SUPPLEMENTARY OBSERVATIONS ON THE SKULL OF
|
|
|
- Claud Young
- 5 years ago
- Views:
Transcription



1 SUPPLEMENTARY OBSERVATIONS ON THE SKULL OF THE FOSSIL PORPOISE ZARHACHIS FLAGELLATOR COPE By Remington Kellogg Of the Bureau of Biological Survey, United States Department of Agriculture During the past ten years a rather large number of vertebrae and other portions of skeletons of cetaceans have been obtained from the Calvert Miocene formation of Maryland. Most of these specimens were obtained by digging into the Calvert Cliffs. During severe storms many tons of sandy clay are dislodged from the face Many of the cliff by the undercutting action of the incoming tide. specimens are destroyed in this manner and at the same time others are exposed. Worn vertebrae and fragments of bones are found at frequent intervals on the beach along the Calvert Cliffs after a storm. With the exception of the zygomatic processes and the condyles, the bones of the skull do not withstand being rolled about by the waves and are soon broken up. When remains of pelagic mammals are found undisturbed in the greenish sandy clay, they are usually well preserved. Complete skeletons of these mammals are rarely found, but broken ends of bones and portions of skulls are often found protruding from the face of the cliff. Norman H. Boss, to whom we are indebted for most of the specimens described in this and the preceding papers, 1 has been extremely fortunate in locating and collecting specimens of fossil porpoises. One of the skulls which he collected during the past year belongs to the Miocene porpoise, Zarhachis flagellator. Although incomplete, this skull supplies most of the structural details which were missing on the other specimen. The locality at which this skull was obtained is less than 2% miles south of the previous discovery. The only certainly known remains of ZarhacMs flagellator from the Atlantic coast province of North America belong to four individuals. One of these is the type specimen, a single anterior caudal vertebra; a fairly complete skeleton which was described in 1924 represents the second specimen; the third is the cranial portion of a skull hereinafter described and figured; and of the fourth the right periotic alone was collected. ' Kellogg, R., A fossil porpoise from the Calvert formation of Maryland. Proc. U. S. Nat. Mus., vol. 63, publ. 2482, pp. 1-39, pis March 26, NO PROCEEDINGS U. S. NATIONAL MUSEUM, VOL. 67, ART f 1 1


2 2 PROCEEDINGS OF THE NATIONAL MUSEUM vol.67 The Calvert formation of Maryland has yielded a rather large number of porpoises and in the main these species have their nearest affinities with those in the Tortonian stage of Europe. A few of them appear to have no counterparts in the European stages or at least none have been described and one of these is Zarhachis. Without entering into a discussion of the fauna as a whole, it is sufficient to observe in the present connection that Zarhachis appears to represent a highly specialized aberrant type whose family allocation is more or less a matter of personal opinion in the light of available data. If it is desirable to associate this porpoise with other genera, it must be done with the understanding that it is merely a matter of convenience. That Zarhachis exhibits certain characters in common with Inia, Lipotes, and Platanista, there can be no doubt, but whether they arose from one or three types of toothed whales can not be demonstrated from the specimens now known. According to our present knowledge, Zarhachis represents a type which can not be referred to any of the recognized families of toothed whales, unless the limits of these families be redefined. A natural grouping of some of the South American Miocene porpoises is not possible at present because of the lack of adequate data on the construction of their skulls. The ancestry of the Zarhachis type of porpoise is completely unknown at present. No fossil porpoises with this type of skull have been described from the Miocene formations of Europe. For this reason there can be little doubt but that it represents a migrant from some other region, probably the south Atlantic, which became associated with more widely distributed types during the latter part of the Miocene period. The skull of Zarhachis is modified in many ways, as will be noted in the descriptive portions of this and the preceding paper. For the purpose of bringing into stronger relief the characters of this porpoise, it is necessary to compare them carefully with living river porpoises. The morphological characters involved in the construction of the skull will form the main subject of this paper. ZARHACHIS FLAGELLATOR Cope Specimen. Cat. No , division of vertebrate paleontology, United States National Museum. The brain case and the proximal portion of the rostrum arc represented; the lachyrmals, jugals, periotics. tympanies, and bones of the inner ear are missing. Fragments of three ribs were found near this skull. Locality. The occurrence of this specimen is as follows: Near latitude 38 38' 45" N., and longitude 76 32' W., on the western shore of Chesapeake Bay, approximately 3% miles south of Chesapeake Beach, Calvert County, Mel. Shown on Patuxent quadrangle or Patuxent folio. No. 152, United States Geological Survey.



3 art. 28 SKULL OF ZARHACHIS FLAGELLATOR KELLOGG 3 Horizon. The specimen was discovered and excavated by Norman H. Boss on December 30, The jagged extremity of the rostrum was found protruding from a layer of greenish sandy clay about 8 feet above the beach at a point less than 50 feet from the end of the cliff north of the Boy Scout camp. Shattuck's zones 6-8 are not clearly differentiated at this point and the specimen came from one of these three zones. SKULL Dorsal view. As mentioned in the preceding paper, 2 the arrangement of the bones which comprise the dorsal surface of the brain case corresponds in a general way with the skull of the Chinese porpoise, Lipotes vexillifer, now living in Tung Ting lake and its tributaries. There are a number of rather obvious differences, as, for instance, the development of a high crest on the extremity of each supraorbital process, the projection of the ascending processes of the premaxillae behind the nasals, the presence of a pair of crescentic orifices for nerves on the posterior walls of the "blow holes," the outward curvature of the vomerine trough to form a portion of the dorso-anterior wall of each nasal passage, and the prolongation of the anterior extremity of the zygomatic process and the postorbital projection of the supraorbital process so that they overlap. In its general features, the skull of Zarhachis is characterized chiefly by the exceedingly long and slender rostrum which comprises more than five-sixths of the total length of the skull, the "upended" supraorbital processes of the frontals, and the pair of crescentic orifices on the posterior walls of the nasal passages below the relatively thick nasals, peculiarities that are most noticeable when contrasted with skulls of other known porpoises. Since the rostrum was fully described in the previous paper, there is no necessity for a detailed discussion of this part of the skull. All of the brain case posterior to the nasal passages and the "upended" supraorbital processes was missing from the first skull (Cat. No , U.S.N.M.). A small fragment of bone which appeared to be a portion of the vertex was found alongside of this skull and it was so placed in the restoration 3 of the brain case. The brain case of the second skull (pi. 1) is in a fair state of preservation and the description that follows has been prepared in the form of a supplement to the preceding paper. Behind the antorbital notches, the maxillae push back over the frontals and expand into relatively thin horizontal plates. The outer border of each maxilla is deflected obliquely upward and inward to conform with the curvature of the "upended" crestlike portion of the» Kellogg, R., A fossil porpoise from the Calvert formation of Maryland. Vroc. U. S Nat. Mus., vol. 03, publ. 2482, pp. 1-39, pis March 26, 1924.» Kellogg, R., Idem, pi. 1.


4 4 PROCEEDINGS OF THE NATIONAL MUSEUM vol. 67 supraorbital process. The maxilla sheathes the internal face of this "upended" process and its outer margin follows the crest of the latter. Behind this process, the maxilla expands horizontally and is applied to the platelike lateral extension of the frontal which in turn contributes the roof for the temporal fossa. The temporal fossae are not visible from a dorsal view and the external margin of the maxilla curves backward from the postorbital projection to the transverse crest of the supraoccipital. At the level of the nasals the whole maxilla curves strongly upward to the transverse crest of the supraoccipital. The dorsal margin of the supraoccipital and the posterior borders of the maxillae together comprise the transverse lambdoid crest. Behind the temporal fossae the lateral margins of the supraoccipital are applied to the posterior borders of the parietals and these combined bones form the thin posteriorly directed lambdoid crest which follows the curvature of the inner and lower margins of each temporal fossa. As in the other skull (Cat, No , U.S.N.M.) the raised convex portions of the premaxillae are widest in front of the premaxillary foramina and taper rapidly posterior to the latter, disappearing in front of the nasal passages. Both premaxillary foramina are compressed dorso-ventrally and of large size; the greatest diameter of the right foramen is 10.5 mm. The right foramen is situated 42 mm. in front of the level of the antorbital notches and the left 37 mm. From each foramen a broad groove passes forward along the internal border of the raised convex outer portion of the premaxilla. On the other skull a groove also extends backward from each foramen in an oblique direction across the premaxilla and terminates near the level of the posterior margin of the "upended" crestlike portion of the supraorbital process. On the second skull the orifice of the premaxillary foramen looks inward and forward. This change in direction of the foramen leaves the posterior margin elevated above the level of the anterior groove and shuts off the posterior groove at The premaxillae are separated by an interval of 17 mm. the source. above the porous pluglike portion of the presphenoid. The presphenoid does not rise to the level of the premaxillae, as in the first skull. Behind the level of the antorbital notches the dorsal aspect of the ascending process of each premaxilla is convexo-concave, the surface sloping from the external to the internal margin. Between the nasal passages and the antorbital notches the internal border of each premaxilla is deflected obliquely downward. The peculiarities of the premaxillae are essentially the same on both skulls. At the level of the anterior margins of the nasals the premaxillae are quite deep, and at the postero-external angle the dorsal surface is raised at least 20 mm. above the horizontally expanded plate of the maxilla. The posterior end of each premaxilla is obliquely truncated,



5 art. 28 SKULL OK ZARHACHIS FLAGELLATOR KELLOGG 5 and their posterior margins form a continuous curve with the posterior border of the protuberance behind the nasal passages. There is a relatively large dorso-ventrally compressed foramen in the maxilla at the posterior extremity of the prcmaxilla which opens into a short, deep groove. In the preceding paper it was stated that "skulls of Lipotes and Inia. may appear more specialized than that of Zarliachis 4 because of the elevation of the vertex and the shifting of the nasals to a vertical position." This opinion was based upon a small fragment of bone which appeared to represent a portion of the vertex. Further on 5 it was said that "this fragment is very important, for it shows that the vertex of the skull was not strongly elevated, or at least no prominent protuberance, like in Inia or Lipotes, was present." On reexamination it has been found that this fragment differs in a number of respects from the vertex of the second skull. The protuberance behind the nasal passages may have changed with age and varied according to sex. Nevertheless certain peculiarities of this fragment are not readily interpreted on the basis of conditions in the second skull, and there are some features which suggest the interpretation given in the preceding paper. This second skull, however, shows that other portions of the attempted restoration are incorrect and also that the protuberance behind the nasal passages is broader than in Inia and Lipotes. The nasal bones do not overhang the nasal passages; their anteroposterior diameter is about equivalent to one-half of their breadth. The nasals are applied to the anterior surfaces of the frontals, as in Lipotes and Inia, but they are proportionately larger bones and actually increase the size of the protuberance behind the nasal passages. From a dorsal view the nasals are almost subtriangular in outline, but the posterior margin is more or less emarginate and the anterior concave. Inferiorly they are overspread by the extremities of the ectethmoids. In the case of Inia and Lipotes the nasals consist of thin plates of bone applied to the anterior surface of the protuberance behind the nasal passages, and their dorsal margins do not extend upward to the level of the dorsal surfaces of the frontals. A narrow interparietal may be present between the crest of the supraoccipital and the protuberance behind the nasal passages. A narrow groove follows the posterior border of this protuberance, and may possibly represent a suture between the combined frontals and the narrow strip of bone which, because of its position, may very well represent the interparietal. An interparietal seems to be present in the skull of Inia geofrensis (Cat. No , U.SN.M.). Both condyles are visible when the skull is viewed from above. ' Kellogg, R., Proc. U. S. Nat. Mus., vol. 63, 1924, p. 7. * Kellogg, R., Idem, p. 13.


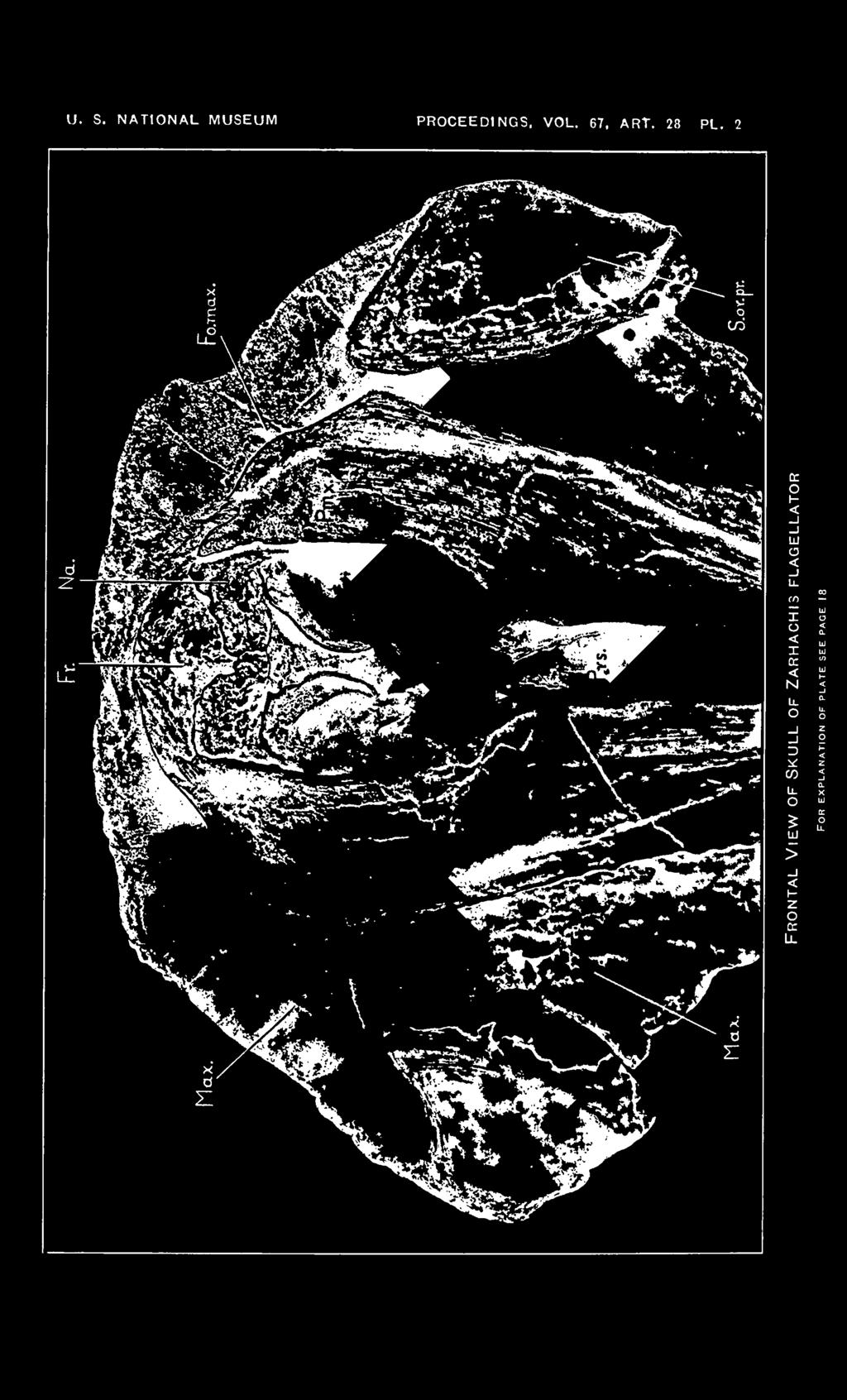
6 6 PROCEEDINGS OF THE NATIONAL MUSEUM vol. 67 The exposure of the frontals on the vertex of the skull appears to be restricted to the protuberance behind the nasal passages. Posterially the frontals abut against a narrow strip of bone which has been referred to as the "interparietal" and anteriorly they are sheathed by the nasals. The protuberance separates the posterior extremities of the maxillae on the vertex of the skull. Laterally the frontals are overspread by the ascending processes of the premaxillae and by the horizontally expanded plates of the maxillae. At a lower level, a thin plate of the frontal projects laterally, forming the roof of the temporal fossa and in front of this each frontal sends out a large supraorbital process which contributes a complete osseous roof for the orbit. The construction of the cresentic foramina below the nasal bones in the skull of Zarhachis (pi. 2) proved very puzzling on first examination. In order to obtain more explicit data on the relations of the bones involved, it seemed advisable to make a comparative examination of the skulls of as many genera of living toothed whales as were available for study. In the course of this review skulls were found in various stages of growth, and these have cleared up whatever uncertainties may have existed in regard to the construction of this portion of the brain case. The interpretations herewith given for the bones inclosing the nasal passages have been confirmed by direct comparison with disarticulated skulls of living dolphins. In a skull of a young Berardius bairdi (Cat. No , division of mammals, U.S.N.M.) from Bering Island the presphenoid rests in the trough of the vomer and the thin lateral walls of the vomer embrace the lower half of this bone. Dorsally, there is a second platelike bone coextensive with the nasal passage which fits into a groove on the upper margin of the lateral wall of the vomer and sheathes the upper half of the presphenoid. One of these platelike bones is present in each nasal passage. These bones fulfill the requirements of the ectethmoids. Posteriorly, each ectethmoid meets the corresponding frontal edge to edge on the outer border of the frontal fontanelle. In older individuals the dorsal margins of the anteriorly directed ectethmoids meet on the mid line and fuse with a perpendicular plate of bone which extends forward from the base of the nasals to a point in front of the nasal passages, thus completely inclosing the presphenoid. The thin longitudinal perpendicular plate either represents the mesethmoid or a dorsal prolongation of the combined ectethmoids. With age, these lateral ectethmoids sheath the internal borders of the frontals and extend upward on the posterior walls of the nasal passages until they meet the anterior margins of the nasal bones from below. Minute foramina are sometimes present in these bones in some of the genera of living porpoises.



7 abt. 28 SKULL OF ZAK 1 1 AC HIS FLAGELLATOR KKLI-Odii ( In most mammals the ethmoid plate ossifies into a median mesethmoid bone bounded below on either side by an ectethmoid. These ectethmoids develop as the cribriform plate. On each side of the perpendicular mesethmoid in skulls of the Miocene porpoises Diochotichus, Ceterhinops, and Squalodon there is a relatively large aperture through which the nasal branch of the ophthalmic division of the fifth cranial nerve passes. In skulls of living porpoises, like Delphinus and Pseudorca, there is a continuous sheet of bone extending upward to the base of the nasals and from which arises the mesial longitudinal perpendicular strip of bone that constitutes the most dorsal portion of the wall between the nasal passages. In each nasal passage in the skull of Lipotes a fissure appears to separate the longitudinal perpendicular plate or mesethmoid superiorly from the laterally placed ectethmoid. Inferiorly, the fissure extends obliquely downward across the posterior wall of each nasal passage. These fissures may represent one of the later stages in the closure of foramina similar to those in the Zarhachis skull. The thin longitudinal bony partition may represent either a dorsal continuation of the combined ectethmoids or the mesethmoid. Most writers in recent years have held that the pluglike porous bone, which rests in the trough of the vomer and terminates the mesorostral gutter, consists of the presphenoid below and the mesethmoid above, the bones being so intimately fused with each other that their limits can not be defined with any degree of accuracy. Such an interpretation of the mesorostral plug would place the mesethmoid below the cribiform plate in most porpoises and in others a portion would actually lie behind it. The flat platelike bone which sheathes the anterior surface of the internal borders of the frontals, conceals the frontal fontanelle, and extends upward to meet the anterior margins of the nasals from below, unquestionably represents the combined ectethmoids in the living porpoises or the cribiform plate of other mammals. The telescoping of the rostral and facial portions of the skull was accompanied by a forward thrust of the presphenoid and a backward thrust of the trough of the vomer, and as a result of one or the other, or possibly both, of the above movements, the lower portions of the ectethmoids were separated on the mid line and overspread the presphenoid laterally. Other bones may be included in these lateral plates and, in the case of Ziphius cavirostris Kernan G refers to them as including the sphenoidal turbinals. A skull of a young Grampus griseus (Cat. No. 15, 773, division of mammals, U.S.N. M.) from Cape Cod, Mass., shows how the Zarhachis type of orifice may have developed. On this young Kernan, John D., The skull of Ziphius cavirostris. Bull. Amer. Mus. Nat. Hist., vol. 38, art. 11, p August 1, 1918.


8 8 PROCEEDINGS OF THE NATIONAL MUSEUM vol. 67 Grampus skull, the dorsal margins of the lateral platelike bones or octethmoids do not meet above the presphenoid as in an older individual and above the level of the latter there is a fissure which extends inward from the internal margin of each ectethmoid to the rudimentary foramen for the nerve. In addition to this fissure, there is a shallow groove leading upward from the foramen and passing obliquely across the upper extremity of the ectethmoid. The ectethmoid has enveloped the foramen. In other mammals, the nasal branch of the ophthalmic division of the fifth cranial nerve > asses outward on each side of the mesethmoid between the latter I and the ectethmoid. A critical examination of the Zarhachis skull (pi. 2) shows that the ectethmoid in the region of the crescentic foramen is more noticeably modified than is the case in the skull of the young Grampus. As in the latter, a fissure extends inward from the internal margin of the ectethmoid to the foramen, but the whole bone is more or less involuted. Each ectethmoid in the region of these crescentic orifices is twisted almost at right angles to the main body of the bone. The curvature of the ectethmoid around the foramen is so unusual in appearance that one might view the semiovoidal rostral border or the margin which is visible on plate 2 as being actually the outer margin of the ectethmoid, and the that border which surrounds the foramen at the bottom or at the caudal end of the deep crescentic aperture is the internal margin. If this is the true explanation, then the flattened condition of the ectethmoid in living porpoises is a later development and is indicative of one of the methods by which the closure of the foramen has been effected, with the accompanying loss of the sense of smell. It should also be noted that the thin perpendicular plate extending from the nasals to a point in front of the nasal passages has been destroyed. The dorsal border of the anterior wall of each nasal passage is very thin and projects dorsally for 15 mm. or more above the porous pluglike portion of the presphenoid. Although a number of genera of toothed whales were studied, no skulls were found in which the construction of the nasal passages corresponded to the Zarhachis type of architecture. In Phocaena (Cat. No. 3659), Tursiops (Cat. No ), and Lipotes (Cat. No ), the dorsal margin of the anterior wall of each nasal passage is formed by a portion of the internal border of the maxilla. In all the toothed whales examined, the maxilla contributes part of the anterior wall of the nasal passage and, with the exception of Inia and Stenodelphis, it always constitutes the upper border of the anterior wall. In Inia and Stenodelphis, however, the premaxillae are thickened dorso-ventrally and closely approximated at the base of the mesorostral gutter; they constitute the upper border of the nasal passages. In Zarhachis, however, at


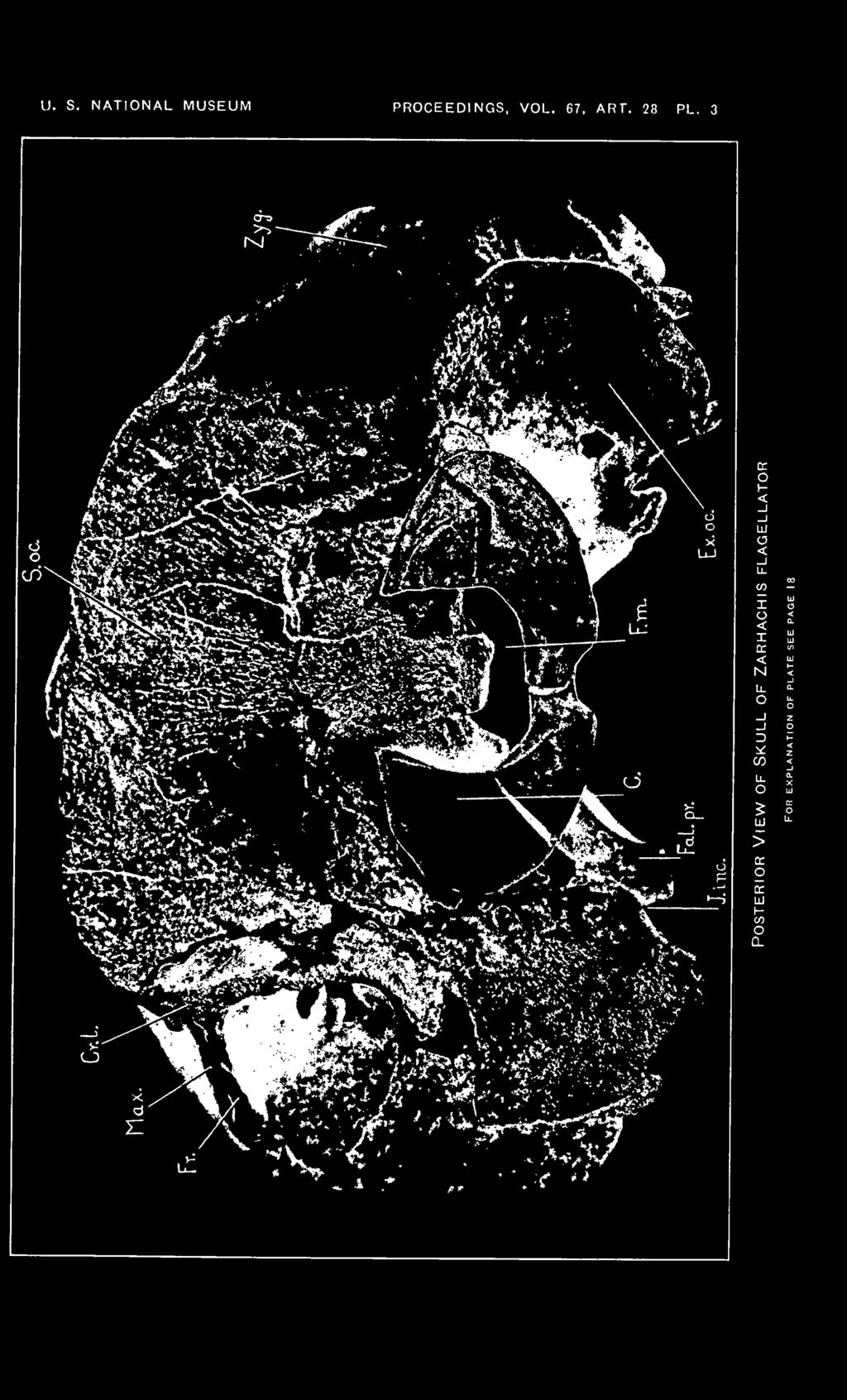
9 abt. 28 SKULL OF ZARHACHIS FLAGELLATOR KELLOGG 9 least half of the upper border of the anterior wall of each nasal passage is contributed by the vomer whose upper margin rolls over or is folded outward so that it meets the maxilla slightly external to the mid line of the nasal passage. The peculiar curvature of the lateral walls of the trough of the vomer around the upper borders of the nasal passages does not appear to be duplicated in any of the living toothed whales. The maxilla contributes the outer wall and about one-half of the dorsal border of the anterior wall. Inferiorly, the palatine extends upward in each nasal passage for a distance equivalent to about three-fourths of the depth of the anterior wall and is inserted between the maxilla and vomer in the shape of a subtriangular wedge. The dorsal margin of the surface of the palatine which takes part in the formation of the nasal passage is emarginate. In front of the nasal passages, the trough of the vomer assumes the shape characteristic of most dolphins. At the level of the anterior margin of the nasal passages, the greatest depth of the trough of the vomer is 98 mm., but at a point 180 mm. in front of these passages, it does not measure more than 25 mm. in depth. For a distance of 75 mm. in front of the nasal passages, the dorsal margins of the lateral walls of the vomer slope forward and downward at a very steep angle. This may be associated with the mesial depression of the internal borders of the premaxillae behind and in front of the premaxillary foramina. Anterior to this depression, the reduction of the lateral walls of the vomer is rather gradual. From a ventral view, the vomer is seen to be split along the mid line of the axial ridge, but this may be abnormal. The axial ridge is wedged in between the opposing faces of the maxillae and in correlation with this compression and with the slope of the internal surface of the maxilla, the floor of the mesorostral gutter is rather narrow posteriorly and it becomes so narrow about 140 mm. in front of the nasal passages that it would appear V_snaped in cross section. The lateral border of the trough of the vomer rolls over and is overspread by the thin margin of the horizontal plate of the premaxilla, which overhangs the mesorostral gutter. Posterior view. The back of the brain case (pi. 3) has been crushed in a dorso-ventral direction and the extent of this crushing is best illustrated on the posterior face of the skull. The lower half of the supraoccipital was too thin to support the great weight of the sediments which overlay the skull, and the bone fractured in many directions. A portion of the supraoccipital was thrust downward into the foramen magnum and it also buckled inward above the condyles. The displacement or amount of crushing at the center of the foramen magnum may equal 20 or 25 mm. Another consequence of this crushing is shown by the buckling of the parietals and frontals in the temporal fossae f 2



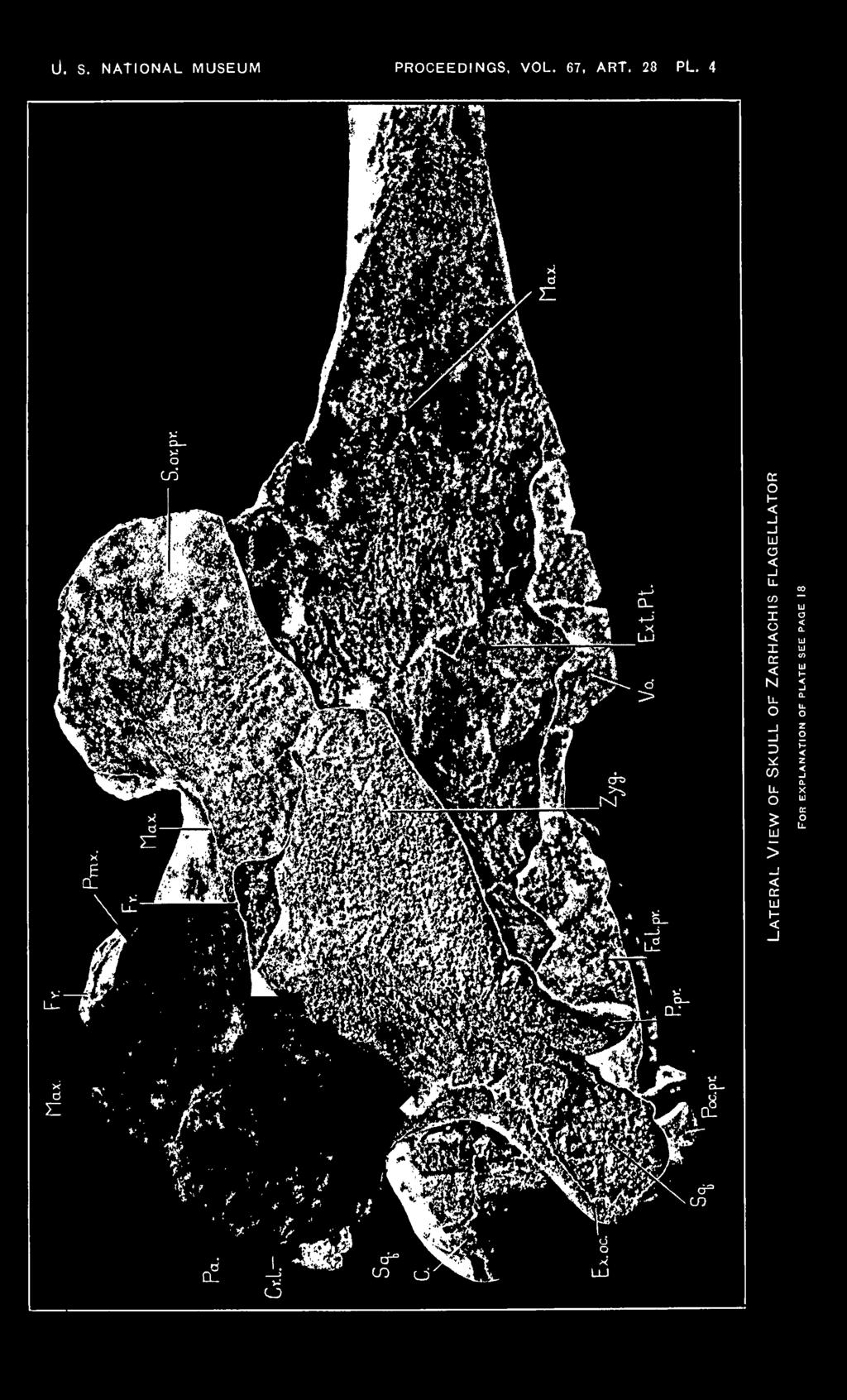
10 10 PROCEEDINGS OF THE NATIONAL MUSEUM vol.67 The posterior face of the Zarhachis skull resembles Lipotes more closely than Inia. The supraoccipital is wider than high, bounded on the sides and at the top by a flaring lambdoid crest. In Lipotes, the supraoccipital is traversed by a distinct median ridge, extending from the lambdoid crest almost to the foramen magnum. A low median crest is present on the upper half of the supraoccipital in Zarhachis. The unusual depth of the thin lambdoid crest is the characteristic peculiarity of the back of the skull. On the inner and lower sides of the temporal fossa the crest curves ventrally and laterally, following the contour of the posterior border of the fossa. At the level of the lower border of the temporal fossa, the crest on the right side is fully 32 mm. deep. The thin lambdoid crest overhangs the exoccipital on each side. The exoccipitals project downward and backward, and are of a different shape than those of Lipotes and Inia, the most noticeable modification in the latter genera being associated with the development of a paroccipital process. In these living river dolphins, the exoccipital is distinctly constricted above the paroccipital process. This process projects outward and backward. In Zarhachis, the exoccipitals are larger, although they are not produced outward far enough to conceal the zygomatic processes from behind. The more detailed features of the exoccipitals are shown on the photograph reproduced on Plate 3. Laterally and anteriorly, the exoccipital is in contact with the squamosal, superiorly it is coalesced with the supraoccipital, while below and internally it fuses with the basioccipital. The deep jugular incisure may mark the junction of the exoccipital with the falcate process of the basioccipital. The foramen magnum originally was probably slightly wider than high. The condyles are semielliptical in outline. They are strongly convex from side to side and are borne on distinct necks. The internal borders of the condyles are sharp edged and concave; the external borders are rounded off. Lateral view. Aside from the relatively large size of the " upended " extremity of the supraorbital process and the massive zygomatic process, the skull (pi. 4) is characterized by a short temporal fossa and a protuberance on the vertex. The protuberance behind the nasal passages is the highest point on the dorsal profile. The transverse crest of the supraoccipital is the next highest point and from this crest the maxilla slopes forward to the base of the rostrum. Above the orbit the extremity of the supraorbital process bends abruptly upward and forms a broad crest. Portions of the outer margin of the maxilla are visible on the upper margin of this crest from a side view and it also sheathes the entire internal surface. The greatest vertical depth of the right supraorbital process is 64 mm. and the, greatest anteroposterior diameter is 96 mm. The external face of the extremity of the supraorbital process is more or


11 art. 28 SKULL OF ZARHACHIS FLAGELLATOR KELLOGG 11 loss flattened and slopes obliquely upward and inward. There is no distinct preorbital apophysis, but the postorbital projection is rather long and attenuate. The orbit is relatively longer than in Lipotes. The lachrymal and jugal are missing. As on the first skull, the zygomatic process of the squamosal is greatly thickened dorso-ventrally in contrast to the long attenuate process of Inia and Lipotes. As viewed from the side, the slope of the posterior half of the dorsal profile of the zygomatic process is very steep ; the inferior profile is biconcave. The anterior extremity is bluntly pointed; the postglenoid process is relatively thin and curves forward. The greatest length of the right zygomatic process along the glenoid face is 98 mm. and the greatest depth near the middle is 57.5 mm. Attention has already been called to the distortion which has resulted from crushing. In addition to the previously mentioned details, it may be noted that the lateral platelike extension of the frontal and the superimposed maxilla have been depressed below their original level. A thin layer of matrix lies between the upper surface of the anterior half of the zygomatic process and the bones mentioned above. The temporal fossa is relatively short and produced backward beyond the level of the main body of the supraoccipital. Within the fossa the parietals and frontals have buckled as mentioned before, producing some irregularities. As in Inia and Lipotes, the parietal is more or less crescentic in shape; it curves around the squamosal on the external wall of the braincase and contributes the posterior border of the temporal fossa and the external plate of the bipartite lambdoid crest, extending downward behind the squamosal until it meets the exoccipital on the lower margin of the temporal fossa. Anteriorly and superiorly, the parietal is suturally united with the frontal and inferiorly it abuts against the alisphenoid. The lateral extremity of the exoccipital is relatively thin and is directed backward. The condyles are large, knoblike in contour, and project beyond the level of the exoccipital. Most of the lower border as well as the anterior end of the external reduplication of the pterygoid is missing, exposing the axial ridge of the vomer. The falcate process of the basioccipital does not project below the postglenoid process. Ventral view. The description of the rostrum in the preceding paper fully covered this part of the skull and it seems unnecessary to repeat some of the peculiarities in the present paper, especially as no additional structural details are shown. It will also be noted that the palatal surface of this skull (pi. 5) is not as well preserved as on the first skull. Some additional details are revealed which necessitate a modification of the interpretation given for the relations of the pterygoids on the first skull.


12 12 PROCEEDINGS OF THE NATIONAL MUSEUM vol. G7 It was stated in the preceding paper that the outer plate of the pterygoid represented the external pterygoid and the internal plate was referred to as the internal pterygoid. This interpretation does not appear to be correct, since both plates are continuous on the right side of the second skull and overspread the inferior face of the basal portion of the alisphenoid; these combined plates and the thin isthmus which unites them form the walls of the longitudinal sinus external to the basisphenoid. In view of the above, it is apparent that the outer plate represents the external reduplication of the pterygoid. The loss of the anterior end of the thin platelike external reduplication of the pterygoid on the right side exposes the pyramidal cavity on the corresponding side of the axial ridge of the vomer for its entire length. On the left side, this thin platelike external reduplication of the pterygoid extends forward 128 mm. in advance of the posterior wall of the nasal passage and contributes the external wall of the pyramidal cavity on the left side of the vomer. Posteriorly, the external reduplication of the left pterygoid extends backward beyond the nasal passages and is bounded by the maxilla anteriorly, by the frontal and alisphenoid superiorly, and by the squamosal posteriorly. The mandibular branch of the trigeminal nerve passes outward through the foramen ovale in the suture between the squamosal and the external reduplication of the pterygoid and follows the trowellike gutter on the external face of the latter. This foramen is situated in approximately the same position as in Platanista. There is a remarkable resemblance between the size and relations of these pterygoids and the corresponding bones in Platanista. The internal plates of the pterygoids are not as well preserved on this specimen as on the first skull. On the latter the thin internal plate of the pterygoid curves around the lower border of the lateral and anterior walls of the corresponding nasal passage and meets the vomer mesially; it then turns almost at right angles to the latter and is closely appressed to the external surface of the trough of the vomer. On skulls of Inia, Lipotes, and Platanista thin plates of the maxillae have overspread the external surface of the trough of the vomer and conceal all but the axial ridge of the latter. On the Zarhachis skull the external surface of the trough of the vomer is not overspread by the maxillae below the level of the palatines. Further comparison with the first skull is scarcely necessary, as it is now apparent that the thin anteriorly directed internal plate of the pterygoid did not by itself form the internal and superior walls of the pyramidal cavity or at least a cavity was not formed which was bounded solely by the internal plate of the pterygoid and its external reduplication as in Platanista. The palatines are not visible from a ventral view when the pterygoids are complete. The sutures defining the limits of the right



13 art. 28 SKULL OP ZARHACHIS FLAGELLATOR KELLOGG 13 palatine are clearly outlined. It is a relatively narrow bone mortised into the maxilla above and bounded externally by the upper margin of the external reduplication of the pterygoid. As now understood, each pyramidal cavity is inclosed on the rear by the internal plate of the pterygoid, on the outside by the external reduplication of the pterygoid, on the inside by a thin internal plate of the pterygoid posteriorly and by the vomer anteriorly, and above by the palatine posteriorly and the maxilla anteriorly. The thin internal plate of the pterygoid does not project as far forward as in Platanista, but otherwise the relations between the palatines and pterygoids are essentially the same in both genera. The external reduplication of the pterygoid is not developed on skulls of Inia and Lipotes, thus exposing the entire alisphenoid. On the ZarhacMs skull the extremity of the alisphenoid is not overspread by the external reduplication of the pter\ T goid and it appears within the temporal fossa in the same relative position as in Inia. In Platanista the extremity of tho alisphenoid is overspread by the external reduplication of the pterygoid. The keel of the trough of the vomer is exposed between the nasal passages and the anterior extremity of the external reduplication of the pterygoid. In its shape and relations with the surrounding bones, the vomer does not differ from that on the first skull. It expands horizontally posterior to the nasal passages, but terminates near the anterior margin of the basisphenoid. The median region of the basicranium is rather broad, bounded on each side, as mentioned above, by a continuous wall formed by the vaginal plate of the pterygoid and the adjoining falcate process of the basioccipital. The surface of the median area between these sloping walls is more or less concave. The basioccipital is a much larger bone than the basisphenoid; the transverse suture between these bones is unusually distinct. The occipital condyles are large and are separated mesially by a deep groove. A deep jugular incisure appears between the internal margin of the exoccipital and the posterior margin of the falcate process of the basioccipital. A small condylar foramen is present on each side between the base of the falcate process of the basioccipital and the condyle. The body of the squamosal contributes a part of the lateral wall of the brain case and its lateral projection or zygomatic process serves as the articular surface for the condyle of the lower jaw. The glenoid articular surface on the zygomatic process curves upward and forward and is concave from side to side. Tho external border of the zygomatic process is prolonged downward, forming a thin crest; the internal border is rounded off. The postglenoid process is short and curves forward. A shallow groove for the external auditory meatus traverses the squamosal behind the postglenoid process. A narrow posteriorly directed process of the squamosal is suturally


14 14 PROCEEDINGS OF THE NATIONAL MUSEUM vol. 6T united with the exoccipital along its external border. To the inside of the postglenoid process and at the origin of the groove for the external auditory meatus, the periotic was attached to the skull. thin plate-like process of the squamosal which is directed inward and downward contributes the external wall of the tympano-periotic fossa; the internal wall is formed by the falcate process of the basioccipital. Between the glenoid process and the internal margin of the glenoid fossa there is a sharply defined longitudinal depression, which curves forward from the base of the groove for the external auditory meatus to the anterior margin of the squamosal. The contour of the ventral surface of the paroccipital process resembles that of Inia and Platanista. There are some minor modifications which are not developed on the paroccipital processes in the last-mentioned genera and of these the thin crest on the anterior border is the most conspicuous. Further comparisons with skulls of the living Iniidae and with Platanista show some interesting structural modifications in the region of the orbitosphenoid. In Inia, the sphenoidal fissure is relatively large, bell-shaped in contour, and bounded laterally and on the rear by the basisphenoid, and anteriorly by the orbitosphenoid. On their outward course, the trochlearis and the ophthalmic division of the trigeminal nerve follow the groove on the ventral surface of an attenuate process of the orbitosphenoid. This groove is rather deep, with well-marked lateral walls; the process as a whole is directed obliquely outward and forward, terminating near the base of the broad channel on the supraorbital process of the frontal. The most remarkable peculiarity of the orbitosphenoid of Inia is the position of the ectal orifice of the optic canal. This orifice appears on the inner wall and about halfway between the anterior and posterior ends of the groove leading forward from the sphenoidal fissure. From this point forward all three nerves occupy the same groove. Within the cranial cavity, the orifice of each optic canal is placed near the inner angle of the sphenoidal fissure and is overhung by a thin platelike process of the orbitosphenoid. The course of the optic canal through the orbitosphenoid parallels the groove leading forward from the sphenoidal fissure. The apex of the process of the orbitosphenoid which contributes the surface of the groove for these nerves is jagged, and projects outward and forward at a lower level than the ventral surface of the supraorbital process. In Lipotes, the sphenoidal fissure is narrower, bounded on the rear and on the outside by the alisphenoid, and on the inside and in front by the orbitosphenoid. The orbitosphenoid of Lipotes has the same general shape as that of Inia and is likewise characterized by a narrow attenuate process which is grooved for the passage of nerves. The course of the optic nerve is slightly modified in comparison with Inia. This nerve does not pierce the orbitosphenoid, A


15 art. 28 SKULL OF ZARHACHIS FLAGELLATOR KELLOGG 15 but follows the groove on the ventral face of the attenuate process of the orbitosphenoid in company with the trochlearis and the ophthalmic division of the trigeminal. This groove is much wider than in Inia, and that portion of the orbitosphenoid which separated the optic canal from the above-mentioned groove has disappeared on the skull of Lipotes. The observations of Anderson on the peculiarities of the orbitosphenoid in the skull of Platanista have been found to agree with specimens in the United States National Museum. This description 7 can be consulted for a more detailed discussion. In Platanista, the sphenoidal fissure is almost closed and the orbitosphenoid is pierced by an extremely small foramen for the optic nerve. The orbitosphenoid has been depressed and thrust backward so that its posterior border slides backward above the anterior margin of the alisphenoid. The ventral surface of the orbitosphenoid is broadly and deeply grooved for the trochlearis and the ophthalmic division of the trigeminal nerve which pass out through the sphenoidal fissure and continue forward along this canallike continuation of the fissure. As viewed through the foramen magnum the cranial orifice of each optic canal is overhung by a thin platelike process of the orbitosphenoid. The minute ectal orifice of the orbitosphenoid portion of the optic canal appears on the inner wall of the canallike continuation of the sphenoidal fissure near the level of the anterior border of the alisphenoid. The upper border of the external reduplication of the pterygoid and a thin platelike process of the frontal which slides forward under the maxilla, as viewed from in front, enclose the passage for the optic nerve from below as it curves forward and outward; the ectal orifice of the optic canal appears near the internal extremity of the lachrymal. The optic canal, as it passes outward along the supraorbital process, is bounded on three sides by the thin plates of the frontal and roofed over by the maxilla. This thin plate like anteriorly directed process of the frontal also conceals the basal half of the supraorbital process from a ventral view. The canallike continuation of the sphenoidal fissure for the passage of nerves is concealed by the pterygoid. Imperfections in the ossification of the walls of the optic canal occur in most skulls of Platanista. In Zarhachis, the course of the optic nerve from the point where it issues from the cranial cavity to the base of the supraorbital process is concealed by the external reduplication of the pterygoid. The edges of the bones surrounding the ectal orifice of the optic canal are broken and jagged on the second skull and no attempt has been made to remove the matrix from this region on the first skull because of its» Anderson, J., Anatomical and Zoological Researches: Comprising an account of the Zoological Results of the Two Expeditions to Western Yunnan in 1868 and London (1878), pp


16 16 PROCEEDINGS OF THE NATIONAL MUSEUM vol.67 fragile condition. Hence one can not be certain what modifications were present around the ectal orifice of the optic nerve near the base of the supraorbital process of the frontal. Both frontals appear to have developed thin plates. It is possible that they may have curved below the optic nerve in the same manner as in Platanista. This point can not be settled on the basis of these two skulls alone. In the preceding paper 8 attention was called to the presence of a peculiar shelf in the temporal fossa above the upper margin of the squamosal. It is now apparent that this shelf was the result of crushing and the consequent buckling of the bones involved and that it is not comparable with a similar groove on the skull of Platanista. The second skull has suffered in the same way on the right side, but to a lesser degree. At the base of the falcate process of the basioccipital and near the posterior margin of the alisphenoid is the partially closed ectal orifice of the canal for the carotid artery. The mandibular branch of the fifth cranial nerve issues through a cleft on the posterior border of the alsiphenoid at a point 11 mm. external to the orifice of the carotid canal and on its forward and outward course crosses the ventral face of this bone, finally emerging in the temporal fossa through the foramen ovale. A postero-external process of the alisphenoid projects backward, meeting the squamosal along its external border, abutting against the exoccipital posteriorly, and uniting with the underlying process of the basioccipital internally. The alisphenoid and the above mentioned bones form the fossa in which periotic and tympanic bones are lodged. A small foramen pierces the alisphenoid above the anterior process of the periotic. Two large foramina appear in this fossa above and internal to the jugular incisure. The ectal orifice of the internal foramen is situated internal to the jugular incisure; it opens at the base and near the posterior margin of the falcate process of the basioccipital. This foramen probably represents the compartment for the nerves in the foramen lacerum posterius. The external foramen is partially inclosed by the exoccipital and may afford a passage for the jugular vein. The foramina within the tympano-periotic fossa have the same general arrangement as on one skull of EurJiinodelpMs bossi 9 which was also obtained from the Calvert formation. In skulls of Platanista, Inia, and Lipotes, a large fissure occupied the area corresponding to the tympano-periotic fossa on the Zarliachis skull. s Ki'llogg, R., Proc. U. S. Nat. Mus., vol. 63, pp R. Kellogg, On the occurrence of remains of fossil porpoises of the genus Eurhinodelphis in North America. Proc. U. S. Nat. Mus., vol. 66, publ. 2563, pi
 165.")


17 aht. 28 SKULL OF ZARHACHIS FLAGELLATOR KELLOGG 17 measurements FOR the skull (in millimeters) Breadth of rostrum at base (between antorbital notches) 163 Greatest breadth of skull across supraorbital processes 260 Greatest breadth of skull across zygomatic processes of squamosals 260 Vertical height of skull (between tips of falcate processes of basioccipital and frontals on vertex of skull) Vertical height of skull (basisphenoid to frontals on vertex of skull 128 Greatest depth of rostrum at level of antorbital notches 89 Greatest distance between outside margins of premaxillae at level of anterior margin of porous pluglike presphenoid 93 Greatest distance between outside margins of premaxillae at level of anterior margin of nasals 130 Greatest breadth of right premaxilla in front of nasal passages 44 Length of frontal plate of right maxilla (antorbital notch to supraoccipital) Greatest breadth of exposed surface of frontal plate of right maxilla behind supraorbital process 66 Least distance across vertex of skull between inner margins of maxillae-. 39 Greatest antero-posterior diameter of supraorbital process of right frontal. 96 Greatest dorso-ventral depth of supraorbital process of right frontal Greatest thickness of "upended" portion of supraorbital process of right frontal 40 Least breadth of braincase between temporal fossae 120 Least distance between temporal fossae as measured between lambdoid crests Distance from center of transverse crest of supraoccipital to upper margin of foramen magnum Greatest breadth of supraoccipital 151 Breadth of foramen magnum Greatest distance between outside margins of occipital condyles 105 Greatest dorso-ventral diameter of right condyle 53 Greatest transverse diameter of right condyle 42 Greatest distance across skull between outside margins of exoccipitals. _ 212 Depth of left exoccipital at extremity Distance from anterior margin of foramen magnum to right nasal passage. 122 Distance from horizontal plate of right maxilla to extremity of right paroccipital process Greatest antero-posterior diameter of zygomatic process of right squamosal 101 Distance between anterior margin of apophysis of right maxilla and posterior face of right condyle 215 Greatest breadth of basioccipital across tips of the falcate processes 101 Distance between anterior margin of foramen magnum and anterior margin of basisphenoid 86 Greatest length of right nasal 14 Greatest breadth of right nasal 26 Greatest length of exposed portion of right frontal on vertex of skull 34 Greatest breadth of exposed portion of right frontal on vertex of skull 28


18 18 PROCEEDINGS OF THE NATIONAL. MUSEUM Vol. 67, abt. 82. EXPLANATION OF PLATES Zarhachis flagellator Cope. Cat. No. 10,911, division of vertebrate paleontology, United States National Museum. Calvert formation, western shore of Chesapeake Bay, about 3% miles south of Chesapeake Beach, Calvert County, Md. Collected by Norman H. Boss, December 30, Plate 1 Dorsal view of skull of Zarhachis flagellator Cope. About Yh natural size. The following abbreviations are used on plates 1 to 5. Bo., basioccipital; Bs., basisphenoid; C, condyle; Cr. I., lambdoid crest; Cr. max., maxillary crest; Ed., ectethmoid; Ex. oc, exoccipital; Ex. pt., external reduplication of the pterygoid; Ex. aud. M., groove for external auditory meatus; Fal. pr., falcate process of basioccipital; F. m., foramen magnum; Fo. c, condylar foramen; Fo. inf., ventral orifice of infraorbital canal; Fo. I. p., foramen lacerum posterius, compartment for vein; Fo. max., maxillary foramen at posterior extremity of the premaxilla; Fo. ov., foramen ovale; Fo. pmx., premaxillary foramen; Fr., frontal; J. inc., jugular incisure; Max., maxilla; N. A., left nasal passage; Na., nasal; Pa., parietal; Pmx., premaxilla; Poc. pr., paroccipital process of exoccipital; P. pr., postglenoid process of squamosal; Prs., presphenoid; Pi., vaginal process of pterygoid; S. oc, supraoccipital; S. or. pr., supraorbital process of frontal; Sq., squamosal; Vo., vomer; Zyg., zygomatic process of squamosal; 1, foramen in alisphenoid above the anterior process of the periotic; 2, passage for mandibular branch of trigeminal nerve in a cleft on posterior border of alisphenoid; S, foramen lacerum posterius, compartment for nerves. Plate 2 Frontal view of skull of Zarhachis flagellator Cope showing position of the crescentic foramina. About % natural size. Plate 3 Posterior view of skull of Zarhachis flagellator Cope. About % natural size. Plate 4 Lateral view of skull of Zarhachis flagellator Cope. About H natural size. Plate 5 Ventral view of skull of Zarhachis flagellator Cope. About % natural size. 18



19 U. S. NATIONAL MUSEUM PROCEEDINGS, VOL. 67, ART. 28 PL. I Dorsal View of Skull of Zarhachis flagellator For explanation of plate see page 18


20 U. S. NATIONAL MUSEUM PROCEEDINGS, VOL. 67, ART. 28 PL. 2


21 U. S. NATIONAL MUSEUM PROCEEDINGS, VOL. 67, ART. 28 PL. 3 O G 5 P


22 U. s. national museum PROCEEDINGS, VOL. 67, ART. 28 PL. 4



23 U. S. NATIONAL MUSEUM PROCEEDINGS, VOL. 67, ART. 28 PL. 5 in Ventral View of Skull of Zarhachis flagellator For explanation of plate see page IJ
AMERICAN MUSEUM NOVITATES Published by
 AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE
 PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA
 A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA MYCTEROSAURUS LONGICEPS S. W. WILLISTON University of Chicago The past summer, Mr. Herman Douthitt, of the University of Chicago paleontological expedition,
A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA MYCTEROSAURUS LONGICEPS S. W. WILLISTON University of Chicago The past summer, Mr. Herman Douthitt, of the University of Chicago paleontological expedition,
Williston, and as there are many fairly good specimens in the American
 56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
WHxVLEBONE WHALE FROM THE CALVERT CLIFFS, MARYLAND.
 DESCRIPTION OF A NEW GENUS AND SPECIES OF WHxVLEBONE WHALE FROM THE CALVERT CLIFFS, MARYLAND. By Remington Kellogg, Of the Bureau of Biological Survey, United States Departm'ent of Agriculture. In the
DESCRIPTION OF A NEW GENUS AND SPECIES OF WHxVLEBONE WHALE FROM THE CALVERT CLIFFS, MARYLAND. By Remington Kellogg, Of the Bureau of Biological Survey, United States Departm'ent of Agriculture. In the
Mammalogy Laboratory 1 - Mammalian Anatomy
 Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
complex in cusp pattern. (3) The bones of the coyote skull are thinner, crests sharper and the
 The bones of the coyote skull are thinner, crests sharper and the") DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons
 Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons 1. Head skeleton of lamprey Cyclostomes are highly specialized in both the construction of the chondrocranium and visceral skeleton.
Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons 1. Head skeleton of lamprey Cyclostomes are highly specialized in both the construction of the chondrocranium and visceral skeleton.
MIOCENE DIATOMACEOUS EARTH OF LOMPOC, CALI- FORNIA.i
 DESCRIPTION OF THE SKULL OF MEGAPTERA MIO- CAENA, A FOSSIL HUMPBACK WHALE FROM THE MIOCENE DIATOMACEOUS EARTH OF LOMPOC, CALI- FORNIA.i By Remington Kellogg. Of the Bureau of Biological Survey, Department
DESCRIPTION OF THE SKULL OF MEGAPTERA MIO- CAENA, A FOSSIL HUMPBACK WHALE FROM THE MIOCENE DIATOMACEOUS EARTH OF LOMPOC, CALI- FORNIA.i By Remington Kellogg. Of the Bureau of Biological Survey, Department
ONLINE APPENDIX 1. Morphological phylogenetic characters scored in this paper. See Poe (2004) for
 for") ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC
 FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the
 Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the") Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
v:ii-ixi, 'i':;iisimvi'\>!i-:: "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi L I E) R.ARY OF THE VERSITY U N I or ILLINOIS REMO
 R.ARY OF THE VERSITY U N I or ILLINOIS REMO") "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
"^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province
 from Dashanpu, Zigong Municipality, Sichuan Province") A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
2. Skull, total length versus length of the presacral vertebral column: (0); extremely elongated neck (e.g. Tanystropheus longobardicus).
; extremely elongated neck (e.g. Tanystropheus longobardicus).") Character list of the taxon-character data set 1. Skull and lower jaws, interdental plates: absent (0); present, but restricted to the anterior end of the dentary (1); present along the entire alveolar
Character list of the taxon-character data set 1. Skull and lower jaws, interdental plates: absent (0); present, but restricted to the anterior end of the dentary (1); present along the entire alveolar
Vol. XIV, No. 1, March, The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S.
 By S.") Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Mammalogy Lab 1: Skull, Teeth, and Terms
 Mammalogy Lab 1: Skull, Teeth, and Terms Be able to: Goals of today s lab Locate all structures listed on handout Define all terms on handout what they are or what they look like Give examples of mammals
Mammalogy Lab 1: Skull, Teeth, and Terms Be able to: Goals of today s lab Locate all structures listed on handout Define all terms on handout what they are or what they look like Give examples of mammals
A New Ceratopsian Dinosaur from the Upper
 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 63. NUMBER 3 A New Ceratopsian Dinosaur from the Upper Cretaceous of Montana, with Note on Hypacrosaurus (With Two Plates) CHARLES W. GILMORE Assistant Curator
SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 63. NUMBER 3 A New Ceratopsian Dinosaur from the Upper Cretaceous of Montana, with Note on Hypacrosaurus (With Two Plates) CHARLES W. GILMORE Assistant Curator
A NEW SPECIES OF TROODONT DINOSAUR FROM THE
 A NEW SPECIES OF TROODONT DINOSAUR FROM THE LANCE FORMATION OF WYOMING By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION The intensive search to which
A NEW SPECIES OF TROODONT DINOSAUR FROM THE LANCE FORMATION OF WYOMING By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION The intensive search to which
 The following text is generated from uncorrected OCR. [Begin Page: Page 1] A NEW CERATOPSIAN DINOSAUR FROM THE UPPER CRETACEOUS OF MONTANA, WITH NOTE ON HYPACROSAURUS ' By CHARLES W. GILMORE assistant
The following text is generated from uncorrected OCR. [Begin Page: Page 1] A NEW CERATOPSIAN DINOSAUR FROM THE UPPER CRETACEOUS OF MONTANA, WITH NOTE ON HYPACROSAURUS ' By CHARLES W. GILMORE assistant
OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE
 OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT This is a report of measurements on the skeleton of a male se1 whale caught in the Antarctic. The skeleton of
OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT This is a report of measurements on the skeleton of a male se1 whale caught in the Antarctic. The skeleton of
OF THE TRIAS THE PHYTOSAURIA
 THE PHYTOSAURIA OF THE TRIAS MAURICE G. MEHL University of Wisconsin Some time ago the writer gave a brief notice of a new genus of phytosaurs of which Angistorhinus grandis Mehl was the type.' It is the
THE PHYTOSAURIA OF THE TRIAS MAURICE G. MEHL University of Wisconsin Some time ago the writer gave a brief notice of a new genus of phytosaurs of which Angistorhinus grandis Mehl was the type.' It is the
New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia
 1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
SUPPLEMENTARY ONLINE MATERIAL FOR. Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor
 http://app.pan.pl/som/app61-ratsimbaholison_etal_som.pdf SUPPLEMENTARY ONLINE MATERIAL FOR Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor Ontogenetic changes in the craniomandibular
http://app.pan.pl/som/app61-ratsimbaholison_etal_som.pdf SUPPLEMENTARY ONLINE MATERIAL FOR Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor Ontogenetic changes in the craniomandibular
Notes on Ceratopsians and Ankylosaurs at the Royal Ontario Museum
 Notes on Ceratopsians and Ankylosaurs at the Royal Ontario Museum Andrew A. Farke, Ph.D. Raymond M. Alf Museum of Paleontology 1175 West Baseline Road Claremont, CA 91711 email: afarke@webb.org Introduction
Notes on Ceratopsians and Ankylosaurs at the Royal Ontario Museum Andrew A. Farke, Ph.D. Raymond M. Alf Museum of Paleontology 1175 West Baseline Road Claremont, CA 91711 email: afarke@webb.org Introduction
THE SKULLS OF ARAEOSCELIS AND CASEA, PERMIAN REPTILES
 THE SKULLS OF REOSCELIS ND CSE, PERMIN REPTILES University of Chicago There are few Permian reptiles of greater interest at the present time than the peculiar one I briefly described in this journal' three
THE SKULLS OF REOSCELIS ND CSE, PERMIN REPTILES University of Chicago There are few Permian reptiles of greater interest at the present time than the peculiar one I briefly described in this journal' three
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO
 A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
CENE RUMINANTS OF THE GENERA OVIBOS AND
 DESCRIPTIONS OF TWO NEW SPECIES OF PLEISTO- CENE RUMINANTS OF THE GENERA OVIBOS AND BOOTHERIUM, WITH NOTES ON THE LATTER GENUS. By James Williams Gidley, Of the United States National Museum. Two interesting
DESCRIPTIONS OF TWO NEW SPECIES OF PLEISTO- CENE RUMINANTS OF THE GENERA OVIBOS AND BOOTHERIUM, WITH NOTES ON THE LATTER GENUS. By James Williams Gidley, Of the United States National Museum. Two interesting
SUPPLEMENTARY INFORMATION
 Character 155, interdental ridges. Absence of interdental ridge (0) shown in Parasaniwa wyomingensis (Platynota). Interdental ridges (1) shown in Coniophis precedens. WWW.NATURE.COM/NATURE 1 Character
Character 155, interdental ridges. Absence of interdental ridge (0) shown in Parasaniwa wyomingensis (Platynota). Interdental ridges (1) shown in Coniophis precedens. WWW.NATURE.COM/NATURE 1 Character
CRANIAL OSTEOLOGY OF SCHIZOTHORAICHTHYS NIGER (MECKEL) MISRA (CYPRINIDAE: SCHIZOTHORACINAE). L NEUROCRANIUM
 MISRA (CYPRINIDAE: SCHIZOTHORACINAE). L NEUROCRANIUM") CRANIAL OSTEOLOGY OF SCHIZOTHORAICHTHYS NIGER (MECKEL) MISRA (CYPRINIDAE: SCHIZOTHORACINAE). L NEUROCRANIUM A. R. YousuF, A. K. PANDIT AND A. R. KHAN Postgraduate Department of Zoology, University of Kashmir,
CRANIAL OSTEOLOGY OF SCHIZOTHORAICHTHYS NIGER (MECKEL) MISRA (CYPRINIDAE: SCHIZOTHORACINAE). L NEUROCRANIUM A. R. YousuF, A. K. PANDIT AND A. R. KHAN Postgraduate Department of Zoology, University of Kashmir,
List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to
 1 Supplementary data CHARACTER LIST List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to characters used by Tchernov et al. (2000), Rieppel, et al. (2002), and Lee
1 Supplementary data CHARACTER LIST List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to characters used by Tchernov et al. (2000), Rieppel, et al. (2002), and Lee
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL
 Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
.56 m. (22 in.). COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition
. COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition") Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.)
") A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.) by Ouyang Hui Zigong Dinosaur Museum Newsletter Number 2 1989 pp. 10-14 Translated By Will Downs Bilby
A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.) by Ouyang Hui Zigong Dinosaur Museum Newsletter Number 2 1989 pp. 10-14 Translated By Will Downs Bilby
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A.
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
A skull without mandihle, from the Hunterian Collection (no.
 4 MR. G. A. BOULENGER ON CHELONIAN REMAINS. [Jan. 6, 2. On some Chelonian Remains preserved in the Museum of the Eojal College of Surgeons. By G. A. Boulenger. [Eeceived December 8, 1890.] In the course
4 MR. G. A. BOULENGER ON CHELONIAN REMAINS. [Jan. 6, 2. On some Chelonian Remains preserved in the Museum of the Eojal College of Surgeons. By G. A. Boulenger. [Eeceived December 8, 1890.] In the course
[Accepted 8th October CONTENTS INTRODUCTION
 183 THE CRANIAL MORPHOLOGY OF A NEW GENUS AND SPECIES OF ICTIDOSAURAN BY A. W. CROMPTON S. A. Museum, Cape Town [Accepted 8th October 19571 (With 7 figures in the text) CONTENTS lntroduction..............
183 THE CRANIAL MORPHOLOGY OF A NEW GENUS AND SPECIES OF ICTIDOSAURAN BY A. W. CROMPTON S. A. Museum, Cape Town [Accepted 8th October 19571 (With 7 figures in the text) CONTENTS lntroduction..............
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature13086 Part I. Supplementary Notes A: Detailed Description of Cotylocara macei gen. et sp. nov. Part II. Table of Measurements for holotype of Cotylocara macei (CCNHM-101) Part III. Supplementary
doi:10.1038/nature13086 Part I. Supplementary Notes A: Detailed Description of Cotylocara macei gen. et sp. nov. Part II. Table of Measurements for holotype of Cotylocara macei (CCNHM-101) Part III. Supplementary
SOME NEW AMERICAN PYCNODONT FISHES.
 SOME NEW AMERICAN PYCNODONT FISHES. By James Williams Gidley, Assistant Curator of Fossil Mammals, United States National Museum. In the United States National Museum are several specimens representing
SOME NEW AMERICAN PYCNODONT FISHES. By James Williams Gidley, Assistant Curator of Fossil Mammals, United States National Museum. In the United States National Museum are several specimens representing
FIRST RECORD OF MESOPLODON DENSIROSTRIS FROM FORMOSA
 FIRST RECORD OF MESOPLODON DENSIROSTRIS FROM FORMOSA TOSHIO KASUYA* AND MASAHARU NISHIWAKI* ABSTRACT Two records of female Mesoplodon densirostris are reported. Comments on the external character, skull
FIRST RECORD OF MESOPLODON DENSIROSTRIS FROM FORMOSA TOSHIO KASUYA* AND MASAHARU NISHIWAKI* ABSTRACT Two records of female Mesoplodon densirostris are reported. Comments on the external character, skull
290 SHUFELDT, Remains of Hesperornis.
 290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
 Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
Temporal lines. More forwardfacing. tubular orbits than in the African forms 3. Orbits larger relative to skull size than in the other genera 2.
 Asian lorises More forwardfacing and tubular orbits than in the African forms 3. Characterized by a marked extension of the ectotympanic into a tubular meatus and a more angular auditory bulla than in
Asian lorises More forwardfacing and tubular orbits than in the African forms 3. Characterized by a marked extension of the ectotympanic into a tubular meatus and a more angular auditory bulla than in
NORTH AMERICA. ON A NEW GENUS AND SPECIES OF COLUBRINE SNAKES FROM. The necessity of recognizing tlie two species treated of in this paper
 ON A NEW GENUS AND SPECIES OF COLUBRINE SNAKES FROM NORTH AMERICA. BY Leonhard Stejneger, and Batrachians. Curator of the Department of Reptiles The necessity of recognizing tlie two species treated of
ON A NEW GENUS AND SPECIES OF COLUBRINE SNAKES FROM NORTH AMERICA. BY Leonhard Stejneger, and Batrachians. Curator of the Department of Reptiles The necessity of recognizing tlie two species treated of
FIELDIANA GEOLOGY NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA
 FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
A NEW SALTICID SPIDER FROM VICTORIA By R. A. Dunn
 Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
Museo di Storia Naturale del Mediterraneo, Via Roma 234, 57125, Livorno, Italy
 bs_bs_banner Zoological Journal of the Linnean Society, 2012, 166, 876 911. With 22 figures Comparative osteology and phylogenetic relationships of Miocaperea pulchra, the first fossil pygmy right whale
bs_bs_banner Zoological Journal of the Linnean Society, 2012, 166, 876 911. With 22 figures Comparative osteology and phylogenetic relationships of Miocaperea pulchra, the first fossil pygmy right whale
Systematic revision of the Miocene long-snouted dolphin Eurhinodelphis longirostris DU Bus, 1872 (Cetacea, Odontoceti, Eurhinodelphinidae)
") BULLETIN DE L INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE SCIENCES DE LA TERRE, 74: 147-174, 2004 BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETENSCHAPPEN AARDWETENSCHAPPEN, 74: 147-174,
BULLETIN DE L INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE SCIENCES DE LA TERRE, 74: 147-174, 2004 BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETENSCHAPPEN AARDWETENSCHAPPEN, 74: 147-174,
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * Dr. L.D. Boonstra. Paleontologist, South African Museum, Cape Town
 THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
AN OSTEOLOGICAL STUDY OF THE CUVIER'S BEAKED WHALE,,ZIPHIUS CAVIROSTRIS, IN THE NORTHWEST PACIFIC
 AN OSTEOLOGICAL STUDY OF THE CUVIER'S BEAKED WHALE,,ZIPHIUS CAVIROSTRIS, IN THE NORTHWEST PACIFIC HIDEO OMURA ABSTRACT Skulls and postcranial skeletons of :(,iphius cavirostris were studied for eleven
AN OSTEOLOGICAL STUDY OF THE CUVIER'S BEAKED WHALE,,ZIPHIUS CAVIROSTRIS, IN THE NORTHWEST PACIFIC HIDEO OMURA ABSTRACT Skulls and postcranial skeletons of :(,iphius cavirostris were studied for eleven
TRACHEMYS SCULPTA. A nearly complete articulated carapace and plastron of an Emjdd A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE,
 A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
APPENDIX. 160 Miscellaneous Intelligence.
 160 Miscellaneous Intelligence. OBITUARY. GENERAL ANDREW A. HUMPHREYS. Brigadier-General Andrew Atkinson Humphreys died in Washington, on the 28th of November last, in the seventy-fourth year of his age.
160 Miscellaneous Intelligence. OBITUARY. GENERAL ANDREW A. HUMPHREYS. Brigadier-General Andrew Atkinson Humphreys died in Washington, on the 28th of November last, in the seventy-fourth year of his age.
ON TWO NEW SPECIMENS OF LYSTROSAURUS-ZONE CYNODONTS
 ON TWO NEW SPECMENS OF LYSTROSAURUS-ZONE CYNODONTS By A. S. Brink ABSTRACT n this paper the skulls of two new specimens of Lystrosaurus-zone cynodonts are described. One is a skull of Notictosaurus luckh1fi
ON TWO NEW SPECMENS OF LYSTROSAURUS-ZONE CYNODONTS By A. S. Brink ABSTRACT n this paper the skulls of two new specimens of Lystrosaurus-zone cynodonts are described. One is a skull of Notictosaurus luckh1fi
Anatomy. Name Section. The Vertebrate Skeleton
 Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
HONR219D Due 3/29/16 Homework VI
 Part 1: Yet More Vertebrate Anatomy!!! HONR219D Due 3/29/16 Homework VI Part 1 builds on homework V by examining the skull in even greater detail. We start with the some of the important bones (thankfully
Part 1: Yet More Vertebrate Anatomy!!! HONR219D Due 3/29/16 Homework VI Part 1 builds on homework V by examining the skull in even greater detail. We start with the some of the important bones (thankfully
Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved
 Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved This was a private report in 2003 on my thoughts on Platecarpus planifrons.
Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved This was a private report in 2003 on my thoughts on Platecarpus planifrons.
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE
 FROM THE") A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
PALEONTOLOGY AND BIOSTRATIGRAPHY OF MONGOLIA
 PALEONTOLOGY AND BIOSTRATIGRAPHY OF MONGOLIA THE JOINT SOVIET-MONGOLIAN PALEONTOLOGICAL EXPEDITION (Transactions, vol. 3) EDITORIAL BOARD: N. N. Kramarenko (editor-in-chief) B. Luvsandansan, Yu. I. Voronin,
PALEONTOLOGY AND BIOSTRATIGRAPHY OF MONGOLIA THE JOINT SOVIET-MONGOLIAN PALEONTOLOGICAL EXPEDITION (Transactions, vol. 3) EDITORIAL BOARD: N. N. Kramarenko (editor-in-chief) B. Luvsandansan, Yu. I. Voronin,
A Fossil Snake (Elaphe vulpina) From A Pliocene Ash Bed In Nebraska
 From A Pliocene Ash Bed In Nebraska") University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
PALEONTOLOGICAL CONTRIBUTIONS
 THE UNIVERSITY OF KANSAS PALEONTOLOGICAL CONTRIBUTIONS August, 1965 Paper 2 A NEW WYOMING PHYTOSAUR By THEODORE H. EATON, JR. [Museum of Natural History, University of Kansas I ABSTRACT The skull of a
THE UNIVERSITY OF KANSAS PALEONTOLOGICAL CONTRIBUTIONS August, 1965 Paper 2 A NEW WYOMING PHYTOSAUR By THEODORE H. EATON, JR. [Museum of Natural History, University of Kansas I ABSTRACT The skull of a
Mammalogy Lecture 8 - Evolution of Ear Ossicles
 Mammalogy Lecture 8 - Evolution of Ear Ossicles I. To begin, let s examine briefly the end point, that is, modern mammalian ears. Inner Ear The cochlea contains sensory cells for hearing and balance. -
Mammalogy Lecture 8 - Evolution of Ear Ossicles I. To begin, let s examine briefly the end point, that is, modern mammalian ears. Inner Ear The cochlea contains sensory cells for hearing and balance. -
Macro-anatomical studies of the African giant pouched rat (Cricetomys gambianus) axial skeleton
 axial skeleton") Standard Scientific Research and Essays Vol1 (10): 221-227, October 2013 http://www.standresjournals.org/journals/ssre Research Article Macro-anatomical studies of the African giant pouched rat (Cricetomys
Standard Scientific Research and Essays Vol1 (10): 221-227, October 2013 http://www.standresjournals.org/journals/ssre Research Article Macro-anatomical studies of the African giant pouched rat (Cricetomys
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/329/5998/1481/dc1 Supporting Online Material for Tyrannosaur Paleobiology: New Research on Ancient Exemplar Organisms Stephen L. Brusatte,* Mark A. Norell, Thomas D.
www.sciencemag.org/cgi/content/full/329/5998/1481/dc1 Supporting Online Material for Tyrannosaur Paleobiology: New Research on Ancient Exemplar Organisms Stephen L. Brusatte,* Mark A. Norell, Thomas D.
LOWER CRETACEOUS OF SOUTH DAKOTA.
 A NEW DINOSAUR, STP^GOSAURUS MARSHl, FROM THE LOWER CRETACEOUS OF SOUTH DAKOTA. By Frederic A. Lucas, Curator, Divisioii of Coiiipnrative Anatomy, in charge, of Section of Vertebrate Fossils. The name
A NEW DINOSAUR, STP^GOSAURUS MARSHl, FROM THE LOWER CRETACEOUS OF SOUTH DAKOTA. By Frederic A. Lucas, Curator, Divisioii of Coiiipnrative Anatomy, in charge, of Section of Vertebrate Fossils. The name
A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev Doklady Akademii Nauk, SSSR 87:
 translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
LEIDY, SHOWING THE BONES OF THE FEET 'AND LIMBS
 CQNTEUBUTIONS FBOM THE MUSEUM OF PALEONTOLOGY (Confindion of Con&&&m froin UB Muaercm of Gcologg) UNIVERSITY OF ' MICHIGAN VOL V, No. 6, pp. 6W3 (e ph.) DEAXMBER 31,1036 A SPECIMEN OF STYLEMYS NEBRASCENSIS
CQNTEUBUTIONS FBOM THE MUSEUM OF PALEONTOLOGY (Confindion of Con&&&m froin UB Muaercm of Gcologg) UNIVERSITY OF ' MICHIGAN VOL V, No. 6, pp. 6W3 (e ph.) DEAXMBER 31,1036 A SPECIMEN OF STYLEMYS NEBRASCENSIS
PEABODY MUSEUM OF NATURAL HISTORY, YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 85 September 21, 1964 A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA STANLEY J. RIEL
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 85 September 21, 1964 A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA STANLEY J. RIEL
THE SKULLS OF THE CATHARTID
 . 272 Vol. 46 THE SKULLS OF THE CATHARTID VULTURES By HARVEY I. FISHER The New World vultures, family Cathartidae, form a heterogeneous group of large birds which is now limited in its range to the Americas.
. 272 Vol. 46 THE SKULLS OF THE CATHARTID VULTURES By HARVEY I. FISHER The New World vultures, family Cathartidae, form a heterogeneous group of large birds which is now limited in its range to the Americas.
SECOND RECORD OF SPECTACLED PORPOISE FROM SUBANTARCTIC SOUTHWEST PACIFIC
 SECOND RECORD OF SPECTACLED PORPOISE FROM SUBANTARCTIC SOUTHWEST PACIFIC R. EWAN FORDYCE Department of Geology, University of Otago, P, 0. Box 56, Dunedin, New Zealand ROBERT H. MATTLIN Fisheries Research
SECOND RECORD OF SPECTACLED PORPOISE FROM SUBANTARCTIC SOUTHWEST PACIFIC R. EWAN FORDYCE Department of Geology, University of Otago, P, 0. Box 56, Dunedin, New Zealand ROBERT H. MATTLIN Fisheries Research
CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN OF RUSSIA AND THE EVOLUTIONARY RELATIONSHIPS OF CASEIDAE
 FROM THE MIDDLE PERMIAN OF RUSSIA AND THE EVOLUTIONARY RELATIONSHIPS OF CASEIDAE") Journal of Vertebrate Paleontology 28(1):160 180, March 2008 2008 by the Society of Vertebrate Paleontology ARTICLE CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN
Journal of Vertebrate Paleontology 28(1):160 180, March 2008 2008 by the Society of Vertebrate Paleontology ARTICLE CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN
NOTE XXXVIII. Three new species of the genus Helota DESCRIBED BY. C. Ritsema+Cz. is very. friend René Oberthür who received. Biet.
 Subshining; HELOTA MARIAE. 249 NOTE XXXVIII. Three new species of the genus Helota DESCRIBED BY C. Ritsema+Cz. The first of these species is very interesting as it belongs to the same section as the recently
Subshining; HELOTA MARIAE. 249 NOTE XXXVIII. Three new species of the genus Helota DESCRIBED BY C. Ritsema+Cz. The first of these species is very interesting as it belongs to the same section as the recently
VARIATION IN MONIEZIA EXPANSA RUDOLPHI
 VARIATION IN MONIEZIA EXPANSA RUDOLPHI STEPHEN R. WILLIAMS, Miami University, Oxford, Ohio In making a number of preparations of proglottids for class study at the stage when sex organs are mature and
VARIATION IN MONIEZIA EXPANSA RUDOLPHI STEPHEN R. WILLIAMS, Miami University, Oxford, Ohio In making a number of preparations of proglottids for class study at the stage when sex organs are mature and
Cranial osteology of the African gerrhosaurid Angolosaurus skoogi (Squamata; Gerrhosauridae) HOLLY A. NANCE
 HOLLY A. NANCE") African Journal of Herpetology, 2007 56(1): 39-75. Herpetological Association of Africa Original article Cranial osteology of the African gerrhosaurid Angolosaurus skoogi (Squamata; Gerrhosauridae) HOLLY
African Journal of Herpetology, 2007 56(1): 39-75. Herpetological Association of Africa Original article Cranial osteology of the African gerrhosaurid Angolosaurus skoogi (Squamata; Gerrhosauridae) HOLLY
DISCOVERY AND CAPTURE
 46 NSHWAK AND KASUYA Hitherto, this species has occurred no other place in the world on 33 28' N. around, San Diego and Casabranca for example. Our news was not only the evidence of this occurrence. When
46 NSHWAK AND KASUYA Hitherto, this species has occurred no other place in the world on 33 28' N. around, San Diego and Casabranca for example. Our news was not only the evidence of this occurrence. When
A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae)
") Genus Vol. 14 (3): 413-418 Wroc³aw, 15 X 2003 A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae) JAROS AW KANIA Zoological Institute, University of Wroc³aw, Sienkiewicza
Genus Vol. 14 (3): 413-418 Wroc³aw, 15 X 2003 A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae) JAROS AW KANIA Zoological Institute, University of Wroc³aw, Sienkiewicza
ZOOLOGISCHE MEDEDELINGEN
 "f ~- >D noitnwz, tito ZOOLOGISCHE MEDEDELINGEN UITGEGEVEN DOOR HET RIJKSMUSEUM VAN NATUURLIJKE HISTORIE TE LEIDEN (MINISTERIE VAN CULTUUR, RECREATIE EN MAATSCHAPPELIJK WERK) Deel 48 no. 25 25 maart 1975
"f ~- >D noitnwz, tito ZOOLOGISCHE MEDEDELINGEN UITGEGEVEN DOOR HET RIJKSMUSEUM VAN NATUURLIJKE HISTORIE TE LEIDEN (MINISTERIE VAN CULTUUR, RECREATIE EN MAATSCHAPPELIJK WERK) Deel 48 no. 25 25 maart 1975
Anatomical description and phylogenetic analysis of Miocene beaked whale from the East African Rift Valley, Kenya
 Southern Methodist University SMU Scholar Collection of Engaged Learning Engaged Learning 4-15-2013 Anatomical description and phylogenetic analysis of Miocene beaked whale from the East African Rift Valley,
Southern Methodist University SMU Scholar Collection of Engaged Learning Engaged Learning 4-15-2013 Anatomical description and phylogenetic analysis of Miocene beaked whale from the East African Rift Valley,
BREVIORA LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1
, A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1") ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
The family Gnaphosidae is a large family
 Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA
 FROM JAVA, INDONESIA") NOTES AND NEWS UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA BY NGUYEN NGOC-HO i) Faculty of Science, University of Saigon, Vietnam Among material recently collected
NOTES AND NEWS UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA BY NGUYEN NGOC-HO i) Faculty of Science, University of Saigon, Vietnam Among material recently collected
J/ieuican JfiLsllm. The Genus Proterix (Insectivora, Erinaceidae) of the Upper Oligocene of North America BY CONSTANCE ELAINE GAWNE1 INTRODUCTION
 of the Upper Oligocene of North America BY CONSTANCE ELAINE GAWNE1 INTRODUCTION") A J/ieuican JfiLsllm PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N. Y. I0024 NUMBER 2 3 I 5 FEBRUARY 28, I968 The Genus Proterix (Insectivora, Erinaceidae)
A J/ieuican JfiLsllm PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N. Y. I0024 NUMBER 2 3 I 5 FEBRUARY 28, I968 The Genus Proterix (Insectivora, Erinaceidae)
Description of Cranial Elements and Ontogenetic Change within Tropidolaemus wagleri (Serpentes: Crotalinae).
.") East Tennessee State University Digital Commons @ East Tennessee State University Electronic Theses and Dissertations 5-2016 Description of Cranial Elements and Ontogenetic Change within Tropidolaemus
East Tennessee State University Digital Commons @ East Tennessee State University Electronic Theses and Dissertations 5-2016 Description of Cranial Elements and Ontogenetic Change within Tropidolaemus
Supplementary Information for: 3D morphometric analysis of fossil canid skulls contradicts
 Supplementary Information for: 3D morphometric analysis of fossil canid skulls contradicts the suggested domestication of dogs during the late Paleolithic Abby Grace Drake 1, * Michael Coquerelle 2,3 Guillaume
Supplementary Information for: 3D morphometric analysis of fossil canid skulls contradicts the suggested domestication of dogs during the late Paleolithic Abby Grace Drake 1, * Michael Coquerelle 2,3 Guillaume
PSYCHE A NEW GENUS AND SPECIES OF SALDIDAE FROM SOUTH AMERICA (HEMIPTERA) BY CARL J. DRAKE AND LUDVIK HOBERLANDT. Iowa State College, Ames
 BY CARL J. DRAKE AND LUDVIK HOBERLANDT. Iowa State College, Ames") PSYCHE Vol. 59 September, 1952 No. 3 A NEW GENUS AND SPECIES OF SALDIDAE FROM SOUTH AMERICA (HEMIPTERA) BY CARL J. DRAKE AND LUDVIK HOBERLANDT Iowa State College, Ames Through the kindness of Dr. P. J.
PSYCHE Vol. 59 September, 1952 No. 3 A NEW GENUS AND SPECIES OF SALDIDAE FROM SOUTH AMERICA (HEMIPTERA) BY CARL J. DRAKE AND LUDVIK HOBERLANDT Iowa State College, Ames Through the kindness of Dr. P. J.
Descriptions of New North American Fulgoridae
 The Ohio State University Knowledge Bank kb.osu.edu Ohio Journal of Science (Ohio Academy of Science) Ohio Journal of Science: Volume 5, Issue 8 (June, 1905) 1905-06 Descriptions of New North American
The Ohio State University Knowledge Bank kb.osu.edu Ohio Journal of Science (Ohio Academy of Science) Ohio Journal of Science: Volume 5, Issue 8 (June, 1905) 1905-06 Descriptions of New North American
DESCRIPTIONS OF NEW AND LITTLE-KNOWN FOSSIL LIZARDS FROM NORTH AMERICA
 PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol.86 Washington: 1938 No. 3042 DESCRIPTIONS OF NEW AND LITTLE-KNOWN FOSSIL LIZARDS FROM NORTH AMERICA By
PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol.86 Washington: 1938 No. 3042 DESCRIPTIONS OF NEW AND LITTLE-KNOWN FOSSIL LIZARDS FROM NORTH AMERICA By
ON THE SCALOPOSAURID SKULL OF OLIVIERIA PARRINGTONI, BRINK WITH A NOTE ON THE ORIGIN OF HAIR
 ON THE SCALOPOSAURID SKULL OF OLIVIERIA PARRINGTONI, BRINK WITH A NOTE ON THE ORIGIN OF HAIR By G. H. Findlay, D.Sc., M.D. (Professor of Dermatology, University of Pretoria; Director, C.S.I.R. Photobiology
ON THE SCALOPOSAURID SKULL OF OLIVIERIA PARRINGTONI, BRINK WITH A NOTE ON THE ORIGIN OF HAIR By G. H. Findlay, D.Sc., M.D. (Professor of Dermatology, University of Pretoria; Director, C.S.I.R. Photobiology
Mar., 1963 RELATIONSHIPS BETWEEN THE BIRDS OF PARADISE AND THE BOWER BIRDS. By WALTER J. BOCK
 Mar., 1963 91 RELATIONSHIPS BETWEEN THE BIRDS OF PARADISE AND THE BOWER BIRDS By WALTER J. BOCK INTRODUCTION Ever since their discovery in the early days of world exploration, the birds of paradise and
Mar., 1963 91 RELATIONSHIPS BETWEEN THE BIRDS OF PARADISE AND THE BOWER BIRDS By WALTER J. BOCK INTRODUCTION Ever since their discovery in the early days of world exploration, the birds of paradise and
PRELIMINARY REPORT ON A CLUTCH OF SIX DINOSAURIAN EGGS FROM THE UPPER TRIASSIC ELLIO T FORMATION, NORTHERN ORANGE FREE STATE. J. W.
 41 Pa/aeont. afr., 22, 41-45 (1979) PRELIMINARY REPORT ON A CLUTCH OF SIX DINOSAURIAN EGGS FROM THE UPPER TRIASSIC ELLIO T FORMATION, NORTHERN ORANGE FREE STATE b y J. W. Kitching ABSTRACT A clutch of
41 Pa/aeont. afr., 22, 41-45 (1979) PRELIMINARY REPORT ON A CLUTCH OF SIX DINOSAURIAN EGGS FROM THE UPPER TRIASSIC ELLIO T FORMATION, NORTHERN ORANGE FREE STATE b y J. W. Kitching ABSTRACT A clutch of
Diurus, Pascoe. sp. 1). declivity of the elytra, but distinguished. Length (the rostrum and tails 26 included) mm. Deep. exception
. declivity of the elytra, but distinguished. Length (the rostrum and tails 26 included) mm. Deep. exception") 210 DIURUS ERYTIIROPUS. NOTE XXVI. Three new species of the Brenthid genus Diurus, Pascoe DESCRIBED BY C. Ritsema+Cz. 1. Diurus erythropus, n. sp. 1). Allied to D. furcillatus Gylh. ²) by the short head,
210 DIURUS ERYTIIROPUS. NOTE XXVI. Three new species of the Brenthid genus Diurus, Pascoe DESCRIBED BY C. Ritsema+Cz. 1. Diurus erythropus, n. sp. 1). Allied to D. furcillatus Gylh. ²) by the short head,
A M E G H I N I A N A. Revista de la Asociación Paleontológia Argentina. Volume XV September-December 1978 Nos. 3-4
 A M E G H I N I A N A Revista de la Asociación Paleontológia Argentina Volume XV September-December 1978 Nos. 3-4 COLORADIA BREVIS N. G. ET N. SP. (SAURISCHIA, PROSAUROPODA), A PLATEOSAURID DINOSAUR FROM
A M E G H I N I A N A Revista de la Asociación Paleontológia Argentina Volume XV September-December 1978 Nos. 3-4 COLORADIA BREVIS N. G. ET N. SP. (SAURISCHIA, PROSAUROPODA), A PLATEOSAURID DINOSAUR FROM
Review of the Miocene long-snouted dolphin Priscodelphinus cristatus. eurhinodelphinids. by Olivier LAMBERT
 BULLETIN DE L INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE SCIENCES DE LA TERRE, 75: 211-235, 2005 BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETENSCHAPPEN AARDWETENSCHAPPEN, 75: 211-235,
BULLETIN DE L INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE SCIENCES DE LA TERRE, 75: 211-235, 2005 BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETENSCHAPPEN AARDWETENSCHAPPEN, 75: 211-235,
Lower Cretaceous Kwanmon Group, Northern Kyushu
 Bull. Kitakyushu Mus. Nat. Hist., 11: 87-90. March 30, 1992 A New Genus and Species of Carnivorous Dinosaur from the Lower Cretaceous Kwanmon Group, Northern Kyushu Yoshihiko Okazaki Kitakyushu Museum
Bull. Kitakyushu Mus. Nat. Hist., 11: 87-90. March 30, 1992 A New Genus and Species of Carnivorous Dinosaur from the Lower Cretaceous Kwanmon Group, Northern Kyushu Yoshihiko Okazaki Kitakyushu Museum
AMERICAN MUSEUM NOVITATES
 AMERICAN MUSEUM NOVITATES Published by Number 749 THE AMERICAN MuewmoF NATURAL HISTORY Oct. 8, 1934 56.9, 72 R (1183: 54) A NEW RHINOCEROS FROM THE SIWALIK BEDS OF INDIA BY EDWIN H. COLBERT INTRODUCTION
AMERICAN MUSEUM NOVITATES Published by Number 749 THE AMERICAN MuewmoF NATURAL HISTORY Oct. 8, 1934 56.9, 72 R (1183: 54) A NEW RHINOCEROS FROM THE SIWALIK BEDS OF INDIA BY EDWIN H. COLBERT INTRODUCTION
Lab 5: Rodentia and Lagomorpha
 Lab 5: Rodentia and Lagomorpha (8 families in B.C.) Sciuridae squirrels (16 species in B.C.) Muridae mice, rats, lemmings, voles (16) Aplodontidae mountain beaver (1) Castoridae beaver (1) Dipodidae jumping
Lab 5: Rodentia and Lagomorpha (8 families in B.C.) Sciuridae squirrels (16 species in B.C.) Muridae mice, rats, lemmings, voles (16) Aplodontidae mountain beaver (1) Castoridae beaver (1) Dipodidae jumping
Cranial osteology and phylogenetic relationships of Hamadasuchus rebouli (Crocodyliformes: Mesoeucrocodylia) from the Cretaceous of Morocco
 from the Cretaceous of Morocco") Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1494 533567 Original Articles HAMADASUCHUS REBOULIH. C. E. LARSSON and H.-D.
Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1494 533567 Original Articles HAMADASUCHUS REBOULIH. C. E. LARSSON and H.-D.
The Discovery of a Tritylodont from the Xinjiang Autonomous Region
 The Discovery of a Tritylodont from the Xinjiang Autonomous Region Ailing Sun and Guihai Cui (Institute of Vertebrate Paleontology, Paleoanthropology, Academia Sinica) Vertebrata PalAsiatica Volume XXVII,
The Discovery of a Tritylodont from the Xinjiang Autonomous Region Ailing Sun and Guihai Cui (Institute of Vertebrate Paleontology, Paleoanthropology, Academia Sinica) Vertebrata PalAsiatica Volume XXVII,
The cranial osteology of Belebey vegrandis (Parareptilia: Bolosauridae), from the Middle Permian of Russia, and its bearing on reptilian evolution
, from the Middle Permian of Russia, and its bearing on reptilian evolution") Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1511 191214 Original Articles RUSSIAN BOLOSAURID REPTILER. R. REISZ ET AL.
Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1511 191214 Original Articles RUSSIAN BOLOSAURID REPTILER. R. REISZ ET AL.
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS
 AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.