ASSESSING THE COMPOSITION OF GREEN TURTLE (Chelonia mydas) FORAGING GROUNDS IN AUSTRALASIA USING MIXED STOCK ANALYSES
|
|
|
- Sydney Floyd
- 5 years ago
- Views:
Transcription
1 ASSESSING THE COMPOSITION OF GREEN TURTLE (Chelonia mydas) FORAGING GROUNDS IN AUSTRALASIA USING MIXED STOCK ANALYSES By MICHAEL PAUL JENSEN B.Sc. (University of Aarhus) (2001) M.Sc. (University of Aarhus) (2005) Institute for Applied Ecology Faculty of Applied Science University of Canberra Australia A thesis submitted in fulfilment of the requirements of the Degree of Doctor of Philosophy at the University of Canberra. March 2010 i
2 Form B Certificate of Authorship of Thesis Except where clearly acknowledged in footnotes, quotations and the bibliography, I certify that I am the sole author of the thesis submitted today entitled - Assessing the composition of green turtle (Chelonia mydas) foraging grounds in Australasia using mixed stock analyses. I further certify that to the best of my knowledge the thesis contains no material previously published or written by another person except where due reference is made in the text of the thesis. The material in the thesis has not been the basis of an award o any other degree or diploma except where due reference is made in the text of the thesis. The thesis complies with University requirements for a thesis as set out in Signature of candidate... Signature of chair of the supervisory panel Copyright.. Date This thesis ( by Michael P. Jensen, 2010) may be freely copied or distributed for private and/or commercial use and study. However, no part of this thesis or the information herein may be included in a publication or referred to in a publication without the written consent of Michael P. Jensen. Any reference to this work must be fully acknowledged. ii
3 ACKNOWLEDGEMENTS This thesis results from three and a half years of research carried out since I came to Australia in June It has been a fantastic ride, with fieldwork at some of the most beautiful places on earth. I have worked with a great number of people, whose contribution in assorted ways to both the research and the preparation of this thesis deserve special mention. It is a pleasure to convey my gratitude to you all in my humble acknowledgment. In the first place, I would like to express my sincere gratitude to my primary supervisor, Nancy FitzSimmons, for her advice and guidance from the onset of this research, as well as giving me extraordinary experiences throughout the whole process. Above all, and most importantly, she provided me unflinching encouragement, trust and support during my time in Australia. Her true passion for science and marine turtles in particular, has made her as a constant oasis of ideas, which exceptionally inspired and enriched my growth as a student, a researcher and as a person. She became my mentor and my friend, and I am indebted to her more than she can know. I gratefully acknowledge also my secondary supervisor, Col Limpus, for his advice, ideas and support. He contains a wealth of knowledge and wisdom when it comes to marine turtles and he has a remarkable way of asking questions that has made me rethink my research and ideas, always to the benefit of the project. During this work I have collaborated with many colleagues for whom I have great regard. I wish to extend my warmest thanks to all those who have helped me with my work, from sending me samples, helping with fieldwork, providing comments and advice, to being a great inspiration. Another special thanks goes to my friend and colleague Ian Bell. In June 2007, he somewhat sceptically, agreed to let a Danish geneticist come along on one of his fieldtrips to the remote northern Great Barrier Reef. Since then, he has contributed so much and given me unconditional support during several fieldtrips, which made him a vital part of this research and so of this thesis. I hope he doesn t regret his decision. His originality and confronting questions have triggered and nourished an intellectual maturity that I will benefit from for a long time to i
4 come. Ian, I am grateful in every possible way and hope to keep up our collaboration in the future. It is a pleasure to pay tribute also to all those who helped provide me with samples. To Mick Guinea and Scott Whiting who have taken time during their own fieldwork to collect genetic samples and to Nick Pilcher who included me in his interesting research of Malaysian green turtles. Many thanks also to Mark Hamann, Frank Loban and Steven Ambar and the rest of the mob up in Torres Strait. I am much indebted to all of them for their valuable contributions, from the planning of the project to the many days of fieldwork. Thanks to Kelly Pendoley for the most recent samples from Western Australia and to Bob Prince for the original WA collection. I would also acknowledge my fellow students and colleagues at the Institute for Applied Ecology; Anett Richter, Kate Hodges, Stewart Pittard, John Roe, Wendy Dimond, David Wong, Carla Eisemberg, Alex Quinn, Anna MsDonald, Tariq Ezaz and Marion Hoehn for their moral support and their friendship. A special thank you goes to Niccy Aitken, first and foremost for her friendship, but I have also benefited from her advice and guidance. She always kindly granted me her time, even to answer some of my naive questions about lab work. She also provided useful comments to the manuscript. I convey special acknowledgement to the office staff Kerrie, Sam and Tara for all their help dealing with travel funds, administration and bureaucratic matters during my stay and in particular with my commute between Canberra, Denmark, the Great Barrier Reef and the rest of the world so I could more easily carry out my research and travels. I am very grateful to Alberto Abreu-Grobois, my masters supervisor, who remains my mentor and provides great inspiration for the way I go about doing research. He always takes time to provide his advice and his bright thoughts are always very fruitful for shaping my ideas and research. I humbly thank my thesis reviewers that in the midst of all their activity, they accepted to be members of the review committee. A big thank you to the turtle gang, Suzanne Livingstone, Jason van de Merwe, Mark Trodoir, Colette Wabnitz, and Dave Wayers for their friendship and for their advice and support. To Sam ii
5 Emerick, thank you for your support and for all your fruitful comments on the manuscript. I would like to thank the sea turtle community for making me part of a very inspirational group of people. Collective and individual acknowledgments are also owed to the many volunteers who took time to help with fieldwork. Thanks to Brian McNeill, Lachlan Duff, Sam Dibella, Tony Mitchell (Barney), Sarah Vargas, Anelise Hahn, Stewart Pittard, Klaus Karlsen, Maren Lyngsgaard and many others too numerous to name. They made fieldwork fun and many new friendships were formed during the many weeks of turtle work. Where would I be without my family? My parents deserve special mention for their steady support. My Father, Henning, is the person who gave me the passion for nature from when I was a child. My Mother, Ulla, is the one who sincerely raised me with her loving support in everything I do. Thanks to my sister Susanne, her husband Cristòbal, and my niece and nephew Isabel and Cris, for being supportive and caring. Without their encouragement and understanding it would have been impossible for me to finish this work. Finally, I would like to thank everybody who contributed to the successful realisation of this thesis, as well as expressing my apology that I could not mention personally each of you one by one. iii
6 TABLE OF CONTENTS ACKNOWLEDGEMENTS... i TABLE OF CONTENTS... iv LIST OF FIGURES... vi LIST OF TABLES... vii ABSTRACT Introduction to the Thesis Sea turtles of the world Lifecycle of Sea Turtles Green turtles (Chelonia mydas) Green turtle population genetics Review: Mixed Stock Analysis and marine turtles Introduction Mixed Stock Analysis MSA and Marine turtles Loggerhead turtles (Caretta caretta) Hawksbill turtles (Eretmochelys imbricata) Green turtles (Chelonia mydas) Complex life history Rookery size and distance Ocean Currents Temporal differences Gender differences Size class differences Limitations with MSA Sample size Source populations Resolution of genetic markers Conservation and Management Influences of larger sample sizes and longer sequences for defining Management Units and estimating stock composition of green turtle foraging grounds Introduction Materials and Methods Sample collection Characterization of mtdna haplotypes Molecular analysis Mixed Stock Analysis Results Rookery diversity Population differentiation Foraging aggregations MSA Discussion Rookery diversity and differentiation Foraging aggregations and MSA Management implications Origin of immature green turtles (Chelonia mydas) at two foraging grounds in Sabah, Malaysia iv
7 4.1 Introduction Materials and Methods Study site and sample collection Characterisation of mtdna haplotypes Mixed Stock Analysis Results Mixed Stock Analysis Discussion Stock composition of green turtle (Chelonia mydas) foraging grounds of the Great Barrier Reef: implications of variation across latitude and size classes Introduction Materials and Methods Study site and sample collection Sample collection Characterisation of mtdna haplotypes Mixed Stock Analysis Migration data from mark-recapture studies Results Haplotype diversity along the GBR foraging grounds Mixed stock analysis and mark-recapture Discussion MSA and mark-recapture tagging data Latitudinal comparisons Age class variation Conservation implications Synopsis Conservation management and implications Future research Modelling REFERENCES APPENDIX APPENDIX APPENDIX v
8 LIST OF FIGURES Figure 1.1. Generalised lifecycle for most species of marine turtles. Species and populations vary mainly in the duration of the different phases. The figure is divided into hatchling/juvenile (light grey line), female (dark grey line) and, male (black line) migration. 6 Figure 2.1. Model of random mixing of pelagic turtles and subsequent random recruitment into benthic foraging grounds that are genetically similar. 22 Figure 2.2. Model of random mixing of pelagic turtles and subsequent selective recruitment into benthic foraging grounds near natal rookeries, leading to genetic structure among foraging grounds. 23 Figure 2.3. Model of non-random mixing of pelagic turtles due to oceanic currents and subsequent selective recruitment into benthic foraging grounds near natal rookeries, leading to genetic structure among foraging grounds and among pelagic turtles in different regions. 24 Figure 2.4. Simulation of a hypothesised foraging aggregation where a) all rookeries are completely differentiated and b) rookeries share common haplotypes. 31 Figure 3.1. Schematic of the mtdna control region in marine turtles and the location of the short sequence used in Dethmers et al. (2006) (primers TCR5 and TCR6) and the long sequence used in this study (primers LTEi9 and H950). 39 Figure 3.2. Sampling locations from nesting sites in eastern Indian Ocean and Southeast Asia with the UPGMA tree of the genetic relationship between rookeries. 41 Figure 3.3. Haplotype network based on maximum parsimony for the short a) and the long b) sequence. 45 Figure 3.4. Rarefraction curves for each Mus. X-axis shows the sample size and Y axis the number of haplotypes. The graph shows the mean and the SD. 50 Figure 4.1. Map showing the 17 genetic stocks used as reference for tracing back the origin of turtles (adapted from Dethmers et al. 2006) and the location of the two study areas, Mantanani Island and Layang Layang Islands. 62 Figure 5.1. Map showing the location of 17 genetically differentiated breeding stocks initially included in the MSA analysis and the six foraging grounds analysed for stock composition 76 Figure 5.2. Map showing the location of the six foraging grounds and the mean relative contribution of ngbr, sgbr/coral Sea and other stocks for each group sampled within the foraging grounds. 90 vi
9 LIST OF TABLES Table 2.1. Studies using Mixed Stock Analysis of marine turtle foraging aggregations; categorised by species and region. 16 Table 3.1. Distribution of C. mydas haplotypes sampled at 15 rookeries in the eastern Indian Ocean and Southeast Asia. 46 Table 3.2. Diversity estimates for 17 green turtle MUs and rookeries, including sample size (n), number of haplotypes (H) haplotype diversity (h) and nucleotide diversity (π). 47 Table 3.3. Analysis of molecular variance (AMOVA) results for 11 green turtle east Indian Ocean Management Units and the Coburg Island rookery 47 Table 3.4. Genetic differentiation (F ST ) among MUs and rookeries based on haplotype frequencies using the long sequence (below diagonal) and estimates of the number of migrants per generation (Nm; above diagonal). 48 Table 3.5. Estimated P values from exact test of population differentiation among east Indian Ocean MUs and the Coburg Peninsula rookeries based on haplotype frequencies using the short sequence (above diagonal) and the long sequence (below diagonal). 49 Table 3.6. Distribution of C. mydas haplotypes sampled at two foraging grounds, Shark Bay and Cocos (Keeling) Islands. 51 Table 3.7. Contribution of regional stocks to two foraging aggregations at Shark Bay and Cocos (Keeling) using the long (L) and short (S) sequence. 53 Table 4.1. Haplotype frequencies of nine nesting populations that either had a mean contribution of ( 1%) in mixed stock analysis to the two foraging grounds (FG) or shared several haplotypes with the FGs. 66 Table 4.2. Estimates of the rookery origin of immature green turtles foraging at Mantanani Island and Layang Layang Island, based on short sequence. 68 Table 4.3. Estimates of the rookery origin of immature green turtles foraging at Mantanani Island and Layang Layang Island based on long sequence. 69 Table 5.1. Haplotype composition of the eight stocks used for the MSA and the six foraging grounds analysed; TS = Torres Strait, CR = Clack Reef, HG = Howicks Group, EB = Edgecombe Bay, SB = Shoalwater Bay, and MB = Moreton Bay, separated by sampling year and size class (A = adults, SA = sub-adults and J = juveniles). 82 Table 5.2. Estimates of haplotype (h) and nucleotide diversity (π) within eight green turtle genetic stocks used for the MSA and six foraging grounds for groupings of adult and immature turtles. 85 vii
10 Table 5.3. Analysis of molecular variance (AMOVA) results for the southern and northern groups of green turtle foraging and nesting sites. 86 Table 5.4. Results from the Bayesian MSA for six green turtle foraging grounds (FG); TS, Torres Strait; CR, Clack Reef; HG, Howicks Group; EB, Edgecombe Bay; SB, Shoalwater Bay and MB, Moreton Bay. 88 viii
11 ABSTRACT Understanding the population dynamics in both breeding and foraging habitats is a vital part of assessing the long-term viability of any species, especially those that are highly migratory. This is particularly true for green turtles, Chelonia mydas, which are long-lived marine turtles that undergo migrations for several years as post hatchlings until they select foraging grounds, and as adults, migrate between their foraging grounds and nesting beaches. Monitoring of populations at the foraging grounds may help detect early signs of population trends that would otherwise take decades to be observed at the nesting beach. In order to gain such insights the connectivity between nesting and foraging habitats must be established. Genetic analysis of rookeries to define discrete populations (stocks), in combination with Mixed Stock Analysis (MSA) based on data from molecular markers, provides an effective approach for estimating the origin of turtles sampled away from their nesting beach. In this thesis, new investigations into the genetic structure of green turtle populations in Australasia were conducted using longer (~780 bp) mitochondrial (mt) DNA sequences, larger sample sizes and new locations. This information provided the baseline data used in Mixed Stock Analyses of the composition of foraging grounds in three regions of Australasia including Western Australia, the Great Barrier Reef (GBR) and Malaysia. In chapter 2, I review what has been learned since the first MSA studies in marine turtles more than a decade ago. Since the early 1990 s, numerous studies used this method to elucidate the rookery origins of young pelagic stage turtles and of older turtles in benthic foraging grounds, in fisheries by-catch and in strandings. These studies have all shown how Mixed Stock Analysis has provided valuable new insights into the distribution of marine turtles, although in most cases the estimates are affected by large uncertainty. Several issues in the effective use of MSA need to be addressed concerning study design, sample sizes and the resolution provided by the genetic marker. Nonetheless, Mixed Stock Analysis holds great potential for monitoring population trends at oceanic and coastal foraging grounds for all size classes. Comparisons of adults and juveniles provide an opportunity to pick up early signs of shifts in the contributions of populations that may indicate population decline (or increase) (e.g., Chapter 5). 1
12 Recent increases in industrial development of coastal island and offshore habitats in Western Australia (WA) have highlighted the need to better understand the dynamics of marine turtle populations in these areas. An analysis of previously sampled populations (Management Units; MUs) and four new rookeries identified two possible new Management Units in this region at Cobourg Peninsula and Cocos (Keeling) Island and grouped Browse Island with the existing MU at Scott Reef and Barrow Island to the large North West Shelf MU. These analyses used a 780 bp sequence of the mtdna control region that encompassed the 386 bp sequence used in a previous study. The longer sequence, larger sample sizes and new locations revealed more than doubled the number of haplotypes (n = 39) than previously observed. However, this made little difference to the population genetic structure as common haplotypes were still shared among population. MSA showed that the majority (>90%) of turtles foraging at Shark Bay were from neighbouring North West Shelf rookeries, while the Cocos (Keeling) foraging ground was composed of turtles mainly from Cocos (~70%), but with some contributions from North West Shelf and possibly Scott Reef MUs. In an investigation of foraging populations in Malaysia, mtdna sequence data were analysed from 81 immature green turtles at two foraging grounds at Mantanani Island and Layang Layang Island located northwest of Sabah, Malaysia. Previously published data from 17 Australasian green turtle populations were used as the baseline data for tracing back the origin of turtles at the two foraging grounds. The majority of these turtles originated from major rookeries in the Malaysia and Philippine Turtle Islands (~30%), and Sarawak (~60%) in north-western Borneo. These same rookeries have a long tradition of using unshaded beach hatcheries that has resulted in the production of mostly female hatchlings. This may have contributed to the 1:4 female biases seen at the two foraging grounds. The implications of hatchery practises at nesting beaches are discussed and the importance of continued monitoring and research at these foraging areas is highly recommended to improve the management of marine turtles in the region. Detailed MSA of green turtle aggregations at six major foraging grounds along the east coast of Australian were combined with data from more than 30 years of mark recapture efforts along the Great Barrier Reef. Overall, the MSA in combination with the mark-recapture data supports a model in which the foraging aggregations are composed of individuals from the two Great Barrier Reef stocks (ngbr, sgbr) with small contributions from other stocks. The north/south 2
13 transect of foraging grounds analysed spanned ~2300 km. Along this transect the main contributor shifted from being predominantly the ngbr stock at foraging grounds in Torres Strait, Clack Reef and the Howicks Group to predominantly the sgbr stock at Edgecombe Bay, Shoalwater Bay and Moreton Bay. At the most northern foraging ground in the Torres Strait, significant shifts in haplotype frequencies between juveniles and adults resulted in major shifts in the estimated stock contributions for these groups. Fewer juveniles originated from the ngbr stock and higher proportion originated from the sgbr and other stocks in comparison to adults. This trend was apparent in the four most northern foraging grounds, even in Edgecombe Bay, which had a predominance of turtles from the sgbr stock. Point estimates of contributions from the ngbr stock dropped from 0.89 in adults to 0.53 in juveniles in Torres, Strait, from 0.69 to 0.49 at Clack Reef, from 0.66 to 0.49 in the Howicks Group and from 0.10 in adults to 0.01 in juveniles at Edgecombe Bay. In contrast, at the Shoalwater Bay foraging ground the opposite was observed, with a drop in contribution from the sgbr stock from 0.98 in adults to 0.84 and 0.85 in juveniles and sub-adults, respectively, and an increase in contributions from other stocks in juveniles and sub-adults. The observed patterns at the various foraging grounds likely resulted from several causes and four possible explanations are explored, the mostly likely of which were that (i) juveniles have shifted foraging grounds as they mature, or that (ii) reduced hatching success from the main ngbr rookery at Raine Island for more than a decade has resulted in reduced recruitment into the ngbr foraging ground. The later possibility suggests a need to take action to conserve the ngbr population The combined strength of data derived from mark-recapture studies, demographic studies to determine sex, maturity and breeding status of the turtles, genetic studies to determine stock composition and satellite telemetry, are needed to provide informed assessments of foraging populations necessary for guiding sustainable management of marine turtles. 3
14 CHAPTER 1 1. Introduction to the Thesis In this thesis, I investigate the use of mitochondrial DNA sequence data to estimate the origin of marine turtles found at foraging grounds. I apply the technique to green turtles (Chelonia mydas) at foraging grounds in different locations throughout Australasia. My results are presented in a series of four chapters, each written as a standalone manuscript for publication. The literature cited is presented in a single reference list at the end of the thesis. This chapter provides a short introduction to marine turtles and the genetic tools that are relevant to the thesis. 1.1 Sea turtles of the world The order testudines, which includes all terrestrial, freshwater and marine turtles, began its evolution in the Triassic period at the beginning of the Mesozoic more than 200 million years ago (Márquez 1990). Sea turtles have existed since the late Jurassic some 150 million years ago, and are amongst the oldest inhabitants of the ocean. Throughout their evolutionary history, this group of animals has been highly diversified with more than 30 now extinct genera (Pritchard 1997; Mortimer et al. 2000). Having now entered the new millennium only seven species remain, all of which are classified under some form of legislation as endangered species. The extant species of sea turtles are classified into two families; the Dermochelidae, which contains only a single species, the leatherback turtle (Dermochelys coriacea), and Cheloniidae, containing the remaining six species; the green turtle (Chelonia mydas), the loggerhead turtle (Caretta caretta), the hawksbill turtle (Eretmochelys imbricata), the flatback turtle (Natator depressus), the Kemp s ridley turtle (Lepidochelys kempii), and the olive ridley turtle (Lepidochelys olivacea) (Pritchard 1997). An eighth taxon, the black turtle or pacific green turtle (Chelonia agassizii), is considered as an independent species or subspecies by some specialists (Pritchard 1997) but phylogenetic studies do not support this view (Naro- Maciel et al. 2008). Despite the fact that only seven species of sea turtles remain and that most are considered endangered or critically endangered (IUCN 2009), they are not considered a relict group (Mortimer et al. 2000). A long history of great cultural and 4
15 economic value to humans, resulting in overexploitation through egg harvesting, hunting, bycatch in fisheries and poor management, have brought many populations to critically low numbers (Witherington & Frazer 2003) Lifecycle of marine turtles All species of marine turtles are long-lived, have a complex life history, and migrate long distances during different life stages. The spatial and temporal scale involved in the marine turtle lifecycle presents a major challenge to research. The marine turtle life cycles are similar among all species with few differences among species, and to some degree, among different populations of the same species. This variation has principally to do with the time individuals spend in the different phases of the life cycle, distances travelling in migrations and the number and size of clutches (Figure 1.1). Female sea turtles reproduce on a one to nine year cycle, depending on the species and population (Miller 1997). Males may breed more frequently (Miller 1997, Limpus 1994). When the time comes, males and females travel variable distances to their mating areas; which can be less than 10 km or over 2000 km away (Mortimer and Carr 1987, Miller 1997). These areas are generally found in the vicinity of the nesting beaches (Kalb et al. 1992, Miller 1997), but can be separated by tens or even hundreds of kilometres (Limpus 1993; FitzSimmons, Limpus, et al. 1997). During the mating season, females often mate with several males, which may, or may not, result in multiple paternity of clutches (Owens 1980, FitzSimmons 1998; Jensen et al. 2006). Mating is generally believed to take place in the month or two prior to the first oviposition of the nesting season (Miller 1997, Mortimer et al. 2000). However, new molecular studies indicate that mating also possibly occurs during the inter-nesting period in areas where breeding males remain in the vicinity of nesting beaches for at least some of the nesting season (Chassin-Noria 2005). Female sea turtles crawl onto tropical or sub-tropical sandy beaches, where they lay between 65 and 180 eggs in a single nesting event, depending on the species. After successfully nesting, they will spend an inter-nesting period of 9-27 days in near-shore waters before returning for a second nesting. This will continue for one to six clutches in a nesting season. At the end of the nesting season the females migrate back to their foraging grounds to feed and regain energy stores for their next nesting season (Miller 1997). 5
16 Figure 1.1. Generalised lifecycle for most species of marine turtles. Species and populations vary mainly in the duration of the different phases. The Figure is divided into hatchling/juvenile (light grey line), female (dark grey line) and, male (black line) migration. Figure adapted from Lanyon et al. (1989). Marine turtle eggs hatch some days after being deposited (Miller 1997). The sex ratio of the hatchlings is dependent on the incubation temperature, with warmer temperatures producing more females, as seen in many reptiles (Ackerman 1997, Miller 1997, Mortimer et al. 2000). Once hatchlings break the surface of the sand, they crawl towards the sea, primarily guided by visual cues (Salmon & Witherington 1995). They move towards the brightest point (which in natural environments is the sea s horizon) and away from tall silhouettes (Miller 1997, Mortimer et al. 2000). Once they reach the water, the hatchlings head out to open sea. In most species, post-hatchling turtles (young pelagic turtles) undertake a passive pelagic migration drifting in oceanic gyres from two to ten or more years, depending on the species (Witherington 1994, Miller 1997, Mortimer et al. 2000). This phase of the sea turtle lifecycle is often referred to as the lost years (Carr 1987), as very little is known about it. After this pelagic stage, older juveniles of most species recruit back into the shallow benthic foraging 6
17 environments of coastal waters (Miller, 1997). In these developmental habitats, some individuals will stay in the same location until they reach puberty, while others may shift several times before settling in one location (Limpus et al. 2003, Miller 1997, Music & Limpus 1997). Once they reach sexual maturity and are ready to breed, both males (FitzSimmons et al. 1997) and females will migrate back to traditional breeding areas near their natal region where they will mate. Following this, females will come on to the beach to nest while males return to their foraging area. Most of what is known about marine turtles is gathered from the short period of their life they spend on the nesting beach. However, numerous studies have used mark-recapture methods and radio and satellite telemetry to investigate marine turtle migration patterns and homing abilities (Cornelius & Robinson 1986, Plotkin et al. 1995, Avens et al. 2003, Bentivegna 2002). These studies have revealed that marine turtles are highly migratory, sometimes migrating thousands of kilometres between foraging areas, breeding areas and nesting beaches. They have also shown that female turtles are philopatric and present an impressive homing ability allowing them to nest on the same beach season after season (Miller 1997). Many of the questions which cannot be answered with traditional methods are now being addressed with molecular techniques. Molecular studies of mitochondrial DNA (mtdna) have shown that many female marine turtles exhibit natal nest site fidelity by returning to the very same beach on which they hatched (Allard et al. 1994, Bowen & Karl 1997). Studies have shown precisions in nest site fidelity on different scales. Mediterranean loggerheads showed population structuring at a scale of 100 km (Schroth et al. 1996). Peare and Parker (1996) found evidence of fine scale natal homing for green turtles at Tortuguero, Costa Rica. There, females nesting closer together were more closely related that those separated by several kilometres. However, they could not replicate these results for the green turtles nesting at Melbourne Beach, FL, USA. Therefore, it is only fair to say that the true precision and extension of natal homing is not yet fully understood. Nonetheless, these and others studies have shown that there is genetic differentiation among females between nesting areas, which is important for informing conservation and management strategies of marine turtles. Also, we now know that the genetic structuring based on bi-parentally inherited nuclear DNA is not as strong as maternally inherited mtdna, indicating male-mediated gene flow between regional nesting beaches (FitzSimmons 1995, Bowen & Karl 1997, FitzSimmons et al. 1997). 7
18 Green turtles (Chelonia mydas) Green turtles are widespread and can be found in all tropical and subtropical seas. Threats to green turtles range from the harvest of turtles and eggs to incidental by-catch by fisheries and entanglement in discarded fishing gear. As a consequence, many populations around the world have been severely impacted and, globally, green turtles are listed as endangered by the International Union for Conservation of Nature and Natural Resources (IUCN, 2009). However, while populations may be declining and depleted in some areas, they are stable or increasing in others, and it has been argued that the conservation status of marine turtle populations should be focused on regions where particular populations are in decline, and not on a global scale (Mrosovsky 2004). In Australia, green turtles are listed as vulnerable under Commonwealth legislation (Environment Protection and Biodiversity Conservation Act 1999). A range of anthropogenic activities impact green turtles in this region such as incidental captures in shark control programs, by-catch in commercial fisheries, ghost net entanglement, and boat strikes (Limpus 2007). Green turtles are also affected by a number of diseases. Fibropapillomas are common among green turtles in locations such as Moreton Bay, Repulse Bay and Edgecombe bay in Queensland and has been reported from the Gulf of Carpentaria, Torres Strait and Western Australia, but seems to absent from offshore reefs (Limpus 2007, Limpus et al , Limpus and Miller 1994). Indigenous harvests occur on the east coast of Queensland, in Torres Strait, the Gulf of Carpentaria, Arnhem Land, the Northern Territory and Western Australia (Limpus & Chatto 2004). The geographic extent and demographic structure of the impact has not been quantified reliably. While the indigenous harvest may affect several species of marine turtles, it is particularly targeted at large adult green turtles. Several monitoring programs active in areas of harvest collect information on numbers of turtles killed, but the origin of these turtles is often unknown. Being able to trace the origins of turtles in areas of harvest is fundamental in order to determine whether populations are under threat from the current level of harvest and other impacts Green turtle population genetics Two of the most important contributions of genetic studies to our understanding of sea turtle biology were 1) the confirmation of natal homing in breeding adults and 2) the use of mixed stock analysis to determine the origins of sea turtles at foraging grounds or along migratory routes. This information is crucial to the conservation and management of these animals. 8
19 Natal homing behaviour of green turtles has allowed the use of the genetic tools for deducing population structure, migration routes and identifying the source population of individual turtles. By returning to their natal region to lay their eggs, nesting females establish a strong genetic differentiation among nesting populations over time (Meylan et al. 1990). This means that single stocks associated with specific rookeries and breeding areas can be identified by sequencing the fast-evolving mitochondrial DNA (mtdna) control region. The resulting differences in haplotype frequencies have been characterised for green turtle breeding populations in the Caribbean (Bass et al. 1996, Encalada et al. 1996, Engstrom et al. 1998, Espinosa et al. 2000), the Mediterranean (Encalada et al. 1996), Africa, (Formia et al. 2006), Ascension Island (Meyland et al. 1990), Southwest Indian Ocean (Bourjea et al. 2007), and the eastern Indian Ocean and Australasia (Dethmers et al. 2006, FitzSimmons et al. 1997, Formia et al. 2006, Moritz et al. 2002, Norman et al. 1994). Seven genetically distinct breeding populations, referred to as Management Units (MUs) (Moritz 1994), have been characterised for 13 green turtle rookeries throughout Australia including; the southern Great Barrier Reef (sgbr) MU, northern Great Barrier Reef (ngbr) MU, Coral Sea MU, Gulf of Carpentaria (GoC) MU, Ashmore Reef MU, Scott Reef MU, and the Northwest Shelf MU. Furthermore, 14 rookeries have been characterised for neighbouring countries (Indonesia, Malaysia, Papua New Guinea, Philippines, Micronesia, and New Caledonia) resulting in identification of a further ten MUs (Dethmers et al. 2006, Moritz et al. 2002). Following characterisation of the mtdna polymorphisms for each MUs in a region, mixed stock analysis (MSA) provides a method of examining the relationships among marine turtle nesting and foraging locations. Tagging studies of nesting females and subsequent recaptures at foraging grounds have shown that green turtles in Australia recruit to foraging grounds from multiple nesting beaches (Limpus, 1993, Limpus et al. 2003, Limpus et al. 1992). By using MSA, it is possible to determine the relative contributions of sources, or stocks, to genetically mixed foraging populations. Lahanas et al. (1998) were the first to determine the contributions of multiple Caribbean green turtle nesting locations to the foraging aggregation in the Bahamas. Since then, MSA has become a widely used tool to determine the composition green turtle foraging grounds (Bass et al. 2006, Luke et al. 2004, Bass et al. 1998, Bass & Witzell, 2000). 9
20 An understanding of the geographic range of marine turtle stocks is extremely important for developing effective management plans. If the range of a marine turtle stock crosses international borders, it could be protected in part, but not all, of its migrational range. While the MSA methodology is a valuable tool, it has its limitations. Small sample sizes, incomplete sampling of rookeries and low genetic differentiation between rookeries all contribute to the large confidence intervals that often accompany MSA estimates. Chapter 2 provides a review of MSA and its application to marine turtle studies. The review summarises the extensive literature on this subject and provides a critical review on the advantages, the limitations and the future of MSA in marine turtles. Two of the biggest limitations to MSA are a lack of genetic structure among source rookeries that results from common haplotypes that are shared across rookeries and also limited sampling of source rookeries. These problems are particularly relevant for the green turtle rookeries in the eastern Indian Ocean and Southeast Asia where two common haplotypes are shared among most populations (Dethmers et al. 2006). This results in large confidence intervals surrounding MSA estimates of foraging green turtles in the region (Dethmers, unpublished). In Chapter 3, I re-assess the genetic structure of green turtle rookeries in the eastern Indian Ocean and Southeast Asia. In order to increase the genetic resolution needed for effective MSA, I reanalyse samples used by Dethmers et al. (2006) using a longer sequence of the mtdna. With the addition of several new green turtle rookeries and increased sample sizes for previous rookeries, the sequences from both the short fragment used by Dethmers et al. (2006) and the new longer fragment were analysed. The genetic population structure of the region is re-evaluated and four new sites included in the revised management units of the region. The effect these additions have on MSA is tested by analysis of two foraging aggregations at Shark Bay and Cocos (Keeling) islands. One obvious advantage of MSA is that it allows for the assessment of the distribution of juvenile turtles across foraging grounds for which little information exists. In Chapter 4, I apply MSA to juvenile foraging aggregations off Sabah Malaysia. A highly female biased sex ratio among foraging immature turtles raises questions about the origin of these turtles. This is the first study to identify the origin of juveniles in Southeast Asia and highlights the importance of monitoring marine turtles at the foraging grounds to evaluate the effects of threats on nesting beaches and foraging grounds. 10
21 It has been suggested that juvenile turtles may tend to forage in the vicinity of their natal nesting colony. If so, how precise is the homing that brings juvenile oceanic migrants back to foraging habitats near their location of origin? If homing is precise, it will carry the expectations of population genetic differences among regional juvenile feeding cohorts, similar to that of nesting populations. The east coast of Australia provides an ideal opportunity to examine the within species regional variation in migratory behaviour to determine how distinct breeding populations are distributed across the available foraging grounds. This is explored in Chapter 5 with special focus on how different stage classes of green turtles are distributed both temporally and spatially. In Chapter 6, I summarise the findings of the thesis and explore the outcomes in the context of what is observed in these species at a global scale. I offer some thoughts on what the future might bring and what direction future research should take to improve the use of MSA in marine turtles. 11
22 Chapter 2 2. Review: Mixed Stock Analysis and marine turtles 2.1 Introduction Migratory marine animals have long puzzled researchers who try to monitor and understand their complex life history. Having various stages of their lifecycle separated by large distances, coupled with long generation times, makes detailed research particularly challenging. The ability to link animals at foraging grounds, or those encountered along migratory routes, back to their breeding habitat is a vital part of effective management and conservation. Several methods have been used to identify the origin of highly migratory species. Stable carbon and oxygen isotopes in the otoliths of Atlantic bluefin tuna, Thunnus thynnus, showed that foraging animals caught by fisheries off the U.S. Atlantic coast originated from spawning grounds in the Mediterranean (Rooker et al. 2008). Molecular markers (mtdna) demonstrated that 46% of whale meat products sold on Japanese markets was minke whale, Balaenoptera acutorostrata scammoni, originating from the depleted and protected Sea of Japan stock (Lukoscheck et al. 2009) Efforts to understand and conserve marine turtle populations face similar challenges. Most species are widespread in tropical and subtropical waters, and hundreds of nesting beaches and foraging grounds produce a complex network of migration routes. As turtles hatch from tropical and subtropical beaches around the world, they spend several years at the mercy of the prevailing currents (Musick & Limpus 1997). As they grow larger, most species recruit back into benthic foraging habitats where they may, or may not, stay for the rest of their lives (Limpus & Walter 1980, Henwood & Ogren 1987, Limpus et al. 1994, Musick & Limpus 1997, Senko et al. 2010). Reaching adulthood, they only leave the foraging area when they make their breeding migration back to their natal region, which can be nearby or several thousand kilometres away (Limpus 2007, 2008). For over nearly half a century, mark-recapture studies have revealed the remarkable migratory feats of marine turtles. We know from these early tagging studies that foraging areas are 12
23 composed of turtles from a mixture of different rookeries (Limpus et al. 1992, Troëng et al. 2005b, Troëng et al. 2005a, Moncada et al. 2006), resulting in the term mixed stocks (Limpus et al. 2009). Both early mark-recapture studies and more recent satellite telemetry studies have revealed links between rookeries and foraging habitats in marine turtles (Bentivegna 2002, Godley et al. 2002, 2003). However, these methods are generally restricted to adult animals and leave out the non-adult portion of the population. While tagging studies remain an important tool for understanding the composition of adult turtles at mixed foraging grounds, molecular techniques have opened up new possibilities. In a study of Atlantic green turtles, Bowen et al. (1992) showed that turtles nesting on Ascension Island and those nesting in Surinam had fixed haplotype differences for mtdna, despite these two populations mixing at shared coastal foraging grounds in Brazil. This was the first proof of natal homing in marine turtles since Archie Carr first proposed the hypothesis in 1967 (Carr 1967). Subsequently, it has been shown that this pattern of mtdna differentiation among rookeries is a common feature amongst all species of marine turtles, albeit to varying degrees (Bowen et al. 1992, Bowen & Karl 1996, Encalada et al. 1996, Bowen et al. 1998, Shanker et al. 2004, Bowen & Karl 2007). When mtdna haplotypes show large frequency shifts among rookeries, they can be used to deduce the natal origin of juvenile and adult turtles captured in migration corridors or at coastal foraging habitats. Marine turtle biologists successfully adopted a method called Mixed Stock Analysis (MSA), which was originally developed for analysing mixed populations of fish. It proved to be easily adapted to marine turtle mixed stocks and has provided valuable new insights into the dispersal of marine turtles (Lahanas et al. 1998, Bass & Witzell 2000, Velez-Zuazo et al. 2008). In this review I will present what has been learned since the first MSA studies in marine turtles more than a decade ago. First, I will present background information on the different methods used for MSA. Next, I will review the large number of studies that have been done for three species of marine turtles, the loggerhead turtle, the hawksbill turtle and the green turtle. I will then highlight some of the main hypotheses about what drives the composition of foraging grounds in marine turtles (rookery size, distance, and ocean currents), and discuss how MSA can be used to better inform management plans. Finally, I will outline problems with applying molecular markers to the estimation of foraging ground composition, and suggest future directions for further improvement of our understanding of the connection between rookeries and foraging habitats in marine turtles. 13
24 2.2 Mixed Stock Analysis Mixed Stock Analysis (MSA) was originally developed to estimate the contributions of genetically differentiated salmon stocks from different rivers to mixed stocks caught in oceanic fisheries (Grant et al. 1980, Pella & Milner 1987). Because salmon and marine turtles share life history traits such as natal homing that result in genetically differentiated breeding stocks and subsequent mixing in foraging habitats, the method has been applicable to marine turtles. Early versions of the method used a Maximum Likelihood (ML) approach to estimate stock composition of a random sample of a mixture from data that consists of the observed genotypic frequencies of the mixture and the observed genotypic frequencies in the source populations. Use of an iteratively re-weighed least square (IRLS) algorithm computes a conditional maximum likelihood estimate of composition using modified weights along with the composition vector from one iteration to the next. An EM algorithm is employed to constrain estimates such that the likelihood function is non-decreasing during the search (Pella & Milner 1987). Later, a Bayesian (Pella & Masuda 2001) model was developed that is considered to have a number of advantages over the ML method. Most importantly the ML assumes that the relative frequency in the source population is known without error. Also, the ML method assumes complete sampling of the possible source population so that a haplotype not found in a source population is assumed to be absent. The reality for marine turtles is that baseline samples are collected from a subset of nesting beaches and sample sizes are often small. They therefore have some uncertainty associated with them Unlike the ML approach, the Bayesian model uses the information from the mixture sample to improve the relative frequencies of the baseline samples. This method treats the baseline relative frequencies as unknown, but with a specified prior distribution. Then, using the baseline and the mixture samples, the posterior prior distribution is set. Importantly, the Bayesian model allows for the presence of rare haplotypes such as singletons or those that occur at < 5% (Pella & Masuda 2001). This means that haplotypes that are actually present in the source populations but not detected because of small sample size are still considered. This 14
25 is particular advantageous in the analysis of turtle mtdna control region data, which is characterized by many rare haplotypes in both source populations and mixed aggregations. Bayesian approaches allow for the incorporation of informed priors that will influence the prior distribution of the relative baseline frequencies. These, are normally educated inputs that help improve the final estimates in situations where common haplotypes affect the baseline sample. For example, the relative size of source populations or the distance to them can be used as priors if these are thought to be proportional to their contribution to the mixture. The Bayesian model therefore provides a means of reducing the negative effects of sampling error (Pella & Milner 1987, Smouse et al. 1990, Bolker et al. 2003) and source population size (Okuyama & Bolker 2005). Most recently Bolker et al. (2007) developed the many-to-many approach specifically with marine turtles in mind. This approach uses a Bayesian hierarchical extension of the Bayesian mixed stock analysis, and applies it to a population framework, allowing for a group of several foraging ground mixtures and source populations to be analysed together. While these methods are computationally and philosophically different, they all use haplotype frequencies to estimate the probability that a sample from the mixed population (e.g. marine turtle foraging ground) has originated from each of the baseline populations. While often far from ideal, under the right conditions, these methods will be able to provide robust estimates of the relative levels of contribution from breeding stocks to a sample of animals caught away from the breeding area. This will hold across all gender and all size classes, something that traditional tracking has yet to achieve. 2.3 MSA and Marine turtles After several studies of marine turtles showed that regional rookeries are genetically differentiated (Bowen et al. 1993, Lahanas et al. 1998, Bourjea et al. 2007, Bowen & Karl 2007, Carreras et al. 2007), it became possible to trace back turtles caught away from their nesting beach. Since the early 1990 s, numerous studies used this method to shed light on the rookery origins of marine turtles in the pelagic stage (Bowen et al. 1995, Bolten et al. 1998), in juvenile benthic foraging grounds (Bass & Witzell 2000, Engstrom et al. 2002, Velez- Zuazo et al. 2008), in adult foraging grounds (Bass et al. 1998, Velez-Zuazo et al. 2008) in 15
26 Table 2.1. Studies using Mixed Stock Analysis of marine turtle foraging aggregations; categorised by species and region. Loggerhead turtle (C. caretta) Green turtle (C. mydas) Hawksbill turtle (E. imbricata) Leatherback turtle (D. coriacea) Atlantic-USA Sears et al Bass et al Bowen et al Roberts et al Reece et al Rankin-Baransky et al Atlantic-East Bolten et al Monzón-Argüello et al Atlantic-Brazil Reis et al Mediterranean Laurent et al Maffucci et al Carreras et al Casale et al Monzón-Argüello et al Pacific Bowen et al Boyle et al Caribbean Engstrom et al Caribbean Bass et al Lahanas et al Luke et al Bolker et al Bjorndal & Bolten 2008) Richardson et al Atlantic-USA Bass et al Bass & Witzell 2000 Atlantic-Brazil Naro-Maciel et al Proietti et al Bjorndal et al Pacific-Hawaii Dutton et al Australia Dethmers unpub. Jensen - Chapter 5 Jensen - Chapter 3 SE Asia Jensen- Chapter 4 Caribbean Bowen et al Bass 1999 Diaz-Fernandez et al Bowen et al Velez-Zuazo et al Blumenthal et al Browne et al Troëng et al. 2005b Richardson et al Australia Broderick et al Bell et al. Unpub. Jensen et al. in prep Indian Ocean Jensen et al. Unpub. Atlantic-Brazil Vargas et al fisheries by-catch (Bowen et al. 1995, Laurent et al. 1998) and in strandings (Rankin- Baransky et al. 2001, Maffucci et al. 2006) (Table 2.1). The different natural history of marine turtle species means that only three of the world s seven species are suitable candidates for MSA, the loggerhead, hawksbill and green turtles. The relevant shared features are genetically differentiated regional rookeries, and spatially 16
27 restricted benthic foraging aggregations that in some areas are easily sampled. Conversely, the leatherbacks, olive ridley and flatback turtles show low divergence among regional nesting populations, making the MSA less effective. In addition, these species inhabit distant, spatially disjunct foraging grounds that are difficult to study. Therefore, the following sections focus on loggerhead, hawksbill and green turtles Loggerhead turtles (Caretta caretta) One of the first studies to use MSA on marine turtles solved an old enigma regarding juvenile loggerhead turtles in Mexico. Large groups of juvenile turtles are found foraging off the coast of Baja California while the nearest known rookeries for this species in the Pacific are found in Japan and Australia. Initially, researchers thought it impossible that the turtles foraging in Mexico could have hatched more than 10,000 kilometres away on the other side of the Pacific Ocean, so they believed an unknown eastern Pacific loggerhead turtle rookery must exist. Subsequently, the mtdna haplotypes from nesting loggerhead turtles collected from rookeries in Japan and Australia were compared with those of juvenile turtles in Mexico. The MSA linked most (>95%) of the eastern Pacific loggerhead turtles to nesting beaches in Japan and possibly Australia (<5%) (Bowen et al. 1995). This demonstrated for the first time the truly remarkable voyages of marine turtles. The Australian haplotype was subsequently found at a Japanese rookery, thus making the requirement of an Australian contribution to the Mexican groups unnecessary (Hatase et al. 2002). The question remained of where loggerhead turtles born in eastern Australia spend their oceanic stage. Recently it has been confirmed that juvenile loggerhead turtles caught in commercial fisheries off the coast of Peru were traced back to nesting beaches in Australia (and New Caledonia) (Boyle et al. 2009). This demonstrated that loggerhead turtles in eastern Australia make similar migrations using the anti-clockwise currents of the South Pacific. These are the only two studies from the Pacific where molecular data has been used to identify the origins of foraging loggerhead turtles. The bulk of mixed stock analysis for loggerhead turtles is from the Atlantic and Mediterranean Oceans (Table 2.1). The Atlantic loggerhead turtles undertake long distance developmental migrations similar to that seen in the Pacific. Juvenile loggerhead turtles found in oceanic foraging aggregations around the Azores and Madeira in the eastern Atlantic are estimated to originate from nesting beaches in Mexico (~10%), south Florida (~70%) and northern Florida to North Carolina (~20%) (Bolten et al. 1998). Some of these eastern Atlantic turtles even pass through the 17
28 Strait of Gibraltar and enter the western Mediterranean, where ~50% of loggerhead turtles caught in pelagic drifting long line fisheries have been shown to originate from eastern Atlantic rookeries and the remaining come from Mediterranean rookeries (Laurent et al. 1998, Carreras et al. 2006). These contributions are roughly proportional to the size of the source rookeries. The post-hatchling turtles spend a number of years in the pelagic stage passively drifting in the prevailing ocean currents before reaching a size where they can safely shift to benthic foraging along coastal areas (Musick & Limpus 1997). As with loggerhead turtles in the Pacific, those originating from western Atlantic rookeries that forage in the eastern Atlantic and Mediterranean traverse back across the Atlantic to recruit into coastal areas along the eastern seaboard of the United States (Laurent et al. 1998). This is supported by the findings that the majority of foraging loggerhead turtles found in neritic habitats throughout the Mediterranean originate from local rookeries (Laurent et al. 1998, Maffucci et al. 2006) and the same is true for those found off the south-eastern United States (Bass et al. 2004, Bowen et al. 2004, Reece et al.2006). Once loggerhead turtles have entered the neritic foraging aggregations, the stock contributions are no longer proportional to the size of the rookeries alone, because these turtles tend to recruit into foraging areas near their natal origin (Sears et al. 1995, Witzell et al. 2002, Bowen et al. 2004, Reece et al. 2006). This led to a juvenile natal homing hypothesis in which post-hatchling loggerhead turtles from different rookeries get mixed in the oceanic stage. Here they undertake long distance dispersal to oceanic foraging areas in the eastern Atlantic and Mediterranean. As they prepare to switch to neritic foraging areas, loggerhead turtles actively return to their region of birth, where they shift to benthic foraging (Bowen et al. 2004, Reece et al. 2006). Although this homing is not as precise as that seen in nesting females, this behaviour still brings juvenile loggerhead turtles back into the vicinity of their natal origin Hawksbill turtles (Eretmochelys imbricata) Hawksbill turtles are found in more tropical waters than the more temperate loggerhead turtles and they display a number of differences. They inhabit tropical foraging grounds to which they show strong fidelity and their feeding ecology keeps them associated with the coral reefs (Limpus 2008). For many years hawksbill turtles were thought to be non migratory (Witzell 1983). However, this hypothesis was refuted by early studies using molecular data to infer distant migratory links among hawksbill turtle breeding and foraging grounds (Broderick et al. 1994, Bowen et al. 1996, Bass 1999). Although their migrations are not 18
29 comparable in scale to those of loggerhead turtles, a number of studies of hawksbill turtle foraging grounds in the Caribbean have shown that rookeries throughout the Caribbean were contributing to these foraging aggregations (Bowen et al. 1996, Bass 1999, Bowen et al. 2007, Velez-Zuazo et al. 2008). Unlike the loggerhead turtles there is little evidence of trans- Atlantic dispersal for Caribbean hawksbill turtles and they seem to be confined within the Caribbean. The complex ocean currents of the Caribbean region differ from the ones influencing the Atlantic loggerhead turtles and they appear to play an important role in generating the different dispersal patterns seen in the Caribbean hawksbill turtles. Furthermore, these studies also support the model whereby a combination of rookeries size and proximity to the foraging ground influences the composition of foraging aggregations (Bass 1999), with the addition of the important influence of ocean currents (Velez-Zuazo et al. 2008, Blumenthal et al. 2009). On the Great Barrier Reef (GBR), it has been suggested that immature hawksbill turtles undertake developmental migrations between different foraging grounds as they mature (Limpus 2008). In the southern GBR, hawksbill foraging aggregations at Heron Island are dominated by immature turtles (Limpus 2008). Conversely, at the northern GBR foraging ground, mainly adult hawksbills are found (Limpus 2008). Recent genetic studies of adult hawksbill turtles foraging at the Howicks Group of islands in the ngbr have showed that the majority (>80%) of the turtles are not from the neighbouring rookery but from the region surrounding the Solomon Islands (Solomon, Papua New Guinea), with only minor contributions from Australian rookeries (Bell et al. unpubl. data) and this is supported by mark-recapture data (Limpus 2008). Further genetic studies are underway to determine the origin of hawksbill turtles at Heron Island that should either support or refute the hypothesis of developmental migration between the sgbr and the ngbr (Jensen et al. unpubl. data) Green turtles (Chelonia mydas) Similar to hawksbill turtles, green turtles are more tropical in their distribution and their posthatchling migrations are thought to be intermediate to those of hawksbill turtles and loggerhead turtles. As observed for hawksbill turtles, green turtle foraging grounds in the Caribbean receive contributions from rookeries throughout the Caribbean region (Lahanas et al. 1998, Luke et al. 2004, Bass et al. 2006, Bolker et al. 2007, Bjorndal & Bolten 2008). Evidence of long distance migration is also observed, with a large (25%) contribution to 19
30 Caribbean foraging grounds coming from Ascension Island (>5500 km distant) (Luke et al. 2004). Green turtles foraging in the southern Atlantic off the coast of Brazil are mainly from Ascension Island but with small contributions from rookeries in the wider Caribbean and possibly from Africa (Bjorndal et al. 2006, Naro-Maciel et al. 2007, Proietti et al. 2009). At the northern extreme, foraging areas in North Carolina are mainly composed of turtles from the east coast of the United States and Mexico, with small contributions from other Caribbean and Atlantic rookeries (Bass et al. 2006). Analyses of green turtles from the Hawaiian archipelago showed that both nesting and foraging aggregations across the entire archipelago all comprise the same genetic stock, with little to no input from other rookeries in the eastern or western Pacific (Dutton et al. 2008). In the Australasian region, recent research by Dethmers et al. (unpubl data) conducted on foraging grounds in the Northern Territory, Aru (to the north) and Ashmore Reef (off Western Australia), indicates a complex mixture of contributing rookeries. The only other research on foraging grounds in this region is reported in this thesis for Malaysia (Chapter 4) the Great Barrier Reef (Chapter 5) and Western Australia (Chapter 3) Complex life history One thing that MSA studies have confirmed is that marine turtles have complex life histories. The patterns seen from MSA differ not only between species but also between different populations or regions within the same species. In some areas, both loggerhead and green turtles show strong fidelity to their neritic foraging area throughout their life (Limpus et al. 1992). In other areas they switch between different developmental habitats and then migrate to a separate adult foraging ground as they reach sexual maturity (Carr et al. 1978, Bjorndal & Bolten 1997, Bjorndal et al. 2003, Godley et al. 2003). A re-occurring theme in MSA of marine turtles has been to try to identify the mechanisms that generate the composition of foraging grounds. Several hypotheses (discussed below) have been proposed to explain the importance of factors such as rookery size, distance between rookeries and foraging grounds (juvenile natal homing), behaviour and ocean currents Rookery size and distance An early study by Bolten (1998) found that rookery contributions to aggregations of pelagic loggerhead turtles were proportional to the size of the rookery. Results from stranded immature loggerhead turtles in the north-eastern United States (Rankin-Baransky et al. 2001), 20
31 and juvenile foraging loggerhead turtles in North Carolina (Bass et al. 2004) are consistent with this model. Similarly, juvenile green turtle foraging aggregations in the Bahamas (Lahanas 1998) appear to originate mostly from large rookeries. It has been suggested that for some species or populations, turtles hatching from different rookeries first become mixed in the pelagic stage and as they mature, they randomly move into the available benthic habitats. Under this scenario the proportion of post hatchlings in the pelagic stage and immature turtles in the benthic habitat is relative to the rookery size of the contributing stocks. Such a pattern of random mixing and recruitment into benthic foraging grounds should result in regional foraging grounds being genetically similar and with no or little genetic structure among them (Figure 2.1). While the model of random recruitment describes the results of some studies (Lahanas et al. 1998), it does not explain the contrasting patterns of many other studies. For example, Bass and Witzell (2000) found significant genetic differences between the juvenile green turtles sampled from a foraging ground in east-central Florida and those sampled at a foraging ground in the Bahamas (Lahanas et al. 1998) despite the two locations being separated by only 350 km. This is not consistent with the expectation of homogenous regional foraging grounds if the composition were driven by rookery size alone. Some researchers have suggested that juvenile turtles recruit to foraging areas that are located near their natal rookeries, as in the case of loggerhead turtles (Laurent et al. 1998, Engstrom et al. 2002, Bass & Witzell 2000). In this scenario, post-hatchling turtles will mix in the pelagic stage but as they mature and recruit back into the benthic habitats along the coast, they actively seek out foraging areas closer to their natal rookeries (Figure 2.2). Bowen et al. (2004) tested this hypothesis of juvenile natal homing behaviour in a study of ten juvenile loggerhead turtle foraging aggregations across the eastern USA. They found evidence for significant population structure across all forging grounds and showed that juvenile loggerhead turtles recruit into benthic foraging grounds closer to their natal rookery. Several studies have corroborated the patterns suggested by juvenile natal homing for green turtles (Chapter 5, Bass & Witzell 2000, Naro-Maciel et al. 2007), loggerhead turtles (Bowen et al. 2004) and hawksbill turtles (Bowen et al. 2007). 21

32 Figure 2.1. Model of random mixing of pelagic turtles and subsequent random recruitment into benthic foraging grounds that are genetically similar. 22

33 Figure 2.2. Model of random mixing of pelagic turtles and subsequent selective recruitment into benthic foraging grounds near natal rookeries, leading to genetic structure among foraging grounds. 23

34 Figure 2.3. Model of non-random mixing of pelagic turtles due to oceanic currents and subsequent selective recruitment into benthic foraging grounds near natal rookeries, leading to genetic structure among foraging grounds and among pelagic turtles in different regions. 24
35 That larger rookeries in a region contribute more turtles to associated foraging grounds is intuitive. Given the strong homing behaviour of adult females, and possibly males (FitzSimmons et al. 1997b) to nest and breed in the vicinity of their natal region, the idea of juvenile homing behaviour is a convincing one. Nonetheless, there are several cases of conflicting results even within the same species when trying to explain foraging ground composition based on rookery size and/or distance alone Ocean Currents While size and distance might largely explain how marine turtles are distributed across foraging grounds, the results are not clearly evident. Disproportionately large or small contributions from some rookeries cannot be explained by these factors alone. Green turtles foraging around Barbados in the West Indies showed large (25%) contributions of turtles from Ascension Island, more than 5500 kilometres away, and substantial contributions (19%) from Tortuguero in Costa Rica, where the rookery is at least an order of magnitude larger and only 2600 kilometres away. There was also a substantial contribution (18.5%) from the distant and much smaller south Florida rookery (Luke et al. 2004). These data indicate that neither distance nor size play a major role in recruitment to this foraging area. The authors suggest that ocean currents offer a plausible explanation to this pattern (see Figure 2.3). Barbados is located at the point where the north equatorial and the south equatorial currents meet and both Ascension Island and south Florida rookeries feed into these two major Atlantic current systems. Costa Rica on the other hand is affected by smaller and more local current systems that would bring fewer post-hatchling turtles towards Barbados. Another convincing example of the role of ocean currents can be found for loggerhead turtles caught in the Mediterranean Sea. Here, foraging aggregations in the western Mediterranean are mainly fed by Atlantic rookeries whereas those in the east are mainly composed of Mediterranean rookeries, thus providing a strong association between water bodies and ocean current systems (Carreras et al. 2006). Likewise, as the South Equatorial Current approaches the east coast of Australia, it splits into the southward East Australian Current and the northward North Queensland Current. This pattern possibly influences the strong partitioning of foraging green turtles between the ngbr and the sgbr (Chapter 5). High resolution ocean current data has been used to model the movement of passively dispersing juvenile hawksbill turtles. These models showed a significant correlation between foraging compositions generated by ocean current models and those from MSA for a number of foraging aggregations throughout 25
36 the Caribbean (Blumenthal et al. 2009). These discrepancies illustrate the complexity of how some or all these factors influence the composition of marine turtle foraging grounds Temporal differences Temporal variation in the genetic structure of nesting stocks may also influence the genetic composition of foraging grounds. Most species of marine turtles have nesting intervals of 2-5 years (Miller 1997) and it has been suggested that sampling from any one year could misrepresent the true genetic composition of the rookery. However, tagging studies have shown that nesting intervals are highly dynamic and turtles that nest together one year won t necessarily nest in the same year during subsequent nesting events (Solow et al. 2002). This should result in strongly homogeneous regional rookeries across short time scales. Furthermore, several studies have found no evidence for temporal differences in green turtles from Costa Rica (Bjorndal et al. 2005), and Australia (Chapter 5), nor for loggerhead turtles in Japan (Hatase et al. 2002), nor hawksbill turtles in Puerto Rico, (Velez-Zuazo et al. 2008). Variation on a larger timescale is more probable and unknown. Temporal variation in the composition of turtles at foraging grounds is another consideration. Foraging aggregations are highly dynamic, especially when composed of multiple rookeries. Seasonal movement is common in both green turtles and loggerhead turtles along the east coast of the United States (Avens & Lohmann 2004) and developmental migrations from strictly juvenile to adult foraging grounds is common in loggerhead turtles (Bolten et al. 1998, Bjorndal et al. 2000, McClellan & Read 2007), has been reported for green turtles (Godley et al. 2003, Bjorndal et al. 2005, Pilcher 2010) and for hawksbill turtles (Whiting & Koch 2006, Grossman et al. 2007, Blumenthal et al. 2009). In other areas, juvenile and adult turtles share foraging grounds and juveniles show strong fidelity to the same area throughout their life (Limpus 1992, Broderick et al. 1994, Limpus et al. 1994). The extent to which these different patterns in the use of foraging grounds, or the specific locations of foraging grounds, are related to temporal variation in the stock composition of foraging aggregations is not well known. Unfortunately, most early studies have limited samples from only one sampling event but temporal variation appears to be limited, at least at a shorter timescales. The few studies that have sampled across several years have found little to no temporal variation. Bass et al. (2004) found no temporal variation in haplotype frequency for immature loggerhead turtles at a North Carolina foraging aggregation sampled over three consecutive 26
37 years. Naro-Maciel (2007) found no temporal variation in green turtle foraging grounds in Brazil, and Velez et al. (2009) found no evidence of temporal variation in a five-year study of hawksbill turtles from Puerto Rico. The only study to report temporal variation in foraging grounds is a 12 year study from a highly dynamic foraging ground for immature green turtles in the Bahamas and found the haplotype frequencies from only one of the years to be significantly different (Bjorndal & Bolten 2008). Nonetheless, marine turtle foraging populations are unlikely to be static. The recruitment of juveniles from several rookeries is a complex process that is affected by variation in output from rookeries caused by variation in nesting numbers, natural catastrophes, predation and human impacts as well as varying ocean currents. These changes at rookeries or in ocean currents are likely to be reflected in foraging ground compositions. Temporal variation in the composition of foraging aggregations is expected if they are comprised of turtles from a large number of rookeries, and for highly dynamic foraging aggregations where juveniles stay for a short amount of time, such as in the Bahamas (Bjorndal & Bolten 2008). This pattern should be masked at foraging grounds that include older turtles and a more diverse range of size classes when sampling across different age groups. Because temporal variation in foraging ground composition violates an assumption of temporal stability when conducting mixed stock analyses, it is important to know if samples are collected across several years and size classes Sexr differences Male green turtles breed more often than females (Limpus 1993) and as a result the amount of energy allocated into reproduction differs between males and females. This suggests that a greater proportion of males might be living in closer proximity to their breeding grounds. Satellite telemetry data from seven nesting females and eight breeding male hawksbill turtles from Mona Island, Puerto Rico showed that males undertook significantly shorter postbreeding migrations (mean = 101 km, n = 8) than females (mean = 867 km, n = 7) (van Dam 2008). Also, foraging areas where turtles from rookeries that are female-biased due to warmer incubation temperature, mix with turtles from cooler male-producing rookeries would be expected to generate different MSA estimates between males and females (see Chapter 4). Bass et al. (1998) found a small difference in the contribution between males and females from different rookeries at a green turtle foraging ground in Nicaragua. However, sample sizes were small (30 for each gender) and the results remain inconclusive. Sex-based dispersal 27
38 remains poorly known in marine turtles. Because marine turtles lack obvious morphological sex characteristics prior to maturity, the gonads of non-adults must be examined using laparoscopy (Miller & Limpus 2003), or via hormonal assays (Diez & Van Dam 2003). This compounds the logistical difficulties in sampling a sufficiently large number of both males and females, especially if sex ratios are highly skewed. As a result of these challenges most studies have been unable to analyse foraging composition by gender Size class differences As mentioned earlier, some foraging grounds are strictly developmental and composed mainly of immature turtles. In a study of hawksbill turtles from Mona Island, Puerto Rico, juvenile hawksbills are likely recruited from other Caribbean rookeries whereas the majority of the adult male turtles originated from Mona Island (Velez-Zuazo et al. 2008). However, these were breeding males and not confirmed residents of the Mona Island foraging aggregation. There are limited studies from foraging grounds with mixed age classes. Most green turtle foraging grounds along the Great Barrier Reef in Australia are shared among all size classes from newly recruited juvenile turtles (~40 cm CCL) to large adults (Limpus 2007). A largescale comparison among size classes at six major foraging aggregations along the east coast of Australia is presented in Chapter Limitations with MSA Mixed Stock Analysis has provided valuable new insights into the distribution of marine turtles but in most cases the estimates are affected by large uncertainty. Several issues in the effective use of MSA need to be addressed concerning study design, sample sizes and the resolution provided by the genetic marker. Ideally, mtdna haplotypes would show unique haplotype shifts among rookeries making it straightforward to assign individuals to their natal rookery. However, marine turtle rookeries rarely have complete fixation of haplotypes and as a result the estimates of foraging ground composition derived from MSA may suffer from lack of resolution or rest on unmet assumptions, clouding our ability to successfully determine contributions. 28
39 Sample size How big a sample size is needed for a robust MSA analysis? Increasing source (rookery) sample size is considered more important than increasing mixture (foraging) sample (Xu et al. 1994). The saturation point where there is sufficient representation of the haplotype frequencies at a rookery varies and depends on the number and variability of the haplotypes, but as long as there is a substantial overlap in haplotype frequencies, large sample sizes will not fix the problem Source populations Probably the most important criterion for a successful MSA is the coverage of all possible source rookeries, especially when populations share widespread haplotypes. This problem should decrease with an increase in the number of rookeries sampled. On the other hand, too many rookeries can also alter the output from the MSA significantly. For example, for loggerhead turtles foraging at Chiriquí lagoon in Panama, inclusion of two Mediterranean rookeries altered the contribution of the large South Florida rookery from 65-71% (estimates from three different programs) to a minimal contribution of 0-11%, and the contribution of the small population on the Florida panhandle from 0 to 23-29% (Engstrom et al. 2002). It such cases it is important to assess the data carefully. In the case of Chiriquí lagoon, the problem was attributed to a common haplotype shared between the Mediterranean and western Atlantic rookeries. CC-A6, a haplotype endemic to the Mediterranean, was not detected in the foraging sample, thus justifying the removal of those rookeries from the analysis. This highlights the fact that results from MSA should be interpreted in light of sampling effort and must be critically evaluated in order to identify caveats in the method or study design. The number of orphan haplotypes, those not observed at the rookeries but seen in foraging grounds, is another indication of inadequate sampling of source populations. Medium frequencies of orphan haplotypes are often indicative of an un-sampled source, while low frequencies of orphan haplotypes are indicative of either an un-sampled source or insufficient sampling of already sampled rookeries. As more rookeries are characterised for mtdna variation, the number of orphan haplotypes seen in foraging aggregations should decrease. 29
40 Resolution of genetic markers The presence of common mtdna haplotypes among populations is another major drawback for marine turtle MSA. Examples of this include the common loggerhead turtle haplotype, CC-A2, found across western Atlantic and Mediterranean rookeries (Bowen et al Carreras et al. 2007), haplotypes C1 and C3 that are shared among green turtle rookeries in the Indo-Pacific (Dethmers et al. 2006, Chapter 3) and the A and F haplotypes that dominate the Caribbean hawksbill turtle rookeries (Velez-Zuazo et al. 2008). As a result, MSA estimates for these regions are surrounded by large errors, thus calling into question how precisely these estimates reflect the true origin of sea turtles aggregated in the foraging areas. To address this issue in this thesis, a longer segment of the mtdna control region has been sequenced and this has increased the resolution of the genetic marker and thereby the power of the MSA in some, but not all cases. In SE Asia, the use of the shorter sequence resulted in unreliable estimates for stock contributions among three neighbouring rookeries (Chapter 4). After sequencing the longer fragment, common haplotypes were resolved into new haplotypes at both the foraging ground and the three rookeries, thus providing more robust estimates of stock contributions (Chapter 4). The Bayesian based MSA allows for the incorporation of prior information in to the analysis. Ideally, the genetic data would be sufficiently powerful that prior information, such as the size of the source population, would make little difference, but this is rarely the case for marine turtle studies. Therefore, knowledge from field studies is essential for producing meaningful results. For example, in identifying source populations it often remains unknown to what extent a lack of genetic structure among rookeries results from contemporary gene flow versus recent shared ancestry. Extensive field studies can provide insights, such as in the GBR, where tagging data indicate limited movement between nesting beaches up to 500 kilometres apart (Limpus 2007). Similarly, mark-recapture studies show that although GBR turtles may use foraging grounds >2000 km distant, most are foraging within 500 km of their breeding grounds (Limpus 2007). Such information can be incorporated as appropriate priors in to the MSA. In most cases such weighted priors result in a narrower confidence interval (Naro Maciel et al. 2007, Bass et al. 2004, Okuyama & Bolker 2005). In cases where haplotypes are shared, using a Bayesian approach with weighted priors also helps. Using priors allows the researcher to incorporate knowledge of the species that provides for a stronger and more biologically meaningful result. 30
 and weighted priors (WP; relative to the size of the rookery).")
. Under the ideal scenario (Figure 2.")

41 Figure 2.4. Simulation of a hypothesised foraging aggregation where a) all rookeries are completely differentiated and b) rookeries share common haplotypes. The MSA was run using BAYES with uniform priors (UP) and weighted priors (WP; relative to the size of the rookery). Dotted line marks the true value of the composition from rookeries R1 (black) = 50%, R2 (grey) = 35%, R3 (white) = 15% and R4 (striped) = 0% This can be illustrated in a simple simulation of two hypothesised foraging aggregations (Figure 2.4). Under the ideal scenario (Figure 2.4a) each contributing stock has no genetic variation and each has a unique haplotype. The MSA for this scenario provides point estimates that are 100% correct for both large and small contributions, and the use of priors does not affect the outcome. In a more realistic marine turtle scenario (Figure 2.4b), where source populations share haplotypes, the MSA tends to underestimate large contributions and 31
42 overestimate small contributions. Using a weighted prior of source population size slightly improves the estimate. This imprecision is expected to increase as the number of source populations and the number of shared haplotypes increased. These examples illustrate the importance of being critical when drawing conclusions from MSA. Ideally, the interpretation of MSA results should use an integrated approach considering demographic, mark-recapture and satellite tracking data if these are available, in order to draw conclusions that are biological meaningful. 2.5 Conservation and Management MSA plays a crucial role in identifying the origins of threatened sea turtle foraging populations. For example, MSA has shown that 50% of loggerhead turtles caught in Mediterranean fisheries originate from rookeries in the South eastern United States (Laurent et al. 1998). Thousands of loggerhead turtles caught in oceanic fisheries in the North Pacific (Wetherall et al. 1993) are derived from nesting beaches in Japan but have their developmental habitat off the coast of Baja California, Mexico (Bowen et al. 1995). In the Caribbean, foraging aggregations are composed of turtles from throughout the Caribbean for green turtles (Luke et al. 2004, Bjorndal & Bolten 2008), loggerhead turtles (Engstrom et al. 2002) and hawksbill turtles (Bass 1999, Velez-Zuazo et al. 2008, Browne et al. 2009) many of which cross international borders. The 1982 United Nations (UN) Convention on the Law of the High Seas states that nations hosting the developmental habitats for migratory marine species hold exclusive fishing rights for these animals (Van dyke 1993). In addition, according to the 1983 UN Convention for the Conservation of Migratory Species (Bonn Convention), it is prohibited to take endangered species during migration on the high seas (Hykle 1992). This means that nations that have either nesting and/or developmental habitats for marine turtles have some level of jurisdiction over these animals on geographically remote foraging areas, even if these are located within the territorial boundaries of another nation. Thus, MSA studies should be designed to provide the foundation for international agreements concerning the protection of marine turtles. 32
43 Mixed Stock Analysis also holds great potential for monitoring population trends at oceanic and coastal foraging grounds for all size classes. Comparisons of adults and juveniles provide an opportunity to pick up early signs of shifts in the contributions of populations that may indicate population decline (or increase) (e.g., Chapter 5). This would provide a very powerful management tool for monitoring population trends at the foraging grounds by allowing time (years or decades) to put effective management actions in place long before the impact would be detected on the nesting beach. However, if estimates are inconsistent and surrounded by large confidence intervals, it will be hard to detect trends on a temporal scale, because estimates cannot be convincingly compared. Changes to foraging grounds as a result of climate change that may affect marine turtle populations have become an important concern (Fuentes et al. 2009, Hamann et al. 2007). Not only might the carrying capacity of foraging grounds be altered, but the currents that transport juveniles to those foraging grounds may shift (Fuentes et al. 2009). Also the sexratio of some turtle populations are expected to be affected by future climate change (Fuentes et al Hamann et al. 2007). In this case, MSA could provide an effective means of identifying both male and female-producing rookeries. MSA and the use of mtdna has revolutionised our understanding of the dynamics of marine turtle foraging grounds and associated migratory behaviour. Previous work has been limited by the lack of resolution and strong conclusions are often hampered by weak estimates. Several studies indicate that MSA estimates are qualitative rather than precise and are most appropriate for general hypothesis testing (Bowen & Karl 2008). We now need to move towards providing estimates that are sufficiently reliable to monitor changes over time and to generate estimates that can be used in population modelling. Future global coverage of rookeries and foraging grounds will help improve estimates and minimise the violation of assumptions of the models (Pella and Milner 1987). The combination of field studies to determine the demography of foraging ground populations, mark recapture studies at both foraging grounds and nesting beaches, satellite tracking (Limpus 2007) and multiple genetic markers (e.g., SNPs and microsatellites) is likely to yield an especially rich body of corroborating data, which none of the techniques alone could provide. 33
44 CHAPTER 3 3. Influences of larger sample sizes and longer sequences for defining Management Units and estimating stock composition of green turtle foraging grounds 3.1 Introduction Understanding the dynamic nature of migratory species in marine environments requires an understanding of the relationships between breeding populations and foraging aggregations. This knowledge is vital if the species is of conservation concern and if threats to the populations occur at both foraging grounds and in breeding areas. Several studies have used molecular markers to elucidate these relationships and inform conservation efforts for marine mammals (e.g., Lukoschek et al. 2009), fish (e.g., Waples et al. 2008) and marine turtles (e.g., Blumenthal et al. 2009). The first step is to identify management units (MU) (Moritz 1994) and determine their boundaries. Identifying those same MUs at foraging grounds where multiple stocks mix is particularly challenging. In marine turtles, sequences of the mitochondrial DNA (mtdna) control region, or d-loop, have been widely used to identify population structure (Bowen et al. 1993, Dethmers et al. 2006) and characterise foraging grounds (e.g., Velez-Zuazo et al. 2006, Blumenthal et al. 2009). Using differences in mtdna haplotype frequencies among rookeries, the proportions of the different haplotypes can be used to quantify the relative contributions of different source populations at habitats where multiple stocks mix (e.g. foraging grounds or migratory corridors). This is done through Mixed Stock Analysis (MSA), which requires a representative and extensive baseline sampling of potential source populations, and large enough differences in the haplotype frequencies among potential sources. In some regions more than one country or jurisdiction shares management responsibilities for marine turtle stocks, and so the identification of MUs is vital. Recent increases in industrial development of coastal island and offshore habitats in Western Australia (WA) have highlighted the need to better understand the dynamics of marine turtle populations in these areas. One species of concern is the green turtle (Chelonia mydas) that 34
45 nests on mainland and island rookeries and uses foraging grounds throughout the region where there are seagrass beds and coral reefs. Relatively little is known about Western Australian green turtle populations in comparison to the intensively studied east coast populations of the Great Barrier Reef (e.g. Limpus & Reed 1985, Limpus et al. 1992, 2000, Limpus 1993, 2007, FitzSimmons et al. 1997, Chaloupka & Limpus 2001, Chaloupka 2002). The most important rookeries in the region include the Lacepede Islands, Barrow Island, Monte Bello Islands (~30 km north of Barrow I.), North West Cape and Browse Island (see Figure 3.2) (Prince, 1994). Additionally, there is scattered nesting along the coast as well as nesting on several small islands and reefs offshore. Some of these sites include the islets of Ashmore Reef, Cartier Island (70 km east of Ashmore Reef), Scott Reef (Sandy Islet) and the Cocos (Keeling) Islands (see Figure 3.2) (Limpus 2007). Previous genetic research has found that the North West Cape and Lacepede Island rookeries form a single genetic stock or Management Unit (MU; Moritz 1994) that presumably includes all mainland and island rookeries found along over 1000 km of coastline (FitzSimmons et al. 1997, Dethmers et al. 2006). This green turtle population is thus one of the largest in the world and the largest remaining in the eastern Indian Ocean (Limpus 2007) and is known as the North West Shelf stock (Dethmers 2006). It has a much larger geographic scope in comparison to other stocks nesting in Australia, such as the two divergent stocks found in the northern and southern Great Barrier Reef (Norman et al. 1994, FitzSimmons et al. 1997, Dethmers et al. 2006). This discrepancy is apparently due to a difference in the geographic distribution of nesting beaches between the two coastlines that has altered opportunities for gene flow among rookeries. In WA there is scattered nesting along the coastline and nearshore islands (Prince 1994, 2000, Limpus 2007), unlike the Great Barrier Reef where rookeries are concentrated in the southern and northern islands (Limpus 2007). If a somewhat relaxed natal philopatry directs females to natal regions rather than specific natal beaches (Norman 1996, FitzSimmons et al. 1997) then this same behaviour in WA appears to have resulted in a large panmictic population. In contrast, some of the island rookeries offshore of WA have been identified through genetic studies as separate populations (Dethmers et al. 2006). Surprisingly, the Scott Reef and Ashmore Reef rookeries were identified as separate populations, even though they are located only 240 km apart. For the turtle nesting here, the genetic differences observed between the Scott and Ashmore Reef rookeries suggests that many of these females may experience a 35
46 stronger natal philopatry, perhaps due to the need to target these isolated reef systems that are located off the edge of the continental shelf and surrounded by deep water. However, sample sizes from Scott and Ashmore reefs were small and no other nearby rookeries were sampled, thus there is a need to do a more detailed genetic analysis of green turtle populations throughout the region. This is needed not only to better understand the geographic extent of the different stocks, but to establish baseline data for determining the composition of turtles at foraging grounds in the region. Very little is known about the foraging aggregations of green turtles in the waters off WA and the potential for increased pressures on these populations from development makes this knowledge crucial for management. There have been no long-term studies of marine turtles on foraging grounds, other than recent collaborative studies that have provided the samples for this and a previous genetic survey of the Ashmore Reef foraging ground (Moritz et al. 2003, Dethmers et al. in prep). This showed that the foraging aggregation at Ashmore Reef was composed by turtles not from the large nesting population at Ashmore itself, but from neighbouring Scott Reef, the North West Shelf and from the Gulf of Carpentaria (Moritz et al. 2003, Dethmers et al. in prep), but estimates were affected by large confidence intervals. The same was the case for several foraging grounds at Fog Bay and the Cobourg Peninsula in the Northern Territory and at Aru (Dethmers et al. in prep). Unfortunately, Mixed Stock Analysis in these studies is hampered by the preponderance of two common and widespread haplotypes. In the SE Asia, Timor and Arafura Sea and East Indian Ocean many of the populations share two very common mtdna haplotypes (called C1 and C3) that make it difficult to accurately assign turtles at foraging grounds to their breeding stock. One solution is to increase the number of markers used, but for marine turtles the maternally-inherited mtdna sequences provides stronger genetic structure than nuclear markers (FitzSimmons et al. 1997, Karl et al. 1992). Another option is to increase the length of the DNA segment being sequenced to add resolution to sequences shared between rookeries. New mtdna primers developed for marine turtles have increased the length of the d-loop sequenced to about 800bp (Abreu-Grobois et al. 2006). For hawksbill turtles in the Caribbean the longer sequence identified a number of new variants in different populations for the most common and widely shared haplotypes (Abreu-Grobois et al. 2006). It remains to be seen if this is also the case for other species, including green turtles, and if so, whether this added variation improves MSA. This will depend on how the new variation is distributed amongst the populations. 36
47 In this study the aims are (i) to reanalyse green turtle stocks in the eastern Indian Ocean and South East Asia that share the common haplotypes C1 and C3 using a longer sequence to see if this adds resolution to the differentiation among rookeries, (ii) to sample key rookeries of green turtles in WA not yet characterized (Barrow Island, Browse Island, the Cocos (Keeling) Islands and Cobourg Peninsula) to determine which stocks, if any, they group with and (iii) to sample the foraging populations at Shark Bay and Cocos (Keeling) Islands and estimate the contributions of different breeding stocks that use these foraging areas and (iv) to gain a better understanding of the migratory dynamics of these populations. 3.2 Materials and Methods Sample collection A total of 15 green turtle rookeries and two foraging aggregations within the eastern Indian Ocean and South East Asia were investigated in this study (Figure 3.2). Previously, 11 of the rookeries have been characterised using a relatively short (384 bp) segment of the mtdna control region (Dethmers et al. 2006). These samples were reanalysed using a different set of primers to amplify approximately 780 bp of the mtdna control region. In addition, the sample size has been increased for a number of those rookeries where samples were available. Overall, sample sizes across the 15 rookeries varied from 11 to 62 (Table 3.1). Four new Australian rookeries were added to the analysis (Figure 3.2). This included the small nesting population at Cocos (Keeling) Islands (n = 19), which represents the western extreme of Australian green turtle rookeries. The Cobourg Peninsula rookery was sampled, and although the sample size was small (n = 11) it filled a gap in the sampling of mainland nesting sites for green turtles along the north coast of Australia. Two islands off the coast of Western Australia were sampled; Browse Island (n = 31), which is located between Scott Reef and mainland Western Australia and Barrow Island, located north of North West Cape (n = 38). To investigate stock composition at foraging grounds, samples from two foraging aggregations were collected from Shark Bay (n = 71) and in the reefs around Cocos (Keeling) Islands (n = 71). The Shark Bay samples were collected in 2002 and the Cocos (Keeling) reef samples in 2004, 2005 and
48 Characterisation of mtdna haplotypes For all new samples, DNA was extracted from tissue samples using a salting-out procedure as described in FitzSimmons (1997). Previously analysed samples were extracted using a variety of extraction methods (chelex, phenol-chloroform and salting out). Extractions were checked for DNA quality and quantity by running 4 µl of DNA through a 1.2% agarose gel and visualizing with CyberSafe stain under UV light. Samples were PCR amplified using the primers LTEi9 (5 GAATAATCAAAAGAGAAGG 3 ) and H950 (5 GTCTCGGATTTAGGGGTTT 3 ) (Abreu-Grobios et al. 2006). These primers amplify a longer (~800 bp) fragment of the mtdna control region that overlaps the 386 bp used by Dethmers et al. (2006) and thereby have the potential to provide additional resolution (Figure 3.1). PCR was set up in a 25 μl reaction containing, 1x reaction buffer, 0.25 mm of each dntp, 1.5 mm MgCl 2, 10 μm of each primer, 1.25 U of Taq polymerase and ~40 ng of template DNA. The PCR was carried out on an Eppendorf Mastercycler ep thermocycler using an initial denaturing step at 94 C for 5 min, followed by 35 cycles of: 45s at 94 C (denaturing), 45s at 52 C (annealing) and 45s at 72 C (extension) and a final extension step of 5min at 72 C. PCR products were analysed for quality and quantity on agarose gels (as above) and all successfully amplified samples were purified prior to sequencing. Sequencing was conducted by Macrogen, Inc (Korea) for both the forward and reverse sequence and the results compiled using the software Geneious Pro (Version 4.7.6). Sequences were aligned using Clustal W (Thompson et al. 1994) implemented within Geneious. The haplotypes were matched against published haplotypes for short and long sequences and unpublished sequences from Australasia. All previously observed sequences were identified by the original name for published short sequences and given a new coding identification for Indo-Pacific green turtles that has the prefix CMIP (Table 3.1). New haplotypes were temporarily designated as New-1, etc, for the short sequence, and given a CMIP identification (e.g. CMIP-01, CMIP-02 etc.) for the longer segment. From these sequences two sets of data were created. In the first all sequences were cropped to include only the short (384 bp) sequence used in previous studies (e.g. Dethmers et al. 2006). The second dataset included the entire 780 bp sequence derived from using the new primers. All the analysis was performed on each data-set to evaluate the difference between the short and long sequences. 38
49 Control Region d-loop bp trna-thr trna-pro trna-phe LTEi9 TCR5 380 bp TCR6 H bp Figure 3.1. Schematic of the mtdna control region in marine turtles and the location of the short sequence used in Dethmers et al. (2006) (primers TCR5 and TCR6) and the long sequence used in this study (primers LTEi9 and H950) Molecular analysis Genetic diversity was measured using haplotype (h) and nucleotide (π) diversities (π; Nei 1987, p. 257) as estimated in ARLEQUIN v (Schneider et al. 2000). Haplotype diversity takes into account the number of haplotypes in a population and their frequency. Nucleotide diversity additionally considers the mean number of nucleotide differences (mutations) between all pairs of haplotypes in the population. Thus higher levels of nucleotide diversity indicate that the haplotypes in a population are more divergent, perhaps for example, due to colonization from different source populations. Estimates for both measures of diversity vary between 0 and 1. Genetic differentiation among sampling locations was tested using three different measures of population differentiation; a conventional F ST test based on haplotype frequencies (Slatkin 1995), a sequence-based F ST test using the Kimuras 2P model and the exact test of population differentiation (Raymond & Rousset). All tests were performed using ARLEQUIN v. 3.11, which provide the P-value and an estimate of the standard error (SE) of the P-value. Pairwise estimates of migration rates (Nm, (Slatkin 1995)) were also calculated using ARLEQUIN v To determine how the molecular variation was partitioned within the populations, an analysis of molecular variance (AMOVA) was performed for both nesting and foraging sites using Arlequin v The pair-wise F ST values (based on haplotype frequencies) obtained from ARLEQUIN were used to construct an UPGMA tree by neighbour joining procedure in PHYLIP 3.68 to display the genetic relationship between rookeries based on the haplotype frequencies. 39
50 Haplotype networks based on statistical parsimony were constructed using the software TCS 1.21 (Clement et al. 2000). Ambiguous connections in the network were resolved using a set of specific criteria. Under neutrality, specific empirical predictions can be made about the most likely connection among haplotypes. Under coalescent theory we expect that the haplotypes with the greatest frequency are older (Crandall & Tempelton 1993). Under that assumption, the expected rank of the haplotype by age is equal to the rank of haplotypes by frequency (Donnelly & Tavare 1996). Furthermore, haplotypes found in high frequency that have been present in the population for a long time are more likely to have originated new haplotypes. This means that frequent haplotypes are typically central in the network whereas rare haplotypes are likely to occur at the tips of the network. It also implies that frequent haplotypes have a greater number of mutational connections (Golding 1987; Excoffier & Langaney 1989). Furthermore it is expected that new haplotypes are more likely to be connected to haplotypes from the same population than to haplotypes from different populations (Watterson 1985; Takahata 1988). Under these assumptions three criteria can be set up. The first is a frequency criterion in which haplotypes are more likely to be connected to high frequency haplotypes. Second is a topological criterion in which haplotypes are more likely to be connected to interior haplotypes than to tip haplotypes. Finally, there is a geographical criterion in which haplotypes are more likely to be connected to haplotypes from the same or neighbouring population than to haplotypes found in distant populations (see Pfenninger & Posada 2002). The program RAREFACT.FOR ( was used to generate intra-sample rarefaction curves. Rarefaction curves provide a way to calculate the number of haplotypes, on average, that would have been expected with different sample sizes. It thereby provides a rough assessment of the sampling intensity needed. If the curve becomes parallel to the x-axis, the observed haplotype frequency should approximate the true level of richness. Otherwise, larger sample sizes can be expected to expose additional variation. 40


51 41 Figure 3.2. Sampling locations from nesting sites in eastern Indian Ocean and Southeast Asia with the UPGMA tree of the genetic relationship between rookeries. Shaded area on both the tree and the map marks the rookeries that are grouped into separate Managements Units. Square represents the two foraging aggregations.
52 Mixed Stock Analysis The contribution of Australasian rookeries to the two foraging aggregations at Shark Bay and Cocos Keeling were estimated under two scenarios; 1) using haplotype frequencies from the short sequence only and 2) using haplotype frequencies from the long sequence. Estimates were performed with a Bayesian approach using the software program Bayes (Pella & Masuda 2001). The Bayesian approach allows for the incorporation of prior knowledge about the source populations (breeding stocks), such as population size. Markov Chain Monte Carlo (MCMC) runs of 50,000 iterations were used for each of the 11 source populations. Each chain was started with a prior expectation of 0.95 from a specific source population and equal (0.05) contribution from the remaining populations. The shrink factor of Gelman & Rubin (1992) was used to assess if the 11 chains had converged. The mixed stock composition was estimated from the mean outputs of chains after 25,000 burn-in steps. To determine the relative contributions of Australasian MUs to the foraging aggregation two models were used. The first used priors based on the relative size of the individual nesting populations as a percentage of the total size of Australasian nesting locations using data included in Dethmers et al. (2006) (Model: Bay1). The second model used uniform priors assuming that each individual rookery has an equal chance of contributing to the foraging aggregation (Model: Bay2). 3.3 Results A total of 15 rookeries and two foraging grounds were analysed in this study. In the following results and discussion section, rookeries and foraging aggregations are treated separately. All data are presented for both the short 384 bp sequence and the long 780 bp sequence of the mtdna control region Rookery diversity Sequencing of the mtdna from 488 nesting turtles revealed 39 haplotypes from the long sequence and 29 for the short sequence (Table 3.1, Appendix 1) in comparison to the 14 haplotypes found previously from these rookeries (Dethmers et al. 2006). In considering the short haplotypes, C3 was the most common and was found in all populations except for Aru and Cobourg Peninsula. There were 13 new short haplotypes and 14 new long haplotypes observed. Five of the long sequences (or four short sequences) were found at Barrow Island and three new haplotypes (long and short) were found at Peninsular Malaysia. Two new 42
53 haplotypes (long and short) were found from the Lacepedes and North West Cape, and a single new haplotype was found at each of Berau Islands, Cobourg Peninsula and West Java. When using the longer sequence, the three most frequent haplotypes C3 (34%), C1 (23%) and D2 (13%) split into five, five and two new haplotypes, respectively. In addition, one rare haplotype New-1 (0.4%) split into two new haplotypes (Table 3.1). A pattern was apparent in which the composition of haplotypes shifted in relation to latitude (Table 3.1). Stocks at latitudes north of -10 S were composed of haplotypes C3, New 10-12, C4, C5, B5, and D2, while stocks south of latitude -10 S were characterized by A2, C1, C2, C3, C9, C19, C25, D5, IND3, New-1-4, New-8 and New-13 (Table 3.1). The divergent A and B haplotypes that characterise the Great Barrier Reef rookeries were not common (< 10%) at any rookery, except for Ashmore Reef, and were only present at 10% in the total sample. B5 was found only from the Sarawak and Peninsula Malaysia stock, A2 was found at low frequency at Ashmore Reef, Scott Reef, Browse Island and Barrow Island, and A3 was found at low frequency at Aru and Scott Reef, but at high frequency (44%) at Ashmore Reefs. Five haplotypes (short sequence) were found for the first time among eastern Australian and South East Asian rookeries that had been described previously from other rookeries. One haplotype (C7) found at Ashmore Reef had been described from a rookery in Papua New Guinea (Dethmers et al. 2006), three haplotypes had previously been observed at Australian foraging grounds (C19 from Field Island, C25 from Ashmore Reef and Field Island and D5 from Cobourg Peninsula; Moritz et al. 2002, Dethmers et al. in prep). These new haplotypes were found at Browse Island (C19) and Barrow Island (C25 and D5). One short haplotype (Ind3) had been previously described from western Indian Ocean rookeries (Formia et al. 2006), but was found for the first time in Australia at Barrow Island. Genetic diversity varied widely among the sampled rookeries, with only two haplotypes (both short and long) found at Aru and Cocos Keeling and up to 11 (short) or 14 (long) haplotypes at Barrow Island (Table 3.1). Haplotype diversity, which considers the frequency of haplotypes varied from 0.07 to 0.76 (mean = 0.48) for the short and 0.07 to 0.83 (mean = 0.54) for the long sequences (Table 3.2). Estimates of nucleotide diversity were low for both the short ( , mean = 0.006) and long ( , mean = 0.006) sequences (Table 3.2). The haplotype network identified C1 (short) and CMIP-04 (long) as being the central haplotypes for the majority of sequences, which reflected the similarity between most of the haplotypes, with the exception of the divergent A and B haplotypes (Figure 3.3). 43
54 Population differentiation Overall, there was significant differentiation (F ST based on haplotype frequencies) among rookeries in all but 9 of 105 pairwise tests for the short and 7 of 105 pairwise tests for the long sequence (P > 0.05). Each pairwise comparison was considered as a discrete hypothesis and thus do not reduce α levels for multiple tests (Table 3.4 and 3.5). Non-significant F ST values for the short sequences grouped Sulu Sea and Sipadan (F ST = 0.046), Peninsula Malaysia and Scott Reef (F ST = 0.032), and Ashmore Reef and Browse Island (F ST = 0.011) in addition to grouping all of Cobourg Peninsula, Lacepedes, Barrow Island and the North West Shelf (P > 0.05) (Table 3.4). When using the longer sequence only Peninsular Malaysia and Scott Reef (F ST = 0.032) grouped in addition to Scott Reef and Browse Island (F ST = 0.007) and all comparisons between Cobourg Peninsula, Lacepedes, Barrow Island and the North West Shelf (P > 0.05). However, given the geographic location of Scott Reef and Peninsular Malaysia, the low F ST value (F ST = 0.032), and significant differentiation shown by the exact tests (Table 3.5) they are kept as separate stocks for further analysis and for the MSA. Although Cobourg Peninsula, did group with the Lacepedes, Barrow Island and North West Cape the sample size for this rookery was low (n = 11) and four of the 11 haplotypes were not found in any of the other three rookeries. In addition it is located more than 2000 kilometres from the nearest of the rookeries in the Lacepedes. The exact tests only grouped Cobourg Peninsula with Barrow Island but not the other two rookeries. For these reasons I consider Cobourg Peninsula to be a separate population until this can be clarified with a larger sample size. The remaining three rookeries at North West Cape, Barrow Island and the Lacepedes were grouped into one MU (North West Shelf) for further analysis (Tables 3.4 and 3.5 and Figure 3.2). After grouping of the Scott/Browse and NWS MUs all pairwise F ST tests were significant (P < 0.05) except for Cobourg Peninsula and NWS (P = 0.36). The exact test gave mostly similar results for the short and long sequences and both suggested non-differentiation between Peninsular Malaysia vs. Cocos (Keeling) and Cocos (Keeling) vs. Scott/Browse MU (Table 3.5). However, given the geographic locations of these rookeries (Figure 3.2), and their differentiation from the West Java rookery this result is likely to be caused by low sample size at Cocos (Keeling) (n = 19), Results from the AMOVA indicated that significant genetic differentiation occurred between latitudinal regions as designated by the Indian Ocean stocks below latitude -10 and Southeast Asia above latitude -10, although this only accounted for 6.5% of the genetic variance when using the long sequences (Table 3.3). Estimates of genetic variance indicated that 32.3% (long 44
 was partitioned within the 11 stocks.")


. a b Figure 3.3. Haplotype network based on maximum parsimony for the short a) and the long b) sequence.")
55 sequences) of variance was distributed among the stocks within these regions and 61.2% of variance (long sequences) was partitioned within the 11 stocks. The estimated variance among regions and among stocks within regions was slightly less for the long sequences, but somewhat greater for the variance within stocks. The rarefaction analysis indicated that most stocks had been adequately sampled as most curves had begun to level off upon reaching the sample size used. The curves of Peninsular Malaysia (n = 29), Aru (n = 28) and the large North West Shelf (n = 114) however indicate that a larger samples size is needed (Figure. 3.4). a b Figure 3.3. Haplotype network based on maximum parsimony for the short a) and the long b) sequence. The most unlikely solutions for ambiguous connections are indicated by the dotted lines. The match between short and long haplotype names are defined in Table 3.1. Black dots indicate haplotype states that are necessary intermediates but were not present in the sample. Each line represents a single mutational step connecting two haplotypes. 45
56 46 South of Lat. -10 North of Lat -10 Long CMIP-01 CMIP-19 CMIP-77 CMIP-78 CMIP-119 CMIP-134 CMIP-69 CMIP-43 CMIP-57 CMIP-26 CMIP-133 CMIP-68 CMIP-66 CMIP-70 CMIP-130 CMIP-131 CMIP-02 CMIP-03 CMIP-05 CMIP-04 CMIP-12 CMIP-16 CMIP-30 CMIP-31 CMIP-10 CMIP-18 CMIP-135 CMIP-120 CMIP-121 CMIP-14 CMIP-13 CMIP-06 CMIP-07 CMIP-17 CMIP-08 CMIP-09 CMIP-11 CMIP-122 CMIP-123 Short C3 New-12 C5 D2 C14 New-11 C4 B5 New-5 New-9 New-10 A3 C7 A2 C1 C9 C19 New-13 New-6 New-7 C25 D5 IND3 New-1 New-2 New-3 New-4 New-8 C2 Table 3.1. Distribution of C. mydas haplotypes sampled at 15 rookeries in the eastern Indian Ocean and Southeast Asia. Short refers to the 384 bp sequence used in previous studies and follows the nomenclature presented in Dethmers et al. (2006), except for new haplotypes. Long refers to the 780 bp fragment used in this study. Shaded areas group together haplotypes that are the same for the short sequence but split into new haplotypes for the long sequence. Note that Scott Reef and Browse Island group into Scott/Browse MU (SR/BroI) and Lacepedes, Barrow Island and North West Cape group into the North West Shelf MU (NWS). Total sample sizes in bold indicate either new sample locations or larger sample sizes in comparison to Dethmers et al. (2006). n WJ BerI SS Sip Sar PM Aru CP CK AR SR/BroI MU SR BroI Total SR/Br NWS MU Lac BaI NWC Total NWS Total all WJ, West Java; BerI, Berau Island; SS, Sulu Sea; Sip, Sipadan; Sar, Sarawak; PM, Peninsular Malaysia; Aru, Aru; CP, Cobourg Peninsula; CK, Cocos (Keeling), Ashmore Reef; SR, Scott Reef; BroI, Browse Island; Lac, Lacepedes; BaI, Barrow Island; NWC, North West Cape and NWS, North West Shelf. 46
57 Table 3.2. Diversity estimates for 17 green turtle MUs and rookeries, including sample size (n), number of haplotypes (H) haplotype diversity (h) and nucleotide diversity (π). MU or Rookery N H h Π West Java MU Long Short Berau Island MU Long Short Sulu Sea MU Long Short Sipadan MU Long Short Sarawak MU Long Short Peninsula Malaysia MU Long Short Aru MU Long Short Cobourg Rookery Long Short Cocos MU Long Short Ashmore Reef MU Long Short Scott Reef Rookery Long Short Browse Island Rookery Long Short Scott/Browse MU Long Short Lacepedes Rookery Long Short Barrow Island Rookery Long Short North west Cape Rookery Long Short North west Shelf MU Long Short Short Mean Long Table 3.3. Analysis of molecular variance (AMOVA) results for 11 green turtle east Indian Ocean Management Units and the Coburg Island rookery. The significance of permutation test (10,000 permutations) are shown for P < 0.05 (*) and P < (***). Regions are divided by the 10 S line of latitude, with the Indian Ocean stocks to the south and the Southeast Asian stocks to the north. % of variance F statistics Among regions Long 4.20 F CT = Short 8.72 F CT = 0.087* Among stocks within Long F SC = 0.333*** regions Short F SC = 0.392*** Within stocks Long F ST = 0.361*** Short F ST = 0.444*** 47
58 48 Table 3.4. Genetic differentiation (F ST ) among MUs and rookeries based on haplotype frequencies using the long sequence (below diagonal) and estimates of the number of migrants per generation (Nm; above diagonal). The significance of permutation test (10,000 permutations) are shown for P > 0.05 (n.s.), P < 0.05 (*) and P < (**). Non-significant values are presented in bold. WJ BerI SS Sip Sar PM Aru CP CK AR SR/BroI NWS WJ MU / BerI MU SS MU Sip MU Sar MU PM MU Aru MU CP rookery CK MU AR MU SR/BroI MU NWS MU * *** * *** *** *** *** *** *** *** *** / *** *** *** *** *** *** *** *** *** *** / *** *** *** *** *** *** *** *** *** / *** * *** *** ** *** ** *** / *** *** *** *** *** *** *** / *** *** * *** * *** / *** *** *** *** *** / *** *** *** n.s / *** * *** / *** *** / *** 1.3 / 48
59 49 Table 3.5. Estimated P values from exact test of population differentiation among east Indian Ocean MUs and the Coburg Peninsula rookeries based on haplotype frequencies using the short sequence (above diagonal) and the long sequence (below diagonal). The significance of permutation tests (10,000 permutations) are shown for P > 0.05 (n.s.), P < 0.05 (*) and P < (**). P values <0.000 are represented by ***. Non-significant P values are presented in bold. WJ BerI SS Sip Sar PM Aru CP CK AR SR/BroI NWS WJ MU / *** *** *** *** *** *** *** *** * ** ** BerI MU / *** *** *** *** *** *** *** *** *** * n.s. SS MU *** *** / *** *** *** *** *** *** *** *** ** Sip MU *** *** *** / *** *** *** *** *** *** *** *** Sar MU *** *** *** *** / *** *** *** PM MU *** *** *** *** *** / *** *** ** 0.15 n.s. *** *** *** *** *** *** Aru MU *** *** *** *** *** *** / *** *** *** *** *** CP rookery *** *** *** *** *** *** *** / *** *** *** CK MU *** *** *** *** *** n.s. AR MU *** SR/BroI MU *** *** *** *** *** *** *** *** *** *** / *** *** *** *** *** *** *** *** *** / *** *** NWS MU *** *** *** *** *** *** *** n.s n.s n.s * *** *** / *** *** *** *** / 49

60 Figure 3.4. Rarefraction curves for each Mus. X axis shows the sample size and Y axis the number of haplotypes. The graph shows the mean and the SD. 50
61 Long CMIP-04 CMIP-30 CMIP-15 CMIP-73 CMIP-75 CMIP-01 CMIP-77 CMIP-78 CMIP-85 CMIP-79 CMIP-76 CMIP-14 CMIP-24 CMIP-25 CMIP-05 CMIP-74 CMIP-80 CMIP-81 CMIP-82 CMIP-83 CMIP-84 Short C1 C3 C9 C21 C25 B4 B7 A2 New-14 New-15 New-16 New-17 New-18 New-19 Table 3.6 Distribution of C. mydas haplotypes sampled at two foraging grounds, Shark Bay and Cocos (Keeling) Islands. Short refers to the 384 bp sequence used in previous studies and follow the nomenclature presented in Dethmers et al. (2006), except for new haplotypes. Long refers to the 780 bp fragment used in this study. Shaded areas group together haplotypes that are the same for the short sequence but split into new haplotypes for the long sequence. Shark Bay Cocos Keeling Foraging aggregations The two foraging aggregations at Shark Bay and Cocos Keeling were comprised of significantly different haplotype frequencies (χ2 = 65.5, P = 0.001) as would be expected for distant foraging populations. Shark Bay consisted of a mixed sample of 71 green turtles in total (33 adult turtles; 12 males and 21 females sampled in 2002 and 38 juvenile turtles also sampled in 2002). These samples comprised 12 haplotypes for the short and 19 haplotypes for the long sequence. The sample was dominated by the short C1 (65%) and C3 (18%) haplotypes and the long CMIP-04 (C1; 59%) and CMIP-01 (C3; 14%) haplotypes. Seven haplotypes were orphan haplotypes that have not been recorded in any breeding stock to date. Six of these were new (short New and long CMIP-74, CMIP-80 and CMIP-81-84) and one (C21) has been previously recorded in foraging turtles at Cobourg Peninsula (Dethmers at el in prep). One individual had a haplotype (long CMIP-25, short B7) that was only found in nesting turtles from the northern Great Barrier Reef stock (Table 3.6 and Chapter 5). There were no significant differences in haplotype frequencies between adult and juvenile turtles for the short (χ2 = 5.83, P = 0.35), or the long (χ2 = 12.04, P = 0.76) sequence and they were pooled for the MSA. The Cocos (Keeling) sample consisted of 71 juvenile green turtles sampled across several years (2004, 2005 and 2009). In contrast to the Shark Bay sample, only four haplotypes were 51
62 identified for both the short and the long sequence. C3 and CMIP-01 (76%) were the most common haplotypes followed by C1 and CMIP-04 (18%). One individual had a haplotype found only outside the eastern Indian Ocean (short B4 and long CMIP-24) in nesting turtles from the northern Great Barrier Reef (Chapter 5) MSA The mixed stock analysis for Shark Bay indicated that this foraging aggregation is composed mainly of turtles from the large North West Shelf MU to the north (mean contribution = , 95% CI = ) using four different models. Overall the analysis gave similar results for the long and the short sequences or when using the two Bayesian models with either rookery size as a prior (Bay1) or when using uniform priors (Bay2) (Table 3.7). The mixed stock analysis for the Cocos (Keeling) foraging turtles was less clear and results were confounded by large confidence intervals. The main contribution was estimated to be coming from the newly sampled Cocos (Keeling) rookery but results varied between the short and the long sequences and between the models (short, Bay1 = 0.79; Bay2 = 0.84 and long, Bay1 = 0.71; Bay2 = 0.57). Estimates for Cocos (Keeling) were surrounded by large CI ranging from 0-1 when using the long sequences, but markedly smaller when using the short sequences (95% CI = ). All models gave similar contribution to the North West Shelf MU (Bay1 = and Bay2 = ). Finally, the MSA using the long sequences assigned a small contribution (Bay1 = 0.12, Bay2 = 0.36) to the Scott Reef/Browse Island MU while MSA using the short sequences estimated a very small (Bay1 = 0.0 and Bay2 = 0.02) contribution from this MU. Again the estimate for the long sequence was surrounded by CI between 0-1. Despite these large confidence intervals all analysis had converged as shown by a Gelman-Rubin shrink factor of less than
63 Table 3.7. Contribution of regional stocks to two foraging aggregations at Shark Bay and Cocos (Keeling) using the long (L) and short (S) sequence. Results are shown for two different models, Bay1 using rookery size as prior information and Bay2 using uniform priors; includes mean and lower 2.5 % and upper 97.5 % confidence intervals. Estimated contributions > 5% are shown in bold. Shark Bay Cocos Keeling Bay1 Bay2 Bay1 Bay2 Stoc k Mean Mean Mean Mean SS L S Sip L S BerI L S Sar L S PM L S Aru L S AR L S SR/ L BerI S NWS L S WJ L S CK L S
64 3.4 Discussion Rookery diversity and differentiation Among all samples analysed in this study 53 different haplotypes were identified for the long sequence and 38 for the short sequence. Three of the most common haplotypes split into new ones. However, in each case, there remained a one common haplotype that had the highest frequency in most populations, accompanied a few rare haplotypes that were often within the same population. It does not appear that this was the result of insufficient sampling at most locations, given that the rarefaction curves suggested that the sample sizes had captured most of the genetic diversity, except for the Peninsular Malaysia, Aru and the Northwest Shelf MUs. Thus the longer sequence added little resolution to the genetic profile of green turtle rookeries throughout the region. The largest effect was seen for Berau Island, Sulu Sea and West Java in South East Asia where haplotype C3 split into new haplotypes in three areas, providing further support for their designation as Management Units. While this adds only small changes to F ST values it may strengthen the precision of MSA in the South East Asian region. This will be explored in Chapter 4. Haplotype diversity increased in half of the populations when using the longer sequence. Among the previously sampled MUs, the mean value (h = 0.61) increased somewhat in comparison to the previous estimate (h = 0.49; Dethmers et al. 2006). The range in haplotype diversities observed are within that found in Southwest Indian Ocean populations, but with a higher mean value (range , mean = 0.37; Bourjea et al. 2006). Nucleotide diversity decreased somewhat in relation to previous estimates (Dethmers et al. 2006), which in part reflects how nucleotide diversity is partitioned along the new section (Abreu-Grobios et al. 2006). Values were similar to estimates from the Southwest Indian Ocean (range ; mean = 0.01; Bourjea et al. 2006). Haplotype diversity within the Northwest Shelf MU was elevated at Barrow Island and North West Cape rookeries in comparison to the more northerly rookery in the Lacepedes. Increased sample sizes and the use of longer sequences in general supported the overall population structure reported in Dethmers et al. (2006). In particular, the differentiation between the Scott Reef and Ashmore Reef populations is supported with the larger samples sizes (n = 35 and 55, respectively). The addition of 31 samples from Browse Island indicated 54
65 that this rookery is genetically differentiated from the Ashmore Reef rookery, ~240 km to the north, but not from Scott Reef rookery, which is ~190 km to the west. These results indicate that the Scott Reef and Browse Island rookeries function as a Management Unit that is different from the Ashmore Reef Management Unit. This interesting result suggests that genetic exchange among populations on islands scattered off the continental shelf is a dynamic process that is likely influenced not only by distance, but by geographic location. Given that Ashmore Reef is located north of Scott Reef and Browse Island, which are at similar latitudes, it may be that latitude, rather than longitude is a stronger cue for natal homing by breeding females in this region. This may be possible given the sensitivity to latitude observed in hatchlings during orientation studies that alter magnetic inclination and intensity (Lohman & Lohman 1994, 1996). It is thought that their ability to orient to magnetic fields (inclination and intensity) not only keeps them within the correct ocean currents during migrations, but that this also forms the basis of an internal map used to direct them back to natal regions as adults (Lohman & Lohman 1998). Variation in magnetic fields due to local geology may also influence navigation. Throughout the Browse and Scott Basins there is variation in the regional geology (Blevin et al. 1997) that is reflected by the presence of one of Australia s largest gas reserves in these basins. Whether these geologic variations are sufficient to act as navigation cues is unknown. There is some evidence that variation in current patterns between Ashmore Reef and Scott Reef/Browse Island at the time of hatchling dispersal has resulted in differential representation of these stocks on foraging grounds to the east (Dethmers et al. in prep), though that is not expected to influence the natal homing ability of adults from those foraging grounds In contrast, the lack of differentiation between the distant Lacepedes and North West Cape rookeries was also supported by increased sample sizes (38 and 25, respectively). While, there was some indication that the new Cobourg Peninsula rookery grouped with both Barrow Island and North West Cape this result was influenced by a small sample size (n = 11) and had four individuals with haplotypes that are not seen in any of the other NWS rookeries. These observations indicate that a larger sample size for this rookery might separate it from this MU. The addition of 38 samples from Barrow Island rookery, located ~50 km offshore, indicated that this rookery groups strongly with the North West Cape rookery, located ~180 km to the south, and somewhat less strongly with the Lacepedes Island rookery, located >800 km to the north and Cobourg Peninsula rookery. 55
66 The final proposal of an MU at the Cocos (Keeling) Islands suffers from a small sample size (n = 19). At present, tests of genetic differentiation suggest that this rookery significantly differentiated from other stocks included in this study Foraging aggregations and MSA The two foraging grounds analysed in this study included one new site at Shark Bay, and the foraging ground at Cocos (Keeling) Island that was previously characterized with a small sample size (n = 35; Dethmers et al. in prep). Shark Bay is the most southern large foraging ground for green turtles in WA and therefore only gets fed by rookeries from the north. It had a large number of haplotypes (n = 19) and showed an overlapping haplotype profile to that observed at Barrow Island and the North West Cape. The MSA results for Shark Bay were fairly robust and indicate that this foraging ground is used mainly (> 90%) by green turtles from the neighbouring North West Shelf MU. This is in agreement with several mark-recapture events of nesting turtles from the NWS MU that use foraging grounds in Shark Bay (Limpus 2004). The observation of a turtle with a northern Great Barrier Reef haplotype in Shark Bay is indicative of rare long distance dispersal of ngbr turtles to foraging aggregations in Western Australia, whether or not this indicates a contemporary or historical event. While such migration feats are rarely documented in green turtles, a small number of tag returns from females tagged while nesting in Western Australia have been recorded as far east as Mornington Island and western Cape York Peninsula in Queensland more than 2500 km away (Limpus 2004). The combination of tag-return data and the high similarity between results generated from the long and short sequence and the Bay1 and Bay2 models also adds confidence to the results. The results for Cocos (Keeling) were ambiguous and surrounded by large confidence intervals. The uncertainty can be seen in the histograms and the trace plots generated by Bayes (data not shown). The trace plots track all chains and should stabilize at the equilibrium posterior distribution. However, for the Scott Reef/Browse, Cocos (Keeling) and North West Shelf stocks they are highly variable and hence generating the large confidence intervals. Unfortunately, the use of the longer sequence has added little resolution to the problem of having a high frequency of shared haplotypes in the MUs (especially C1 and C3) and hence 56
67 this continues to generate large confidence intervals. The estimates derived from the MSA in this region must be carefully analysed and should only be used as indicative of general trends of green turtle migratory behaviour. Therefore, especially the MSA from the Cocos (Keeling) foraging ground should be interpreted with caution. Those results suggest that this foraging aggregation is occupied mainly by turtles from the Cocos (Keeling) stock, but also from the North West Shelf MU and possibly the Scott Reef/Browse Island MU. This would be a quite different dynamic than that observed for the nearest foraging ground that has been studied at Ashmore Reef. There, MSA results suggest that few turtles using that foraging ground originated from the Ashmore Reef rookery (Dethmers et al. in prep). Satellite telemetry studies of six adult females nesting at Cocos (Keeling) showed that all six returned to foraging grounds within the Cocos (Keeling) less than 50 km away (Whiting et al. 2007), supporting the short distance migration for most turtles suggested by the MSA results. In addition a tag-return was recorded from a female green turtle foraging at Cocos (Keeling) that has previously been recorded nesting at Ningaloo Reef not far from North West Cape (S. Whiting pers. comm.). Together, the limited tagging data and the MSA analysis corroborate a scenario where the majority of turtles foraging at Cocos (Keeling) are from the local breeding stock. The tag return from Ningaloo supports the estimate of a ~10% contribution from the North West Shelf Management implications There are two key points regarding management issues for marine turtles in which genetic studies provide unique insights. First, management of marine turtles needs to be based on identified breeding populations, the Management Units, because these reflect groupings of turtles that have existed since the recent past and among which limited genetic exchange occurs, atleast for the mtdna. Thus, there is little migration of individuals among the Management Units that would allow for compensation of mortality occurring at a particular MU. Second, management of marine turtles must take into account the entire geographic range encompassed by an MU, including the foraging grounds used by those turtles. For this, flipper tagging and satellite tagging also provide information on the geographic scope of foraging grounds used by particular rookeries, but unless these studies are done on a comprehensive scale, the insights are somewhat limited. Genetic studies have been shown to greatly enhance this information by providing estimates of the proportion of each stock represented at particular foraging grounds (Velez-Zuazo et al. 2008, Bjorndal & Bolten 2008, Blumenthal et al. 2009). In this way, mortality at foraging grounds can be proportionally 57
68 attributed to each MU. Continuation of the genetic sampling and analyses of foraging ground aggregations off the Western Australian coast is particularly important given the scarcity of mark-recapture studies at these foraging grounds. For the green turtle populations of Western Australia and offshore waters, we have identified four MUs. First, the Northwest Shelf MU is a mainland/nearshore stock that encompasses at least 1000 km of coastline and which is one of the world s largest remaining green turtle populations. Management of this stock must consider the cumulative impacts that may adversely affect this large population. Not only do cumulative impacts need to include the range of rookeries within this MU, but the analysis of foraging grounds indicates that turtles from this MU utilize foraging grounds from Shark Bay, to the Cocos (Keeling) Islands and across the Northern Territory to Torres Strait (Chapter 5). Thus, there is a need to also monitor a diversity of foraging grounds used by this MU to assess impacts and sources of mortality. For the Scott Reef/Browse Island and Ashmore Reef MUs, there is genetic divergence over much smaller geographic ranges. This indicates that impacts to rookeries in either of these regions will potentially be affecting an entire MU. Thus, although an adverse impact to the Scott Reef rookery may be somewhat buffered by nearby rookeries, if impacts also encompass Browse Island, or other smaller nearby rookeries, then the entire MU may be at risk. At present we do not know the complete geographic scope of either MU. Additional sampling of nearby rookeries, including Cartier Reef and Maret Island would provide a better indication of the geographic scope of the offshore MUs. The analysis of foraging grounds indicates that turtles from the Scott Reef/Browse Island MU use foraging grounds that range from Cocos (Keeling) Island to the west, to Field Island in the east (Dethmers et al. in prep), but there are large errors with the estimates. Further flipper tagging of turtles at these MUs and satellite tagging could help determine the geographic scope of foraging grounds used by these MUs. This information is important for successful implementation of management and conservations strategies for this remote region of Australia. 58
69 CHAPTER 4 4. Origin of immature green turtles (Chelonia mydas) at two foraging grounds in Sabah, Malaysia. 4.1 Introduction Green turtles, Chelonia mydas, are one of the most widely distributed species of marine turtle and although they are the most abundant large herbivore in many shallow marine habitats, various populations are at risk from a variety of threats including many that are unintentional. Throughout southeast Asia, large (>2000 nests per year) regional rookeries are found along the coasts of Indonesia, eastern Malaysia (Sabah and Sarawak Turtle Islands), Peninsular Malaysia and the Tawi-Tawi Turtle Islands, and the Philippines (Figure 4.1) (Shanker & Pilcher 2003, Pilcher 2007). Several smaller rookeries (~ nests/year) are scattered throughout the region (Pilcher 2007). Green turtles in SE Asia have long suffered severe impacts from fisheries, cultural harvests and a large and widespread harvest of eggs causing major declines in many populations (Limpus et al. 2001, Shanker & Pilcher 2003). While many of these populations remain depleted, some are showing signs of recovery as a result of long-term conservation efforts such as at Sabah, Malaysia. However, there are also other, less obvious, threats impacting these populations. For example, at many rookeries, in particular at the Turtle Islands complex and Sarawak, there is a long tradition of relocating nests to unshaded beach hatcheries for incubation (Tiwol & Cabanban 2000). Although protecting these nests is important, many of these relocated nests experience higher temperatures than they otherwise would, thereby unintentionally altering the sex of marine turtle hatchlings, as determined by Temperature-dependant Sex Determination (TSD). The critical period of sex determination occurs during the middle third of the incubation period, when higher nest temperatures produce females (Mrosovsky 1980, 1994). Despite several studies pointing out the importance of shading hatcheries to control nest temperatures (van de Merwe et al. 2005, 2006), unshaded hatcheries were used for several decades and today the hatcheries only receive partial shading (Basintal & Lakim 1993, Pilcher 2000). As a result, millions of female hatchlings, but only few males have been released from these regions (Tiwol & Cabanban 59
70 2000). Such a strong bias is likely to have profound impacts on the population trends in the coming years as the adult populations become increasingly female biased (Limpus et al. 2001). It has been suggested that concerns should be raised if sex ratios of hatchlings across all nesting beaches for a particular stock, approach 1M: 4F (Poloczanska et al. 2009). Therefore, information derived from monitoring demographic parameters of the nesting populations and hatchling sex ratios from rookeries throughout Southeast Asia is crucial for informing conservation efforts, as is the need to monitor turtle populations at the foraging grounds. Fortunately, most nesting beaches have been identified and monitoring or hatchery management programs are established at many rookeries. Considerably less is known about the foraging ground populations throughout this region. Studies of the demographics and dynamics of populations at various foraging grounds throughout the region are needed to allow identification of trends and potential threats to all age-classes of the population. Hormonal assays (Diez & Van Dam 2003) or laparoscopic examinations (Miller & Limpus 2003) can be used to determine gender in immature turtles and sex ratios across size classes and are thus important components of demographic studies. Several long-term foraging ground mark recapture studies have shown the value of providing detailed sex and age-class-specific demographic information (Chaloupka & Limpus 1997, Limpus & Chaloupka 1997, Chaloupka & Limpus 2001, Chaloupka & Limpus 2002, Pilcher 2010), however, trends in a population may not be detected for several decades. For example, the effect of predation and egg harvest on the nesting beach might not become apparent until several decades later, given the years taken by green turtles to mature (Chaloupka et al. 2004). Likewise, the effects of increased sand temperature on hatchling sex ratios, from either global warming or hatchery practices, will not be detected at the nesting beach until decades later. Such declines may be observed much earlier by monitoring different size classes at the foraging grounds. Therefore, the many beach monitoring programs, while extremely important and logistically easier to carry out than foraging studies, do not provide a complete picture of marine turtle population viability. In a recent foraging ground study in Malaysia, Pilcher (2010), found that the shallow waters surrounding Mantanani Islands (off Kinabalu, Borneo) served as a developmental habitat for immature green turtles, with turtles recruiting at around 38 cm in curved carapace length (CCL). He also noted that turtles larger than 62 cm CCL were generally absent from these areas, thus suggesting a residence period for immature turtles of six to seven years. Importantly, he also found that this foraging aggregation of 60
71 immature green turtles had a highly female-biased sex ratio of 1M: 4F, indicating a possible link with the biased sex ratios produced at many hatcheries in the region. If there are adult females at the foraging grounds, then opportunistic recapture of females tagged on nesting beaches can establish the connection between rookeries and foraging grounds by females undertaking their breeding migrations. However, such detection typically requires tagging thousands of adult turtles and considerable effort to re-capture adults at foraging grounds. This effort is reflected in long term mark-recapture studies that have revealed strong links between nesting and foraging aggregations such as in the Great Barrier Reef (Limpus 2007), Tortuguero, Costa Rica (Troëng et al. 2005b) and Hawaii (Balazs et al. 1987). More recently, large-scale satellite tagging projects of breeding female turtles have and are being used to establish the links between rookeries and the foraging grounds they use (Godley et al. 2002, Craig et al. 2004, van de Merwe et al. 2009). Notwithstanding the benefit of such information, understanding the links between turtle breeding populations and the foraging grounds they use is only the first step. What is needed for more effective species management is knowledge of the contribution of each regional breeding population to each foraging area they use. With this information it is possible to consider in more detail the variation in growth rates and sex-ratios at the different foraging grounds in order to model the dynamics of the population (Chaloupka 2004). Knowing the contribution of each regional population to each foraging ground allows for an assessment of the overall level of risk faced by a population resulting from threats at both nesting beaches and foraging grounds (Chapter 6, Reece et al. 2006). For example, linking populations that may be affected by various anthropogenic activities, such as fisheries or subsistence harvest, hundreds or even thousands of kilometres away from foraging grounds (Bowen et al. 1995, Laurent et al. 1998, Boyle et al. 2009) will allow for better species management outcomes. Such information on the origins of turtles at foraging grounds can be obtained through the use of genetic analyses. Mixed stock analysis (MSA) using mitochondrial DNA (mtdna) haplotype frequency differences among nesting populations provides the background information to link foraging aggregations back to their rookery of origin. This analysis employs either a maximum likelihood or a Bayesian algorithm to estimate the contribution of source populations resulting in the best fit to the observed haplotype frequencies from the mixed foraging aggregate (Pella & Masuda 2001, Okuyama & Bolker 2005). The use of genetic markers thereby provides a more powerful complement to traditional mark-recapture 61
 and the location of the two study areas, Mantanani Island and Layang Layang Islands. techniques that can be applied to all gender and size classes.")
72 Figure 4.1. Map showing the 17 genetic stocks used as reference for tracing back the origin of turtles (adapted from Dethmers et al. 2006) and the location of the two study areas, Mantanani Island and Layang Layang Islands. techniques that can be applied to all gender and size classes. The number of studies using MSA has increased in the last decade (Chapter 2), and is now an integrated part of most marine turtle foraging ground studies. However, the use of MSA of turtles at foraging grounds has been limited within the Australasian region (i.e., Broderick et al. 1994), although several new studies have recently been completed (Dethmers et al. in prep, Bell et al. in prep, this thesis). In this study I used mtdna sequencing techniques to study immature green turtles at the foraging site studied by Pilcher (2010) and a second site also off the coast of Borneo, Malaysia, to assess the natal origin of turtles utilising these foraging grounds. This study provides a test of whether the aggregations of turtles that have observed female-biased sex ratios originate from rookeries with a long tradition of relocating nests into unshaded hatcheries. 62
73 4.2 Materials and Methods Study site and sample collection The two study sites around Mantanani Island (N 6 71, E ), and Layang Layang (N 7 40, E ) are located northwest of Sabah, Malaysia, and separated from each other by approximately 290 km (Figure 4.1). Turtles were captured around Mantanani Island in December 2006, January, April and October 2007, January, September and December 2008, and February Captures at Layang Layang were carried out only in October Immature green turtles were captured by rodeo-style capture (Limpus & Reed 1985) in the shallow (> 30 cm) to deeper (2.5 m) water. All captured turtles were flipper-tagged, weighed and measured for Curved Carapace Length (CCL; ± 0.1 cm) and a skin biopsy taken and stored in NaCl saturated 20% DMSO. Gender was determined for all first time captures using laparoscopy procedures (Miller & Limpus 2003). Turtles were returned to the sea within one to two hours of capture Characterisation of mtdna haplotypes DNA was extracted using the salting out method described by FitzSimmons et al. (1995). DNA was resuspended in LTE buffer (0.1 mm EDTA, 10 mm Tris, ph 7.5). Primers LTEi9 (GGGAATAATCAAAAGAGAAGG-3 ) and H950 (GTCTCGGATTTAGG GGTTT-3 ) (Abreu-Grobois et al. 2006) were used to amplify a ~780-bp fragment of the mtdna control region. PCR was performed in a 25 μl reaction containing 1x reaction buffer (1.5 mm MgCl2), 0.25 mm of each dntp, 10 μm of each primer and 1.25 U of Taq polymerase and ~40 ng of template DNA. The PCR protocol consisted of a 5 min denaturing step (94 C) followed by 35 cycles of: 45 s at 94 C (denaturing), 45 s at 52 C (annealing) and 45 s at 72 C (extension) and a final extension step of 5 min at 72 C. PCR products were purified using a PEG cleanup procedure and nucleotide sequences were determined for both strands of the PCR amplification products at the Macrogen sequencing facility (Macrogen Inc., Seoul, Korea). Geneious Pro ( was used to generate a single consensus strand from the aligned forward and reverse sequences for each sample. All sequences were aligned using Clustal W as implemented in Geneious and corrected by eye when necessary. Haplotypes were identified by running a search against a collated database of known green turtle haplotypes. If no matching haplotypes were found, the BLAST search procedure was used 63
74 to search the Genbank database ( If no matching haplotypes were found the sequence was identified as being unique and given a name. Each new sequence was compared with previously published haplotypes of the mtdna control region based on shorter sequences. If there was an identical sequence match with the shorter segment, the original nomenclature for the short sequence was used and a new name was given to the longer sequence. Naming of new haplotypes followed a standardised nomenclature for Indo-Pacific green turtles using the prefix CMIP-followed by the next sequential number. Following haplotype identification, a randomised chi-square test was performed using the program CHIRXC (Zaykin & Pudovkin 1993) to determine if there were significant shifts in haplotype frequencies between the two foraging grounds and between different years Mixed Stock Analysis Initial baseline information for the MSA was assembled using mtdna haplotype frequencies of 17 genetically differentiated Australasian green turtle breeding stocks (Dethmers et al. 2006) and data from Chapters 3 and 5; Figure 4.1 and Table 4.1). For this initial analysis, new sequences were cropped to include only the 384-bp region reported in Dethmers et al. (2006). A second, baseline was set up using of 12 breeding stocks from the eastern Indian Ocean (Chapter 3) and the ngbr (Chapter 5) that had been characterised for the entire 780-bp sequence. A Bayesian approach using the program BAYES (Pella & Masuda 2001) was used to estimate the most likely origin of each individual and the contribution of the different Australasian rookeries to the foraging grounds studied. This analysis was conducted using both uniform and weighted priors (assumptions). In the analysis using uniform priors, each of the rookeries was equally likely to contribute individuals to the foraging aggregations. For the weighted prior analysis, the potential contributions of different rookeries to the foraging aggregations were weighted relative to the size of the rookery. Thus, large rookeries were assumed to have contributed more individuals to the two aggregations. A total of 50,000 Markov Chain Monte Carlo (MCMC) steps were run for 17 chains using the short sequence and 13 chains using the long sequence, each with different starting points. A burn-in of 25,000 runs was used to calculate the posterior distribution. The Gelman and Rubin 64
75 shrink factor diagnostic was computed to test that all chains had converged as indicated by a shrink factor less than 1.2 for each chain (Pella & Masuda 2001). Individuals with haplotypes not observed in any of the nesting rookeries were removed from the analysis by the program as these were uninformative. 4.3 Results Of the 91 green turtle tissues sampled only 81 samples were successfully amplified by PCR (Table 4.1, see also Appendix 2) due to the degradation of ten samples. Samples were collected from Mantanani Island (2006: females = 23 and males = 3; 2007: females = 24 and males = 9; 2008: females = 11 and males = 3) and Layang Layang (2006: females = 12 and males = 6). The sex ratio for all turtles caught was 1M: 4F (n = 73) at Mantanani and 1M : 2F (n = 18) at Layang Layang and at both areas combined the sex ratio was 1M : 3.3F. A total of 12 haplotypes were identified for the short fragment of the mtdna control region of which three were previously undescribed. These were found in one (CMIP-71), two (CMIP-72) and three (CMIP-70) individuals and comprised 7.1% of the sampled individuals. The most common were haplotypes C3 found in 21.2 %, D2 found in 18.8% and C4 found in 16.5% of the samples (Table 4.1). One individual had a haplotype (B7) only found previously in northern Great Barrier Reef (ngbr) rookeries. Five individuals had a haplotype (CmP19) not found within the baseline samples, however this haplotype has been found at low frequency (> 3%) at a rookery in Taiwan (P. Dutton, pers. com). Two haplotypes (D2 and C3) split up into two and three new haplotypes, respectively, when using the entire 780 bp sequence (Table 4.1 and Appendix A). There was no evidence for temporal variation in haplotype composition between turtles sampled in different years at Mantanani [ (χ2 = 12.68, P = 0.20), (χ2 = 8.75, P = 0.31) or (χ2 = 4.31, P = 0.94)], nor between Layang Layang (2006) and Mantanani [2006 (χ2 = 7.26, P = 0.61), 2007 (χ2 = 8.12, P = 0.69), and 2008 (χ2 = 4.5, P = 0.87)]. When sampling years were combined the two foraging areas also had similar genetic composition (χ2 = 9.50, P = 0.596). Furthermore, there was no difference in haplotype frequencies between males and females (χ2 = 17.09, P = 0.094) thus the data were pooled for all further analysis. 65
76 Table 4.1. Haplotype frequencies of nine nesting populations that either had a mean contribution of ( 1%) in mixed stock analysis to the two foraging grounds (FG) or shared several haplotypes with the FGs. Abbreviations are; ngbr, northern Great Barrier Reef; Mic, Micronesia; PNG, Papua New Guinea; PM, Peninsular Malaysia; Sar, Sarawak; SS, Sulu Sea; Ber, Berau; Sip, Sipadan; and Aru, and the two foraging aggregations Man, Mantanani and LL, Layang Layang. Haplotypes new is based on the ~780 bp fragment using primers LTEi9 and H950, short is based on a ~380 bp fragment used by Dethmers et al. (2006) and other are haplotypes reported by different authors that correspond to the old haplotypes. Nesting Populations FGs ngbr Mic PNG PM Sar SS Ber Sip Aru Man LL Total Haplotypes New Old Other CMIP-28 J CMIP-24 B CMIP-34 C CMIP-13 D CMIP-22 B7 CmP CMIP-21 B CMIP-05 A CMIP-29 B E CMIP-02 A3 CMJ8 4 CmP A A CMIP-01 1 CMIP-119 CMIP-67 C3 1 CMJ CMIP-68 C CMIP-66 B CMIP-57 CMIP-43 D2 2 CMJ CMIP-69 C CMIP-26 C14 CMJ CMIP-132 A CMIP-03 C CMIP-56 - CmP CMIP CMIP CMIP Pop Size Haplotype C3 was split into three new haplotypes (CMIP-01, n = 18; CMIP-67, n = 2 and CMIP-119, n = 1) when using the longer sequence. 2 Haplotype D2 was split into two new haplotypes (CMIP-57, n = 11 and CMIP-43, n = 6) when using the longer sequence. 3 Dutton unpublished, 4 Hamabata et al Pop sizes are drawn from Basintal & Lakim 1993, Pilcher & Basintal 2000, Palma 1993, Limpus et al. 2001, de Silva 1982, Leh 1993, Adnyana et al
77 Mixed Stock Analysis The initial mixed stock analysis using the short sequence and all rookeries indicated that most rookeries were likely contributors to the foraging ground as they had an upper 95% CI larger than zero. This was true for both the estimates using uniform and weighted priors (Table 4.2). However, when using uniform priors, the estimates for contributions from Sipadan were disproportionally large relative to those from the Sulu Sea given the very small size of the Sipadan population in comparison to that of the Sulu Sea population. When rookery size was accounted for as a weighted prior, the estimated contribution from Sipadan decreased relative to that from the Sulu Sea. Nonetheless, the confidence intervals (CI) for all estimates were large and the lower 2.5% CI included zero in all cases except for Sarawak (mean = 31%, 95% CI = 16-54%). This likely reflects the presence of a number of haplotypes in the region that are broadly distributed among rookeries. To test this, Pearson correlation coefficients were calculated for the MCMC sample of the individual stock proportions (Pella & Masuda 2001). High posterior correlations can sometimes be attributed to slow convergence (indicated by a large shrink factor). Indeed, a high posterior correlation between Sulu Sea, Berau and Sipadan rookeries indicated that in order to provide more reliable estimates of the contributions of these stocks, these rookeries should be combined to determine a regional estimate. To do this, the program estimates baseline stock proportions individually, then pools the estimates of chosen stocks and estimates the uncertainty for this pooled estimate. When the three rookeries were grouped (East Borneo group) to provide a regional estimate, the CI decreased, it did not include zero, and the mean contribution from those three rookeries combined was 47%, (95% CI = 24-77%). Together, the East Borneo group and Sarawak were estimated as being the origins of 78% of the foraging ground turtles sampled. The next highest mean estimate of contribution was 13% from Aru, but as with all remaining estimates, a zero value is included within the 95% CI making the estimates too uncertain. Given that the initial MSA showed no contribution from outside the 13 breeding stocks that had been characterised for the longer sequence, only these 13 stocks were used in the a second analysis. MSA using the data from the longer sequence and a weighted prior analysis showed similar results to that of the short sequence, but with a few interesting differences (Table 4.3). First, the contribution from Aru dropped from 13% to 0% while the contribution from Ashmore Reef increased from 1% to 7%. These estimates all had large CIs. The contribution from Sarawak increased from 31% to 60%. Previous ambiguous results for Sulu Sea, Sipadan and Berau Island appeared to be resolved using the longer sequence. The larger Sulu Sea was 67
78 identified as the main contributor (29%) from eastern Borneo, with little or no contributions from the much smaller stocks at Sipadan and Berau Island. Given that the use of the longer sequence added some resolution to the genetic structure by splitting up common haplotypes C3 and D2 at both foraging and regional breeding stocks the discussion will focus on the estimates from the long sequence using weighted priors. Table 4.2. Estimates of the rookery origin of immature green turtles foraging at Mantanani Island and Layang Layang Island, based on Bayesian estimations that incorporated 17 major rookeries as potential source populations and using the shorter 384 bp sequence. Haplotypes found at the foraging grounds but not in any of the baseline rookeries were excluded from the analysis and their proportion reported as unknown origin. The posterior probability estimates, median and the 95% confidence intervals are shown for estimates using both weighed priors (using rookery size as a prior) as well as a uniform prior. Rookeries Weighed prior Uniform prior Media Media Mean n 2.5 % 97.5 % Mean n 2.5 % 97.5 % ngbr 1% 1% 0% 5% 1% 0% 0% 4% Coral Sea 0% 0% 0% 0% 0% 0% 0% 1% sgbr 0% 0% 0% 1% 0% 0% 0% 1% New Caledonia 0% 0% 0% 0% 0% 0% 0% 1% Micronesia 0% 0% 0% 4% 1% 0% 0% 6% PNG 1% 0% 0% 12% 4% 3% 0% 15% Penin. Malaysia 4% 0% 0% 31% 3% 0% 0% 25% Sarawak 31% 30% 16% 54% 27% 26% 15% 42% East Borneo group* 47% 45% 24% 77% 47% 47% 12% 70% Sulu Sea 28% 32% 0% 56% 18% 20% 0% 47% Sipadan 18% 0% 0% 76% 26% 23% 0% 68% Berau 2% 0% 0% 41% 2% 0% 0% 33% Aru 13% 5% 0% 27% 12% 13% 0% 25% Gulf of Carpentaria 0% 0% 0% 0% 0% 0% 0% 2% Ashmore Reef 1% 0% 0% 13% 2% 0% 0% 17% Scott Reef/Browse 0% 0% 0% 0% 0% 0% 0% 5% West Java 0% 0% 0% 5% 2% 0% 0% 20% Cocos (Keeling) 0% 0% 0% 4% 0% 0% 0% 4% Northwest Shelf 1% 1% 0% 5% 0% 0% 0% 1% Unknown origin 14% *Regional estimates for Sulu Sea, Berau and Sipadan combined. 68
79 Table 4.3 Estimates of the rookery origin of immature green turtles foraging at Mantanani Island and Layang Layang Island, based on Bayesian estimations using 13 major rookeries characterised for the longer 780 bp sequence as a baseline. Haplotypes found at the foraging grounds but not in any of the baseline rookeries were excluded from the analysis and their proportion reported as unknown origin. The posterior probability estimates, median and the 95% confidence intervals are shown for estimates using both weighed priors (using rookery size as a prior) as well as a uniform prior. Rookeries Weighed prior Uniform prior Media Media Mean n 2.5 % 97.5 % Mean n 2.5 % 97.5 % ngbr Penin. Malaysia Sarawak Sulu Sea Sipadan Berau Aru Cobourg Penin Ashmore Reef Scott Reef/Browse West Java Cocos (Keeling) Northwest Shelf Unknown origin 14% 4.4 Discussion Green turtles sampled in foraging grounds at two locations off north-western Sabah were all immature turtles that presumably use these areas as developmental habitat. While a few adult turtles have been observed in the deeper water surrounding the reef, data suggest this foraging area is unsuitable for larger turtles (Pilcher 2010). Where the turtles go once they reach ~60 cm CCL is unknown but they likely take up residence in nearby foraging habitats suitable for adult turtles. This contrasts with many foraging grounds on the Great Barrier Reef where both immature and adult turtles are found foraging on the same reef (Limpus & Reed 1985). Genetic analyses of these two foraging ground populations show they are genetically similar and that both are comprised of turtles from multiple rookeries. The majority (60%; 95% CI = 45-79%) originate from Sarawak in north-western Borneo, with another large portion (29%; 95% CI = %) of turtles originating from Sulu Sea on the east coast of Borneo. Many of these sites have a long tradition of relocating eggs to unshaded beach hatcheries, thereby resulting in highly female-biased hatchling production (Tiwol & Cabanban 2000). 69
80 An individual with haplotype (B7 or CMIP-22), only found in ngbr rookeries, provided an unanticipated result and some additional insights. This result may indicate that a small proportion of turtles may have originated from nesting beaches more than 3800 kilometres to the southeast. A few green turtles, flipper-tagged while nesting in the ngbr, have been recovered from foraging grounds in eastern Indonesia 2770 kilometres from the beach where they were tagged (Limpus 2007). This suggests that oceanic currents can carry migrating post-hatchlings north and westwards from the ngbr into waters of the Banda and Celebes Sea and that some eventually take up residence there. Another turtle, tagged while foraging on Clack Reef in the ngbr, was recorded nesting on a beach in the Marshall Islands 3,800 km away (Limpus et al. 2009). It remains inconclusive whether some ngbr nesting turtles travel to foraging grounds in the South China Sea or alternatively if the B7/CMIP-22 haplotype exists undetected in rookeries closer to Sabah. The five turtles with haplotype CmP19 are likely to originate from an unknown (or not-yet-sampled) rookery. This haplotype has been found in turtles nesting in Taiwan, but at a low frequency (0.025, n = 40; P. Dutton pers. comm.). Given that the most frequent haplotype from Taiwan rookery (P. Dutton pers. comm.) was not found among our foraging assemblage makes it an unlikely source population When interpreting the results of MSA, consideration must be given to whether all rookeries in the region have been sampled, or if the sample sizes from any given rookery may be too small to give a good representation of haplotype frequencies, especially rare haplotypes. These shortcomings apply to this study as evidenced by the proportion (14%) of haplotypes found among the foraging aggregation but not identified in the baseline samples. Although the baseline sample covers the majority of the main nesting populations in the region, there are gaps in the coverage, especially from the region north of Borneo. Many small to medium sized nesting events have been recorded for areas including Vietnam (Hamann et al. 2006), Taiwan (Cheng et al. 2009), and China (Shizheng & Shi 2009). Recent satellite tagging studies indicate that the north-western Sabah foraging grounds are indeed composed of turtles coming from numerous rookeries for which we have no genetic data including; Vietnam, (Hamann et al. 2006), Terengganu (van de Merwe et al. 2009) and Taiwan (Cheng et al. 2009). A more comprehensive sampling of the baseline rookeries is needed for the region and should be a priority for future work. 70
81 Shared haplotypes can reduce the power of the MSA. This is observed in locations with haplotype C3 that is shared between Peninsular Malaysia, Sarawak, Sulu Sea, Berau and Sipadan rookeries. Haplotypes D2 and C5 are shared between Sulu Sea, Berau and Sipadan rookeries, and haplotype C14 is shared between Berau, Sipadan and Aru rookeries. This can result in large uncertainty surrounding the estimates for these populations, as observed for analyses using the short sequence. Uncertainty caused by small sample sizes or shared haplotypes can sometimes be decreased by either pooling samples or producing group estimates as was done for the short sequence data. This can be useful especially for geographically adjacent and genetically similar stocks, such as Sipadan, Sulu Sea and Berau. Another method is to increase the resolution of the genetic markers. Analysis of the entire ~780 bp mtdna region in the foraging ground samples resulted in identification of several new haplotypes via splitting of the two most common and widespread haplotypes. This analysis; 1) resulted in substantial shifts in contributions, as seen in the point estimate for Sarawak that nearly doubled, 2) it, resolved the ambiguities between Sulu Sea, Sipadan and Berau Island, indicating that the Sulu Sea population supplied an estimated 30% of turtles to the foraging ground and 3) it yielded estimates with smaller confidence intervals. Although the two foraging aggregations studied here showed no significant differentiation in haplotype frequencies, they do differ in their sex ratio, with the Mantanani sample being more female biased (1M:4F) than that at Layang Layang (1M:2F). However, it remains inconclusive whether this is a true pattern or the effect of small sample sizes. If the Sarawak and Sulu Sea rookeries are indeed the main sources of recruitment into these foraging aggregations, and they are producing almost entirely females, different contribution estimates (from Sarawak, Sulu Sea, Berau, Sipadan) would be expected between males and females. In order to test this, an increased sample size of > 60 individuals is needed for both males and females at the two foraging areas. Nonetheless, given the high female bias and the estimate that the majority of turtles at the two foraging grounds likely originates from the main hatcheries at Sarawak, Sipadan and Sulu Sea, it is clear that the problem of sex-biased hatchling production needs to be addressed. The data provided by this study emphasise the need for long-term management plans that address historical impacts as well as contemporary ones, and the importance of foraging ground studies as a record of the viability of the population. There is a need for management practices to adapt and reflect the biological needs of the turtles themselves. In hatchery 71
82 operations, factors such as nest depth and shading can be manipulated to influence nest temperature and thereby hatchling sex (van de Merwe et al. 2005, 2006). Clearly, nesting beach projects are very important for marine turtle conservation and recovery, but those beaches also benefit from the protection of regional foraging aggregations and what happens in one area directly impacts the other. Understanding the factors that drive the foraging ground demographic structure and composition allows for a forward-looking model to predict trends. This research provides initial data towards a better understanding of connectivity between nesting and foraging habitats for green turtles in a region that lags behind the intensively-studied Caribbean, Atlantic and Australian populations. It also highlights the strengths of using genetic data to estimate the origin of turtles at foraging grounds, and the limitations that need to be addressed in future work. 72
83 CHAPTER 5 5. Stock composition of green turtle (Chelonia mydas) foraging grounds of the Great Barrier Reef: implications of variation across latitude and size classes 5.1 Introduction Migratory species present unique and complex challenges for conservation. Movement of animals between nations, and therefore jurisdictions, complicates legislative and regulatory conservation policies that are effective within single nations (Miller 2000, Campbell et al. 2009). The threats faced by migratory species at foraging grounds may be quite different from those at breeding grounds, which increases the level of understanding required by conservation managers to sustain populations. Additionally, migration itself can bring with it special risks that can be exacerbated by human-induced changes to the environment. For example, salmon spawning in Californian rivers may be affected by fisheries in the open ocean thousands of kilometres away (Miller 2000), and human-made dams block salmon and steelhead migration to breeding grounds above the dams (Williams et al. 2008, Waples et al. 2008, Waples et al. 2009). For sea turtles, long migrations between foraging and nesting regions expose them to anthropogenic threats at both nesting beaches and foraging grounds. At nesting beaches animals or eggs can be harvested, and at foraging grounds, turtles are killed in subsistence or commercial harvests, incidental by-catch in fisheries and from pollution and destruction of habitat (Lutcavage et al. 1996, Limpus 2007)., Additionally, impacts of climate change are expected to affect habitat quality and availability at nesting beaches as well as foraging grounds (Hamann et al. 2007, Hawkes et al. 2009, Poloczanska et al. 2009). Biologists and managers must be able to identify populations and to fully understand the stock boundaries of endangered species to effectively work towards survival of the populations. Green turtles (Chelonia mydas) are the most abundant species of marine turtles in Australia (Limpus 2007). They nest on mainland and island beaches, and two of the largest breeding 73
84 populations in the world are found along Australia s Great Barrier Reef; the southern Great Barrier Reef (sgbr) genetic stock and the northern Great Barrier Reef (ngbr) genetic stock (Figure 5.1) (Limpus et al. 2000, Dethmers et al. 2006). Green turtles that hatch from these regions spend several years drifting with major ocean currents before they recruit back into coastal foraging areas around coral reefs and seagrass beds at a size of cm, curved carapace length (CCL) (Limpus 2007). While some populations of marine turtles shift habitat as they pass through different life stages, most green turtle populations in Australia show high fidelity to their choice of foraging ground (Musick & Limpus 1997, Limpus 2007). The east coast of Australia is home to one of the oldest continuous monitoring programs of both nesting and foraging marine turtles in the world. These foraging ground mark recapture programs have provided important detailed sex and age-class-specific demographic information and have revealed links between foraging and nesting habitats,limpus & Reed 1985, Limpus 1993, 2007, Limpus et al. 1992, 1994, 2000), but they are logistically difficult and resource intensive. Furthermore, they typically only connect foraging grounds to nesting beaches for mature females undertaking their breeding migrations, failing to capture the male and non-adult component of the population. In some areas, both ends of the breeding migrations of adult males have been determined by capturing breeding males at mating areas or around nesting beaches (Limpus 1993), or by satellite tracking of male turtles (Limpus 1993, Hays G. et al. 2001, van de Merwe et al. 2009). The use of genetic markers provides an alternative to traditional mark-recapture techniques that can be applied to all gender and size classes. Using Mixed Stock Analysis (MSA), the relative proportion of genetically differentiated nesting populations represented at a given foraging ground can be estimated (Bass et al. 1998, 2006, Lahanas et al. 1998, Bass & Witzell 2000, Luke et al. 2004, Bowen et al. 2007). This not only provides valuable information about migration pathways and the geographic range of the stocks, but also on the nesting populations that are likely to be impacted by potential threats at the foraging grounds. In this way, mortality at foraging grounds can be proportionally attributed to each breeding stock. For example, the application of MSA revealed that by-catch in Mediterranean fisheries impacted breeding populations of loggerhead turtles on nesting beaches in Florida and Mexico (Laurent et al. 1998). 74
85 Despite more than 40 years of monitoring the Great Barrier Reef foraging populations, little is known about how juvenile green turtles are distributed across available foraging grounds and how the composition of foraging grounds changes though time. Particularly within the northern Great Barrier Reef (ngbr), sea turtle populations are impacted by subsistence harvest from Indigenous communities, both at nesting beaches and at foraging grounds. Detailed monitoring of foraging populations allows for early detection of biological trends and potential threats to marine turtles without the lag time required for those trends to manifest in nesting adult populations (Chapter 2). This paper presents a detailed MSA of green turtle aggregations at six major foraging grounds along the east coast of Australian (Figure 5.1). The genetic results are combined with data from more than 30 years of mark recapture efforts along the Great Barrier Reef to address the following three questions: 1) Does the information obtained by traditional flipper tagging for adult females match estimates derived from MSA? 2) Are there differences in the stock composition of foraging grounds among size classes? 3) Is there temporal variation in the stock composition of foraging grounds between turtles sampled in the early 1990s and turtles sampled from 2005 and 2009? This study is the first description of the complex spatial patterns of variation in foraging populations resulting from the interaction of different lifehistory components of a marine turtle population. Information of this nature is critical for the design and implementation of scientifically based conservation to underpin management policies for sea turtles. 5.2 Materials and Methods Study site and sample collection This study investigates the composition of six major green turtle foraging grounds along the extent of the Great Barrier Reef Marine Park, Queensland, Australia from the Torres Strait in the north to Moreton Bay in the south and encompassing more than 2300 km. Approximate locations of the sample areas are provided in Figure 5.1, and additional information about each area is provide below, as listed from north to south. 75
. Torres Strait (TS) (S 10 29.59, E 142 10.44 ) is located between the tip of the Cape York Peninsula and Papua New Guinea.")
86 Figure 5.1. Map showing the location of 17 genetically differentiated breeding stocks initially included in the MSA analysis and the six foraging grounds analysed for stock composition. Modified from Dethmers et al. (1996). Torres Strait (TS) (S , E ) is located between the tip of the Cape York Peninsula and Papua New Guinea. The southern area of Torres Strait marks the northern extreme of the Great Barrier Reef Marine Park and consists of a cluster of islands and reefs. Torres Strait links the Coral Sea to the Arafura Sea and is influenced by strong currents passing through the Strait. An agreement between Australia and Papua New Guinea, known as the Torres Strait Treaty, governs the management of the Torres Strait region and was established to resolve issues of sovereignty and maritime boundaries, to recognise and protect the traditional way of life and livelihood of Traditional Inhabitants, and to protect the regions marine environment and resources. The Torres Strait Fisheries Act 1985 gives effect to Australia s obligations under the Treaty in managing the fisheries resources including marine turtles. Clack Reef (CR) (S , E ) is located within the ngbr and is one of several green turtle foraging sites that has been the subject of a long-term mark-recapture program. Clack Reef has a large reef platform supporting seagrass areas. Clack Island is a 76
87 significant sacred site and is an important place in Aboriginal ritual and mythology. It is the traditional centre for male ritual activities and therefore has restricted access. The Howicks Group (HG) (S , E ) is a group of islands consisting of 16 reefs, sand cays and several remnant islands. Most reefs within the Howicks Group have large sand covered reef platforms with active coral growth restricted to their margins, but three of the 16 reef support oval-shaped and vegetated sand cays. Edgecombe Bay (EB) (S , E ) is located in the central section of the Great Barrier Reef Marine Park. Large patches of seagrass beds are located along the fringe of the Bay before it rapidly drops off to depths of up to 6 m. Coral reef can be found in small areas along the eastern and the western sections of the Bay. Shoalwater Bay (SB) (S , E ) is also located in the central section of the Great Barrier Reef Marine Park and supports a large foraging population of green turtles. This large embayment contains large intertidal seagrass beds with mangroves on the landward fringe and represents one of the least impacted foraging populations in Australia due to its protection as a military training area. Moreton Bay (MB) (S , E ) is located outside the Great Barrier Reef Marine Park and represents the most southern large green turtle foraging ground in Queensland. Moreton Bay is a large embayment with significant seagrass and mangrove areas. Turtles are often found foraging across the seagrass flats and into the mangroves with the rising tide and fall back to deeper water at low tide Sample collection To establish the baseline data for the nesting populations, additional samples from the ngbr were analysed for comparison with previous haplotype frequencies (Dethmers et al. 2006). This was necessary because a re-analysis of a random sample suggested that a few ngbr samples reported in Dethmers et al. (2006) were incorrectly assigned to haplotype using denaturing gradient gel electrophoresis.. Therefore, the original 53 nesting turtles collected in 1990 from the ngbr were re-sequenced (Dethmers et al. 2006). In addition, samples from 32 new nesting females collected from Raine Island (ngbr) in 2008 were analysed to assess 77
88 whether there was temporal variability in the genetic structure of the ngbr nesting population (Table 5.1). Foraging ground samples were collected from a combination of different size classes; juvenile (35-55 cm CCL), sub-adult (56-86 cm CCL) and adult turtles (> 86 cm CCL). Contemporary samples were collected between and historical samples were collected in the early 1990s. This allowed for the assessment of the foraging ground composition across size classes as well as temporal variability. Overall, the genetic sample (n = 967) consisted of 98 juvenile (16 in 1992 and 82 in ) and 146 adults (88 in 1992and 58 in ) from Torres Strait; 24 juveniles ( ), 22 sub-adults ( ) and 45 adults ( ) from Clack Reef; 84 juveniles ( ) and 79 adults ( ) from the Howicks Group; 83 juveniles ( ) and 85 adults ( ) from Edgecombe Bay; 42 juveniles ( ), 45 subadults ( ) and 88 adults ( ) from Shoalwater Bay and 80 juveniles (1990) and 46 sub-adults (1990) from Moreton Bay (Table 5.2). All turtles were captured by rodeo-style capture (see Limpus & Reed 1985) in the shallow (> 30 cm) to deeper (< 5 m) water with the exception of some adult turtles from Torres Strait collected in A subset of these samples (n = 67) originated from harvested turtles sampled at the Daru markets in Papua New Guinea, but all turtles were taken from within the Torres Strait. Furthermore, they were collected outside the nesting season to minimise the chance of collecting non-resident turtles undertaking their breeding migration. All captured turtles were flipper-tagged, measured for curved carapace length (CCL; ± 0.1 cm) and a skin biopsy was taken and stored in sodium chloride (NaCl) saturated 20% DMSO. The gender of all adult turtles was determined by sexspecific morphological characteristics. The identification of gender in immature green turtles relies on examination of the reproductive organs via laparoscopy (Miller & Limpus 2003) and was only identified for immature turtles caught in the early 1990s, and a small proportion of juveniles from the Torres Strait caught in Characterisation of mtdna haplotypes DNA was extracted using a salting out method as described in FitzSimmons et al. (1995). DNA was re-suspended in a low concentration TE buffer (0.1 mm EDTA, 10 mm Tris, ph 7.5). The primers LTEi9 (GGGAATAATCAAAAGAGAAGG-3 ) and H950 (GTCTCGGATTTAGGGGTTT-3 ) (Abreu-Grobois et al., 2006) were used to amplify a ~780-bp fragment of the mtdna control region. PCR was performed in a 25 μl reaction containing, 1 x Reaction buffer, 0.25 mm of each dntp, 10 μm of each primer, 1.25 U of 78
89 Taq polymerase and ~40 ng of template DNA. The PCR protocol consisted of a 5 min denaturation step (94 C) followed by 35 cycles of: 45 s at 94 C (denaturing), 45 s at 52 C (annealing) and 45 s at 72 C (extension) and a final extension step of 5 min at 72 C. PCR product was purified using a PEG cleanup procedure and sequencing of both forward and reverse strands was carried out by Macrogen (Macrogen Inc., Seoul, Korea). Geneious Pro ( was used to generate a single consensus strand from the aligned forward and reverse sequences. All sequences were aligned using Clustal W as implemented in Geneious and corrected by eye when necessary. Haplotypes were identified by running a search against known green turtle haplotypes. If similar haplotypes were not found using the initial search, a BLAST analysis was used to run a search against the Genbank database ( If still no similar haplotypes were identified, the sequence was identified as being unique and named accordingly. To assess the genetic diversity of each group (i.e., juveniles and adults within and among foraging grounds), haplotype (h) and nucleotide diversity (π; Nei 1987) were estimated, and an analysis of molecular variance (AMOVA) was conducted using Arlequin version 3.0 (Excoffier et. al 2005). In addition, a randomised chi-square test (chirxc; Zaykin & Pudovkin 1993) was used to determine if there were significant shifts in haplotype frequencies among groups within each foraging ground, between foraging grounds, and between foraging grounds and the ngbr and sgbr rookeries Mixed Stock Analysis Baseline information for the mixed stock analysis (MSA) was derived from the mtdna haplotype frequencies of 17 genetically differentiated green turtle breeding stocks in Australasia (Dethmers et al. 2006), in combination with new samples from Raine Island. All new sequences were cropped to include only the 384 bp region reported in Dethmers et al. (2006). Two different MSA approaches were used to estimate the most likely origin of each individual and the contribution of the different Australasian nesting rookeries to the foraging grounds. One approach used the program Bayes (Pella & Masuda 2001) and the other used the many-to-many analysis (Bolker et al. 2007) using the software package R. Both use a similar Bayesian approach but while BAYES can only analyse one foraging aggregation at a time, the many-to-many approach puts the analysis into a meta-population context and can include multiple foraging grounds into one analysis. Both methods were conducted using either uniform or weighted priors. When using uniform priors it is assumed that each rookery 79
90 has the same likelihood of contributing individuals to the foraging aggregations. When using weighted priors, the contributions of different rookeries are weighted relative to the size of the rookery. This assumes that larger rookeries contribute more individuals to foraging aggregations. Because the Bayesian analysis has a tendency to overestimate a small or zero contribution, when using a high number of possible sources the precision of the estimates can be affected, especially if many are unlikely contributors. Therefore, an initial run of the MSA using all 17 breeding stocks as possible sources was conducted in BAYES to assess the contribution from rookeries more than 2000 km away. This resulted in nine rookeries being excluded as being unlikely contributors (upper 97.5 CI < 0.02). Hence the final analysis was simplified by using only eight source populations (Table 5.1). Using these eight rookeries, eight chains were run each with different starting points. Each chain was completed using 20,000 Markov Chain Monte Carlo (MCMC) steps and a burn-in of 10,000 runs to calculate the posterior distribution. The Gelman and Rubin shrink factor diagnostic was calculated to test that the posterior probability distribution of all chains had converged (shrink factor > 1.2) (Pella & Masuda 2001). Individuals with haplotypes not observed in any of the nesting rookeries were removed from the analysis by the program as uninformative. MSA was conducted for each group (juveniles, sub-adults, and/or adults) within each foraging aggregation. To obtain the best possible estimates four different analyses were run; two using BAYES, with either uniform (BAY1) or weighted (BAY2) priors and two using the many-to-many analysis with either uniform (MTM1) or weighted (MTM2) priors Migration data from mark-recapture studies For comparison with the MSA analyses, migration data from flipper tagging was compiled from the Queensland Parks and Wildlife Marine Turtle Research Database for TS, CR, SB and MB. These data included tag recoveries from turtles tagged at breeding areas and recaptured at their foraging areas or vice versa. 80
91 5.3 Results Sequence data were obtained from a 740-bp fragment, spaning most of the mitochondrial control region for a total of 967 foraging turtles and 85 nesting turtles. This region contained the 384-bp segment previously sampled in the regional population genetic analysis of Australasian green turtles (Dethmers et al. 2006) (Figure 5.1) and all analyses were done using only the short fragment Haplotype diversity along the GBR foraging grounds A total of 67 haplotypes were identified. The four most common haplotypes were A2 (50%), B1 (19%), B3 (8%) and B7 (6%), all of which were found in high frequency within the ngbr and/or sgbr breeding stock (Table 5.1). A large number of haplotypes (n = 41) were previously undescribed (orphan haplotypes) and had not been found in any rookery sampled previously. These were all found in low frequency ( %) and made up less than 6% of the total sample. Within some sample groups, these haplotypes made up a large proportion of the haplotypes. The proportion of these orphan haplotypes was highest (10-15%) in the TS sample, excluding juveniles from , for which the sample size was low. Orphan haplotypes were similarly in high frequencies among juveniles in CR (13%) and the HG (10%) and low to moderate for all other groups (0-10%) (Table 5.1). A large range in genetic diversity was observed, with the lowest values found in the southernmost foraging grounds. Nucleotide and haplotype diversity was lowest in MB and SB (h = , and π = ), they increased to the north and were highest in TS (h = , and π = ) (Table 5.2). Haplotype A2 was the most common in the southernmost foraging grounds and accounted for approximately 80% of foraging turtles at MB and SB. Along with the latitudinal change in diversity, haplotype A2 decreased to account for only 12% of haplotypes at TS while the most common haplotype, B1, increased to account for 39% of the haplotypes (Table 5.1). To test for hierarchical structuring among the southern (MB, SB, EB and the sgbr breeding stock) and northern (HG, CR, TS and the ngbr breeding stock) the AMOVA was run using these two groups and showed that the among-region variance (FCT) component was large (0.46) (Table 5.3). 81
92 82 ngbr-1990 ngbr-2008 Coral Sea sgbr New Caledonia Micronesia PNG Aru GoC TS-A TS-A TS-J TS-J CR-A CR-SA CR-J HG-A HG-J EB-A EB-J SB-A SB-SA SB-J MB-SA MB-J Table 5.1. Haplotype composition of the eight stocks used for the MSA and the six foraging grounds analysed; TS = Torres Strait, CR = Clack Reef, HG = Howicks Group, EB = Edgecombe Bay, SB = Shoalwater Bay, and MB = Moreton Bay, separated by sampling year and size class (A = adults, SA = sub-adults and J = juveniles). Sample size (N) and the total number of haplotypes in the foraging aggregations not recorded in any nesting population (orphan haplotypes) are shown. Rookeries Foraging grounds N B B B B J CMIP CMIP CMIP C J B A C C C A A A
93 83 Table 5.1 continued E1/E C C C Orphan Haplotypes Total C D CmP A CmP C CMIP CMIP CMIP C CMIP CMIP CMIP CMIP CMIP CMIP CMIP CMIP CMIP CMIP CMIP CMIP CMIP CMIP CMIP
94 84 Table 5.1 continued CMIP CMIP CMIP CMIP CMIP CMIP CMIP CMIP CMIP CMIP CMIP CMIP CmI CMIP CMIP CMIP CMIP Cmp
95 Table 5.2. Estimates of haplotype (h) and nucleotide diversity (π) within eight green turtle genetic stocks used for the MSA and six foraging grounds for groupings of adult and immature turtles. Rookery n Hap H SE π SE ngbr Coral Sea sgbr New Caledonia Micronesia PNG Aru GoC Foraging grounds TS-Adults TS-Juveniles CR-Adults CR-Immature HG-Adults HG-Juveniles EB-Adults EB-Juveniles SB-Adults SB-Immature MB-Sub-Adult MB-Juveniles Pairwise comparisons of variation in haplotype frequencies between years and gender classes within each foraging ground were not significant for any pairwise comparison except in the TS, where significant differences were found between juveniles and adults sampled in (χ2 = 41.16, P = 0.021), and between the juvenile and adult samples (χ2 = 50.68, P = 0.003). Pairwise comparison of the overall haplotype frequencies among all foraging grounds showed that both TS juveniles as well as TS adults were significantly different from all other foraging grounds (P < 0.05). The only non-significant differences in haplotype frequencies among foraging aggregations were between neighbouring CR and the HG (χ2 = 23.98, P = 0.36), neighbouring EB and SB (χ2 = 13.30, P = 0.54) and neighbouring SB and MB (χ2 = P = 0.183). Because no strong groupings were observed the various MSA were run for each foraging ground separately. 85
96 Table 5.3. Analysis of molecular variance (AMOVA) results for the southern and northern groups of green turtle foraging and nesting sites. The southern group included MB-J, MB-SA, SB-I, SB-A, EB-J, EB-A and the sgbr breeding stock. The northern group included HG-J, HG-A, CR-I, CR-A, TS-J ( ), TS-A and the ngbr breeding stock. The significance of permutation test (10,000 permutations) are shown for P < 0.05 (*), P < 0.01 (**) and P < (***) Variance Component % of variation F-Statistics Among regions 46.2 F (northern and southern) CT = *** Among groups within region 3.4 F SC = ** Within groups 50.4 F ST = *** Tests for haplotype frequency differences among foraging grounds and the ngbr stock gave varied results. TS-juveniles from displayed significantly different haplotype frequencies from the ngbr stock (χ2 = P = 0.000), but this was not true for haplotype frequencies between TS adults and ngbr stock (χ2 = P = 0.589), nor for juveniles from early 1990 s and the ngbr stock (χ2 = P = 0.959). All other foraging aggregations had significantly different haplotype frequencies when compared with the ngbr stock (P < 0.05). Haplotype frequencies of foraging samples in comparison to the sgbr stock were all significantly different (P < 0.05), except between the sgbr stock and SB (χ2 = P = 0.37) and MB (χ2 = P = 0.36), located roughly 200 km and 500 km respectively from the main nesting sites for the sgbr stock (Capricorn-Bunker Group) Mixed stock analysis and mark-recapture Due to high similarity between the sgbr and the Coral Sea stocks (F ST = 0.099, P = 0.049), the MSA was unable to distinguish between them and thus the estimates for these stocks were surrounded by high confidence intervals (95% CI) that included zero in all cases. Combining these two into a regional estimate resulted in a decrease of the CI. The mixed stock analysis suggested that the majority of foraging turtles from all six foraging grounds most likely originated from the ngbr or the sgbr/coral Sea stocks (Table 5.4). Both the weighted and uniform estimates gave similar results for the MSA and BAYES. The estimates using uniform priors appeared to assign a greater proportion to small contributing stocks than did the estimates using weighted (rookery size) priors. The many-to-many analysis provided similar estimates to BAYES in most tests, but it assigned a higher 86
97 contribution to small rookeries outside the GBR. This was especially evident when using uniform priors. Given the high accordance between the mark-recapture estimates (Table 5.4 and Figure 5.2) and both the BAYES analyses and, to a somewhat lesser extent, the many-tomany analyses with weighted priors analyses, they were chosen as the most reliable estimates. Given the greater support from the mark-recapture estimates for the BAYES analyses using weighted priors (BAY2), those results are presented in more detail below. The five breeding stocks of Aru, the Gulf of Carpentaria (GoC), Micronesia, New Caledonia and Papua New Guinea (PNG) were estimated to have made only small contributions to the foraging aggregations and these have been grouped into the category other. For individual estimates from these rookeries, see Appendix 3. Even though only the haplotype frequencies of juvenile turtles sampled from TS in were significantly different from adults within the same foraging ground, the MSA was run for all age classes and years individually to determine whether any trends were evident in the stock contributions estimated for the different groups across all foraging grounds Torres Strait Estimates from the weighted MSA showed similar contributions of adult turtles originating from the ngbr stock sampled in the early 1990s (0.89 ngbr, 95% CI ), and those sampled in 2009 (0.86 ngbr, CI ) (Table 5.4 and Figure 5.2). The estimates for juveniles collected in showed a lower contribution from the ngbr stock (0.53 ngbr, 95% CI ) in comparison with both groups of adults. Although the sample size (n = 16) for juveniles collected in was too small for a reliable MSA estimate, all 16 individuals had haplotypes that characterise the ngbr breeding stock. Among all samples there was only a low contribution from the sgbr/cs group (< 0.12 sgbr/cs) and from other stocks (< 0.02 other) except for juveniles sampled in Estimates for that sample indicate that the sgbr/cs group contributed 21% (95% CI ) and the contribution from other stocks combined was 31%. Most of these were estimated to be coming from GoC (0.14) and PNG (0.08). The mark-recapture data (n = 352) from adult turtles resident at TS indicated a predominance (0.91) of ngbr turtles, with small (0.06) contributions from the sgbr/cs, similar to the MSA results for adults. A low frequency (0.03) of tag returns from other stocks came from The Lacepedes (n = 1), Western Australia, Solomon Islands (n = 1) and New Caledonia (n = 3). 87
98 88 Table 5.4. Results from the Bayesian MSA for six green turtle foraging grounds (FG); TS, Torres Strait; CR, Clack Reef; HG, Howicks Group; EB, Edgecombe Bay; SB, Shoalwater Bay and MB, Moreton Bay. Each FG is divided into adults (A), sub-adults (SA), and juveniles (J) if sampled, year collected and sample size (N). MSA was calculated using 17 regional breeding stocks as possible sources, but for simplicity only the two main contributors are listed. The combined contributions of the remaining 14 stocks are compiled into the Other category. The 95% confidence intervals (CI) are shown in parentheses. Contributing stocks ngbr sgbr/coral Sea Other Flat prior Weighted prior Flat prior Weighted prior Flat prior Weighted prior FG Stage Year N Tag BAY1 MTM1 BAY2 MTM2 Tag BAY1 MTM1 BAY2 MTM2 Tag BAY1 MTM1 BAY2 MTM2 A Flipper tag % 6% 3% A % 77 % 89 % 88 % 8 % 7 % 9 % 4 % 6 % 2 % 16 % (72-97) (66-86) (77-99) (79-99) (0-18) (0-17) (0-19) (0-7) (0-23) (0-10) 8 % TS J NA NA NA NA NA NA NA NA NA NA NA NA 85 % 74 % 86 % 87 % 11 % 9 % 12 % 6 % 4 % 2 % A % 7 % (70-98) (60-86) (73-99) (75-94) (0-25) (0-21) (0-25) (0-10) (0-21) (0-8) J % 53 % 56 % 19 % 12 % 21 % 13% 31 % 26 % (37-64) (32-56) (39-66) (43-70) (0-33) (0-27) 44 % (10-33) (3-24) (17-57) (15-40) 31 % CR A Flipper tag A SA+J % 23% 12% 66% (47-82) 37 % (0-66) 55 % (40-70) 43 % (29-59) 69 % (53-85) 49 % (8-70) 73 % (57-86) 6 (41-77) 23 % (1-40) 26 % (0-57) 14 % (1-32) 19 % (1-44) 24 % (11-40) 37 % (0-60) 13 % (1-20) 23 % (8-38) 12 % (1-43) 38 % (0-99) 31 % 38 % 7 % (0-21) 14 % (0-86) 14 % 17 % HG A J % (49-76) 48 % (35-60) 57 % (44-68) 43 % (31-54) 66 % (52-79) 49 % (37-61) 7 (57-82) 52 % (39-65) 3 (12-44) 47 % (33-60) 18 % (1-37) 29 % (3-50) 32 % (18-46) 48 % (35-61) 2 (8-32) 36 % (23-50) 7 % (0-31) 5 % (0-16) 25 % 28 % 2 % (0-13) 3 % (0-10) 1 12 % 88
99 89 Table 5.4 continued Contributing stocks ngbr sgbr/coral Sea Other Stage Year N Tag BAY1 MTM1 BAY2 MTM2 Tag BAY1 MTM1 BAY2 MTM2 Tag BAY1 MTM1 BAY2 MTM2 EB A J % (0-16) (0-2) 9 % (3-17) 1 % (0-5) 1 (2-19) 1 % (0-5) 12 % (5-21) 2 % (0-6) 82 % (61-93) 88 % (72-97) 42 % (4-77) 44 % (5-79) 85 % (64-94) 92% (73-99) 77 % (66-87) 83 % (71-92) 11 % (0-37) 11 % (2-27) 49 % 65 % 5 % (0-30) 7 % (0-26) 11 % 15 % SB A Flipper tag A SA J % 96 % 3 % 1 % (0-3) 4 % (0-15) (0-4) 2 % (0-6) 6 % (1-15) 3 % (0-9) 2 % (0-6) 3 % (1-19) 3 % (0-10) 2 % (0-7) 9 % (2-23) 4 % (0-13) 98 % (91-99) 8 (55-94) 8 (50-95) 73 % (34-94) 34 % (2-69) 29 % (2-67) 98 % (94-99) 84 % (62-96) 85 % (58-99) 91 % (83-96) 72 % (55-85) 73 % (56-86) 2 % (0-8) 15 % (2-42) 19 % (4-50) 25 % 6 68 % <1% (0-3) 8 % (0-32) 12 % (0-40) 7% 19 % 23 % MB A Flipper tag SA J % 94 % 4 % 3 % (0-12) (0-3) 3 % (0-9) 2 % (0-6) 6% (0-15) 2 % (0-6) 4 % (0-14) 2 % (0-9) 89 % (71-99) 89 % (74-97) 42 % (4-80) 44 % (9-76) 92 % (80-93) 95 % (82-99) 8 (66-91) 83 % (70-92) 7 % (0-26) 11 % (2-26) 55 % 54 % 2 % (0-13) 3 % (0-16) 16 % 15 % 89

100 Figure 5.2. Map showing the location of the six foraging grounds and the mean relative contribution of ngbr, sgbr/coral Sea and other stocks for each group sampled within the foraging grounds. Estimates are based on the model BAY2 using weighted priors. Groups are based on size categories of adults (A), sub-adults (SA) and juveniles (J) and for different years of sampling (designated by the last two digits of the year). Clack Reef Due to low sample sizes of juvenile (n = 24) and sub-adult (n = 22) turtles from Clack Reef these were combined into an estimate for immature turtles (n = 46). Contribution estimates from the ngbr stock at Clack Reef were similarly high for adults (0.69 ngbr, 95% CI ), albeit being slightly lower than that for TS. Estimated contributions from the sgbr/cs (0.24 sgbr/cs, 95% CI ) and other rookeries (0.07 other, 95% CI ) was higher than for adults observed at TS. Non-adult turtles showed a lower contribution from the ngbr stock (0.49 ngbr, CI ), and increased contribution from the sgbr/cs group (0.37 sgbr/cs, 95% CI ) and other stocks (0.14 other, 95% CI ) relative to adults. However, the sample size for the immature turtles was small (n = 46) and the estimates had large CIs. The estimates of stock contribution (0.65 ngbr,
101 sgbr/cs, 0.12 other) derived from mark-recapture data (n = 48) matched that from MSA for adult turtles. Tag returns from the other stocks came from the Papua New Guinea, New Caledonia, Solomon Islands and the Marshall Islands. The Howicks group Estimated stock contributions for the HG foraging aggregation were similar to those of Clack Reef. The estimates suggest that around twice as many adult turtles originate from the ngbr stock in comparison to the sgbr/cs group, with a small contribution from stocks outside the GBR (0.66 ngbr, 95% CI ; 0.32 sgbr/cs, 95% CI , and 0.02 other, 95% CI ). Similar to the more northern foraging grounds, the estimated stock contribution from the ngbr among juveniles dropped from (0.49 ngbr, 95% CI ), and increased for the sgbr/cs group (0.48 sgbr/cs, 95% CI ). In contrast, only small contributions from other stocks (0.03 other, 95% CI ) was indicated for the juvenile turtles. Edgecombe Bay MSA estimates for EB indicated the main contribution shifted to the sgbr/cs group for both adults (0.85 sgbr/cs, 95% CI ) and juveniles (0.92 sgbr/cs 95% CI ). A low contribution from the ngbr stock was indicated for juveniles (0.01, 95% CI ), but again this was slightly higher for adults (0.10, 95% CI ). There was a low contribution from other stocks for both adults (0.05 other, 95% CI ) and juveniles (0.07 other, 95% CI ) Shoalwater Bay In SB, the MSA indicated that the foraging aggregation was dominated by turtles from the sgbr/cs group for adults (0.98, 95% CI ), sub-adults (0.84, 95% CI ) and juveniles (0.85, 95% CI ). Estimated contributions from the ngbr stock were small for all size classes (< 0.03). Contributions from other stocks were equally small for adult (<.01, 95% CI ), but somewhat higher for sub-adults (0.08, 95% CI ) and juveniles (0.12, 95% CI ). The mark-recapture data (n = 166) indicated that most (0.96) adult turtles were from the sgbr stock, with only small (0.01) contributions from the ngbr stock and from the Coral Sea and New Caledonia (0.03, other). 91
102 Moreton Bay The MB foraging aggregation was estimated to be composed of turtles from the sgbr/cs group for both sub-adults (0.92, 95% CI ) and juveniles (0.95, 95% CI ). Small contributions were estimated from the ngbr stock (< 0.06) and other stocks (< 0.03). Mark-recapture data (n = 122) turtles resident at MB indicated a large contribution (0.94) from the sgbr stock and small contributions from the ngbr stock (0.02) and New Caledonia and Vanuatu (0.04, other). 5.4 Discussion MSA and mark-recapture tagging data Mixed Stock Analysis (MSA) estimates for marine turtles are generally surrounded by large uncertainty and point estimates are used in a qualitative rather than an absolute manner. This means that comparisons of estimates among groups or foraging grounds are generally not feasible. However, research in eastern Australia has presented a unique opportunity to apply an integrated approach combining MSA with more than 30 years of mark-recapture data (Limpus 2007), which has allowed greater confidence in the point estimates generated by the MSA. Overall, the MSA in combination with the mark-recapture data supports a model in which the foraging aggregations are composed of individuals from the two Great Barrier Reef stocks (ngbr, sgbr) with small contributions from other stocks. The BAYES (weighted priors) analysis suggest small (> 0.01 and < 0.14) contributions from Aru, New Caledonia, GoC, PNG and Micronesia and the mark-recapture data specify contributions from Western Australia, New Caledonia, GoC, PNG, Vanuatu and the Marshal Islands. MSA depends on the relative distinctiveness of the haplotypes present within the stocks and the coverage of all possible contributing stocks, and performs generally well in estimating the probability of turtles coming from specific stocks that are often comprised of several rookeries. Mark-recapture studies however, provide a direct link between individual turtles and specific nesting beaches, but rarely provide an accurate estimate of the proportion of all contributing stocks, especially in remote regions with little monitoring. When used together tagging and genetic studies will complement each other and provide a more robust measure of stock assessment in foraging area studies. 92
103 Latitudinal comparisons The north/south transect of foraging grounds analysed spanned ~2300 km. Along this transect the main contributor shifted from being predominantly the ngbr stock at foraging grounds in Torres Strait, Clack Reef and the Howicks Group to predominantly the sgbr stock at Edgecombe Bay, Shoalwater Bay and Moreton Bay. Similar results have been found using flipper tagging. Limpus et al. (2003) examined the relative proportion of tag recoveries of ngbr and sgbr stocks of green turtles in 1 o latitudinal blocks along the east coast of Australian and showed a consistent trend from ngbr stock prevalence in the north to sgbr stock dominance in the south. The area where the two stocks are equal must be located somewhere between HG and EB and should be seen in the light of major ocean currents affecting the region. The Southern Equatorial Current (SEC) brings water westwards into the Coral Sea and is likely to affect green hatchlings emerging from rookeries in New Caledonia and the Coral Sea Platform. The SEC then divides as it approaches the east Queensland coast at about -18º latitude and brings entrained post-hatchlings either southwards within the East Australian Current (EAC) or northwards within the North Queensland Current (Boyle 2006). This suggests that post-hatchling green turtles emerging from rookeries in the sgbr are carried southward with the flow of the EAC. As the EAC turns away from the Australian east coast, post-hatchlings are carried northward again and remain in the warm water eddies within the Coral and Tasman seas. This is supported by stranding records in southern Queensland and northern New South Wales for all size classes up to the size observed when juveniles recruit back into coastal foraging grounds (Boyle 2006). Hatchlings emerging from the ngbr and any hatchlings carried north with the division of the SEC would be affected by the surface waters in the Gulf of Papua. The currents and eddies affecting the ngbr and Gulf of Papua region are complex and vary both spatially and temporally. Post hatchlings in this region may travel into the Torres Strait, remain within the gyre in the Gulf of Papua, enter the Solomon Sea, or be directed southwards into the EAC (Boyle 2006). Given the results from the MSA and tagging data, the most likely routes for post-hatchlings in the ngbr are to remain within the circulation of the northern Coral Sea (the North Queensland Current), or to enter the Solomon Sea, travelling within numerous gyres in the region until they recruit back into coastal foraging areas. 93
104 This could explain why the southern-most foraging aggregation is stocked almost exclusively by turtles from the sgbr for both sub-adults and juveniles. The ngbr foraging aggregations on the other hand have a more diverse mixture and had a greater number of haplotypes. The presence of haplotype C9 (n = 4) which is found only on Scott Reef and North West Shelf, Western Australia (see Chapter 3) suggests that a small number of turtles embark on longdistance migration events. This is corroborated by mark-recapture data that show records of small contributions from the Coral Sea, New Caledonia, and the Lacepedes, Western Australia (Limpus 2007). The high number of orphan haplotypes limits the MSA and suggests that there are gaps in the coverage of source populations. This is further supported by the mark-recapture data showing contributions from Solomon Island, Vanuatu and the Marshal islands (Limpus 2007). It is likely that sampling of these breeding populations would reveal the origin of the orphan haplotypes found in the GBR foraging aggregations Age class variation At the most northern foraging ground in the Torres Strait, significant shifts in haplotype frequencies between recently sampled juveniles and all sampled adults results in major shifts in the estimated stock contributions for these groups. It appears that fewer juveniles have come from the ngbr stock and higher proportions have come from the sgbr and other stocks. This trend is apparent in the four most northern foraging grounds, including Edgecombe Bay, for which the MSA indicated a predominance of turtles from the sgbr stock. Point estimates of contributions from the ngbr stock dropped from 0.89 in adults to 0.53 in juveniles in the TS, from 0.69 to 0.49 at Clack Reef, 0.66 to 0.49 in the Howicks Group and from 0.10 in adults to 0.01 in juveniles at Edgecombe Bay. In most cases this was countered by increases from both the sgbr stock and those from other stocks, though the estimated contribution of other stocks was too weak to be meaningful for the Howicks Group and Edgecombe Bay. In contrast, at the Shoalwater Bay foraging ground the opposite was observed, with a drop in contribution from the sgbr stock from 0.98 in adults to 0.84 and 0.85 in juveniles and sub-adults respectively, and an increase in contributions from other stocks. At the most southern large foraging ground in Moreton Bay, the only comparison that could be made was between the mark-recapture data from adults and the MSA for sub-adults and juveniles. Here there was little indication of any differences among the groups. The varied patterns were observed at the various foraging grounds likely resulted from several causes and four possible explanations are explored below. These are: 1) there has been a shift 94
105 in haplotype frequencies in the ngbr rookeries over time, 2) juveniles have shifted foraging grounds as they mature, 3) there may be a recent population increase in the sgbr stock, as has been indicated by population modelling and 4) there is evidence to suggest that there has been reduced hatching success from the main ngbr rookery at Raine Island for more than a decade resulting in reduced recruitment into the ngbr foraging ground from this stock. The temporal sampling from the Torres Strait foraging ground provides a test of whether there has been be a shift in haplotype frequencies for the ngbr rookeries over time. Most reference data was sampled in the early 1990s, almost 20 years ago and haplotype profiles of the breeding stock could have changed over that time period. However, absence of difference in the haplotype frequencies from turtles nesting on Raine Island (ngbr) in 1992 and 2008 suggests temporal stability of the nesting populations. This idea has also been supported in other regions including green turtles in Costa Rica (Bjorndal et al. 2005), loggerhead turtles in Japan (Hatase et al. 2002) and hawksbill turtles in Puerto Rico, (Velez-Zuazo et al. 2008). Have juveniles shifted foraging grounds as they mature? In some areas adult and juvenile foraging habitats are geographically separated from each other, and immature turtles may shift through a number of different foraging grounds as they mature (Lutcavage & Musick 1985, Musick & Limpus 1997). However, in regions such as the Great Barrier Reef juvenile and adult foraging habitats are overlapping or coincide (Limpus & Reed 1985, Limpus et al. 1992). Additionally, long-term mark-recapture studies on the Great Barrier Reef suggest that green turtles across all size classes show strong fidelity to their foraging area and have limited movement between foraging grounds (Limpus & Walter 1980, Limpus et al. 1994, Limpus & Chaloupka 1997). Nonetheless, the results from MSA of juvenile and adult turtles support a model in which immature green turtles from several regional stocks recruit into shallow areas along the east coast of Australia. As they mature a small portion of the turtles originating from distant rookeries shift foraging grounds and become residents at foraging grounds closer to their natal rookery. This may occur at numbers too low to allow identification in mark recapture studies, especially for turtles shifting to remote foraging grounds where there is little mark-capture effort. Several other mark-recapture studies have found that immature green turtles may undertake extensive developmental migrations (Bjorndal & Bolten 1997, Musick & Limpus 1997). In the Bahamas, immature green turtles tagged on foraging grounds will remain in the coastal juvenile foraging habitats for several years before emigrating to other foraging areas 95
106 throughout the Greater Caribbean (Bjorndal et al. 2003). In Brazil, some immature green turtles show strong fidelity to their foraging area for extended periods of time while others would exhibit long distance shifts (> 100 km) (Godley et al. 2003). These migrations to different foraging and developmental areas may be in response to differing food abundance and quality and/or population density (Bjorndal et al. 2000). There is some evidence from population modelling that there has been a recent population increase in the sgbr stock (Chaloupka & Limpus 2001), which in part could explain the results. An increase in the sgbr should result in an overall reduction in the relative frequency of all other groups (assuming these are not increasing as well). However, there is not only an increase in the frequency of the sgbr contribution among the juvenile turtles but also an increase in turtles from rookeries outside the GBR (mainly New Caledonia, Aru, GoC and PNG). This suggests that either option 2) or a decrease in ngbr recruitment and a subsequently an increase in the relative frequency of turtles from other rookeries are the more likely hypotheses. It has been strongly suggested that flooding of nests has significantly contributed to reduced hatching success on Raine Island over the last 15 years (Limpus et al. 2000). Flooding of nests from a rise in the water table following heavy rain, large tides or from storm surges have all contributed to this problem. This has resulted in nests being flooded in the lower laying areas of the beach. In February 1997, 93 holes were dug within each habitat type around the island to determine the impact of the flooding on egg development and hatching success (Limpus et al. 2000). All nests sampled within these sites indicated that no eggs had survived the inundation of flooding. Since 1996, flooding of much of the nesting habitat has been a regular event and it is likely that during wet years with broad scale flooding, a very low percentage of nests on Raine Island would produce hatchlings. This supports the pattern observed at the foraging grounds and could be explain the reduction in recruitment of ngbr juveniles at the ngbr foraging grounds Conservation implications The Great Barrier Reef supports some of the largest nesting and foraging populations of green turtles in the world. Understanding how these foraging grounds are connected to regional rookeries is a vital aspect of implementing effective management plans. This study suggests 96
107 that using both conventional mark-recapture programs and MSA together could be a useful management tool for migratory species that present challenges for managers due to spatial and temporal constraints. Historically, mark-recapture studies have revealed the migrations of many adult green turtles that use foraging areas along the east coast of Australia. In this study, the combination of MSA and mark-recapture data show a shift in contribution from the two GBR breeding stocks along a north-south transect. In the north (Torres Strait, Clack Reef and the Howicks Group), foraging adults are principally coming from the ngbr rookeries whereas in the south (Moreton Bay, Shoalwater Bay and Edgecombe Bay) foraging adult turtles are principally coming from sgbr rookeries, demonstrating that the majority of turtles chose to forage within 600 km of their natal rookery. The shift in composition between juveniles and adults is likely the result of either juvenile developmental migration, where juveniles from distant stocks shift to more local foraging areas as they reach adulthood, or a reduced hatching success at Raine Island. These are complex interactions and at present there is insufficient data to determine the relative importance of these two hypotheses. If reduced hatching success at Raine Island were the main force driving the observed pattern, significant changes in the impact of anthropogenic mortality on regional rookeries foraging at ngbr foraging grounds could be expected in the future. Reductions in the numbers of ngbr turtles leads to increases in the relative frequencies of sgbr and other rookeries at ngbr foraging aggregations. Under this scenario, impacts on both ngbr and sgbr rookeries could be expected to increase during the next 20 years, emphasising the need for long term management strategies and continuation of population monitoring at both foraging and nesting sites. Although the sgbr population may be increasing, (Chaloupka & Limpus 2001), results of modelling the population dynamics of this stock indicate that it is quite susceptible to the harvest of adult turtles (Chaloupka 2004), The combined strength of data derived from mark-recapture studies, demographic studies to determine sex, maturity and breeding status of the turtles, genetic studies to determine stock composition and satellite telemetry, are needed to provide informed assessments of foraging populations necessary for guiding sustainable management of marine turtles. 97
108 CHAPTER 6 6. Synopsis. Green turtles have been globally listed as endangered according to IUCN criteria (IUCN, 2009). In Australia, the green turtle is listed as vulnerable under the Commonwealth legislation (Environment Protection and Biodiversity Conservation Act 1999). The Recovery Plan for Marine Turtles in Australia (Australia, 2003) describes threats to Australian populations and provides objectives to secure their survival. A set of action plans have been formed that aim to reduce detrimental impacts to populations and to promote the perseverance and recovery of Australian populations of marine turtles. One aim of this action plan is the genetic identification of Australian marine turtle populations in support of the development of programs and protocols to monitor marine turtle populations in Australian waters, assess the size and status of those populations and the causes of their mortality, and address information gaps. The identification of genetically differentiated populations serves two purposes. First, it allows us to identify the populations that are genetically isolated from each other. These stocks reflect groupings of turtles that have existed since the recent past and for which limited genetic exchange occurs. Thus, little migration of individuals occurs among these Management Units (MUs; Moritz 1998) that would allow for compensation of mortality occurring at a particular MU. Therefore, MUs should receive specific management attention if under threat. Designation of the geographic boundaries of rookeries that comprise MU is important for conservation planning, but this alone does not take into account the full scope of the marine turtle habitat used by migratory and foraging animals within each MU. Therefore management of marine turtles must take into account the entire geographical range of the MU, including foraging grounds used by those turtles. By characterising breeding stocks using mitochondrial (mt)dna sequence data as genetic markers, the origin of turtles sampled away from their nesting habitat and be determined using mixed stock analysis (MSA) (Pella & Masouda 2001). By determining the proportional stock contributions to foraging grounds 98
109 mortality at the foraging grounds can be attributed to each MU. These studies not only provide valuable information about migration pathways and the geographical range of stocks but also in identifying nesting populations likely to be impacted by possible threats at the foraging grounds. In this study I have confirmed the previous designations of green turtle MUs in Australasia (Moritz et al. 2002, Dethmers et al. 2006) using increased sample sizes and increased genetic resolution through the sequencing of a longer fragment of the mtdna for some MUs. Through sampling additional rookeries, two of these have been designated to existing MUs and two appear to be unique MUs. This has led to increased attention being given to these rookeries for further studies and sampling. This baseline information on the genetic structure among MUs has then been used to conduct mixed stock analyses of foraging grounds off Western Australia, Malaysia and along a transect running the length of the Great Barrier Reef. In order to improve estimates from MSA I used a 780 bp sequence from the mtdna control region as genetic marker to assess whether this would add resolution to the genetic structure of the Australasian region. Application of this marker to the analysis of green turtle population genetic structure across three regions allowed a thorough assessment of the longer sequence. This also provided reference data for this and future analysis of foraging grounds in the region. Use of the longer sequence did provide greater resolution of genetic structure among some populations and also improved some MSA estimates, but this impact was variable. The variation in genetic structure, in terms of the extent to which there were common and widespread haplotypes among the breeding populations, and sample sizes had a large influence on the robustness of MSA estimates. Source populations for the Shark Bay foraging ground in Western Australia were well characterised but affected by shared haplotypes. In Malaysia there was also the problem of shared haplotypes as well as limited sampling of source populations, especially towards the north. In contrast, the potential source populations for Great Barrier Reef foraging grounds displayed a high genetic differentiation (Dethmers et al. 2006, Chapter 5). These contrasts in the study areas highlighted the limitations of using mixed stock analysis (e.g., Chapter 3). But they also showed that even small increases in genetic structure among populations found when using the longer sequence, 99
110 which uncovered variation within the common haplotypes, can improve MSA estimates as was seen in the Malaysian foraging grounds. The relatively high precision in the MSA analyses for GBR foraging grounds was enabled by the genetic structure among the contributing stocks and extensive sampling. Importantly, traditional mark-recapture data could be used to help select the most appropriate statistical approach to use with the MSA. This study revealed differences in the contributions of source populations to the juveniles versus adults (and/or sub-adults) components of the foraging aggregations. This was an unexpected result that demonstrates the operation of dynamic processes. It is possible that developmental migration is occurring on a larger scale than indicated by tagging data (Limpus 2007), in which maturing turtles migrate to foraging grounds closer to their natal regions. Alternatively, and of great concern, is that the data also suggest that there has been a decrease in the proportion of ngbr juvenile turtles recruiting to the Great Barrier Reef foraging grounds. There is support for this hypothesis from observations of a lack of hatchling emergence from one of the world s largest green turtle rookeries at Raine Island in the ngbr (Limpus et al. 2003). From a management perspective, this later hypothesis must be considered and further tested with field studies or we may witness a severe decline in the population in the future. Conservation management and implications The study of immature green turtles in Malaysia highlighted how MSA can be a powerful tool to monitor juvenile foraging aggregations by helping to detect early signs of demographic change. In this case, MSA was used to demonstrate a link between increased sex-bias at foraging grounds with poor hatchery practices at the source populations, and to highlight a major conservation concern for these populations. Similarly, the comparison of juvenile versus adult (and/or) sub adult turtles in the GBR foraging grounds demonstrated demographic changes in the source populations. In the GBR this may in part be occurring through undocumented developmental migrations, or more seriously, from a prolonged decrease in juvenile recruitment from the ngbr rookeries. These results are ground breaking in how they have provided a window into the future for the regional rookeries, by seeing decades ahead. For the GBR populations, in addition to a possible decline in the future numbers of nesting turtles, if the observed trends in stock contribution continues then Indigenous harvest of turtles in the ngbr will begin to harvest proportionally more turtles from the sgbr. This may significantly increase the impact on the sgbr population in the 100
111 future. Biologists and managers must be able to identify and monitor these changes across all age classes to effectively plan for ensuring survival of the populations. Considerable interest has been generated by this research and it has already caused increased concern for these populations. Further work is in progress to incorporate these data on stock contributions and juvenile and adult dispersal to develop spatially explicit matrix population models to evaluate the impact of harvest and other threats on long-term trends in population change. The model will be used to evaluate different management scenarios and thereby provide detailed, stock-specific management recommendations to local communities to promote effective conservation of Australian green turtles. Future research Demographic studies of foraging grounds and genetic sampling should be continued to monitor temporal variation as found in the ngbr. The study of green turtles along the Great Barrier Reef highlights the power of these methods, especially when combined with longterm demographic, mark-recapture and satellite data. The complete stock structure including boundaries of distinct populations needs to be understood, not only for adult turtles but for all age classes. Without this knowledge the effects of threats on nesting beaches, foraging grounds and migratory corridors cannot be evaluated for individual populations. Effort should be made to increase the sampling of rookeries, especially from northern SE Asia and from the South Pacific. For some analyses, increasing genetic resolution could be helped by resequencing individuals from potential contributing stocks that have the common haplotype, using the longer sequence. Alternatively, the application of a diversity of genetic markers may provide increased resolution. Application of nuclear microsatellites to investigate the stock contributions to green turtle foraging grounds in the Northern Territory using assignment-based methods (Cornuet et al. 1999) were problematic as there was poor self-assignment of nesting turtles to their rookeries (McCann 2003). There is a continuous need for new methods as limitations in previous ones come to light. A relatively new set of markers, known as Single Nucleotide Polymorphisms (SNPs), are rapidly becoming the marker of choice for many applications in population ecology, evolution and conservation genetics (Morin et al. 2004). SNPs result from single basepair mutations and they are more frequent in the genome than microsatellites. This allows for the discovery and screening of much larger number of loci. They offer more 101
112 rapid and accurate data than microsatellites. Recently, the first set of SNP markers were developed for marine turtles (Roden et al. 2009a, 2009b). Preliminary results are promising and their utility as a population diagnostic marker appears to be at least equal to that of microsatellite markers. The use of multiple SNP markers is expected to provide more detailed information because they allow for sampling from many more regions of an organisms genome. If an ongoing trial of SNP utility in Australian green turtles is promising, then we will be the first to use this novel genetic technique to address the important issue of anthropogenic mortality of marine turtles. Modelling Even with all the considerable effort required to obtain this genetic information, the stock identification and proportional contributions from stocks at foraging grounds needs to be incorporated into population models that include parameters related to breeding grounds as well as foraging grounds. Creating population models that can reliably assess the potential effects of harvest on specific populations can provide the information needed to bring together Indigenous communities, managers, biologists and politicians to prioritise needs and attempt workable solutions. Because many marine turtle species are an important cultural heritage with respect to Aboriginal and Torres Straight Islanders customary rights of harvest, the goals to preserve this precious resource in a sustainable manner are of primary importance for Australia. The genetic data from this project in combination with existing genetic data, data from markrecapture studies and over 30 years of life history data of the Great Barrier Reef (GBR) populations will be used to develop spatially explicit matrix population models to evaluate the impact of harvest and other threats on long-term rates of population change. Although some data have been collected on these harvests, few attempts have been made to describe quantitatively the level of harvest. This is an important area of future research. It is essential that data on mortality in various habitats are included in demographic models and management plans of both northern and southern GBR populations. Although this future research takes a broad regional approach, the results will provide specific, local management recommendations based on the combined genetic and field data. 102
113 REFERENCES Abreu-Grobois F.A., Horrocks, J.A., Formia A., Dutton P., Leroux R., Velez-Zuazo X., Soares L., & Meylan (2006) New mtdna D-loop primers which work for a variety of marine turtle species may increase the resolution of mixed stock analysis. In: Frick M., A. Panagopoulous, A. F. Rees & K. Williams (eds.) Proceedings of the 26 th Annual Symposium on Sea Turtle Biology and Conservation. International Sea Turtle Society, Athens, Greece, Available from /abreu_grobois_etal_new_dloop_primers.pdf. Ackerman, R.A. (1997) The Nest Environment and the Embryonic Development of Sea Turtles. In The Biology of Sea Turtles, eds. P.L. Lutz and J.A. Musick, pp CRC Marine Science Series, CRC Press, Inc., Boca Raton, Florida. Adnyana, W., Soede, L.P., Gearheart, G. & Halim, M. (2008) Status of green turtle (Chelonia mydas) nesting and foraging populations of Berau, East Kalimantan, Indonesia, including results from tagging and telemetry. Indian Ocean Turtle Newsletter 7:2-11. Allard, M.W., Miyamoto, M.M., Bjorndal, K.A., and Bowen, B.W. (1994) "Support for natal homing in Green turtles from mitochondrial DNA sequences." Copeia 1994: Avens, L. & Lohmann, K.J. (2004) Navigation and seasonal migratory orientation in juvenile sea turtles. Journal of Experimental Biology 207: Balazs, G.H., Forsyth, R.G. & Kam, A.K.H. (1987) Preliminary assessment of habitat utilization by Hawaiian green turtles in their resident foraging pastures. NOAA Tech Memo. NOAA-TM-NMFS-SWFC-71. Basintal, P. & Lakim, M. (1993) Status and management of sea turtles at Turtle Islands Park. In A. Nacu, R. Trono, J.A. Palma, D. Torres and F Agas Jr. (eds.), Proceedings of the First ASEAN Symposium Workshop on Marine Turtle Conservation. pp WWF-USAID, Manila, Philippines. Bass, A.L., Epperly, S.P. & Braun-McNeill, J. (2006) Green turtle (Chelonia mydas) foraging and nesting aggregations in the Caribbean and Atlantic: impact of currents and behavior on dispersal. Journal of Heredity 97:346. Bass, A.L. (1999) Genetic analysis to elucidate the natural history and behavior of hawksbill turtles (Eretmochelys imbricata) in the wider Caribbean: a review and re-analysis. Chelonian Conservation and Biology Bass, A.L., Epperly, S.P. & Braun-McNeill, J. (2004) Multi-year analysis of stock composition of a loggerhead turtle (Caretta caretta) foraging habitat using maximum likelihood and Bayesian methods. Conservation Genetics 5: Bass, A.L., Lagueux, C.J. & Bowen, B.W. (1998) Origin of green turtles, Chelonia mydas, at "Sleeping Rocks" off the northeast coast of Nicaragua. Copeia
114 Bass, A.L. & Witzell, W.N. (2000) Demographic Composition of Immature Green Turtles (Chelonia mydas) from the East Central Florida Coast: Evidence from mtdna Markers. Herpetologica 56: Bentivegna, F. (2002) Intra-Mediterranean migrations of loggerhead sea turtles (Caretta caretta) monitored by satellite telemetry. Marine biology 141: Bjorndal, K.A., Bolten, A.B. & Chaloupka, M.Y. (2003) Survival probability estimates for immature green turtles Chelonia mydas in the Bahamas. Marine Ecology Progress Series 252: Bjorndal, K.A., Bolten, A.B. & Chaloupka, M.Y. (2005) Evaluating trends in abundance of immature green turtles, Chelonia mydas, in the greater Caribbean. Ecological Applications 15: Bjorndal, K.A. & Bolten, A.B. (1997) Developmental migrations of juvenile green turtles in the Bahamas. in J. A. Keinath, D. E. Barnard, J. A. Musick, and B. A. Bell, comps. Proc. 15th Ann.. Symp. Sea Turtle Biol. Conserv. NOAA Tech. Memo., NMFS- SEFSC-387. pp Bjorndal, K.A., Bolten, A.B. & Martins, H.R. (2000) Somatic growth model of juvenile loggerhead sea turtles Caretta caretta: duration of pelagic stage. Marine Ecology Progress Series 202: Bjorndal, K.A. & Bolten, A.B. (2008) Annual variation in source contributions to a mixed stock: implications for quantifying connectivity. Molecular Ecology 17: Bjorndal, K.A., Bolten, A.B., Moreira, L., Bellini, C. & Marcovaldi, M.Â. (2006) Population Structure and Diversity of Brazilian Green Turtle Rookeries Based on Mitochondrial DNA Sequences. Chelonian Conservation and Biology 5:262. Bjorndal, K.A., Bolten, A.B. & Troëng, S. (2005) Population structure and genetic diversity in green turtles nesting at Tortuguero, Costa Rica, based on mitochondrial DNA control region sequences. Marine Biology 147: Blumenthal, J.M., Austin, T.J., Bell, C.D.L., Bothwell, J.B., Broderick, A.C., Ebanks-Petrie, G., Gibb, J.A., Luke, K.E., Olynik, J.R., Orr, M.F., Solomon, J.L. & Godley, B.J. (2009) Ecology of Hawksbill Turtles, Eretmochelys imbricata, on a Western Caribbean Foraging Ground. Chelonian Conservation and Biology 8:1-10. Blumenthal, J., Abreu-Grobois, F., Austin, T., Broderick, A., Bruford, M., Coyne, M., Ebanks-Petrie, G., Formia, A., Meylan, P., Meylan, A. & Godley, B. (2009) Turtle groups or turtle soup: dispersal patterns of hawksbill turtles in the Caribbean. Molecular Ecology 18: Bolker, B.M., Okuyama, T., Bjorndal, K.A. & Bolten, A.B. (2007) Incorporating multiple mixed stocks in mixed stock analysis: 'many-to-many' analyses. Molecular Ecology 16:
115 Bolten, A.B., Bjorndal, K.A., Martins, H.R. & al., E. (1998) Transatlantic developmental migrations of loggerhead sea turtles demonstrated by mtdna sequence analysis. Ecological Applications 8:1-7. Bourjea, J., LapèGue, S., Gagnevin, L., Broderick, D., Mortimer, J.A., Ciccione, S., Roos, D., Taquet, C. & Grisel, H. (2007) Phylogeography of the green turtle, Chelonia mydas, in the Southwest Indian Ocean. Molecular Ecology 16: Bowen, B.W., Abreu-Grobois, F.A., Balazs, G.H., Kamezaki, N., Limpus, C.J. & Ferl, R.J. (1995) Trans-Pacific migrations of the loggerhead turtle (Caretta caretta) demonstrated with mitochondrial DNA markers. Proc Natl Acad Sci U S A 92: Bowen, B.W., Avise, J.C., Richardson, J.I., Meylan, A.B., Margaritoulis, D.P. & R., H.S. (1993) Population structure of loggerhead turtles (Caretta caretta) in the Northwestern Atlantic ocean and Mediterranean sea. Conservation Biology 7: Bowen, B.W., Bass, A.L., Garcia-Rodriguez, A., Diez, C.E., Van Dam, R.P., Bolten, A.B., Bjorndal, K.A., Miyamoto, M.M. & Ferl, R.J. (1996) Origin of hawksbill turtles in a Caribbean feeding area as indicated by genetic markers. Ecological Applications 6: Bowen, B.W., Clark, A.M., Abreu-Grobois, F.A., Chaves, A., Reichart, H.A. & Ferl, R.J. (1998) Global Phylogeography of the ridley sea turtles (Lepidochelys spp) as inferred from mitochondrial DNA sequences. Genetica 101: Bowen, B.W., Grant, W.S., Hillis-Starr, Z., Shaver, D.J., Bjorndal, K.A., Bolten, A.B. & Bass, A.L. (2007) Mixed-stock analysis reveals the migrations of juvenile hawksbill turtles (Eretmochelys imbricata) in the Caribbean Sea. Molecular Ecology 16: Bowen, B.W. & Karl, S.A. (1996) Population structure, Phylogeography, and molecular evolution. The Biology of Sea Turtles. pp CRC Press, Boca Raton, FL. Bowen, B.W. & Karl, S.A. (2007) Population genetics and phylogeography of sea turtles. Molecular Ecology 16: Bowen, B.W., Bass, A.L., Chow, S., Bostrom, M., Bjorndal, K.A., Bolten, A.B., Okuyama, T., Bolker, B.M., Epperly, S., Lacasella, E., Shaver, D., Dodd, M., Hopkins-Murphy, S.R., Musick, J.A., Swingle, M., Rankin-Baransky, K., Teas, W., Witzell, W.N. & Dutton, P.H. (2004) Natal homing in juvenile loggerhead turtles (Caretta caretta). Molecular Ecology 13: Bowen, B.W., Meylan, A.B., Ross, J.P., Limpus, C.J., Balazs, G.H. & Avise, J.C. (1992) Global Population Structure and Natural History of the Green Turtle (Chelonia mydas) in Terms of Matriarchal Phylogeny. Evolution 46: Boyle, M., FitzSimmons, N., Limpus, C., Kelez, S., Velez-Zuazo, X. & Waycott, M. (2009) Evidence for transoceanic migrations by loggerhead sea turtles in the southern Pacific Ocean. Proceedings of the Royal Society B: Biological Sciences 276:
116 Boyle, M. (2006) Post-hatchling sea turtle biology. PhD thesis. James Cook University. Broderick, D., Moritz, C., Miller, J.D., Guinea, M., Prince, R.I.T. & Limpus, C...J... (1994) Genetic studies of the hawksbill turtle Eretmochelys imbricata: evidence for multiple stocks in Australian waters. Pacific Conservation Biology 1: Browne, D., Horrocks, J. & Abreu-Grobois, F. (2009) Population subdivision in hawksbill turtles nesting on Barbados, West Indies, determined from mitochondrial DNA control region sequences. Conservation Genetics DOI: /s Campbell, L.M., Silver, J.J., Gray, N.J., Ranger, S., Broderick, A., Fisher, T., Godfrey, M.H., Gore, S., Jeffers, J., Martin, C., McGowan, A., Richardson, P., Sasso, C., Slade, L. & Godley, B. (2009) Co-management of sea turtle fisheries: Biogeography versus geopolitics. Marine Policy 33: Carr, A.F. (1967) So excellent a fishe: a natural history of sea turtles. Scribner, New York, New York. Carr, A.F. (1987) New perspectives on the pelagic stage of sea turtle development. Conservation Biology 1: Carr, A.F., Carr, M.F. & Meylan, A.B. (1978) The ecology and migrations of sea turtles, 7. The West Caribbean green turtle colony. Bulletin of the American Museum of Natural History, 162. Carreras, C., Pascual, M., Cardona, L., Aguilar, A., Margaritoulis, D., Rees, A., Turkozan, O., Levy, Y., Gasith, A., Aureggi, M. & Khalil, M. (2007) The genetic structure of the loggerhead sea turtle ( Caretta caretta ) in the Mediterranean as revealed by nuclear and mitochondrial DNA and its conservation implications. Conservation Genetics 8: Carreras, C., Pont, S., Maffucci, F., Pascual, M., Barceló, A., Bentivegna, F., Cardona, L., Alegre, F., SanFélix, M., Fernández, G. & Aguilar, A. (2006) Genetic structuring of immature loggerhead sea turtles (Caretta caretta) in the Mediterranean Sea reflects water circulation patterns. Marine Biology 149: Casale, P., Freggi, D., Gratton, P., Argano, R. & Oliverio, M. (2008) Mitochondrial DNA reveals regional and interregional importance of the central Mediterranean African shelf for loggerhead sea turtles (Caretta caretta). Scientia Marina 72: McCann, M.J. (2002) Identifying the origins of green turtles (Chelonia mydas) feeding in northern Australia from genetic markers. Honours Thesis, University of Canberra. Chaloupka, M. (2004) Southern Great Barrier Reef green sea turtle (Chelonia mydas) stock: consequences of local sex-biased harvesting. Species Conservation and Management: Case Studies. pp Oxford University Press. Chaloupka, M. & Limpus, C. (2001) Trends in the abundance of sea turtles resident in southern Great Barrier Reef waters. Biological Conservation 102:
117 Chaloupka, M., Limpus, C. & Miller, J. (2004) Green turtle somatic growth dynamics in a spatially disjunct Great Barrier Reef metapopulation. Coral Reefs 23: Chaloupka, M. (2002) Stochastic simulation modelling of southern Great Barrier Reef green turtle population dynamics. Ecological modelling 148: Chaloupka, M.Y. & Limpus, C.J. (2002) Survival probability estimates for the endangered loggerhead sea turtle resident in Southern Great Barrier Reef waters. Marine Biology 140: Chaloupka, M. & Limpus, C. (1997) Robust statistical modelling of hawksbill sea turtle growth rates (southern Great Barrier Reef). Marine Ecology Progress Series 146:1-8. Chassin-Noria, O. (2005) Paternity studies on sea turtles, old questions, new information required. Oral presentation at the 25th Sea Turtle Symposium, Savannah, Georgia USA January Cheng, I., Huang, C.T., Hung, P.Y., Ke, B.Z., Kuo, C.W. & Fong, C.L. (2009) Ten years of monitoring the nesting ecology of the green turtle, Chelonia mydas, on Lanyu (Orchid Island), Taiwan. Zoological Studies 48: Clement, M., Posada, D. & Crandall, K.A. (2000) TCS: a computer program to estimate gene genealogies. Molecular Ecology Cornelius S.E, and Robinson DC (1986) Post-nesting movements of female olive ridley turtles tagged in Costa Rica. Vida Silvestre Neotropical 1: Cornuet, J.M., Piry, S., Luikart, G., Estoup, A.& Solignac, M. (1999) New methods employing multi locus genotypes to select or exclude populations as origins of individuals. Genetics 153: Craig, P., Parker, D., Brainard, R., Rice, M. & Balazs, G. (2004) Migrations of green turtles in the central South Pacific. Biological Conservation 116: Crandall, K.A. & Tempelton, A.R. (1993) Empirical tests of some predictions from coalescent theory with applications to intraspecific phylogeny reconstruction. Genetics 134: Dethmers, K. (2010) Ecology and Phylogeography of an Australasian green turtle population: a case study for conservation from Aru.PhD Thesis, Radboud University, Holland. Dethmers, K., Broderick, D., Moritz, C., Fitzsimmons, N., Limpus, C., Lavery, S., Whiting, S., Guinea, M., Prince, R. & Kennett, R. (2006) The genetic structure of Australasian green turtles (Chelonia mydas): exploring the geographical scale of genetic exchange. Molecular Ecology 15:
118 Diaz-Fernandez, R., Okayama, T., Uchiyama, T., Carrillo, E., Espinosa, G., Marquez, R., Diez, C. & Koike, H. (1999) genetic sourcing for the hawksbill turtle, Eretmochelys imbricata, in the northern Caribbean region. Chelonian Conservation and Biology 3: Diez, C.E. & Van Dam, R.P. (2003) Sex Ratio of an Immature Hawksbill Sea turtle Aggregation at Mona Island, Puerto Rico. Journal of Herpetology 37: Donnelly, P. & Tavare, S. (1996) The ages of alleles and a coalescent. Adv. Appl. Probab.18. Dutton, P.H., Balazs, G.H., LeRoux, R.A., Murakawa, S.K.K., Zarate, P. & Martínez, L.S. (2008) Composition of Hawaiian green turtle foraging aggregations: mtdna evidence for a distinct regional population. Endangered species research 5: Encalada, S.E., Lahanas, P.N., Bjorndal, K.A., Bolker, B.M., Miyamoto, M.M. & Bowen, B.W. (1996) Phylogeography and population structure of the Atlantic and Mediterranean green turtle Chelonia mydas: a mitochondrial DNA control region sequence assessment. Molecular Ecology 5: Engstrom, T.N., Meylan, P.A. & Meylan, A.B. (2002) Origin of juvenile loggerhead turtles (Caretta caretta) in a tropical developmental habitat in Caribbean Panama. Animal Conservation 5: Excoffier, L. & Langaney, A. (1989) Origin and differentiation of human mitochondrial DNA. Am. J. Hum. Genetics 44: Excoffier, L., Laval, G. & Schneider, S. (2005) Arlequin ver. 3.0: An integrated software package for population genetics data analysis. Evolutionary Bioinformatics Online 1: FitzSimmons, N.N. (1998) Single paternity of clutches and sperm storage in the promiscuous green turtle (Chelonia mydas). Molecular Ecology 7: FitzSimmons, N.N., Moritz, C., Limpus, C.J., Pope, L. & Prince, R. (1997a) Geographic structure of the mitochondrial and nuclear gene polymorphisms in Australian green turtle populations and male-biased gene flow. Genetics 147: FitzSimmons, N., Limpus, C., Norman, J., Goldizen, A., Miller, J. & Moritz, C. (1997b) Philopatry of male marine turtles inferred from mitochondrial DNA markers. Proceedings of the National Academy of Sciences of the United States of America 94: FitzSimmons, N., Moritz, C. & Moore, S. (1995) Conservation and dynamics of microsatellite loci over 300 million years of marine turtle evolution. Molecular Biology and Evolution 12: Formia, A., Godley, B., Dontaine, J. & Bruford, M. (2006) Mitochondrial DNA diversity and Phylogeography of endangered green turtle (Chelonia mydas) populations in Africa. Conservation Genetics 7:
119 Fuentes, M., Hamann, M. & Lukoschek, V. ( 2009). Marine Reptiles and Climate Change. A Marine Climate Change Impacts and Adaptation Report Card for Australia Fuentes, M., Hamann, M. & Limpus, C.J. (2010) Vulnerability of sea turtle nesting grounds to climate change. Global Change Biology. DOI /j x Gelman, A. & Rubin, D.B. (1992) Inference from iterative simulation using multiple sequences. Statistical Science 7: Godley, B.J., Lima, E., Åkesson, S., Broderick, A.C., Glen, F., Godfrey, M.H., Luschi, P. & Hays, G.C. (2003) Movement patterns of green turtles in Brazilian coastal waters described by satellite tracking and flipper tagging. Marine Ecology Progress Series 253: Godley, B.J., Richardson, S., Broderick, A.C., Coyne, M.S., Glen, F. & Hays, G.C. (2002) Long-term satellite telemetry of the movements and habitat utilisation by green turtles in the Mediterranean. Ecography 25: Golding, G.B. (1987) The detection of deleterious selection using ancestors inferred from a evolutionary history. Genetic Research 49: Grant W.S., Milner, G.B., Krasnowski P. & Utter F. M. (1980) Use of biochemical genetic variation for identification of sockeye salmon (Oncorrhynchus nerka) stocks in Cook inlet, Alaska. Can. J. Fish. Aquat. Sci. 37: Grossman, A., Bellini, C., Fallabrino, A., Formia, A., Mba, J.M. & Obama, C. (2007) Second TAMAR-tagged hawksbill recaptured in Corisco Bay, West Africa. Marine Turtle Newsletter 116:26. Hamabata, T., Nishida, S., Kamezaki, N. & Koike, H. (2009) Genetic structure of populations of the green turtle (Chelonia mydas) in Japan using mtdna control region sequences. Bulletin of the graduate school of social and cultural studies, Kyushu University 15: Hamann, M., Limpus, C.J. & Read, M.A. (2007) Vulnerability of marine reptiles in the Great Barrier Reef to climate change. In, Climate change and the Great Barrier Reef. Great Barrier Reef Marine Park Authority and The Australian Greenhouse Office. Hamann, M., Cuong, C.T., Hong, N.D., Thuoc, P. & Thuhien, B.T. (2006) Distribution and abundance of marine turtles in the Socialist Republic of Viet Nam. Biodiversity and Conservation 15: Hatase H., Kinoshita M., Bando T., Kamezaki N., Sato K., Y., M., K., G., K., O., Y., N., Takeshita H. & Sakamoto W. (2002) Population structure of loggerhead turtles, Caretta caretta, nesting in Japan: bottlenecks on the Pacific population. Marine Biology 141: Hawkes, L., Broderick, A., Godfrey, M. & Godley, B. (2009) Climate change and marine turtles. Endangered Species Research 7:
120 Hays G., Broderick A., Glen F., Godley B. & Nichols W. (2001) The movements and submergence behaviour of male green turtles at Ascension Island. Marine Biology 139: Henwood, T.A. & Ogren, L.H. (1987) Distribution and migration of immature Kemp's ridley turtles (Lepidochelys kempi) and green turtles (Chelonia mydas) off Florida, Georgia and South Carolina. Northeast Gulf Science, 9, 153. IUCN. (2009) IUCN Red List of Threatened Species. Version Downloaded on 01 March Jensen, M.P., Abreu-Grobois, F.A., Frydenberg, J. & Loeschcke, V. (2006) Microsatellites provide insight into contrasting mating patterns in arribada vs. non-arribada olive ridley sea turtle rookeries. Molecular Ecology 15: Kalb, H., Valverde, R., & Owens, D. W. (1992) What is the reproductive patch of the olive ridley sea turtle? In Proceedings of the 12th Annual Workshop on Sea Turtle Conservation and Biology. Jekyll Island, Georgia pp Lahanas, P.N., Bjorndal, K.A., Bolten, A.B., Encalada, S.E., Miyamoto, M.M., Valverde, R.A. & Bowen, B.W. (1998) Genetic composition of a green turtle (Chelonia mydas) feeding ground population: evidence for multiple origins. Marine Biology 130: Lanyon, J. M., Limpus, C.M. & Marsh, H. (1989) Dugongs and turtles grazers in the seagrass system. In. Biology of seagrasses. A.W.D. Larkun, A.J. McComb & S.A. Shepherd. Elsevier, New York, Pp Laurent, L., Casale, P., Bradai, M.N., Godley, B.J., Gerosa, G., Broderick, A.C., Schroth, W., Schierwater, B., Levy, A.M., Freggi, D., Abd el-mawla, E.M., Hadoud, D.A., Gomati, H.E., Domingo, M., Hadjichristophorou, M., Kornaraky, L., Demirayak, F. & Gautier, C. (1998) Molecular resolution of marine turtle stock composition in fishery bycatch: a case study in the Mediterranean. Molecular Ecology 7: Leh, P. (1993) The 1992 green turtle conservation program at the Sarawak Turtle Islands. In A. Nacu, R. Trono, J.A. Palma, D. Torres and F Agas Jr. (eds.), Proceedings of the First ASEAN Symposium Workshop on Marine Turtle Conservation. pp WWF-USAID, Manila, Philippines. Limpus, C. (1992) The hawksbill turtle, Eretmochelys imbricata, in Queensland: population structure within a southern Great Barrier Reef feeding ground. Wildlife Research 19: Limpus, C.J. (1993) The green turtle, Chelonia mydas, in Queensland - breeding males in the southern Great Barrier Reef. Wildlife Research 20: Limpus, C.J. (2007) A biological review of Australian marine turtles: 2. Green turtle (Chelonia mydas). Flatback turtle, Natator depressus. 110
121 Limpus, C. J. (2008) A biological review of Australian marine turtles: 3. Hawksbill Turtle (Eretmochelys imbricata) (Linnaeus). Limpus, C. & Chaloupka, M. (1997) Nonparametric regression modelling of green sea turtle growth rates (southern Great Barrier Reef). Marine Ecology Progress Series 149: Limpus, C. & Nicholls, N. (1988) The southern oscillation regulates the annual numbers of green turtles (Chelonia mydas) breeding around northern Australia. Australian wildlife research 15: Limpus C.J. & Reed, P. (1985a) The loggerhead turtle, Caretta caretta, in Queensland observations on internesting behavior. Australian wildlife research 12: Limpus, C.J. & Reed, P.C. (1985b) The green turtle, Chelonia mydas, in Queensland: A preliminary description of the population structure in a coral reef feeding ground. Biology of Australasian frogs and reptiles, ed. by Gordon Grigg, Richard Shine and Harry Ehmann. pp Royal Zoological Society of New South Wales. Limpus, C.J. & Walter, D.G. (1980) The growth of immature green turtles (Chelonia mydas) under natural conditions. Herpetologica 36: Limpus, C.J., Miller, J., Parmenter, C., Reimer, D., McLachlan, N. & Webb, R. (1992) Migration of green (Chelonia mydas) and loggerhead (Caretta caretta) turtles to and from eastern Australian rookeries. Wildlife Research 19: Limpus, C.J., Couper, P.J. & Read, M.A. (1994) The green turtle, Chelonia mydas, in Queensland: population structure in a warm temperate feeding area. Memoirs of the Queensland museum 35: Limpus, C.J., Al-Ghais, S.M., Mortimer, J.A. & Pilcher, N.J. (2001) Marine turtles in the Indian Ocean and Southeast Asian region: Breeding, distribution, migration and population trends. Convention on Migratory Species, Manila, Philippines. Limpus, C. J., Miller, J.D., Parmenter, C.J. & Limpus, D.J. (2000) The green turtle, Chelonia mydas, population of Raine Island and the Northern Great Barrier Reef: Memoirs of the Queensland museum 49: Limpus, C.J., Bell, I. & Miller, J.D. (2009) Mixed stocks of green turtles foraging on Clack Reef, Northern Great Barrier Reef identified from long term tagging studies. Marine Turtle Newsletter 123:3-5. Luke, K., Horrocks, J.A., LeRoux, R.A. & Dutton, P.H. (2004) Origins of green turtle ( Chelonia mydas ) feeding aggregations around Barbados, West Indies. Marine Biology 144: Lutcavage, M.E., Plotkin, P.T., Witherington, B.E. & Lutz, P.L. (1996) Human impacts on sea turtle survival. The Biology of Sea Turtles (eds. Lutz P.L., Musick J.A.). p CRC Press, Boca Raton, FL. 111
122 Lutcavage, M. & Musick, J.A. (1985) Aspects of the Biology of Sea Turtles in Virginia. Copeia, Lukoscheck V., Funahashi N., Lavery S., Dalebout M.L., Cipriano F., Baker C.S. (2009) High proportion of protected minke whale sold on Japanese markets is due to illegal, unreported or unregulated exploitation. Animal Conservation 12: Maffucci, F., Kooistra, W. & Bentivegna, F. (2006) Natal origin of loggerhead turtles, Caretta caretta, in the neritic habitat off the Italian coasts, Central Mediterranean. Biological Conservation 127: Márquez, M.R. (1990) Sea Turtles of the World. An annotated and illustrated catalog of sea turtle species known to date. FAO Fisheries Synopsis No. 125:1-81. McClellan, C.M. & Read, A.J. (2007) Complexity and variation in loggerhead sea turtle life history. Biology Letters 3:592. Meyland, A.B., Bowen, B.W. & Avise, J.C., A genetic test of the natal homing versus social facilitation models for green turtle migration. Science 248: van de Merwe, J., Ibrahim, K. & Whittier, J.M. (2005) Effects of hatchery shading and nest depth on the development and quality of Chelonia mydas hatchlings: implications for hatchery management in Peninsular; Malaysia. Australian Journal of Zoology 53: van de Merwe, J., Ibrahim, K. & Whittier, J.M. (2006) Effects of nest depth, shading, and metabolic heating on nest temperatures in sea turtle hatcheries. Chelonian Conservation and Biology 5: van de Merwe, J., Ibrahim, K., Lee, S. & Whittier, J. (2009) Habitat use by green turtles (Chelonia mydas) nesting in Peninsular Malaysia: local and regional conservation implications. Wildlife Research 36: Miller, J.D. (1997) Reproduction in sea turtles. The biology of sea turtles. pp CRC Press. Inc, Boca Raton, Florida. Miller, J.D. & Limpus, C.J. (2003) Ontogeny of marine turtle gonads. In: P. Lutz, J. Musick and J, Wyneken (eds). Biology of Sea Turtles. CRC Press, Boca Raton. Miller, K. (2000) Pacific Salmon Fisheries: Climate, Information and Adaptation in a Conflict-Ridden Context. Climatic Change 45:61, 37. Moncada, F., Abreu-Grobois, F.A., Muhlia-Melo, A., Bell, C., Tröeng, S., Bjorndal, K.A., Bolten, A.B., Meylan, A.B., Zurita, J., Espinosa, G. & others. (2006) Movement patterns of green turtles (Chelonia mydas) in Cuba and adjacent Caribbean waters inferred from flipper tag recaptures. Journal of herpetology 40: Moritz, C. (1994). "Defining evolutionarily significant units for conservation." TREE 9:
123 Moritz, C., Broderick D., Dethmers, K., FitzSimmons, N.N. and Limpus C.J. (2002). Population genetics of Southeast Asian and Western Pacific green turtles, Chelonia mydas. Final Report to UNEP/CMS. Monzón-Argüello, C., Rico, C., Carreras, C., Calabuig, P., Marco, A. & López-Jurado, L.F. (2009) Variation in spatial distribution of juvenile loggerhead turtles in the eastern Atlantic and western Mediterranean Sea. Journal of Experimental Marine Biology and Ecology 373: Mortimer, J.A., Donnelly, M. & Plotkin, P.T. (2000) Sea Turtles. In Seas at The Millennium: An Environmental Evaluation (Edited by C. Sheppard). Elsevier Ltd. pp., Mortimer A.J., & Carr A. (1987) Reproduction and migrations of the Ascension Islands green turtle (Chelonia mydas). Copeia, 1: Morin, P.A., Luikart, G., Wayne, R.K., SNP-Workshop Group (2004) SNPs in Ecology, evolution and conservation. Trends in Ecology & Evolution 19: Mrosovsky, N. (1980) Thermal Biology of Sea Turtles. American Zoologist 20: Mrosovsky, N. (1994) Sex ratios of sea turtles. Journal of Experimental Zoology 270: Musick J.A. & Limpus,C.J. (1997) Habitat utilization and migration in juvenile sea turtles. The Biology of Sea Turtles (eds. Lutz P.L., Musick J.A.). pp CRC Press, Boca Raton, FL. Naro-Maciel, E., Le, M., FitzSimmons, N.N. & Amato, G. (2008) Evolutionary relationships of marine turtles: A molecular phylogeny based on nuclear and mitochondrial genes. Molecular Phylogenetics and Evolution 4: Naro-Maciel, E., Becker, J.H., Lima, E.H.S.M., Marcovaldi, M.A. & DeSalle, R. (2007) Testing Dispersal Hypotheses in Foraging Green Sea Turtles (Chelonia mydas) of Brazil. J Hered 98: Nei, M. (1987) Molecular Evolutionary Genetics. Columbia University Press, New York, NY, USA. Norman, J.A. (1996) Conservation genetics of the green turtle Chelonia mydas L. PhD Thesis. University of Queensland. Norman, J.A., Moritz, C. and Limpus, C.J. (1994). "Mitochondrial DNA control region polymorphisms: genetic markers for ecological studies of marine turtles." Molecular Ecology 3: Okuyama, T. & Bolker, B.M. (2005) Combining genetic and ecological data to estimate sea turtle origins. Ecological Applications 15: Owens, D. W. (1980) The comparative Reproductive physiology of Sea Turtles. American Zoologist 20:
124 Palma, P. (1993) Marine turtle conservation in the Philippines. In A. Nacu, R. Trono, J.A. Palma, D. Torres and F Agas Jr. (eds.), Proceedings of the First ASEAN Symposium Workshop on Marine Turtle Conservation. pp WWF-USAID, Manila, Philippines. Pella, J. & Masuda, M. (2001) Bayesian methods for analysis of stock mixtures from genetic characters. Fisheries Bulletin 99: Pella J.J. & Milner G. B. (1987) Use of genetic marks in stock composition analysis. In Population genetics and fisheries management, eds Ryman N. & Utter F. University of Washington Press, Seattle, pp Peare, T., and Parker, P.G. (1996) Local genetic structure within two rookeries of Chelonia mydas (the green turtle)." Heredity 77: Pfenninger, M. & Posada, D. (2002) Phylogeographic history of the land snail Candidula unifasciata (Helicellinae, Stylommatophora): fragmentation, corridor migration, and secondary contact. Evolution 56: Pilcher, N.J. (2010) Population structure and growth of immature green turtles at Mantanani, Sabah, Malaysia. Pilcher, N.J. (2007) Regional Action Plan for the Conservation of Marine Turtles and their Habitats in the Sulu-Sulawesi Seascape. Conservation International Philippines. Pilcher, N.J. & Basintal, P. (2000) Reproductive biology of the green turtle Chelonia mydas in Sabah, Malaysia. Asian Journal of Tropical Biology 4: Plotkin PT, Byles RA, David RC, and Owens DW (1995) Independent versus socially facilitated oceanic migrations of the olive ridley, Lepidochelys olivacea. Marine Biology 122: Poloczanska, E.S., Limpus, C.J. & Hays, G.C. (2009) Vulnerability of Marine Turtles to Climate Change. D. W. Sims, editor: Advances in Marine Biology. pp Burlington: Academic Press. Prince, R.I.T. (1994) Status of the Western Australian marine turtle populations: the Western Australian Marine Turtle Project In 'Proceedings of the Marine Turtle Conservation Workshop' (Compiled by Russell James) pp (Australian National Parks and Wildlife Service; Canberra.). Prince, R.I.T. (2000) The Western Australian Marine Turtle Project. Sea turtles of the Indo- Pacific: Research, Management and Conservation. ASEAN ACADEMIC Press, Kuala Lumpur. Pritchard, P.C.H. (1997) Evolution, Phylogeny, and current status. The Biology of Sea Turtles. ed. Peter L. Lutz and John A. Musick. pp CRC Press, Boca Raton. 114
125 Proietti, M.C., Lara-Ruiz, P., Reisser, J.W., Pinto, L.D.S., Dellagostin, O.A. & Marins, L.F. (2009) Green turtles (Chelonia mydas) foraging at Arvoredo Island in Southern Brazil: genetic characterization and mixed stock analysis through mtdna control region haplotypes. Genetics and Molecular Biology 32. Rankin-Baransky, K., Williams, C.J., Bass, A.L., Bowen, B.W. & Spotila, J.R. (2001) Origin of loggerhead turtles stranded in the northeastern United States as determined by mitochondrial DNA analysis. Journal of Herpetology 35:638. Raymond, M. & Rousset, F. An exact test for population differentiation. Evolution 49: Reece, J.S., Ehrhart, L.M. & Parkinson, C.L. (2006) Mixed stock analysis of juvenile loggerheads (Caretta caretta) in Indian River Lagoon, Florida: implications for conservation planning. Conservation Genetics 7: Reis, E.C., Soares, L.S., Vargas, S.M., Santos, F.R., Young, R.J., Bjorndal, K.A., Bolten, A.B. & Lôbo-Hajdu, G. (2009) Genetic composition, population structure and phylogeography of the loggerhead sea turtle: colonization hypothesis for the Brazilian rookeries. Conservation Genetics. DOI: /s Richardson, P.B., Bruford, M.W., Calosso, M.C., Campbell, L.M., Clerveaux, W., Formia, A., Godley, B.J., Henderson, A.C., McClellan, K., Newman, S., Parsons, K., Pepper, M., Ranger, S., Silver, J.J., Slade, L. & Broderick, A.C. (2009) Marine Turtles in the Turks and Caicos Islands: Remnant Rookeries, Regionally Significant Foraging Stocks, and a Major Turtle Fishery. Chelonian Conservation and Biology 8: Roberts, M., Anderson, C., Stender, B., Segars, A., David Whittaker, J., Grady, J. & Quattro, J. (2005) Estimated contribution of Atlantic Coastal loggerhead turtle nesting populations to offshore feeding aggregations. Conservation Genetics 6: Roden, S.E., Dutton, P.H., & Morin, P.A. (2009a) Development of single nucleotide polymorphisms (SNP s) using AFLP fragment isolation technique for Chelonia mydas. Journal of Heredity, in press Roden, S.E., Dutton, P.H. & Morin, P.A. (2009b) Characterization of single nucleotide polymorphism markers for the green sea turtle (Chelonia mydas). Molecular Ecology Resources 9: Rooker, J., Secor, D., DeMetrio, G., Kaufman, A., Belmonte Ríos, A. & Ticina, V. (2008) Evidence of trans-atlantic movement and natal homing of bluefin tuna from stable isotopes in otoliths. Marine Ecology Progress Series 368: Salmon, M., Witherington, B. E (1995) Artificial lighting and seafinding behavior by loggerhead hatchlings: evidence for lunar modulation Copeia, Schroth, W., Streit, B., and Schierwater, B. (1996) Evolutionary handicap for turtles. Nature 384:
126 Sears, C., Bowen, B.W., Chapman, R.W., Galloway, S.B., Hopkins-Murphy, S.R. & Woodley, C.M. (1995) Demographic composition of the feeding population of juvenile loggerhead sea turtles (Caretta caretta) off Charleston, South Carolina: evidence from mitochondrial DNA markers. Marine Biology 123: Senko, J., López-Castro, M.C., Koch, V. & Nichols, W.J. (2010) Immature east Pacific green turtles (Chelonia mydas) use multiple foraging areas off the Pacific Coast of Baja California Sur, Mexico: First evidence from mark-recapture data. Pacific Science 64: Shanker, K., Ramadevi, J., Choudhury, B.C., Singh, L. & Aggarwal, R.K. (2004) Phylogeography of olive ridley turtles (Lepidochelys olivacea) on the east coast of India: implications for conservation theory. Molecular Ecology 13: Shanker, K. & Pilcher, N.J. (2003) Marine Turtle Conservation in South and Southeast Asia: Cause for hope or hopeless cause? Marine Turtle Newsletter 100: Shizheng, W. & Shi, H. (2009) Sea Turtle Conservation Status in Paracel Islands and Case Analysis of Sea Turtle Conservation in Hainan Province. In Chan, E.H., N. Pilcher and K. Hiew Report of the Workshop on Regional Regional Cooperation to Address Direct Capture of Sea Turtles. pp Kuala Terengganu, Malaysia. Universiti Malaysia Terengganu Press. de Silva, G. (1982) The status of sea turtle populations in east Malaysia and the South China Sea. In: Biology and Conservation of Sea Turtles (K.A. Bjorndal, ed.). pp Smithsonian Institution Press, Washington, D.C. Slatkin, M. (1995) A Measure of Population Subdivision Based on Microsatellite Allele Frequencies. Genetics 139: Solow, A.R., Bjorndal, K.A. & Bolten, A.B. (2002) Annual variation in nesting numbers of marine turtles: the effect of sea surface temperature on re-migration intervals. Ecology Letters 5: Takahata, N. (1988) The coalescent in two partially isolated diffusion populations. Genet. Res. 52: Tiwol, C.M. & Cabanban, A.S. (2000) All female hatchlings from the open-beach hatchery at Gulisaan Island, Turtle Islands park, Sabah. Sea turtles of the Indo-Pacific: Research, Management and Conservation. Asean Academic Press, Kuala Lumpur. Troëng, S., Dutton, P.H. & Evans, D. (2005a) Migration of hawksbill turtles (Eretmochelys imbricate) from Tortuguero, Costa Rica. Ecography 28: Troëng, S., Evans, D.R., Harrison, E. & Lagueux, C.J. (2005b) Migration of green turtles Chelonia mydas from Tortuguero, Costa Rica. Marine Biology 148: Vargas, S.M., Araujo, F.C., Monteiro, D.S., Estima, S.C., Almeida, A.P., Soares, L.S. & Santos, F.R. (2008) Genetic diversity and origin of Leatherback turtles (Dermochelys coriacea) from the Brazilian coast. Journal of Heredity 99:
127 Velez-Zuazo, X., Ramos, W.D., Van Dam, R.P., Diez, C.E., Abreu-Grobois, A.[. & McMillan, W.O. (2008) Dispersal, recruitment and migratory behaviour in a hawksbill sea turtle aggregation. Molecular Ecology 17: Waples, R.S., Beechie, T. & Pess, G.R. (2009) Evolutionary History, Habitat Disturbance Regimes, and Anthropogenic Changes: What Do These Mean for Resilience of Pacific Salmon Populations? Ecology and Society 14:3. Waples, R.S., Zabel, R.W., Scheuerell, M.D. & Sanderson, B.L. (2008) Evolutionary responses by native species to major anthropogenic changes to their ecosystems: Pacific salmon in the Columbia River hydropower system. Molecular Ecology 17: Watterson, G.A. (1985) The genetic divergence of two populations. Theor. Popul. Biol 27: Wetherall, J.A., Balazs, G.H., Tokunaga, R.A. & Yong, M.Y.Y. (1993) in International North Pacific Fisheries Commission symposium on biology, distribution, and stock assessment of species caught in the high seas driftnet fisheries in the north Pacific Ocean. Eds, Ito, J., Shaw, W. & Burgner, R.L. (int. North Pacific Fisheries Commission, Vancouver, Canada). Bull. No. 53: Whiting, S. & Koch, A.U. (2006) Oceanic movement of a benthic foraging juvenile hawksbill turtle from the Cocos (Keeling) Islands. Marine Turtle Newsletter 112: Whiting S. D., Murray, W., Macrae I., Thorn R., Chongkin, M., Kock A. (2008) Nonmigratory breeding by isolated green sea turtles (Chelonia mydas) in the Indian Ocean: biological and conservation implications. Naturwissenschaften 95: Williams, J.G., Zabel, R.W., Waples, R.S., Hutchings, J.A. & Connor, W.P. (2008) Potential for anthropogenic disturbances to influence evolutionary change in the life history of a threatened salmonid. Evolutionary Applications 1: Witherington B.E., and Frazer N.B. (2003) Social and Economic Aspects of Sea Turtle Conservation. In The Biology of Sea Turtles, Volume II, eds Lutz PL, Musick JA, and Wyneken, J, pp CRC Marine Science Series, CRC Press, Inc., Boca Raton, Florida. Witzell, W.N. (1983) Synopsis of biological data on the hawksbill turtle Eretmochelys imbricate (Linnaeus, 1766). FAO fisheries synopsis 137:1-73. Witzell, W.N., Bass, A.L., Bresette, M.J., Singewald, D.A. & Gorham, J.C. (2002) Origin of immature loggerhead sea turtles (Caretta caretta) at Hutchinson Island, Florida: evidence from mtdna markers. Fisheries Bulletin 100: Zaykin, D.V. & Pudovkin, A.I. (1993) Two Programs to Estimate Significance of {chi}2 Values Using Pseudo-Probability Tests. Journal of Heredity 84:
128 APPENDIX 1 Appendix 1. All sequence used in Chapter 3 for the 780 bp amplified by primers LTEi9 and H950. Grey area identifies the 384 bp sequence used by Dethmers et al
129 119

130 120
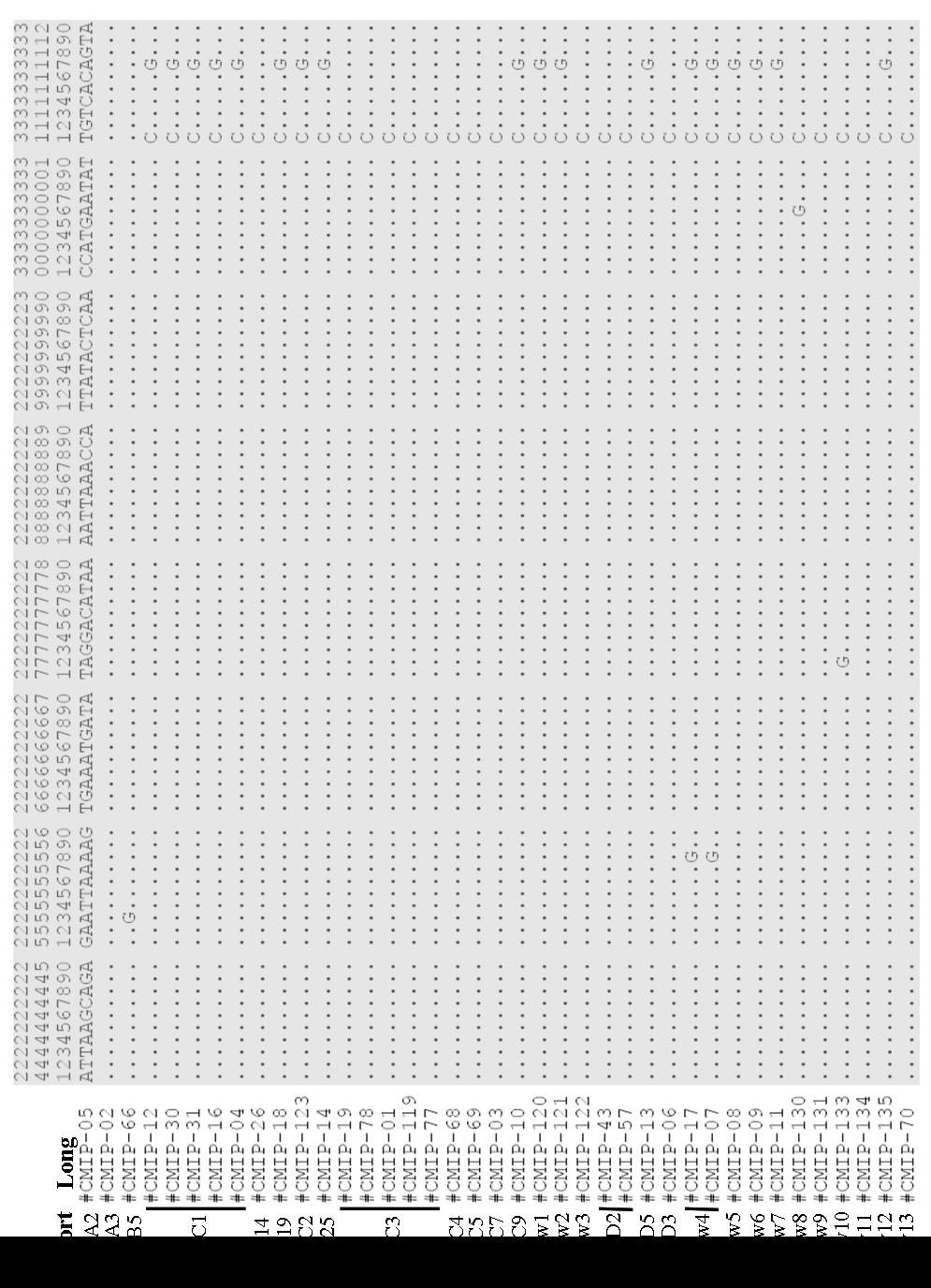
131 121

132 122

133 123

134 124
MARINE TURTLE GENETIC STOCKS OF THE INDO-PACIFIC: IDENTIFYING BOUNDARIES AND KNOWLEDGE GAPS NANCY N. FITZSIMMONS & COLIN J. LIMPUS
 MARINE TURTLE GENETIC STOCKS OF THE INDO-PACIFIC: IDENTIFYING BOUNDARIES AND KNOWLEDGE GAPS NANCY N. FITZSIMMONS & COLIN J. LIMPUS 7 th MEETING OF SIGNATORY STATES, INDIAN SOUTH-EAST ASIAN MARINE TURTLE
MARINE TURTLE GENETIC STOCKS OF THE INDO-PACIFIC: IDENTIFYING BOUNDARIES AND KNOWLEDGE GAPS NANCY N. FITZSIMMONS & COLIN J. LIMPUS 7 th MEETING OF SIGNATORY STATES, INDIAN SOUTH-EAST ASIAN MARINE TURTLE
BIODIVERSITY CONSERVATION AND HABITAT MANAGEMENT Vol. II Initiatives For The Conservation Of Marine Turtles - Paolo Luschi
 INITIATIVES FOR THE CONSERVATION OF MARINE TURTLES Paolo Luschi Department of Biology, University of Pisa, Italy Keywords: sea turtles, conservation, threats, beach management, artificial light management,
INITIATIVES FOR THE CONSERVATION OF MARINE TURTLES Paolo Luschi Department of Biology, University of Pisa, Italy Keywords: sea turtles, conservation, threats, beach management, artificial light management,
INTRODUCTION OBJECTIVE METHOD IDENTIFICATION OF NATAL ORIGIN SEA TURTLES AT BRUNEI BAY / LAWAS FORAGING HABITATS
 REGIONAL MEETING ON CONSERVATION AND MANAGEMENT OF SEA TURTLE FORAGING HABITATS IN SOUTHEAST ASIAN WATERS - OCTOBER 0 AnCasa Hotel & Spa Kuala Lumpur IDENTIFICATION OF NATAL ORIGIN SEA TURTLES AT BRUNEI
REGIONAL MEETING ON CONSERVATION AND MANAGEMENT OF SEA TURTLE FORAGING HABITATS IN SOUTHEAST ASIAN WATERS - OCTOBER 0 AnCasa Hotel & Spa Kuala Lumpur IDENTIFICATION OF NATAL ORIGIN SEA TURTLES AT BRUNEI
INTRODUCTION OBJECTIVE REGIONAL ANALYSIS ON STOCK IDENTIFICATION OF GREEN AND HAWKSBILL TURTLES IN THE SOUTHEAST ASIAN REGION
 The Third Technical Consultation Meeting (3rd TCM) Research for Stock Enhancement of Sea Turtles (Japanese Trust Fund IV Program) 7 October 2008 REGIONAL ANALYSIS ON STOCK IDENTIFICATION OF GREEN AND HAWKSBILL
The Third Technical Consultation Meeting (3rd TCM) Research for Stock Enhancement of Sea Turtles (Japanese Trust Fund IV Program) 7 October 2008 REGIONAL ANALYSIS ON STOCK IDENTIFICATION OF GREEN AND HAWKSBILL
PARTIAL REPORT. Juvenile hybrid turtles along the Brazilian coast RIO GRANDE FEDERAL UNIVERSITY
 RIO GRANDE FEDERAL UNIVERSITY OCEANOGRAPHY INSTITUTE MARINE MOLECULAR ECOLOGY LABORATORY PARTIAL REPORT Juvenile hybrid turtles along the Brazilian coast PROJECT LEADER: MAIRA PROIETTI PROFESSOR, OCEANOGRAPHY
RIO GRANDE FEDERAL UNIVERSITY OCEANOGRAPHY INSTITUTE MARINE MOLECULAR ECOLOGY LABORATORY PARTIAL REPORT Juvenile hybrid turtles along the Brazilian coast PROJECT LEADER: MAIRA PROIETTI PROFESSOR, OCEANOGRAPHY
Conservation Sea Turtles
 Conservation of Sea Turtles Regional Action Plan for Latin America and the Caribbean Photo: Fran & Earle Ketley Rare and threatened reptiles Each day appreciation grows for the ecological roles of sea
Conservation of Sea Turtles Regional Action Plan for Latin America and the Caribbean Photo: Fran & Earle Ketley Rare and threatened reptiles Each day appreciation grows for the ecological roles of sea
Final Report for Research Work Order 167 entitled:
 Final Report for Research Work Order 167 entitled: Population Genetic Structure of Marine Turtles, Eretmochelys imbricata and Caretta caretta, in the Southeastern United States and adjacent Caribbean region
Final Report for Research Work Order 167 entitled: Population Genetic Structure of Marine Turtles, Eretmochelys imbricata and Caretta caretta, in the Southeastern United States and adjacent Caribbean region
Convention on the Conservation of Migratory Species of Wild Animals
 MEMORANDUM OF UNDERSTANDING ON THE CONSERVATION AND MANAGEMENT OF MARINE TURTLES AND THEIR HABITATS OF THE INDIAN OCEAN AND SOUTH-EAST ASIA Concluded under the auspices of the Convention on the Conservation
MEMORANDUM OF UNDERSTANDING ON THE CONSERVATION AND MANAGEMENT OF MARINE TURTLES AND THEIR HABITATS OF THE INDIAN OCEAN AND SOUTH-EAST ASIA Concluded under the auspices of the Convention on the Conservation
2. LITERATURE REVIEW
 2. LITERATURE REVIEW 2.1. General remarks of seaturtle Overall, there are seven living species of seaturtles distributed worldwide (Marquez-M, 1990). They are Green turtle (Chelonia mydas), Hawksbill turtle
2. LITERATURE REVIEW 2.1. General remarks of seaturtle Overall, there are seven living species of seaturtles distributed worldwide (Marquez-M, 1990). They are Green turtle (Chelonia mydas), Hawksbill turtle
Since 1963, Department of Fisheries (DOF) has taken up a project to breed and protect sea Turtles on Thameehla island.
 has taken up a project to breed and protect sea Turtles on Thameehla island.") Thameehla (Diamond) Island Marine Turtle Conservation and Management Station, Ayeyawady Region, Myanmar Background Thameehla Island is situated between the Bay of Bengal and the Gulf of Mottama (Gulf of
Thameehla (Diamond) Island Marine Turtle Conservation and Management Station, Ayeyawady Region, Myanmar Background Thameehla Island is situated between the Bay of Bengal and the Gulf of Mottama (Gulf of
Bycatch records of sea turtles obtained through Japanese Observer Program in the IOTC Convention Area
 Bycatch records of sea turtles obtained through Japanese Observer Program in the IOTC Convention Area Kei Okamoto and Kazuhiro Oshima National Research Institute of Far Seas Fisheries, Japan Fisheries
Bycatch records of sea turtles obtained through Japanese Observer Program in the IOTC Convention Area Kei Okamoto and Kazuhiro Oshima National Research Institute of Far Seas Fisheries, Japan Fisheries
Study site #2 the reference site at the southern end of Cleveland Bay.
 CHRISTINE HOF / WWF-AUS We all made our way from various parts of Queensland to our reference site at Cleveland Bay in order to sample the environment and turtles for the Rivers to Reef to Turtles (RRT)
CHRISTINE HOF / WWF-AUS We all made our way from various parts of Queensland to our reference site at Cleveland Bay in order to sample the environment and turtles for the Rivers to Reef to Turtles (RRT)
Green Turtle (Chelonia mydas) nesting behaviour in Kigamboni District, United Republic of Tanzania.
 nesting behaviour in Kigamboni District, United Republic of Tanzania.") Green Turtle (Chelonia mydas) nesting behaviour in Kigamboni District, United Republic of Tanzania. Lindsey West Sea Sense, 32 Karume Road, Oyster Bay, Dar es Salaam, Tanzania Introduction Tanzania is
Green Turtle (Chelonia mydas) nesting behaviour in Kigamboni District, United Republic of Tanzania. Lindsey West Sea Sense, 32 Karume Road, Oyster Bay, Dar es Salaam, Tanzania Introduction Tanzania is
EYE PROTECTION BIFOCAL SAFETY GLASSES ANSI Z87.1 ANSI Z87.1 ANSI Z87.1 SAFETY GOGGLE MODEL # TYG 400 G SAFETY GOGGLE MODEL # TYG 405 SAFETY GOGGLE
 EYE PROTECTION TY700-F Bifocal Safety Glasses EN166 TY701-SF Safety Glasses EN166 Removeable & soft foam inner frame provides comfortable fit Anti-fog and anti-scratch treated lenses Trendy & Sporty style,
EYE PROTECTION TY700-F Bifocal Safety Glasses EN166 TY701-SF Safety Glasses EN166 Removeable & soft foam inner frame provides comfortable fit Anti-fog and anti-scratch treated lenses Trendy & Sporty style,
ABSTRACT. Ashmore Reef
 ABSTRACT The life cycle of sea turtles is complex and is not yet fully understood. For most species, it involves at least three habitats: the pelagic, the demersal foraging and the nesting habitats. This
ABSTRACT The life cycle of sea turtles is complex and is not yet fully understood. For most species, it involves at least three habitats: the pelagic, the demersal foraging and the nesting habitats. This
Andaman & Nicobar Islands
 Map showing and Nicobar Dr. A. Murugan Suganthi Devadason Marine Research Institute 44-Beach Road, Tuticorin-628 001, India Tel.: +91 461 2336488; Fax: +91 461 2325692 & Nicobar Location: 6 45 N to 13
Map showing and Nicobar Dr. A. Murugan Suganthi Devadason Marine Research Institute 44-Beach Road, Tuticorin-628 001, India Tel.: +91 461 2336488; Fax: +91 461 2325692 & Nicobar Location: 6 45 N to 13
Aspects in the Biology of Sea Turtles
 Charting Multidisciplinary Research and Action Priorities towards the Conservation and Sustainable Management of Sea Turtles in the Pacific Ocean: A Focus on Malaysia Malaysia s Natural Heritage Aspects
Charting Multidisciplinary Research and Action Priorities towards the Conservation and Sustainable Management of Sea Turtles in the Pacific Ocean: A Focus on Malaysia Malaysia s Natural Heritage Aspects
Marine Turtle Research Program
 Marine Turtle Research Program NOAA Fisheries Southwest Fisheries Science Center La Jolla, CA Agenda Item C.1.b Supplemental Power Point Presentation 2 September 2005 Marine Turtle Research Program Background
Marine Turtle Research Program NOAA Fisheries Southwest Fisheries Science Center La Jolla, CA Agenda Item C.1.b Supplemental Power Point Presentation 2 September 2005 Marine Turtle Research Program Background
HOWICK GROUP FIELD RESEARCH
 HOWICK GROUP FIELD RESEARCH UPDATE #6 The Rivers to Reef to Turtles Project We embarked on our second Rivers to Reef to Turtles Project (RRT) Field Trip to the offshore, very remote and isolated part of
HOWICK GROUP FIELD RESEARCH UPDATE #6 The Rivers to Reef to Turtles Project We embarked on our second Rivers to Reef to Turtles Project (RRT) Field Trip to the offshore, very remote and isolated part of
Sea Turtles in the Middle East and South Asia Region
 Sea Turtles in the Middle East and South Asia Region MTSG Annual Regional Report 2018 Editors: Andrea D. Phillott ALan F. Rees 1 Recommended citation for this report: Phillott, A.D. and Rees, A.F. (Eds.)
Sea Turtles in the Middle East and South Asia Region MTSG Annual Regional Report 2018 Editors: Andrea D. Phillott ALan F. Rees 1 Recommended citation for this report: Phillott, A.D. and Rees, A.F. (Eds.)
University of Canberra. This thesis is available in print format from the University of Canberra Library.
 University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
PROJECT DOCUMENT. Project Leader
 Thirty-seventh Meeting of the Program Committee Southeast Asian Fisheries Development Center Sunee Grand Hotel & Convention Center, Ubon Ratchathani, Thailand 1-3 December 2014 WP03.1d-iii Program Categories:
Thirty-seventh Meeting of the Program Committee Southeast Asian Fisheries Development Center Sunee Grand Hotel & Convention Center, Ubon Ratchathani, Thailand 1-3 December 2014 WP03.1d-iii Program Categories:
Sea Turtle Conservation in Seychelles
 Sea Turtle Conservation in Seychelles by Jeanne A. Mortimer, PhD Presentation made to participants of the Regional Workshop and 4 th Meeting of the WIO-Marine Turtle Task Force Port Elizabeth, South Africa
Sea Turtle Conservation in Seychelles by Jeanne A. Mortimer, PhD Presentation made to participants of the Regional Workshop and 4 th Meeting of the WIO-Marine Turtle Task Force Port Elizabeth, South Africa
MANAGING MEGAFAUNA IN INDONESIA : CHALLENGES AND OPPORTUNITIES
 MANAGING MEGAFAUNA IN INDONESIA : CHALLENGES AND OPPORTUNITIES By Dharmadi Agency for Marine and Fisheries Research Ministry of Marine Affairs and Fisheries Republic of Indonesia MEGAFAUNA I. SEA TURTLES
MANAGING MEGAFAUNA IN INDONESIA : CHALLENGES AND OPPORTUNITIES By Dharmadi Agency for Marine and Fisheries Research Ministry of Marine Affairs and Fisheries Republic of Indonesia MEGAFAUNA I. SEA TURTLES
MARINE TURTLE LANDING, HATCHING, AND PREDATION IN TURTLE ISLANDS PARK (TIP), SABAH
, SABAH") MARINE TURTLE LANDING, HATCHING, AND PREDATION IN TURTLE ISLANDS PARK (TIP), SABAH Coastal and Marine Resources Management in the Coral Triangle-Southeast Asia (TA 7813-REG) Technical Report FINAL REPORT:
MARINE TURTLE LANDING, HATCHING, AND PREDATION IN TURTLE ISLANDS PARK (TIP), SABAH Coastal and Marine Resources Management in the Coral Triangle-Southeast Asia (TA 7813-REG) Technical Report FINAL REPORT:
HAWKSBILL SEA TURTLE POPULATION MONITORING
 HAWKSBILL SEA TURTLE POPULATION MONITORING CAHUITA NATIONAL PARK COSTA RICA, 2007 1 PROJECT INFORMATION FOR APPLICANTS WELCOME! Didiher Chacón-Chaverri Project Director Joana Hancock Research Coordinator
HAWKSBILL SEA TURTLE POPULATION MONITORING CAHUITA NATIONAL PARK COSTA RICA, 2007 1 PROJECT INFORMATION FOR APPLICANTS WELCOME! Didiher Chacón-Chaverri Project Director Joana Hancock Research Coordinator
PROJECT DOCUMENT. This year budget: Project Leader
 Thirty-sixth Meeting of the Program Committee Southeast Asian Fisheries Development Center Trader Hotel, Penang, Malaysia 25-27 November 2013 WP03.1d-iii PROJECT DOCUMENT Program Categories: Project Title:
Thirty-sixth Meeting of the Program Committee Southeast Asian Fisheries Development Center Trader Hotel, Penang, Malaysia 25-27 November 2013 WP03.1d-iii PROJECT DOCUMENT Program Categories: Project Title:
Tagging Study on Green Turtle (Chel Thameehla Island, Myanmar. Proceedings of the 5th Internationa. SEASTAR2000 workshop) (2010): 15-19
 (2010): 15-19") Title Tagging Study on Green Turtle (Chel Thameehla Island, Myanmar Author(s) LWIN, MAUNG MAUNG Proceedings of the 5th Internationa Citation SEASTAR2000 and Asian Bio-logging S SEASTAR2000 workshop) (2010):
Title Tagging Study on Green Turtle (Chel Thameehla Island, Myanmar Author(s) LWIN, MAUNG MAUNG Proceedings of the 5th Internationa Citation SEASTAR2000 and Asian Bio-logging S SEASTAR2000 workshop) (2010):
Study site #3 the primary site at the southern end of Upstart Bay.
 CHRISTINE HOF / /WWF-AUS We all made our way from various parts of Queensland to our primary study site at Upstart Bay in order to sample the environment and turtles for the Rivers to Reef to Turtles (RRT)
CHRISTINE HOF / /WWF-AUS We all made our way from various parts of Queensland to our primary study site at Upstart Bay in order to sample the environment and turtles for the Rivers to Reef to Turtles (RRT)
Who Really Owns the Beach? The Competition Between Sea Turtles and the Coast Renee C. Cohen
 Who Really Owns the Beach? The Competition Between Sea Turtles and the Coast Renee C. Cohen Some Common Questions Microsoft Word Document This is an outline of the speaker s notes in Word What are some
Who Really Owns the Beach? The Competition Between Sea Turtles and the Coast Renee C. Cohen Some Common Questions Microsoft Word Document This is an outline of the speaker s notes in Word What are some
CHARACTERISTIC COMPARISON. Green Turtle - Chelonia mydas
 5 CHARACTERISTIC COMPARISON Green Turtle - Chelonia mydas Green turtles average 1.2m to 1.4m in length, are between 120kg to 180kg in weight at full maturity and found in tropical and sub-tropical seas
5 CHARACTERISTIC COMPARISON Green Turtle - Chelonia mydas Green turtles average 1.2m to 1.4m in length, are between 120kg to 180kg in weight at full maturity and found in tropical and sub-tropical seas
THE SPATIAL DYNAMICS OF SEA TURTLES WITHIN FORAGING GROUNDS ON ELEUTHERA, THE BAHAMAS
 Earthwatch 2016 Annual Field Report TRACKING SEA TURTLES IN THE BAHAMAS THE SPATIAL DYNAMICS OF SEA TURTLES WITHIN FORAGING GROUNDS ON ELEUTHERA, THE BAHAMAS Annabelle Brooks, MSc REPORT COMPLETED BY:
Earthwatch 2016 Annual Field Report TRACKING SEA TURTLES IN THE BAHAMAS THE SPATIAL DYNAMICS OF SEA TURTLES WITHIN FORAGING GROUNDS ON ELEUTHERA, THE BAHAMAS Annabelle Brooks, MSc REPORT COMPLETED BY:
American Samoa Sea Turtles
 American Samoa Sea Turtles Climate Change Vulnerability Assessment Summary An Important Note About this Document: This document represents an initial evaluation of vulnerability for sea turtles based on
American Samoa Sea Turtles Climate Change Vulnerability Assessment Summary An Important Note About this Document: This document represents an initial evaluation of vulnerability for sea turtles based on
Final Report. Nesting green turtles of Torres Strait. Mark Hamann, Justin Smith, Shane Preston and Mariana Fuentes
 Final Report Nesting green turtles of Torres Strait Mark Hamann, Justin Smith, Shane Preston and Mariana Fuentes Nesting green turtles of Torres Strait Final report Mark Hamann 1, Justin Smith 1, Shane
Final Report Nesting green turtles of Torres Strait Mark Hamann, Justin Smith, Shane Preston and Mariana Fuentes Nesting green turtles of Torres Strait Final report Mark Hamann 1, Justin Smith 1, Shane
Maternal Effects in the Green Turtle (Chelonia mydas)
") Maternal Effects in the Green Turtle (Chelonia mydas) SUBMITTED BY SAM B. WEBER TO THE UNIVERSITY OF EXETER AS A THESIS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY IN BIOLOGY; 8 TH JUNE 2010 This thesis is
Maternal Effects in the Green Turtle (Chelonia mydas) SUBMITTED BY SAM B. WEBER TO THE UNIVERSITY OF EXETER AS A THESIS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY IN BIOLOGY; 8 TH JUNE 2010 This thesis is
Dugong and Marine Turtle Knowledge Handbook February 2005
 PART 2b: MARINE TURTLES 56 Introduction 56 Concerns 56 Biology of marine turtles 60 Names and Classification of Marine Turtles 64 Loggerhead Turtle 65 Green Turtle 69 Hawksbill Turtle 74 Olive Ridley Turtle
PART 2b: MARINE TURTLES 56 Introduction 56 Concerns 56 Biology of marine turtles 60 Names and Classification of Marine Turtles 64 Loggerhead Turtle 65 Green Turtle 69 Hawksbill Turtle 74 Olive Ridley Turtle
The Seal and the Turtle
 The Seal and the Turtle Green Sea Turtle (Chelonia mydas) Weight: Length: Appearance: Lifespan: 300-350 pounds (135-160 kg) for adults; hatchlings weigh 0.05 lbs (25 g) 3 feet (1 m) for adults; hatchlings
The Seal and the Turtle Green Sea Turtle (Chelonia mydas) Weight: Length: Appearance: Lifespan: 300-350 pounds (135-160 kg) for adults; hatchlings weigh 0.05 lbs (25 g) 3 feet (1 m) for adults; hatchlings
Sea Turtles and Longline Fisheries: Impacts and Mitigation Experiments
 Sea Turtles and Longline Fisheries: Impacts and Mitigation Experiments Yonat Swimmer, Mike Musyl, Lianne M c Naughton, Anders Nielson, Richard Brill, Randall Arauz PFRP P.I. Meeting Dec. 9, 2003 Species
Sea Turtles and Longline Fisheries: Impacts and Mitigation Experiments Yonat Swimmer, Mike Musyl, Lianne M c Naughton, Anders Nielson, Richard Brill, Randall Arauz PFRP P.I. Meeting Dec. 9, 2003 Species
Sea Turtles and the Environmental Management of Industrial Activities in. North West Western Australia. Kellie Lee Pendoley
 Sea Turtles and the Environmental Management of Industrial Activities in North West Western Australia Kellie Lee Pendoley This thesis is presented for the degree of Doctor of Philosophy Murdoch University
Sea Turtles and the Environmental Management of Industrial Activities in North West Western Australia Kellie Lee Pendoley This thesis is presented for the degree of Doctor of Philosophy Murdoch University
Status of leatherback turtles in Australia
 Status of leatherback turtles in Australia by Colin Limpus 1. The legal protection status for leatherback turtles In Australia, wildlife management is the responsibility of both the Federal and State and
Status of leatherback turtles in Australia by Colin Limpus 1. The legal protection status for leatherback turtles In Australia, wildlife management is the responsibility of both the Federal and State and
RWO 166. Final Report to. Florida Cooperative Fish and Wildlife Research Unit University of Florida Research Work Order 166.
 MIGRATION AND HABITAT USE OF SEA TURTLES IN THE BAHAMAS RWO 166 Final Report to Florida Cooperative Fish and Wildlife Research Unit University of Florida Research Work Order 166 December 1998 Karen A.
MIGRATION AND HABITAT USE OF SEA TURTLES IN THE BAHAMAS RWO 166 Final Report to Florida Cooperative Fish and Wildlife Research Unit University of Florida Research Work Order 166 December 1998 Karen A.
What I learned from Limpus, Carter. Quantifying a Nesting Season. and Hamann (2001) and. Sussing out. Identifying
 and. Sussing out. Identifying") What I learned from Limpus, Carter and Hamann (2001) and Quantifying a Nesting Season Identifying Sussing out Michael Guinea School of Environment Charles Darwin University Darwin 0909 Whole of season
What I learned from Limpus, Carter and Hamann (2001) and Quantifying a Nesting Season Identifying Sussing out Michael Guinea School of Environment Charles Darwin University Darwin 0909 Whole of season
Rookery on the east coast of Penins. Author(s) ABDULLAH, SYED; ISMAIL, MAZLAN. Proceedings of the International Sy
 ABDULLAH, SYED; ISMAIL, MAZLAN. Proceedings of the International Sy") Temperature dependent sex determina Titleperformance of green turtle (Chelon Rookery on the east coast of Penins Author(s) ABDULLAH, SYED; ISMAIL, MAZLAN Proceedings of the International Sy Citation SEASTAR2000
Temperature dependent sex determina Titleperformance of green turtle (Chelon Rookery on the east coast of Penins Author(s) ABDULLAH, SYED; ISMAIL, MAZLAN Proceedings of the International Sy Citation SEASTAR2000
REPORT / DATA SET. National Report to WATS II for the Cayman Islands Joe Parsons 12 October 1987 WATS2 069
 WATS II REPORT / DATA SET National Report to WATS II for the Cayman Islands Joe Parsons 12 October 1987 WATS2 069 With a grant from the U.S. National Marine Fisheries Service, WIDECAST has digitized the
WATS II REPORT / DATA SET National Report to WATS II for the Cayman Islands Joe Parsons 12 October 1987 WATS2 069 With a grant from the U.S. National Marine Fisheries Service, WIDECAST has digitized the
Recognizing that the government of Mexico lists the loggerhead as in danger of extinction ; and
 RESOLUTION URGING THE REPUBLIC OF MEXICO TO END HIGH BYCATCH MORTALITY AND STRANDINGS OF NORTH PACIFIC LOGGERHEAD SEA TURTLES IN BAJA CALIFORNIA SUR, MEXICO Recalling that the Republic of Mexico has worked
RESOLUTION URGING THE REPUBLIC OF MEXICO TO END HIGH BYCATCH MORTALITY AND STRANDINGS OF NORTH PACIFIC LOGGERHEAD SEA TURTLES IN BAJA CALIFORNIA SUR, MEXICO Recalling that the Republic of Mexico has worked
Dr Kathy Slater, Operation Wallacea
 ABUNDANCE OF IMMATURE GREEN TURTLES IN RELATION TO SEAGRASS BIOMASS IN AKUMAL BAY Dr Kathy Slater, Operation Wallacea All sea turtles in the Caribbean are listed by the IUCN (2012) as endangered (green
ABUNDANCE OF IMMATURE GREEN TURTLES IN RELATION TO SEAGRASS BIOMASS IN AKUMAL BAY Dr Kathy Slater, Operation Wallacea All sea turtles in the Caribbean are listed by the IUCN (2012) as endangered (green
Gulf and Caribbean Research
 Gulf and Caribbean Research Volume 16 Issue 1 January 4 Morphological Characteristics of the Carapace of the Hawksbill Turtle, Eretmochelys imbricata, from n Waters Mari Kobayashi Hokkaido University DOI:
Gulf and Caribbean Research Volume 16 Issue 1 January 4 Morphological Characteristics of the Carapace of the Hawksbill Turtle, Eretmochelys imbricata, from n Waters Mari Kobayashi Hokkaido University DOI:
A brief report on the 2016/17 monitoring of marine turtles on the São Sebastião peninsula, Mozambique
 A brief report on the 2016/17 monitoring of marine turtles on the São Sebastião peninsula, Mozambique 23 June 2017 Executive summary The Sanctuary successfully concluded its 8 th year of marine turtle
A brief report on the 2016/17 monitoring of marine turtles on the São Sebastião peninsula, Mozambique 23 June 2017 Executive summary The Sanctuary successfully concluded its 8 th year of marine turtle
Research and Management Techniques for the Conservation of Sea Turtles
 Research and Management Techniques for the Conservation of Sea Turtles Prepared by IUCN/SSC Marine Turtle Specialist Group Edited by Karen L. Eckert Karen A. Bjorndal F. Alberto Abreu-Grobois M. Donnelly
Research and Management Techniques for the Conservation of Sea Turtles Prepared by IUCN/SSC Marine Turtle Specialist Group Edited by Karen L. Eckert Karen A. Bjorndal F. Alberto Abreu-Grobois M. Donnelly
click for previous page SEA TURTLES
 click for previous page SEA TURTLES FAO Sheets Fishing Area 51 TECHNICAL TERMS AND PRINCIPAL MEASUREMENTS USED head width (Straight-line distances) head prefrontal precentral carapace central (or neural)
click for previous page SEA TURTLES FAO Sheets Fishing Area 51 TECHNICAL TERMS AND PRINCIPAL MEASUREMENTS USED head width (Straight-line distances) head prefrontal precentral carapace central (or neural)
Marine reptiles review of interactions and populations Final Report
 Marine reptiles review of interactions and populations Final Report October 2016 Prepared for Department of Conservation Project Code: POP2015-06 Project No: 4658 Project start date: 14 September 2015
Marine reptiles review of interactions and populations Final Report October 2016 Prepared for Department of Conservation Project Code: POP2015-06 Project No: 4658 Project start date: 14 September 2015
Prepared by Christine Hof and Dr Ian Bell
 Prepared by Christine Hof and Dr Ian Bell Acknowledgements We gratefully thank Kelly Forester from Helipower for his piloting expertise and local knowledge, and for Queens Beach Action Group for ground
Prepared by Christine Hof and Dr Ian Bell Acknowledgements We gratefully thank Kelly Forester from Helipower for his piloting expertise and local knowledge, and for Queens Beach Action Group for ground
Allowable Harm Assessment for Leatherback Turtle in Atlantic Canadian Waters
 Maritimes Lead: Stock Status Report 2004/035 Allowable Harm Assessment for in Atlantic Canadian Waters Background The leatherback turtle (Dermochelys coriacea) is designated as endangered by the Committee
Maritimes Lead: Stock Status Report 2004/035 Allowable Harm Assessment for in Atlantic Canadian Waters Background The leatherback turtle (Dermochelys coriacea) is designated as endangered by the Committee
BRITISH INDIAN OCEAN TERRITORY (BIOT) BIOT NESTING BEACH INFORMATION. BIOT MPA designated in April Approx. 545,000 km 2
 BIOT NESTING BEACH INFORMATION. BIOT MPA designated in April Approx. 545,000 km 2") BRITISH INDIAN OCEAN TERRITORY (BIOT) BIOT Dr Peter Richardson, Marine Conservation Society (MCS), UK BIOT MPA designated in April 2010. Approx. 545,000 km 2 Green turtle (Chelonia mydas): Estimated 400
BRITISH INDIAN OCEAN TERRITORY (BIOT) BIOT Dr Peter Richardson, Marine Conservation Society (MCS), UK BIOT MPA designated in April 2010. Approx. 545,000 km 2 Green turtle (Chelonia mydas): Estimated 400
Title Temperature among Juvenile Green Se.
 Title Difference in Activity Correspondin Temperature among Juvenile Green Se TABATA, RUNA; WADA, AYANA; OKUYAMA, Author(s) NAKAJIMA, KANA; KOBAYASHI, MASATO; NOBUAKI PROCEEDINGS of the Design Symposium
Title Difference in Activity Correspondin Temperature among Juvenile Green Se TABATA, RUNA; WADA, AYANA; OKUYAMA, Author(s) NAKAJIMA, KANA; KOBAYASHI, MASATO; NOBUAKI PROCEEDINGS of the Design Symposium
Proceedings of the 2nd Internationa. SEASTAR2000 Workshop) (2005):
 (2005):") TitleSeasonal nesting of green turtles a Author(s) YASUDA, TOHYA; KITTIWATTANAWONG, KO KLOM-IN, WINAI; ARAI, NOBUAKI Proceedings of the 2nd Internationa Citation SEASTAR2 and Asian Bio-logging S SEASTAR2
TitleSeasonal nesting of green turtles a Author(s) YASUDA, TOHYA; KITTIWATTANAWONG, KO KLOM-IN, WINAI; ARAI, NOBUAKI Proceedings of the 2nd Internationa Citation SEASTAR2 and Asian Bio-logging S SEASTAR2
Research and Management Techniques for the Conservation of Sea Turtles
 Research and Management Techniques for the Conservation of Sea Turtles Prepared by IUCN/SSC Marine Turtle Specialist Group Edited by Karen L. Eckert Karen A. Bjorndal F. Alberto Abreu-Grobois M. Donnelly
Research and Management Techniques for the Conservation of Sea Turtles Prepared by IUCN/SSC Marine Turtle Specialist Group Edited by Karen L. Eckert Karen A. Bjorndal F. Alberto Abreu-Grobois M. Donnelly
UPSTART BAY FIELD RESEARCH
 UPSTART BAY FIELD RESEARCH UPDATE #5 The Rivers to Reef to Turtles Project On 14 June we all met at our reference site in Upstart Bay to sample the environment and turtles for the Rivers to Reef to Turtles
UPSTART BAY FIELD RESEARCH UPDATE #5 The Rivers to Reef to Turtles Project On 14 June we all met at our reference site in Upstart Bay to sample the environment and turtles for the Rivers to Reef to Turtles
SEA TURTLE MOVEMENT AND HABITAT USE IN THE NORTHERN GULF OF MEXICO
 SEA TURTLE MOVEMENT AND HABITAT USE IN THE NORTHERN GULF OF MEXICO Kristen M. Hart, Ph.D., Research Ecologist, USGS Wetland and Aquatic Research Center, Davie, FL Margaret M. Lamont, Ph.D., Biologist,
SEA TURTLE MOVEMENT AND HABITAT USE IN THE NORTHERN GULF OF MEXICO Kristen M. Hart, Ph.D., Research Ecologist, USGS Wetland and Aquatic Research Center, Davie, FL Margaret M. Lamont, Ph.D., Biologist,
POP : Marine reptiles review of interactions and populations
 POP2015-06: Marine reptiles review of interactions and populations Dan Godoy Karearea Consultants Department of Conservation CSP technical working group presentation: research results 22 September 2016
POP2015-06: Marine reptiles review of interactions and populations Dan Godoy Karearea Consultants Department of Conservation CSP technical working group presentation: research results 22 September 2016
OVERVIEW OF THE RESEARCH ACTIVITIES AT SEAFDEC-MFRDMD
 OVERVIEW OF THE RESEARCH ACTIVITIES AT SEAFDEC-MFRDMD INTERNATIONAL SEMINAR S-2 Sustainable Production of Fisheries Resources in Southeast Asia 15 December 2011 INTRODUCTION MFRDMD is the 4 th department
OVERVIEW OF THE RESEARCH ACTIVITIES AT SEAFDEC-MFRDMD INTERNATIONAL SEMINAR S-2 Sustainable Production of Fisheries Resources in Southeast Asia 15 December 2011 INTRODUCTION MFRDMD is the 4 th department
Final Report The People s Trust for Endangered Species Project: Conservation genetics and migratory patterns of sea turtles in Southern Brazil
 Final Report The People s Trust for Endangered Species Project: Conservation genetics and migratory patterns of sea turtles in Southern Brazil Project Team M.Sc. Maíra Carneiro Proietti M.Sc. Júlia Wiener
Final Report The People s Trust for Endangered Species Project: Conservation genetics and migratory patterns of sea turtles in Southern Brazil Project Team M.Sc. Maíra Carneiro Proietti M.Sc. Júlia Wiener
DRAFT Kobe II Bycatch Workshop Background Paper. Sea Turtles
 IOTC-2010-WPEB-Inf11 DRAFT Kobe II Bycatch Workshop Background Paper Sea Turtles In addition to other anthropogenic activities such as egg predation, directed harvest, and coastal development, the incidental
IOTC-2010-WPEB-Inf11 DRAFT Kobe II Bycatch Workshop Background Paper Sea Turtles In addition to other anthropogenic activities such as egg predation, directed harvest, and coastal development, the incidental
Dive-depth distribution of. coriacea), loggerhead (Carretta carretta), olive ridley (Lepidochelys olivacea), and
, loggerhead (Carretta carretta), olive ridley (Lepidochelys olivacea), and") 189 Dive-depth distribution of loggerhead (Carretta carretta) and olive ridley (Lepidochelys olivacea) sea turtles in the central North Pacific: Might deep longline sets catch fewer turtles? Jeffrey J.
189 Dive-depth distribution of loggerhead (Carretta carretta) and olive ridley (Lepidochelys olivacea) sea turtles in the central North Pacific: Might deep longline sets catch fewer turtles? Jeffrey J.
The state of conservation of sea turtles in the Mediterranean- case study of Greece
 The state of conservation of sea turtles in the Mediterranean- case study of Greece Panagiota Theodorou Conservation Coordinator ARCHELON, the Sea Turtle Protection Society of Greece Greece www.archelon.gr
The state of conservation of sea turtles in the Mediterranean- case study of Greece Panagiota Theodorou Conservation Coordinator ARCHELON, the Sea Turtle Protection Society of Greece Greece www.archelon.gr
Adani. Appendix B11 Marine Turtle Nesting Surveys, Abbot Beach
 Adani Appendix B11 Marine Turtle Nesting Surveys, Abbot Beach Terminal 0 Environmental Impact Statement Abbot Point Coal Terminal 0 EIS Adani Table of Contents Section 1 Marine Turtle Nesting Surveys Abbot
Adani Appendix B11 Marine Turtle Nesting Surveys, Abbot Beach Terminal 0 Environmental Impact Statement Abbot Point Coal Terminal 0 EIS Adani Table of Contents Section 1 Marine Turtle Nesting Surveys Abbot
MARINE TURTLE RESOURCES OF INDIA. Biotechnology, Loyola College, Chennai National Biodiversity Authority, Chennai
 MARINE TURTLE RESOURCES OF INDIA M.C. John Milton 1 and K. Venkataraman 2 1 P.G. & Research Department of Advanced Zoology and Biotechnology, Loyola College, Chennai - 600 034 2 National Biodiversity Authority,
MARINE TURTLE RESOURCES OF INDIA M.C. John Milton 1 and K. Venkataraman 2 1 P.G. & Research Department of Advanced Zoology and Biotechnology, Loyola College, Chennai - 600 034 2 National Biodiversity Authority,
Interaction Between Sea Turtle and Human Activities: A Survey on Local Communities at Kuala Lawas off Brunei Bay. 2.0 OBJECTIVES 1.
 Regional Meeting on Conservation and Management of Sea Turtle Foraging Habitats in Southeast Asian Water. (Japanese Trust Fund V Program) 22-24 October 13 AnCasa Hotel, Kuala Lumpur. Interaction Between
Regional Meeting on Conservation and Management of Sea Turtle Foraging Habitats in Southeast Asian Water. (Japanese Trust Fund V Program) 22-24 October 13 AnCasa Hotel, Kuala Lumpur. Interaction Between
Sea Turtle, Terrapin or Tortoise?
 Sea Turtles Sea Turtle, Terrapin or Tortoise? Based on Where it lives (ocean, freshwater or land) Retraction of its flippers and head into its shell All 3 lay eggs on land All 3 are reptiles Freshwater
Sea Turtles Sea Turtle, Terrapin or Tortoise? Based on Where it lives (ocean, freshwater or land) Retraction of its flippers and head into its shell All 3 lay eggs on land All 3 are reptiles Freshwater
Profile of the. CA/OR Drift Gillnet Fishery. and its. Impacts on Marine Biodiversity
 Profile of the CA/OR Drift Gillnet Fishery and its Impacts on Marine Biodiversity Todd Steiner Turtle Island Restoration Network History of CA/OR Drift Gillnet Fishery 1977 S. CA coastal harpoon & set
Profile of the CA/OR Drift Gillnet Fishery and its Impacts on Marine Biodiversity Todd Steiner Turtle Island Restoration Network History of CA/OR Drift Gillnet Fishery 1977 S. CA coastal harpoon & set
Sea Turtle Conservation
 Sea Turtle Conservation Volunteer Information Guide Index Introduction 2 Sample Volunteer Schedule 9 Volunteer 3 What s Next? 10 Roles and Commitments 5 Recommended Pre-Departure Reading 11 Our Commitment
Sea Turtle Conservation Volunteer Information Guide Index Introduction 2 Sample Volunteer Schedule 9 Volunteer 3 What s Next? 10 Roles and Commitments 5 Recommended Pre-Departure Reading 11 Our Commitment
May 7, degrees and no sign of slowing down, the clearing of Jamursba Medi Beach in
 May 7, 1984. 95 degrees and no sign of slowing down, the clearing of Jamursba Medi Beach in the Bird s Head Peninsula, Indonesia, reveals a gold sand beach and vast outstretches of turquoise water. The
May 7, 1984. 95 degrees and no sign of slowing down, the clearing of Jamursba Medi Beach in the Bird s Head Peninsula, Indonesia, reveals a gold sand beach and vast outstretches of turquoise water. The
Nature Watch. Tracking Turtles through Time and Space. Kartik Shanker I ARTICLE
 Nature Watch Tracking Turtles through Time and Space Kartik Shanker Marine turtles are known to migrate several thousands of kilometres between their feeding and breeding grounds.. These migrations have
Nature Watch Tracking Turtles through Time and Space Kartik Shanker Marine turtles are known to migrate several thousands of kilometres between their feeding and breeding grounds.. These migrations have
Green Turtle (Chelonia mydas) Foraging and Nesting Aggregations in the Caribbean and Atlantic: Impact of Currents and Behavior on Dispersal
 Foraging and Nesting Aggregations in the Caribbean and Atlantic: Impact of Currents and Behavior on Dispersal") Journal of Heredity 2006:97(4):346 354 doi:10.1093/jhered/esl004 Advance Access publication June 16, 2006 Green Turtle (Chelonia mydas) Foraging and Nesting Aggregations in the Caribbean and Atlantic:
Journal of Heredity 2006:97(4):346 354 doi:10.1093/jhered/esl004 Advance Access publication June 16, 2006 Green Turtle (Chelonia mydas) Foraging and Nesting Aggregations in the Caribbean and Atlantic:
Morning Census Protocol
 Morning Census Protocol Playa Norte Marine Turtle Conservation Click to edit Master subtitle style & Monitoring Programme All photographic images within are property of their copyrights and may only be
Morning Census Protocol Playa Norte Marine Turtle Conservation Click to edit Master subtitle style & Monitoring Programme All photographic images within are property of their copyrights and may only be
Types of Data. Bar Chart or Histogram?
 Types of Data Name: Univariate Data Single-variable data where we're only observing one aspect of something at a time. With single-variable data, we can put all our observations into a list of numbers.
Types of Data Name: Univariate Data Single-variable data where we're only observing one aspect of something at a time. With single-variable data, we can put all our observations into a list of numbers.
B E L I Z E Country Report. WIDECAST AGM FEB 2, 2013 Linda Searle ><> Country Coordinator
 B E L I Z E Country Report WIDECAST AGM FEB 2, 2013 Linda Searle > Country Coordinator OVERVIEW Happy Anniversary! Belize Sea Turtle Conservation Network Turtle Projects Historical Importance Threats
B E L I Z E Country Report WIDECAST AGM FEB 2, 2013 Linda Searle > Country Coordinator OVERVIEW Happy Anniversary! Belize Sea Turtle Conservation Network Turtle Projects Historical Importance Threats
INDIA. Sea Turtles along Indian coast. Tamil Nadu
 Dr. A. Murugan Suganthi Devadason Marine Research Institute 44-Beach Road, Tuticorin-628 001 Tamil Nadu, India Tel.: +91 461 2323007, 2336487 Fax: +91 461 2325692 E-mail: muruganrsa@sancharnet sancharnet.in
Dr. A. Murugan Suganthi Devadason Marine Research Institute 44-Beach Road, Tuticorin-628 001 Tamil Nadu, India Tel.: +91 461 2323007, 2336487 Fax: +91 461 2325692 E-mail: muruganrsa@sancharnet sancharnet.in
Green turtles in the Gulf of Venezuela
 Green turtles in the Gulf of Venezuela Gaby Montiel-Villalobos Kate Rodríguez-Clark Hector Barrios-Garrido Alberto Abreu-Grobois, Rodrigo Lazo WIDECAST AGM Baltimore, MD February 2, 2013 Instituto Venezolano
Green turtles in the Gulf of Venezuela Gaby Montiel-Villalobos Kate Rodríguez-Clark Hector Barrios-Garrido Alberto Abreu-Grobois, Rodrigo Lazo WIDECAST AGM Baltimore, MD February 2, 2013 Instituto Venezolano
Endangered Species Origami
 Endangered Species Origami For most of the wild things on Earth, the future must depend upon the conscience of mankind ~ Dr. Archie Carr, father of modern marine turtle biology and conservation Humpback
Endangered Species Origami For most of the wild things on Earth, the future must depend upon the conscience of mankind ~ Dr. Archie Carr, father of modern marine turtle biology and conservation Humpback
A Reading A Z Level R Leveled Book Word Count: 1,564. Sea Turtles
 A Reading A Z Level R Leveled Book Word Count: 1,564 Sea Turtles SeaTurtles Table of Contents Introduction...4 Types of Sea Turtles...6 Physical Appearance...12 Nesting...15 Hazards....20 Protecting Sea
A Reading A Z Level R Leveled Book Word Count: 1,564 Sea Turtles SeaTurtles Table of Contents Introduction...4 Types of Sea Turtles...6 Physical Appearance...12 Nesting...15 Hazards....20 Protecting Sea
GENETIC STRUCTURE AND DIVERSITY OF GREEN TURTLES (Chelonia mydas) FROM TWO ROOKERIES IN THE SOUTH CHINA SEA
 FROM TWO ROOKERIES IN THE SOUTH CHINA SEA") Journal of Sustainability Science and Management Special Issue Number 1: The International Seminar on the Straits of Malacca ISSN: 1823-8556 Penerbit UMT GENETIC STRUCTURE AND DIVERSITY OF GREEN TURTLES
Journal of Sustainability Science and Management Special Issue Number 1: The International Seminar on the Straits of Malacca ISSN: 1823-8556 Penerbit UMT GENETIC STRUCTURE AND DIVERSITY OF GREEN TURTLES
Sixth Meeting of the IAC Conference of the Parties
 Sixth Meeting of the IAC Conference of the Parties The Sixth Meeting of the IAC Conference of the Parties (COP6) was held in Galapagos, Ecuador, from June 26-28, 2013. The meeting discussed proposals for
Sixth Meeting of the IAC Conference of the Parties The Sixth Meeting of the IAC Conference of the Parties (COP6) was held in Galapagos, Ecuador, from June 26-28, 2013. The meeting discussed proposals for
Tour de Turtles: It s a Race for Survival! Developed by Gayle N Evans, Science Master Teacher, UFTeach, University of Florida
 Tour de Turtles: It s a Race for Survival! Developed by Gayle N Evans, Science Master Teacher, UFTeach, University of Florida Length of Lesson: Two or more 50-minute class periods. Intended audience &
Tour de Turtles: It s a Race for Survival! Developed by Gayle N Evans, Science Master Teacher, UFTeach, University of Florida Length of Lesson: Two or more 50-minute class periods. Intended audience &
Diane C. Tulipani, Ph.D. CBNERRS Discovery Lab July 15, 2014 TURTLES
 Diane C. Tulipani, Ph.D. CBNERRS Discovery Lab July 15, 2014 TURTLES How Would You Describe a Turtle? Reptile Special bony or cartilaginous shell formed from ribs Scaly skin Exothermic ( cold-blooded )
Diane C. Tulipani, Ph.D. CBNERRS Discovery Lab July 15, 2014 TURTLES How Would You Describe a Turtle? Reptile Special bony or cartilaginous shell formed from ribs Scaly skin Exothermic ( cold-blooded )
Marine Turtle Surveys on Diego Garcia. Prepared by Ms. Vanessa Pepi NAVFAC Pacific. March 2005
 Marine Turtle Surveys on iego Garcia Prepared by Ms. Vanessa Pepi NAVFAC Pacific March 2005 Appendix K iego Garcia Integrated Natural Resources Management Plan April 2005 INTROUCTION This report describes
Marine Turtle Surveys on iego Garcia Prepared by Ms. Vanessa Pepi NAVFAC Pacific March 2005 Appendix K iego Garcia Integrated Natural Resources Management Plan April 2005 INTROUCTION This report describes
Congratulations on the completion of your project that was supported by The Rufford Small Grants Foundation.
 The Rufford Small Grants Foundation Final Report Congratulations on the completion of your project that was supported by The Rufford Small Grants Foundation. We ask all grant recipients to complete a Final
The Rufford Small Grants Foundation Final Report Congratulations on the completion of your project that was supported by The Rufford Small Grants Foundation. We ask all grant recipients to complete a Final
COCA-LOCA : Connectivity of Loggerhead turtle (Caretta caretta) in Western Indian Ocean, implementation of local and regional management measures
 in Western Indian Ocean, implementation of local and regional management measures") COCA-LOCA : Connectivity of Loggerhead turtle (Caretta caretta) in Western Indian Ocean, implementation of local and regional management measures Mayeul Dalleau, Maxime Lalire, Céline Tardy, Suaad Al Harthi,
COCA-LOCA : Connectivity of Loggerhead turtle (Caretta caretta) in Western Indian Ocean, implementation of local and regional management measures Mayeul Dalleau, Maxime Lalire, Céline Tardy, Suaad Al Harthi,
BBRG-5. SCTB15 Working Paper. Jeffrey J. Polovina 1, Evan Howell 2, Denise M. Parker 2, and George H. Balazs 2
 SCTB15 Working Paper BBRG-5 Dive-depth distribution of loggerhead (Carretta carretta) and olive ridley (Lepidochelys olivacea) turtles in the central North Pacific: Might deep longline sets catch fewer
SCTB15 Working Paper BBRG-5 Dive-depth distribution of loggerhead (Carretta carretta) and olive ridley (Lepidochelys olivacea) turtles in the central North Pacific: Might deep longline sets catch fewer
UPSTART BAY FIELD RESEARCH
 WWF-AUS / SEAN HOOBIN UPSTART BAY FIELD RESEARCH UPDATE #12 The Rivers to Reef to Turtles Project We all met again at our primary study site in Upstart Bay to sample the environment and turtles for the
WWF-AUS / SEAN HOOBIN UPSTART BAY FIELD RESEARCH UPDATE #12 The Rivers to Reef to Turtles Project We all met again at our primary study site in Upstart Bay to sample the environment and turtles for the
NETHERLANDS ANTILLES ANTILLAS HOLANDESAS
 THE AD HOC DATA REPORT EL REPORTE DE DATOS AD HOC FOR THE COUNTRY OF POR EL PAIS DE NETHERLANDS ANTILLES ANTILLAS HOLANDESAS PREPARED BY/ PREPARADO POR GERARD VAN BUURT Western Atlantic Turtle Symposium
THE AD HOC DATA REPORT EL REPORTE DE DATOS AD HOC FOR THE COUNTRY OF POR EL PAIS DE NETHERLANDS ANTILLES ANTILLAS HOLANDESAS PREPARED BY/ PREPARADO POR GERARD VAN BUURT Western Atlantic Turtle Symposium
GUIDELINES FOR APPROPRIATE USES OF RED LIST DATA
 GUIDELINES FOR APPROPRIATE USES OF RED LIST DATA The IUCN Red List of Threatened Species is the world s most comprehensive data resource on the status of species, containing information and status assessments
GUIDELINES FOR APPROPRIATE USES OF RED LIST DATA The IUCN Red List of Threatened Species is the world s most comprehensive data resource on the status of species, containing information and status assessments
Costa Rica Turtle Conservation
 Costa Rica Turtle Conservation Visit the tropical beaches of Costa Rica and play your part in the conservation and preservation of some of the ocean's most recognisable inhabitants, turtles. Set between
Costa Rica Turtle Conservation Visit the tropical beaches of Costa Rica and play your part in the conservation and preservation of some of the ocean's most recognisable inhabitants, turtles. Set between
Representation, Visualization and Querying of Sea Turtle Migrations Using the MLPQ Constraint Database System
 Representation, Visualization and Querying of Sea Turtle Migrations Using the MLPQ Constraint Database System SEMERE WOLDEMARIAM and PETER Z. REVESZ Department of Computer Science and Engineering University
Representation, Visualization and Querying of Sea Turtle Migrations Using the MLPQ Constraint Database System SEMERE WOLDEMARIAM and PETER Z. REVESZ Department of Computer Science and Engineering University
Great Barrier Reef. By William Lovell, Cade McNamara, Ethan Gail
 Great Barrier Reef By William Lovell, Cade McNamara, Ethan Gail Marine biome Characteristics Covers about 70% of earth one cup of salt per gallon of water Over 1 million species discovered Importance Provides
Great Barrier Reef By William Lovell, Cade McNamara, Ethan Gail Marine biome Characteristics Covers about 70% of earth one cup of salt per gallon of water Over 1 million species discovered Importance Provides
Green turtle (Chelonia mydas) mixed stocks in the southwestern Atlantic, as revealed by
 mixed stocks in the southwestern Atlantic, as revealed by") Green turtle (Chelonia mydas) mixed stocks in the southwestern Atlantic, as revealed by mtdna haplotypes and drifter trajectories. Maíra Carneiro Proietti 1 ; Júlia Wiener Reisser 1 ; Paul Gerhard Kinas
Green turtle (Chelonia mydas) mixed stocks in the southwestern Atlantic, as revealed by mtdna haplotypes and drifter trajectories. Maíra Carneiro Proietti 1 ; Júlia Wiener Reisser 1 ; Paul Gerhard Kinas
Green Turtles in Peninsular Malaysia 40 YEARS OF SEA TURTLE CONSERVATION EFFORTS: WHERE DID WE GO WRONG? Olive Ridley Turtles in Peninsular Malaysia
 40 YEARS OF SEA TURTLE CONSERVATION EFFORTS: WHERE DID WE GO WRONG? (Did we go wrong?) Green Turtles in Peninsular Malaysia Lessons learnt and the way forward By Kamaruddin Ibrahim (TUMEC, DoFM) Dionysius
40 YEARS OF SEA TURTLE CONSERVATION EFFORTS: WHERE DID WE GO WRONG? (Did we go wrong?) Green Turtles in Peninsular Malaysia Lessons learnt and the way forward By Kamaruddin Ibrahim (TUMEC, DoFM) Dionysius
Part II: Species and species groups. Chapter 15. Vulnerability of marine reptiles in the Great Barrier Reef to climate change
 Chapter 15 Vulnerability of marine reptiles in the Great Barrier Reef to climate change Mark Hamann, Colin J Limpus and Mark A Read 15.1 Introduction 15.1.1 Marine reptiles Marine reptiles are an important
Chapter 15 Vulnerability of marine reptiles in the Great Barrier Reef to climate change Mark Hamann, Colin J Limpus and Mark A Read 15.1 Introduction 15.1.1 Marine reptiles Marine reptiles are an important
Status of leatherback turtles in India
 Indian Ocean SouthEast Asian Leatherback Turtle Assessment IOSEA Marine Turtle MoU 2006 Status of leatherback turtles in India By BC Choudhury 1. The legal protection status for leatherback turtles 1.1.
Indian Ocean SouthEast Asian Leatherback Turtle Assessment IOSEA Marine Turtle MoU 2006 Status of leatherback turtles in India By BC Choudhury 1. The legal protection status for leatherback turtles 1.1.
Universities Research Journal 2011, Vol. 4, No. 2
 Universities Research Journal 2011, Vol. 4, No. 2 Comparative Study on Hatching Rate and Incubation Period of Sea Turtles from Kadongalay Island and Thameehla Island in Ayeyrawady Region and Oyster Island
Universities Research Journal 2011, Vol. 4, No. 2 Comparative Study on Hatching Rate and Incubation Period of Sea Turtles from Kadongalay Island and Thameehla Island in Ayeyrawady Region and Oyster Island