Evolution, multiple acquisition, and localization of endosymbionts in bat flies. corresponding author: Katharina Dittmar;
|
|
|
- Roland Douglas
- 5 years ago
- Views:
Transcription
1 AEM Accepts, published online ahead of print on 22 February 2013 Appl. Environ. Microbiol. doi: /aem Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 Evolution, multiple acquisition, and localization of endosymbionts in bat flies (Diptera: Hippoboscoidea: Streblidae and Nycteribiidae) Solon F. Morse 1,2, Sarah E. Bush 3, Bruce D. Patterson 4, Carl W. Dick 5, Matthew E. Gruwell 6, Katharina Dittmar 1,2 corresponding author: Katharina Dittmar; kd52@buffalo.edu 1 Department of Biological Sciences, University at Buffalo (SUNY), Buffalo, NY, 14260, USA, kd52@buffalo.edu 2 Graduate Program for Ecology, Evolution and Behavior, University at Buffalo, (SUNY) Buffalo, NY, 14260, USA, smorse@buffalo.edu 3 Department of Biology, University of Utah, Salt Lake City, UT 84112, bush@biology.utah.edu 4 Department of Zoology, Field Museum of Natural History, Chicago IL 60605, USA, bpatterson@fieldmuseum.org 5 Department of Biology, Western Kentucky University, Bowling Green, KY 42101, USA, carl.dick@wku.edu 6 School of Science, Penn State Erie, The Behrend College, Erie, PA 16568, USA, meg26@psu.edu
2 Abstract Bat flies are a diverse clade of obligate ectoparasites on bats. Like most blood-feeding insects they harbor endosymbiotic prokaryotes, but the origins and nature of these symbioses are still poorly understood. To expand the knowledge of bacterial associates in bat flies, the diversity and evolution of the dominant endosymbionts in six of eight nominal subfamilies of bat flies (Streblidae, Nycteribiidae) were studied. Furthermore, the localization of endosymbionts and their transmission across developmental stages within the family Streblidae were explored. Results show diverse microbial associates in bat flies, with at least four ancestral invasions of distantly related microbial lineages throughout bat fly evolution. Phylogenetic relationships support the presence of at least two novel symbiont lineages (here: Clades B and D), and extend the geographic and taxonomic range of a previously documented lineage (Candidatus Aschnera chinzeii; here: Clade A). Although these lineages show reciprocally monophyletic clusters with several bat fly host clades, their phylogenetic relationships generally do not reflect current bat fly taxonomy or phylogeny. However, within some endosymbiont clades, congruent patterns of symbiont-host divergence are apparent. Other sequences identified in this study fall into the widely distributed, highly invasive, insect-associated Arsenophonus lineage, and may be the result of symbiont replacements and/or transient infections (here: Clade C). Vertical transmission of endosymbionts of clades B and D is supported by fluorescent signal (FISH) and microbial DNA detection across developmental stages. Fluorescent bacterial signal is consistently localized within structures resembling bacteriomes, although their anatomical position differs by host fly
3 clade. In summary, results suggest an obligate host-endosymbiont relationship for three of the four known symbiont clades associated with bat flies (Clades A, B, and D).
4 Introduction Bat flies are a diverse group of obligate ectoparasites that feed exclusively on bat blood (1, 2). Based on current taxonomy, they comprise two families, the Nycteribiidae and the Streblidae (3). As blood-feeders they rely on a nutritionally deficient diet, hence they are expected to have formed symbiotic relationships with mutualistic bacteria (4-9). Recent studies have examined several genera of Streblidae and Nycteribiidae for the presence of endosymbionts. Trowbridge, et al. (6) documented an Arsenophonus-like bacteria in several fly species belonging to the genus Trichobius (Streblidae, Trichobiinae). This was subsequently corroborated by Lack, et al. (10) for several populations of Trichobius major, and by Nováková, et al. (11) for an unknown Trichobius species. Arsenophonus spp. are widespread Gammaproteobacteria that show a great diversity of symbiotic relationships with their arthropod hosts, ranging from reproductive parasites, facultative symbionts of unknown function, to vertically transmitted obligate mutualists (7, 11-19). In addition to mutualistic roles, Lack, et al. (10) hypothesized that Arsenophonus manipulates host reproduction in T. major. Nováková, et al. (11) and Hosokawa, et al. (20) documented endosymbionts in several genera and species of Nycteribiinae (Nycteribiidae). Although similar in 16S rrna gene sequences to Arsenophonus, they were ultimately assigned to a distinct lineage that is sister to Candidatus Riesia pediculicola, an endosymbiont found in primate lice. Based on phylogenetic characteristics, Hosokawa, et al. (20) proposed the name Candidatus Aschnera chinzeii for this clade. The presence of vertical transmission and an overall evolutionary concordance between host and symbiont suggest an obligate association with its host (20). Although the specific function of Candidatus Aschnera
5 chinzeii is unknown, the closely related Candidatus Riesia pediculicola is essential for the survival of its louse host, providing B complex vitamins that are lacking in the host s blood diet (21). Despite previous efforts, no comprehensive studies of bat fly microbiomes exist that reflect the taxonomic diversity of their fly hosts, hindering our understanding of the diversity, function and evolution of endosymbiotic associations in bat flies. Bat flies, like their bat hosts, are cosmopolitan in distribution, and more than 500 species are known to parasitize many genera of bats. They are members of the Hippoboscoidea, a group of highly specialized blood-feeding Diptera, and as such are allied with the tsetse flies (Glossinidae) and the louse flies (Hippoboscidae) (22). Streblidae is composed of five subfamilies, the New World Nycterophiliinae, Streblinae, and Trichobiinae, and the Old World Nycteriboscinae and Ascodipterinae (2, 23). Nycteribiidae currently contains three subfamilies: the Old World Cyclopodiinae and Archinycteribiinae, and the cosmopolitan Nycteribiinae (2). Bat flies are found chiefly in tropical and subtropical regions, with a fewer number of temperate representatives. Current phylogenetic consensus suggests that the Streblidae are paraphyletic, with the monophyletic Nycteribiidae included within Old World Streblidae (3, 22). Bat flies are not only taxonomically and morphologically diverse, but also have a peculiar reproductive biology. Like all Hippoboscoidea, they are adenotrophically viviparous: a single larva develops in the female, fed by secretions from milk glands, which are highly modified accessory glands. Third-instar larvae of streblid and nycteribiid bat flies are deposited as sessile prepupae onto a substrate, like a cave wall, where they immediately pupate (24, 25). This pupiparity makes horizontal transfer of
6 symbionts unlikely, as there is no feeding stage outside the body of the maternal parent. Hosokawa, et al. (20) observed symbiont infection in the maternal milk gland in several members of the Nycteribiinae, suggesting uterine transmission of bacteria to the larvae through milk gland secretions, which also has been documented for tsetse flies (26). To expand our knowledge of the evolution and transmission of endosymbiotic bacterial associates in bat flies, we explored their prevalence and diversity in six of the eight bat fly subfamilies. Specifically, we sought to determine if previously identified bat fly endosymbionts are consistently present across families and subfamilies throughout their geographic distribution. Furthermore, we explored the evolutionary relationships among identified endosymbiotic clades, and showed endosymbiont localization and transmission across developmental stages within the family Streblidae. Downloaded from on October 23, 2018 by guest
7 Materials and methods Samples. Specimens were collected in microfuge tubes containing 96% ethanol, and stored at -80 C for further use. Representatives of 42 species were included in this study, with an average of 1.9 specimens per species (min:1; max:5). Samples included representative members of both known families of bat flies: Nycteribiidae (17 species) and Streblidae (25 species). Six of the eight described subfamilies of bat flies were included in this study, including: a) the streblid subfamilies Nycteriboscinae, Streblinae, Trichobiinae and Nycterophiliinae, and b) the nycteribiid subfamilies Cyclopodiinae and Nycteribiinae. Two subfamilies, Archinycteribiinae (Nycteribiidae) and Ascodipterinae (Streblidae) are not represented for lack of specimens. Samples were collected across a wide geographic and host-bat distribution (Table 1). In order to test for vertical transmission of symbionts in New World Streblidae, pupae from Trichobius frequens, T. intermedius, and Nycterophilia cf. coxata were collected either from pupal deposition sites on cave walls, or were obtained by capturing adult female flies on glue traps suspended from poles placed in the cave. Captured females subsequently deposited pupae directly on the trap (27). Pupae collected in this manner were 6-12 hours post-deposition, and had minimal exposure to the larger cave environment. To compare infection rates between male and female bat flies, 64 T. frequens samples from the same Puerto Rican population were sexed and tested for symbiont presence, as were 33 Puerto Rican Nycterophilia cf. coxata flies. DNA extraction. DNA was extracted from whole bat fly adults and pupae using the Qiagen DNeasy kit and protocol. For adult flies, the abdomen of each sample was pierced so as to maintain exoskeleton integrity to allow for mounting, identification and sexing
8 post-extraction. Pupae were washed in 3% hydrogen peroxide solution and dissected out of the puparium under sterile conditions. DNA concentration of each sample was quantified using a spectrophotometer (Nanodrop, ND-1000, Wilmington, DE). PCR amplification of bacterial 16S rrna and grol genes. A 1.3-kb fragment of the eubacterial 16S rrna gene (SSU rrna) was amplified via PCR using the general eubacterial 16S primers 16Sa1 (5'- AGA GTT TGA TCM TGG CTC AG -3') and 16Sb1 (5'- TAC GGY TAC CTT GTT ACG ACT T -3') following the PCR protocol outlined in Fukatsu and Nikoh (28). An 850-bp region of the chaperonin grol gene was amplified using the primers groel-2f (5'- ATG GGB GCT CAA ATG GTK AAA -3') and groel- 2R (5'- CTC TTT CAT TTC AAC TTC NGT BGC A -3') following the protocols outlined in Hosokawa and Fukatsu (29) and Hosokawa, et al. (20). Because some grol sequences did not amplify using these primers, additional custom primers were designed: the forward primer groel196f (5'- TTY GAR AAT ATG GGW GCH CAA ATG -3') and reverse primers groel840r (5'- DCC AGG AGC YTT NAC AGC AG -3') and groel1247r (5'- CHA CHA CHC CTT CTT CHA CHG C -3'). PCR was performed in a final volume of 25 µl containing 11 µl H 2 O, 2 µl 50 mm MgCl 2, 3 µl 10x Taq buffer, 2.5 µl 10 mm dntp mix, 2.5 µl loading dye, 1 µl (10pmol/µl) each primer, 1 µl of template DNA, and 0.2 µl of Taq DNA polymerase (Promega, Madison, WI, U.S.A.). The amplification conditions were 2 min at 95 C followed by 30 cycles of 1 min at 95 C, 1 min at 55 C, and 1.15 min at 72 C, with a final extension at 72 C for 10 min. Negative controls (lacking template DNA) were included in all amplifications. Positive PCR samples were cloned into the pcr 2.1 vector using the TOPO-TA Cloning Kit following the instructions of the manufacturer (Invitrogen). At least sixteen
9 insert-positive colonies per cloned sample (range 16-30) were sent to the High Throughput Genomics Unit (Seattle, WA) for cleaning and sequencing using the original forward primer and the standard vector M13 reverse primer supplied with the TOPO-TA kit. Evolutionary analyses. Raw sequences were edited and assembled using Geneious Pro 5.6. Taxonomic affinities of the sequences were identified using NCBI s BLASTn search. QIIME was used to check for chimeric sequences. As outgroups for these analyses we chose bacteria previously used in similar studies, which were clearly identified as evolutionarily distant to endosymbionts in this analysis. Sequences were aligned with MAFFT (30) as implemented in Geneious Pro 5.6. Bacterial 16S rrna gene sequences contained highly conserved regions interspersed with divergent regions and were therefore aligned using the E-INS-i algorithm, which is optimized for aligning sequences with multiple conserved domains and long gaps (30); grol sequences were aligned using the G-INS-i algorithm, which is optimized for aligning sequences with global homology (30). The 16S rrna sequence alignments exhibited several ambiguously aligned regions: the GBLOCKS program (31) was used to remove these poorly aligned positions and to obtain unambiguous sequence alignments, with parameters allowing for less strict blocks, gaps within the final blocks, and less strict flanking positions. The 3 rd codon position was excluded from our analysis of the grol gene to avoid problems associated with saturation. Evolutionary models were selected for each gene using the Akaike Information Criterion (AIC) as implemented in jmodeltest (32). Relationships of bacterial 16S rrna genes were explored using a network approach, which assesses the underlying structure, ambiguity and conflict in the data that
10 a bifurcating tree might miss (33). Analyses were conducted with SplitsTree 4.0 (34), using the GTR evolutionary model in conjunction with the agglomerating NeighborNet algorithm. For comparative purposes data were combined with select GenBank entries of invertebrate gammaproteobacterial symbionts and free-living Gammaproteobacteria. Phylogenetic analyses were conducted on samples represented by grol and 16S DNA, using Maximum Likelihood (ML) and Bayesian methods. ML analysis was done using PhyML Version 3 (35), as implemented in Geneious Pro 5.6. Node support was assessed by bootstrap analyses with 1000 bootstrap replicates. Bayesian topologies were obtained through MrBayes (36); posterior probabilities were used to gauge nodal support. Relative rate tests of lineage evolution were performed using RRTree on the grol dataset (37). Tests involved the comparisons of major bat fly symbiont lineages (see results) to each other, resulting in three comparisons (Table 1). Whole-mount fluorescent in situ hybridization (FISH) and histology. Fluorescent in situ hybridization (FISH) was carried out on pupae and adults of T. frequens, as well as adults of N. cf. coxata, following the protocols outline in Koga, et al. (38). Both genera are members of the widely distributed New World Streblidae, which had not yet been studied using FISH techniques. Hybridization was performed using the eubacterial probe EUB338 (5'-GCTGCCTCCCGTAGGAGT-3') labeled with Alexafluor 647 [5' end]. Acetone- or ethanol-preserved adults and pupae were fixed in Carnoy's solution (chloroform:ethanol:glacial acetic acid, 6:3:1), following the removal of legs in adults to facilitate penetration of fixative. Pupae were dissected out of the puparium before fixation. All specimens were incubated in 150 µl of hybridization buffer (20 mm Tris
11 HCl, ph 8.0; 0.9 M NaCl; 0.01% sodium dodecyl sulfate; 30% formamide) containing 50 pmol/ml of probe along with DAPI at 42 C overnight. The samples were then washed with hybridization buffer made from 50% formamide at 42 for 15 min, followed by a 1 SSC (0.15 M NaCl, 15 mm sodium citrate) wash at 42 for 10 min and two 1x PBS washes at room temperature for 30 min each. The samples were mounted on glass microscope slides (VWR) using SlowFade mounting medium (Molecular Probes, Eugene, Oregon, USA). Mounted specimens were visualized under a ZEISS LSM 710 laser confocal microscope. In order to identify the organismal source of the fluorescence in Trichobius flies, representative specimens were sectioned and stained. Bat flies were fixed in freshly made 4% PFA + 0.1% Triton X-100 for 4-12 h for histological sectioning. After fixation, specimens were transferred through a dehydrating ethanol series, starting with 30% ETOH, and ending with final storage in 96% ETOH. Specimens were embedded in Periplast Paraffin following previous protocols. After dorsal-to-ventral sectioning, slides were stained following the Luxol Fast Blue protocol. Sections were examined using a 40X, oil-immersion objective on a ZEISS Phase Contrast Fluorescence Inverted Microscope. However, Nycterophilia specimens are laterally compressed and very small (~1.65 mm total length), making embedding and sectioning untenable. Because specimens are fairly transparent, we used light microscopy on whole specimens to identify internal abdominal structures associated with FISH localization. Results
12 Sequences. Microbial DNA was detected in all samples studied in this effort. We obtained 68 unique microbial 16S DNA and 50 unique microbial grol gene sequences from the adult flies, the majority of which exhibited a high degree of similarity to sequence representatives of Candidatus Riesia, Arsenophonus, or Candidatus Aschnera chinzeii when identified by BLASTn. Exceptions are bacterial sequences of Nycterophilia and the Trichobius caecus group, which had Providencia sequences as their best hits (89% pairwise similarity with Providencia spp. (e-value = 0.0). Other sequences identified as part of the bat fly microbiome showed high similarity to the alphaproteobacteria Bartonella (39), Wolbachia, and Rickettsia. These sequences were detected in a subset of samples (15.1% of adult samples, combined). The 16S rrna gene sequences for the symbionts of clades A, B and D (Figure 1) contained an AT bias of 48 50%, 48 52%, and 51 53%, respectively. All three clades fall within in the range of other insect symbionts obtained from GenBank. Clade C (Arsenophonus) symbionts exhibited a lower AT bias, ranging from 45 47%. There was also an AT bias in grol sequences for these symbiont clades: clades A and B both ranged from 64 69%, while clade D ranged from 67-70%. Clade C (Arsenophonus) exhibited a range of values from 58-64%, with values at the high end (63-64%) for Arsenophonus symbionts associated to the monophyletic Eucampsipoda symbiont clade. This compares to 61-69% for other endosymbionts and 48-57% for free-living bacteria. Representative population samples from Trichobius frequens (Streblidae) showed that 82% (31/38) of teneral (freshly emerged, unfed) females and 65% (17/26) of teneral males tested positive for clade B symbionts, using either the 16S or grol primers. Proportions of infected males and females were not significantly different (chi-square
13 test; χ 2 = 2.131; P > 0.15). In specimens of Nycterophilia cf. coxata (Nycterophiliinae), all females and males were infected with Clade D symbiont [100%]. Using PCR to amplify either the 16S rrna or grol genes, we detected Arsenophonus infections in 85% (17/22) of T. frequens pupae, 87% (7/8) of T. intermedius pupae, and 100% (3/3) pupae of Nycterophilia cf. coxata flies. Phylogenetic and Network analyses. Phylogenetic analyses of the concatenated data revealed that bat fly endosymbionts fall into four major divergent clades (named A through D), each of which receives strong nodal support. These clades were congruently recovered by both ML and Bayesian methodologies (Figure 3; ML trees presented). The clades are also present as well-delineated clusters in the network analysis, although the overall network appears unresolved and scattered at the base (Figure 1). Clade A is equivalent to Candidatus Aschnera chinzeii (20). It contains samples from the subfamily Nycteribiinae, including the genera Basilia, Nycteribia, Penicillidia, and Phthiridium (Figures 1, 3). Samples in this clade closely reflect evolutionary relationships of their bat fly hosts (20). Clade A is restricted to the Old World, with samples from Japan, China, Malaysia, the Philippines, and Slovenia. Although this clade is recovered as a sister-group to Candidatus Riesia pediculicola in the ML and Bayesian analyses, this relationship is poorly supported. In fact, in the network analysis the Candidatus Riesia clade allies closely to clade D (Figure 1, see below). Clade B constitutes a novel bacterial clade that unites endosymbiont sequences from the New World subfamilies Trichobiinae and Streblinae (Figures 1, 3), including samples from Puerto Rico, Mexico, the Dominican Republic, French Guyana, and the U.S. The monophyly of clades A and B together with Candidatus Riesia pediculicola is
14 moderately supported in both ML and Bayesian phylogenies, and relatively long branches in clades A and B (Figure 3) indicate high average sequence divergence from Candidatus Riesia and Clade A and B. Clade C, which is recovered with high support in the ML and Bayesian reconstruction, appears scattered and unresolved in the network analyses (Figures 1, 2). Because of the high similarity of bat fly symbiont sequences in this clade to the representative species of Arsenophonus (A. triatomarum, and A. nasoniae), we suggest these belong to that genus. This clade also contains most of the symbiont sequences obtained from the closely related Hippoboscidae (ked and louse flies) (Figures 1, 2, 3). Microbial sequences detected in the Old World subfamilies Cyclopodiinae and Nycteriboscinae fall into this clade, although within-clade relationships remain obscure due to lack of statistical support. Although this group mostly unites symbiont sequences from Old World bat flies (Philippines, China, Europe), a few New World species are interspersed among them, most notably the two New World representatives of the cosmopolitan nycteribiid genus Basilia (Nycteribiidae) and singular samples from the Neotropical genera Paradyschiria and Anatrichobius (Streblidae: Trichobiinae) (Figure 2). Clade D includes two novel subclades from the New World streblid subfamily Nycterophiliinae and the New World Trichobiinae (T. caecus group; (40)). Within each clade, endosymbiont sequences cluster by host species. Strong patterns of co-divergence have been found in the Nycterophiliinae subclade (41). Tests of relative rates of lineage evolution in grol revealed that if considered separately, bat fly clades A+B+Candidatus Riesia and clade D showed significantly
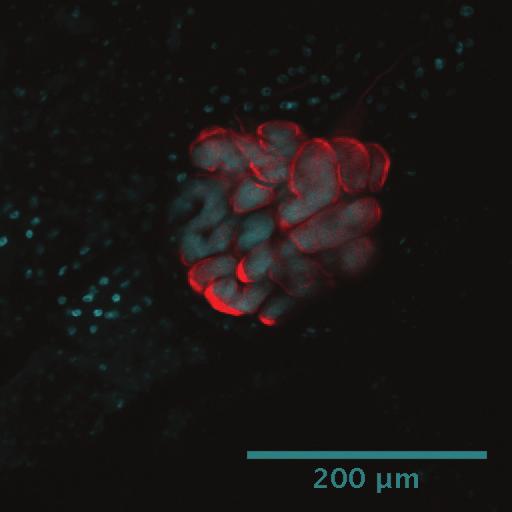
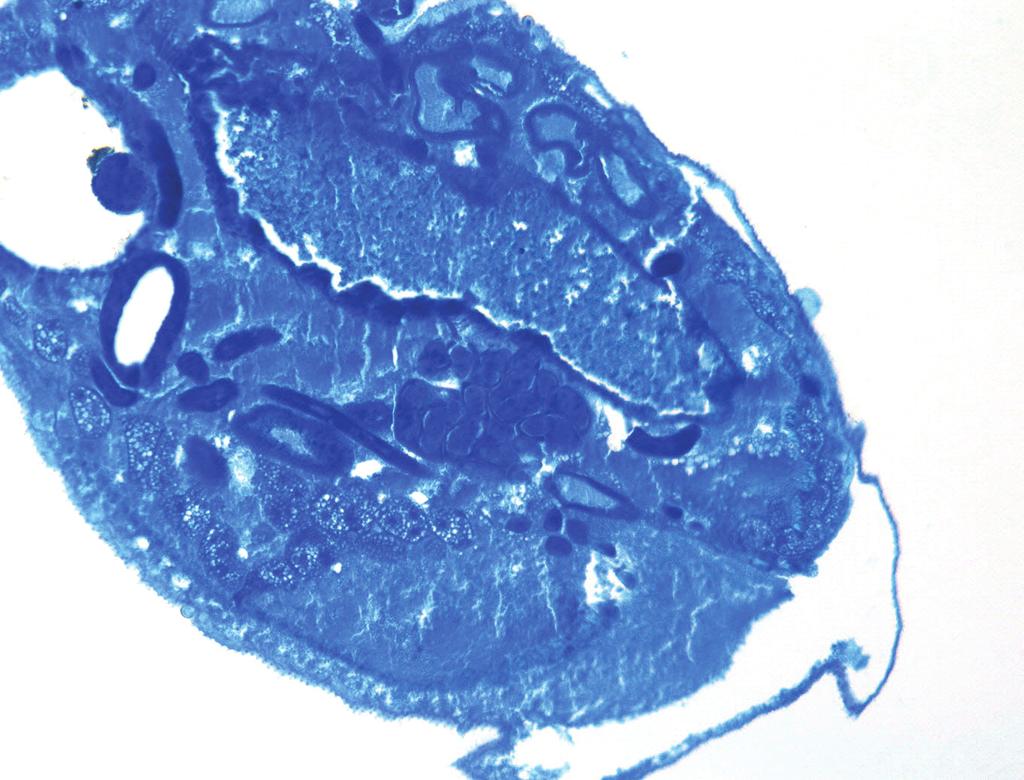
15 higher rates of evolution compared to Arsenophonus sensu stricto (lineage C) (Table 1). No significant differences were detected when comparing clades A+B+Candidatus Riesia and clade C to each other. Dissections and in-situ hybridization. To localize endosymbionts of the family Streblidae, whole mounts of pupae, males and females of T. frequens (Clade B), and males and females of N. cf. coxata (Clade D) were labeled with fluorescent probes. Because a general eubacterial probe was used, potentially all bacterial associates (including Bartonella, Wolbachia, and Rickettsia, when present) were highlighted. Results showed singular, localized fluorescence in the abdomen of both species, with well-defined boundaries. In adult Trichobius flies, the fluorescent signal was located in the dorsal mid-section of the abdomen extending to or slightly beyond the height of spiracle 3 (Figure 4b). The location and strength of the signal was consistent across all specimens tested (12), and did not vary by sex (6 males and 6 females). Based on the sections obtained from the Trichobius specimens, their bacteriome seems associated to the dorsal anterior midgut region, but not directly connected to the midgut (Figure 4e). In pupae, fluorescence was detected immediately after deposition (age: 36 hrs, image not shown) and appeared clustered in the abdominal region at the mid-point of pupal development (Figure 4d, pupa 12 days old; (27)). In Nycterophilia flies, the signal occupied the anterior, ventral section of the abdomen, below the ventral portion of sternite 1 (Figure 5a,b). Fluorescent signal was only detected in females (see Discussion). Light microscopy of blood-fed flies showed the signal to be anatomically closely associated to the base of an organ resembling the crop (Figure 5d).
16 In both species, the localization of the probe signal demonstrated an intracellular presence of bacteria. Bacteria occurred in hypertrophied cells (bacteriocytes), which clustered into a single bacteriome. In Trichobius, the average size of the bacteriocytes was 55±14.9 µm (N=16 bacteriocytes measured from 3 females). The bacteriome formed a disk-shaped, single cell layer structure, consisting of 7 13 cells (Figure 4c). In Nycterophilia, the average size of the bacteriocytes was 39±5.7 µm (N=6 bacteriocytes measured from 3 females). The bacteriome consisted of 1-3 aggregated cells (Figure 5c). Discussion Acquisition and evolution of endosymbionts in bat flies. Given the distribution of bat fly families and subfamilies across the endosymbiont phylogeny, it becomes clear that bat flies independently acquired heterogeneous symbionts multiple times throughout their evolutionary history. Based on the clades in the ML phylogeny (16S rrna and grol genes), we suggest at least one ancestral acquisition for the Nycteribiidae (clades A; Figure 3); and three events for the Streblidae (clades B and D for Trichobiinae, clade D for Nycterophiliinae; Figure 3). This is also supported by the extreme AT bias and long branch-length exhibited by clades A, B and D, suggesting that the endosymbiotic lifestyle is ancient, evolution was particularly rapid, or both (42) (Table 1). Our calculation of obligate symbiont acquisition events considers only well-supported clades of host families, which are represented by multiple genera. This is a conservative estimate, and it is conceivable that additional data will increase this count, especially considering possible replacement events in clade C (Figure 2).
17 Our data confirmed the presence of a previously identified, exclusively Old World monophyletic clade of obligate bat fly endosymbionts, known as Candidatus Aschnera chinzeii (clade A) (20). However, the geographic distribution of this clade encompasses a much broader range than Japan (as documented by Hosokawa, et al. (20)), extending into mainland Asia and the Palearctic. The results of this study support suggestions of cospeciation between Nycteribiinae and their endosymbionts, with some conspicuous exceptions. Endosymbiont sequences obtained from two Basilia spp. (Nycteribiinae) fall outside clade A, and cluster into the widely distributed Arsenophonus clade C (Figure 2). Interestingly, the Basilia spp. in question align with the New World representatives of this genus. Despite being collected in different localities (US and Panama), their Arsenophonus sequences show 99.2% pairwise similarity, suggesting a possible endosymbiont replacement in New World Basilia (and Nycteribiinae), as well as a geographic structuring of endosymbiont communities. Furthermore, two endosymbiont sequences from Nycteribia spp. [Nycteribiinae; geographic locality unknown; GenBank accession numbers FJ and FJ265804; (11)] fall outside of Clade A (Figure 2), again suggesting endosymbiont replacements within another geographically wide-ranging genus. Alternatively, these clade C sequences may represent transient, local infections. Because of limited sampling throughout the known geographic ranges of Basilia and Nycteribia, it remains ambiguous whether this is truly a replacement rather than a transient infection, and the frequency of such replacements remains unclear. Streblid endosymbionts identified by Trowbridge, et al. (6), Lack, et al. (10), Nováková, et al. (11) were previously assigned to the genus Arsenophonus. In our analysis, they form their own, novel monophyletic lineage (clade B), which is distinct but
18 allied to Candidatus Riesia pediculicola (Figure 3). Therefore, in a manner similar to Candidatus Aschnera chinzeii, this clade should be considered distinct from both Candidatus Riesia and Arsenophonus. The prior assessment of these sequences as Arsenophonus is an example of how studies involving small numbers of sequences limit our understanding of microbial taxonomy. The uniqueness of this clade is further supported by high sequence divergence and the significantly faster evolutionary rate along the branch leading to the ancestral node of lineage B, when compared to the Arsenophonus clade (Table 1). Specifically, clade B contains endosymbionts of bat flies in the subfamilies Trichobiinae and Streblinae, with the striking exception of those of the T. caecus group, which supports clade D symbionts (see below). Within clade B, three poorly supported endosymbiont clades emerge, uniting bacterial sequences obtained from the bat fly genera Aspidoptera, Strebla, Megistopoda, Paratrichobius, and Trichobius (Figure 3). These subclades correspond roughly with the bat fly species divisions (series 1 through 3) of Wenzel and Tipton (1), but more data are needed to warrant a claim for co-speciation. The poorly supported relationships between clades A and B and Candidatus Riesia obscure their evolutionary history, although it is likely that clade A and B symbiont associations are independent events, and date back to a common ancestor in their respective host clades (Figure 3). Clade C includes type Arsenophonus sequences, symbiont sequences from Hippoboscidae, ticks, and other invertebrates (11), and an array of symbiont sequences from the geographically widespread Old World Cyclopodiinae, Nycteriboscinae, some Nycteribia, as well as New World Basilia (both Nycteribiinae) (Figure 1). The lack of well-defined subclusters in the network (Figure 1, 2) and poor support for subclade nodes
19 in the phylogenetic analyses (Figure 3) make inferences about specific relationships and symbiont acquisitions within this clade impossible at this time (e.g. Leptocyclopodia spp.). Some endosymbiont subclades seem to exhibit patterns of reciprocal monophyly on a bat fly genus level (i.e. Eucampsipoda spp., Figure 3). Arsenophonus is clearly a complex lineage, composed of a number of horizontally transmitted clades of unknown function with several vertically transmitted monophyletic clades restricted to specific groups of arthropods (11). Some bat fly associated Arsenophonus sequences show short branches (i.e. Brachytarsina sp., Megastrebla sp.), suggesting a recent, possibly horizontal acquisition, and possibly transient infections (Figure 3). Others, such as endosymbiont sequences associated with Eucampsipoda spp. have relatively longer branches possibly indicating an older association with this genus or more rapid endosymbiont evolution (Figure 3, Table 1). That heterogeneous Arsenophonus clades have repeatedly established endosymbiotic relationships with diverse groups of bat flies, to the apparent exclusion of other bacterial groups, suggests that Arsenophonus has characteristics making it a particularly successful invader and/or especially persistent once it has invaded. The type strain of Arsenophonus, A. nasonia, is a widespread parasite with a large genome and substantial metabolic capabilities (43), is able to transmit horizontally between species (44), and can be cultured in cell-free media. Other Arsenophonus strains (e.g. Arsenophonus arthropodicus in the louse fly Pseudolynchia canariensis) have been successfully cultured in insect cell lines (19). Furthermore, the A. nasonia genome encodes elements of the type III secretion system (45), commonly used by pathogenic bacteria to invade host cells. These characteristics may contribute to
20 repeated invasions of novel insect lineages, where, once established, Arsenophonus may persist, and sometimes adopt the characteristics of a mutualist. Clade D unites two novel subclades associated with the exclusively New World subfamily Nycterophiliinae, and the T. caecus group of Trichobiinae, respectively. The long branches dividing the subclades from each other suggest independent acquisition events of endosymbionts in ancestral host fly lineages (Figures 1, 3). It is notable that species belonging to the T. caecus group [including T. bilobus, T. caecus, T. galei, T. johnsonae, T. machadoallisoni, T. wenzeli, and T. yunkeri (46)] and Nycterophilia bat flies overlap substantially in host distribution, mostly occurring on ambient temperature and hot-cave-roosting bats in the families Natalidae and Mormoopidae, as well as some Phyllostomidae. The relatively close phylogenetic relationship between symbionts in these two host clades is more likely due to ecological shifts by their bat fly hosts to specialize on bats roosting in hot caves (41), rather than to host phylogeny, as there is currently no evidence of shared ancestry between the T. caecus group of Trichobius and Nycterophiliinae. Bacteriomes and vertical transmission of endosymbionts in bat flies. We detected endosymbionts in pupae and adults for both Nycterophilia (clade D) and Trichobius (clade B) bat flies. Based on the viviparous reproductive strategy of bat flies, which entails the intra-uterine development of the larva, and the deposition of a non-motile, non-feeding pupa (see introduction), the presence of endosymbiont DNA across developmental stages provides strong evidence for vertical endosymbiont transmission in clades B and D. This is further supported by the fact that pupae collected from glue traps (age 0-12 hrs), with no prior cave wall contact showed genetically identical
21 endosymbiont DNA to that of their maternal parents, as well as fluorescent signal (see FISH results). In addition, the same bat fly species collected from geographically distant, and ecologically diverse caves contain genetically highly similar endosymbiont DNA (e.g. Figure 3; T. intermedius), suggesting patterns of vertical inheritance, rather than de novo acquisition from the environment. This now supports vertical transmission for three of four major endosymbiont clades associated with bat flies (20). Previous studies on bat fly endosymbiont localization by in-situ hybridization concentrated on representative nycteribiid hosts of Candidatus Aschnera chinzeii (clade A). Specifically, both male and female Nycteribia and male Penicillidia bat flies showed loose clusters of bacteriocytes scattered in different locations of their abdominal cavities (20). In Nycteribia, this seems to be a general trend across species (20). In contrast, female Penicillidia were shown to house symbionts in the milk gland, and in a novel, paired bacteriome at the position of the lateral sclerotized plates of sternite 5. Based on these observations, Hosokawa, et al. (20) hypothesized the evolution of different symbiotic organs in closely related bat flies that were infected by the same, evolutionarily conserved endosymbiont. In representative streblid host flies of endosymbiont clades B (Trichobius) and D (Nycterophilia), however, results show singular bacteriomes, with no difference in localization between the sexes in Trichobius. Although no fluorescence was detected in male Nycterophilia flies, PCR detected endosymbionts in 100% of all males. Therefore, the negative results could be due to our small sample size of males [5] coupled with sample preservation problems, or to a low-level infection in males that is difficult to detect by in situ methods. The general location of the bacteriome in the Trichobius abdomen is similar to that observed in the related tsetse flies (Glossinidae), but unlike
22 Wigglesworthia glossinidia, the primary endosymbiont of Trichobius does not appear to reside in the epithelial lining of the midgut (9). This suggests that heterogeneous endosymbiont clades have differential cellular preferences in hosts. Wigglesworthia endosymbionts and Clade B symbionts in bat flies are not closely related (Figure 1), but may be functionally and immunologically similar. Although the location of the bacteriocytes in Nycterophilia flies is clearly associated with the crop, it remains unclear at this point if it is actually part of the crop (Figure 5). Currently, there are no examples of similar endosymbiont locations from other insects, and further studies with more specimens are necessary to resolve this issue. Biological function of endosymbionts in bat flies. The high prevalence of symbionts of clades A, B, and D in bat fly populations, their high AT-bias, the reciprocal monophyly and co-divergence of symbionts and fly hosts in clades and subclades, the localization of symbionts in bacteriomes, as well as their vertical transmission in these clades, all suggest an obligatory association of symbiont clades A, B, and D with their bat fly hosts. Based on their close phylogenetic relationship to Glossinidae and their similar bloodfeeding habits, it is possible that these heterogeneous symbiont clades also engage in a nutritional symbiosis, which, in a manner similar to the Wigglesworthia symbiont in tsetse flies, may influence bat fly longevity, digestion, productivity and vector competence (i.e., Bartonella) (9, 39). Furthermore, the high prevalence of infection in both males and females across clades A (20), B, and D suggest that these symbionts likely do not produce a male-killing phenotype, as has been identified in Arsenophonus nasoniae, although prevalence in both males and females does not preclude other forms of reproductive manipulation.
23 Clade C symbionts (Arsenophonus) may also be obligate symbionts in some bat flies, or, alternatively, may represent local, transient infections. The reciprocal monophyly of symbionts and species of Eucampsipoda may be an indication of evolutionarily stable associations, but this connection remains tenuous. Documented phenotypes for Arsenophonus in insects include son-killing (12, 47, 48), obligate mutualism (5, 18, 21), and presumably horizontally-transmitted associates with unknown but possibly mutualistic roles in plant-feeding insects such as whiteflies (14) and aphids (15), and blood-feeding arthropods such as ticks (49), triatomine bugs (7), and louse flies (6, 11, 19). This research expands our understanding of the evolution of endosymbiotic associations in bat flies. Because of a (still) limited sampling across the taxonomic diversity, geographic range and life history stages of bat flies, many aspects of symbiontbat fly associations remain to be discovered. Acknowledgements This research was supported by NSF Grants DEB , DEB , and DEB awarded to K.D., C.D., and B.P.; NSF SGER awarded to KD; and DEB awarded to S.B., R. Brown, D. Clayton, and R. Moyle. For field-work support, we thank the Field Museum s Council on Africa, the Barbara Brown Fund, IDP/FMNH African Training Fund, and Bud and Onnolee Trapp, as well as Armando Rodriguez, Megan L. Porter, Daniel Fong, John Jasper, Jack Wood, and Emily DiBlasi. The Zeiss LSM 710 In Tune Confocal Microscope used for FISH imaging was purchased through NSF Major Research Instrumentation grant DBI (University at Buffalo).
24 References Wenzel RL, Tipton VJ The streblid batflies of Panama (Diptera: Calypterae: Streblidae), p In Wenzel RL, Tipton VJ (ed.), Ectoparasites of Panama. Field Mus. Nat. Hist., Chicago. 2. Dick CW, Patterson BD Bat flies: obligate ectoparasites of bats, p In Morand S, Krasnov BR, Poulin R (ed.), Micromammals and Macroparasites. Springer. 3. Dittmar K, Porter ML, Murray S, Whiting MF Molecular phylogenetic analysis of nycteribiid and streblid bat flies (Diptera: Brachycera, Calyptratae): implications for host associations and phylogeographic origins. Mol Phylogenet Evol 38: Dergousoff SJ, Chilton NB Detection of a new Arsenophonus-type bacterium in Canadian populations of the Rocky Mountain wood tick, Dermacentor andersoni. Exp Appl Acarol 52: Perotti MA, Allen JM, Reed DL, Braig HR Host-symbiont interactions of the primary endosymbiont of human head and body lice. FASEB (Fed Am Soc Exp Biol) J 21: Trowbridge RE, Dittmar K, Whiting MF Identification and phylogenetic analysis of Arsenophonus- and Photorhabdus-type bacteria from adult Hippoboscidae and Streblidae (Hippoboscoidea). J Invertebr Pathol 91: Hypsa V, Dale C In vitro culture and phylogenetic analysis of "Candidatus Arsenophonus triatominarum," an intracellular bacterium from the triatomine bug, Triatoma infestans. Int J Syst Bacteriol 47: Hosokawa T, Koga R, Kikuchi Y, Meng X-Y, Fukatsu T Wolbachia as a bacteriocyte-associated nutritional mutualist. Proc Natl Acad Sci U S A 107: Pais R, Lohs C, Wu Y, Wang J, Aksoy S The obligate mutualist Wigglesworthia glossinidia influences reproduction, digestion, and immunity processes of its host, the tsetse fly. Appl Environ Microbiol 74: Lack JB, Nichols RD, Wilson GM, Van Den Bussche RA Genetic signature of reproductive manipulation in the phylogeography of the bat fly, Trichobius major. J Hered 102: Nováková E, Hypsa V, Moran NA Arsenophonus, an emerging clade of intracellular symbionts with a broad host distribution. BMC Microbiol 9: Gherna RL, Werren JH, Weisburg W, Cote R, Woese CR, Mandelco L, Brenner DJ Arsenophonus nasoniae gen. nov., sp. nov., the causative agent of the son-killer trait in the parasitic wasp Nasonia vitripennis. Int J Syst Evol Microbiol 41: Subandiyah S, Nikoh N, Tsuyumu S, Somowiyarjo S, Fukatsu T Complex endosymbiotic microbiota of the citrus psyllid Diaphorina citri (Homoptera: Psylloidea). Zool Sci (Tokyo) 17:
25 Thao ML, Baumann P Evidence for multiple acquisition of Arsenophonus by whitefly species (Sternorrhyncha: Aleyrodidae). Curr Microbiol 48: Russell JA, Latorre A, Sabater-Muñoz B, Moya A, Moran NA Sidestepping secondary symbionts: widespread horizontal transfer across and beyond the Aphidoidea. Mol Ecol 12: Bressan A, Sémétey O, Arneodo J, Lherminier J, Boudon-Padieu E Vector transmission of a plant-pathogenic bacterium in the Arsenophonus clade sharing ecological traits with facultative insect endosymbionts. Phytopathology 99: Clay K, Klyachko O, Grindle N, Civitello D, Oleske D, Fuqua C Microbial communities and interactions in the lone star tick, Amblyomma americanum. Mol Ecol 17: Allen JM, Reed DL, Perotti MA, Braig HR Evolutionary relationships of "Candidatus Riesia spp.," endosymbiotic enterobacteriaceae living within hematophagous primate lice. Appl Environ Microbiol 73: Dale C, Beeton M, Harbison C, Jones T, Pontes M Isolation, pure culture, and characterization of "Candidatus Arsenophonus arthropodicus," an intracellular secondary endosymbiont from the hippoboscid louse fly Pseudolynchia canariensis. Appl Environ Microbiol 72: Hosokawa T, Nikoh N, Koga R, Sato M, Tanahashi M, Meng XY, Fukatsu T Reductive genome evolution, host-symbiont co-speciation and uterine transmission of endosymbiotic bacteria in bat flies. ISME J 6: Sasaki-Fukatsu K, Koga R, Nikoh N, Yoshizawa K, Kasai S, Mihara M, Kobayashi M, Tomita T, Fukatsu T Symbiotic bacteria associated with stomach discs of human lice. Appl Environ Microbiol 72: Petersen FT, Meier R, Kutty SN, Wiegmann BM The phylogeny and evolution of host choice in the Hippoboscoidea (Diptera) as reconstructed using four molecular markers. Mol Phylogenet Evol 45: Maa T, Peterson B Hippoboscidae. In McAlpine J, Peterson B, Shewell G, Teskey H, Vockeroth J, Wood D (ed.), Manual of Nearctic Diptera (Volume 2). Research Branch, Agriculture Canada. 24. Fritz GN Biology and ecology of bat flies (Diptera: Streblidae) on bats in the genus Carollia. J Med Entomol 20: Overal WL Host-relations of the batfly, Megistopoda aranea (Diptera-- Streblidae) in Panamá. Univ Kansas Sci Bull 52: Attardo GM, Lohs C, Heddi A, Alam UH, Yildirim S, Aksoy S Analysis of milk gland structure and function in Glossina morsitans: milk protein production, symbiont populations and fecundity. J Insect Physiol 54: Dittmar K, Morse S, Gruwell M, Mayberry J, DiBlasi E Spatial and temporal complexities of reproductive behavior and sex ratios: a case from parasitic insects. PloS One 6:e Fukatsu T, Nikoh N Two intracellular symbiotic bacteria from the mulberry psyllid Anomoneura mori (Insecta, Homoptera). Appl Environ Microbiol 64:
26 Hosokawa T, Fukatsu T Nardonella endosymbiont in the West Indian sweet potato weevil Euscepes postfasciatus (Coleoptera: Curculionidae). Appl Entomol Zool 45: Katoh K, Kuma K-i, Toh H, Miyata T MAFFT version 5: improvement in accuracy of multiple sequence alignment. Nucleic Acids Res 33: Talavera G, Castresana J Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst Biol 56: Posada D jmodeltest: Phylogenetic model averaging. Mol Biol Evol 25: Salemi M, Vandamme A-M, Lemey P The phylogenetic handbook : a practical approach to phylogenetic analysis and hypothesis testing. Cambridge University Press, Cambridge, UK ; New York. 34. Huson DH, Bryant D Application of phylogenetic networks in evolutionary studies. Mol Biol Evol 23: Guindon S, Dufayard J-F, Lefort V, Anisimova M, Hordijk W, Gascuel O New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol 59: Ronquist F, Huelsenbeck JP MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19: Robinson-Rechavi M, Huchon D RRTree: relative-rate tests between groups of sequences on a phylogenetic tree. Bioinformatics 16: Koga R, Tsuchida T, Fukatsu T Quenching autofluorescence of insect tissues for in situ detection of endosymbionts. Appl Entomol Zool 44: Morse SF, Olival KJ, Kosoy M, Billeter S, Patterson BD, Dick CW, Dittmar K Global distribution and genetic diversity of Bartonella in bat flies (Hippoboscoidea, Streblidae, Nycteribiidae). Infect Genet Evol 12: Wenzel RL The streblid batflies of Venezuela (Diptera: Streblidae). Brigham Young University Science Bulletin-Biological Series 20: Morse S, Dick CW, Patterson BD, Dittmar K Some like it hot Evolution and ecology of novel endosymbionts in bat flies of cave-roosting bats (Hippoboscoidea, Nycterophiliinae). Appl Environ Microbiol 78: Moran NA, McCutcheon JP, Nakabachi A Genomics and evolution of heritable bacterial symbionts. Annu Rev Genet 42: Darby AC, Choi JH, Wilkes T, Hughes MA, Werren JH, Hurst GD, Colbourne JK Characteristics of the genome of Arsenophonus nasoniae, son-killer bacterium of the wasp Nasonia. Insect Mol Biol 19 Suppl 1: Duron O, Wilkes TE, Hurst GDD Interspecific transmission of a malekilling bacterium on an ecological timescale. Ecol Lett 13: Wilkes TE, Darby AC, Choi J-H, Colbourne JK, Werren JH, Hurst GDD The draft genome sequence of Arsenophonus nasoniae, son-killer bacterium of Nasonia vitripennis, reveals genes associated with virulence and symbiosis. Insect Molecular Biology 19 Suppl 1:59-73.
27 Dick CW, Graciolli G Checklist of world Streblidae (Diptera: Hippoboscoidea). 47. Ferree PM, Avery A, Azpurua J, Wilkes T, Werren JH A bacterium targets maternally inherited centrosomes to kill males in Nasonia. Curr Biol 18: Balas MT, Lee MH, Werren JH Distribution and fitness effects of the son-killer bacterium in Nasonia. Evol Ecol 10: Grindle N, Tyner JJ, Clay K, Fuqua C Identification of Arsenophonustype bacteria from the dog tick Dermacentor variabilis. J Invertebr Pathol 83: Downloaded from on October 23, 2018 by guest
28 Table Captions Table 1. Relative rate test for comparing the molecular evolutionary rates of grol gene sequences between the bat fly symbiont clades A+B and D with the Arsenophonus clade C. See text and Figures 1 or 3 for clade designations. Downloaded from on October 23, 2018 by guest
29 Figure Legends Figure 1. 16S rrna gene neighbor-net including sequences of nycteribiid and streblid bat fly endosymbionts, other insect symbionts, and various free-living bacteria. Bat fly symbiont clades are labeled A-D (see text). Figure 2. Inset of clade C (Arsenophonus) 16S rrna gene neighbor-net including sequences of nycteribiid and streblid bat fly endosymbionts (see text). Figure 3. Phylogenetic tree of bat fly endosymbionts in the gammaproteobacteria as inferred from concatenated 16S rrna and grol genes. A maximum likelihood tree (ML) is shown. Numbers at nodes represent bootstrap values/ Bayesian posterior probabilities (Bayesian topology not shown). Major bat fly symbiont subclades are labeled A-D (see text). Ancestral endosymbiont acquisition events are marked with circles on branches. Red: Streblidae. Blue: Nycteribiidae. Asterisks denote bacterial names; all other names refer to the host of the symbionts. Figure 4. Localization of endosymbiont signal in adult female Trichobius bat flies (Streblidae: Trichobiinae) by fluorescent in situ hybridization. Samples are DAPI stained. Cyan signals specify host insect nuclei, whereas red signals indicate the endosymbiont. (a) An external dorsal view of an adult female using transmitted light. (b) The same view as in (a) showing the bacteriome (red) in the dorsal anterior portion of the abdomen. (c) An enlarged view of the bacteriome showing individual bacteriocytes (red). (d) A 12-
30 day-old pupa removed from its pupal case. The specimen is facing right; developing wings can be seen above and below. The pupal bacteriome (red) is visible in the abdomen. (e) Luxol Fast Blue-stained section of an adult female abdomen showing the position of the bacteriome in relation to the midgut. The specimen is dorso-ventrally sectioned. Figure 5. Localization of the endosymbiont signal in adult female Nycterophilia bat fly (Streblidae: Nycterophiliinae) by fluorescent in situ hybridization. Samples are DAPI stained. Cyan signals specify host insect nuclei, whereas red signals indicate endosymbionts. (a) An external lateral view of an adult female using transmitted light. (b) The same view as in (a) showing the bacteriome (red) in the ventral anterior of the abdomen. (c) An enlarged view of the bacteriome (red) showing individual bacteriocytes. (d) A transmitted light image of a male specimen showing the position of abdominal structures.
31 Clade D Nycterophiliinae, Trichobiinae Clade A Nycteribiinae Candidatus Aschnera chinzei Clade B Streblinae, Trichobiinae Vibrio fischeri Candidatus Blochmannia ulcerosus Candidatus Kleidoceria schneideri Buchnera aphidicola Buchnera aphidicola Candidatus Ishikawaella capsulata Ishikawaella symbiont of Megacopta cribraria Enterobacteriaceae bacterium EpuTsukuba Downloaded from Streblidae Streblinae, Trichobiinae Nycteriboscinae Nycterophiliinae Nycteribiidae Nycteribiinae Cyclopodiinae Other Hippoboscoidea Hippoboscidae Glossinidae Other bacterial symbionts Arsenophonus sp. Candidatus Riesia pediculicola other insect symbionts Clade C (Arsenophonus) Providencia stuartii Providencia vermicola Proteus mirabilis Xenorhabdus kozodoii Photorhabdus asymbiotica Enterobacteriaceae bacterium SesTsukuba Photorhabdus luminescens Escherichia coli Candidatus Regiella insecticola Regiella symbiont of Sitobion miscanthi Midgut symbiont of Parastrachia japonensis Candidatus Regiella insecticola Candidatus Hamiltonella defensa Candidatus Hamiltonella defensa 0.01 on October 23, 2018 by guest
32 Clade C Arsenophonus Nycteriboscinae Brachytarsina Megastrebla Streblinae Anatrichobius Paradyschiria Cyclopodiinae Dipseliopoda Eucampsipoda Leptocyclopodia Nycteribiinae Basilia Nycteribia Other Symbionts Symbionts of Hippoboscidae Arsenophonus triatominarum Arsenophonus nasoniae
33 Streblidae Streblinae, Trichobiinae Nycteriboscinae Nycterophiliinae 1 0 0/100 Nycteribia pygmaea 1 0 0/100 Nycteribia pleuralis 9 6/ /97 Nycteribiidae Nycteribiinae Cyclopodiinae Other Hippoboscoidea Hippoboscidae 7 0/96 * /98 * / /100 Nycteribia allotopa 1 0 0/95 Phthiridium cf. tonkinensis [JAE1224] 1 0 0/100 Phthiridium sp. 2 [P220] Phthiridium sp. 3 [P479] 1 0 0/100 Phthiridium hindlei / / C D 8 1/100 B / A Candidatus Aschnera chinzeii 1 0 0/100 Basilia truncata 1 0 0/100 Basilia nattereri [ZAG03] 1 0 0/100 Basilia rybini 9 4/ / /100 Penicillidia jenynsii Penicillidia oceanica [JAE978] Penicillidia monoceros 1 0 0/100 Penicillidia dufourii [CHI05] 1 0 0/100 Candidatus Riesia pediculicola * 8 8/* Trichobius intermedius [PSU1303] 8 1/* Trichobius intermedius [MEX06] Trichobius intermedius [MEX10] 9 8/100 Trichobius intermedius [EHA49-T] 9 8/100 Trichobius intermedius [DR05076] Aspidoptera falcata [TK135285] / * Trichobius parasiticus [MEX2009F1] / Trichobius parasiticus [MEX08] Trichobius parasiticus [MEX05] 9 6 / / / Trichobius neotropicus [DR ] 9 5 / Trichobius neotropicus [DR ] Trichobius dugesoides [MN114] 7 7 / / Megistopoda aranea [EHA49-M] Paratrichobius sp. [FG10] Strebla diphyllae [MEX15.1] / Trichobius frequens [SM ] / Trichobius frequens [BFPaperpupa1] / Trichobius frequens [SM ] Trichobius cf. cernyi [KD ] 9 4 / / 9 5 Trichobius sp. 3 [MEX2009E3] Trichobius major [CWD998] 9 2 / Leptocyclopodia n. sp. 1 [JAE3270] Leptocyclopodia n. sp. 1 [JAE1033] 9 9 / 9 4 Brachytarsina sp. 1 [P4635] Megastrebla sp. [P4511.2] 9 6 / * Ornithomyia fringillina [MCP ] 8 7 / Arsenophonus nasoniae * Leptocyclopodia ferrarii ssp. [P414] Leptocyclopodia brevicula [P2466] 8 3 / Basilia boardmani [JAW05] Arsenophonus triatominarum * * / Lipoptena cervi [DK] / Lipoptena depressa [WSPA2000] / Eucampsipoda cf. latisterna [P639] 7 0 / * Eucampsipoda cf. latisterna [P636] / Eucampsipoda africana [BDP4416] 8 2 / Eucampsipoda inermis [P4496] Eucampsipoda inermis [JAE514] Nycterophilia cf. coxata [FE21] Nycterophilia coxata [DR05256] 8 5 / * Nycterophilia coxata [MEX2009G3] / Nycterophilia n. sp. [DR ] Nycterophilia parnelli [MEX2009A4.2] Trichobius n. sp. 1 [MEX17] / Trichobius cf. yunkeri [FG08] Trichobius n. sp. 2 [MEX02] Providencia stuartii * Photorhabdus asymbiotica * Proteus mirabilis * Trichobius caecus species group Trichobius dugesii species group Trichobius major species group 0.02



34 b c midgut e d wings abdomen bacteriome a




35 a b Downloaded from c d gut crop on October 23, 2018 by guest
Lecture 11 Wednesday, September 19, 2012
 Lecture 11 Wednesday, September 19, 2012 Phylogenetic tree (phylogeny) Darwin and classification: In the Origin, Darwin said that descent from a common ancestral species could explain why the Linnaean
Lecture 11 Wednesday, September 19, 2012 Phylogenetic tree (phylogeny) Darwin and classification: In the Origin, Darwin said that descent from a common ancestral species could explain why the Linnaean
David A Wilkinson, Olivier Duron, Colette Cordonin, Yann Gomard, Beza Ramasindrazana, Patrick Mavingui, Steven M Goodman, Pablo Tortosa
 The bacteriome of bat flies (Nycteribiidae) from the Malagasy region: a community shaped by host ecology, bacterial transmission mode, and host-vector specificity. David A Wilkinson, Olivier Duron, Colette
The bacteriome of bat flies (Nycteribiidae) from the Malagasy region: a community shaped by host ecology, bacterial transmission mode, and host-vector specificity. David A Wilkinson, Olivier Duron, Colette
CLADISTICS Student Packet SUMMARY Phylogeny Phylogenetic trees/cladograms
 CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes)
") Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
Introduction to phylogenetic trees and tree-thinking Copyright 2005, D. A. Baum (Free use for non-commercial educational pruposes) Phylogenetics is the study of the relationships of organisms to each other.
Research Note. A novel method for sexing day-old chicks using endoscope system
 Research Note A novel method for sexing day-old chicks using endoscope system Makoto Otsuka,,1 Osamu Miyashita,,1 Mitsuru Shibata,,1 Fujiyuki Sato,,1 and Mitsuru Naito,2,3 NARO Institute of Livestock and
Research Note A novel method for sexing day-old chicks using endoscope system Makoto Otsuka,,1 Osamu Miyashita,,1 Mitsuru Shibata,,1 Fujiyuki Sato,,1 and Mitsuru Naito,2,3 NARO Institute of Livestock and
The effects of diet upon pupal development and cocoon formation by the cat flea (Siphonaptera: Pulicidae)
") June, 2002 Journal of Vector Ecology 39 The effects of diet upon pupal development and cocoon formation by the cat flea (Siphonaptera: Pulicidae) W. Lawrence and L. D. Foil Department of Entomology, Louisiana
June, 2002 Journal of Vector Ecology 39 The effects of diet upon pupal development and cocoon formation by the cat flea (Siphonaptera: Pulicidae) W. Lawrence and L. D. Foil Department of Entomology, Louisiana
Insect vectors. Dr. Carmen E. Rexach Micro 1 Mt SAC Biology Department Internet version
 Insect vectors Dr. Carmen E. Rexach Micro 1 Mt SAC Biology Department Internet version Biological vs mechanical transmission Mechanical Pathogen is picked up from a source and deposited on another location
Insect vectors Dr. Carmen E. Rexach Micro 1 Mt SAC Biology Department Internet version Biological vs mechanical transmission Mechanical Pathogen is picked up from a source and deposited on another location
Comparing DNA Sequences to Understand Evolutionary Relationships with BLAST
 Comparing DNA Sequences to Understand Evolutionary Relationships with BLAST INVESTIGATION 3 BIG IDEA 1 Lab Investigation 3: BLAST Pre-Lab Essential Question: How can bioinformatics be used as a tool to
Comparing DNA Sequences to Understand Evolutionary Relationships with BLAST INVESTIGATION 3 BIG IDEA 1 Lab Investigation 3: BLAST Pre-Lab Essential Question: How can bioinformatics be used as a tool to
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
The Making of the Fittest: LESSON STUDENT MATERIALS USING DNA TO EXPLORE LIZARD PHYLOGENY
 The Making of the Fittest: Natural The The Making Origin Selection of the of Species and Fittest: Adaptation Natural Lizards Selection in an Evolutionary and Adaptation Tree INTRODUCTION USING DNA TO EXPLORE
The Making of the Fittest: Natural The The Making Origin Selection of the of Species and Fittest: Adaptation Natural Lizards Selection in an Evolutionary and Adaptation Tree INTRODUCTION USING DNA TO EXPLORE
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A.
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Methicillin-Resistant Staphylococcus aureus
 Methicillin-Resistant Staphylococcus aureus By Karla Givens Means of Transmission and Usual Reservoirs Staphylococcus aureus is part of normal flora and can be found on the skin and in the noses of one
Methicillin-Resistant Staphylococcus aureus By Karla Givens Means of Transmission and Usual Reservoirs Staphylococcus aureus is part of normal flora and can be found on the skin and in the noses of one
A comparison of placental tissue in the skinks Eulamprus tympanum and E. quoyii. Yates, Lauren A.
 A comparison of placental tissue in the skinks Eulamprus tympanum and E. quoyii Yates, Lauren A. Abstract: The species Eulamprus tympanum and Eulamprus quoyii are viviparous skinks that are said to have
A comparison of placental tissue in the skinks Eulamprus tympanum and E. quoyii Yates, Lauren A. Abstract: The species Eulamprus tympanum and Eulamprus quoyii are viviparous skinks that are said to have
GEODIS 2.0 DOCUMENTATION
 GEODIS.0 DOCUMENTATION 1999-000 David Posada and Alan Templeton Contact: David Posada, Department of Zoology, 574 WIDB, Provo, UT 8460-555, USA Fax: (801) 78 74 e-mail: dp47@email.byu.edu 1. INTRODUCTION
GEODIS.0 DOCUMENTATION 1999-000 David Posada and Alan Templeton Contact: David Posada, Department of Zoology, 574 WIDB, Provo, UT 8460-555, USA Fax: (801) 78 74 e-mail: dp47@email.byu.edu 1. INTRODUCTION
PCR detection of Leptospira in. stray cat and
 PCR detection of Leptospira in 1 Department of Pathology, School of Veterinary Medicine, Islamic Azad University, Shahrekord Branch, Shahrekord, Iran 2 Department of Microbiology, School of Veterinary
PCR detection of Leptospira in 1 Department of Pathology, School of Veterinary Medicine, Islamic Azad University, Shahrekord Branch, Shahrekord, Iran 2 Department of Microbiology, School of Veterinary
Ch 1.2 Determining How Species Are Related.notebook February 06, 2018
 Name 3 "Big Ideas" from our last notebook lecture: * * * 1 WDYR? Of the following organisms, which is the closest relative of the "Snowy Owl" (Bubo scandiacus)? a) barn owl (Tyto alba) b) saw whet owl
Name 3 "Big Ideas" from our last notebook lecture: * * * 1 WDYR? Of the following organisms, which is the closest relative of the "Snowy Owl" (Bubo scandiacus)? a) barn owl (Tyto alba) b) saw whet owl
Supplemental Information. Discovery of Reactive Microbiota-Derived. Metabolites that Inhibit Host Proteases
 Cell, Volume 168 Supplemental Information Discovery of Reactive Microbiota-Derived Metabolites that Inhibit Host Proteases Chun-Jun Guo, Fang-Yuan Chang, Thomas P. Wyche, Keriann M. Backus, Timothy M.
Cell, Volume 168 Supplemental Information Discovery of Reactive Microbiota-Derived Metabolites that Inhibit Host Proteases Chun-Jun Guo, Fang-Yuan Chang, Thomas P. Wyche, Keriann M. Backus, Timothy M.
History of Lineages. Chapter 11. Jamie Oaks 1. April 11, Kincaid Hall 524. c 2007 Boris Kulikov boris-kulikov.blogspot.
 History of Lineages Chapter 11 Jamie Oaks 1 1 Kincaid Hall 524 joaks1@gmail.com April 11, 2014 c 2007 Boris Kulikov boris-kulikov.blogspot.com History of Lineages J. Oaks, University of Washington 1/46
History of Lineages Chapter 11 Jamie Oaks 1 1 Kincaid Hall 524 joaks1@gmail.com April 11, 2014 c 2007 Boris Kulikov boris-kulikov.blogspot.com History of Lineages J. Oaks, University of Washington 1/46
RICKETTSIA SPECIES AMONG TICKS IN AN AREA OF JAPAN ENDEMIC FOR JAPANESE SPOTTED FEVER
 RICKETTSIA SPECIES AMONG TICKS IN AN AREA OF JAPAN ENDEMIC FOR JAPANESE SPOTTED FEVER Makoto Kondo 1, Katsuhiko Ando 2, Keiichi Yamanaka 1 and Hitoshi Mizutani 1 1 Department of Dermatology, 2 Department
RICKETTSIA SPECIES AMONG TICKS IN AN AREA OF JAPAN ENDEMIC FOR JAPANESE SPOTTED FEVER Makoto Kondo 1, Katsuhiko Ando 2, Keiichi Yamanaka 1 and Hitoshi Mizutani 1 1 Department of Dermatology, 2 Department
Laboratory 7 The Effect of Juvenile Hormone on Metamorphosis of the Fruit Fly (Drosophila melanogaster)
") Laboratory 7 The Effect of Juvenile Hormone on Metamorphosis of the Fruit Fly (Drosophila melanogaster) (portions of this manual were borrowed from Prof. Douglas Facey, Department of Biology, Saint Michael's
Laboratory 7 The Effect of Juvenile Hormone on Metamorphosis of the Fruit Fly (Drosophila melanogaster) (portions of this manual were borrowed from Prof. Douglas Facey, Department of Biology, Saint Michael's
Bi156 Lecture 1/13/12. Dog Genetics
 Bi156 Lecture 1/13/12 Dog Genetics The radiation of the family Canidae occurred about 100 million years ago. Dogs are most closely related to wolves, from which they diverged through domestication about
Bi156 Lecture 1/13/12 Dog Genetics The radiation of the family Canidae occurred about 100 million years ago. Dogs are most closely related to wolves, from which they diverged through domestication about
Evolutionary Trade-Offs in Mammalian Sensory Perceptions: Visual Pathways of Bats. By Adam Proctor Mentor: Dr. Emma Teeling
 Evolutionary Trade-Offs in Mammalian Sensory Perceptions: Visual Pathways of Bats By Adam Proctor Mentor: Dr. Emma Teeling Visual Pathways of Bats Purpose Background on mammalian vision Tradeoffs and bats
Evolutionary Trade-Offs in Mammalian Sensory Perceptions: Visual Pathways of Bats By Adam Proctor Mentor: Dr. Emma Teeling Visual Pathways of Bats Purpose Background on mammalian vision Tradeoffs and bats
Bacterial whole genome sequencing in clinical microbiology, infection control and public health. Julian Parkhill. FIS, Birmingham, November 2013
 Bacterial whole genome sequencing in clinical microbiology, infection control and public health Julian Parkhill FIS, Birmingham, November 2013 Falling costs of genomics 2003 Cost/genome Throughput 60,000
Bacterial whole genome sequencing in clinical microbiology, infection control and public health Julian Parkhill FIS, Birmingham, November 2013 Falling costs of genomics 2003 Cost/genome Throughput 60,000
Why Don t These Drugs Work Anymore? Biosciences in the 21 st Century Dr. Amber Rice October 28, 2013
 Why Don t These Drugs Work Anymore? Biosciences in the 21 st Century Dr. Amber Rice October 28, 2013 Outline Drug resistance: a case study Evolution: the basics How does resistance evolve? Examples of
Why Don t These Drugs Work Anymore? Biosciences in the 21 st Century Dr. Amber Rice October 28, 2013 Outline Drug resistance: a case study Evolution: the basics How does resistance evolve? Examples of
A Mitochondrial DNA Phylogeny of Extant Species of the Genus Trachemys with Resulting Taxonomic Implications
 NOTES AND FIELD REPORTS 131 Chelonian Conservation and Biology, 2008, 7(1): 131 135 Ó 2008 Chelonian Research Foundation A Mitochondrial DNA Phylogeny of Extant Species of the Genus Trachemys with Resulting
NOTES AND FIELD REPORTS 131 Chelonian Conservation and Biology, 2008, 7(1): 131 135 Ó 2008 Chelonian Research Foundation A Mitochondrial DNA Phylogeny of Extant Species of the Genus Trachemys with Resulting
UNIT III A. Descent with Modification(Ch19) B. Phylogeny (Ch20) C. Evolution of Populations (Ch21) D. Origin of Species or Speciation (Ch22)
 B. Phylogeny (Ch20) C. Evolution of Populations (Ch21) D. Origin of Species or Speciation (Ch22)") UNIT III A. Descent with Modification(Ch9) B. Phylogeny (Ch2) C. Evolution of Populations (Ch2) D. Origin of Species or Speciation (Ch22) Classification in broad term simply means putting things in classes
UNIT III A. Descent with Modification(Ch9) B. Phylogeny (Ch2) C. Evolution of Populations (Ch2) D. Origin of Species or Speciation (Ch22) Classification in broad term simply means putting things in classes
Species: Panthera pardus Genus: Panthera Family: Felidae Order: Carnivora Class: Mammalia Phylum: Chordata
 CHAPTER 6: PHYLOGENY AND THE TREE OF LIFE AP Biology 3 PHYLOGENY AND SYSTEMATICS Phylogeny - evolutionary history of a species or group of related species Systematics - analytical approach to understanding
CHAPTER 6: PHYLOGENY AND THE TREE OF LIFE AP Biology 3 PHYLOGENY AND SYSTEMATICS Phylogeny - evolutionary history of a species or group of related species Systematics - analytical approach to understanding
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1
 Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
Gliding Motility Assay for P. berghei Sporozoites
 Gliding Motility Assay for P. berghei Sporozoites Important Notes: 1. For all dilutions (including antibodies and sporozoites), always make slightly more than needed. For instance, if you need 200 µl sporozoites
Gliding Motility Assay for P. berghei Sporozoites Important Notes: 1. For all dilutions (including antibodies and sporozoites), always make slightly more than needed. For instance, if you need 200 µl sporozoites
Modern Evolutionary Classification. Lesson Overview. Lesson Overview Modern Evolutionary Classification
 Lesson Overview 18.2 Modern Evolutionary Classification THINK ABOUT IT Darwin s ideas about a tree of life suggested a new way to classify organisms not just based on similarities and differences, but
Lesson Overview 18.2 Modern Evolutionary Classification THINK ABOUT IT Darwin s ideas about a tree of life suggested a new way to classify organisms not just based on similarities and differences, but
Phylogeny Reconstruction
 Phylogeny Reconstruction Trees, Methods and Characters Reading: Gregory, 2008. Understanding Evolutionary Trees (Polly, 2006) Lab tomorrow Meet in Geology GY522 Bring computers if you have them (they will
Phylogeny Reconstruction Trees, Methods and Characters Reading: Gregory, 2008. Understanding Evolutionary Trees (Polly, 2006) Lab tomorrow Meet in Geology GY522 Bring computers if you have them (they will
Bioinformatics: Investigating Molecular/Biochemical Evidence for Evolution
 Bioinformatics: Investigating Molecular/Biochemical Evidence for Evolution Background How does an evolutionary biologist decide how closely related two different species are? The simplest way is to compare
Bioinformatics: Investigating Molecular/Biochemical Evidence for Evolution Background How does an evolutionary biologist decide how closely related two different species are? The simplest way is to compare
Testing Phylogenetic Hypotheses with Molecular Data 1
 Testing Phylogenetic Hypotheses with Molecular Data 1 How does an evolutionary biologist quantify the timing and pathways for diversification (speciation)? If we observe diversification today, the processes
Testing Phylogenetic Hypotheses with Molecular Data 1 How does an evolutionary biologist quantify the timing and pathways for diversification (speciation)? If we observe diversification today, the processes
The Evolution of Human-Biting Preference in Mosquitoes
 Got Blood? The Evolution of Human-Biting Preference in Mosquitoes by Gary H. Laverty Department of Biological Sciences University of Delaware, Newark, DE Part I A Matter of Preference So, what do we do
Got Blood? The Evolution of Human-Biting Preference in Mosquitoes by Gary H. Laverty Department of Biological Sciences University of Delaware, Newark, DE Part I A Matter of Preference So, what do we do
Evolution in dogs. Megan Elmore CS374 11/16/2010. (thanks to Dan Newburger for many slides' content)
") Evolution in dogs Megan Elmore CS374 11/16/2010 (thanks to Dan Newburger for many slides' content) Papers for today Vonholdt BM et al (2010). Genome-wide SNP and haplotype analyses reveal a rich history
Evolution in dogs Megan Elmore CS374 11/16/2010 (thanks to Dan Newburger for many slides' content) Papers for today Vonholdt BM et al (2010). Genome-wide SNP and haplotype analyses reveal a rich history
Cladistics (reading and making of cladograms)
") Cladistics (reading and making of cladograms) Definitions Systematics The branch of biological sciences concerned with classifying organisms Taxon (pl: taxa) Any unit of biological diversity (eg. Animalia,
Cladistics (reading and making of cladograms) Definitions Systematics The branch of biological sciences concerned with classifying organisms Taxon (pl: taxa) Any unit of biological diversity (eg. Animalia,
ERG on multidrug-resistant P. falciparum in the GMS
 ERG on multidrug-resistant P. falciparum in the GMS Minutes of ERG meeting Presented by D. Wirth, Chair of the ERG Geneva, 22-24 March 2017 MPAC meeting Background At the Malaria Policy Advisory Committee
ERG on multidrug-resistant P. falciparum in the GMS Minutes of ERG meeting Presented by D. Wirth, Chair of the ERG Geneva, 22-24 March 2017 MPAC meeting Background At the Malaria Policy Advisory Committee
6. The lifetime Darwinian fitness of one organism is greater than that of another organism if: A. it lives longer than the other B. it is able to outc
 1. The money in the kingdom of Florin consists of bills with the value written on the front, and pictures of members of the royal family on the back. To test the hypothesis that all of the Florinese $5
1. The money in the kingdom of Florin consists of bills with the value written on the front, and pictures of members of the royal family on the back. To test the hypothesis that all of the Florinese $5
Development and improvement of diagnostics to improve use of antibiotics and alternatives to antibiotics
 Priority Topic B Diagnostics Development and improvement of diagnostics to improve use of antibiotics and alternatives to antibiotics The overarching goal of this priority topic is to stimulate the design,
Priority Topic B Diagnostics Development and improvement of diagnostics to improve use of antibiotics and alternatives to antibiotics The overarching goal of this priority topic is to stimulate the design,
Guidelines for Laboratory Verification of Performance of the FilmArray BCID System
 Guidelines for Laboratory Verification of Performance of the FilmArray BCID System Purpose The Clinical Laboratory Improvement Amendments (CLIA), passed in 1988, establishes quality standards for all laboratory
Guidelines for Laboratory Verification of Performance of the FilmArray BCID System Purpose The Clinical Laboratory Improvement Amendments (CLIA), passed in 1988, establishes quality standards for all laboratory
1 In 1958, scientists made a breakthrough in artificial reproductive cloning by successfully cloning a
 1 In 1958, scientists made a breakthrough in artificial reproductive cloning by successfully cloning a vertebrate species. The species cloned was the African clawed frog, Xenopus laevis. Fig. 1.1, on page
1 In 1958, scientists made a breakthrough in artificial reproductive cloning by successfully cloning a vertebrate species. The species cloned was the African clawed frog, Xenopus laevis. Fig. 1.1, on page
Phylogeographic assessment of Acanthodactylus boskianus (Reptilia: Lacertidae) based on phylogenetic analysis of mitochondrial DNA.
 based on phylogenetic analysis of mitochondrial DNA.") Zoology Department Phylogeographic assessment of Acanthodactylus boskianus (Reptilia: Lacertidae) based on phylogenetic analysis of mitochondrial DNA By HAGAR IBRAHIM HOSNI BAYOUMI A thesis submitted in
Zoology Department Phylogeographic assessment of Acanthodactylus boskianus (Reptilia: Lacertidae) based on phylogenetic analysis of mitochondrial DNA By HAGAR IBRAHIM HOSNI BAYOUMI A thesis submitted in
TOPIC CLADISTICS
 TOPIC 5.4 - CLADISTICS 5.4 A Clades & Cladograms https://upload.wikimedia.org/wikipedia/commons/thumb/4/46/clade-grade_ii.svg IB BIO 5.4 3 U1: A clade is a group of organisms that have evolved from a common
TOPIC 5.4 - CLADISTICS 5.4 A Clades & Cladograms https://upload.wikimedia.org/wikipedia/commons/thumb/4/46/clade-grade_ii.svg IB BIO 5.4 3 U1: A clade is a group of organisms that have evolved from a common
A Unique Approach to Managing the Problem of Antibiotic Resistance
 A Unique Approach to Managing the Problem of Antibiotic Resistance By: Heather Storteboom and Sung-Chul Kim Department of Civil and Environmental Engineering Colorado State University A Quick Review The
A Unique Approach to Managing the Problem of Antibiotic Resistance By: Heather Storteboom and Sung-Chul Kim Department of Civil and Environmental Engineering Colorado State University A Quick Review The
Your web browser (Safari 7) is out of date. For more security, comfort and the best experience on this site: Update your browser Ignore
 is out of date. For more security, comfort and the best experience on this site: Update your browser Ignore") Your web browser (Safari 7) is out of date. For more security, comfort and the best experience on this site: Update your browser Ignore Activitydevelop EXPLO RING VERTEBRATE CL ASSIFICATIO N What criteria
Your web browser (Safari 7) is out of date. For more security, comfort and the best experience on this site: Update your browser Ignore Activitydevelop EXPLO RING VERTEBRATE CL ASSIFICATIO N What criteria
ASVCP quality assurance guidelines: veterinary immunocytochemistry (ICC)
") ASVCP quality assurance guidelines: veterinary immunocytochemistry (ICC) Version 1.0 (Approved 11/2017) Developed by the American Society for Veterinary Clinical Pathology (ASVCP) Quality Assurance and
ASVCP quality assurance guidelines: veterinary immunocytochemistry (ICC) Version 1.0 (Approved 11/2017) Developed by the American Society for Veterinary Clinical Pathology (ASVCP) Quality Assurance and
ANTIBIOTIC SENSITIVITY PATTERN OF YERSINIA ENTEROCOLITICA ISOLATED FROM MILK AND DAIRY PRODUCTS*
 Short Communication ANTIBIOTIC SENSITIVITY PATTERN OF YERSINIA ENTEROCOLITICA ISOLATED FROM MILK AND DAIRY PRODUCTS* T.R.Pugazhenthi 1, A. Elango 2, C. Naresh Kumar 3, B. Dhanalakshmi 4 and A. Bharathidhasan
Short Communication ANTIBIOTIC SENSITIVITY PATTERN OF YERSINIA ENTEROCOLITICA ISOLATED FROM MILK AND DAIRY PRODUCTS* T.R.Pugazhenthi 1, A. Elango 2, C. Naresh Kumar 3, B. Dhanalakshmi 4 and A. Bharathidhasan
26. The Relationships between Oxygen Consumption and Duration o f Pupal-Adult Development in the Silkworm Bombyx mandarina
 134 Proc. Japan Acad., 69, Ser. B (1993) [Vol. 69(B), 26. The Relationships between Oxygen Consumption and Duration o f Pupal-Adult Development in the Silkworm Bombyx mandarina By Weide SHEN and Kunikatsu
134 Proc. Japan Acad., 69, Ser. B (1993) [Vol. 69(B), 26. The Relationships between Oxygen Consumption and Duration o f Pupal-Adult Development in the Silkworm Bombyx mandarina By Weide SHEN and Kunikatsu
Evolution. Evolution is change in organisms over time. Evolution does not have a goal; it is often shaped by natural selection (see below).
.") Evolution Evolution is change in organisms over time. Evolution does not have a goal; it is often shaped by natural selection (see below). Species an interbreeding population of organisms that can produce
Evolution Evolution is change in organisms over time. Evolution does not have a goal; it is often shaped by natural selection (see below). Species an interbreeding population of organisms that can produce
Application of sewage in pisciculture in order to augment fish production has been an
 Conclusions Application of sewage in pisciculture in order to augment fish production has been an ancient practice in India and other countries like i.e. China, Egypt and Europe. Possible health hazard
Conclusions Application of sewage in pisciculture in order to augment fish production has been an ancient practice in India and other countries like i.e. China, Egypt and Europe. Possible health hazard
Martin Chénier, Ph.D. Microbiology. Antibiotics in Animal Production: Resistance and Alternative Solutions
 Faculty of Agricultural and Environmental Sciences Department of Food Science, Department of Animal Science Martin Chénier, Ph.D. Microbiology Antibiotics in Animal Production: Resistance and Alternative
Faculty of Agricultural and Environmental Sciences Department of Food Science, Department of Animal Science Martin Chénier, Ph.D. Microbiology Antibiotics in Animal Production: Resistance and Alternative
Reintroducing bettongs to the ACT: issues relating to genetic diversity and population dynamics The guest speaker at NPA s November meeting was April
 Reintroducing bettongs to the ACT: issues relating to genetic diversity and population dynamics The guest speaker at NPA s November meeting was April Suen, holder of NPA s 2015 scholarship for honours
Reintroducing bettongs to the ACT: issues relating to genetic diversity and population dynamics The guest speaker at NPA s November meeting was April Suen, holder of NPA s 2015 scholarship for honours
Ecology of RMSF on Arizona Tribal Lands
 Ecology of RMSF on Arizona Tribal Lands Tribal Vector Borne Disease Meeting M. L. Levin Ph.D. Medical Entomology Laboratory Centers for Disease Control mlevin@cdc.gov Rocky Mountain Spotted Fever Disease
Ecology of RMSF on Arizona Tribal Lands Tribal Vector Borne Disease Meeting M. L. Levin Ph.D. Medical Entomology Laboratory Centers for Disease Control mlevin@cdc.gov Rocky Mountain Spotted Fever Disease
Resolving Confusion in the Use of Concepts and Terminology in Intra-Puparial
 1 Martín-Vega et al.: Terminology in 8 Daniel Martín-Vega 2 intra-puparial development studies 9 Department of Life Sciences 3 10 11 Natural History Museum Cromwell Road 4 Journal of Medical Entomology
1 Martín-Vega et al.: Terminology in 8 Daniel Martín-Vega 2 intra-puparial development studies 9 Department of Life Sciences 3 10 11 Natural History Museum Cromwell Road 4 Journal of Medical Entomology
INHERITANCE OF BODY WEIGHT IN DOMESTIC FOWL. Single Comb White Leghorn breeds of fowl and in their hybrids.
 440 GENETICS: N. F. WATERS PROC. N. A. S. and genetical behavior of this form is not incompatible with the segmental interchange theory of circle formation in Oenothera. Summary.-It is impossible for the
440 GENETICS: N. F. WATERS PROC. N. A. S. and genetical behavior of this form is not incompatible with the segmental interchange theory of circle formation in Oenothera. Summary.-It is impossible for the
Bio 1B Lecture Outline (please print and bring along) Fall, 2006
 Fall, 2006") Bio 1B Lecture Outline (please print and bring along) Fall, 2006 B.D. Mishler, Dept. of Integrative Biology 2-6810, bmishler@berkeley.edu Evolution lecture #4 -- Phylogenetic Analysis (Cladistics) -- Oct.
Bio 1B Lecture Outline (please print and bring along) Fall, 2006 B.D. Mishler, Dept. of Integrative Biology 2-6810, bmishler@berkeley.edu Evolution lecture #4 -- Phylogenetic Analysis (Cladistics) -- Oct.
ESCHERICHIA COLI RESISTANCE AND GUT MICROBIOTA PROFILE IN PIGS RAISED WITH DIFFERENT ANTIMICROBIAL ADMINISTRATION IN FEED
 ESCHERICHIA COLI RESISTANCE AND GUT MICROBIOTA PROFILE IN PIGS RAISED WITH DIFFERENT ANTIMICROBIAL ADMINISTRATION IN FEED Caroline Pissetti 1, Jalusa Deon Kich 2, Heather K. Allen 3, Claudia Navarrete
ESCHERICHIA COLI RESISTANCE AND GUT MICROBIOTA PROFILE IN PIGS RAISED WITH DIFFERENT ANTIMICROBIAL ADMINISTRATION IN FEED Caroline Pissetti 1, Jalusa Deon Kich 2, Heather K. Allen 3, Claudia Navarrete
Prof. Neil. J.L. Heideman
 Prof. Neil. J.L. Heideman Position Office Mailing address E-mail : Vice-dean (Professor of Zoology) : No. 10, Biology Building : P.O. Box 339 (Internal Box 44), Bloemfontein 9300, South Africa : heidemannj.sci@mail.uovs.ac.za
Prof. Neil. J.L. Heideman Position Office Mailing address E-mail : Vice-dean (Professor of Zoology) : No. 10, Biology Building : P.O. Box 339 (Internal Box 44), Bloemfontein 9300, South Africa : heidemannj.sci@mail.uovs.ac.za
Fig Phylogeny & Systematics
 Fig. 26- Phylogeny & Systematics Tree of Life phylogenetic relationship for 3 clades (http://evolution.berkeley.edu Fig. 26-2 Phylogenetic tree Figure 26.3 Taxonomy Taxon Carolus Linnaeus Species: Panthera
Fig. 26- Phylogeny & Systematics Tree of Life phylogenetic relationship for 3 clades (http://evolution.berkeley.edu Fig. 26-2 Phylogenetic tree Figure 26.3 Taxonomy Taxon Carolus Linnaeus Species: Panthera
soft ticks hard ticks
 Ticks Family Argasidae soft ticks Only 4 genera of Argasidae Argas, Ornithodoros, Otobius (not covered) and Carios (not covered) Family Ixodidae hard ticks Only 4 genera of Ixodidae covered because of
Ticks Family Argasidae soft ticks Only 4 genera of Argasidae Argas, Ornithodoros, Otobius (not covered) and Carios (not covered) Family Ixodidae hard ticks Only 4 genera of Ixodidae covered because of
17.2 Classification Based on Evolutionary Relationships Organization of all that speciation!
 Organization of all that speciation! Patterns of evolution.. Taxonomy gets an over haul! Using more than morphology! 3 domains, 6 kingdoms KEY CONCEPT Modern classification is based on evolutionary relationships.
Organization of all that speciation! Patterns of evolution.. Taxonomy gets an over haul! Using more than morphology! 3 domains, 6 kingdoms KEY CONCEPT Modern classification is based on evolutionary relationships.
Understanding and prevention of transmission of antibiotic resistance between bacterial populations and One Health reservoirs
 Priority Topic D - Transmission Understanding and prevention of transmission of antibiotic resistance between bacterial populations and One Health reservoirs The overarching goal of this priority topic
Priority Topic D - Transmission Understanding and prevention of transmission of antibiotic resistance between bacterial populations and One Health reservoirs The overarching goal of this priority topic
How to load and run an Agarose gel PSR
 How to load and run an Agarose gel PSR Agarose gel electrophoresis is the most effective way of separating DNA fragments of varying sizes ranging from100 bp to 25 kb. This protocol divided into three stages:
How to load and run an Agarose gel PSR Agarose gel electrophoresis is the most effective way of separating DNA fragments of varying sizes ranging from100 bp to 25 kb. This protocol divided into three stages:
Accepted Manuscript. News & Views. Primary feather vane asymmetry should not be used to predict the flight capabilities of feathered fossils
 Accepted Manuscript News & Views Primary feather vane asymmetry should not be used to predict the flight capabilities of feathered fossils Xia Wang, Robert L. Nudds, Colin Palmer, Gareth J. Dyke PII: S2095-9273(17)30453-X
Accepted Manuscript News & Views Primary feather vane asymmetry should not be used to predict the flight capabilities of feathered fossils Xia Wang, Robert L. Nudds, Colin Palmer, Gareth J. Dyke PII: S2095-9273(17)30453-X
Dynamic evolution of venom proteins in squamate reptiles. Nicholas R. Casewell, Gavin A. Huttley and Wolfgang Wüster
 Dynamic evolution of venom proteins in squamate reptiles Nicholas R. Casewell, Gavin A. Huttley and Wolfgang Wüster Supplementary Information Supplementary Figure S1. Phylogeny of the Toxicofera and evolution
Dynamic evolution of venom proteins in squamate reptiles Nicholas R. Casewell, Gavin A. Huttley and Wolfgang Wüster Supplementary Information Supplementary Figure S1. Phylogeny of the Toxicofera and evolution
Product Performance Test Guidelines OPPTS Treatments to Control Pests of Humans and Pets
 United States Environmental Protection Agency Prevention, Pesticides and Toxic Substances (7101) EPA 712 C 98 411 March 1998 Product Performance Test Guidelines OPPTS 810.3300 Treatments to Control Pests
United States Environmental Protection Agency Prevention, Pesticides and Toxic Substances (7101) EPA 712 C 98 411 March 1998 Product Performance Test Guidelines OPPTS 810.3300 Treatments to Control Pests
About Ticks and Lyme Disease
 About Ticks and Lyme Disease Ticks are small crawling bugs in the spider family. They are arachnids, not insects. There are hundreds of different kinds of ticks in the world. Many of them carry bacteria,
About Ticks and Lyme Disease Ticks are small crawling bugs in the spider family. They are arachnids, not insects. There are hundreds of different kinds of ticks in the world. Many of them carry bacteria,
Caecilians (Gymnophiona)
") Caecilians (Gymnophiona) David J. Gower* and Mark Wilkinson Department of Zoology, The Natural History Museum, London SW7 5BD, UK *To whom correspondence should be addressed (d.gower@nhm. ac.uk) Abstract
Caecilians (Gymnophiona) David J. Gower* and Mark Wilkinson Department of Zoology, The Natural History Museum, London SW7 5BD, UK *To whom correspondence should be addressed (d.gower@nhm. ac.uk) Abstract
Pesky Ectoparasites. Insecta fleas, lice and flies. Acari- ticks and mites
 Pesky Ectoparasites Parasite control should be at the forefront of every pet owner s life as all animals have the propensity to contract numerous ones at one stage or another. They are a challenge to the
Pesky Ectoparasites Parasite control should be at the forefront of every pet owner s life as all animals have the propensity to contract numerous ones at one stage or another. They are a challenge to the
Volume 2 Number 1, July 2012 ISSN:
 Volume 2 Number 1, July 2012 ISSN: 229-9769 Published by Faculty of Resource Science and Technology Borneo J. Resour. Sci. Tech. (2012) 2: 20-27 Molecular Phylogeny of Sarawak Green Sea Turtle (Chelonia
Volume 2 Number 1, July 2012 ISSN: 229-9769 Published by Faculty of Resource Science and Technology Borneo J. Resour. Sci. Tech. (2012) 2: 20-27 Molecular Phylogeny of Sarawak Green Sea Turtle (Chelonia
recent extinctions disturb path to equilibrium diversity in Caribbean bats
 Log-likelihood In the format provided by the authors and unedited. recent extinctions disturb path to equilibrium diversity in Caribbean bats Luis Valente, 2, rampal S. etienne 3 and Liliana M. Dávalos
Log-likelihood In the format provided by the authors and unedited. recent extinctions disturb path to equilibrium diversity in Caribbean bats Luis Valente, 2, rampal S. etienne 3 and Liliana M. Dávalos
The melanocortin 1 receptor (mc1r) is a gene that has been implicated in the wide
 is a gene that has been implicated in the wide") Introduction The melanocortin 1 receptor (mc1r) is a gene that has been implicated in the wide variety of colors that exist in nature. It is responsible for hair and skin color in humans and the various
Introduction The melanocortin 1 receptor (mc1r) is a gene that has been implicated in the wide variety of colors that exist in nature. It is responsible for hair and skin color in humans and the various
Mechanisms and Pathways of AMR in the environment
 FMM/RAS/298: Strengthening capacities, policies and national action plans on prudent and responsible use of antimicrobials in fisheries Final Workshop in cooperation with AVA Singapore and INFOFISH 12-14
FMM/RAS/298: Strengthening capacities, policies and national action plans on prudent and responsible use of antimicrobials in fisheries Final Workshop in cooperation with AVA Singapore and INFOFISH 12-14
THF EGG. OUTLINE LIFE-HISTORY OF THE CHRY$OMELID GAS TROIDEA CYANEA MELSHEIMER.
 6 PSYCHE [February OUTLINE LIFE-HISTORY OF THE CHRY$OMELID GAS TROIDEA CYANEA MELSHEIMER. BY A. A. GIR_&ULT, WASHINGTON, D. C. DURING late June, 1907, adults of this species were observed feeding on the
6 PSYCHE [February OUTLINE LIFE-HISTORY OF THE CHRY$OMELID GAS TROIDEA CYANEA MELSHEIMER. BY A. A. GIR_&ULT, WASHINGTON, D. C. DURING late June, 1907, adults of this species were observed feeding on the
PARASITOLOGICAL EXAMINATIONS CATALOGUE OF SERVICES AND PRICE LIST
 INSTITUTE OF PARASITOLOGY Biomedical Research Center Seltersberg Justus Liebig University Giessen Schubertstrasse 81 35392 Giessen Germany Office: +49 (0) 641 99 38461 Fax: +49 (0) 641 99 38469 Coprological
INSTITUTE OF PARASITOLOGY Biomedical Research Center Seltersberg Justus Liebig University Giessen Schubertstrasse 81 35392 Giessen Germany Office: +49 (0) 641 99 38461 Fax: +49 (0) 641 99 38469 Coprological
Multiplex real-time PCR for the passive surveillance of ticks, tick-bites, and tick-borne pathogens
 Multiplex real-time PCR for the passive surveillance of ticks, tick-bites, and tick-borne pathogens Guang Xu, Stephen Rich Laboratory of Medical Zoology University of Massachusetts Amherst TICKS ARE VECTORS
Multiplex real-time PCR for the passive surveillance of ticks, tick-bites, and tick-borne pathogens Guang Xu, Stephen Rich Laboratory of Medical Zoology University of Massachusetts Amherst TICKS ARE VECTORS
BioSci 110, Fall 08 Exam 2
 1. is the cell division process that results in the production of a. mitosis; 2 gametes b. meiosis; 2 gametes c. meiosis; 2 somatic (body) cells d. mitosis; 4 somatic (body) cells e. *meiosis; 4 gametes
1. is the cell division process that results in the production of a. mitosis; 2 gametes b. meiosis; 2 gametes c. meiosis; 2 somatic (body) cells d. mitosis; 4 somatic (body) cells e. *meiosis; 4 gametes
The Essentials of Ticks and Tick-borne Diseases
 The Essentials of Ticks and Tick-borne Diseases Presenter: Bobbi S. Pritt, M.D., M.Sc. Director, Clinical Parasitology Laboratory Co-Director, Vector-borne Diseases Laboratory Services Vice Chair of Education
The Essentials of Ticks and Tick-borne Diseases Presenter: Bobbi S. Pritt, M.D., M.Sc. Director, Clinical Parasitology Laboratory Co-Director, Vector-borne Diseases Laboratory Services Vice Chair of Education
Evolution of Birds. Summary:
 Oregon State Standards OR Science 7.1, 7.2, 7.3, 7.3S.1, 7.3S.2 8.1, 8.2, 8.2L.1, 8.3, 8.3S.1, 8.3S.2 H.1, H.2, H.2L.4, H.2L.5, H.3, H.3S.1, H.3S.2, H.3S.3 Summary: Students create phylogenetic trees to
Oregon State Standards OR Science 7.1, 7.2, 7.3, 7.3S.1, 7.3S.2 8.1, 8.2, 8.2L.1, 8.3, 8.3S.1, 8.3S.2 H.1, H.2, H.2L.4, H.2L.5, H.3, H.3S.1, H.3S.2, H.3S.3 Summary: Students create phylogenetic trees to
Protocol for fabrication of microcompartments for long-term culture and imaging of small C. elegans larvae. Henrik Bringmann, March 2011.
 Protocol for fabrication of microcompartments for long-term culture and imaging of small C. elegans larvae Henrik Bringmann, March 2011. 1 Step-by-Step Protocol Step1 : Preparing a humidity dish (see illustration
Protocol for fabrication of microcompartments for long-term culture and imaging of small C. elegans larvae Henrik Bringmann, March 2011. 1 Step-by-Step Protocol Step1 : Preparing a humidity dish (see illustration
The Friends of Nachusa Grasslands 2016 Scientific Research Project Grant Report Due June 30, 2017
 The Friends of Nachusa Grasslands 2016 Scientific Research Project Grant Report Due June 30, 2017 Name: Laura Adamovicz Address: 2001 S Lincoln Ave, Urbana, IL 61802 Phone: 217-333-8056 2016 grant amount:
The Friends of Nachusa Grasslands 2016 Scientific Research Project Grant Report Due June 30, 2017 Name: Laura Adamovicz Address: 2001 S Lincoln Ave, Urbana, IL 61802 Phone: 217-333-8056 2016 grant amount:
Title: Phylogenetic Methods and Vertebrate Phylogeny
 Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
3. records of distribution for proteins and feeds are being kept to facilitate tracing throughout the animal feed and animal production chain.
 CANADA S FEED BAN The purpose of this paper is to explain the history and operation of Canada s feed ban and to put it into a broader North American context. Canada and the United States share the same
CANADA S FEED BAN The purpose of this paper is to explain the history and operation of Canada s feed ban and to put it into a broader North American context. Canada and the United States share the same
muscles (enhancing biting strength). Possible states: none, one, or two.
. Possible states: none, one, or two.") Reconstructing Evolutionary Relationships S-1 Practice Exercise: Phylogeny of Terrestrial Vertebrates In this example we will construct a phylogenetic hypothesis of the relationships between seven taxa
Reconstructing Evolutionary Relationships S-1 Practice Exercise: Phylogeny of Terrestrial Vertebrates In this example we will construct a phylogenetic hypothesis of the relationships between seven taxa
Wes Watson and Charles Apperson
 Wes Watson and Charles Apperson Ticks are not insects! Class Acarina Order Parasitiformes Family Argasidae soft ticks (5 genera) Family Ixodidae hard ticks (7 genera) Genus Dermacentor 30 species Amblyomma
Wes Watson and Charles Apperson Ticks are not insects! Class Acarina Order Parasitiformes Family Argasidae soft ticks (5 genera) Family Ixodidae hard ticks (7 genera) Genus Dermacentor 30 species Amblyomma
Biodiversity and Distributions. Lecture 2: Biodiversity. The process of natural selection
 Lecture 2: Biodiversity What is biological diversity? Natural selection Adaptive radiations and convergent evolution Biogeography Biodiversity and Distributions Types of biological diversity: Genetic diversity
Lecture 2: Biodiversity What is biological diversity? Natural selection Adaptive radiations and convergent evolution Biogeography Biodiversity and Distributions Types of biological diversity: Genetic diversity
What are taxonomy, classification, and systematics?
 Topic 2: Comparative Method o Taxonomy, classification, systematics o Importance of phylogenies o A closer look at systematics o Some key concepts o Parts of a cladogram o Groups and characters o Homology
Topic 2: Comparative Method o Taxonomy, classification, systematics o Importance of phylogenies o A closer look at systematics o Some key concepts o Parts of a cladogram o Groups and characters o Homology
Note: The following article is used with permission of Dr. Sonia Altizer.
 PROFESSIONAL BUTTERFLY FARMING PART I - By Nigel Venters (Contributing Author: Dr. Sonia Altizer) Note: The following article is used with permission of Dr. Sonia Altizer. Monarch Health Program, University
PROFESSIONAL BUTTERFLY FARMING PART I - By Nigel Venters (Contributing Author: Dr. Sonia Altizer) Note: The following article is used with permission of Dr. Sonia Altizer. Monarch Health Program, University
Outline 4/25/2009. Cytauxzoonosis: A tick-transmitted parasite of domestic and wild cats in the southeastern U.S. What is Cytauxzoonosis?
 Cytauxzoonosis: A tick-transmitted parasite of domestic and wild cats in the southeastern U.S. Michelle Rosen Center for Wildlife Health Department of Forestry, Wildlife, & Fisheries What is Cytauxzoonosis?
Cytauxzoonosis: A tick-transmitted parasite of domestic and wild cats in the southeastern U.S. Michelle Rosen Center for Wildlife Health Department of Forestry, Wildlife, & Fisheries What is Cytauxzoonosis?
Formation of Proximal and Anterior Limb Skeleton Requires Early Function of Irx3 and Irx5 and Is Negatively Regulated by Shh Signaling
 Developmental Cell, Volume 29 Supplemental Information Formation of Proximal and Anterior Limb Skeleton Requires Early Function of Irx3 and Irx5 and Is Negatively Regulated by Shh Signaling Danyi Li, Rui
Developmental Cell, Volume 29 Supplemental Information Formation of Proximal and Anterior Limb Skeleton Requires Early Function of Irx3 and Irx5 and Is Negatively Regulated by Shh Signaling Danyi Li, Rui
Antibiotics: Conflict and Communication in Microbial Communities
 Antibiotics: Conflict and Communication in Microbial Communities Antibiotics mediate species interactions in natural habitats, affecting the dynamics of microbial coevolution Daniel C. Schlatter and Linda
Antibiotics: Conflict and Communication in Microbial Communities Antibiotics mediate species interactions in natural habitats, affecting the dynamics of microbial coevolution Daniel C. Schlatter and Linda
Topical prevention and treatment of ticks, fleas, mosquitoes, biting flies and lice for monthly use on dogs and puppies 7 weeks of age and older
 BAYER HEALTHCARE LLC Animal Health Division P.O. BOX 390, SHAWNEE MISSION, KS, 66201-0390 Customer Service Tel.: 800-633-3796 Customer Service Fax: 800-344-4219 Website: www.bayer-ah.com Every effort has
BAYER HEALTHCARE LLC Animal Health Division P.O. BOX 390, SHAWNEE MISSION, KS, 66201-0390 Customer Service Tel.: 800-633-3796 Customer Service Fax: 800-344-4219 Website: www.bayer-ah.com Every effort has
COMMITTEE FOR VETERINARY MEDICINAL PRODUCTS
 The European Agency for the Evaluation of Medicinal Products Veterinary Medicines and Information Technology EMEA/CVMP/005/00-FINAL-Rev.1 COMMITTEE FOR VETERINARY MEDICINAL PRODUCTS GUIDELINE FOR THE TESTING
The European Agency for the Evaluation of Medicinal Products Veterinary Medicines and Information Technology EMEA/CVMP/005/00-FINAL-Rev.1 COMMITTEE FOR VETERINARY MEDICINAL PRODUCTS GUIDELINE FOR THE TESTING
Early warning for Lyme disease: Lessons learned from Canada
 Early warning for Lyme disease: Lessons learned from Canada Nick Hume Ogden, National Microbiology Laboratory @ Saint-Hyacinthe Talk outline The biology of Lyme disease emergence in the context of climate
Early warning for Lyme disease: Lessons learned from Canada Nick Hume Ogden, National Microbiology Laboratory @ Saint-Hyacinthe Talk outline The biology of Lyme disease emergence in the context of climate
Consequences of Antimicrobial Resistant Bacteria. Antimicrobial Resistance. Molecular Genetics of Antimicrobial Resistance. Topics to be Covered
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
COMPARING DNA SEQUENCES TO UNDERSTAND EVOLUTIONARY RELATIONSHIPS WITH BLAST
 Big Idea 1 Evolution INVESTIGATION 3 COMPARING DNA SEQUENCES TO UNDERSTAND EVOLUTIONARY RELATIONSHIPS WITH BLAST How can bioinformatics be used as a tool to determine evolutionary relationships and to
Big Idea 1 Evolution INVESTIGATION 3 COMPARING DNA SEQUENCES TO UNDERSTAND EVOLUTIONARY RELATIONSHIPS WITH BLAST How can bioinformatics be used as a tool to determine evolutionary relationships and to
MID 23. Antimicrobial Resistance. Consequences of Antimicrobial Resistant Bacteria. Molecular Genetics of Antimicrobial Resistance
 Antimicrobial Resistance Molecular Genetics of Antimicrobial Resistance Micro evolutionary change - point mutations Beta-lactamase mutation extends spectrum of the enzyme rpob gene (RNA polymerase) mutation
Antimicrobial Resistance Molecular Genetics of Antimicrobial Resistance Micro evolutionary change - point mutations Beta-lactamase mutation extends spectrum of the enzyme rpob gene (RNA polymerase) mutation
Grasshopper Dissection
 Grasshopper Dissection External Observation Locate the head, thorax, and abdomen. Observe the head. Locate the two compound eyes and the three simple eyes. 1. Why do you think grasshoppers have two types
Grasshopper Dissection External Observation Locate the head, thorax, and abdomen. Observe the head. Locate the two compound eyes and the three simple eyes. 1. Why do you think grasshoppers have two types
7.013 Spring 2005 Problem Set 2
 MIT Department of Biology 7.013: Introductory Biology - Spring 2005 Instructors: Professor Hazel Sive, Professor Tyler Jacks, Dr. Claudette Gardel NAME TA 7.013 Spring 2005 Problem Set 2 FRIDAY February
MIT Department of Biology 7.013: Introductory Biology - Spring 2005 Instructors: Professor Hazel Sive, Professor Tyler Jacks, Dr. Claudette Gardel NAME TA 7.013 Spring 2005 Problem Set 2 FRIDAY February
Lecture 6: Fungi, antibiotics and bacterial infections. Outline Eukaryotes and Prokaryotes Viruses Bacteria Antibiotics Antibiotic resistance
 Lecture 6: Fungi, antibiotics and bacterial infections Outline Eukaryotes and Prokaryotes Viruses Bacteria Antibiotics Antibiotic resistance Lecture 1 2 3 Lecture Outline Section 4 Willow and aspirin Opium
Lecture 6: Fungi, antibiotics and bacterial infections Outline Eukaryotes and Prokaryotes Viruses Bacteria Antibiotics Antibiotic resistance Lecture 1 2 3 Lecture Outline Section 4 Willow and aspirin Opium