Review of the Miocene long-snouted dolphin Priscodelphinus cristatus. eurhinodelphinids. by Olivier LAMBERT
|
|
|
- Todd Ball
- 5 years ago
- Views:
Transcription
 and phylogeny among eurhinodelphinids by Olivier LAMBERT L a m b e rt, O., 2005. Review o f the Miocene long-snouted dolphin Priscodelphinus cristatus DU Bus, 1872 (Cetacea, Odontoceti) and phylogeny among eurhinodelphinids.")
 (Cetacea, Odontoceti, Eurhinodelphinidae), recognized in the area o f Antwerp (North o f Belgium,")
1 BULLETIN DE L INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE SCIENCES DE LA TERRE, 75: , 2005 BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETENSCHAPPEN AARDWETENSCHAPPEN, 75: , 2005 Review of the Miocene long-snouted dolphin Priscodelphinus cristatus d u Bus, 1872 (Cetacea, Odontoceti) and phylogeny among eurhinodelphinids by Olivier LAMBERT L a m b e rt, O., Review o f the Miocene long-snouted dolphin Priscodelphinus cristatus DU Bus, 1872 (Cetacea, Odontoceti) and phylogeny among eurhinodelphinids. Bulletin de l'institut royal des Sciences naturelles de Belgique, Sciences de la Terre 75: , 4 pis., 10 figs., 3 tables; Bruxelles-Brussel, March 31, ISSN A bstract The Miocene long-snouted dolphin species Eurhinodelphis cristatus (sensu A b el, 1902) (Cetacea, Odontoceti, Eurhinodelphinidae), recognized in the area o f Antwerp (North o f Belgium, southern margin o f the North Sea Basin) and the east coast o f the USA (Maryland and Virginia), is re-described, including several undescribed specimens associated with ear bones and teeth. The systematic affinities o f this species with other members o f the family Eurhinodelphinidae are investigated, leading to its inclusion in the new genus Xiphiacetus, together with the species Eurhinodelphis bossi (sensu K e llo g g, 1925). The asymmetry o f the skull is discussed, especially for a strange specimen with an asymmetrical basicranium interpreted as the consequence o f pathology. Thanks to recently found specimens, the stratigraphie range o f Xiphiacetus cristatus might be extended in the Upper Miocene, widening the Eurhinodelphinidae distribution, previously limited to an Upper Oligocene - Middle Miocene interval. The description in X. cristatus o f several structures related to the telescoping o f the skull, hearing, and the feeding apparatus, considered as derived compared to other eurhinodelphinids, allows suggestions concerning evolutionary trends inside the family. A cladistic analysis is undertaken in a way to examine the phylogenetic relationships between the best-known eurhinodelphinid genera. The main results o f the analysis are a sister-group relationship between Schizodelphis + Xiphiacetus and Ziphiodelphis + (Mycteriacetus + Argyrocetus), and a more sternward Eurhinodelphis. This topology leads to some hypotheses concerning the evolution o f the habitat of the eurhinodelphinids. An appendix contains remarks about the systematic status o f several taxa usually included in, or related to, the family Eurhinodelphinidae. Key-words: Eurhinodelphinidae, taxonomy, Xiphiacetus cristatus, evolutionary trends, phylogeny R ésum é L espèce de dauphin longirostre miocène Eurhinodelphis cristatus (sensu A b el, 1902) (Cetacea, Odontoceti, Eurhinodelphinidae), identifiée dans la région d Anvers (nord de la Belgique, bord sud du Bassin de la Mer du Nord) et sur la côte est des Etats-Unis (Maryland et Virginie), est re-décrite, en incluant plusieurs spécimens non encore décrits, comprenant des os de l oreille et des dents. Les affinités systématiques de cette espèce avec les autres membres de la famille Eurhinodelphinidae sont investiguées, menant à son inclusion dans le nouveau genre Xiphiacetus, auquel est également rapportée l espèce Eurhinodelphis bossi (sensu K e llo g g, 1925). L asymétrie du crâne est commentée, en particulier pour un étrange spécimen muni d un basicrâne asymétrique interprété comme la conséquence d une pathologie. Grâce à des spécimens récemment découverts, l extension stratigraphique de Xiphiacetus cristatus pourrait être prolongée dans le Miocène supérieur, allongeant la distribution des Eurhinodelphinidae préalablement limitée à un intervalle Oligocène supérieur - Miocène moyen. La description chez X. cristatus de plusieurs structures liées au téléscopage du crâne, à l'audition, et à l appareil nutritif, considérées comme dérivées par rapport aux autres eurhinodelphinidés, permet quelques suggestions à propos de tendances évolutives au sein de la famille. Une analyse cladistique est entreprise afin d examiner les relations de parenté entre les genres les mieux connus d eurhinodelphinidés. Les résultats principaux de l analyse sont une relation de groupes-frères entre Schizodelphis + Xiphiacetus et Ziphiodelphis + (Mycteriacetus + Argyrocetus), et une position plus basale pour Eurhinodelphis. Cette topologie permet quelques hypothèses concernant l évolution de l habitat des eurhinodelphinidés. Un appendice contient quelques remarques à propos du statut systématique de plusieurs taxa habituellement inclus dans, ou apparentés à, la famille Eurhinodelphinidae. Mots-clefs: Eurhinodelphinidae, taxinomie, Xiphiacetus cristatus, tendances évolutives, phylogénie Introduction In 1872, d u Bus shortly described several species in the genera of long-snouted dolphins E urhinodelphis d u Bus, 1867 and P riscodelphinus L e id y, 1851 (Cetacea, Odontoceti, Eurhinodelphinidae), from the Miocene of Antwerp (North of Belgium), among them, the species P riscodelphinus pro d u ctu s and P. cristatus. The new combination E urhinodelphis cristatus was proposed by A b e l (1902) on the basis of eight skulls and partial skulls referred to these species; the holotype of E. cristatus is associated with all the cervical and several thoracic vertebrae and ribs. Later, the presence of the species was suspected in the Miocene of Portugal ( M a t a, ), and recognized in the Miocene of the eastern coast of the USA, Calvert Formation ( M y r ic k, 1979, unpublished thesis; M u iz o n, 1988a), and of Italy (B ia n u c c i et al., 1994). Additional specimens from Belgium and a comparison with specimens from the Calvert Formation allow a more detailed description and a systematic revision of E. crista-


2 212 Olivier LAMBERT tus (sensu A b e l, ). As suggested by L a m b e r t ( ), this species is referred here to a new genus together with E. bossi K e l l o g g, The asymmetry of the skull is discussed, especially for a strange individual with an asymmetric basicranium. The stratigraphie data obtained with several recently found Belgian skulls might lead to a refining of the stratigraphie range o f the species. Comments are given about evolutionary trends observed among the eurhinodelphinids. The general phylogenetic study of the odontocetes by M u iz o n ( ) briefly commented the relationships inside the family Eurhinodelphinidae; a more detailed study, using parsimony software, is undertaken, taking account of the new taxonomy within the family. M aterial and m ethods Abbreviations. IRSNB: Institut royal des Sciences naturelles de Belgique, Brussels; M: Fossil mammals collection o f types and figured specimens from the IRSNB; MGPD: Museo di Geología e Paleontología dell Universita di Padova, Italy; MNHN: Muséum national d Histoire naturelle, Paris, France; MP: Museo di Storia Naturale e del Territorio Certosa di Calci dell Università di Pisa, Italy; USNM: United States National Museum o f Natural History, Smithsonian Institution, Washington DC, USA; YPM: Peabody Museum of Natural History, Yale University, New Haven, USA; ZMA: Zoologisch Museum Amsterdam, The Netherlands. Terminology. The terminology for cranial, ear bones and vertebral anatomy is mainly taken from: F o r d y c e (1994), K a s u y a (1973), M u iz o n (1984, 1988a), and R o m m e l (1990). The orientations of the tympanic bulla and periotic are simplified in the descriptions compared to the actual anatomical position on the basicranium. The long axis of the tympanic is considered as anteroposterior, with ventral surfaces of inner and outer posterior prominences indicating the horizontal plane. The anterior direction of the periotic is given by the longitudinal axis of the anterior process, and the horizontal ventral plane by the surface contacting the most ventral points of pars cochlearis and anterior process. Schematic drawings illustrating the measurements on the skull o f eurhinodelphinids are in L a m b e r t (2004, fig. D- T axonom y Order Cetacea B r is s o n, 1762 Suborder Odontoceti F lo w e r, 1867 Superfamily Eurhinodelphinoidea M u iz o n, 1988a Family Eurhinodelphinidae A b e l, 1901 Xiphiacetus n. gen. Etymology. From Xiphias, swordfish, and cetus, whale in ancient Greek; Xiphiacetus because of a rostrum longer than the mandible, a feature characterizing the swordfish. Gender: masculine. Type species. X. cristatus ( d u Bus, 1872) Included species. X. cristatus and X. bossi ( K e l l o g g, 1925) Diagnosis. Eurhinodelphinid genus differing from: - the closest genus Schizodelphis G e r v a is, 1861 in: more robust skull with more progressive elevation of the premaxillae towards the vertex; medial plate of the maxilla along the vertex less concave and less erected; thicker supraorbital process; rostrum generally relatively wider at its base; longer fossa for the postorbital lobe of the pterygoid sinus, usually longer than half the width of the orbit roof. - Ziphiodelphis D a l P ia z, 1908 in: narrower and thicker triangular part of the premaxilla medially to the premaxillary foramen, lacking the more regular flatness and lateral slope seen in Ziphiodelphis', mesorostral groove widely open at that level; posterolaterally shorter plate of the maxilla along the vertex, giving the posterodorsal outline of the skull a more rounded aspect in lateral view; longer and narrower vertex (width less than 30 per cent of the postorbital width); nasal longer than wide lacking an anterodorsal projection. - Eurhinodelphis DU Bus, 1867 (sensu L a m b e r t, 2004) in: maxillary part of the rostrum relatively shorter; dorsomedian portion o f the supraoccipital shield concave; less elevated and wider paroccipital process of the exoccipital with lower occipital condyles (ventral margin of the condyles much lower than the level of the floor of the temporal fossa); distinctly concave premaxillary sac fossa; zygomatic process o f the squamosal flatter in lateral view and wider in ventral view; presence of a fossa for the postorbital lobe of the pterygoid sinus on the ventral surface of the supraorbital process; pars cochlearis of the periotic anteromedially rounded; inner posterior prominence of the tympanic distinctly narrower than the outer prominence. - Argyrocetus L y d e k k e r, 1893 in: relatively wider face (ratio between the bizygomatic width of the skull and the length of the cranium from the antorbital notch to the occipital condyles > 1); more elevated vertex with nasal longer than wide, at the same level or lower than the frontal; more erected supraoccipital shield (Appendix 1 for comments about the content of the genus Argyrocetus). - Macrodelphinus W il s o n, 1935 in: more elevated and more transversely compressed vertex with nasal as long as wide or longer than wide, and frontal as long as, or shorter than the nasal; supraoccipital shield closer to the vertical. - Mycteriacetus L a m b e r t, 2004 in: relatively wider cranium (ratio between the bizygomatic width of the skull and the length of the face from the antorbital notch to the occipital condyles > 1); more elevated vertex with nasal at the same level or lower than the frontal, and frontal as long as, or shorter than the nasal; more erected supraoccipital shield.
 to Priscodelphinus, but that genus is now restricted to its type-species and holotype - a series of six undiagnostic")
 * 1872 Priscodelphinus cristatus d u B u s, p. 497. v. 1880 Priscodelphinus cristatus V a n B e n e d e n & G e r v a is, p. 495. v. 1902 Eurhinodelphis cristatus Ab el, pi.")
3 Review of Priscodelphinus cristatus and phylogeny 213 Comment. The holotype of Xiphiacetus cristatus was originally referred by d u B u s (1872) to Priscodelphinus, but that genus is now restricted to its type-species and holotype - a series of six undiagnostic vertebrae from New Jersey (P. harlani, L e id y, 1851)- and considered as Odontoceti incertae sedis (e.g., F o r d y c e & M u iz o n, 2001). Xiphiacetus cristatus (d u Bus, 1872) * 1872 Priscodelphinus cristatus d u B u s, p v Priscodelphinus cristatus V a n B e n e d e n & G e r v a is, p v Eurhinodelphis cristatus Ab el, pi. 15, figs 1, 2; pi. 16; pi. 17, fig. 3. v Eurhinodelphis cristatus A bf.l, pi. 19, fig. 3; pi. 20, figs 1, 2, 18-20; pi. 21, figs 2, 3, 9; pi. 22, figs 1, 2; pi. 23, figs Emended diagnosis. This species differs from Xiphiacetus bossi in the following combination of characters: major thickening of the maxilla on the supraorbital process; posterior margin of the maxilla on the cranium notched by a forwards indentation of the frontal and the supraoccipital laterally to the vertex; vertex acute and short in lateral view constituted by the posterodorsal edge of the frontals and the supraoccipital, vertical on its mediodorsal portion against the frontals; trapezoid dorsal surface of the frontals on the vertex plane and anteriorly sloping; relatively lower temporal fossa (less than 20 per cent of the bizygomatic width); deep and long fossa for the postorbital lobe o f the pterygoid sinus. Holotype. IRSNB 3234-M.361, a p a r tia l s k u ll la c k in g te e th, e a r b o n e s a n d b a s ic r a n iu m, a s s o c ia te d w ith th e s e v e n c e r v ic a l v e r te b r a e, s e v e n th o r a c ic v e r te b r a e, a n d s e v e ra l r ib s (fo u n d b e tw e e n 1861 a n d 1863, in d iv id u a l 1 in A b e l, 1902, p i. 15, f ig s 1, 2; se v e ra l p o s t- c r a n ia l elements are figured in A b e l, 1931, pi. 19, fig. 3; pi. 20, figs 1,2,18-20; pi. 21, figs 2, 3, 9; pi. 22, figs 1, 2; pi. 23, figs 8-10). Locus typicus. The holotype was found in Antwerp (Fig. 1), and the locality cited by A b e l (1902) is? 4e Section. This locality corresponds to a portion of the fortification belt built in the 1860 s around Antwerp (see map in V a n d e n B r o e c k, 1874), north of Berchem, southeastern suburb of Antwerp. However, the question mark of Abei lowers the interest of that information. Stratum typicum. No data are available for the holotype. However, several skulls from Antwerp and the skulls from the Calvert Formation (east coast of USA) are accompanied by more precise stratigraphie information. The species is limited to the Miocene, mainly Middle Miocene, with a possible extension in the Belgian Upper Miocene (see stratigraphie remarks below). Referred Belgian specimens. IRSNB 3241-M. 1893, a well preserved partial skull (found in 4e Section, Antwerp; individual 2 in A b e l, 1902, pi. 16); IRSNB 3242, fragments of skull (found in? 4e Section, Antwerp, ; individual 4 in A b e l, 1902, pi. 17, fig. 3); IRSNB 3237-M.1894, partial skull (found in? 4e Section, Antwerp, ; individual 5 in A b e l, 1902); IRSNB 3240-M.1895, partial skull (found in 4e Section, Antwerp, ; individual 6 in A b e l, 1902); IRSNB M.1896, partial skull (found in 1978 by G. Paredis in Borgerhout, eastern suburb of Antwerp, near the Stenen Brug bridge, during the construction of the motorway around the city); IRSNB 3227-M.1897, partial skull (with a label Eurhinodelphis cristatus, Abei, 1904 ); IRSNB 3236, partial skull (found in Antwerp area); IRSNB 8243-M.1898, partial skull (with a label Eurhinodelphis cristatus - Et.; Anversien, lettre M, 6 E Southern North Sea 10 Km The Netherlands Antwerp Kessel 51 N Belgium Fig. 1 Map of northern Belgium and location o f the two localities of the Belgian specimens o f Xiphiacetus cristatus, Antwerp and Kessel. Inset: southern North Sea Basin. Dashed line: southern limit of Neogene deposits in Belgium (modified after T a v e r n ie r & de H e in z e lin, 1963).
; IRSNB 3589, a fragment of the right side of the vertex (found in the Antwerp area); IRSNB M.")
; IRSNB 3245, fragmentary rostrum [found in Antwerp, 1861-1863, individual 17 of E urhinodelphis longirostris (sensu A b e l, 1902)];")
4 214 Olivier LAMBERT Loc.: Kessel-Expl. fév., mars, Reg. - I. G ; Kessel: Prov. Antwerp); IRSNB 8243, vertex and left supraorbital process (with a label isolé - Terrain Boldérien - Loc. Kessel - le 18 mars I. G. n 8243 ); IRSNB 3589, a fragment of the right side of the vertex (found in the Antwerp area); IRSNB M.1899, the maxillary part of a right supraorbital process (found in the Antwerp area); IRSNB M.1900, a right supraorbital process (found in the Antwerp area); IRSNB M. 1901, a right supraorbital process (found in the Antwerp area); IRSNB 3245, fragmentary rostrum [found in Antwerp, , individual 17 of E urhinodelphis longirostris (sensu A b e l, 1902)]; IRSNB M.1902, a fragmented right tympanic (found associated to a partial skull in 1974 by L. Peeters, near the locality of IRSNB M.1896 (Deume bridge)]. R eferred A m erican specim ens. The skulls from the Calvert Formation referred by M y r ic k (1979) to E urhinodelphis cristatus: USNM 21303, USNM , USNM 13470, USNM 24856, USNM , USNM 13436, USNM 21363, and USNM 21360; E. w hitm orei: USNM 25666; E. a sh b y i: USNM and USNM (partially suggested in M u iz o n, 1988a; detailed in L a m b e r t, 2004). F urther com m ents on specim ens p revio u sly reported to the species. IRSNB 3243-M.367 is a partial skull lacking the apical part of the rostrum, nasals and ear bones, from 4 Section, Vieux-Dieu (= Oude God, Mortsel). It is the holotype and only specimen of P riscodelphinus p ro d u c tus sensu d u Bus 1872, figured by V a n B e n e d e n & G e r v a is (1880, pi. 58, fig. 3), and revised by A b e l (1902) in E urhinodelphis cristatus (individual 3, pi. 18, fig. 3). It is referred to X iphiacetus bossi, as well as the fragments of skull IRSNB 3497 [ 4e Section, also placed by A b e l (1902) in E urhinodelphis cristatus, individual 7] ( L a m b e r t, in press). IRSNB 3443-M.368, a right squamosal with the corresponding paroccipital process of the exoccipital (from 3e section, Antwerp, individual 8 of E urhinodelphis cristatus in A b e l, 1902), is too fragmentary to be included in a defined species of eurhinodelphinid; it shows actually more similarities with E. longirostris, and is referred here to Eurhinodelphinidae aff. E urhinodelphis. Its association with three vertebrae (two cervicals and one thoracic) is judged doubtful. The partial skull of an odontocete from the late Miocene of Portugal referred to E urhinodelphis cf. cristatus by M a t a ( , fig. 1, pi. 1-3) lacks all diagnostic characters of the species and shares affinities with the kentriodontids ( E s t e v e n s, 2003; L a m b e r t, 2004). B ia n u c c i et al. (1994) identified a partial skull from the Miocene of the Pietra leccese (Apulia, Italy) as E urh in od elp his cristatus. Its state o f preservation precludes a specific attribution; it is referred here to X ip h ia cetus aff. bossi, according to the observations in L a m b e r t (2004). D escription S ku ll (PI. 1, PI. 2, Figs 1-3, PI. 3, Fig. 1; Figs 2-4) Prem axilla. The premaxilla is longer than the maxilla anteriorly, a feature observed on the small skull IRSNB M.1896: the premaxilla-maxilla suture reaches the ventral margin of the rostrum 450 mm anteriorly to its base (PI. 1, Fig. 2). However, this rostrum lacks its anterior portion, and consequently the length of the premaxillary portion cannot be evaluated. On better-preserved skulls from the Calvert Formation, the premaxilla-maxilla suture is completely fused anteriorly, precluding a quantification of the relative lengths of the maxilla and the premaxilla on the rostrum. In particular, the large skull USNM has a nearly complete rostrum with a rostral length of 970 mm, and a total length of the skull of 1170 mm (Fig. 2; Table 1), but without indications of the anterior part of the premaxilla-maxilla suture. A deep longitudinal groove follows the rostral maxilla-premaxilla suture from ca. 15 mm anterior to the antorbital notch, at the exit o f a dorsal infraorbital foramen; the groove last alveoli Fig. 2 Reconstruction o f the skull o f Xiphiacetus cristatus in dorsal view, mainly based on the nearly complete specimen USNM 21363, from the Calvert Formation, Middle Miocene of the east coast o f the USA. Scale bar = 200 mm.


5 Review o f Priscodelphinus cristatus and phylogeny 215 Table 1 Measurements (mm) on the skulls of Xiphiacetus cristatus from IRSNB and USNM. e indicates estimate, complete and no data. The numbers of the measurements refer to L am bert, 2004, fig nearly Holotype IRSNB M. 361 IRSNB M.1893 IRSNB M.1896 IRSNB M.1897 IRSNB M total length skull length base rostrum-anterior maxilla length anterior orbit-posterior skull length anterior supraoccipital-anterior orbit el length orbit e77 6. length temporal fossa - e width base rostrum el23 - el width premaxillae at base rostrum e80 e76 e IRSNB M width skull at level of postorbital processes e e width skull at level of zygomatic processes e width bony nares - 32 e30 e width nasals e34 e33 e maximal posterior premaxillary width el 13 e90 - e minimal posterior distance between maxillae 46 e40 e width between ventromedial margins exoccipitals el width between lateral margin of occipital condyles width between inner margins of occipital condyles height cranium height base rostrum - 71 e47 - e height temporal fossa height ventral margin of occipital condyles height occipital condyles e48-44 USNM USNM USNM USNM USNM USNM USNM USNM USNM USNM e200 _ 186 _ 171 _ e e77 - e65 - e el e e e49 e31 e e e e e el e l e
, innervating/vascularizing the anterior of the rostrum.")

6 216 Olivier LAMBERT nearly reaches the apex of the rostrum. Additionally to the eurhinodelphinids, a similar groove is present in other groups of Miocene long-snouted dolphins (e.g., platanistids, eoplatanistids) and in extant river dolphins (e.g., Platanista, Pontoporia), innervating/vascularizing the anterior of the rostrum. The slight posterior widening of the premaxilla on the rostrum increases mm before the base of the rostrum, together with a considerable flattening of the bone. In this widened area, the median portion of the premaxilla is often depressed relatively to the lateral portion; the degree of medial deepening is however variable. The premaxillary sac fossa is usually well excavated, and its elevation towards the vertex is only pronounced from the mid-length of the orbit. The posterior apex of the premaxilla contacts the frontal, as a thin process between the nasal and the median margin of the maxilla. The premaxilla is sometimes thick and wide at the level of the posterior margin of the bony nares. Maxilla. The maxilla is poorly exposed in dorsal view of the rostrum, with a slight lateral bulge ca. 200 mm anteriorly to the base of the rostrum. The lateral margins of the maxillae diverge towards the antorbital notches, widening the base of the rostrum; the lateral margin of the maxilla is extended on the supraorbital process by a short longitudinal crest, medial to the antorbital notch. On the skull USNM 21363, alveoli with a mean diameter of 8 mm and separation septa of 5-7 mm are present on the first 740 mm of the rostrum, interrupting 240 mm before the apex. This condition confirms that the apex of the rostrum was only constituted by the edentulous premaxillae, as it is the case in Eurhinodelphis cocheteuxi DU Bus, 1867, E. longirostris du Bus, 1872, m á Xiphiacetus bossi. Number and spacing of the alveoli are intraspeciflcally variable; for instance, on the smaller IRSNB M.1896, the bases of the crowns of successive anterior teeth nearly contact, with a diameter of 5 mm for the alveoli. This variability might be related to the age of the individuals. A striking feature o f Xiphiacetus cristatus is the frequent strong thickening of the maxilla on the supraorbital process, forming a longitudinally elongated dome, posterolateral to the antorbital notch. The development of the dome is variable, rarely absent, and sometimes reaching mm of height in lateral view (e.g., IRSNB M.1899, PI. 1, Figs 7-8). This structure stresses the median depression of the face due to the medial slope of the flat dorsal surface of the premaxillae. The dome shows similarities with the elevated frontal at the level of the supraorbital process on the Miocene platanistid Pomatodelphis, wider and lower than in the other Miocene platanistid Zarhachis (see K e l l o g g, 1924, 1959). This dome might correspond to an area of origin for rostral and facial muscles as seen in Hyperoodon-, the very high maxillary crest medial to the antorbital notch provides to this extant ziphiid genus a different orientation for the rostral and pars anterointernus muscles, inserting on the melon ( S c h e n k k a n, 1973). A b e l (1905, p. 118) suspected the elevation of the maxilla on the supraorbital processes of Xiphiacetus cristatus to be linked to a sexual dimorphism (males with thicker maxillae, in a way somewhat similar to Hyperoodon). M y r ic k (1979) could not find specimens lacking the thickened maxilla among American specimens that he referred to X. cristatus. Therefore, either all the American specimens represent males, or there is no sexual dimorphism for that feature. M y r ic k (1979) could however detect a good correlation between the relative age of the specimen and the elevation of its maxilla. Concerning the Belgian specimens, the maxilla is distinctly thicker in larger animals; the smallest skulls IRSNB M.1896 and IRSNB 3237-M.1894 lack a clear elevation, and the largest IRSNB 3241-M and IRSNB 3227-M.1897 have a high maxillary dome. The maxillary crests of Hyperoodon are also known to increase in size allometrically (H e y n in g, 1989). The posterior margin of the maxilla goes barely beyond the anterodorsal margin of the supraoccipital; the acute transverse crest is thus roughly rectilinear. This crest, made of the compression of the maxilla and frontal against the supraoccipital shield, is notched by a distinct indentation of the supraoccipital and frontal on the dorsal edge of the maxilla, laterally to the vertex (= maxillary fold sensu M y r ic k, 1979). This structure, absent in other eurhinodelphinid taxa, is present on all the skulls of Xiphiacetus cristatus for which this area is preserved, including USNM and USNM , respectively holotypes of Eurhinodelphis whitmorei and E. ashbyi sensu M y r ic k (1979). The maxilla sends a slightly concave median strip towards the frontal part of the vertex, which pinches, with the nasal, the posterior projection of the premaxilla. This median plate of the maxilla is less straightened up and more covering than in E. cocheteuxi, for which it reaches a vertical position. Nasal. The morphology of the nasal is variable; it is either longer than wide, or roughly square, always narrower and lower than the frontal. Its dorsal surface progressively curves anteroventrally, without distinct angle. The posterior vertical plate of the mesethmoid projects only weakly dorsally from the ventral margin of the nasals, which are therefore only partially retained anteriorly. Frontal. The dorsal surface of the frontal on the vertex is smooth and flat, anteriorly sloping, often with an acute and high posterodorsal edge, continuous with the transverse crest on the maxilla/supraoccipital. The degree o f penetration of the frontals between the nasals is variable. In ventral view, a deep and long fossa for the postorbital lobe of the pterygoid sinus incises the frontal along the posterior wall of the optic canal, in an anterolateral direction. Laterally, this fossa ends mm before the lateral margin of the supraorbital process. In some specimens, the fossa is so deep in the frontal (up to 10 mm) that it nearly reaches the maxilla dorsally. If present, the fossa is usually shallower in Xiphiacetus


7 Review o f Priscodelphinus cristatus and phylogeny 217 antorbital notch lacrimal maxilla infraorbital foramen?fossa for preorbital lobe of pterygoid sinus optic foramen fossa for postorbital lobe of sinus pterygoid cerebral cavity squamosal glenoid surface falciform process tympanosquamosal recess spiny process postglenoid process paroccipital process of exoccipital frontal palatine posterior lacerate foramen mark of pterygoid pterjp;oid sinus ossa vomer choana orbitosphenoid pterygoid sinus fossa in alisphenoid pterygoid? ventral aperture for foramen pseudo-ovale PMNVj foramen ovale carotid foramen parietal basioccipital crest occipital condyles Fig. 3 Schematic drawing of the right side of the basicranium of Xiphiacetus cristatus IRSNB 3241-M. 1893, Miocene of Antwerp, in ventral view. bossi, and laterally shorter in Schizodelphis spp. and Ziphiodelphis spp., never as developed as in X. cristatus; it is absent in Eurhinodelphis spp. Palatine. The anterior margin of the palatine is either wide and blunt, ca. 40 mm anterior to the antorbital notches, or slightly longer and more pointed. The lateral margin is roughly rectilinear. Pterygoid. The pterygoid sinus fossa excavates the pterygoid roughly until the level of the antorbital notches (with intraspecific variation; the fossa ends sometimes just posterior to the notches). The lateral lamina of the pterygoid is complete and strong, contacting the falciform process of the squamosal. No hamular process of the pterygoid is preserved on any of the specimens of this species. Jugal-lacrimal. The jugal-lacrimal complex is visible in lateral view, anterior to the preorbital process of the frontal, and sending a short posterior process between frontal and maxilla. In ventral view, the complex forms the U-shaped ventral part of the antorbital notch. The posterolateral suture between lacrimal and frontal follows the anterior margin of the poorly individualized and narrow optic groove. The lacrimal is excavated by a shallow fossa, which might have held the preorbital lobe of the pterygoid sinus. Supraoccipital. The dorsomedian portion of the supraoccipital is vertically applied on the elevated posterior margin of the frontals. More ventrally, this concave plate is less inclined, posteroventrally bending towards the occipital condyles. The lateral parts of the bone are convex, more protuberant. Exoccipital. The paroccipital process of the exoccipital is wide and low, giving a low position to the occipital condyles compared to Eurhinodelphis spp.; their ventral margin is much lower than the level of the floor of the temporal fossa. Basioccipital. The basioccipital basin is deep, limited by basioccipital crests forming an angle of about 55. The posteroventral extremity of the basioccipital crest is thick, and reaches the ventral level o f the exoccipital. Squamosal. The zygomatic process of the squamosal is high and long; the elongated postglenoid process is slightly more robust, with a rounded apex. The temporal fossa is reduced, longer than high, with the roof roughly at the same vertical level than the roof of the orbit. In



8 218 Olivier LAMBERT ventral view, the glenoid surface is wide and medially margined by a deep tympanosquamosal recess. The recess extends for a short distance dorsally to the glenoid surface, and dorsally to the base of the falciform process. The long anterior portion of the recess is supported by the zygomatic process, which is anteromedially widened. The falciform process of the squamosal is anteromedially directed, differing from the more laterally bent process in Eurhinodelphis cocheteuxi. Alisphenoid. The foramen ovale, anterolateral to the carotid foramen at the posterior margin of the alisphenoid, is anterolaterally followed by the path for the mandibular nerve V3 (PMNV3). This PMNV3 is a deep groove, which leads to a foramen piercing the falciform process of the squamosal in a laterodorsal direction, and emerging in the base of the temporal fossa (= foramen pseudo-ovale sensu F o r d y c e, 1994). This last part of the path is variable: for instance, on the skull IRSNB 3241-M (PI. 2, Fig. 1C; Fig. 3) the sulcus shows a bifurcation, with a second branch more anteriorly directed, probably exiting in the temporal fossa ventrally to the lateral lamina of the pterygoid. The PMNV3 ventrally overhangs a dorsoposteriorly extended round and smooth fossa on the alisphenoid (pterygoid sinus fossa in alisphenoid sensu F o r d y c e, 1994). This fossa, laterodorsally deepening, is connected to the fossa for the postorbital lobe of the pterygoid sinus on the frontal, confirming the functional interpretation o f this latter. Parietal. Several small foramina pierce the parietal posteriorly to the foramen ovale, but their position and number are variable and the correlation with the foramina identified by F o r d y c e (1994) in Waipatia was not successful. Asymmetry. The vertex is distinctly asymmetric: the suture between the frontals is posteriorly deflected on the left side. Asymmetry is also observed on the relative development of the two maxillae along the vertex. For instance, the large fragmentary skull IRSNB M.1897 has a right maxilla much higher and wider than the left, forming a large transverse protuberance between the vertex and the indentation of the supraoccipital and frontal described above. Furthermore, the deflection of the suture between the frontals is much pronounced on that skull; the left frontal is strongly narrowed posteriorly, and its posterodorsal edge is deeply excavated, forming a wide valley (PI. 3, Fig. 1A; Fig. 4). On this skull, the strong asymmetry of the vertex is curiously associated to asymmetric squamosals, an unusual feature in the odontocetes ( M e a d, 1975; H e y n in g, 1989). In lateral view, the left zygomatic process is more than 10 mm shorter than the right (measured from the rostral apex o f the zygomatic process to the ventral tip of the postglenoid process); it is also narrower in ventral view, and the roof of the tympanosquamosal recess is open on its anterior part, isolating the zygomatic process from the lateral wall of the cerebral cavity (PI. 3, Fig. IB). FIo w e l l (1925) suggested that the asymmetry observed in some pinnipeds, at the level of the shape and size of bony elements involved in the insertion of masseter and temporal muscles, could be caused by injury or disease, producing a weakening of these muscles on one side of the skull relatively to the other. The earlier in the life of the animal the injury occurs, the more the asymmetry is pronounced. On IRSNB 3227-M.1897, the clear difference of size and shape between the two squamosals, bones implied in the articulation of the mandible and in the insertion of masticatory muscles, might be explained by the development, before the end of the growth, of a deficiency of the masticatory muscles of the left side. In that case, the pathological asymmetry of the basicranium would be independent from the asymmetrical vertex. Teeth (PI. 2, Figs 3A-3C) Nine distal teeth are attached to the maxillae of the small skull IRSNB M The teeth have an average total length of 16 mm (6 mm for the root), and a maximal width at the base of the crown of 5 mm. The teeth nearly contact each other by the tip of the root and the base of the crown. The root is strongly flattened transversely with a roughly rectangular profile in lateral view, and an elongated dorsoposterior comer projecting towards the preceding tooth. The base of the crown is a slightly longitudinally elongated thick ring, quickly distally narrowing. The rest of the crown is longitudinally flattened. The median margin of the crown is rectilinear, sometimes slightly concave on the distal portion, while the lateral margin is parallel to the median for the first third of the crown, followed by a distinct angle towards the tip. Among extant odontocetes, the teeth of Pontoporia blainvillei show the most striking resemblances; for example, loose maxillary teeth from P. blainvillei ZMA (PI. 2, Fig. 4) show several characters in common with Xiphiacetus cristatus: small and numerous teeth frontal fragment of. premaxilla maxilla right frontal supraoccipital left frontal mesethmoid bony nares indentation of frontal maxilla prcmaxilla Fig. 4 Posterior part o f the cranium o f Xiphiacetus cristatus IRSNB 3227-M.1897, Miocene o f Antwerp, in anterodorsal view, showing the strong asymmetry of the bones o f the vertex. The nasals are lost.


 with a tympanic bulla. A recently discovered Belgian skull is also associated with a right tympanic bulla IRSNB M.1902. Periotic.")
9 Review o f Priscodelphinus cristatus and phylogeny 219 close one to the other (some of them were still attached with each other by the base of the crown); strongly transversely flattened root with a rectangular lateral aspect and a posterodorsal projection; thickened base of the crown; crown longitudinally flattened with the median margin rectilinear and an angle on the lateral margin. The similarities at the level of the teeth between those two species, both bearing an elongated rostrum and an individualized neck, might have a palaeoecological meaning (feeding behaviour), and should probably be interpreted as homoplasies. Ear bones (PI. 3, Figs 2-5; Fig. 5) The partial skull USNM is associated with a periotic, the skull USNM with a fragment of periotic, and the skull USNM (considered by M y r ic k, 1979 as Eurhinodelphis ashbyi and referred here to Xiphiacetus cristatus) with a tympanic bulla. A recently discovered Belgian skull is also associated with a right tympanic bulla IRSNB M Periotic. The right periotic USNM 21360, only lacking the medioposterior part of the pars cochlearis, has a total length of 35 mm. Its general morphology is close to Eurhinodelphis cocheteuxi, except its slenderness. A wide and deep articular facet, laterally and medially limited by longitudinal crests, excavates the ventral surface of the anterior process; the median crest is more pronounced. An anteroextemal sulcus is posterolaterally curving, incising the anterior process before the large lateral tuberosity. The latter bears a large mallear fossa, pars cochlearis fovea epitubaria anterior articular facçt fenestra ovalis suip^dial muscle fossa _ antero-extemal sulcus -... lateral tuberosity' mallear fossa' fenestra ovalis fossa incudis facial sulcus ß \ posterior articular facet hiatus epitympanicus Fig. 5 Periotics of Xiphiacetus cristatus from the Calvert Formation, Middle Miocene o f the east coast of the USA. A. ventral view o f the right periotic USNM 21360, with a portion o f the pars cochlearis missing. B. ventral view of the right periotic USNM 21363, lacking most of its pars cochlearis and anterior process, and showing the shortened posterior process. A anterior to a shallow but well-defined fossa incudis. The hiatus epitympanicus is pronounced, deeply cutting the lateral margin of the bone, without any articular rim. The dorsal protuberance of the anterior process is separated from the pars cochlearis by a transverse depression leaving from the median contact between the anterior process and the pars cochlearis towards the hiatus epitympanicus, on the dorsal face of the bone. The median surface of the anterior process is slightly depressed, cut by two small longitudinal sulci along the anterior articular facet (anterointemal sulci sensu F o r d y c e, 1994), and pierced by numerous tiny apical foramina. The anteromedial comer of the pars cochlearis is rounded in ventral view, and the anterior portion is narrower in medial view than in Eurhinodelphis cocheteuxi. The partially preserved internal auditory meatus, including the tractus spiralis foraminosus and the foramen singulare, is anteriorly pointed; its tip contains the dorsal opening of the facial canal; it is extended anteriorly by a narrow fissure nearly reaching the contact between the pars cochlearis and the anterior process. The posterior process is relatively long beyond the posteromedian limit of the stapedial muscle fossa, with a length corresponding to 70 per cent of the length of the pars cochlearis (measured from the anterior wall to the posteromedian limit of the stapedial muscle fossa). The concave posterior articular facet is posterolateroventrally curved. The dorsal face of the posterior process is roughly keeled: a wide longitudinal protuberance runs from the tip o f the process until the internal auditory meatus. Only the posterior process and the posterior part of the pars cochlearis are preserved on the right periotic associated with the skull USNM This fragment fits the periotic USNM for the shape of the pars cochlearis and the base of the posterior process, but the posterior process, which is complete, is much shorter, less than 37 per cent of the length of the pars cochlearis, and associated to a stronger posteroventral curve of its dorsal margin (PI. 3, Fig. 3; Fig. 5). The length of the posterior process is thus variable among the individuals of this species. A similar condition may also be present in Xiphiacetus bossi: the periotic USNM has a long posterior process (73 per cent of the length of the pars cochlearis), and the periotic USNM has a short posterior process (26 per cent of the length of the pars cochlearis). The progressive loss of bony contact between the periotic and the basicranium among the Delphinida (sensu M u iz o n, 1988b), leading to a better isolation of the ear bones from the skull, is related to the shortening o f the posterior process that contacts the squamosal and exoccipital on more sternward groups o f Cetacea. The reduction of the process is for instance particularly pronounced among the families Pontoporiidae and Iniidae (about 28 per cent of the length of the pars cochlearis in Pontoporia and 15 per cent in Inia). Tympanic bulla. None of the two known tympanic bullae of Xiphiacetus cristatus is complete: on both of them the posterior process is lacking. The left tympanic USNM
.")
, wider and less pointed than the one of Eurhinodelphis cocheteuxi.")
10 220 Olivier LAMBERT , although lacking a part of the outer lip and the sigmoid process, is better preserved in its anterior part. It has a total length of 35 mm and a maximal width of 21 mm, slightly smaller than IRSNB M.1902, with a width of 23 mm (PI. 3, Figs 4,5). The anterior extremity is rounded and wide in ventral view, similarly to Ziphiodelphis abeli, Z. sigmoideus, and Mycteriacetus bellunensis ( D \l P ia z, 1977, pi. 3, figs 6-14; P il l e r i, 1985, pi. 45, fig. a, pi. 48, fig. c), wider and less pointed than the one of Eurhinodelphis cocheteuxi. However, this character seems variable within the genus Xiphiacetus: tympanic bullae o fx. cristatus IRSNB M and X. bossi USNM are relatively more pointed (PI. 3, Fig. 4A; M u iz o n, 1988a, fig. 6a). The distinct median groove is widening and shallowing anteriorly, anterolaterally turning, and ending after 25 mm. It separates the outer posterior prominence from a narrower and posteriorly shorter inner posterior prominence. In E. cocheteuxi, both prominences are sub equal in width, and the median groove is deeper. The condition of Xiphiacetus, also observed in Mycteriacetus, Schizodelphis, and Ziphiodelphis, might constitute a synapomorphy, differentiating those genera from Eurhinodelphis (see phylogeny below). In medial view, the dorsal margin of the involucrum is high and parallel to the ventral margin for the posterior half of its length. After that, it is cut by a strong indentation, followed by a short platform and a progressive lowering until the anterior end of the bone. The ventral margin of the involucrum rises on the last anterior centimetre. The sigmoid process, only preserved in IRSNB M. 1902, is transversely oriented, with a weakly angulated posteroventral corner. Mandible The partial skull USNM is associated to a fragment of mandible of 305 mm, including the anterior of the rami (one third of the total length) and the posterior of the symphysis. The width and height of the mandible at the symphysis respectively equal 38 and 24 mm. The alveolar groove contains about 45 alveoli on a length of 265 mm, with a diameter of 3-5 mm. The lateral surface of the mandible is hollowed by a deep longitudinal groove. Post-cranial skeleton (PI. 4) The vertebrae associated to the holotype IRSNB M.361, seven cervicals and seven thoracics, were described and figured by A b e l (1931). The free cervicals bear long centra (Table 2), with proportions close to the extant river dolphins Platanista and Inia, indicating a long and flexible neck. The atlas is long and robust; the anterior articular surfaces are deeply concave; the dorsal transverse process is slightly longer and stronger than the ventral. The transverse process of the axis is robust; the neural spine is dorsally divided in a vertical thinner blade and a stronger dorsoposterior projection. The triangular neural arch of the five last cervicals bears a well developed pre- and postzygapophysis. The cervical c4 of X. cristatus IRSNB 3234-M.361 is close to the cervical (?c5) Table 2 Measurements (mm) on the cervical (cl-c7) and thoracic (tl-tx) vertebrae of Xiphiacetus cristatus IRSNB 3234-M.361 (holotype), Miocene o f Antwerp region. The thoracic tx does not directly follow t6; at least one vertebra o f the series is missing, e indicates estimate, '+ nearly complete and no data. Some of the measurements are illustrated on Plate 4. The centrum length on the atlas and the axis (cl, c2) is the maximum longitudinal length between anterior and posterior articular surfaces. On the six first thoracic vertebrae, the posterior centrum width is considerably larger than the anterior centrum width because of the presence o f the diapophysis (articular surface with the tuberculum o f the rib). cl c2 c3 c 4 c5 c6 c7 tl t2 t3 t4 t5 t6 tx 1. cen tru m length anterior centrum w idth - - e e63 3. anterior centrum height e po sterio r cen trum w idth posterior cen trum height w id th acro ss tran sv erse p ro cesses 112 el 34 e l2 6 e l2 6 e e l e l4 l 141 e l e l total h eight o f vertebra height neural arch e29 9. h eig h t neural spine _
, but with a more dorsoventrally flattened centrum, and a longer transverse process with a lower parapophysis. The robust parapophysis of c6 is much elongated, 1.")

11 Review o f Priscodelphinus cristatus and phylogeny 221 of Xiphiacetus bossi USNM 8842 figured by K e l l o g g (1925, pi. 6, figs 3, 6), but with a more dorsoventrally flattened centrum, and a longer transverse process with a lower parapophysis. The robust parapophysis of c6 is much elongated, 1.5 times longer than the parapophysis of c5, and more than twice longer than the plate-like parapophysis of c7. A similarly longer parapophysis of c6 is observed in the archaeocete Dorudon atrox', this parapophysis is one of the origins for the longus coli muscle, which inserts on the ventral side of the centra of the first cervicals, and which acts mainly as a flexor of the neck ( U h e n, 2003). This condition further supports the high flexibility of the neck of Xiphiacetus cristatus. The thoracics are generally similar to X. bossi USNM 8842 ( K e l l o g g, 1925, pi. 7-9), also with large pre- and postzygapophyses. The neural spine of t5 is slightly posteriorly projecting, contrary to the known anterior thoracics of X. bossi. Concerning the articulation of the ribs, the strong transverse process bears a wide parapophysis, and the centrum of at least the six first thoracics bears a diapophysis considerably increasing the posterior width of the centrum. No forelimb element of Xiphiacetus cristatus is known, contradicting A b e l (1931). R em arks on the stratigraphie range o f Xiphiacetus cristatus The specimens of Xiphiacetus cristatus from the east coast of the USA were all found in the Calvert Formation, more precisely in the beds 12, 13, and 14 as defined by S h a t t u c k ( ) ( M y r ic k, ), dated from the Middle Miocene ( V e r t e u il & N o r r is, , fig. 4 ). All the Belgian skulls of Xiphiacetus cristatus found during the 1860 s were collected without precise stratigraphie and location data. None of them has a preservation similar enough to specimens o f Eurhinodelphis cocheteuxi, from the Antwerp Sands (late Early to Middle Miocene), to imply a similar origin. The partial skulls IRSNB 8243-M.1898 and IRSNB 8243 were discovered in Kessel, 18 km southeast o f Antwerp (Fig. 1 ), during the building of a fort, in February-March In the area of Kessel, the only Neogene unit recorded under the Quaternary layer is the member of the Antwerp Sands (see H u y g e b a e r t & N o l f, 1979, Kessel is close to the drilling GD 11 in figs 1, 3). The specimens IRSNB M.1896 and IRSNB M.1902 were found in the Sables à FIétérocètes, the former more or less one metre above the upper Glycymeris layer o f the Antwerp Sands (G. Paredis, pers. comm. 2002; L. Peeters, pers. comm. 2003). A section close to the place of discovery of the two skulls (Stenen Brug, I S. B.) was described by D e M e u t e r et al. (1976, p. 10; flg. 16); no Sables à Hétérocètes are described in that section, where the Antwerp Sands are overlaid with reworked Deume Sands and Kattendijk Sands, respectively from the Upper Miocene and Lower Pliocene. However, IRSNB M.1896 is a fragile specimen, with slender fragments of the rostrum preserved, and several teeth in situ', it is therefore difficult to imagine this skull having undergone a phase of reworking. Non-reworked sediments might therefore be present above the Antwerp Sands, alternately with reworked Upper Miocene sediments, and under reworked Lower Pliocene deposits. The nonreworked deposits would correspond to the Sables à Hétérocètes. G l i b e r t & d e H e in z e lin (1955) and D e M e u t e r & L a g a (1976) include the Sables à Hétérocètes in the Upper Miocene Diest Formation, a formation represented in Antwerp by the Deume Sands. Even if microfossils from new samples of the non-reworked levels would help to clarify the problem, Xiphiacetus cristatus is probably recorded from levels at least younger than the Antwerp Sands. The chronological range o f the family Eurhinodelphinidae was usually proposed from the Late Oligocene to the Middle Miocene ( F o r d y c e & B a r n e s, 1994; F o r d y c e & M u iz o n, 2001). Those newly identified Belgian skulls of Xiphiacetus cristatus might extend that range into the Late Miocene, pending additional stratigraphie information. E volution ary con sid erations For several characters, Xiphiacetus cristatus seems particularly derived compared to other eurhinodelphinids. The progressive telescoping of the skull, moving the bony nares posterodorsally, is a process that can be traced among the major lineages of odontocetes ( M i l l e r, 1923). In X. cristatus, the supraoccipital shield is nearly vertical against an elevated and acute posterior margin of the vertex, laterally followed by an acute transverse crest. This morphology indicates a pronounced posterodorsal shift of the bony nares, longitudinally pinching the vertex (Fig. 6). The height of the temporal fossa of X. cristatus is reduced compared to other eurhinodelphinids and taxa probably related to the family (e.g., Eoplatanista D a l P ia z, 1916 and Argyrocetus joaquinensis K e l l o g g, 1932) (Fig. 6). When calculating the ratio between the height of the temporal fossa and the bizygomatic width of the skull among eurhinodelphinoid taxa (Eurhinodelphinidae + Eoplatanistidae, M u iz o n, 1988a), the following trend is observed (Fig. 7). The ratio varies between 0.44 and 0.35 among the two species of Eoplatanista, and equals 0.37 for the holotype o f 'Argyrocetus' joaquinensis. It decreases to 0.28 for the holotypes o f Eurhinodelphis longirostris and Mycteriacetus bellunensis, between 0.28 and 0.26 for Ziphiodelphis (Z. abeli and Z. sigmoideus), 0.25 for the holotypes of Eurhinodelphis cocheteuxi and Schizodelphis sulcatus, a mean of 0.22 for the specimens of Schizodelphis from the USNM (specific taxonomy not always resolved and several specimens slightly dorsoventrally compressed) and for Xiphiacetus bossi, and finally no more than 0.20 for X. cristatus. As previously mentioned, few data are available about the total length o f the mandible for the eurhinodelphinids.


, Ziphiodelphis spp. (4), Schizodelphis spp. (11), Xiphiacetus bossi (5), and X. cristatus (5).")




12 222 Olivier LAMBERT Eoplatanista Argyrocetus' M ycteriacetus mandible mandible Eurhinodelphis Ziphiodelphis Xiphiacetus Fig. 6 Schematic left lateral view o f the cranium for several long-snouted dolphins, showing the variation o f the degree of telescoping and o f the height of the temporal fossa. A. Eoplatanista gresalensis MGPD B. Argyrocetus joaquinensis USNM (mod. from K e llo g g, 1932). C. Mycteriacetus bellunensis MGPD D. Eurhinodelphis cocheteuxi IRSNB 3252-M.294. E. Ziphiodelphis sigmoideus MGPD F. Xiphiacetus cristatus IRSNB M Not to scale. o CL. C l. Taxa Fig. 7 Diagram illustrating the trend towards a decrease o f the ratio between height of temporal fossa and bizygomatic width among eurhinodelphinoid taxa. Interval bars indicate more than one specimen: Eoplatanista spp. (5), Ziphiodelphis spp. (4), Schizodelphis spp. (11), Xiphiacetus bossi (5), and X. cristatus (5). For Argyrocetus' joaquinensis, Eurhinodelphis longirostris, E. cocheteuxi, and Mycteriacetus bellunensis, only the holotype was included. Several skulls from the USNM were too compressed dorsoventrally to be adequately used. Q. O. Nevertheless, the ratio between bizygomatic width of the skull and length o f the mandible increases from Eoplatanista spp. (mandible as long as rostrum) to Xiphiacetus spp., with Argyrocetus patagonicus and Mycteriacetus bellunensis as intermediaries. Because the temporal fossa is the main area of origin for the temporalis muscles, elevating the mandible, the reduction of the size of the temporal fossa, probably associated to less powerful muscles, might be partially related to the shortening of the mandible. However, it is likely that the way the animal feeds - size and type of prey, kind of grasping process - is another factor related to the muscular development, and thus to the size of the temporal fossa. For example, suction feeding upon squids asks for a less powerful mandible than predation upon marine mammals. The temporal fossa of the extant ziphiid Ziphius cavirostris, suspected to feed by sucking in the preys ( H e y n in g & M e a d, 1996), is considerably reduced compared to the large fossa of the killer whale Orcinus orca. The shortening of the posterior process of the periotic m Xiphiacetus is discussed above, and this feature, linked to a better isolation of the ear bones relatively to the basicranium, is derived compared to the condition in Eurhinodelphis, Schizodelphis, and Ziphiodelphis. At the level o f the pterygoid sinuses, Xiphiacetus cristatus is the eurhinodelphinid species with the deepest and longest fossa for the postorbital lobe o f the sinus: the


13 Review of Priscodelphinus cristatus and phylogeny 223 sinus does not reach the roof of the orbit in Eurhinodelphis - or does not excavate it, the fossa is laterally shorter in Schizodelphis and Ziphiodelphis, and it is shallower in Xiphiacetus bossi. The expansion of the pterygoid sinus in the orbit and the temporal fossa is a character used by M u iz o n (1991) to group the superfamily Eurhinodelphinoidea with the Delphinida sensu M u iz o n, 1988b. It is suggested here that this feature progressively appears within the family Eurhinodelphinidae, together with the shortening of the posterior process of the periotic; those two characters should therefore be considered as homoplasies, occurring independently in at least two different lineages (Eurhinodelphinidae and Delphinida). Another character particularly specialized in X. cristatus is the shape of the teeth. The condition, probably ontogenetically variable, is close to the extant Pontoporia, and is likely derived, even if only few data are available on the teeth of other eurhinodelphinids. A ll those anatom ical observations, from diverse areas o f the skull, m ost o f them functionally related to the acoustic and feeding system s, indicate that Xiphiacetus cristatus is m ore specialized am ong eurhinodelphinids. T he Stratigraphie data and the palaeontological record are fragm entary and lack precision, but a correlation betw een the m ore derived features o f that species and its possibly longer fossil record m ight be inform ative. Phylogeny Analysis The only previous phylogenetic study illustrating the eurhinodelphinid relationships is the one of M u iz o n (1991), isolating the crownward Schizodelphis + (Ziphiodelphis + Eurhinodelphis) from Argyrocetus by the sub vertical occipital shield, and Ziphiodelphis + Eurhinodelphis by the wider occipital shield and shorter cerebral skull *-12*-17* « * * Eoplatanista (MED) 13-14* Argyrocetus (SA) 7**-8 Mycteriacetus (MED) 16' 13 Squalodon Ziphiodelphis (MED) Xiphiacetus (NS+EU+MED) Schizodelphis (NS+EU+MED) Eurhinodelphis (NS) Fig. 8 Most parsimonious cladogram illustrating the relationships between eurhinodelphinid genera. Tree length 32 steps; C.I. 0.75; R.I Numbers at each node refer to characters discussed in the text and listed in Appendix 2. Change from state 0 or 1 to state 2 ( ); reversal from state 1 to state 0 (*); reversal from state 2 to state 1 (**). Abbreviations between brackets indicate the region of origin: EU, east coast of the USA; MED, Mediterranean; NS, North Sea Basin; SA, east coast of South America. Taking into account the systematic revision inside the family ( L a m b e r t, 2004; this study), a cladistic analysis examining the phylogenetic relationships between the best-known eurhinodelphinid genera Eurhinodelphis (E. cocheteuxi and E. longirostris), Mycteriacetus (M. bellunensis), Schizodelphis (S. morckhoviensis, IS. barnesi, and Schizodelphis sp. from the USNM), Xiphiacetus {X. cristatus and X. bossi), and Ziphiodelphis (Z. abeli and Z. sigmoideus), the more fragmentarily known Argyrocetus patagonicus, and the eoplatanistid Eoplatanista was carried out with the parsimony program PAUP, version 4.0 beta 10 ( S w o f f o r d, 1998). A matrix of 21 characters (excluding uninformative characters and all treated as unweighted and unordered, see Appendix 2 for the description of the characters) for the seven taxa (Table 3) was built on the basis of direct observations of most o f the specimens, except the holotype of Argyroce- Table 3 Data matrix o f 21 characters for one outgroup and seven analysed taxa. All characters with multiple states are treated as unordered. Primitive state, 0 ; derived states, 1, 2 ; variable between 0 and 1, a ; missing character, 7. Description o f the characters in Appendix 2. Characters Taxa Squalodon Eoplatanista 0 a a ? Argyrocetus 1? 1 0?? 2 0 1?? 0 1 0? 7 7???? Mycteriacetus ? 0? Ziphiodelphis Xiphiacetus n. gen a a 1 Eurhinodelphis a Schizodelphis a 1 1 a a 1 a

, with the main eurhinodelphinid localities.")
.")
14 224 Olivier LAMBERT North Sea North Pacific Macrodelphinus kelloggi O Eurhinodelphis cocheteuxi E. longirostris 0 Xiphiacetus cristatus X. hossi Schizodelphis morckhoviensis North Atlantic Xiphiacetus cristatus X. bossi Schizodelphis morckhoviensis Schizodelphis sp. Argyrocetus patagonicus Schizodelphis sulcatus S j j Medite rranean Xiphiacetus aff. bossi I Ziphiodelphis abeli Z sigmoideus M ycteriacetus bellunensis Fig. 9 Palaeogeographic map o f the North Atlantic and Mediterranean realms during Middle Miocene (mod. from S m ith et al., 1994), with the main eurhinodelphinid localities. Several o f the species placed on the map are dated from Early or Late Miocene; at those times, the outline o f the coasts differed a while from the reconstruction presented here. The locality of Argyrocetus patagonicus is outside the map, in the Early Miocene o f Patagonia, Argentina. tus patagonicus (figured in L y d e k k e r, 1893). One outgroup was chosen a priori: the squalodontid Squalodon. The poorly known large eurhinodelphinid Macrodelphinus W ils o n, 1935 and the two species usually thought to belong to the same genus than Argyrocetus patagonicus, A.' joaquinensis K e l l o g g, 1932 and A. bakersfieldensis ( W ils o n, 1935), were not included in the analysis and their status is briefly discussed in Appendix 1. The result of the performed heuristic search is a unique minimal cladogram of 32 steps (C.I. 0.75; R.I. 0.73), presented in Figure 8. The main features of that cladogram are a sister-group relationship between Schizodelphis + Xiphiacetus and Ziphiodelphis + (Mycteriacetus + Argyrocetus), and a more sternward Eurhinodelphis. The close relationship between Mycteriacetus bellunensis and Argyrocetus patagonicus was already suggested by M u iz o n (1988a), C o z z u o l (1996), and B ia n u c c i & L a n d in i (2002), while the general topology of the cladogram differs from the tree of M u iz o n (1991) because of a larger number of characters and modifications concerning the generic allocation o f the different species. This cladogram also differs from the consensus tree of L a m b e r t (in press), giving hypothetical relationships between most of the Neogene odontocete families; in that study, using a larger number of characters, and only including three eurhinodelphinid taxa and no eoplatanistid, Eurhinodelphinidae is not monophyletic: Eurhinodelphis cocheteuxi is a more crownward, sister-group o f the ziphiids. Discussion The relatively isolated basal position of Eurhinodelphis obtained here allows establishing an interesting parallel with the palaeogeography of the family. The two species of Eurhinodelphis are currently only known from the North Sea Basin. On the other hand, several species of Schizodelphis and Xiphiacetus are described from both sides of the North Atlantic: in the North Sea Basin and along the eastern coast of the USA; Schizodelphis and Xiphiacetus are also known from the Mediterranean (Fig. 9). Species of the phylogenetically closely related Xiphiacetus and Schizodelphis might share a more pelagic habitat compared to Eurhinodelphis. The ancestral habitat could therefore have been more coastal, or even estuarine [one of the oldest records of eurhinodelphinids is from fluvial-lacustrine deposits of the Late Oligocene of Australia ( F o r d y c e, 1983)], with a progressive invasion of wider areas of the continental shelf until the mid- Miocene climatic optimum. From a morphological point


15 Review o f Priscodelphinus cristatus and phylogeny 225 of view, the similarities of the eurhinodelphinids with the river dolphins at the level of the length of the rostrum, bearing a deep lateral groove, and of the length of the neck, support the hypothesis of relatively slow-swimming shallow-water dwellers. The sensitive edentulous anterior part of the rostrum, longer than the mandible, could correspond to an efficient tool used to forage by burrowing on the sediments of the bottom to detect and disturb preys. Similar arguments were produced to propose a burrowing behaviour for Eurhinosaurus longirostris, a Jurassic ichthyosaur with a rostrum twice as long as the mandible (R ie s s, 1986; G o d e f r o i t, 1996). Ziphiodelphis and Mycteriacetus are for now only known from Mediterranean. A more southern origin (Mediterranean) for the clade Ziphiodelphis + (Mycteriacetus + Argyrocetus), in warmer climate, could have made possible a later dispersion in the southern hemisphere (South Atlantic for Argyrocetus patagonicus). A ck now ledgm ents I would like to thank the director o f the IRSNB, D. Cahen, and the Department o f Palaeontology for allowing me to study the collection of fossil odontocetes. G. Bianucci (Pisa), D.J. Bohaska (Washington D.C.), Mariagabriella Fornasiero and Letizia Del Favero (Padua), G. Paredis and L. Peeters (Deume, Belgium), and P.J.H. Van Bree (Amsterdam), kindly provided access to fossils in their care. We gratefully acknowledge L. Peeters for having presented the tympanic bulla (IRSNB M. 1902) to the IRSNB. Discussions with C. de Muizon (Paris) were very helpful. Critical comments o f G. Bianucci and C. de Muizon improved previous versions o f the manuscript. W. Miseur is thanked for photographic work. This paper is a part of a PhD thesis at the Free University o f Bmssels (ULB), granted by the Fonds pour la Formation à la Recherche dans Industrie et dans l Agriculture (FRIA). The visits to the MGPD, MP, and USNM collections were financed by a grant o f the Fonds National de la Recherche Scientifique (FNRS). References A b e l, O., Les dauphins longirostres du Boldérien (Miocène supérieur) des environs d Anvers. I. Mémoires du Musée Royal d Histoire Naturelle de Belgique, 1: A b el, O., Les dauphins longirostres du Boldérien (Miocène supérieur) des environs d Anvers. II. Mémoires du Musée Royal d 'Histoire Naturelle de Belgique, 2: A b e l, O., Les Odontocètes du Boldérien (Miocène supérieur) des environs d Anvers. Mémoires du Musée Royal d Histoire Naturelle de Belgique, 3: A b el, O., Das Skelett der Eurhinodelphiden aus dem Obermiozän von Antwerpen. Mémoires du Musée Royal d'histoire Naturelle de Belgique, 48: B a rnes, L. G., Outline o f Eastern North Pacific fossil cetacean assemblage. Systematic Zoology, 25 (4): B ianucci, G. & L a n d in i, W., Change in diversity, ecological significance and biogeographical relationships of the Mediterranean Miocene toothed whale fauna. Geobios, 35, mémoire spécial 24: B ia nucci G., L a n d in i, W. & V a róla, A., New remains o f Cetacea Odontoceti from the «Pietra leccese» (Apulia, Italy). Bollettino della Societa Paleontológica Italiana, 33 (2): B risso n, M.-J., Regnum Animale in classes IX distributum, sine synopsis methodica. Theodorum Haak, Paris, 296 pp. C a brera, A., Cetáceos fósiles del Museo de La Plata. Revista del Museo de La Plata, 29: C o zzu o l, M. A., The record o f aquatic mammals in southern South America. Münchner Geowissenschaftliche Abhandlungen, 30 (A): D al P ia z, G., Sui Vertebrati delle arenarie mioceniche di Belluno. Atti della Accademia Scientifica veneto-trentino-istriana, classe 1, 5: D al P ia z, G., Gli Odontoceti del Miocene Bellunese. Introduzione generale - Parte Prima-Seconda. Memorie d ell Istituto Geologico della Reale Università di Padova, 4: D al P ía z, G., Gli Odontoceti del Miocene Bellunese. Parte Quinta-Décima Cyrtodelphis, Acrodelphis, P r o d e lp h i nus, Ziphiodelphis, Scaldicetus. Memorie dell 'Istituto Geologico delia Reale Università di Padova, Allegato al vol. 4(1916): D e M eu ter, F. J. & L a g a, P., Lithostratigraphy and biostratigraphy based on benthonic foraminifera o f the Neogene deposits of northern Belgium. Bulletin de la Société belge de Géologie, 85: D e M eu ter, F. J., W o u ters, K. & R in g ele, A., Lithostratigraphy o f Miocene sediments from temporary outcrops in the Antwerp city area. Service Géologique de Belgique, Professional Paper, 3: du Bus, B.A.L., Sur quelques Mammifères du Crag d Anvers. Bulletin de l Académie Royale des Sciences de Belgique, 24: d u Bus, B.A.L., Mammifères nouveaux du Crag d Anvers. Bulletin de l Académie Royale des Sciences de Belgique, 34: E stev en s, M., Mamíferos marinhos do Miocénico da Península de Setúbal. Ciências da Terra (UNL), N úm ero Especial V (CD-ROM): A60-A63. F lo w er, W. H., Description o f the skeleton o f Inia geoffrensis and the skull o f Pontoporia blainvillei, with remarks on the systematic position of these animals in the Order Cetacea. Transactions o f the Zoological Society o f London, 6: F o r d y c e, R. E., Redescription o f Early Miocene dolphin Phocaenopsis mantelli Huxley, 1859 (Odontoceti incertae sedis). New Zealand Journal o f Geology and Geophysics, 24 (4): F o rdyce, R. E., Rhabdosteid dolphins (Mammalia: Cetacea) from the Middle Miocene, Lake Frome area, South Australia. Alcheringa, 7: F o rdyce, R. E., Waipatia maerewhenua, new genus and new species (Waipatiidae, new family), an archaic late Oligocène dolphin from New Zealand. In: B er ta, A. &. D em ér é, T.A. (Editors), Contributions in Marine Mammal Paleontology Honoring Frank C. Whitmore, Jr., Proceedings o f the San Diego Society o f Natural History, 29:
, Secondary Adaptation o f Tetrapods to Life in Water. Verlag Dr.")

16 226 Olivier LAMBERT F o rdyce, R. E. & B a r n es, L. G., The evolutionary history o f whales and dolphins. Annual Review o f Earth and Planetary Science, 22: F o rdyce, R. E. & M u izo n, C. d e, Evolutionary history of cetaceans: a review. In: M a zin, J.-M. & d e B u ffr én il, V. (Editors), Secondary Adaptation o f Tetrapods to Life in Water. Verlag Dr. Friedrich Pfeil, München, Germany, pp G ervais, P., Sur différentes espèces de vertébrés fossiles observés pour la plupart dans le Midi de la France. Mémoires de l 'Académie des Sciences et des Lettres de Montpellier, 16: G libert, M. & de H ein zelin, J., La faune et l âge Miocène supérieur des Sables de Deume. Bulletin de l 'Institut Royal des Sciences Naturelles de Belgique, 31 (71): G o d efroit, P., Biodiversité des reptiles marins du Jurassique inférieur belgo-luxembourgeois. Bulletin de la Société belge de Géologie, 104 (1-2): H ey n in g, J. E., Comparative facial anatomy of beaked whales (Ziphiidae) and a systematic revision among the families o f extant Odontoceti. Contributions in Science, Natural History Museum o f Los Angeles County, 405: H ey n in g, J. E. & M ea d, J. G., Suction feed in g in b eak ed w hales: m orphological and o bservational evidence. Contributions in Science, Natural History Museum o f Los Angeles County, 464: H o w ell, A. B., Asymmetry in the skulls o f mammals. Proceedings o f the United States National Museum, 67: H u y g eba ert, B. & N o lf, D., Otolithes de téléostéens et biostratigraphie des Sables de Zonderschot (Miocène moyen de la Belgique). Mededelingen Werkgroep Tertiair Kwartair Geologie, 16 (2): Ka su y a, T., S y stem atic consid eratio n o f recent to o th ed w h ales based on the m o rp h o lo g y o f ty m p ano-p erio tic bone. Scientific Reports o f the Whales Research Institute, Tokyo, 25: K ello g g, R., A fossil porpoise from the C alv ert F orm a tio n o f M aryland. Proceedings o f the United States National Museum, 63 (14): K ello g g, R., On the occurrence of remains of fossil porpoises of the genus Eurhinodelphis in North America. Proceedings o f the United States National Museum, 66 (26): K e llo g g, R., A Miocene long-beaked porpoise from California. Smithsonian Miscellaneous Collections, 87 (2): K ello gg, R., Description of the skull of Pomatodelphis inaequalis Allen. Bulletin o f the Museum o f Comparative Zoology, Cambridge, 121: L a m bert, O., Systematic revision o f the Miocene longsnouted dolphin Eurhinodelphis longirostris DU Bus, 1872 (Cetacea, Odontoceti, Eurhinodelphinidae). Bulletin de l Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre ', 74: L a m bert, O., in press. Phylogenetic affinities o f the longsnouted dolphin Eurhinodelphis (Cetacea, O dontoceti) from the M iocene o f A ntwerp. Palaeontology. L eid y, J., Proceedings o f the Academy o f Natural Sciences o f Philadelphia, 5: L y d e k k er, R., On Zeuglodont and other Cetacean remains from the Tertiary o f the Caucasus. Proceedings o f the Geological Society o f London, 1892: L y d e k k er, R., Contribution to the knowledge o f the fossil vertebrates of Argentina. Part II. Cetacean skulls from Patagonia. Anales del Museo de La Plata, 1893: M a ta, C.R. d a, Nota preliminar sobre um delfinideo (Eurhinodelphis cf. cristatus) do Miocénico do Penedo, a Norte do Cabo Espichel. Boletim do Museu e Laboratorio Mineralógico e Geológico da Faculdade de Ciëncias, Universidade de Lisboa, 9: M c h ed lid ze, G. A., General features o f the palaeobiological evolution o f Cetacea. {Osnovnye Cherty Paleobiologicheskoi Istorii Kitoobraznykh}. Metsniereba Publishers, Tbilisi. Translated from the Russian in 1984 by Amerind Publishing Co. Pvt. Ltd., New Delhi, 139 pp. M ea d, J. G., Anatomy of the external nasal passages and facial complex in the Delphinidae (Mammalia: Cetacea). Smithsonian Contribution to Zoology, 207: M iller, G.S., J r, The telescoping of the cetacean skull. Smithsonian Miscellaneous Collections, 76 (5): M u izo n, C. d e, Les vertébrés de la Formation Pisco (Pérou). Deuxième partie: Les odontocètes (Cetacea, Mammalia) du Pliocène inférieur du Sud-Sacaco. Travaux de l Institut français d Etudes Andines, 27: M u izo n, C. d e, 1988a. Le polyphylétisme des Acrodelphidae, odontocètes longirostres du Miocène européen. Bulletin du Muséum National d'histoire Naturelle, Paris, section C, (4), 10, (1): M u izo n, C. d e, 1988b. Les relations phylogénétiques des Delphinida. Annales de Paléontologie, 74 (4): M u izo n, C. d e, A new Ziphiidae (Cetacea) from the Early Miocene o f Washington State (USA) and phylogenetic analysis of the major groups o f odontocetes. Bulletin du Muséum National d Histoire Naturelle, Paris, section C, (4), 12, (3-4): M y rick, A.C., Variation, taphonomy and adaptation o f the Rhabdosteidae (= Eurhinodelphidae) (Odontoceti, Mammalia) from the Calvert Formation o f Maryland and Virginia. PhD Dissertation, University of California, Los Angeles, (University Microfilms International, Ann Arbor, Michigan), 347 pp. (unpublished). P illeri, G., The Miocene Cetacea o f the Belluno sandstones (Eastern Southern Alps). Memorie di Scienze Geologiche, 36: R ie s s, J., Fortbewegungsweise, Schwimmbiophysik und Phylogenie der Ichthyosaurier. Palaeontographica, A, 192: Rom m el, S., Osteology o f the Bottlenose dolphin. In: S. L ea th erw oo d & R. R. R eeves (Editors), The Bottlenose dolphin. Academic Press, San Diego, pp S ch en k k a n, E. J., On the comparative anatomy and function o f the nasal tract in odontocetes (Mammalia, Cetacea). Bijdragen tot de Dierkunde, 43: S h a ttu ck, G. B., Geological and paleontological relations, with a review of earlier investigations. In: C la rk, W. B. (Editor), Maryland Geological Survey, Miocene. Johns Hopkins Press, Baltimore, 543 pp. S m ith, A. G., S m ith, D. G. & F u n n ell, B. M., Atlas of Mesozoic and Cenozoic Coastlines. Cambridge University Press, 109 pp. S w o ffo rd, D. L PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4. Sinauer Associates, Sunderland, Massachusetts.

17 Review o f Priscodelphinus cristatus and phylogeny 227 T a v ernier, R. & de H ein zelin, J., Introduction au Néogène de la Belgique. Memoire de la Société belge de Géologie, de Paléontologie et d 'Hydrologie, 8: U h en, M. D., Form, function, and anatomy of Dorudon atrox (Mammalia, Cetacea): an archaeocete from the Middle to Late Eocene o f Egypt. Papers on Paleontology, 34: V an B en ed en, P.- J. & G ervais, P., Ostéographie des cétacés vivants et fossiles. Arthus Bertrand, Paris, 634 pp. V anden B ro eck E., Esquisse géologique et paléontologique des dépôts pliocènes des environs d Anvers, première partie. Annales de la Société Malacologique de Belgique, 9 (2): V erteu il, L. de & N o r r is, G., Miocene dinoflagellate stratigraphy and systematics o f Maryland and Virginia. Micropaleontology, suppl. 42: W ilso n, L. E., Miocene marine mammals from the Bakersfield region, California. Bulletin o f the Peabody Museum o f Natural History, 4: Olivier L a m bert (boursier F.R.I.A. jusqu octobre 2004) Département de Paléontologie Institut royal des Sciences naturelles de Belgique Rue Vautier 29, B-1000 Brussels Belgium 01ivier.Lambert@ naturalsciences.be Typescript submitted: July 10, 2004 Revised typescript received: October 15, 2004
.")
.")
18 228 Olivier LAMBERT Appendix 1 Comments on several taxa previously included in, or related to, the family Eurhinodelphinidae (reviews by F o rdyce, 1981; M u izo n, 1988a, 1991; Fo rdy ce & M u izo n, 2001). Iniopsis caucasica L y d ekk er, 1892 The study o f the systematic affinities o f that probable primitive eurhinodelphinid, from the Late Oligocene o f Georgia and Azerbaidzhán, would necessitate direct observation o f the specimens (especially the ear bones and the basicranium, see M c h ed lid ze, 1976, pi. 9, 10). Protodelphinus capellinii D al P ia z, 1977 The only species o f the genus Protodelphinus was described by D al P iaz (1977) on the basis of a partial mandible MGPD 26182, small fragments of rostrum, a right periotic-tympanic bulla set MGPD and several teeth from the Early Miocene of north-eastern Italy (Libano Sandstone, Bolzano quarry). In this paper, D al P iaz considered the species as a primitive member o f the family Delphinidae, an attribution also supported by P illeri (1985) who described a new mandibular fragment, possibly belonging to a new species from the same genus. The ear bones lead M u izo n (1991) to suggest that Protodelphinus might be a eurhinodelphinid (at least a eurhinodelphinoid, Fo rdy ce & M u izo n, 2001). Those ear bones show clear eurhinodelphinid affinities (e.g., the long and pointed anterior process of the periotic bearing an anterior articular facet and the indentation on the high dorsal margin o f the involucrum o f the tympanic), even if several differences (size, shape of the pars cochlearis o f the periotic, proportions o f the tympanic) with the known eurhinodelphinids from the Belluno Sandstone, Ziphiodelphis and Mycteriacetus, preclude a specific attribution. However, some characters of the mandible provide differences with the eurhinodelphinids: the lateral surface lacks a longitudinal groove, even at the beginning of the symphysis; the ventral margin is distinctly angulated at mm anteriorly to the beginning o f the symphysis, indicating a dorsal elevation o f that margin; the forwards decrease of the height o f the symphyseal portion is stronger than in eurhinodelphinids. These three characters might indicate a shorter symphyseal portion of the mandible compared to the known eurhinodelphinids. Mistakes could therefore be suspected in the associations o f isolated bones suggested by D a l P iaz (1916, 1977), as already discussed by M u izo n (1988a) concerning Dalpiazina ombonii. In a deposit relatively so concentrated in fossil odontocete fragments as the Bolzano quarry, an association of ear bones and a mandible, while the basicranium is lacking, seems rather doubtful. In the present case, as the mandible does not fit the diagnostic characters seen on the ear bones, the association should be rejected. It is suggested here to remove the ear bones MGPD from the holotype, and to refer them to Eurhinodelphinidae incertae sedis. The mandible and the associated teeth show similarities with kentriodontid delphinoids. Therefore, the holotype o f Protodelphinus capellinii is restricted to the mandible MGPD and associated teeth MGPD 26186, and that species is provisionally referred to Odontoceti aff. Delphinoidea. Argyrocetus L y d ek k er, 1893 The poorly diagnosed genus Argyrocetus is currently including three species: the type-species A. patagonicus L ydek k er, 1893 from the Late Oligocene-Early Miocene o f Patagonia, Argentina; A. joaquinensis K el lo g g, 1932 and A. bakersfieldensis (W ilso n, 1935), both from the Early Miocene of California (see B a rn es, 1976). Those species are less extensively known than the eurhinodelphinids from Europe and the east coast o f the USA. Only one specimen is described for the species A. patagonicus; the anterior part o f the rostrum is poorly preserved in A. patagonicus, and absent in both Californian species; no ear bones and no teeth are described for the genus; and the mandible is known for A. patagonicus only. Argyrocetus patagonicus L y d ekk er, 1893 The species was first briefly described by L y d ek k er (1893), particularly stressing the shape o f the nasals, projecting over the bony nares where they end with a nearly straight transverse edge. Additionally to a scale mistake in the pictures o f L y d ek k er (1893, pi. 5) - the reduction is not o f 2/3 but 1/2, C a brera (1926) provided further information: while the rostrum is preserved anteriorly on a shorter distance than the mandible, he maintained that the premaxillae are anteriorly longer than the maxillae, estimating the premaxillary part of the rostrum at 200 mm for a total length o f the skull o f at least 854 mm. This premaxillary length corresponds to the distance between the anterior preserved extremity of the maxilla and the apex of the articulated mandible on the left side picture of L ydekker (1893, pi. 5). Thus the opinion o f C a brera (1926) is that the anterior preserved extremity o f the maxilla corresponds to the real apex of that bone, and that the mandible was 200 mm longer than the maxillary part o f the rostrum. C a brera (1926) also proposed that this missing anterior premaxillary part o f the rostrum did not bear teeth, because the corresponding portion of the mandible seems to lack individualized alveoli. With this incomplete and dorsoventrally crushed rostrum (exaggerating the dorsal opening o f the vomerian gutter, following C a b r era, 1926), it is not possible to guess if the premaxillae were longer than the mandible, the condition observed in at least Schizodelphis, Xiphiacetus, and Ziphiodelphis. However, the probably edentulous premaxillae extending further anteriorly than the maxillae for a substantial distance maintain Argyrocetus patagonicus in the family Eurhinodelphinidae. The close relationship between A. patagonicus and Mycteriacetus bellunensis obtained in the above phylogenetic analysis further supports that hypothesis. 'Argyrocetus'joaquinensis K ello gg, 1932 In his systematic discussion about this species, K ello gg (1932) evaluated the similarities o f the holotype of 'A. joaquinensis sensu K ello g g, 1932 with the eurhinodelphinids known at that time; because o f resemblances with A. patagonicus at the level of the supraoccipital shield, the elevation of the vertex, and the proportions o f the skull, he tentatively referred with considerable hesitation the Californian skull to the same genus. The only areas that are well preserved in both A. patagonicus and A. joaquinensis are the supraoccipital shield and the nasals and frontals on the vertex. The supraoccipital seems roughly similar in both species, notwithstanding the more acute aspect of the lambdoid crest in A. joaquinensis. The strange vertex o f the latter, however, strongly differs from that of A. patagonicus; following K ello g g (1932), the nasals are much longer than wide, they are posteriorly thicker, and they do not overhang the bony nares - a feature considered by L y dekk er (1893) as the main characteristic of A. patagonicus. The attribution to the genus Argyrocetus is therefore doubtful. Furthermore, when



19 Review of Priscodelphinus cristatus and phylogeny 229 comparing the holotype o f A. ' joaquinensis to other eurhinodelphinid taxa, clear differences appear in lateral view: the large temporal fossa (see Fig. 6) is much more developed dorsoposteriorly, higher relatively to the vertex than in Eurhinodelphis, Mycteriacetus, Schizodelphis, Xiphiacetus, and Ziphiodelphis, and the zygomatic process is longer anterodorsally. This condition is interestingly similar to the Italian eoplatanistid Eoplatanista (sensu M u izo n, 1988a; e.g., Eoplatanista gresalensis M G PD in P illeri, 1985, pi. 57, fig. a). The latter differs however in the wider and flatter vertex, posteriorly followed by the nearly horizontal median portion o f the supraoccipital. Because of the lack o f information about the rostrum and the mandible, and the absence o f obvious similarities with the known eurhinodelphinids, the holotype o f 'A.' joaquinensis should not be definitely included in that family. Other specimens referred to A. joaquinensis by B arnes (1976), i.e. the holotypes o f Doliodelphis littlei W ilson, 1935 and Eurhinodelphis extensus W ilso n, 1935, do not provide more information about the systematic status o f the species; they could even belong to another taxon [e.g., apart from its size, the holotype o f Doliodelphis littlei seems more similar to Argyrocetus' bakersfieldensis (W ilso n, 1935)]. 'Argyrocetus bakersfieldensis (W ilso n, 1935). This species, first assigned to the genus Acrodelphis by W ilson (1935), was revised by B arnes (1976) in the genus Argyrocetus. A. bakersfieldensis shares with the type species A. patagonicus the anterodorsal projection o f the nasals above the bony nares, with an acute anterior edge. The frontals are relatively longer on the more transversely compressed vertex (Fig. 10), but those differences might be easily explained by intrageneric variation. However, the shape of the supraoccipital shield differs significantly: in A. bakersfieldensis, after a small step at the contact with the frontal, the concave mediodorsal portion of the occipital becomes nearly horizontal. The shield is too incomplete to follow the slope more posteriorly, but that morphology, contradicting the schematic reconstruction o f the supraoccipital proposed by W ilson (1935, fig. 11), differs from the roughly regularly sloping supraoccipital shield of A. patagonicus. The condition o f A.' bakersfieldensis reminds Eoplatanista, for which the anteromedian portion o f the supraoccipital roughly follows the horizontal plane o f the frontal on the Argyrocetus bakersfieldensis supraoccipital frontal maxilla----- nasal, bony nares premaxilla. Macrodelphinus Fig. 10 Schematic drawings o f the vertex of Argyrocetus' bakersfieldensis YPM and Macrodelphinus kelloggi YPM Not to scale. vertex, often with a small step between the frontal and the supraoccipital (e.g., E. gresalensis MGPD 26409, P illeri, 1985, pi. 55). This feature gives Eoplatanista a more convex posterodorsal area o f the cranial skull, compared to the eurhinodelphinids (Fig. 6). The preserved fragments o f the roof of the temporal fossa o f the holotype of Argyrocetus' bakersfieldensis indicate a high fossa, similar to Eoplatanista and higher than in known eurhinodelphinids. Other characters differentiating Eoplatanista from the eurhinodelphinids, as the narrower and thicker preorbital process and the high and acute palate keel, are however absent in Argyrocetus' bakersfieldensis. To summarize, two of the three species usually included in Argyrocetus, A. 'joaquinensis and 'A. ' bakersfieldensis, are not enough similar to the type species to be referred to that eurhinodelphinid genus. Furthermore, because the specimens lack the areas of the skull where diagnostic characters o f the family Eurhinodelphinidae are found, and because o f similarities with the monogeneric family Eoplatanistidae, their familial attribution could not be resolved. Both of them are referred to Odontoceti aff. Eurhinodelphinoidea. Macrodelphinus kelloggi W ilso n, 1935 The holotype o f this large Early Miocene Californian species is composed of the cranium, including the left preorbital process, the base o f the rostrum, and a more anterior rostral portion bearing teeth. A right occipital condyle, an incomplete left tympanic bulla, a scapula, a fragment o f atlas, and several additional vertebrae were tentatively referred to that specimen by W ilson (1935), who described a second specimen: the basal part o f a rostrum associated with the left body o f a mandible and several post-cranial bones. A left humerus, first identified by W ilso n (1935) as belonging to a cetothere, was also referred to the species Macrodelphinus kelloggi by B a rnes (1976). In the description o f the holotype, W ilson (1935, p. 29) gave the following indication: The maxilla is broad at the base of the rostrum; and at the anterior extremity, as preserved, it descends abruptly laterally from the premaxilla. Taking account o f the reconstruction o f W ilso n (1935, fig. 4) and o f the height o f the rostrum at the preserved extremity, the missing apical portion might be relatively long, and the abrupt descent might indicate that the maxilla was significantly shorter than the premaxilla. In addition, M. kelloggi exhibits premaxillae thick and wide at the level of the posterior margin of the bony nares. The surface occupied by the frontals on the vertex is larger, relatively to the short and wide nasals, than in any other eurhinodelphinid (Fig. 10). This condition reminds the vertex morphology o f the more basal odontocetes Waipatia and Squalodon, and likely indicates a primitive state among eurhinodelphinids. The slope and concavity of the supraoccipital shield show rather good similarities with Argyrocetus patagonicus, and the nasals are slightly elevated anterodorsally. Apart from the absence of a clear indentation at mid-length o f the dorsal margin of the involucrum, the description o f the tympanic bulla o f Macrodelphinus kelloggi follows the morphology o f the eurhinodelphinid tympanies, with a broad, shallow and wide median groove, and the dorsal margin o f the thick involucrum strongly anteriorly lowering. All those characteristics o f the skull and the tympanic support the inclusion, first proposed by B arnes (1976), o f M. kelloggi in the family Eurhinodelphinidae, probably in a basal position.
, present (1). 2.")
 - < 1 (1). 4.")
20 230 Olivier LAMBERT Appendix 2 List of the characters used in the cladistic analysis. 1. Anterior portion o f the rostrum constituted by the only premaxillae, forming more than 10 per cent o f the total length o f the rostrum, and lacking alveoli: absent (0), present (1). 2. Mesorostral groove: widely open at the level o f the antorbital notches (0) - nearly closed (1). 3. Proportions o f the cranium: ratio between bizygomatic width and longitudinal distance from antorbital notch to occipital condyles > 1 (0) - < 1 (1). 4. Telescoping o f the skull: ratio between longitudinal distance from anterior margin o f the supraoccipital to the occipital condyles and bizygomatic width o f the skull > 0.30 (0)-< 0.30(1). 5. Preorbital process o f the frontal in lateral view: abruptly dorsoventrally thickened (0) - weakly thickened, almost as thin as the supraorbital area (1). 6. Concavity o f the premaxillary sac fossa: fossa flat or weakly concave (0) - strongly concave (1). 7. Dorsal surface of the nasal: lower or at the same level than the frontal on the vertex (0) - higher than the frontal without anterodorsal projection (1) - projecting anterodorsally (2). 8. Proportions of the nasal: wider than long (0) - as long as wide or longer than wide ( 1 ). 9. Relative length o f the frontal on the vertex: distinctly longer than the nasal (0) - same length or shorter than the nasal (1). 10. Width of the vertex: ratio between minimum distance separating the maxillae across the vertex and postorbital width of the skull < 0.20 (0) - between 0.20 and 0.30 (1) - > 0.30 (2). 11. Dorsomedian portion o f the supraoccipital shield: roughly flat or convex (0) - longitudinally concave (1). This character does not involve the degree of development o f the more lateral lambdoidal crests. 12. Supraoccipital shield sub vertical on its portion above the occipital condyles: no (0) - yes (1). 13. Height of the occipital condyles: ventral margin o f the condyles much lower than the floor o f the temporal fossa (0) - approximately at the level of the floor of the temporal fossa (1). Additionally to Eurhinodelphis sp., Argyrocetus patagonicus seems to have the derived elevated condyles. 14. Roughly complete covering o f the temporal fossa by the frontal-maxilla plate: no, squamosal widely visible in dorsal view (0) - yes (1). 15. Height of the temporal fossa: ratio between height o f the fossa and bizygomatic width > 0.30 (0) - < 0.30 (1). 16. Fossa for the postorbital lobe of the pterygoid sinus on the orbit roof: no fossa reaching the orbit roof (0) - small fossa laterally limited (1) - longer fossa usually excavating at least half the width o f the orbit roof (2). 17. Pterygoid sinus fossa in the alisphenoid dorsoposteriorly excavated above the base of the falciform process: no (0) - yes (1). This condition is often difficult to observe; it is at least present on some specimens o f Schizodelphis, Xiphiacetus, and Ziphiodelphis. 18. Anterior articular facet o f the periotic: nearly flat or weakly excavated (0) - wide and deep (1). 19. Shape o f the pars cochlearis of the periotic: angulated outline, trapezoidal to rectangular in ventral view (0) - rounded, especially anteromedially (1). 20. Tymanic bulla anteriorly pointed in ventral view, with a progressive narrowing: no, abrupt narrowing (0) - yes (1). 21. Inner posterior prominence of the tympanic bulla: roughly as wide as the outer prominence in ventral view (0) - distinctly narrower (1). Even if the prominences are less distinct because of the lack o f a deep median furrow in Eoplatanista, the widths are similar.
. f: frontal; me: mesethmoid; n: nasal; pmx: premaxilla; smx: maxilla. Fig. 2 Xiphiacetus cristatus IRSNB M.1896,?Late Miocene of Antwerp region.")

21 Review o f Priscodelphinus cristatus and phylogeny 231 E xplanation of plates P l a t e Fig. 1 Skull o f Xiphiacetus cristatus IRSNB 3234-M.361 (holotype), Miocene o f Antwerp region. Dorsal view. The labels on the skull are from A bel (1902). f: frontal; me: mesethmoid; n: nasal; pmx: premaxilla; smx: maxilla. Fig. 2 Xiphiacetus cristatus IRSNB M.1896,?Late Miocene of Antwerp region. Detail of the rostrum in left lateral view, showing the descent of the maxilla-premaxilla suture. Fig. 3 Xiphiacetus cristatus IRSNB 3240-M.1895, Miocene of Antwerp region. Posterior part o f the cranium in dorsal view. Fig. 4 Xiphiacetus cristatus IRSNB 3237-M.1894, Miocene of Antwerp region. Cranium in dorsal view. Fig. 5 Xiphiacetus cristatus IRSNB 8243-M.1898, Miocene of Kessel. Cranium in dorsal view. Fig. 6 Skull o f Xiphiacetus cristatus IRSNB 3241-M.1893, Miocene o f Antwerp region. Right lateral view. The labels on the skull are from A bel (1902). f: frontal; pi: palatine; pmx: premaxilla; smx: maxilla; sq: squamosal; v: vomer. Fig. 7 Isolated supraorbital process o f the right maxilla o f Xiphiacetus cristatus IRSNB M. 1899, Miocene o f Antwerp region. Lateral view. Fig. 8 Isolated supraorbital process of the right frontal-maxilla o f Xiphiacetus cristatus IRSNB M.1900, Miocene of Antwerp region. Lateral view. Scale bars for Figs 1-8 = 50 mm. P la te 2 Fig. 1 Skull o f Xiphiacetus cristatus IRSNB 3241-M.1893, Miocene of Antwerp region. A. ventral view. B. posterior view. C. detail o f the right side o f the cranium in ventral view. Scale bar for A-C = 50 mm. The labels on the skull are from A bel (1902). exo: exoccipital; pi: palatine; smx: maxilla; v: vomer. Fig. 2 Right supraorbital process of Xiphiacetus cristatus IRSNB M.1901, Miocene of Antwerp region. Ventral view showing the fossa for the postorbital lobe of the pterygoid sinus. Scale bar = 50 mm. Fig. 3 Maxillary teeth o f Xiphiacetus cristatus IRSNB M.1896,?Late Miocene of Antwerp region. A. anterolateral view of a series of seven teeth. B. lingual view o f the same series. Scale bar for A-B = 20 mm. C. posterior view of another tooth. Scale bar = 10 mm. Fig. 4 Maxillary teeth o f Pontoporia blainvillei ZMA The first on the left is in anterior or posterior view, the four others in lingual or labial view, all o f them with the crown towards the top of the plate. Scale bar = 10 mm. P late 3 Fig. 1 Skull of Xiphiacetus cristatus IRSNB 3227-M. 1897, Miocene o f Antwerp region. A. posterior part of the cranium in anterodorsal view. B. partial basicranium in anteroventral view, showing the asymmetry o f the squamosals (right larger than left). Scale bars = 50 mm. Fig. 2 Right periotic of Xiphiacetus cristatus USNM 21360, from the Calvert Formation, Middle Miocene o f the east coast of the USA, with a portion o f the pars cochlearis missing. A. lateral view. B. medial view. C. dorsal view. D. ventral view. Fig. 3 Posterior portion of the right periotic of Xiphiacetus cristatus USNM 21363, Calvert Formation, Middle Miocene of the east coast o f the USA. Ventral view, showing the shortened posterior process. Scale bar for Figs 2-3 = 10 mm. Fig. 4 Right tympanic bulla o f Xiphiacetus cristatus IRSNB M.1902,?Late Miocene o f Antwerp region. A. ventral view. B. medial view. C. dorsal view. Fig. 5 Left tympanic bulla o f Xiphiacetus cristatus USNM , Calvert Formation, Middle Miocene of the east coast of the USA. A. ventral view. B. medial view. Scale bar for Figs 4-5 = 20 mm. P late 4 Cervical and thoracic vertebrae o f Xiphiacetus cristatus IRSNB 3234-M.361 (holotype), Miocene of Antwerp region. A. atlas (cl) in dorsal view. B. atlas in anterior view. C. seven cervicals (cl-c7) in left lateral view. Note the ventrolaterally elongated parapophysis o f c6. D. axis (c2) in anterior view. E. third cervical (c3) in anterior view. F. six first thoracics (t 1-t6) in left lateral view. G. first thoracic (tl) in left lateral view. H. first thoracic in anterior view, illustrating measurements 6-9. I. first thoracic in dorsal view. Scale bars for A, B, D, E, G, H, I = 20 mm, for C, F = 50 mm.

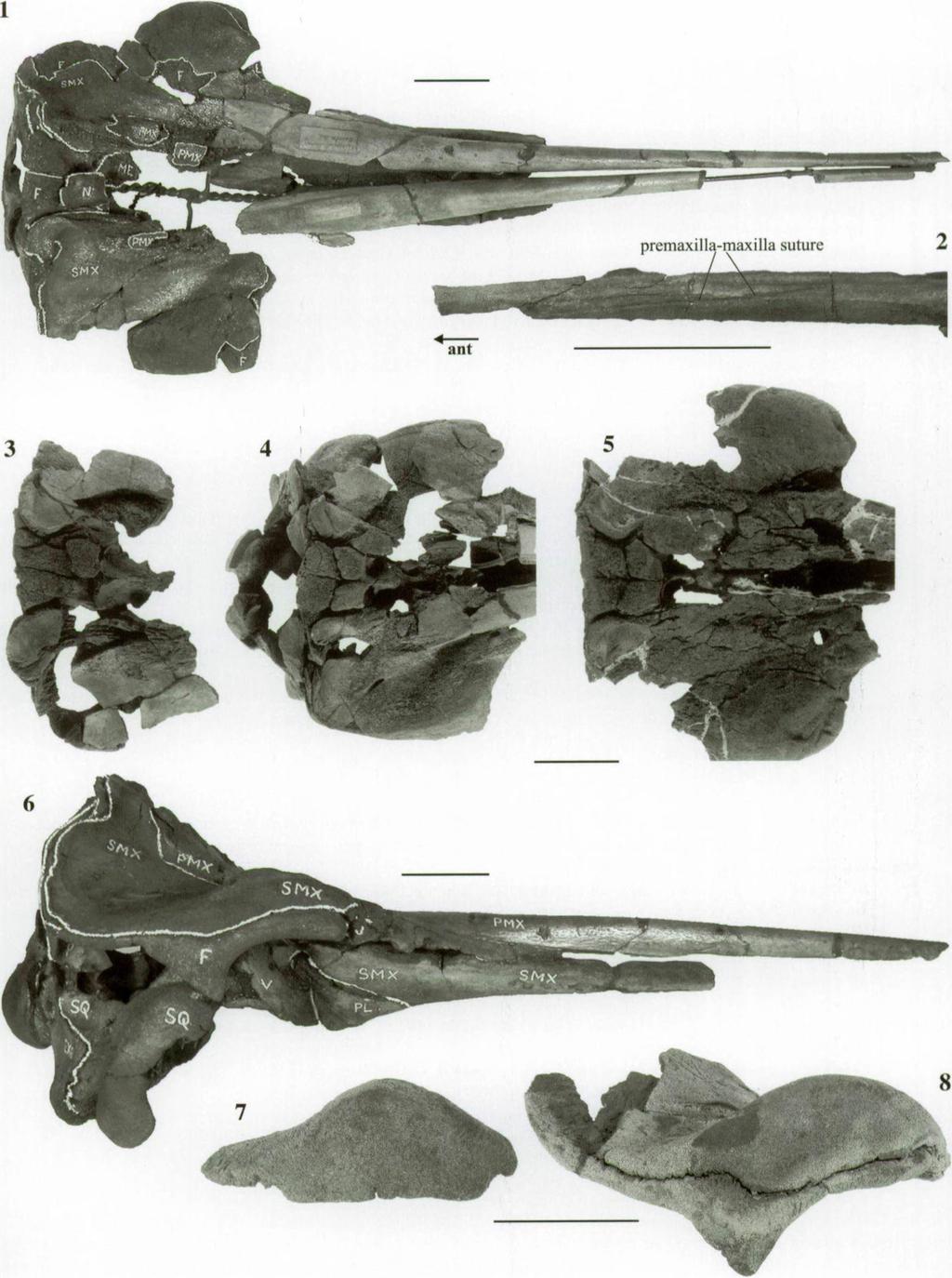
22 232 Olivier LAMBERT premaxilla-maxilla suture P la te 1



23 Review of Priscodelphinus cristatus and phylogeny 233 A fossa for f postorbital lobe of pterygoid sinus P l a t e 2


24 234 Olivier LAMBERT P l a t e 3


25 Review o f Priscodelphinus cristatus and phylogeny 6 P la te 4 235
Systematic revision of the Miocene long-snouted dolphin Eurhinodelphis longirostris DU Bus, 1872 (Cetacea, Odontoceti, Eurhinodelphinidae)
") BULLETIN DE L INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE SCIENCES DE LA TERRE, 74: 147-174, 2004 BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETENSCHAPPEN AARDWETENSCHAPPEN, 74: 147-174,
BULLETIN DE L INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE SCIENCES DE LA TERRE, 74: 147-174, 2004 BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETENSCHAPPEN AARDWETENSCHAPPEN, 74: 147-174,
A new archaic homodont toothed cetacean (Mammalia, Cetacea, Odontoceti) from the early Miocene of Peru
 from the early Miocene of Peru") A new archaic homodont toothed cetacean (Mammalia, Cetacea, Odontoceti) from the early Miocene of Peru Olivier LAMBERT Institut royal des Sciences naturelles de Belgique, D.O. Terre et Histoire de la Vie,
A new archaic homodont toothed cetacean (Mammalia, Cetacea, Odontoceti) from the early Miocene of Peru Olivier LAMBERT Institut royal des Sciences naturelles de Belgique, D.O. Terre et Histoire de la Vie,
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
2. Skull, total length versus length of the presacral vertebral column: (0); extremely elongated neck (e.g. Tanystropheus longobardicus).
; extremely elongated neck (e.g. Tanystropheus longobardicus).") Character list of the taxon-character data set 1. Skull and lower jaws, interdental plates: absent (0); present, but restricted to the anterior end of the dentary (1); present along the entire alveolar
Character list of the taxon-character data set 1. Skull and lower jaws, interdental plates: absent (0); present, but restricted to the anterior end of the dentary (1); present along the entire alveolar
AMERICAN MUSEUM NOVITATES Published by
 AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province
 from Dashanpu, Zigong Municipality, Sichuan Province") A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
First record of a platanistid (Cetacea, Odontoceti) in the North Sea Basin: a review of Cyrtodelphis Abel, 1899 from the Miocene of Belgium
 in the North Sea Basin: a review of Cyrtodelphis Abel, 1899 from the Miocene of Belgium") First record of a platanistid (Cetacea, Odontoceti) in the North Sea Basin: a review of Cyrtodelphis Abel, 1899 from the Miocene of Belgium Olivier Lambert 1 1 Institut royal des Sciences naturelles de
First record of a platanistid (Cetacea, Odontoceti) in the North Sea Basin: a review of Cyrtodelphis Abel, 1899 from the Miocene of Belgium Olivier Lambert 1 1 Institut royal des Sciences naturelles de
ONLINE APPENDIX 1. Morphological phylogenetic characters scored in this paper. See Poe (2004) for
 for") ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature13086 Part I. Supplementary Notes A: Detailed Description of Cotylocara macei gen. et sp. nov. Part II. Table of Measurements for holotype of Cotylocara macei (CCNHM-101) Part III. Supplementary
doi:10.1038/nature13086 Part I. Supplementary Notes A: Detailed Description of Cotylocara macei gen. et sp. nov. Part II. Table of Measurements for holotype of Cotylocara macei (CCNHM-101) Part III. Supplementary
Mammalogy Laboratory 1 - Mammalian Anatomy
 Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
SUPPLEMENTARY OBSERVATIONS ON THE SKULL OF
 SUPPLEMENTARY OBSERVATIONS ON THE SKULL OF THE FOSSIL PORPOISE ZARHACHIS FLAGELLATOR COPE By Remington Kellogg Of the Bureau of Biological Survey, United States Department of Agriculture During the past
SUPPLEMENTARY OBSERVATIONS ON THE SKULL OF THE FOSSIL PORPOISE ZARHACHIS FLAGELLATOR COPE By Remington Kellogg Of the Bureau of Biological Survey, United States Department of Agriculture During the past
YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
 Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia
 1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC
 FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the
 Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the") Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE
 PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA
 A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA MYCTEROSAURUS LONGICEPS S. W. WILLISTON University of Chicago The past summer, Mr. Herman Douthitt, of the University of Chicago paleontological expedition,
A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA MYCTEROSAURUS LONGICEPS S. W. WILLISTON University of Chicago The past summer, Mr. Herman Douthitt, of the University of Chicago paleontological expedition,
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE
 FROM THE") A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
SUPPLEMENTARY ONLINE MATERIAL FOR. Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor
 http://app.pan.pl/som/app61-ratsimbaholison_etal_som.pdf SUPPLEMENTARY ONLINE MATERIAL FOR Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor Ontogenetic changes in the craniomandibular
http://app.pan.pl/som/app61-ratsimbaholison_etal_som.pdf SUPPLEMENTARY ONLINE MATERIAL FOR Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor Ontogenetic changes in the craniomandibular
Mammalogy Lab 1: Skull, Teeth, and Terms
 Mammalogy Lab 1: Skull, Teeth, and Terms Be able to: Goals of today s lab Locate all structures listed on handout Define all terms on handout what they are or what they look like Give examples of mammals
Mammalogy Lab 1: Skull, Teeth, and Terms Be able to: Goals of today s lab Locate all structures listed on handout Define all terms on handout what they are or what they look like Give examples of mammals
List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to
 1 Supplementary data CHARACTER LIST List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to characters used by Tchernov et al. (2000), Rieppel, et al. (2002), and Lee
1 Supplementary data CHARACTER LIST List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to characters used by Tchernov et al. (2000), Rieppel, et al. (2002), and Lee
complex in cusp pattern. (3) The bones of the coyote skull are thinner, crests sharper and the
 The bones of the coyote skull are thinner, crests sharper and the") DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
Williston, and as there are many fairly good specimens in the American
 56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.)
") A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.) by Ouyang Hui Zigong Dinosaur Museum Newsletter Number 2 1989 pp. 10-14 Translated By Will Downs Bilby
A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.) by Ouyang Hui Zigong Dinosaur Museum Newsletter Number 2 1989 pp. 10-14 Translated By Will Downs Bilby
FIELDIANA GEOLOGY NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA
 FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
Museo di Storia Naturale del Mediterraneo, Via Roma 234, 57125, Livorno, Italy
 bs_bs_banner Zoological Journal of the Linnean Society, 2012, 166, 876 911. With 22 figures Comparative osteology and phylogenetic relationships of Miocaperea pulchra, the first fossil pygmy right whale
bs_bs_banner Zoological Journal of the Linnean Society, 2012, 166, 876 911. With 22 figures Comparative osteology and phylogenetic relationships of Miocaperea pulchra, the first fossil pygmy right whale
v:ii-ixi, 'i':;iisimvi'\>!i-:: "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi L I E) R.ARY OF THE VERSITY U N I or ILLINOIS REMO
 R.ARY OF THE VERSITY U N I or ILLINOIS REMO") "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
"^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
A Fossil Snake (Elaphe vulpina) From A Pliocene Ash Bed In Nebraska
 From A Pliocene Ash Bed In Nebraska") University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
Anatomical description and phylogenetic analysis of Miocene beaked whale from the East African Rift Valley, Kenya
 Southern Methodist University SMU Scholar Collection of Engaged Learning Engaged Learning 4-15-2013 Anatomical description and phylogenetic analysis of Miocene beaked whale from the East African Rift Valley,
Southern Methodist University SMU Scholar Collection of Engaged Learning Engaged Learning 4-15-2013 Anatomical description and phylogenetic analysis of Miocene beaked whale from the East African Rift Valley,
SUPPLEMENTARY INFORMATION
 Character 155, interdental ridges. Absence of interdental ridge (0) shown in Parasaniwa wyomingensis (Platynota). Interdental ridges (1) shown in Coniophis precedens. WWW.NATURE.COM/NATURE 1 Character
Character 155, interdental ridges. Absence of interdental ridge (0) shown in Parasaniwa wyomingensis (Platynota). Interdental ridges (1) shown in Coniophis precedens. WWW.NATURE.COM/NATURE 1 Character
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A.
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons
 Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons 1. Head skeleton of lamprey Cyclostomes are highly specialized in both the construction of the chondrocranium and visceral skeleton.
Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons 1. Head skeleton of lamprey Cyclostomes are highly specialized in both the construction of the chondrocranium and visceral skeleton.
Vol. XIV, No. 1, March, The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S.
 By S.") Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Anatomy. Name Section. The Vertebrate Skeleton
 Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
TRACHEMYS SCULPTA. A nearly complete articulated carapace and plastron of an Emjdd A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE,
 A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
WHxVLEBONE WHALE FROM THE CALVERT CLIFFS, MARYLAND.
 DESCRIPTION OF A NEW GENUS AND SPECIES OF WHxVLEBONE WHALE FROM THE CALVERT CLIFFS, MARYLAND. By Remington Kellogg, Of the Bureau of Biological Survey, United States Departm'ent of Agriculture. In the
DESCRIPTION OF A NEW GENUS AND SPECIES OF WHxVLEBONE WHALE FROM THE CALVERT CLIFFS, MARYLAND. By Remington Kellogg, Of the Bureau of Biological Survey, United States Departm'ent of Agriculture. In the
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO
 A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
Cranial osteology of the African gerrhosaurid Angolosaurus skoogi (Squamata; Gerrhosauridae) HOLLY A. NANCE
 HOLLY A. NANCE") African Journal of Herpetology, 2007 56(1): 39-75. Herpetological Association of Africa Original article Cranial osteology of the African gerrhosaurid Angolosaurus skoogi (Squamata; Gerrhosauridae) HOLLY
African Journal of Herpetology, 2007 56(1): 39-75. Herpetological Association of Africa Original article Cranial osteology of the African gerrhosaurid Angolosaurus skoogi (Squamata; Gerrhosauridae) HOLLY
A M E G H I N I A N A. Revista de la Asociación Paleontológia Argentina. Volume XV September-December 1978 Nos. 3-4
 A M E G H I N I A N A Revista de la Asociación Paleontológia Argentina Volume XV September-December 1978 Nos. 3-4 COLORADIA BREVIS N. G. ET N. SP. (SAURISCHIA, PROSAUROPODA), A PLATEOSAURID DINOSAUR FROM
A M E G H I N I A N A Revista de la Asociación Paleontológia Argentina Volume XV September-December 1978 Nos. 3-4 COLORADIA BREVIS N. G. ET N. SP. (SAURISCHIA, PROSAUROPODA), A PLATEOSAURID DINOSAUR FROM
HONR219D Due 3/29/16 Homework VI
 Part 1: Yet More Vertebrate Anatomy!!! HONR219D Due 3/29/16 Homework VI Part 1 builds on homework V by examining the skull in even greater detail. We start with the some of the important bones (thankfully
Part 1: Yet More Vertebrate Anatomy!!! HONR219D Due 3/29/16 Homework VI Part 1 builds on homework V by examining the skull in even greater detail. We start with the some of the important bones (thankfully
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * Dr. L.D. Boonstra. Paleontologist, South African Museum, Cape Town
 THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL
 Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA
 FROM JAVA, INDONESIA") NOTES AND NEWS UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA BY NGUYEN NGOC-HO i) Faculty of Science, University of Saigon, Vietnam Among material recently collected
NOTES AND NEWS UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA BY NGUYEN NGOC-HO i) Faculty of Science, University of Saigon, Vietnam Among material recently collected
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/329/5998/1481/dc1 Supporting Online Material for Tyrannosaur Paleobiology: New Research on Ancient Exemplar Organisms Stephen L. Brusatte,* Mark A. Norell, Thomas D.
www.sciencemag.org/cgi/content/full/329/5998/1481/dc1 Supporting Online Material for Tyrannosaur Paleobiology: New Research on Ancient Exemplar Organisms Stephen L. Brusatte,* Mark A. Norell, Thomas D.
An Early Miocene long-snouted marine platanistid dolphin (Mammalia, Cetacea, Odontoceti) from the Korneuburg Basin (Austria)
 from the Korneuburg Basin (Austria)") Beitr. Paläont., 27: 407-418, Wien 2002 An Early Miocene long-snouted marine platanistid dolphin (Mammalia, Cetacea, Odontoceti) from the Korneuburg Basin (Austria) by Lawrence G. Barnes* B arnes, L. G.
Beitr. Paläont., 27: 407-418, Wien 2002 An Early Miocene long-snouted marine platanistid dolphin (Mammalia, Cetacea, Odontoceti) from the Korneuburg Basin (Austria) by Lawrence G. Barnes* B arnes, L. G.
BREVIORA LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1
, A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1") ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
OF THE TRIAS THE PHYTOSAURIA
 THE PHYTOSAURIA OF THE TRIAS MAURICE G. MEHL University of Wisconsin Some time ago the writer gave a brief notice of a new genus of phytosaurs of which Angistorhinus grandis Mehl was the type.' It is the
THE PHYTOSAURIA OF THE TRIAS MAURICE G. MEHL University of Wisconsin Some time ago the writer gave a brief notice of a new genus of phytosaurs of which Angistorhinus grandis Mehl was the type.' It is the
.56 m. (22 in.). COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition
. COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition") Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev Doklady Akademii Nauk, SSSR 87:
 translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE
 OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT This is a report of measurements on the skeleton of a male se1 whale caught in the Antarctic. The skeleton of
OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT This is a report of measurements on the skeleton of a male se1 whale caught in the Antarctic. The skeleton of
The cranial osteology of Belebey vegrandis (Parareptilia: Bolosauridae), from the Middle Permian of Russia, and its bearing on reptilian evolution
, from the Middle Permian of Russia, and its bearing on reptilian evolution") Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1511 191214 Original Articles RUSSIAN BOLOSAURID REPTILER. R. REISZ ET AL.
Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1511 191214 Original Articles RUSSIAN BOLOSAURID REPTILER. R. REISZ ET AL.
Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved
 Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved This was a private report in 2003 on my thoughts on Platecarpus planifrons.
Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved This was a private report in 2003 on my thoughts on Platecarpus planifrons.
A new species of sauropod, Mamenchisaurus anyuensis sp. nov.
 A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
A New Ceratopsian Dinosaur from the Upper
 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 63. NUMBER 3 A New Ceratopsian Dinosaur from the Upper Cretaceous of Montana, with Note on Hypacrosaurus (With Two Plates) CHARLES W. GILMORE Assistant Curator
SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 63. NUMBER 3 A New Ceratopsian Dinosaur from the Upper Cretaceous of Montana, with Note on Hypacrosaurus (With Two Plates) CHARLES W. GILMORE Assistant Curator
MIOCENE DIATOMACEOUS EARTH OF LOMPOC, CALI- FORNIA.i
 DESCRIPTION OF THE SKULL OF MEGAPTERA MIO- CAENA, A FOSSIL HUMPBACK WHALE FROM THE MIOCENE DIATOMACEOUS EARTH OF LOMPOC, CALI- FORNIA.i By Remington Kellogg. Of the Bureau of Biological Survey, Department
DESCRIPTION OF THE SKULL OF MEGAPTERA MIO- CAENA, A FOSSIL HUMPBACK WHALE FROM THE MIOCENE DIATOMACEOUS EARTH OF LOMPOC, CALI- FORNIA.i By Remington Kellogg. Of the Bureau of Biological Survey, Department
Notes on Ceratopsians and Ankylosaurs at the Royal Ontario Museum
 Notes on Ceratopsians and Ankylosaurs at the Royal Ontario Museum Andrew A. Farke, Ph.D. Raymond M. Alf Museum of Paleontology 1175 West Baseline Road Claremont, CA 91711 email: afarke@webb.org Introduction
Notes on Ceratopsians and Ankylosaurs at the Royal Ontario Museum Andrew A. Farke, Ph.D. Raymond M. Alf Museum of Paleontology 1175 West Baseline Road Claremont, CA 91711 email: afarke@webb.org Introduction
A skull without mandihle, from the Hunterian Collection (no.
 4 MR. G. A. BOULENGER ON CHELONIAN REMAINS. [Jan. 6, 2. On some Chelonian Remains preserved in the Museum of the Eojal College of Surgeons. By G. A. Boulenger. [Eeceived December 8, 1890.] In the course
4 MR. G. A. BOULENGER ON CHELONIAN REMAINS. [Jan. 6, 2. On some Chelonian Remains preserved in the Museum of the Eojal College of Surgeons. By G. A. Boulenger. [Eeceived December 8, 1890.] In the course
Cranial osteology and phylogenetic relationships of Hamadasuchus rebouli (Crocodyliformes: Mesoeucrocodylia) from the Cretaceous of Morocco
 from the Cretaceous of Morocco") Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1494 533567 Original Articles HAMADASUCHUS REBOULIH. C. E. LARSSON and H.-D.
Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1494 533567 Original Articles HAMADASUCHUS REBOULIH. C. E. LARSSON and H.-D.
NOTES ON THE FIRST SKULL AND JAWS OF RIOJASAURUS INCERTUS (DINOSAURIA, PROSAUROPODA, MELANOROSAURIDAE) OF THE LATE TRIASSIC OF LA RIOJA, ARGENTINA
 OF THE LATE TRIASSIC OF LA RIOJA, ARGENTINA") NOTES ON THE FIRST SKULL AND JAWS OF RIOJASAURUS INCERTUS (DINOSAURIA, PROSAUROPODA, MELANOROSAURIDAE) OF THE LATE TRIASSIC OF LA RIOJA, ARGENTINA José F. Bonaparte and José A. Pumares translated by Jeffrey
NOTES ON THE FIRST SKULL AND JAWS OF RIOJASAURUS INCERTUS (DINOSAURIA, PROSAUROPODA, MELANOROSAURIDAE) OF THE LATE TRIASSIC OF LA RIOJA, ARGENTINA José F. Bonaparte and José A. Pumares translated by Jeffrey
Description of Cranial Elements and Ontogenetic Change within Tropidolaemus wagleri (Serpentes: Crotalinae).
.") East Tennessee State University Digital Commons @ East Tennessee State University Electronic Theses and Dissertations 5-2016 Description of Cranial Elements and Ontogenetic Change within Tropidolaemus
East Tennessee State University Digital Commons @ East Tennessee State University Electronic Theses and Dissertations 5-2016 Description of Cranial Elements and Ontogenetic Change within Tropidolaemus
A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province
 A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province by Hu Shaojin (Kunming Cultural Administrative Committee, Yunnan Province) Vertebrata PalAsiatica Vol. XXXI, No. 1
A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province by Hu Shaojin (Kunming Cultural Administrative Committee, Yunnan Province) Vertebrata PalAsiatica Vol. XXXI, No. 1
 The following text is generated from uncorrected OCR. [Begin Page: Page 1] A NEW CERATOPSIAN DINOSAUR FROM THE UPPER CRETACEOUS OF MONTANA, WITH NOTE ON HYPACROSAURUS ' By CHARLES W. GILMORE assistant
The following text is generated from uncorrected OCR. [Begin Page: Page 1] A NEW CERATOPSIAN DINOSAUR FROM THE UPPER CRETACEOUS OF MONTANA, WITH NOTE ON HYPACROSAURUS ' By CHARLES W. GILMORE assistant
VERTEBRATA PALASIATICA
 41 2 2003 2 VERTEBRATA PALASIATICA pp. 147 156 figs. 1 5 1) ( 100044), ( Parakannemeyeria brevirostris),,, : ( Xiyukannemeyeria),,, Q915. 864 60 Turfania (,1973), Dicynodon (, 1973 ; Lucas, 1998), (Lystrosaurus)
41 2 2003 2 VERTEBRATA PALASIATICA pp. 147 156 figs. 1 5 1) ( 100044), ( Parakannemeyeria brevirostris),,, : ( Xiyukannemeyeria),,, Q915. 864 60 Turfania (,1973), Dicynodon (, 1973 ; Lucas, 1998), (Lystrosaurus)
SOME NEW AMERICAN PYCNODONT FISHES.
 SOME NEW AMERICAN PYCNODONT FISHES. By James Williams Gidley, Assistant Curator of Fossil Mammals, United States National Museum. In the United States National Museum are several specimens representing
SOME NEW AMERICAN PYCNODONT FISHES. By James Williams Gidley, Assistant Curator of Fossil Mammals, United States National Museum. In the United States National Museum are several specimens representing
PALEONTOLOGY AND BIOSTRATIGRAPHY OF MONGOLIA
 PALEONTOLOGY AND BIOSTRATIGRAPHY OF MONGOLIA THE JOINT SOVIET-MONGOLIAN PALEONTOLOGICAL EXPEDITION (Transactions, vol. 3) EDITORIAL BOARD: N. N. Kramarenko (editor-in-chief) B. Luvsandansan, Yu. I. Voronin,
PALEONTOLOGY AND BIOSTRATIGRAPHY OF MONGOLIA THE JOINT SOVIET-MONGOLIAN PALEONTOLOGICAL EXPEDITION (Transactions, vol. 3) EDITORIAL BOARD: N. N. Kramarenko (editor-in-chief) B. Luvsandansan, Yu. I. Voronin,
Title: Phylogenetic Methods and Vertebrate Phylogeny
 Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
A NEW SALTICID SPIDER FROM VICTORIA By R. A. Dunn
 Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
NAUSHONIA PAN AMEN SIS, NEW SPECIES (DECAPODA: THALASSINIDEA: LAOMEDIIDAE) FROM THE PACIFIC COAST OF PANAMA, WITH NOTES ON THE GENUS
 FROM THE PACIFIC COAST OF PANAMA, WITH NOTES ON THE GENUS") 5 October 1982 PROC. BIOL. SOC. WASH. 95(3), 1982, pp. 478-483 NAUSHONIA PAN AMEN SIS, NEW SPECIES (DECAPODA: THALASSINIDEA: LAOMEDIIDAE) FROM THE PACIFIC COAST OF PANAMA, WITH NOTES ON THE GENUS Joel
5 October 1982 PROC. BIOL. SOC. WASH. 95(3), 1982, pp. 478-483 NAUSHONIA PAN AMEN SIS, NEW SPECIES (DECAPODA: THALASSINIDEA: LAOMEDIIDAE) FROM THE PACIFIC COAST OF PANAMA, WITH NOTES ON THE GENUS Joel
A New Pterosaur from the Middle Jurassic of Dashanpu, Zigong, Sichuan
 A New Pterosaur from the Middle Jurassic of Dashanpu, Zigong, Sichuan by Xinlu He (Chengdu College of Geology) Daihuan Yang (Chungking Natural History Museum, Sichuan Province) Chunkang Su (Zigong Historical
A New Pterosaur from the Middle Jurassic of Dashanpu, Zigong, Sichuan by Xinlu He (Chengdu College of Geology) Daihuan Yang (Chungking Natural History Museum, Sichuan Province) Chunkang Su (Zigong Historical
NEW INFORMATION ON THE CRANIUM OF BRACHYLOPHOSAURUS CANADENSIS (DINOSAURIA, HADROSAURIDAE), WITH A REVISION OF ITS PHYLOGENETIC POSITION
, WITH A REVISION OF ITS PHYLOGENETIC POSITION") Journal of Vertebrate Paleontology 25(1):144 156, March 2005 2005 by the Society of Vertebrate Paleontology NEW INFORMATION ON THE CRANIUM OF BRACHYLOPHOSAURUS CANADENSIS (DINOSAURIA, HADROSAURIDAE), WITH
Journal of Vertebrate Paleontology 25(1):144 156, March 2005 2005 by the Society of Vertebrate Paleontology NEW INFORMATION ON THE CRANIUM OF BRACHYLOPHOSAURUS CANADENSIS (DINOSAURIA, HADROSAURIDAE), WITH
A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae)
") Genus Vol. 14 (3): 413-418 Wroc³aw, 15 X 2003 A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae) JAROS AW KANIA Zoological Institute, University of Wroc³aw, Sienkiewicza
Genus Vol. 14 (3): 413-418 Wroc³aw, 15 X 2003 A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae) JAROS AW KANIA Zoological Institute, University of Wroc³aw, Sienkiewicza
Diurus, Pascoe. sp. 1). declivity of the elytra, but distinguished. Length (the rostrum and tails 26 included) mm. Deep. exception
. declivity of the elytra, but distinguished. Length (the rostrum and tails 26 included) mm. Deep. exception") 210 DIURUS ERYTIIROPUS. NOTE XXVI. Three new species of the Brenthid genus Diurus, Pascoe DESCRIBED BY C. Ritsema+Cz. 1. Diurus erythropus, n. sp. 1). Allied to D. furcillatus Gylh. ²) by the short head,
210 DIURUS ERYTIIROPUS. NOTE XXVI. Three new species of the Brenthid genus Diurus, Pascoe DESCRIBED BY C. Ritsema+Cz. 1. Diurus erythropus, n. sp. 1). Allied to D. furcillatus Gylh. ²) by the short head,
THE SKULLS OF ARAEOSCELIS AND CASEA, PERMIAN REPTILES
 THE SKULLS OF REOSCELIS ND CSE, PERMIN REPTILES University of Chicago There are few Permian reptiles of greater interest at the present time than the peculiar one I briefly described in this journal' three
THE SKULLS OF REOSCELIS ND CSE, PERMIN REPTILES University of Chicago There are few Permian reptiles of greater interest at the present time than the peculiar one I briefly described in this journal' three
Results of Prof. E. Stromer's Research Expedition in the Deserts of Egypt
 Proceedings of the Royal Bavarian Academy of Science Mathematical-physical Division Volume XXVIII, Paper 3 Results of Prof. E. Stromer's Research Expedition in the Deserts of Egypt II. Vertebrate Remains
Proceedings of the Royal Bavarian Academy of Science Mathematical-physical Division Volume XXVIII, Paper 3 Results of Prof. E. Stromer's Research Expedition in the Deserts of Egypt II. Vertebrate Remains
ZOOLOGISCHE MEDEDELINGEN
 "f ~- >D noitnwz, tito ZOOLOGISCHE MEDEDELINGEN UITGEGEVEN DOOR HET RIJKSMUSEUM VAN NATUURLIJKE HISTORIE TE LEIDEN (MINISTERIE VAN CULTUUR, RECREATIE EN MAATSCHAPPELIJK WERK) Deel 48 no. 25 25 maart 1975
"f ~- >D noitnwz, tito ZOOLOGISCHE MEDEDELINGEN UITGEGEVEN DOOR HET RIJKSMUSEUM VAN NATUURLIJKE HISTORIE TE LEIDEN (MINISTERIE VAN CULTUUR, RECREATIE EN MAATSCHAPPELIJK WERK) Deel 48 no. 25 25 maart 1975
A REDESCRIPTION OF THE HOLOTYPE OF CALLIANASSA MUCRONATA STRAHL, 1861 (DECAPODA, THALASSINIDEA)
") Crustaceana 52 (1) 1977, E. J. Brill, Leiden A REDESCRIPTION OF THE HOLOTYPE OF CALLIANASSA MUCRONATA STRAHL, 1861 (DECAPODA, THALASSINIDEA) BY NASIMA M. TIRMIZI Department of Zoology, University of Karachi,
Crustaceana 52 (1) 1977, E. J. Brill, Leiden A REDESCRIPTION OF THE HOLOTYPE OF CALLIANASSA MUCRONATA STRAHL, 1861 (DECAPODA, THALASSINIDEA) BY NASIMA M. TIRMIZI Department of Zoology, University of Karachi,
FIRST RECORD OF MESOPLODON DENSIROSTRIS FROM FORMOSA
 FIRST RECORD OF MESOPLODON DENSIROSTRIS FROM FORMOSA TOSHIO KASUYA* AND MASAHARU NISHIWAKI* ABSTRACT Two records of female Mesoplodon densirostris are reported. Comments on the external character, skull
FIRST RECORD OF MESOPLODON DENSIROSTRIS FROM FORMOSA TOSHIO KASUYA* AND MASAHARU NISHIWAKI* ABSTRACT Two records of female Mesoplodon densirostris are reported. Comments on the external character, skull
The family Gnaphosidae is a large family
 Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
DISCOVERY OF GENUS PLATOLENES (COLEOP TERA : TENEBRIONIDAE) FROM INDIA WITH DESCRIPTION OF TWO NEW SPECIES G. N. SABA
 FROM INDIA WITH DESCRIPTION OF TWO NEW SPECIES G. N. SABA") Rec. zool. Surv. India, 85(3) : 433-437,1988 DISCOVERY OF GENUS PLATOLENES (COLEOP TERA : TENEBRIONIDAE) FROM INDIA WITH DESCRIPTION OF TWO NEW SPECIES By G. N. SABA Zoological Survey of India M-Block,
Rec. zool. Surv. India, 85(3) : 433-437,1988 DISCOVERY OF GENUS PLATOLENES (COLEOP TERA : TENEBRIONIDAE) FROM INDIA WITH DESCRIPTION OF TWO NEW SPECIES By G. N. SABA Zoological Survey of India M-Block,
CRANIAL OSTEOLOGY OF SCHIZOTHORAICHTHYS NIGER (MECKEL) MISRA (CYPRINIDAE: SCHIZOTHORACINAE). L NEUROCRANIUM
 MISRA (CYPRINIDAE: SCHIZOTHORACINAE). L NEUROCRANIUM") CRANIAL OSTEOLOGY OF SCHIZOTHORAICHTHYS NIGER (MECKEL) MISRA (CYPRINIDAE: SCHIZOTHORACINAE). L NEUROCRANIUM A. R. YousuF, A. K. PANDIT AND A. R. KHAN Postgraduate Department of Zoology, University of Kashmir,
CRANIAL OSTEOLOGY OF SCHIZOTHORAICHTHYS NIGER (MECKEL) MISRA (CYPRINIDAE: SCHIZOTHORACINAE). L NEUROCRANIUM A. R. YousuF, A. K. PANDIT AND A. R. KHAN Postgraduate Department of Zoology, University of Kashmir,
SUBFAMILY THYMOPINAE Holthuis, 1974
 click for previous page 29 Remarks : The taxonomy of the species is not clear. It is possible that 2 forms may have to be distinguished: A. sublevis Wood-Mason, 1891 (with a synonym A. opipara Burukovsky
click for previous page 29 Remarks : The taxonomy of the species is not clear. It is possible that 2 forms may have to be distinguished: A. sublevis Wood-Mason, 1891 (with a synonym A. opipara Burukovsky
A NEW SPECIES OF A USTROLIBINIA FROM THE SOUTH CHINA SEA AND INDONESIA (CRUSTACEA: BRACHYURA: MAJIDAE)
") 69 C O a g r ^ j^a RAFFLES BULLETIN OF ZOOLOGY 1992 40(1): 69-73 A NEW SPECIES OF A USTROLIBINIA FROM THE SOUTH CHINA SEA AND INDONESIA (CRUSTACEA: BRACHYURA: MAJIDAE) H P Waener SMITHSONIAN INSTITUTE
69 C O a g r ^ j^a RAFFLES BULLETIN OF ZOOLOGY 1992 40(1): 69-73 A NEW SPECIES OF A USTROLIBINIA FROM THE SOUTH CHINA SEA AND INDONESIA (CRUSTACEA: BRACHYURA: MAJIDAE) H P Waener SMITHSONIAN INSTITUTE
CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN OF RUSSIA AND THE EVOLUTIONARY RELATIONSHIPS OF CASEIDAE
 FROM THE MIDDLE PERMIAN OF RUSSIA AND THE EVOLUTIONARY RELATIONSHIPS OF CASEIDAE") Journal of Vertebrate Paleontology 28(1):160 180, March 2008 2008 by the Society of Vertebrate Paleontology ARTICLE CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN
Journal of Vertebrate Paleontology 28(1):160 180, March 2008 2008 by the Society of Vertebrate Paleontology ARTICLE CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN
A new carnosaur from Yongchuan County, Sichuan Province
 A new carnosaur from Yongchuan County, Sichuan Province by Dong Zhiming Institute of Vertebrate Palaeontology and Palaeoanthropology, Academia Sinica Zhang Yihong, Li Xuanmin, and Zhou Shiwu Chongqing
A new carnosaur from Yongchuan County, Sichuan Province by Dong Zhiming Institute of Vertebrate Palaeontology and Palaeoanthropology, Academia Sinica Zhang Yihong, Li Xuanmin, and Zhou Shiwu Chongqing
A new kentriodontid (Cetacea: Delphinoidea) from the Middle Miocene of Hungary
 from the Middle Miocene of Hungary") Mitt. Mus. Nat.kd. Berl., Geowiss. Reihe 8 (2005), 53 73 / DOI 10.1002/mmng.200410004 A new kentriodontid (Cetacea: Delphinoidea) from the Middle Miocene of Hungary Emese Kazár* Geological Institute of
Mitt. Mus. Nat.kd. Berl., Geowiss. Reihe 8 (2005), 53 73 / DOI 10.1002/mmng.200410004 A new kentriodontid (Cetacea: Delphinoidea) from the Middle Miocene of Hungary Emese Kazár* Geological Institute of
Temporal lines. More forwardfacing. tubular orbits than in the African forms 3. Orbits larger relative to skull size than in the other genera 2.
 Asian lorises More forwardfacing and tubular orbits than in the African forms 3. Characterized by a marked extension of the ectotympanic into a tubular meatus and a more angular auditory bulla than in
Asian lorises More forwardfacing and tubular orbits than in the African forms 3. Characterized by a marked extension of the ectotympanic into a tubular meatus and a more angular auditory bulla than in
35. DATA REPORT: CRETACEOUS OSTRACODES FROM HOLES 865A AND 866A (MID-PACIFIC MOUNTAINS) 1. Renée Damotte 2
 1. Renée Damotte 2") Winterer, E.L., Sager, W.W., Firth, J.V., and Sinton, J.M. (Eds.), 1995 Proceedings of the Ocean Drilling Program, Scientific Results, Vol. 143 35. DATA REPORT: CRETACEOUS OSTRACODES FROM HOLES 865A AND
Winterer, E.L., Sager, W.W., Firth, J.V., and Sinton, J.M. (Eds.), 1995 Proceedings of the Ocean Drilling Program, Scientific Results, Vol. 143 35. DATA REPORT: CRETACEOUS OSTRACODES FROM HOLES 865A AND
( M amenchisaurus youngi Pi, Ouyang et Ye, 1996)
") 39 4 2001 10 V ERTEBRATA PALASIATICA pp. 266 271 fig. 1,pl. I ( 643013), ( M amenchisaurus hochuanensis),,, Q915. 864 1995 12 31 (ZDM0126) ( M amenchisau rus hochuanensis Young et Chao, 1972),,, ZDM0126
39 4 2001 10 V ERTEBRATA PALASIATICA pp. 266 271 fig. 1,pl. I ( 643013), ( M amenchisaurus hochuanensis),,, Q915. 864 1995 12 31 (ZDM0126) ( M amenchisau rus hochuanensis Young et Chao, 1972),,, ZDM0126
AN OSTEOLOGICAL STUDY OF THE CUVIER'S BEAKED WHALE,,ZIPHIUS CAVIROSTRIS, IN THE NORTHWEST PACIFIC
 AN OSTEOLOGICAL STUDY OF THE CUVIER'S BEAKED WHALE,,ZIPHIUS CAVIROSTRIS, IN THE NORTHWEST PACIFIC HIDEO OMURA ABSTRACT Skulls and postcranial skeletons of :(,iphius cavirostris were studied for eleven
AN OSTEOLOGICAL STUDY OF THE CUVIER'S BEAKED WHALE,,ZIPHIUS CAVIROSTRIS, IN THE NORTHWEST PACIFIC HIDEO OMURA ABSTRACT Skulls and postcranial skeletons of :(,iphius cavirostris were studied for eleven
Florida, Gainesville, Florida, 32611, U.S.A. b Smithsonian Tropical Research Institute, Ancon, Republic of Panama,
 This article was downloaded by: [78.22.97.164] On: 04 May 2013, At: 14:02 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer
This article was downloaded by: [78.22.97.164] On: 04 May 2013, At: 14:02 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer
TERRIER BRASILEIRO (Brazilian Terrier)
") 04.07.2018/ EN FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) SECRETARIAT GENERAL: 13, Place Albert 1 er B 6530 Thuin (Belgique) FCI-Standard N 341 TERRIER BRASILEIRO (Brazilian Terrier) 2 TRANSLATION:
04.07.2018/ EN FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) SECRETARIAT GENERAL: 13, Place Albert 1 er B 6530 Thuin (Belgique) FCI-Standard N 341 TERRIER BRASILEIRO (Brazilian Terrier) 2 TRANSLATION:
BRAZILIAN TERRIER (Terrier Brasileiro)
") FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) SECRETARIAT GENERAL: 13, Place Albert 1er B 6530 Thuin (Belgique) 06.09.2013 / EN FCI-Standard N 341 BRAZILIAN TERRIER (Terrier Brasileiro) This illustration
FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) SECRETARIAT GENERAL: 13, Place Albert 1er B 6530 Thuin (Belgique) 06.09.2013 / EN FCI-Standard N 341 BRAZILIAN TERRIER (Terrier Brasileiro) This illustration
Major cranial changes during Triceratops ontogeny John R. Horner 1, * and Mark B. Goodwin 2
 273, 2757 2761 doi:10.1098/rspb.2006.3643 Published online 1 August 2006 Major cranial changes during Triceratops ontogeny John R. Horner 1, * and Mark B. Goodwin 2 1 Museum of the Rockies, Montana State
273, 2757 2761 doi:10.1098/rspb.2006.3643 Published online 1 August 2006 Major cranial changes during Triceratops ontogeny John R. Horner 1, * and Mark B. Goodwin 2 1 Museum of the Rockies, Montana State
A new dolphin from the early Miocene of Patagonia, Argentina: Insights into the evolution of Platanistoidea in the Southern Hemisphere
 A new dolphin from the early Miocene of Patagonia, Argentina: Insights into the evolution of Platanistoidea in the Southern Hemisphere MARIANA VIGLINO, MÓNICA R. BUONO, CAROLINA S. GUTSTEIN, MARIO A. COZZUOL,
A new dolphin from the early Miocene of Patagonia, Argentina: Insights into the evolution of Platanistoidea in the Southern Hemisphere MARIANA VIGLINO, MÓNICA R. BUONO, CAROLINA S. GUTSTEIN, MARIO A. COZZUOL,
CENE RUMINANTS OF THE GENERA OVIBOS AND
 DESCRIPTIONS OF TWO NEW SPECIES OF PLEISTO- CENE RUMINANTS OF THE GENERA OVIBOS AND BOOTHERIUM, WITH NOTES ON THE LATTER GENUS. By James Williams Gidley, Of the United States National Museum. Two interesting
DESCRIPTIONS OF TWO NEW SPECIES OF PLEISTO- CENE RUMINANTS OF THE GENERA OVIBOS AND BOOTHERIUM, WITH NOTES ON THE LATTER GENUS. By James Williams Gidley, Of the United States National Museum. Two interesting
PALEONTOLOGICAL CONTRIBUTIONS
 THE UNIVERSITY OF KANSAS PALEONTOLOGICAL CONTRIBUTIONS August, 1965 Paper 2 A NEW WYOMING PHYTOSAUR By THEODORE H. EATON, JR. [Museum of Natural History, University of Kansas I ABSTRACT The skull of a
THE UNIVERSITY OF KANSAS PALEONTOLOGICAL CONTRIBUTIONS August, 1965 Paper 2 A NEW WYOMING PHYTOSAUR By THEODORE H. EATON, JR. [Museum of Natural History, University of Kansas I ABSTRACT The skull of a
4. Premaxilla: Foramen on the lateral surface of the premaxillary body (Yates 2007 ch. 4) 0 absent 1 present
 0 absent 1 present") The character matrix used as a basis for this study is that of Yates et al (2010) which is modified from the earlier matrix used by Yates (2007). This matrix includes characters acquired and/or modified
The character matrix used as a basis for this study is that of Yates et al (2010) which is modified from the earlier matrix used by Yates (2007). This matrix includes characters acquired and/or modified
PICCOLO LEVRIERO ITALIANO
 FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) SECRETARIAT GENERAL: 13, Place Albert 1 er B 6530 Thuin (Belgique) 17.12.2015/ EN FCI-Standard N 200 PICCOLO LEVRIERO ITALIANO (Italian Sighthound) 2 TRANSLATION:
FEDERATION CYNOLOGIQUE INTERNATIONALE (AISBL) SECRETARIAT GENERAL: 13, Place Albert 1 er B 6530 Thuin (Belgique) 17.12.2015/ EN FCI-Standard N 200 PICCOLO LEVRIERO ITALIANO (Italian Sighthound) 2 TRANSLATION:
University of Iowa Iowa Research Online
 University of Iowa Iowa Research Online Theses and Dissertations Spring 2016 A reassessment of the late Eocene - early Oligocene crocodylids Crocodylus megarhinus Andrews 1905 and Crocodylus articeps Andrews
University of Iowa Iowa Research Online Theses and Dissertations Spring 2016 A reassessment of the late Eocene - early Oligocene crocodylids Crocodylus megarhinus Andrews 1905 and Crocodylus articeps Andrews
MUNIDOPSIS ALBATROSSAB, A NEW SPECIES OF DEEP-SEA GALATHEIDAE (DECAPODA, ANOMURA) FROM THE EASTERN PACIFIC OCEAN
 FROM THE EASTERN PACIFIC OCEAN") MUNIDOPSIS ALBATROSSAB, A NEW SPECIES OF DEEP-SEA GALATHEIDAE (DECAPODA, ANOMURA) FROM THE EASTERN PACIFIC OCEAN BY WILLIS E. PEQUEGNAT and LINDA H. PEQUEGNAT Department of Oceanography, Texas A & M University,
MUNIDOPSIS ALBATROSSAB, A NEW SPECIES OF DEEP-SEA GALATHEIDAE (DECAPODA, ANOMURA) FROM THE EASTERN PACIFIC OCEAN BY WILLIS E. PEQUEGNAT and LINDA H. PEQUEGNAT Department of Oceanography, Texas A & M University,