the underlying differentiating layer. In the proxirnal side (p) a labelled cell (arrow) is seen in contact with the pale flat cells. Arrowhead points to an interscale capillary. x 850 Fig. 25. Lamphropolis. Hinge region (H) with well keratinized (K) epiderrnis at stage 5-6, six days post-injection. In the distal side (d) labelled cells (arrows) are visible under the cornified layer and in the underlying differentiating (pre-a) layer. Two labelled cells (arrows) are also seen in the flat proximal side (p). Arrowheads on rnelanocytes under the basal layer. x 850 Fig. 26. Lamphropolis. Area by the tip of a well keratinized (k) scale, six days post-injection. In the distal (dorsal) side of the scale (d) labelled cells (arrows) are located in the differentiating a-layer. In the proximal side (p), near the scale tip, two flat labelled cells (arrows) are incorporated into the keratinized layer. The arrowhead points to the tip of the next scale. x 800 of the new scale (Fig. 26). No labelled cells were seen in the germinative layer at this stage and only occasional labelled cells were located in the maturing layer just above the germinative layer or in the scale tip. Also, the flat proximal side of the forming scale contained some labelled cells (Figs ). After 6-8 days, labelled flat cells in the proximal side were seen in the basal and upper narrow layers facing the B-cells of the distal side (Fig. 28). The latter labelled cells sometimes showed a diminished number of trace granules. Twelve-fourten and twenty days post-injection Occasional labelled cells were seen after this period both in the apical non-scaled epidermis of the regenerating tail as well as in more proximal regions where more or less differentiated scales were present. In general, the labelling was quite diluted over the nuclei and labelled cells were observed essentially in the apical prekeratinized layers of the outer shedding layer that covered the forming scales; the so called «wound epithelium and lacunar layern (Maderson et al., 1978; Figs. 1, 29,30). Rare labelled cells appeared near the keratinized layers, at the tip of the new scales, whilst rare labelled cells were seen in some layers of the deeper intermediate and hinge regions. After twenty days post-injection, there was almost no trace of radioactivity in the skin. Later, up to 50 days of tail regeneration in the present study, regenerated scales continued to lengthen in their dorsal outer side, so that they became less and less overlapped (Figs. 21, 31), finally resulting in the hinge region and a short portion of the scale being covered by the proximal new scales. Discussion Cell multiplication and morphogenesis of regenerating scales The present autoradiographical analysis throws new light on the mechanism of scale morphogenesis in the regenerating tail of lizards (see Figs. 1, 32a). In fact, both in Anolis and in Lampropholis, the downgrowth of epidermal papillae into the dermis seems initially to derive from a more or less localized cell multiplication in alternated spots along the regenerating apical epidermis. The high Germinative %L in these areas with respect to the epidermis in the tail tip (apical cup, see Fig. 1) suggests that the epidermis grows faster in these regions and that a folding process may take place. These results Fig. 27. Lamphropolis. Medial part of regenerating scale at stage 6. After 8 days post-injection, labelled cells are visible in the forrning a-layer (a) under the thick B-layer. Note the hypertrophy of the pale basal layer of the distal side (d) and the underlying rnelanocytes. The arrow points to a labelled cell in the proximal side. x 900 Fig. 28. Lamphropolis. Hinge region with a distal side at stage 2-3. In this section after 8 days post-injection, three labelled cells are visible in flat layers of the proximal side and one (arrowhead) has reached the forming B-layer of the distal side (d). x 800 Fig. 29. Lamphropolis. Apical interscale region (p indicates the forrning proximal side of a scale, d points to the distal side of the other scale). After 12 days post-injection, weakly-labelled cells are visible in the <,lacunar tissue,> (S) under the externa1 -wound epidermis>>. x 830 Fig. 30. Lamphropolis. Another interscale region showing four labelled cells (arrows) in the spinosus (dacunar tissue)> (S), 12 days post-injection. d points to the distal side of a scale. x 830 Fig. 31. Lamphropolis. General view of a new scale in the proximal region of a regenerating tail after 50 days of regeneration. The superficial wellkeratinized dorsal side (S) of the scale has lengthened rnuch more than the ventral side (v). Arrowheads points to the shedding <cwound epitheliurn,,. H, hinge regions. de, dermis. m, rnuscles. x 170

12 130 Autoradiographical study of regenerating lizard scales
13 Autoradiographical study of regenerating lizard scales Fig. 32a Fig.32b Fig. 32. Schematic drawing of cell proliferatiion (a) and movement (b) during scale morphogenesis and at the apex of the regenerating tail (c). Fig. 32a features 6 arbitrary stages of scale morphogenesis and dots represent areas where labelled cells (and mitosis, M) are more frequent. Arrows point to the forming B-layer where the medial splitting will take place. In Fig. 32b the long cu~ed arrows indicate the direction of cell migration while the symbols (A,., e, e) refer to the periods to reach that level. In Fig. 32c the movements (arrows) of epidermal cells in the apical region of the regenerating tail are indicated. Legends: B, basal layer. d, dista1 side, p, proximal side. do, dorsal side. v, ventral side. H, hinge region. W, wound epidermis. L, lacunar tissue. ms, medial splitting. A, 4-5 hours. m, 2-4 days., 6-8 days. e days. 4-i-W :intercalar cell division :upperward cell division. Fig. 32c -:downward cell division. :cell hypertrophy. t : cell flattening.

14 Autoradiographical study of regenerating lizard scales accord well with those of Cox (1969), who found a Labelling Index of 5-15 in the epidermis of the regenerating tail in Anolis. The morphogenetic mechanism that determines the downgrowth of the regenerating epidermis appears to be different from that of the normal development of scales, where an epidermal outgrowth is produced (Maderson, 1965; Bryant and Bellairs, 1967; Dhouailly and Maderson, 1984). In their study Lih and Maneelly (1969) hypothesized that the soft dermis of the regenerating tail was a more permissive environment to epidermal invagination than the outer harder cornified wound epithelium (which is strengthened by cell junctions). The specific localization of dermal melanocytes essentially under the distal side of the forming scale. gives some indications that the dermis may play a role in this process, but further study is required. After the initial invagination of the linear epidermal surface of the apical epidermis, the distal side of the invaginations keeps most of the dividing cells, whilst in the proximal side cell proliferation decreases. Though the mechanism that is responsible for the latter phenomenon is unknown, this results in some morphogenetic effects which are completely different from those in hair development (see Bryant and Bellairs, 1 967). First, the distal side will legthen much more than the proximal side because of the intercalar cell multiplication. As a results of this process, the distal side keeps growing for a long time and will extends itself in order to form the future dorsal outer side of the regenerated scale. This is shown by the greater %L still present in the distal side of proximal scales in more advanced stages of development (Fig. 1 a,b). Secondly, the asymmetncal growth of the two sides will result in a backward tilting movement of the axes of the forming scale. so that the orientation will set the hinge region more proximal than the growing tip of the new scale (Figs 1, 13, 21, 32). Therefore the new scales will be initially superimposed and later, after the splitting in the middle of the scales, the continuous growth of the distal dorsal part will extend the new scale (Figs. 21, 31). Thirdly, the cells in the proximal side, where a lower cell multiplication occurs, become flatter and probably stretch each other in order to keep pace with the rapid distension of the opposite distal side. The proximal side (future inner-ventral side of the new scale) will result as a flat layer of pale stretched cells, as is typical for this region in snakes and lizards where a simplified cell stratification occurs (Roth and Jones, 1967). Though the ventral side of the scale shows a basal layer, this latter will generate layers of flat cells. Conversely, the distal side appears thicker and the basal layer is typically made up of polygonal, often hypertrophied cells. Their multiplication produces the characteristic cell stratification of the normal epidermis that follows the typical sequence of the shedding cycle (Landmann, 1979; Maderson, 1985). During the intercalar lengthening of the dorsal side of the regenerating scale, a concomitant upward epidermal stratification occurs. This process leads to the formation of a B, (probably of a mesos) and a-layers, lacunar and clear layers toward the axial region of the forming scale. The differentiation of an outermost Oberhautchen layer in the middle of the regenerating scale is a problem that will be solved only by means of electron microscopy. These differentiating layers have been observed in the distal side, from the hinge region to the tip of the new scale. Conversely, this typical sequence is not seen over the flat cells of the basal layer of the proximal side over which other flat and long pale cells are stratified. The nature of these cells is not known at the moment but further studies with the electron microscope should clarify this problem. Timing and epithelial cell migration During epidermal and scale regeneration the following general consideration on the timing and direction of migrating epithelial cells are drawn from the present study (Fig. 32b,c). By the tip of the regenerating tail, the proliferating epidermis mostly supplies cells that move distally and upwards, as represented in Fig. 32c. This has been suggested by the stretched appearance of normal or labelled cells in the prekeratinized layers, which are oriented in an oblique fashion after 6 days post-injection (Figs. 22, 23). In more proximal regions (0.5 mm from the tail tip), the epidermal cells do not contribute so much to tip lengthening, and cell multiplication is mostly directed upwards to the epidermal surface (Fig. 32c). The intense cell multiplication in these areas determines the downgrowth movement leading to the formation of new scales. In the distal and proximal sides of scales, cell multiplication in the basal layer is intercalar and some daughter cells remain in this layer whilst a few others migrate upwards to the media1 axis of the forming scale and downwards, thus lowering the hinge region. In fact, lower labelled cells in the basal layer were seen 2-4 and even 6 days post-injection. In the proximal side, the cells directed toward the media1 axis of the forming scale produce, in 6-8 days, a thin cornifying layer that faces the thicker keratinized layer of the opposite distal side. In the distal side after days postinjection, labelled cells appear localized according to the stage of the shedding cycle of the regenerating scale in which the 3~-thymidine was injected. As previously reported, in normal skin of the lizard Anolis (Flaxman, 1972; Flaxman and Maderson, 1973) and the snake Constrictor (Dowining and Roth, 1974) after 1 and 24 hours from the administration of 3 ~ - thymidine, labelled cells were observed only in the basal layer, despite the stage of the shedding cycle in which

15 A utoradiographical study o f regenera ting lizard scales the in-jection was done. This has been confirmed in the present study after 4-5 hours post injection. Both in lizards and snakes, cells differentiate following the order of formation in the shedding cycle after the injection of 3~-thymidine. In normal epidermis, the migration of the labelled cells from the basal layer to the cornified upper layers does not take place before 3-5 days and the labelling index of the basal layer increases from less than 10% to 40-50% during this period (Flaxman, 1972; Flaxman and Maderson, 1973). When the injection of 3~-thymidine is done at the early-mid resting stage (stage 1 according to Maderson, 1985; Landmann, 1986) only the Oberhautchen, B and mesos layers of the outer epidermal generation are completed, but not the a-layers. Therefore, days later labelled cells appeared in the forming a, clear and lacunar stages of the outer epidermal generation. These were the first labelled cells that appeared. When the injection was delayed to the mid-late resting stage, labelled cells appeared in the clear layer of the outer epidermal generation and in the Oberhautchen and B layers of the inner epidermal generation. Labelled cells in the mesos and a cells of the inner epidermal generation appeared when 3~-thymidine was administered during the renewal phase (Flaxman, 1972). In the present study a single pulse of 3~-thymidine to an animal with scales in a proximo-dista1 gradient of development (Fig. la-c). would label cells of the distal side in different intrinsic situation of the shedding cycle. However, cell behaviour and kinetics should resemble renewal in normal epidermis. Furthermore, over the first outer generation of a forming scale a provisional wound and lacunar epidermis are also present (Maderson and Roth, 1972), which constitute the shiny, soft, and unscaled epidermis of the young regenerating tail (Shah and Chakko, 1967; Maderson et al., 1978). The present observations have shown that in the apical non-scaled epidermis and in the distal side of the forming scales (before the first shed of the wound and lacunar epidermis), epidermal cells have already moved from the basal layer into the living layer above in 2 days. The labelled cells that still remain in the basal layer often show a reduction of radioactivity, indicating that part of the tracer has been incorporated in daughter cells that have moved one-two layers upwards. After 4 days, labelled cells reached 3-4 cell layers above the germinative (spinosus cells) and after 6-8 days post-injection they reached the cornifying outer layers, both in the apical, prescaling and scaling epidermis. After 6-8 days post-injection or later, labelled cells tended to disappear in the apical epidermis of the tail tip. In the prescaling and scaling epidermis after 6-8 days post-injection, labelled cells appeared as pre-b cells, premesos or pre-a cells, according to the stage of scale differentiation present at the moment of the injection of the tracer. Owing to the proximo-dista1 gradient of scale morphogenesis, at the moment of 3~-thymidine injection, the distal side of the regenerating scales were at stages 2-4. and come even during stage 5 of the shedding cycle (Bryant and Bellairs, 1967; Maderson, 1985). Despite this, labelled cells reached the differentiating cornified layers (B or a) after 6-8 days while a few labelled cells were still observed in the basal layer, generally with a reduced number of trace granules with respect to 4-5 hours post-injection. In the more proximal scales (stages 5-6) when the B layer was forming during the period of the injection, labelled cells appeared only in the underlying a forming layers (maybe also the mesos layer). After days post-injection rare labelled cells were still seen in the basal and intermediate differentiating layers (pre-b and pre-a), while weakly labelled tlat cells were limited to the interscale lacunar and outermost wound layers before shedding. This suggests that these superficial cells have moved toward the surface from the deeper regions of the forming scale after being pushed out by the progressive formation of the underlying keratinized layers (Fig. 32b). The low number of trace granules seen in these cells is probably due to the labelling dilution by some multiplication during the upward movement from the lower epidermal layers. In the non-scaled epidermis some cell divisions were observed among spinosus cells, up to three cell layers above the germinative stratum. After 20 days post-injection the lack of radioactivity in more differentiated (proximal) scales was due to the disruption of the nuclei in the cells that reached the cornified layers and to the first shedding of the regenerating epidermis in the proximal regions of the regenerating tail, after days of tail regeneration. Acknowledgements. The study on Anolis was initiated in the USA (Department of Biological Sciences, University of Illinois, Chicago) during an italian CNR scholarship period and the UIC financia1 support. The tissues of Lampropholis were collected in Australia (University of Sydney, Department of Histology and Embryology) during a scholarship period offered by the Australian DEET and the ltalian MAE. Many thanks to Mr. Claudio Friso for the drawings, Dr. Paolo Romandini for assistence with statistics and Miss Tamsin Braisher for reading the English text. References Alexander N.J. (1970). Comparison of a and O Keratin in reptiles. Z. Zellforsch. 11 0, Alexander N.J. and Parakkal P.F. (1969). Formation of a and O type keratin in lizard epidermis during the molting cycle. Z. Zellforsch. 1 01, Bryant S.V. and Bellairs A. d'a. (1967). Tail regeneration in the lizards Anguis fragilis and Lacerta dugesii J. Linn. Soc. (Zool) 46, Cox P.G. (1969). Some aspects of tail regeneration in the lizard Anolis carolinensis. 1) A description based on histology and autoradio-
16 A utoradiographical study of regenerating lizard scales graphy. J. Exp. Zool. 171, Dhouailly D. and Maderson P.F.A. (1984). Ultrastructural obse~ations on the ernbryonic developrnent of the integurnent of Lacerta muralis (Lacertilia, Reptilia). J. Morphol. 179, Downing S.W. and Roth S.I. (1974). The derivation of the cells of the epiderrnal strata of the boa constrictor (Constrictor constrictor). J. Inv. Dermatol. 62, Flaxrnan A.B. (1972). Cell differentiation and its control in the vertebrate epidermis. Arn. Zool. 12, Flaxman B.A. and Maderson P.F.A. (1973). Relationship between pattern of cell rnigration frorn the germina1 layer and changing pattern of differentiation in the lizard epidermis. J. Exp. Zool. 183, Hughes A. and New D. (1959). Tail regeneration in the gecknonid lizard, Sphaerodactylus. J. Ernbryol. Exp. Morphol. 7, Landrnann L. (1979). Keratin forrnation and barrier mechanisrns in the epidermis of Natrix natrix (Reptilia: serpentes): an ultrastructural study. J. Morphol. 162, Landrnann L. (1986). The skin of reptiles, epidermis and dermis. In: Biology of the integurnent. Vol. 2. Bereiter-Hann J., Matoltsy A.G. and Sylvia-Richards K. (eds). Springer Verlag. Liu H.-C. and Maneely R.B. (1969). Obse~ations on the developrnent and regeneration of tail epidermis in Hemidactilus bowringi (Gray). Acta Anat. 72, Maderson P.F.A. (1965). The ernbryonic development of the squarnate integurnent. Acta Zool. 46, Maderson P.F.A. (1971). The regeneration of caudal epidermal specializations in Lygodactylus picturatus keniensis (Gekkonidae, Lacertilia). J. Morphol. 134, Maderson P.F.A. (1985). Sorne developmental problerns of the reptilian integurnent. In: Biology of Reptilia. Vol. 14. Gans C., Billet F. and Maderson P.F.A. (eds). John Wiley and Sons. New York. pp Maderson P.F.A. and Roth S.I. (1972). A histological study of early cutaneous wound healing in lizards in vivo and in vitro. J. Exp. Zool. 180, Maderson P.F.A., Flaxrnan B.A., Roth S.I. and Szabo G. (1972). Ultrastructural contributions to the identification of cell types in the lizard epiderrnal generation. J. Morphol. 136, Maderson P.F.A., Baranowitz S. and Roth S.I. (1978). A histological study of the long response to trauma of squarnate integurnent. J. Morphol. 157, Quattrini D. (1952). Ricerche anatorniche e sperirnentali sulla autotornia della coda delle lucertole. 1) Dinarnica del1 'autotornia e conseguence su1 tegumento (osse~azioni nella Lacerta sicula Raf.). Arch. Zool. Ital. 37, Quattrini D. (1954). Piano di autotomia e rigenerazione nella coda dei sauri. Arch. Ital. Anat. Ernbriol. 59, Roth S.I. and Jones W.A. (1967). The ultrastructure and enzyrnatic activity of the boa constrictor (Constrictor constrictotj skin during the resting phase. J. Ultrastruc. Res. 18, Shah R.V. and Chakko T.V. (1967). Histological 0bSeNati0flS on the normal and regenerating tail of the house lizard, Hemidactylus flaviviridis. J. Anirn. Morphol. Physiol. 15, Weakley B.S. (1981). A beginner handbook in biological electron rnicroscopy. Churchill-Livingstone. Werner Y.L. (1967). Regeneration of specialized scales in tails of Teratoscincus (Reptilia: Gekkonidae). Senck. Biol. 48, Accepted September 17, 1993
Development, comparative morphology and cornification of reptilian claws in relation to claws evolution in tetrapods
 Contributions to Zoology, 78 (1) 25-42 (2009) Development, comparative morphology and cornification of reptilian claws in relation to claws evolution in tetrapods Lorenzo Alibardi 1, 2 1 Dipartimento di
Contributions to Zoology, 78 (1) 25-42 (2009) Development, comparative morphology and cornification of reptilian claws in relation to claws evolution in tetrapods Lorenzo Alibardi 1, 2 1 Dipartimento di
EFFECTS OF TEMPERATURE ON GROWTH IN THE REGENERATING TAIL OF THE SCINCID LIZARD, MABUYA STRIATA. Accepted: June 1977
 EFFECTS OF TEMPERATURE ON GROWTH IN THE REGENERATING TAIL OF THE SCINCID LIZARD, MABUYA STRIATA D K MAGON Department of Zoology. Kenyatta University College. Box 43844. Nairobi. Kenya Accepted: June 1977
EFFECTS OF TEMPERATURE ON GROWTH IN THE REGENERATING TAIL OF THE SCINCID LIZARD, MABUYA STRIATA D K MAGON Department of Zoology. Kenyatta University College. Box 43844. Nairobi. Kenya Accepted: June 1977
BEAK AND FEATHER DYSTROPHY IN WILD SULPHUR-CRESTED COCKATOOS (CACATUA GALERITA)
") BEAK AND FEATHER DYSTROPHY IN WILD SULPHUR-CRESTED COCKATOOS (CACATUA GALERITA) Author(s): Steven McOrist, Douglas G. Black, David A. Pass, Peter C. Scott, and John Marshall Source: Journal of Wildlife
BEAK AND FEATHER DYSTROPHY IN WILD SULPHUR-CRESTED COCKATOOS (CACATUA GALERITA) Author(s): Steven McOrist, Douglas G. Black, David A. Pass, Peter C. Scott, and John Marshall Source: Journal of Wildlife
A comparison of placental tissue in the skinks Eulamprus tympanum and E. quoyii. Yates, Lauren A.
 A comparison of placental tissue in the skinks Eulamprus tympanum and E. quoyii Yates, Lauren A. Abstract: The species Eulamprus tympanum and Eulamprus quoyii are viviparous skinks that are said to have
A comparison of placental tissue in the skinks Eulamprus tympanum and E. quoyii Yates, Lauren A. Abstract: The species Eulamprus tympanum and Eulamprus quoyii are viviparous skinks that are said to have
A Scanning Electron Microscopic Study of Eggshell Surface Topography of Leidynema portentosae and L. appendiculatum (Nematoda: Oxyuroidea)
") The Ohio State University Knowledge Bank kb.osu.edu Ohio Journal of Science (Ohio Academy of Science) Ohio Journal of Science: Volume 88, Issue 5 (December, 1988) 1988-12 A Scanning Electron Microscopic
The Ohio State University Knowledge Bank kb.osu.edu Ohio Journal of Science (Ohio Academy of Science) Ohio Journal of Science: Volume 88, Issue 5 (December, 1988) 1988-12 A Scanning Electron Microscopic
Lesson 7. References: Chapter 6: Chapter 12: Reading for Next Lesson: Chapter 6:
 Lesson 7 Lesson Outline: Embryonic Origins of the Dermis Specializations of the Dermis o Scales in Fish o Dermal Armour in Tetrapods Epidermal/Dermal Interactions o Feathers o Hair o Teeth Objectives:
Lesson 7 Lesson Outline: Embryonic Origins of the Dermis Specializations of the Dermis o Scales in Fish o Dermal Armour in Tetrapods Epidermal/Dermal Interactions o Feathers o Hair o Teeth Objectives:
Development of the Intestinal Villi Associated
 Development of the Intestinal Villi Associated with the Increased Epithelial Cell Mitosis in Chickens Koh-en YAMAUCHI, Eiji NAKAMURA and Yutaka ISSHIKI Laboratory of Animal Science, Faculty of Agriculture,
Development of the Intestinal Villi Associated with the Increased Epithelial Cell Mitosis in Chickens Koh-en YAMAUCHI, Eiji NAKAMURA and Yutaka ISSHIKI Laboratory of Animal Science, Faculty of Agriculture,
Keywords: lizards; limb regeneration; FGF administration; histology; 5BrdU-immunohistochemistry
 Article FGFs Treatment on Amputated Lizard Limbs Stimulate the Regeneration of Long Bones, Opening New Avenues for Limb Regeneration in Amniotes: A Morphological Study Lorenzo Alibardi Comparative Histolab
Article FGFs Treatment on Amputated Lizard Limbs Stimulate the Regeneration of Long Bones, Opening New Avenues for Limb Regeneration in Amniotes: A Morphological Study Lorenzo Alibardi Comparative Histolab
DEVELOPMENT OF THE HEAD AND NECK PLACODES
 DEVELOPMENT OF THE HEAD AND NECK Placodes and the development of organs of special sense L. Moss-Salentijn PLACODES Localized thickened areas of specialized ectoderm, lateral to the neural crest, at the
DEVELOPMENT OF THE HEAD AND NECK Placodes and the development of organs of special sense L. Moss-Salentijn PLACODES Localized thickened areas of specialized ectoderm, lateral to the neural crest, at the
Lesson 6. References: Chapter 6: Reading for Next Lesson: Chapter 6:
 Lesson 6 Lesson Outline: General Features of the Integument Embryonic Origins of the Epidermis Specializations of the Epidermis o Glands o Keratin and Stratum Corneum Objectives: At the end of this lesson
Lesson 6 Lesson Outline: General Features of the Integument Embryonic Origins of the Epidermis Specializations of the Epidermis o Glands o Keratin and Stratum Corneum Objectives: At the end of this lesson
HISTOPATHOLOGY. Introduction:
 Introduction: HISTOPATHOLOGY Goats and sheep are the major domestic animal species in India. Much of the economy of the country has been depend upon the domestication of these animals. Especially economy
Introduction: HISTOPATHOLOGY Goats and sheep are the major domestic animal species in India. Much of the economy of the country has been depend upon the domestication of these animals. Especially economy
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11046 Supplementary Figure 1: Images of PB-positive cells in the subepidermal region (a-i) Representative images of PB positive cells in the subepidermis of the upper beak of the pigeon.
doi:10.1038/nature11046 Supplementary Figure 1: Images of PB-positive cells in the subepidermal region (a-i) Representative images of PB positive cells in the subepidermis of the upper beak of the pigeon.
Anat. Labor. of Prof. H. SETO, Tohoku University, On the Sensory Terminations Formed along the Ductus
 Anat. Labor. of Prof. H. SETO, Tohoku University, Sendai. On the Sensory Terminations Formed along the Ductus Pancreaticus in Cat. The existence of PACINIan bodies in the pancreas of mammals, especially
Anat. Labor. of Prof. H. SETO, Tohoku University, Sendai. On the Sensory Terminations Formed along the Ductus Pancreaticus in Cat. The existence of PACINIan bodies in the pancreas of mammals, especially
Tail regeneration in the lizards Anguis fragiris and Lacerta dugesii
 J. Linn.Soc. (Zool.) 46,310,~. 297 With 1 Plute and 3 text-figures Printed in Great Britaitt April. 1967 Tail regeneration in the lizards Anguis fragiris and Lacerta dugesii BY SUSAX V. BRYANT AND A. d'a.
J. Linn.Soc. (Zool.) 46,310,~. 297 With 1 Plute and 3 text-figures Printed in Great Britaitt April. 1967 Tail regeneration in the lizards Anguis fragiris and Lacerta dugesii BY SUSAX V. BRYANT AND A. d'a.
Morphology and Ultrastructure of Possible Integumentary Sense Organs in the Estuarine Crocodile (Crocodylus porosus)
") JOURNAL OF MORPHOLOGY 229:315-324 (1996) Morphology and Ultrastructure of Possible Integumentary Sense Organs in the Estuarine Crocodile (Crocodylus porosus) KATE JACKSON, DAVID G. BUTLER, AND JOHN H.
JOURNAL OF MORPHOLOGY 229:315-324 (1996) Morphology and Ultrastructure of Possible Integumentary Sense Organs in the Estuarine Crocodile (Crocodylus porosus) KATE JACKSON, DAVID G. BUTLER, AND JOHN H.
Phylum Platyhelminthes Flatworms
 Phylum Platyhelminthes Flatworms The Acoelomates The acoelomates are animals that lack a coelom. Acoelomates lack a body cavity, and instead the space between the body wall and the digestive tract is filled
Phylum Platyhelminthes Flatworms The Acoelomates The acoelomates are animals that lack a coelom. Acoelomates lack a body cavity, and instead the space between the body wall and the digestive tract is filled
Vol. XIV, No. 1, March, The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S.
 By S.") Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Cornification in developing claws of the common Australian skink (Lampropholis guichenoti) (Squamata, Lacertidae)
 (Squamata, Lacertidae)") Italian Journal of Zoology ISSN: 1125-0003 (Print) 1748-5851 (Online) Journal homepage: http://www.tandfonline.com/loi/tizo20 Cornification in developing claws of the common Australian skink (Lampropholis
Italian Journal of Zoology ISSN: 1125-0003 (Print) 1748-5851 (Online) Journal homepage: http://www.tandfonline.com/loi/tizo20 Cornification in developing claws of the common Australian skink (Lampropholis
The epidermis of reptiles and birds (sauropsids)
") ORIGINAL PAPER Soft epidermis of a scaleless snake lacks beta-keratin M. Toni, L. Alibardi Department of Biology, University of Bologna, Bologna, Italy 07 European Journal of Histochemistry Beta-keratins
ORIGINAL PAPER Soft epidermis of a scaleless snake lacks beta-keratin M. Toni, L. Alibardi Department of Biology, University of Bologna, Bologna, Italy 07 European Journal of Histochemistry Beta-keratins
,,, THE MORPHOLOGY AND MORPHOMETRY OF THE PECTEN OCULI IN DIURNAL AND NOCTURNAL BIRDS: A
 ,,, THE MORPHOLOGY AND MORPHOMETRY OF THE PECTEN OCULI IN DIURNAL AND NOCTURNAL BIRDS: A COMPARATIVE STUDY" BY llijama, S.G., B. V. M. (NBI), Department of Veteri nary Anatomy, University of I\Jairobi.
,,, THE MORPHOLOGY AND MORPHOMETRY OF THE PECTEN OCULI IN DIURNAL AND NOCTURNAL BIRDS: A COMPARATIVE STUDY" BY llijama, S.G., B. V. M. (NBI), Department of Veteri nary Anatomy, University of I\Jairobi.
Prof. Dr. F. BECK, Howard Florey Institute, University of Melbourne, Parkville, 3000 Melbourne, Victoria, Australia
 Reviews and critical articles covering the entire field of normal anatomy (cytology, histology, cyto- and histochemistry, electron microscopy, macroscopy, experimental morphology and embryology and comparative
Reviews and critical articles covering the entire field of normal anatomy (cytology, histology, cyto- and histochemistry, electron microscopy, macroscopy, experimental morphology and embryology and comparative
SCANNING electron - microscopy has
 Characteristics of the Absorptive Surface of the Small Intestine of the Chicken from 1 Day to 14 Weeks of Age 1 R. C. BAYER, C. B. CHAWAN, F. H. BIRD AND S. D. MUSGRAVE Department of Animal and Veterinary
Characteristics of the Absorptive Surface of the Small Intestine of the Chicken from 1 Day to 14 Weeks of Age 1 R. C. BAYER, C. B. CHAWAN, F. H. BIRD AND S. D. MUSGRAVE Department of Animal and Veterinary
DLS Sample Preparation Guide
 DLS Sample Preparation Guide The Leica TCS SP8 DLS is an innovative concept to integrate the Light Sheet Microscopy technology into the confocal microscope. Due to its unique optical architecture samples
DLS Sample Preparation Guide The Leica TCS SP8 DLS is an innovative concept to integrate the Light Sheet Microscopy technology into the confocal microscope. Due to its unique optical architecture samples
A Lymphosarcoma in an Atlantic Salmon (Salmo salar)
") A Lymphosarcoma in an Atlantic Salmon (Salmo salar) Authors: Paul R. Bowser, Marilyn J. Wolfe, and Timothy Wallbridge Source: Journal of Wildlife Diseases, 23(4) : 698-701 Published By: Wildlife Disease
A Lymphosarcoma in an Atlantic Salmon (Salmo salar) Authors: Paul R. Bowser, Marilyn J. Wolfe, and Timothy Wallbridge Source: Journal of Wildlife Diseases, 23(4) : 698-701 Published By: Wildlife Disease
Frog Dissection Information Manuel
 Frog Dissection Information Manuel Anatomical Terms: Used to explain directions and orientation of a organism Directions or Positions: Anterior (cranial)- toward the head Posterior (caudal)- towards the
Frog Dissection Information Manuel Anatomical Terms: Used to explain directions and orientation of a organism Directions or Positions: Anterior (cranial)- toward the head Posterior (caudal)- towards the
Vertebrates. Vertebrate Characteristics. 444 Chapter 14
 4 Vertebrates Key Concept All vertebrates have a backbone, which supports other specialized body structures and functions. What You Will Learn Vertebrates have an endoskeleton that provides support and
4 Vertebrates Key Concept All vertebrates have a backbone, which supports other specialized body structures and functions. What You Will Learn Vertebrates have an endoskeleton that provides support and
Flatworms Flatworms Platyhelminthes dorsoventrally free-living planarian parasitic fluke tapeworm label three body layers ectoderm mesoderm
 Flatworms Flatworms are in the phylum Platyhelminthes. Flatworms are flattened dorsoventrally (top to bottom). The group includes the freshwater, free-living planarian and the parasitic fluke and tapeworm.
Flatworms Flatworms are in the phylum Platyhelminthes. Flatworms are flattened dorsoventrally (top to bottom). The group includes the freshwater, free-living planarian and the parasitic fluke and tapeworm.
A quantitative study of hair growth using mouse and rat vibrissal follicles
 /. Embryol. exp. Morph. Vol. 72, pp. 209-224, 1982 209 Printed in Great Britain Company of Biologists Limited 1982 A quantitative study of hair growth using mouse and rat vibrissal follicles I. Dermal
/. Embryol. exp. Morph. Vol. 72, pp. 209-224, 1982 209 Printed in Great Britain Company of Biologists Limited 1982 A quantitative study of hair growth using mouse and rat vibrissal follicles I. Dermal
RATE OF SCUTE ANNULI DEPOSITION OF EASTERN BOX TURTLES (TERRAPENE CAROLINA CAROLINA) HELD IN CAPTIVITY AND IN THEIR NATURAL HABITAT
 HELD IN CAPTIVITY AND IN THEIR NATURAL HABITAT") Ana Maria Caputo December 4, 2007 RATE OF SCUTE ANNULI DEPOSITION OF EASTERN BOX TURTLES (TERRAPENE CAROLINA CAROLINA) HELD IN CAPTIVITY AND IN THEIR NATURAL HABITAT Eastern box turtles (terappene Carolina
Ana Maria Caputo December 4, 2007 RATE OF SCUTE ANNULI DEPOSITION OF EASTERN BOX TURTLES (TERRAPENE CAROLINA CAROLINA) HELD IN CAPTIVITY AND IN THEIR NATURAL HABITAT Eastern box turtles (terappene Carolina
Reptile Identification Guide
 Care & preservation of Surrey s native amphibians and reptiles Reptile Identification Guide This identification guide is intended to act as an aid for SARG surveyors. Adder, Vipera berus A short, stocky
Care & preservation of Surrey s native amphibians and reptiles Reptile Identification Guide This identification guide is intended to act as an aid for SARG surveyors. Adder, Vipera berus A short, stocky
Short-term Water Potential Fluctuations and Eggs of the Red-eared Slider Turtle (Trachemys scripta elegans)
") Zoology and Genetics Publications Zoology and Genetics 2001 Short-term Water Potential Fluctuations and Eggs of the Red-eared Slider Turtle (Trachemys scripta elegans) John K. Tucker Illinois Natural History
Zoology and Genetics Publications Zoology and Genetics 2001 Short-term Water Potential Fluctuations and Eggs of the Red-eared Slider Turtle (Trachemys scripta elegans) John K. Tucker Illinois Natural History
Gross and histological studies of digestive tract of broilers during postnatal growth and development
 J. Bangladesh Agril. Univ. 10(1): 69 77, 2012 ISSN 1810-3030 Gross and histological studies of digestive tract of broilers during postnatal growth and development M. Nasrin, M. N. H. Siddiqi, M. A. Masum
J. Bangladesh Agril. Univ. 10(1): 69 77, 2012 ISSN 1810-3030 Gross and histological studies of digestive tract of broilers during postnatal growth and development M. Nasrin, M. N. H. Siddiqi, M. A. Masum
General morphology of the oral cavity of the Nile crocodile, Crocodylus niloticus (Laurenti, 1768). I. Palate and gingivae
. I. Palate and gingivae") Onderstepoort Journal of Veterinary Research, 70:281 297 (2003) General morphology of the oral cavity of the Nile crocodile, Crocodylus niloticus (Laurenti, 1768). I. Palate and gingivae J.F. PUTTERILL
Onderstepoort Journal of Veterinary Research, 70:281 297 (2003) General morphology of the oral cavity of the Nile crocodile, Crocodylus niloticus (Laurenti, 1768). I. Palate and gingivae J.F. PUTTERILL
Title. CitationJapanese Journal of Veterinary Research, 24(1-2): 37. Issue Date DOI. Doc URL. Type. File Information
: 37. Issue Date DOI. Doc URL. Type. File Information") Title DISTRIBUTION OF LYMPHATIC TISSUES IN DUCK CAECA Author(s)KITAMURA, Hirokazu; SUGIMURA, Makoto; HASHIMOTO, Yos CitationJapanese Journal of Veterinary Research, 24(1-2): 37 Issue Date 1976-05 DOI 10.14943/jjvr.24.1-2.37
Title DISTRIBUTION OF LYMPHATIC TISSUES IN DUCK CAECA Author(s)KITAMURA, Hirokazu; SUGIMURA, Makoto; HASHIMOTO, Yos CitationJapanese Journal of Veterinary Research, 24(1-2): 37 Issue Date 1976-05 DOI 10.14943/jjvr.24.1-2.37
Avian Reproductive System Female
 extension Avian Reproductive System Female articles.extension.org/pages/65372/avian-reproductive-systemfemale Written by: Dr. Jacquie Jacob, University of Kentucky For anyone interested in raising chickens
extension Avian Reproductive System Female articles.extension.org/pages/65372/avian-reproductive-systemfemale Written by: Dr. Jacquie Jacob, University of Kentucky For anyone interested in raising chickens
THE ROLE OF WATER IN THE EVOLUTION OF THE TERRESTRIAL VERTEBRATES
 26 THE ROLE OF WATER IN THE EVOLUTION OF THE TERRESTRIAL VERTEBRATES BY J. GRAY, M.A., King's College, Cambridge. (From the Zoological Laboratory, Cambridge.) (Received igth January 1928.) (With Three
26 THE ROLE OF WATER IN THE EVOLUTION OF THE TERRESTRIAL VERTEBRATES BY J. GRAY, M.A., King's College, Cambridge. (From the Zoological Laboratory, Cambridge.) (Received igth January 1928.) (With Three
Wound types and healing part three: classification of injuries
 Vet Times The website for the veterinary profession https://www.vettimes.co.uk Wound types and healing part three: classification of injuries Author : Louise O Dwyer Categories : RVNs Date : July 1, 2010
Vet Times The website for the veterinary profession https://www.vettimes.co.uk Wound types and healing part three: classification of injuries Author : Louise O Dwyer Categories : RVNs Date : July 1, 2010
Transition from embryonic to adult epidermis in reptiles occurs by the production of corneous beta-proteins
 Int. J. ev. iol. 58: 829-839 (2014) doi: 10.1387/ijdb.140325la www.intjdevbiol.com Transition from embryonic to adult epidermis in reptiles occurs by the production of corneous beta-proteins LORENZO LIRI*
Int. J. ev. iol. 58: 829-839 (2014) doi: 10.1387/ijdb.140325la www.intjdevbiol.com Transition from embryonic to adult epidermis in reptiles occurs by the production of corneous beta-proteins LORENZO LIRI*
Morphology of cat vomeronasal organ non-sensory epithelium during postnatal development
 Original rticle pissn 2093-3665 eissn 2093-3673 Morphology of cat vomeronasal organ non-sensory epithelium during postnatal development Sanaa. M. Elgayar, Heba M. Saad-Eldin, Ola. Haussein epartment of
Original rticle pissn 2093-3665 eissn 2093-3673 Morphology of cat vomeronasal organ non-sensory epithelium during postnatal development Sanaa. M. Elgayar, Heba M. Saad-Eldin, Ola. Haussein epartment of
Intestinal linear foreign body
 Vet Times The website for the veterinary profession https://www.vettimes.co.uk Intestinal linear foreign body Author : Sally Birch Categories : Companion animal, Vets Date : February 6, 2017 Your first
Vet Times The website for the veterinary profession https://www.vettimes.co.uk Intestinal linear foreign body Author : Sally Birch Categories : Companion animal, Vets Date : February 6, 2017 Your first
A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae)
") Genus Vol. 14 (3): 413-418 Wroc³aw, 15 X 2003 A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae) JAROS AW KANIA Zoological Institute, University of Wroc³aw, Sienkiewicza
Genus Vol. 14 (3): 413-418 Wroc³aw, 15 X 2003 A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae) JAROS AW KANIA Zoological Institute, University of Wroc³aw, Sienkiewicza
NOTE XXXVIII. Three new species of the genus Helota DESCRIBED BY. C. Ritsema+Cz. is very. friend René Oberthür who received. Biet.
 Subshining; HELOTA MARIAE. 249 NOTE XXXVIII. Three new species of the genus Helota DESCRIBED BY C. Ritsema+Cz. The first of these species is very interesting as it belongs to the same section as the recently
Subshining; HELOTA MARIAE. 249 NOTE XXXVIII. Three new species of the genus Helota DESCRIBED BY C. Ritsema+Cz. The first of these species is very interesting as it belongs to the same section as the recently
Histo-Morphological Study on the Footpad of Ostrich (Struthio camelus) In Relation to Locomotion
 In Relation to Locomotion") Histo-Morphological Study on the Footpad of Ostrich (Struthio camelus) In Relation to Locomotion S.A.A. El-Gendy a, Amira Derbalah b, M.E.R. Abu El-Magd c a Department of Anatomy and Embryology, b Histology,
Histo-Morphological Study on the Footpad of Ostrich (Struthio camelus) In Relation to Locomotion S.A.A. El-Gendy a, Amira Derbalah b, M.E.R. Abu El-Magd c a Department of Anatomy and Embryology, b Histology,
Seasonal Variations of yeso sika Deer Skin and its Vegetable Tanned Leather
 Seasonal Variations of yeso sika Deer Skin and its Vegetable Tanned Leather Shigeharu Fukunaga, Akihiko Yoshie, Ikuo Yamakawa, Fumio Nakamura Laboratory of Animal By-product Science, Graduate School of
Seasonal Variations of yeso sika Deer Skin and its Vegetable Tanned Leather Shigeharu Fukunaga, Akihiko Yoshie, Ikuo Yamakawa, Fumio Nakamura Laboratory of Animal By-product Science, Graduate School of
The Armyworm in New Brunswick
 The Armyworm in New Brunswick Mythimna unipuncta (Haworth) Synonym: Pseudaletia unipuncta (Haworth) ISBN 978-1-4605-1679-9 Family: Noctuidae - Owlet moths and underwings Importance The armyworm attacks
The Armyworm in New Brunswick Mythimna unipuncta (Haworth) Synonym: Pseudaletia unipuncta (Haworth) ISBN 978-1-4605-1679-9 Family: Noctuidae - Owlet moths and underwings Importance The armyworm attacks
A NEW SALTICID SPIDER FROM VICTORIA By R. A. Dunn
 Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
Morphology of the femoral glands of the lizard Iguana iguana (linnaeus, 1758) (reptilia, iguanidae)
 (reptilia, iguanidae)") A. Ferreira Femoral glands of lizard 97 ARTIGO ARTICLE Morphology of the femoral glands of the lizard Iguana iguana (linnaeus, 1758) (reptilia, iguanidae) Morfologia das glândulas femorais do lagarto Iguana
A. Ferreira Femoral glands of lizard 97 ARTIGO ARTICLE Morphology of the femoral glands of the lizard Iguana iguana (linnaeus, 1758) (reptilia, iguanidae) Morfologia das glândulas femorais do lagarto Iguana
Name Class Date. After you read this section, you should be able to answer these questions:
 CHAPTER 14 4 Vertebrates SECTION Introduction to Animals BEFORE YOU READ After you read this section, you should be able to answer these questions: How are vertebrates different from invertebrates? How
CHAPTER 14 4 Vertebrates SECTION Introduction to Animals BEFORE YOU READ After you read this section, you should be able to answer these questions: How are vertebrates different from invertebrates? How
Taxonomy. Chapter 20. Evolutionary Development Diagram. I. Evolution 2/24/11. Kingdom - Animalia Phylum - Chordata Class Reptilia.
 Taxonomy Chapter 20 Reptiles Kingdom - Animalia Phylum - Chordata Class Reptilia Order Testudines - turtles Order Crocodylia - crocodiles, alligators Order Sphenodontida - tuataras Order Squamata - snakes
Taxonomy Chapter 20 Reptiles Kingdom - Animalia Phylum - Chordata Class Reptilia Order Testudines - turtles Order Crocodylia - crocodiles, alligators Order Sphenodontida - tuataras Order Squamata - snakes
I another of a genetically different breed of fowl or species of bird that the
 GENOTYPIC CONTROL OF FEATHER COLOR PATTERN AS DEMONSTRATED BY THE EFFECTS OF A SEX- LINKED GENE UPON THE MELANOPHORES* B. H. WILLIER AND MARY E. RAWLES Department of Biology, The Johns Hopkins University,
GENOTYPIC CONTROL OF FEATHER COLOR PATTERN AS DEMONSTRATED BY THE EFFECTS OF A SEX- LINKED GENE UPON THE MELANOPHORES* B. H. WILLIER AND MARY E. RAWLES Department of Biology, The Johns Hopkins University,
Changes in hair growth characteristics following the wounding of vibrissa follicles in the hooded rat
 J. Embryol. exp. Morph. 83, 81-93 (1984) 81 Printed in Great Britain The Company of Biologists Limited 1984 Changes in hair growth characteristics following the wounding of vibrissa follicles in the hooded
J. Embryol. exp. Morph. 83, 81-93 (1984) 81 Printed in Great Britain The Company of Biologists Limited 1984 Changes in hair growth characteristics following the wounding of vibrissa follicles in the hooded
By H. G. JOHNSTON, Ames, Iowa.
 Dec., 19930 Bulletin of the Brooklyn Entomological Society 295 FOUR NEW SPECIES OF MIRIDAE FROM TEXAS (HEMIPTERA).* By H. G. JOHNSTON, Ames, Iowa. Phytocoris conspicuus n. sp. This species is readily distinguished
Dec., 19930 Bulletin of the Brooklyn Entomological Society 295 FOUR NEW SPECIES OF MIRIDAE FROM TEXAS (HEMIPTERA).* By H. G. JOHNSTON, Ames, Iowa. Phytocoris conspicuus n. sp. This species is readily distinguished
8/19/2013. Topic 14: Body support & locomotion. What structures are used for locomotion? What structures are used for locomotion?
 Topic 4: Body support & locomotion What are components of locomotion? What structures are used for locomotion? How does locomotion happen? Forces Lever systems What is the difference between performance
Topic 4: Body support & locomotion What are components of locomotion? What structures are used for locomotion? How does locomotion happen? Forces Lever systems What is the difference between performance
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE
 FROM THE") A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
HISTOLOGY OF MAMMARY GLAND DURING LACTATING AND NON-LACTATING PHASES OF MADRAS RED SHEEP WITH SPECIAL REFERENCE TO INVOLUTION
 International Journal of Science, Environment and Technology, Vol. 5, No 3, 2016, 991 996 ISSN 2278-3687 (O) 2277-663X (P) HISTOLOGY OF MAMMARY GLAND DURING LACTATING AND NON-LACTATING PHASES OF MADRAS
International Journal of Science, Environment and Technology, Vol. 5, No 3, 2016, 991 996 ISSN 2278-3687 (O) 2277-663X (P) HISTOLOGY OF MAMMARY GLAND DURING LACTATING AND NON-LACTATING PHASES OF MADRAS
An introduction to ear cytology in small animal patients
 Vet Times The website for the veterinary profession https://www.vettimes.co.uk An introduction to ear cytology in small animal patients Author : Ariane Neuber Categories : RVNs Date : November 1, 2009
Vet Times The website for the veterinary profession https://www.vettimes.co.uk An introduction to ear cytology in small animal patients Author : Ariane Neuber Categories : RVNs Date : November 1, 2009
THE MICROSCOPE PATHOGEN IDENTIFICATION
 CONTENTS 5 ABOUT THE AUTHOR 5 ACKNOWLEDGEMENTS 6 OVERVIEW 6 What is the Purpose of this Book? 6 What are the Limitations of Light Microscopy as a Diagnostic Tool? 7 When Should I Contact a Veterinarian?
CONTENTS 5 ABOUT THE AUTHOR 5 ACKNOWLEDGEMENTS 6 OVERVIEW 6 What is the Purpose of this Book? 6 What are the Limitations of Light Microscopy as a Diagnostic Tool? 7 When Should I Contact a Veterinarian?
Total Distribution of Taste Buds on the Tongue of the Pup
 The Ohio State University Knowledge Bank kb.osu.edu Ohio Journal of Science (Ohio Academy of Science) Ohio Journal of Science: Volume 4, Issue 6 (November, 194) 194-11 Total Distribution of Taste Buds
The Ohio State University Knowledge Bank kb.osu.edu Ohio Journal of Science (Ohio Academy of Science) Ohio Journal of Science: Volume 4, Issue 6 (November, 194) 194-11 Total Distribution of Taste Buds
Plating the PANAMAs of the Fourth Panama Carmine Narrow-Bar Stamps of the C.Z. Third Series
 Plating the PANAMAs of the Fourth Panama Carmine Narrow-Bar Stamps of the C.Z. Third Series by Geoffrey Brewster The purpose of this work is to facilitate the plating of CZSG Nos. 12.Aa, 12.Ab, 13.A, 14.Aa,
Plating the PANAMAs of the Fourth Panama Carmine Narrow-Bar Stamps of the C.Z. Third Series by Geoffrey Brewster The purpose of this work is to facilitate the plating of CZSG Nos. 12.Aa, 12.Ab, 13.A, 14.Aa,
Animal Coverings Facilitated
 Animal Coverings Facilitated Students will explore various animal coverings with their senses, with help from a high-powered microscope. Description: Explore fur, feathers and scales like never seen before
Animal Coverings Facilitated Students will explore various animal coverings with their senses, with help from a high-powered microscope. Description: Explore fur, feathers and scales like never seen before
Minnesota_mammals_Info_9.doc 11/04/09 -- DRAFT Page 1 of 64. Minnesota mammals
 Minnesota_mammals_Info_9.doc 11/04/09 -- DRAFT Page 1 of 64 Minnesota mammals This is a short guide to Minnesota mammals, with information drawn from Hazard s Mammals of, Walker s Mammals of the World,
Minnesota_mammals_Info_9.doc 11/04/09 -- DRAFT Page 1 of 64 Minnesota mammals This is a short guide to Minnesota mammals, with information drawn from Hazard s Mammals of, Walker s Mammals of the World,
JOURNAL OF. RONALD W. HODGES Systematic Entomology Laboratory, USDA, % U.S. National Museum of Natural History, MRC 168, Washington, D.C.
 JOURNAL OF THE LEPIDOPTERISTS' Volume 39 1985 SOCIETY Number 3 Journal of the Lepidopterists' Society 39(3), 1985, 151-155 A NEW SPECIES OF TlLDENIA FROM ILLINOIS (GELECHIIDAE) RONALD W. HODGES Systematic
JOURNAL OF THE LEPIDOPTERISTS' Volume 39 1985 SOCIETY Number 3 Journal of the Lepidopterists' Society 39(3), 1985, 151-155 A NEW SPECIES OF TlLDENIA FROM ILLINOIS (GELECHIIDAE) RONALD W. HODGES Systematic
EXOTIC CLINICAL PATHOLOGY
 Brittney Exarhos, LVT, RVT Toledo Zoo and Aquarium 2700 Broadway St. Toledo OH 43609 EXOTIC CLINICAL PATHOLOGY Veterinary technicians in a zoo setting often spend a lot of time in the lab. They must have
Brittney Exarhos, LVT, RVT Toledo Zoo and Aquarium 2700 Broadway St. Toledo OH 43609 EXOTIC CLINICAL PATHOLOGY Veterinary technicians in a zoo setting often spend a lot of time in the lab. They must have
The family Gnaphosidae is a large family
 Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
Transformed centrioles In adult and aged cat pinealocytes
 Transformed centrioles In adult and aged cat pinealocytes J. L. Calvo. J. Boya*. J. E. Garcia-Mauriño and D. Rancaño Department of Histology. Faculty of Medicine. University Complutense, 28040 Madrid.
Transformed centrioles In adult and aged cat pinealocytes J. L. Calvo. J. Boya*. J. E. Garcia-Mauriño and D. Rancaño Department of Histology. Faculty of Medicine. University Complutense, 28040 Madrid.
Exotic Hematology Lab Leigh-Ann Horne, LVT, CWR Wildlife Center of Virginia
 Exotic Hematology Lab Leigh-Ann Horne, LVT, CWR Wildlife Center of Virginia lhorne@wildlifecenter.org Anne Lynch, LVT Cedarcrest Animal Clinic amllvt9@gmail.com Introduction While the general set-up for
Exotic Hematology Lab Leigh-Ann Horne, LVT, CWR Wildlife Center of Virginia lhorne@wildlifecenter.org Anne Lynch, LVT Cedarcrest Animal Clinic amllvt9@gmail.com Introduction While the general set-up for
Morphologic study of dog flea species by scanning electron microscopy
 Scientia Parasitologica, 2006, 3-4, 77-81 Morphologic study of dog flea species by scanning electron microscopy NAGY Ágnes 1, L. BARBU TUDORAN 2, V. COZMA 1 1 University of Agricultural Sciences and Veterinary
Scientia Parasitologica, 2006, 3-4, 77-81 Morphologic study of dog flea species by scanning electron microscopy NAGY Ágnes 1, L. BARBU TUDORAN 2, V. COZMA 1 1 University of Agricultural Sciences and Veterinary
Electron Microscopic Observations on Ciliated Epithelium of Tracheal Organ Cultures Infected with Bordetella bronchiseptica
 Microbiol. Immunol. Vol. 33 (2), 111-121, 1989 Electron Microscopic Observations on Ciliated Epithelium of Tracheal Organ Cultures Infected with Bordetella bronchiseptica Kachiko SEKIYA,*,1 Yutaka FUTAESAKU,2
Microbiol. Immunol. Vol. 33 (2), 111-121, 1989 Electron Microscopic Observations on Ciliated Epithelium of Tracheal Organ Cultures Infected with Bordetella bronchiseptica Kachiko SEKIYA,*,1 Yutaka FUTAESAKU,2
Body Parts and Products (Sessions I and II) BROWARD COUNTY ELEMENTARY SCIENCE BENCHMARK PLAN
 BROWARD COUNTY ELEMENTARY SCIENCE BENCHMARK PLAN") activities 22&23 Body Parts and Products (Sessions I and II) BROWARD COUNTY ELEMENTARY SCIENCE BENCHMARK PLAN Grade K Quarter 3 Activities 22 & 23 SC.F.1.1.1 The student knows the basic needs of all living
activities 22&23 Body Parts and Products (Sessions I and II) BROWARD COUNTY ELEMENTARY SCIENCE BENCHMARK PLAN Grade K Quarter 3 Activities 22 & 23 SC.F.1.1.1 The student knows the basic needs of all living
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
PLASMODIUM MODULE 39.1 INTRODUCTION OBJECTIVES 39.2 MALARIAL PARASITE. Notes
 Plasmodium MODULE 39 PLASMODIUM 39.1 INTRODUCTION Malaria is characterized by intermittent fever associated with chills and rigors in the patient. There may be enlargement of the liver and spleen in the
Plasmodium MODULE 39 PLASMODIUM 39.1 INTRODUCTION Malaria is characterized by intermittent fever associated with chills and rigors in the patient. There may be enlargement of the liver and spleen in the
Morphology of Shells From Viable and Nonviable Eggs of the Chinese Alligator (Alligator sinensis)
") ~ JOURNAL OF MORPHOLOGY 222:103-110 (1994) Morphology of Shells From Viable and Nonviable Eggs of the Chinese Alligator (Alligator sinensis) CAROLE S. WINK AND RUTH M. ELSEY Department of Anatomy, Louisiana
~ JOURNAL OF MORPHOLOGY 222:103-110 (1994) Morphology of Shells From Viable and Nonviable Eggs of the Chinese Alligator (Alligator sinensis) CAROLE S. WINK AND RUTH M. ELSEY Department of Anatomy, Louisiana
Biology. Slide 1of 50. End Show. Copyright Pearson Prentice Hall
 Biology 1of 50 2of 50 Phylogeny of Chordates Nonvertebrate chordates Jawless fishes Sharks & their relatives Bony fishes Reptiles Amphibians Birds Mammals Invertebrate ancestor 3of 50 A vertebrate dry,
Biology 1of 50 2of 50 Phylogeny of Chordates Nonvertebrate chordates Jawless fishes Sharks & their relatives Bony fishes Reptiles Amphibians Birds Mammals Invertebrate ancestor 3of 50 A vertebrate dry,
Growth and Development. Sex determination Development: embryogenesis and morphogenesis Metamorphosis
 Herp Development Growth and Development Sex determination Development: embryogenesis and morphogenesis Metamorphosis Growth and Development Sex determination Development: embryogenesis and morphogenesis
Herp Development Growth and Development Sex determination Development: embryogenesis and morphogenesis Metamorphosis Growth and Development Sex determination Development: embryogenesis and morphogenesis
Mesosomes are a definite event in antibiotic-treated Staphylococcus aureus ATCC 25923
 Tropical Biomedicine 24(1): 105 109 (2007) Mesosomes are a definite event in antibiotic-treated Staphylococcus aureus ATCC 25923 Santhana Raj, L. 1*, Hing, H.L. 2, Baharudin Omar 2, Teh Hamidah, Z. 1,
Tropical Biomedicine 24(1): 105 109 (2007) Mesosomes are a definite event in antibiotic-treated Staphylococcus aureus ATCC 25923 Santhana Raj, L. 1*, Hing, H.L. 2, Baharudin Omar 2, Teh Hamidah, Z. 1,
Module Egg. MODULE NO. 25: Internal Quality of Egg
 Module Egg MODULE NO. 25: Internal Quality of Egg Quality Quality : Degree of excellence Those conditions and characteristics that consumers want, and are willing to pay for, are, in a broad sense, factors
Module Egg MODULE NO. 25: Internal Quality of Egg Quality Quality : Degree of excellence Those conditions and characteristics that consumers want, and are willing to pay for, are, in a broad sense, factors
Protocol for fabrication of microcompartments for long-term culture and imaging of small C. elegans larvae. Henrik Bringmann, March 2011.
 Protocol for fabrication of microcompartments for long-term culture and imaging of small C. elegans larvae Henrik Bringmann, March 2011. 1 Step-by-Step Protocol Step1 : Preparing a humidity dish (see illustration
Protocol for fabrication of microcompartments for long-term culture and imaging of small C. elegans larvae Henrik Bringmann, March 2011. 1 Step-by-Step Protocol Step1 : Preparing a humidity dish (see illustration
Title. Author(s)Takahashi, Ryoichi. CitationInsecta matsumurana, 14(1): 1-5. Issue Date Doc URL. Type. File Information
Takahashi, Ryoichi. CitationInsecta matsumurana, 14(1): 1-5. Issue Date Doc URL. Type. File Information") Title Some Aleyrodidae from Mauritius (Homoptera) Author(s)Takahashi, Ryoichi CitationInsecta matsumurana, 14(1): 1-5 Issue Date 1939-12 Doc URL http://hdl.handle.net/2115/9426 Type bulletin File Information
Title Some Aleyrodidae from Mauritius (Homoptera) Author(s)Takahashi, Ryoichi CitationInsecta matsumurana, 14(1): 1-5 Issue Date 1939-12 Doc URL http://hdl.handle.net/2115/9426 Type bulletin File Information
Gross and Microscopic Features of the Interdigital Sinus in the Barbados Black Belly Sheep in Trinidad
 Original Research Article International Journal of Current Research in Medical Sciences ISSN: 2454-5716 www.ijcrims.com Volume 2, Issue 7-2016 SOI: http://s-o-i.org/1.15/ijcrms-2016-2-7-4 Gross and Microscopic
Original Research Article International Journal of Current Research in Medical Sciences ISSN: 2454-5716 www.ijcrims.com Volume 2, Issue 7-2016 SOI: http://s-o-i.org/1.15/ijcrms-2016-2-7-4 Gross and Microscopic
Class Reptilia Testudines Squamata Crocodilia Sphenodontia
 Class Reptilia Testudines (around 300 species Tortoises and Turtles) Squamata (around 7,900 species Snakes, Lizards and amphisbaenids) Crocodilia (around 23 species Alligators, Crocodiles, Caimans and
Class Reptilia Testudines (around 300 species Tortoises and Turtles) Squamata (around 7,900 species Snakes, Lizards and amphisbaenids) Crocodilia (around 23 species Alligators, Crocodiles, Caimans and
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS
 AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Funkhouser, W. D., 1927. New Australian Membracidae (Homoptera). Records of the Australian Museum 15(5): 305 312, plate xxvi. [6 April 1927]. doi:10.3853/j.0067-1975.15.1927.817
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Funkhouser, W. D., 1927. New Australian Membracidae (Homoptera). Records of the Australian Museum 15(5): 305 312, plate xxvi. [6 April 1927]. doi:10.3853/j.0067-1975.15.1927.817
Dolichopeza reidi nov.sp., a new crane fly species from Lord Howe Island, New South Wales, Australia (Diptera: Tipulidae)
") Linzer biol. Beitr. 49/1 727-731 28.7.2017 Dolichopeza reidi nov.sp., a new crane fly species from Lord Howe Island, New South Wales, Australia (Diptera: Tipulidae) Günther THEISCHINGER Abstract: Dolichopeza
Linzer biol. Beitr. 49/1 727-731 28.7.2017 Dolichopeza reidi nov.sp., a new crane fly species from Lord Howe Island, New South Wales, Australia (Diptera: Tipulidae) Günther THEISCHINGER Abstract: Dolichopeza
DRAFT TANZANIA STANDARD
 Hatching eggs Specification DRAFT TANZANIA STANDARD TANZANIA BUREAU OF STANDARDS 1 Hatching eggs Specification TBS/AFDC 22 (5271) P3 0 FOREWORD This Tanzania standard was developed due to rapid increase
Hatching eggs Specification DRAFT TANZANIA STANDARD TANZANIA BUREAU OF STANDARDS 1 Hatching eggs Specification TBS/AFDC 22 (5271) P3 0 FOREWORD This Tanzania standard was developed due to rapid increase
SEMESTER ONE 2007 INFECTION and IMMUNITY GRADUATE ENTRY PROGRAMME PARASITOLOGY PRACTICAL 9 Dr TW Jones NEMATODES
 SEMESTER ONE 2007 INFECTION and IMMUNITY GRADUATE ENTRY PROGRAMME PARASITOLOGY PRACTICAL 9 Dr TW Jones NEMATODES Objectives After this class I expect you to be able to: 1. Describe and recognise the range
SEMESTER ONE 2007 INFECTION and IMMUNITY GRADUATE ENTRY PROGRAMME PARASITOLOGY PRACTICAL 9 Dr TW Jones NEMATODES Objectives After this class I expect you to be able to: 1. Describe and recognise the range
EGG STAGE. 1. How many eggs does a female Monarch usually lay on one milkweed plant? Given a choice, what age plant, or leaves, does she prefer?
 EGG STAGE 1. How many eggs does a female Monarch usually lay on one milkweed plant? Given a choice, what age plant, or leaves, does she prefer? 2. The egg stage lasts 1-3 days. Look at the egg that you
EGG STAGE 1. How many eggs does a female Monarch usually lay on one milkweed plant? Given a choice, what age plant, or leaves, does she prefer? 2. The egg stage lasts 1-3 days. Look at the egg that you
Animal Form and Function. Amphibians. United by several distinguishing apomorphies within the Vertebrata
 Animal Form and Function Kight Amphibians Class Amphibia (amphibia = living a double life) United by several distinguishing apomorphies within the Vertebrata 1. Skin Thought Question: For whom are integumentary
Animal Form and Function Kight Amphibians Class Amphibia (amphibia = living a double life) United by several distinguishing apomorphies within the Vertebrata 1. Skin Thought Question: For whom are integumentary
KS1 Baby Animals. Marwell Wildlife Colden Common Winchester Hampshire SO21 1JH
 Marwell Wildlife Colden Common Winchester Hampshire SO21 1JH Marwell is a limited liability company registered in England and Wales under no. 1355272. The company is a registered charity, no. 275433. VAT
Marwell Wildlife Colden Common Winchester Hampshire SO21 1JH Marwell is a limited liability company registered in England and Wales under no. 1355272. The company is a registered charity, no. 275433. VAT
Liver and Gallbladder Morphology of the juvenile Nile crocodile, Crocodylus niloticus (Laurenti, 1768)
") Liver and Gallbladder Morphology of the juvenile Nile crocodile, Crocodylus niloticus (Laurenti, 1768) by ERNA VAN WILPE Submitted in partial fulfilment of the requirements for the degree MSc DEPARTMENT
Liver and Gallbladder Morphology of the juvenile Nile crocodile, Crocodylus niloticus (Laurenti, 1768) by ERNA VAN WILPE Submitted in partial fulfilment of the requirements for the degree MSc DEPARTMENT
Key words: Plasmodium, Kentropyx calcarata, Brazil, merogony, gametocytes, ultrastructure
 FOLIA PARASITOLOGICA 49: 2-8, 2002 Fine structure of erythrocytic stages of a Plasmodium tropiduri-like malaria parasite found in the lizard Kentropyx calcarata (Teiidae) from north Brazil Ilan Paperna
FOLIA PARASITOLOGICA 49: 2-8, 2002 Fine structure of erythrocytic stages of a Plasmodium tropiduri-like malaria parasite found in the lizard Kentropyx calcarata (Teiidae) from north Brazil Ilan Paperna
Breed Characteristics Overall Impression The German Long faced
 Text: N. Akkerman, The Netherlands Photos: Uwe Held, Germany In General The German Long Faced Tumbler is closely related to the English Magpie. Not because of recent crossings, no, they both spring from
Text: N. Akkerman, The Netherlands Photos: Uwe Held, Germany In General The German Long Faced Tumbler is closely related to the English Magpie. Not because of recent crossings, no, they both spring from
Dogs. WORD BANK: blind, cattle, companions, countries, guard, hunt, sleds, warn. Level 2.0, Story 1. Copyright 2012 Read Naturally, Inc.
 Dogs Level 2.0, Story 1 1 2 3 4 5 7 8 3 people or friends you spend a lot of time with 5 find and kill animals for food 7 watch something closely in order to keep it safe 8 unable to see 1 areas of land
Dogs Level 2.0, Story 1 1 2 3 4 5 7 8 3 people or friends you spend a lot of time with 5 find and kill animals for food 7 watch something closely in order to keep it safe 8 unable to see 1 areas of land
Lacerta viridis. Functional anatomy of the lungs of the green lizard, (Accepted 18 February 1977)
") J. Anat. (1978), 125, 2, pp. 421-431 421 With 9 figures Printed in Great Britain Functional anatomy of the lungs of the green lizard, Lacerta viridis C. MEBAN Department of Anatomy, The Queen's University
J. Anat. (1978), 125, 2, pp. 421-431 421 With 9 figures Printed in Great Britain Functional anatomy of the lungs of the green lizard, Lacerta viridis C. MEBAN Department of Anatomy, The Queen's University
TWO NEW PINE-FEEDING SPECIES OF COLEOTECHNITES ( GELECHIIDAE )
") Journal of the Lepidopterists' Society 32(2), 1978, 118-122 TWO NEW PINE-FEEDING SPECIES OF COLEOTECHNITES ( GELECHIIDAE ) RONALD W. HODGES l AND ROBERT E. STEVENS2 ABSTRACT. Two new species of moths,
Journal of the Lepidopterists' Society 32(2), 1978, 118-122 TWO NEW PINE-FEEDING SPECIES OF COLEOTECHNITES ( GELECHIIDAE ) RONALD W. HODGES l AND ROBERT E. STEVENS2 ABSTRACT. Two new species of moths,
1. Examine the specimens of sponges on the lab table. Which of these are true sponges? Explain your answers.
 Station #1 - Porifera 1. Examine the specimens of sponges on the lab table. Which of these are true sponges? Explain your answers. 2. Sponges are said to have an internal special skeleton. Examine the
Station #1 - Porifera 1. Examine the specimens of sponges on the lab table. Which of these are true sponges? Explain your answers. 2. Sponges are said to have an internal special skeleton. Examine the
1. If possible, place the class based on loss of pigment (bleaching) from the skin.
 from the skin.") 4-H Poultry Judging Past egg production (reasons class) Interior egg quality candling Interior egg quality - broken out Exterior egg quality Poultry carcass parts identification Poultry carcass quality
4-H Poultry Judging Past egg production (reasons class) Interior egg quality candling Interior egg quality - broken out Exterior egg quality Poultry carcass parts identification Poultry carcass quality
Diversity of Animals
 Classifying Animals Diversity of Animals Animals can be classified and grouped based on similarities in their characteristics. Animals make up one of the major biological groups of classification. All
Classifying Animals Diversity of Animals Animals can be classified and grouped based on similarities in their characteristics. Animals make up one of the major biological groups of classification. All
When? Why? and How?: Some Speculations on the Evolution of the Vertebrate Integument
 AM. ZOOLOCIST, 12:159-171 (1972). When? Why? and How?: Some Speculations on the Evolution of the Vertebrate Integument PAUL F. A. MADERSON Biology Department, Brooklyn College, Brooklyn, New York 11210
AM. ZOOLOCIST, 12:159-171 (1972). When? Why? and How?: Some Speculations on the Evolution of the Vertebrate Integument PAUL F. A. MADERSON Biology Department, Brooklyn College, Brooklyn, New York 11210
Medical Department PHYSIOLOGICAL EAR CLEANSER
 PHYSIOLOGICAL EAR CLEANSER Their ears are fragile, take care! Structure of the external ear Pinna Ear canal External ear Border Collie Jack Russel Inner ear? Tympanic membrane Middle ear Bearded Collie
PHYSIOLOGICAL EAR CLEANSER Their ears are fragile, take care! Structure of the external ear Pinna Ear canal External ear Border Collie Jack Russel Inner ear? Tympanic membrane Middle ear Bearded Collie
Effects of Natural Selection
 Effects of Natural Selection Lesson Plan for Secondary Science Teachers Created by Christine Taylor And Mark Urban University of Connecticut Department of Ecology and Evolutionary Biology Funded by the
Effects of Natural Selection Lesson Plan for Secondary Science Teachers Created by Christine Taylor And Mark Urban University of Connecticut Department of Ecology and Evolutionary Biology Funded by the
Fishes, Amphibians, Reptiles
 Fishes, Amphibians, Reptiles Section 1: What is a Vertebrate? Characteristics of CHORDATES Most are Vertebrates (have a spinal cord) Some point in life cycle all chordates have: Notochord Nerve cord that
Fishes, Amphibians, Reptiles Section 1: What is a Vertebrate? Characteristics of CHORDATES Most are Vertebrates (have a spinal cord) Some point in life cycle all chordates have: Notochord Nerve cord that