UNIVtKSlT v C p. ILLINOIS srary AT URBANA-CHAMPAIGN L ZOLOGY
|
|
|
- Augusta Reeves
- 5 years ago
- Views:
Transcription

1


2 UNIVtKSlT v C p ILLINOIS srary I AT URBANA-CHAMPAIGN L ZOLOGY CO CO

3
4



5 /kjjuh^^i IUHMT FIELDIANA: GEOLOGY A Continuation of the GEOLOGICAL SERIES of FIELD MUSEUM OF NATURAL HISTORY VOLUME 41 FIELD MUSEUM OF NATURAL HISTORY CHICAGO, U.S.A. ^ ^\fr o<&\.v.* *

6


7 TABLE OF CONTENTS The Morphology and Relationships of the Cretaceous Teleost Apsopelix. By Susan Teller-Marshall and David Bardack 1 The Deseadan, Early Oligocene, Mt supialia of South America. By Bryan Patterson and Larry G. Marshall 37

8 \;



9 6E010SY LH // FIELDIANA Geology U0 Published by Field Museum of Natural History Volume 41, No. 1 September 29, 1978 The Morphology and Relationships of the Cretaceous Teleost Apsopelix Susan Teller-Marshall Department of Biological Sciences University of Illinois at Chicago Circle Chicago, Illinois and David Bardack Department of Biological Sciences University of Illinois at Chicago Circle Chicago, Illinois and Research Associate Field Museum of Natural History INTRODUCTION The fossil fishes described in this paper were first reported over a hundred years ago and have been assigned to systematic positions in different teleostean cohorts at one time or another since their discovery. Often, specimens from new localities or those preserved in an unusual manner were given new generic and specific names, adding to the confused taxonomy of these fishes. This work began as an investigation of specimens from a new locality in Texas. These specimens were preserved three dimensionally and were particularly well suited for acid preparation. Later it became evident that a complete description required examination of additional specimens and that the nomenclature of these specimens was in need of extensive revision. Apsopelix was described by Cope (1871) who assigned the name to a single, fragmentary specimen (AMNH 1602) from the "Benton" Library of Congress Catalog Card No.: ISSN Publication
 divided.")
 stressed the similarities among Pelycorapis, Syllaemus, Apsopelix, and a fourth genus, Leptichthys Stewart (1899).")
10 2 FIELDIANA: GEOLOGY, VOLUME 41 formation (Upper Cretaceous) of Kansas. Later Cope (1875, 1877) described two other genera which he believed were similar to Apsopelix: Syllaemus and Pelycorapis. The latter genus, which contained the species P. varius and P. berycinus, was later (Dunkle, 1958) divided. Pelycorapis varius was assigned to Thrissopater, which is a synonym of Pachyrhizodus (Forey, 1977). Woodward (1901, 1903, 1907) stressed the similarities among Pelycorapis, Syllaemus, Apsopelix, and a fourth genus, Leptichthys Stewart (1899). Jordan (1924) placed Leptichthys and Pelycorapis within the genus Apsopelix and assigned this genus to the Syllaemidae. Finally, Dunkle (1958) argued that Helmintholepis Cockerell (1919) and Paleoclupea Dante (1942) were synonyms of Pelycorapis berycinus, that the berycinus species and the Apsopelix-Syllaemus-Leptichthys group share "distinctive as well as general" characters. Dunkle (1958) then tentatively assigned P. berycinus to Apsopelix, the oldest available name. Most recently, Patterson and Rosen (1977) reported that they could detect no differences between Syllaemus, Pelycorapis berycinus, Apsopelix, Leptichthys, Helmintholepis, and Paleoclupea, and treated Apsopelix anglicus (Dixon) as the valid name for this material. Our investigation included examination of specimens referred to Paleoclupea, Leptichthys, Syllaemus, Pelycorapis, and Apsopelix, as well as new material and supports the argument that all of these fossils represent the same fish, Apsopelix anglicus. The present investigation centers on the morphology of Apsopelix which until now has received only cursory description. This account includes features previously undescribed and pursues in further detail features which have been inadequately considered. An attempt is made to focus on those derived characters, present in Apsopelix, which will permit comparison of this genus with major teleostean groups. In the past, Apsopelix has been assigned to the Percesoces 1 (Cope, 1875), Mugilidae (Stewart, 1900), Crossognathidae (Woodward, 1901; Patterson and Rosen, 1977), Syllaemidae (Cragin, 1901; Green, 1957), Pelycorapidae (Cragin, 1901), Clupeidae (Stewart, 1900; Woodward, 1903, 1907; Jordan, 1923; Berg, 1940), and Apsopelicidae (Romer, 1966). Apsopelix is known from North America (Colorado, South Dakota, Kansas, Texas), England, and France. Part of the European material is Albian, but otherwise Apsopelix is restricted to the geologic interval between the Cenomanian and Santonian, possibly



11 TELLER-MARSHALL & BARDACK: APSOPELIX 3 Campanian (but see p. 4) stages of the Upper Cretaceous. It is a unique fish. The antorbitals, scales, and pelvic bones are larger than those in other teleosts of comparable size. The cranial roofing bones are distinctive, and the maxilla comprises nearly all of the dermal upper jaw. Thus, specimens of Apsopelix are easily recognized. The combined advantages of three-dimensional preservation of some fossil material and of acid preparation techniques have facilitated a complete account of the skull. Remaining specimens elucidate most of the post-cranial skeleton, but the caudal skeleton is not complete in any specimen examined. This incomplete preservation of the caudal skeleton is characteristic of Apsopelix, but enough information is available from these specimens to permit some conclusions about the caudal morphology. The morphological adaptations of Apsopelix include features associated with microphagous feeding, particularly the long gill rakers. The gut must have been very long, judging from the remote position of the anal fin. The body was more robust than laterally compressed. These clues indicate that the diet was probably in large part comprised of material difficult to digest, such as plankton. ACKNOWLEDGEMENTS For information concerning Apsopelix we thank Peter L. Forey and Colin Patterson, British Museum of Natural History; and Donn E. Rosen, American Museum of Natural History. The drawings were prepared by Karen Baker Al Lerio and the manuscript typed by Mrs. Beverly Woodard. SPECIMENS EXAMINED Material from several museums has been prepared and studied. Abbreviations include: AMNH, American Museum of Natural History; FMNH, Field Museum of Natural History; KU, University of Kansas Museum of Natural History; SMM, Sternberg Memorial Museum; USNM, United States National Museum; UT, University of Texas Vertebrate Paleontological Collections. 1 The suborder Percesoces was once used to unite the Crossognathidae, Ammodytidae, Scombresocidae, Atherinidae, Mugilidae, and Sphyraenidae. See Woodward, 1901, Part IV, p. 347 for diagnosis.


12 4 FIELDIANA: GEOLOGY, VOLUME 41 AMNH 1602, fish body FMNH PF7463, three-dimensional head and anterior of body, Eagle Ford Formation, Dallas County, Texas, Texas Industries Quarry, southwest of intersection of Chalk Hill Road and Dallas-Ft. Worth Turnpike. KU 309, whole fish; KU 310, fish without tail; KU 318, head and pectoral girdle; KU 519, whole fish, Smoky Hill Chalk Member, Niobrara Formation, western Kansas. KU 18, fish without tail, Fencepost Limestone Bed at top of Greenhorn Formation, western Kansas. KU 882, three-dimensional head and anterior of body, Jetmore or Lincoln limestone levels of Greenhorn Formation, Mitchell County, Kansas, 1 mile northeast of Scotsville. SMM 7958, whole fish, Smoky Hill Chalk Member, Niobrara Formation, Kansas. USNM 16725, three-dimensional head and anterior part of body, Pierre Shale, Chamberlain, South Dakota (as per catalogue, but horizon and location uncertain as there is no evidence of the matrix similar to that containing the fossil at this location.) UT 848, posterior part of head and anterior part of body, Upper Cretaceous (horizon uncertain), Williamson County, Texas, 1 3 A miles east of Coupland in branch of Bushy Creek. UT , whole fish, Austin Group, Ector Member, Fannin County, Texas, Savoy Pit 4 miles south of Savoy on farm road 1752, then east three-quarters of a mile on unpaved road. UT , whole fish, basal Austin Chalk, Grayson County, Texas west of farm road 1417 where it passes radar tower of Perry AFB. SYSTEMATIC DESCRIPTION Subdivision Teleostei (sensu ir'atterson, 1975) Order incertae sedis Family Crossognathidae Patterson and Rosen, 1977 Genus Apsopelix Cope, 1871 Apsopelix Cope, 1871, p. 424; Jordan, 1924, p. 230, pi. XX; Dunkle, 1958, p Type species: A. sauriformis (monotypy). Syllaemus Cope, 1875, p. 180; Stewart, 1900, p. 384, pi. LXXII fig. 2; Woodward, 1901, p. 350; Cragin, 1901, p. 26, pi. I; Woodward, 1903, p. 88, pi. XX, pi. XXI figs. 1-2; Chabanaud, 1930, p. 646, pi. LXVIII; Green, 1957, p. 40, figs. 1-2; Wenz, 1965, p. 13, figs. 3-5, pi. II. Type species: S. latifrons (monotypy). Pelycorapis Cope (not Cope, 1874), 1877, p Type species: Thrissopater varius (monotypy). Leptichthys Stewart, 1890, p. 78; Stewart, 1900, p. 372, pi. LXXII fig. 1; Jordan, 1924, p. 231, pi. XXI. Type species: L. agilis (monotypy).
. Diagnosis. Teleost fishes in which the trunk is fusiform, and head length about equal to greatest depth. Head tapers ventrally.")


13 TELLER-MARSHALL & BARDACK: APSOPELIX 5 Helmintholepis Cockerell, 1919, p. 176, pi. XXXII, fig. 4. Type species: H. vermiculatus (monotypy). Paleoclupea Dante, 1942, p. 340, figs Type species: P. dakotensis (monotypy). Diagnosis. Teleost fishes in which the trunk is fusiform, and head length about equal to greatest depth. Head tapers ventrally. Orbit diameter per cent head length, nearly equal in length to ethmoid region; sclerotic ring ossified, snout pointed. Rostral fused with underlying mesethmoid, shield-like anteriorly with paired splints passing back below frontals. Cranial roof arched transversely with a median depression where frontals meet parietals; parietals wider than long, medially united. Roofed posttemporal fossae confluent above cranial vault, extending forward to orbitosphenoid; deep dilatator and subtemporal fossae; orbitosphenoid forms vertical plate across upper third of orbit; basisphenoid present; parasphenoid forms angle at ascending wings and extends posteriorly beyond braincase. Occipital condyle formed by basioccipital and exoccipitals. Hyomandibular vertical with posteriorly directed fossa below neurocranial head. Palate with ossified processes which meet lateral and median ethmoid elements. Lower jaw articulation lies beneath anterior third of orbit. Premaxilla about one-tenth of upper jaw; maxilla arcuate, about eight times longer than maximum depth; two supramaxillae. Coronoid height of mandible contained 2 1 /2 times in mandible length. Angular and articular fused, retroarticular distinct. Parasphenoid toothless. Basihyal with a pair of tooth plates; gill rakers long and slender. No bone-enclosed ethmoid commissure; preopercular and infraorbital sensory canals with short, bone-enclosed branches; lateral line conspicuous. Dermosphenotic and supraorbitals large; antorbital nearly covers ethmoid region; posterior infraorbitals extend to preopercular, no suborbitals. Preopercular expanded ventroposteriorly with radiating ridges on expanded surface; operculum and suboperculum smooth, about equal in size. Scales cycloid, rows on one side at midbody; height of exposed portion of each scale about twice its length. About 40 preural centra, about 12 caudal centra; each centrum with a single lateral ridge, neural arches fused to all but anterior centra; epineural, epipleural, and epicentral intermusculars present. Pectoral fins with about 15 rays; pelvic fins abdominal with at least 12 rays, pelvic bone broadly expanded. Dorsal fin with at least 13 rays, originating about midway between snout and posterior end of body; anal fin short and very close to caudal origin; caudal fin deeply forked with narrow
 Calamopleurus y anglicus Dixon, 1850, p. 375, pi. XXXII, figs. 11-12. Apsopelix sauriformis Cope, 1871, p. 424; Jordan, 1924, p. 230, pi. XX; Dunkle, 1958, p. 271.")


14 6 FIELDIANA: GEOLOGY, VOLUME 41 base; fin rays cross hypurals at steep angle. Uroneurals modified posteriorly, anterior uroneurals cover lateral faces of ural and first two preural centra. Apsopelix anglicus (Dixon) Calamopleurus y anglicus Dixon, 1850, p. 375, pi. XXXII, figs Apsopelix sauriformis Cope, 1871, p. 424; Jordan, 1924, p. 230, pi. XX; Dunkle, 1958, p Syllaemus latifrons Cope, 1875, p. 181; Stewart, 1900, p. 384, pi. LXXII, fig. 2; Woodward, 1901, p. 351; Cragin, 1901, p. 27, pi. I. Pelycorapis berycinus Cope, 1877, p Calamopleurus anglicus Woodward, 1888, p Leptichthys agilis Stewart, 1899, p. 78; Stewart, 1900, p. 372, pi. LXXII, fig. 1; Jordan, 1924, p. 231, pi. XXI. Syllaemus anglicus Woodward, 1901, p. 351; Woodward, 1903, p. 89, pi. XX, pi. XXI 1-2; Chabanaud, 1930, p. 646, pi. LXVIII. figs. Helmintholepis vermiculatus Cockerell, 1919, p. 176, pi. XXXII, fig. 4. Paleoclupea dakotensis Dante, 1942, p. 340, figs Syllaemus hanifii Green, 1957, p. 40, figs Syllaemus albiensis Wenz, 1965, p. 13, figs. 3-5, pi. II. Diagnosis. As for genus, only species. Proportions (as a percentage of standard length): head length 23-25, greatest depth 23-25, prepectoral 28-29, prepelvic 71-75, predorsal 41-42, preanal Description. General features: The head is triangular in lateral view with the posterior border being gently curved. Dorsally the head forms a entrally. Maximum head * relatively broad, flat surface and tapers depth behind the lower jaw articulation equals about 65 per cent of the head length. Diameter of the orbit is slightly less than the preorbital distance and represents about 30 per cent of the head length. The jaw articulation lies beneath and slightly anterior to the middle of the orbit. Three specimens preserved in three-dimensional form probably closely retain the original shape of the fish. Examination of these suggests that the body was deeper than wide, ovoid in cross-section (at least at mid-body), and somewhat wider ventrally than dorsally. Scales with their exposed area twice as high as long cover the body. Both the pectoral and pelvic fins insert ventrally. 'Calamopleurus was founded by Agassiz on fragments of an entirely different fish from the Upper Cretaceous of Brazil. See Woodward, 1903, p. 89.


: The neurocranium is 2V2 times as long as deep; its maximum")
15 TELLER-MARSHALL & BARDACK: APSOPELIX ', I O,Asp V n ;v Epo Soc Pto Fig. 1. Apsopelix anglicus, dorsal view of the neurocranium. For abbreviations see p. 30. Pelvic fins are abdominal, the dorsal and anal fins are short. The caudal fin is slender and deeply forked. NEUROCRANIUM (figs. 1-5): The neurocranium is 2V2 times as long as deep; its maximum width across the pterotics equals nearly
, subtemporal fossae (fig. 3, stf), and dilatator fossae (fig. 3, df) are well developed.")

16 8 FIELDIANA: GEOLOGY, VOLUME 41 three-fourths the total neurocranial length. The postorbital part of the braincase is short, accounting for less than one-third of the total neurocranial length. Maximum neurocranial depth occurs just behind the orbit. The posttemporal fossae (fig. 3, ptf), subtemporal fossae (fig. 3, stf), and dilatator fossae (fig. 3, df) are well developed. The posttemporal fossae are confluent above the cranial vault and extend well forward, probably to the orbitosphenoid. As in megalopids, the vertical depth of these fossae has resulted in a pronounced convexity of the skull roof in the otic region. In posterior view (fig. 4) the neurocranium slopes ventrolateral^ from the midline so that its dorsal surface describes an arc of 130 around an axis at the middle of the basioccipital. The frontals (fig. 1, Fr) are flat sheets of bone, narrow anteriorly, then widening gradually to contact the parietals and pterotics posteriorly. The frontals are joined by a straight suture and form a median depression in the cranial roof (fig. 1) just anterior to the fronto-parietal juncture. Each frontal shows a line of irregularly shaped ridges and pits marking the path of the supraorbital sensory canal. These lines diverge anteriorly. The parietals (figs. 1-2, Pa) meet in the dorsal midline at a wavy suture. They are wider than long and taper laterally, extending more than half the distance from their joint suture to the lateral edge of the cranial roof. They are also marked with ridges and pits, including a prominent, centrally located pit line (fig. 2, apl). Each pterotic (figs. 1-4, Pto) forms the dorsolateral corner of the neurocranium and makes up the lateral wall plus part of the dorsal and ventral walls of the posttemporal fossa. In dorsal view the dermopterotics meet the frontal, parietal, and epioccipital with irregularly shaped sutures. The lateral surface of the autopterotic is deeply excavated to form, along with the autosphenotic, the dilatator fossa. This fossa is roofed by the frontal and more posteriorly by a lateral extension of the dermopterotic. Beneath this fossa, the hyomandibular facet (fig. 3, fhm) slopes anteroventrally and continues forward onto the autosphenotic. The facet is slightly constricted midway along its length. Beneath the hyomandibular facet is a deep subtemporal fossa (fig. 3, stf), its long axis sloping anteroventrally. The autopterotic forms the roof of this fossa. The epioccipitals (figs. 1, 4, Epo) are visible in dorsal and posterior views. They contact the parietals anteriorly, the supraoccipital


 is visible in dorsal and posterior views.")
17 TELLER-MARSHALL & BARDACK: APSOPELIX Fig. 2. Apsopelix anglicus, dorsal view of the posterior third of the neurocranium. For abbreviations see p. 30. medially, and the pterotics laterally. The dorsal surface of each epioccipital tapers and is produced posteriorly over the dorsomedial corner of the posttemporal fossa. The epioccipitals are covered dorsally by large supratemporals (fig. 2, Stt). A transverse groove along the anterior margin of the epioccipitals and supraoccipital marks the line of contact between the supratemporals and neurocranium. In posterior view the epioccipitals contact the supraoccipital medially and the exoccipitals ventrally. In this view they are square-shaped, with the lateral margin and ventrolateral corner thickened to form a rounded ridge (fig. 4). This ridge continues onto the exoccipital. The epioccipitals contribute to the dorsomedial and most of the medial wall of the posttemporal fossa. The supraoccipital (figs. 1, 4, Soc) is visible in dorsal and posterior views. Dorsally it is wider than long and exhibits a small, median, posteriorly directed prong. In posterior view it is pentagonal, and about as deep as wide. Together with the exoccipitals, the supraoccipital is gently concave in posterior view. The exoccipitals (figs. 3, 4, Exo) each exhibit a triradiate set of


18 es 1 3 a I '> «? O Q- o 1X1


; the remainder of these foramina probably carried blood vessels and the supratemporal branches of IX and X. The intercalars (figs. 3, 4, Ic) are small and barely visible in posterior view.")
19 TELLER-MARSHALL & BARDACK: APSOPELIX 11 Soc Pto Fig. 4. Apsopelix anglicus, posterior view of the neurocranium. For abbreviations see p. 30. rounded ridges in posterior view. The dorsally directed ridge is sutured to a ventrolateral^ thickened arm of the epioccipital. The laterally directed ridge meets the intercalar and autopterotic where these bones form the floor of the posttemporal fossa, while the ventromedially directed ridge contacts the basioccipital and contributes to the occipital condyle. Lateral to the foramen magnum is the foramen for the vagus nerve trunk (fig. 4, X), while the glossopharyngeal nerve trunk probably emerged through one of the numerous foramina on the posterolateral surface of the exoccipital (fig. 3); the remainder of these foramina probably carried blood vessels and the supratemporal branches of IX and X. The intercalars (figs. 3, 4, Ic) are small and barely visible in posterior view. Each does, however, extend forward to meet the prootic along the margin of the subtemporal fossa. The basioccipital (figs. 3, 4, Boc) lies beneath the exoccipitals and is covered laterally by the parasphenoid. In lateral view the surface of the basioccipital is concave. These depressions may have housed anterior extensions of the swimbladder. The prootics (fig. 3, Pro) are deeper than long in lateral view and

.")
 and for the hyomandibular branch of VII (fig. 3, fhm VII).")
20 12 FIELDIANA: GEOLOGY, VOLUME 41 expanded dorsally to meet the autopterotics, autosphenotics, and pterosphenoids. Ventrally each prootic forms a thin sheet overlapped laterally by ascending wings of the parasphenoid. There is a slender, posteriorly directed arm which forms the ventroposterior wall of the subtemporal fossa and contacts the intercalar, while the main body of the prootic contributes to the anterior wall of this fossa. Ventral to the base of this posteriorly directed arm is the foramen for posterior exit of the lateral head vein (fig. 3, fhv). More anteriorly on the lateral surface of the prootic are two additional foramina for the orbitonasal artery and palatine branch of VII (fig. 3, foa; fpal VII) and for the hyomandibular branch of VII (fig. 3, fhm VII). The anterior surface of the prootic shows a triangular pattern of foramina of equal size; two lateral foramina mark the exit of the trigeminal nerve and the anterior exit of the lateral head vein, while the more medial foramen marks the exit of the oculomotor nerve. The autosphenotics (fig. 1, 3, Asp) are triangular in dorsal view, rectangular in orbital view, and produced laterally to a prominent spine. The dorsal and posterior surfaces are concave, while the orbital surface is nearly flat and vertical. This latter face is pierced by a foramen for the otic branch of the facial nerve. Behind the spine a pocket on the posterior face of the autosphenotic (fig. 3, flap), oriented more ventrally than horizontally, probably marks the origin of part of the levator arcus palatini (cf. Forey, 1973a, p. 1,308). The dorsal surface of the autosphenotic contributes to the dilatator fossa (fig. 3, df). The pterosphenoids (fig. 3, Pts) are anteriorly facing sheets which form the margin of the optic foramen with their medial edges. Lateral and dorsal to the optic foramen is a small opening within each pterosphenoid probably for the trochlear. The orbitosphenoid (fig. 3, Ors) extends forward in front of the pterosphenoids. It is a vertical sheet of bone longer than deep and extending more than half the distance to the anterior margin of the orbit. The dorsal part of this sheet is enlarged where the olfactory tract lay, and a foramen at the anterior end of the bone marks the exit of this tract. The basisphenoid is Y-shaped in orbital view; the upper arms of the Y form a gentle yolk shaped edge, and the vertical stem of the Y is longer than the arms. The stem contacts the parasphenoid below, while the arms touch the prootics and pterosphenoids above. The parasphenoid (fig. 3, Par) forms an obtuse angle beneath its


. There is a narrow ventral vomer and a pair of stout lateral ethmoids (fig. 3, Le) which form the anterior boundary of the orbit.")
21 TELLER-MARSHALL & BARDACK: APSOPELIX 13 Fig. 5. Apsopetix anglicus, dorsal view of the ethmoid region showing the relationships of the median ethmoid (Me), palatine process (pme), and maxilla (Mx). For abbreviations, see p. 30. junction with the prootic. Anteriorly it is a slender, triangular strut deeply cleft for receipt of the vomer. Behind the point where the ascending wings of the parasphenoid meet the prootics, the parasphenoid deepens, increasing the size of the posterior myodome, and extends ventrally as an elongated process which passes beneath and posterior to the basioccipital. The parasphenoid appears to be toothless. The ethmoid region is well ossified. It consists of a single dorsal median ethmoid (figs. 1, 5, Me), probably the result of fusion between a smooth dermal rostral and the underlying peri and endochondral ossifications (see Patterson, 1970 for a discussion of the ethmoid region and Nelson, 1969, p. 8 regarding use of the word "fusion"). There is a narrow ventral vomer and a pair of stout lateral ethmoids (fig. 3, Le) which form the anterior boundary of the orbit. The posterior extent of the vomer could not be determined directly; however, KU 882 shows a ventral cleft between anterior arms of the parasphenoid presumably occupied by the vomer. This cleft disappears beneath the middle of the orbit, but it was filled by matrix posteriorly so that it is likely that the ossified vomer extended only as far as the posterior edge of the ethmoid region. Patterson


22 14 FIELDIANA: GEOLOGY, VOLUME 41 and Rosen (1977) cite a partially paired vomerine tooth patch in Apsopelix, but none of the specimens examined in this study show evidence of vomerine teeth. The median ethmoid bifurcates posteriorly, each half tapering beneath the frontals. Anterior to the bifurcation the ethmoid widens, extending anteroventrally to abut on either side against a dorsally directed process from the palatine (figs. 5-6, pme). The most anterior part of the ethmoid is narrow and biconcave to receive the anterior ends of the maxillae. There is no evidence of a bone enclosed ethmoid commissure. The lateral ethmoids are set back some distance from the median ethmoid since the ethmoid region at least equals the orbit in length. Anteriorly, the lateral ethmoid forms the posterior wall of the nasal capsule, then increases in depth toward the orbit, spanning the distance between the frontal above and the parasphenoid below. A small depression on the posterior surface of the lateral ethmoid, at the ventrolateral corner, receives a dorsal process from the palate (fig. 6, pie). HYOPALATINE SERIES (figs. 5-7): Elements of the hyopalatine series are similar in their relationships to those of other lower teleosts, but some of the bones have distinct characteristics. Each hyomandibular (fig. 6, Hm) is vertical with an expanded neurocranial head. The narrow shaft has a groove along its anterior edge. The medial side of this groove extends forward as a thin sheet of bone. Just below the neurocranial head is a large posterior fossa which contained the origin of the dilatator operculi. In front of this fossa are numerous small foramina. Forey (1975, pp , fig. 12) has contrasted the hyomandibular architecture and muscle origins in Elops and two clupemorphs. As far as the levator arcus palatini and the dilatator operculi are concerned, the skeletal features of Apsopelix indicate that the muscle arrangements probably differed from that in Elops and other early teleosts. But the arrangement of these muscles in Apsopelix was not as in clupeomorphs either, since in Apsopelix the dermosphenotic is large, there is no ridge on the frontal, and no anterodorsal process on the hyomandibular (cf. Forey, 1975, fig. 12b). In Apsopelix there appears to have been a shift of at least part of the dilatator operculi to an origin on the hyomandibular, freeing the posterior edge of the autosphenotic for origin of levator arcus


. The metapterygoid (fig. 6, Mpt) is an elongate bone, saddleshaped where its anterior edge overlaps the endopterygoid (fig. 6, Enpt).")
23 TELLER-MARSHALL & BARDACK: APSOPELIX 15 Fig. 6. Apsopetix anglicus, right hyopalatine series in lateral view. For abbreviations see p. 30. palatini fibers in order to meet the demands of a microphagous feeding mode that requires complex movements and increased efficiency of the palate abductors. So, while both Apsopelix and clupeomorphs exhibit morphological adaptations associated with the microphagous feeding mode, the adaptations of Apsopelix differ from those of clupeomorphs and are not indicative of a close relationship. Between the shaft and the anterior bony sheet of the hyomandibular are two foramina, one of which probably marks the exit of the mandibular and preopercular branches of the facial nerve (fig. 6, fmp VII). The metapterygoid (fig. 6, Mpt) is an elongate bone, saddleshaped where its anterior edge overlaps the endopterygoid (fig. 6, Enpt). Posteriorly the metapterygoid joins the lower portion of the hyomandibular, but the suture is difficult to follow. The ventral edge of the metapterygoid lies along the dorsal border of the quadrate. Each ectopterygoid (fig. 6, Ecpt) is boomerang-shaped and lies along the anterior edge of the quadrate. It then curves anteriorly, continuing along the lateral edge of the endopterygoid (fig. 6, Enpt). None of the pterygoid elements appear to bear dentition.
 stems from the posterior part of the palatine as in Albula vulpes (Forey, 1973b, fig. 79) or from the anterior part of the ectopterygoid as in Clupea harengus (Kirchoff, 1958, figs.")

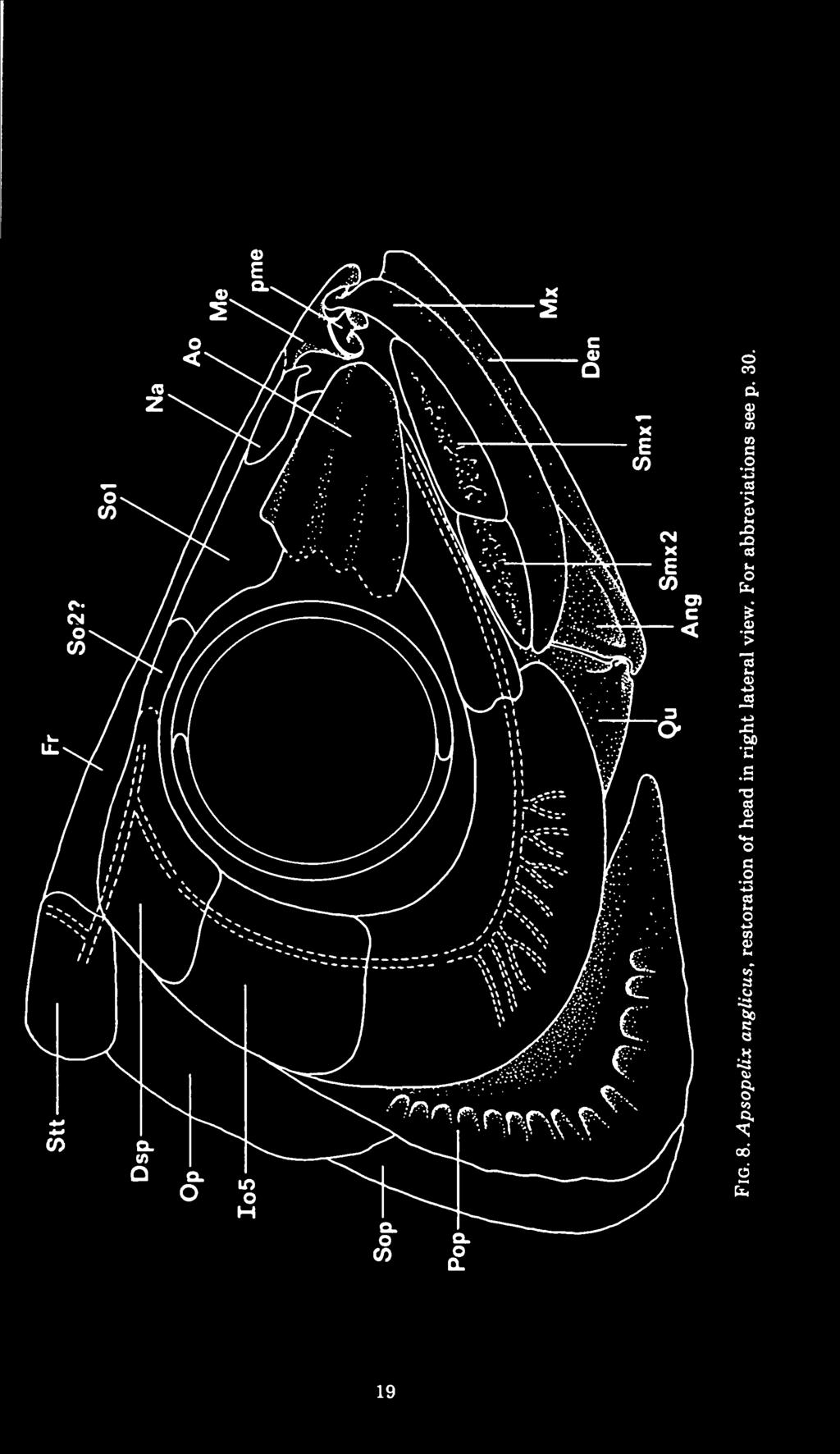
24 16 FIELDIANA: GEOLOGY, VOLUME 41 The anterior part of the palate shows no sutures on the specimens examined and there is some difficulty determining whether the lateral ethmoid process of the palate (fig. 6, pie) stems from the posterior part of the palatine as in Albula vulpes (Forey, 1973b, fig. 79) or from the anterior part of the ectopterygoid as in Clupea harengus (Kirchoff, 1958, figs. 11, 15, 16). The palatine (fig. 6, Pal) is well ossified throughout its length and bears an anterior process (figs. 5-6, pme) which extends between the median ethmoid and the maxilla. This process is fused to the palatine in FMNH PF7463. It appears to be a separate ossification in USNM 16725, but it is intimately associated with the dermopalatine and is not an ethmoid element (cf. Wenz, 1965, fig. 3). This same process is an ossification of the autopalatine in Albula vulpes. SMM 7958 shows a number of conical teeth on the palatine; these teeth are larger than the similarly shaped teeth on the jaw elements. The quadrate (fig. 6, Qu) is broadly expanded dorsally to meet the pterygoids. Posteriorly it bears a deep, broad notch for the symplectic (fig. 6, Sy). The quadrate condyle lies beneath the orbit. Behind the condyle a notch in the quadrate receives the postarticular process of the angular (fig. 7). Upper jaw (figs. 5, 8): The upper jaw consists of a premaxilla, maxilla, and two supramaxillae. A premaxilla has never been described in Apsopelix. Of the specimens examined in this study only one, KU 18, retains a fragment of A it. small, laminar bone, it bears minute acuminate teeth. The premaxilla probably fits into a notch at the anterior end of the maxilla (fig. 5), but its true extent and shape remain unknown. Rart? Fig. 7. Apsopelix anglicus lateral (left) and medial (right) views of the lower jaw articulation. For abbreviations see p. 30.

 is the longer of the two. Neither of these bones bears an anterior projection as in many other lower teleosts, but rather the posterior supramaxilla barely overlies its anterior partner.")
: The lower jaw is short and deep.")
25 TELLER-MARSHALL & BARDACK: APSOPELIX 17 The maxilla (figs. 5, 8, Mx) extends from beneath the anterior end of the orbit forward to articulate with the median ethmoid. It is a sabre-shaped bone, nearly uniform in depth throughout its length, and about 12 times longer than deep. Dentition consists of a single row of minute marginal teeth similar to those on the premaxilla, 0.5 mm. from base to tip in FMNH PF The anterior supramaxilla (fig. 8, Smx 1) is the longer of the two. Neither of these bones bears an anterior projection as in many other lower teleosts, but rather the posterior supramaxilla barely overlies its anterior partner. UT is preserved with the mouth open and shows that the supramaxillae do not separate as much as in clupeoids, but tend to function as a unit. Both supramaxillae exhibit rugose ornamentation along their entire length and are as deep as the maxilla at their deepest point. Lower jaw (figs. 7-8): The lower jaw is short and deep. Maximum between one-third and one-half total length and occurs depth is about midway along this length. The ventral margins of both the dentary (figs. 7-8, Den) and angular (figs. 7-8, Ang) curve medially to form a narrow shelf marked with pores for the mandibular sensory canal. The number of these pores is uncertain, but there appear to be at least two on the posterior part of the dentary and one on the angular. Mandibular dentition consists of a single row of minute teeth, like those on the maxilla, along the dentary margin. The dentaries meet in a short anterior symphysis without curving much inward. Each dentary receives the angular in a deep posterior notch and extends back above and below it (fig. 7). A postarticular process of the angular extends behind the facet for the quadrate condyle, although this structure is broken off easily and is missing in all but one of the specimens examined, KU 318. There is a small, distinct retroarticular visible in KU 318, USNM 16725, and UT Whether this element takes part in or is excluded from the articular facet could not be determined from these specimens. The medial surface of the angular shows a pocket filled by what appears to be an articular in KU 318 (fig. 7, Art). Patterson and Rosen (1977) state that in BMNH the mandibular sensory canal opening is in the medial position, the angular and articular are fused, and the retroarticular is free except on the surface of the facet for the quadrate where all three bones appear to be fused.



26 18 FIELDIANA: GEOLOGY, VOLUME 41 Hyoid and branchial ARCHES: Some elements of the hyoid and branchial arches are preserved in FMNH PF 7463, including one anterior ceratohyal, hypohyals, basihyal, the anterior epibranchials and ceratobranchials, one hypobranchial, and one basibranchial. The anterior ceratohyal is rectangular with a gently concave ventral edge and a centrally located foramen. Two equally developed hypohyals meet the anterior edge of the ceratohyal. The basihyal ossified only posteriorly. A pair of tooth plates lies on its oral surface and extends forward beyond the ossified element. Each tooth plate is about one-fifth as wide as long and covers the lateral third of the basihyal, leaving the center of that bone without teeth. There are 10 longitudinal rows of small, posteriorly directed tooth bases per tooth plate. The basihyal is the only element of the hyoid series with dentition preserved. Along the oral edge of the epibranchials and ceratobranchials are a series of gently curved gill rakers, about 8 mm. in length. There are no tooth plates present on any of the branchial arches. Impressions of 12 branchiostegal rays are present in UT , but the irregular nature of the spaces between these grooves indicates that there were more rays. Dermal bones of the cheek (fig. 8): The infraorbitals are preserved in FMNH PF 7463 and USNM There is a large antorbital followed by at least three distinct infraorbitals plus a dermosphenotic. The postorbital components of this series extend back as far as the preoperculum. The antorbital (fig. 8, Ao) is wedge shaped, expanded posteriorly, and tapering to a blunt anterior edge. There are two anteroposteriorly oriented grooves on the antorbital, the lower probably associated with the infraorbital sensory canal as it entered the antorbital, while the more dorsal probably housed the antorbital branch of that canal. The dorsal edge of the antorbital overlaps the first supraorbital. The infraorbitals are so fragmented that it is difficult to determine their exact number. The primitive condition of the teleostean infraorbital series probably included seven separate bones from the antorbital to the dermosphenotic (Nelson, 1969). There appears to have been a reduction in the number of infraorbital bones in Apsopelix. However, there is some question as to where the implied fusion has occurred. Wenz (1965, fig. 5, p. 18) figured Apsopelix with an antorbital plus five infraorbitals, including the dermosphenotic.


27 eo o eo J3 00 as gf x

 between the large infraorbital and the dermosphenotic, implying that the large element is a fusion of Io 2 + 3+4.")
28 20 FIELDIANA: GEOLOGY, VOLUME 41 There is a large infraorbital covering the posteroventral region of the cheek and marked by several branches of the infraorbital sensory canal. Above this largest bone in the series are figured two more bones plus the dermosphenotic, implying that the large bone is a fusion of the infraorbitals In USNM and FMNH PF 7463, the large infraorbital covering the posteroventral area of the cheek and marked by branches of the infraorbital sensory canal is easily recognized, but there appears to be only one additional bone (fig. 8, Io5) between the large infraorbital and the dermosphenotic, implying that the large element is a fusion of Io If Wenz's (1965) interpretation is correct, then it is possible that the element referred to in our paper as Io5 is actually a fusion of Io 4+5. The dermosphenotic is an elongate, triangular bone overlying the autosphenotic and covering the dilatator fossa. It is smooth where it meets the infraorbitals, but ornamented with ridges and pits where it contacts the frontal. This ornamentation marks the meeting of the temporal and infraorbital canal. The infraorbital sensory canal traverses a path through the center of the infraorbital bones. There is at least one long, anteriorly expanded supraorbital (fig. 8, SO 1) and possibly a second, smaller supraorbital evident only in USNM The first supraorbital covers the anterior margin of the orbit posteriorly and extends forward medial to the antorbital. The second supraorbital (fig. 8, SO 2) spans the distance between the posterior edge of the first supraorbital and the anterior edge of the dermosphenotic. The nasal is a small, rod-shaped bone (fig. 8, Na) above and anterior to the first supraorbital in USNM It is in line with the anterior part of the supraorbital sensory canal on the frontal and shows evidence of a bone enclosed canal. The nasals appear to taper anteriorly, but this could be an artifact of preservation. Opercular series (fig. 8): The preoperculum (fig. 8, Pop) expands ventraliy from a relatively narrow dorsal end. The surface of the expanded portion is marked by radiating ridges which end posteriorly as fine lines, each ridge giving rise to about four lines. Pores of the preopercular sensory canal are evident along the anterior margin of these radiating ridges. The sensory canal does not run along the anterior margin of the bone, but rather through its center, particularly at the angle (cf. Forey, 1973b, Elops, fig. 6, p. 19).
 is also smooth and about the same size and shape as the operculum.")
: The scales are large and cycloid. Faint anteroposteriorly oriented folds and closely spaced circuli are visible on the exposed surface.")
 discussed the scales of Apsopelix in detail.")
29 TELLER-MARSHALL & BARDACK: APSOPELIX 21 The operculum (fig. 8, Op) is smooth and without ornamentation. Its width is contained in its length 1 Vb times. It has a gently curved posterior border, and is barely overlapped anteriorly by the preoperculum. The suboperculum (fig. 8, Sop) is also smooth and about the same size and shape as the operculum. Most of its surface is covered by the expanded ventral portion of the preoperculum. The interoperculum is covered by the preoperculum except for its ventral edge. It is about as long as the suboperculum. Scales (fig. 10): The scales are large and cycloid. Faint anteroposteriorly oriented folds and closely spaced circuli are visible on the exposed surface. A complete scale is longer than deep, the deepest portion being posterior to the center. There are longitudinal rows of scales present on each side. The sixth transverse row from the dorsal fin carries the lateral line, which consists of a series of simple tube-like openings. Dunkle (1958, pp ) discussed the scales of Apsopelix in detail. Paired fins and their supports (figs. 2, 9, 10): The paired fins are in the primitive position for teleosts, the pectorals inserting low on the flank and the pelvics set back in the abdominal region. There are at least 15 rays on the broad-based pectoral fins and at least 12 rays on the pelvic fins (fig. 10). The supratemporals (fig. 2, Stt) are scale-like sheets of bone which overlap the supraoccipital, epioccipitals, and pterotics in dorsal view. Their anterior surface has coarse rugae which follow the path of the supratemporal commissure and its posteriorly directed branches. The supratemporals meet only at their anterior edges on the dorsal midline. The posttemporals (fig. 2, Ptt) and supracleithra are rod-shaped bones. The supracleithrum is longer than the posttemporal. The lateral line traverses the posttemporal near its posterodorsal margin along the anterior half of its length. The supracleithrum overlaps a considerable portion of the cleithrum, and reaches the level of the ventral margin of the orbit ventrally. Each cleithrum curves anteroventrally and medially around the gill chamber and meets its complement in the ventral midline just anterior to this chamber. The lateral surface of the cleithrum is expanded above the pectoral fin and there appears to be at least one scale-like postcleithrum just above the pectoral fin base. Only the posterior end of the scapula has been observed; it has a rounded ridge which forms a point of attachment for the first pec-



30 22 FIELDIANA: GEOLOGY, VOLUME 41 Fig. 9. Apsopelix anglicus, ventral view of right pelvic plate. A = anterior; M= medial. toral ray and the first radial. The coracoid contacts the medial edge of the scapula posteriorly and the inner face of the cleithrum more anteriorly. Where the coracoid meets the scapula it forms a ridge continuous with the scapular ridge and supports the majority of the fin rays via the remaining radials. The coracoids meet in the ventral midline to form a median ventral keel. This keel extends forward, but does not reach the cleithral symphysis. Posteriorly the keel extends back beyond the scapulacoracoid ridge. The mesocoracoid extends dorsally from its basal contact with the scapula and coracoid, forming an arch. Its dorsal end touches the medial surface of the cleithrum. There are four radials, the most medial being longest. They are simple struts, slightly expanded at their distal ends. The prepelvic distance is about three-fourths of the standard length. Each pelvic fin is attached to a long, broad triangular pelvic bone (fig. 9) which extends anteriorly from the origin of the fin for a distance of about four abdominal vertebrae. There is a thickened ridge along the posterior edge of each pelvic bone. The two bones meet in a median symphysis along the ventral midline. This massive, broad pelvic bone is charactertistic of Apsopelix. M


 original description of AMNH 1602.")
31 TELLER-MARSHALL & BARDACK: APSOPELIX 23 Median fins (fig. 10): Of those specimens studied in which the whole fish is preserved, the ventral surface is exposed and the body twisted. The preservation of the median and caudal fins is poor. KU 309, KU 519, and SMM 7958, in which the dorsal fin is preserved, and UT , in which the anal fin is preserved, show the best median fin preservation of the specimens examined in this study. The dorsal fin is short, with about rays, the longest being most anterior. The predorsal length is about 40 per cent of the standard length. The first dorsal ray is the longest and most robust. The anal fin of Apsopelix is short and the preanal length is about four-fifths of the standard length. UT shows a few anal rays, but otherwise this fin is not known in Apsopelix apart from Cope's (1871) original description of AMNH Only the first anal ray is preserved in UT , but examination of photos of AMNH 1602 show the anal fin with rays. The caudal fin is deeply forked, with a narrow base. The number of branched principal rays appears to be nine above and eight below, but such counts are difficult to obtain in the specimens examined and may be in error. Vertebral column and caudal skeleton (fig. 10): There are about 40 preural centra; nearly 30 are abdominal. The centra are longer than deep, somewhat constricted at their centers, and pierced for passage of the notochord. Each centrum is marked by a single lateral ridge. The neural arches of the anteriormost centra are autogenous, but otherwise they appear to be fused with the centra as are the haemal arches. Pleural ribs are associated with at least 20 centra, being absent from the first few vertebrae, but the nature of the rib articulations has not been preserved in the specimens examined. KU 18 shows numerous intermuscular bones. Above the pleural ribs are epipleural intermusculars which are forked anteriorly. Epipleurals also appear to be associated with haemal arches as far back as the 36th centrum. There appear to be short, straight epicentral intermusculars associated with the more anterior abdominal centra. Anteriorly forked epineural intermusculars are associated with the neural arches as far back as the 36th centrum. The quality of caudal preservation in Apsopelix specimens is generally poor. It is fragmentary at best in the North American specimens, but Patterson and Rosen (1977) state that enough of it is visible in AMNH 8330 and in several British Museum specimens to



32 3 05 o 24
, differing only in having the neural and haemal spines more strongly inclined and pressed together plus increased hypurostegy so that the proximal ends of the upper and lower principal rays")


33 TELLER-MARSHALL & BARDACK: APSOPELIX 25 show that it is almost identical with that of Crossognathus (cf. Patterson and Rosen, 1977, fig. 21, p. 114), differing only in having the neural and haemal spines more strongly inclined and pressed together plus increased hypurostegy so that the proximal ends of the upper and lower principal rays almost meet. When compared with Patterson and Rosen's figure of the caudal skeleton of Crossognathus, the fragmentary caudal material from KU 309, KU 519, SMM 7958, and UT supports In Apsopelix uroneurals 1 their conclusions. and 2 extend forward to the second preural centrum and the third uroneural extends forward to the first preural centrum. There are, in addition to these elongate uroneurals, a posterior set of shorter uroneurals, although the exact number cannot be determined in the specimens examined. There are more than six hypurals, but the exact number is uncertain. (There are nine hypurals in Crossognathus.). The first and second hypurals are fused or partly fused in all of the specimens examined for this study. The parhypural is fused to the first preural centrum. It and the haemal spines of PU^ and PU 3 are broadly expanded. According to Patterson and Rosen (1977), there are caudal scutes present in Crossognathus. There is no evidence of caudal scutes in the North American specimens of Apsopelix examined in this study. DISCUSSION A number of distinctive morphological features in Apsopelix (the large scales, antorbitals and pelvic bones, the form and proportion of the dermal upper jaw elements, the arrangement and ornamentation of the cranial roof bones) have facilitated identification and the proper assignment of specimens previously described as Syllaemus, Pelycorapis, Leptichthys, Helmintholepis, and Paleoclupea. However, these features along with nearly all others known in Apsopelix have little value as clues to the position of this genus among the teleostean fishes. It exhibits a host of characters primitive for teleosts, a few characters which seem to be unique to the genus Apsopelix (posteriorly directed fossa on the hyomandibular, premaxilla so reduced as to be nearly lost, basihyal with a pair of tooth plates along its lateral margins only, antorbital which nearly covers the ethmoid region, broadly expanded pelvic bone), and no suite of characters that unequivocally indicate to which major teleost group it is most closely related. Of the major teleost groups, it is least likely that Apsopelix 's relationships are to be sought among the euteleosts. It was placed
.")

34 26 FIELDIANA: GEOLOGY, VOLUME 41 within the Percesoces 1 (Woodward, 1901) and Mugilidae (Stewart, 1900) on the mistaken assumption that the major element of the upper jaw was a premaxilla. Apsopelix shares none of the specializations unique to euteleosts (Patterson, 1970; Nelson, 1973a; Patterson and Rosen, 1977). Certain features present in Apsopelix were thought by previous investigators (Stewart, 1900; Woodward, 1903, 1907; Jordan, 1923; Chabanaud, 1930; Berg, 1940) to be indicative of a relationship with clupeomorphs. These include: 1) laterally compressed body, 2) form and position of fins, 3) features associated with microphagous feeding (reduced dentition on the dermal jaws; short, deep lower jaw; curved maxilla with a flat articular head; large, mobile supramaxillae; long gill rakers). Character 1 has proved to be either an artifact of preservation or an error of interpretation. Three-dimensionally preserved specimens of Apsopelix indicate that the body of this fish was ovoid or even pear-shaped in cross-section. This hypothesis is reinforced by the fact that the flattened specimens have been dorsoventrally compressed. A laterally compressed fish would be more likely to come to rest on its side and be preserved laterally compressed rather than dorsoventrally compressed as is the case in Apsopelix. Character 2, the form and position of the fins, is present in Apsopelix, clupeids, and engraulids. It is also present in elopids, albulids, and (with the addition of the adipose fin) salmonids, osmerids, and other lower teleosts. Thus, the form and position of the fins is an adaptation to mode of life and found in several primitive teleost groups. The third group of characters, those associated with microphagous feeding, form a functional complex to facilitate a feeding mode that developed a number of times in the course of teleost evolution. Among living teleosts the epibranchial organ, a structural adaptation to the microphagous feeding mode, has developed independently in at least four lineages of teleosts (Nelson, 1967). So it is reasonable to assume that the occurrence of this whole complex of characters, while valuable in reinforcing other evidence of a relationship, is not by itself sufficient to prove a connection between Apsopelix and clupeomorphs. Until recently two characters previously unknown in Apsopelix, the presence of epipleural intermusculars and a ventral keel on the 1 See footnote, p. 3
 and the keeled parasphenoid is characteristic of North American specimens of Pachyrhizodus, a genus which")


35 TELLER-MARSHALL & BARDACK: APSOPELIX 27 parasphenoid coupled with the extension of this bone posterior to the braincase, were also associated with clupeomorphs. Epipleural intermuscular bones are found at least in some osteoglossomorphs (Patterson, 1975) and the keeled parasphenoid is characteristic of North American specimens of Pachyrhizodus, a genus which Forey (1973b, 1977) associates with salmoniform euteleosts. Since these characters are not exclusive to clupeomorphs, it is unlikely that their presence in Apsopelix indicates clupeomorph affinities. In summary, Apsopelix has been associated previously with clupeomorphs on the basis of characters which can be shown to have been erroneously described, plesiomorphic for teleosts, or convergent. Derived characters shared by clupeomorphs include a swim-bladder-ear connection involving penetration of the exoccipital and prootic, a temporal foramen between the frontal and parietal in the apex of the posttemporal fossa (Greenwood et al., 1966), supratemporal commissure associated with the supraoccipital (Patterson, 1970), a pleurostylar caudal skeleton (Greenwood, 1968; Gosline, 1971; but see Forey, 1975), fusion of the angular and articular with the exclusion of the retroarticular from the articular facet (Nelson, 1973b), and fusion of tooth plates lb 1-3 and Cb 5 (Nelson, 1967). Apsopelix exhibits none of these characters so its relationships lie elsewhere. Wenz (1965) and Romer (1966) assigned Apsopelix to the Elopoidei. Apsopelix shares a large number of primitive features with living members of the elopiform fishes (see Forey, 1973b, pp ), but these characters are found in members of all major teleost groups and are of no use in determining relationships. Patterson and Rosen (1977) cite four shared, derived characters for the cohort Elopomorpha: a leptocepholus larva, angular and retroarticular bones fused, rostral and prenasal ossicles, and a compound neural arch formed in cartilage over PU1 and Ul. The first character is of no help in determining relationships of fossil fishes; the retroarticular is not fused with the angular in Apsopelix, and no rostral or prenasal ossicles are present in Apsopelix. The neural arches of the first ural and first preural centra are not preserved in the North American specimens of Apsopelix examined for this study, but in the similar Crossognathus there are separate neural arches over Ul and PU1. Patterson and Rosen (1977) cite some resemblances between the posttemporal fossae of Apsopelix and those of megalopids, but state that in Apsopelix the condition is not precisely the same as in

 suggest convergence of this character.")
36 28 FIELDIANA: GEOLOGY, VOLUME 41 recent megalopids. Other braincase similarities noted by Patterson and Rosen include the large myodome, shape of the intercalar, strongly ossified ethmoid region, and a deep pit in the lateral surface of the posterior part of the basioccipital suggesting the possibility of a simple otophysic connection. None of these similarities is convincing. Differences in the formation of the posttemporal fossae between megalopids and Apsopelix (see Patterson and Rosen, 1977 p. 132) suggest convergence of this character. The enlarged myodome characteristic of Apsopelix and megalopids generally occurs in conjunction with a deep neurocranium and is not exclusive to megalopids. The intercalar of North American specimens of Apsopelix does not resemble that of Eocene megalopids (cf. figs. 3, 4 with Forey, 1973b, figs. 36, 37, 42, 43) as stated by Patterson and Rosen (1977). It is a proportionately smaller bone in Apsopelix without the posterior expansion present in Eocene megalopids. A heavily ossified mesethmoid is primitive that an for teleosts (Patterson, 1970). Finally, it seems likely otophysic connection per se must be looked upon as a primitive teleostean character (Greenwood, 1973). In summary, there are no derived characters of elopomorphs shared with Apsopelix. Characters of the braincase shared by megalopids and Apsopelix can be more reasonably explained as convergent or primitive for teleosts, since the structure of the lower jaw and caudal skeleton of Apsopelix opposes elopomorph relationships. Thus, the probability of finding relationships of Apsopelix within the elopomorphs fades. Part of the problem in comparing Apsopelix to the osteoglossomorphs is that this group is so diverse. Among those derived characters which are known in at least some osteoglossomorphs (see Greenwood et al., 1966, 1967; Greenwood, 1967, 1970, 1971, 1973; Nelson, 1968, 1969, 1972) there is only one which appears to be shared with Apsopelix. That character, fusion of Io (Nelson, 1969), has yet to be firmly established as present in Apsopelix. Given that there is a fusion of Io in Apsopelix, this character alone (found also in Chanos and cypriniformes) is not sufficient to indicate a relationship. One similarity between Apsopelix and at least some fossil osteoglossomorphs could be the shape and disposition of the premaxilla. Judging from the relationship of the maxilla to the median ethmoid in Apsopelix, the premaxilla probably overlapped the anterior end of the maxilla and extended down over it to form the


 place Apsopelix with Crossognathus as a nominal family incertae sedis within the group containing")
37 TELLER-MARSHALL & BARDACK: APSOPELIX 29 anteriormost border of the upper jaw as in Ichthyodectes ctenodon and Xiphactinus audax (see Bardack, 1965, fig. 16, p. 58, fig. 9, p. 45). These three fishes also share an ossified malleolar process on the anterior end of the palatine, but this ossification appears to be a response to a functional need and is also present in some members of the elopiforms (Forey, 1973b). This discussion has reviewed the four major teleost groups and argued against assigning Apsopelix to any one of these groups. The original hope of the authors was that more information concerning the morphology of Apsopelix would facilitate the discovery of its systematic relationships. The accumulation of this information has instead precipitated arguments against a systematic assignment of Apsopelix to any presently well-defined group. In their analysis of the relationships of fossil and Recent teleosts, Patterson and Rosen (1977) place Apsopelix with Crossognathus as a nominal family incertae sedis within the group containing Tharsis dubius and extant teleosts. That is, Apsopelix shares with Tharsis dubius the three features it 1 exhibits which are synapomorphies with Recent teleosts, but does not show any derived characters typical of a Recent teleost group. We agree with Patterson and Rosen (1977) and feel that it is preferable that Apsopelix be left incertae sedis rather than erroneously associated with a teleost group on the basis of convergent or plesiomorphous characters. GEOLOGIC AND GEOGRAPHIC RANGE The oldest Apsopelix specimen known is a single fish from the Albian of France (Wenz, 1965). By Cenomanian time Apsopelix ranged throughout the North Temperate Euroamerican Region (Kauffman, 1973). It apparently did not enter other regions. Morphological features of Apsopelix remain constant throughout its geologic as well as geographic range. The Late Cretaceous specimens from the Western Interior do not differ significantly from the English, French (Patterson and Rosen, 1977), or Gulf Coast specimens. The morphological stability of Apsopelix in time supports the idea of punctuated equilibria (Eldridge and Gould, 1972; Gould and Eldrige, 1977), the concept of speciation as a rare event that punc- 'These three features are: loss of suborbital bones, presence of epipleural intermuscular bones, and uroneurals distributed in two series, i.e., the uppermost uroneurals overlap and he at an angle to the longer anterior ones.



38 30 FIELDIANA: GEOLOGY, VOLUME 41 tuates a system in homeostatic equilibrium. Essentially, we have only the stable stage of evolution in this group. The geographic distribution of Apsopelix coupled with the morphological similarity of individuals throughout its range is reasonable when viewed from the perspective of the configuration of continents and oceans in Cretaceous times (see Kauffman, 1973, fig. 2, p. 356). The greater proximity of the European and American continents during the Jurassic and Early Cretaceous combined with a similar biologic source area in the continuous north temperate seaway resulted in a close similarity of Northern European and North American bivalve faunas (Kauffman, 1973) and would support the similarity of Northern European and North American Apsopelix. These fishes appear to have been specifically adapted to interior marine environments 1 of the north temperate realm, to have maintained a set of morphological adaptations to this type of environment through time, and to have become extinct with the disappearance of this type of environment. Ang apl Ao Art Asp Bsp Boc Den df Dsp Ecpt Enpt Epo Exo fhm fhmvii ABBREVIATIONS angular anterior pit line antorbital articular autosphenotic basisphenoid basioccipital dentary dilatator fossa dermosphenotic ectopterygoid endopterygoid epioccipital exoccipital hyomandibular facet foramen for hyomandibular trunk of facial nerve 'Specimens from Texas localities near the Western Interior/Gulf Atlantic Coast boundary are found with invertebrates and in sediments that are characteristic of interior marine rather than coastal shelf environments.



39 fhv TELLER-MARSHALL & BARDACK: APSOPELIX 31


40 32 FIELDIANA: GEOLOGY, VOLUME 41 Bardack, David REFERENCES Anatomy and evolution of Chirocentrid fishes. Paleontol. Contr. Univ. Kansas, 40, pp Berg.L. S Classification of fishes both recent and fossil. Trudy Zool. Inst., Leningrad 5,2, 517 pp. Chabanaud, P Sur un poisson telesteen du Turonien d'indre-et-loire. Extrait Bull. Soc. geol. France, 4 e ser., t. XXX, pp COCKERELL.T.D.A Some American Cretaceous fish scales. U.S. Geol. Surv. Prof. Pap. 120, pp , pis Cope.E.D On the fossil reptiles and fishes of the Cretaceous rocks of Kansas. U.S. Geol. Surv. Wyoming and Portions of the Contiguous Territories, 2nd Ann. Rept., pp The vertebrata of the Cretaceous formations of the West. Rept. U.S. Geol. Surv. Terr., no. 2, pp , pis. 55, 10 figs Report on the geology of the region of the Judith River, Montana, and on vertebrate fossils obtained on or near the Missouri River. Bull. U.S. Geol. Geog. Surv., 3, (art. 29), pp , 5 pis. Cragin.F.W A study of some teleosts from the Russell Substage of the Platte Cretaceous Series. Col. College Stud., 9, pp , pis Dante, J. H Description of fossil fishes from the Upper Cretaceous of North America. Amer. Jour. Sci., 240, pp , 2 pis. Dixon, D.H The geology and fossils of the Tertiary and Cretaceous formations of Sussex. F. Dixon, London, 422 pp. 40 pis. Dunkle.D.H Three North American Cretaceous fishes. Proc. U.S. Nat. Mus., 108, no. 3401, pp Eldredge, N., and S. J. Gould Punctuated equilibria: an alternative to phyletic gradualism, pp In: Schopf, T. J. M., ed., Models in paleobiology. Freeman, Cooper and Co., San Francisco, Calif. Forey, P. L. 1973a. A primitive Clupeomorph fish from the Middle Cenomanian of Hakel, Lebanon. Canad. Jour. Earth Sci., 10, no. 8, pp. 1,302-1, b. A revision of the elopiform fishes fossil and recent. Bull. Brit. Mus. (Nat. Hist.) Geol, London, Supp. 10, 222 pp., 92 figs.
. Bull. Brit. Mus. (Nat. Hist.) Geol., London, 28, no. 2, pp.")


41 TELLER-MARSHALL & BARDACK: APSOPELIX A fossil clupeomorph fish from the Albian of the Northwest Territories of Canada, with notes on cladistic relationships of clupeomorphs. Jour. Zool., London, pp The osteology of Notelops Woodward, Rhacolepis Agassiz and Pachyrhizodus Dixon (Pisces: Teleostei). Bull. Brit. Mus. (Nat. Hist.) Geol., London, 28, no. 2, pp , 39 figs. Gardiner, B.G Certain palaenoiscoid fishes and the evolution of the snout in actinopterygians. Bull. Brit. Mus. Nat. Hist. (Geol.), London, 8, pp , 2 pis., 20 figs. Gosline, W. A Some osteological features of modern lower teleostean fishes. Smithson. Misc. Coll., Washington, 142, 3, pp. 1-42, 8 figs Functional morphology and classification of teleostean fishes. University Press of Hawaii, Honolulu, pp. ix-208. Gould, S. J. and N. Eldredge Punctuated equilibria: the tempo and mode of evolution reconsidered. Paleobiology, 3, no. 2, pp Green, M Syllaemus from the Carlile Formation of South Dakota. Proc. South Dakota Acad. Sci., XXXVI, pp Greenwood, P. H The caudal fin skeleton in osteoglossoid fishes. Ann. Mag. Nat. Hist., (13) 9, pp The osteology and relationships of the Denticipitidae, a family of clupeomorph fishes. Bull. Brit. Mus. (Nat. Hist.) Zool, London, 16, pp On the genus Lycoptera and its relationship with the Family Hiodontidae (Pisces, Osteoglossomorpha). Bull. Brit. Mus., Nat. Hist. (Zool.), London, 19, pp Hyoid and ventral gill arch musculature in osteoglossomorph fishes. Bull. Brit. Mus. Nat. Hist. (Zool.), London, 22, pp Interrelationships of osteoglossomorphs, pp In: Greenwood, P. H., R. S. Miles, and C. Patterson eds.. Interrelationships of fishes, Academic Press, Supp. No. 1, Zool. Jour. Linnean Soc., London, vol. 53. Greenwood, P. H., G. S. Myers. D. E. Rosen, and S. H. Weitzman Named main divisions of teleostean fishes. Proc. Biol. Soc., Washington, 80, pp Greenwood, P. H., D. E. Rosen, S. H. Weitzman, and G. S. Myers Phyletic studies of teleostean fishes with a provisional classification of living forms. Bull. Amer. Mus. Nat. Hist., 131, pp , pi , 32 charts. Jordan, D. S A classification of fishes including families and genera as far as known. Standford Univ. Publ., Biol. Sci., Palo Alto, 3, pp A collection of fossil fishes in the University of Kansas from the Niobrara Formation of the Cretaceous. Kansas Univ. Sci. Bull., XV, no. 2, pp


42 34 FIELDIANA: GEOLOGY, VOLUME 41 Kauffman, E. G Cretaceous bivalvia, pp In: Hallam A., ed., Atlas of paleobiogeography, Elsevier Co., Amsterdam. Kirchhoff, H Funktionell-antatomische Untersuchung des Visceralapparates von Clupea harengus, L. Zool. Jb. Anat. Band 76, Heft 4, pp , Germany. Nelson, G. J Gill arches of teleostean fishes of the Family Clupeidae. Copeia, pp GUI arches of some teleostean fishes of the Division Osteoglossomorpha. Jour. Linn. Soc. (Zool.), 47, pp Infraorbital bones and their bearing on the phylogeny and geography of osteoglossomorph fishes. Amer. Mus. Novit., Observations on the gut of the Osteoglossomorpha. Copeia, pp a. Notes on the structure and relationships of certain Cretaceous and Eocene fishes. Amer. Mus. Novit., b. Relationships of clupeomorphs, with remarks on the structure of the lower jaw in fishes, pp In: Greenwood, P. H., R. S. Miles, and C. Patterson, eds., Interrelationships of fishes, Academic Press, Supp. No. 1, Zool. Jour. Linnean Soc, London, vol. 53. Nybelin, O Notes on the reduction of the sensory canal system and of the canal-bearing bones in the snout of higher actinopterygian fishes. Ark. Zool., Stockholm, (2) 19, pp , 4 figs. Patterson, C Two Upper Cretaceous salmoniform fishes from Lebanon. Bull. Brit. Mus. (Nat. Hist.) Geol, London, 19, pp , 48 figs., 5 pis Interrelationships of holosteans, pp In: Greenwood, P.H., R.S. Miles, and C. Patterson, eds., Interrelationships of fishes. Academic Press, Supp. No. 1, Zool. Jour. Linnean Soc, London, vol. 53. Patterson, C. and D. E. Rosen Review of Ichthyodectiform and other Mesozoic teleost fishes and the theory and practice of classifying fossils. Bull. Amer. Mus. Nat. Hist., 158, art. 2, pp , 53 figs. ROMER, A. S Vertebrate paleontology. Univ. Chicago Press, 3rd ed., pp. viii pp., 433 figs. Stewart, A Leptichthys, a new genus of fishes from the Cretaceous of Kansas. Amer. Geol, 24, pp Teleosts of the Upper Cretaceous. Kansas Univ. Geol. Surv., 6, pt. 2, pp , 40 pis., 6 figs. Wenz, S Les poissons Albiens de Vallantigny (Aube). Ann. Paleontol., fasc. 1, pp. 3-23, 2 pis.

 London, 4, pp.")
 London, pp.")
43 TELLER-MARSHALL & BARDACK: APSOPELIX 35 Woodward, A. S A synopsis of the vertebrate fossils of the English chalk. Proc. Geol. Assn., London, 10, pp Catalogue of the fossil fishes in the British Museum (Natural History) London, 4, pp. xxxviii pages, 19 pis., 22 figs Fossil fishes of the English chalk, Part II. Palaeontogr. Soc. (Monogr.) London, pp , pis. XIV-XX Fossil fishes of the English chalk, Part III. Palaeontogr. Soc. (Monogr.) London, pp , pis. XXI-XXVI.
44

45



46
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
REVISION OF THE GENUS MARTINICHTHYS, MARINE FISH (TELESOSTEI, TSELFATIIFORMES) FROM THE LATE CRETACEOUS OF KANSAS (UNITED STATES)
 FROM THE LATE CRETACEOUS OF KANSAS (UNITED STATES)") 1 REVISION OF THE GENUS MARTINICHTHYS, MARINE FISH (TELESOSTEI, TSELFATIIFORMES) FROM THE LATE CRETACEOUS OF KANSAS (UNITED STATES) TAVERNE L., 2000. Revision of the genus Martinichthys, marine fish (Teleostei,
1 REVISION OF THE GENUS MARTINICHTHYS, MARINE FISH (TELESOSTEI, TSELFATIIFORMES) FROM THE LATE CRETACEOUS OF KANSAS (UNITED STATES) TAVERNE L., 2000. Revision of the genus Martinichthys, marine fish (Teleostei,
Osteology of the Clupeiform fish, genus Hyperlophus (II)
") Bull. Kitakyushu Mas. Nat. Hist., 4: 77-102. December 31, 1982 Osteology of the Clupeiform fish, genus Hyperlophus (II) Yoshitaka Yabumoto Kitakyushu Museum of Natural History, Nishihonmachi, Yahatahigashiku,
Bull. Kitakyushu Mas. Nat. Hist., 4: 77-102. December 31, 1982 Osteology of the Clupeiform fish, genus Hyperlophus (II) Yoshitaka Yabumoto Kitakyushu Museum of Natural History, Nishihonmachi, Yahatahigashiku,
VERTEBRATA PALASIATICA
 VERTEBRATA PALASIATICA ONLINE SUPPLEMENTARY MATERIAL Panxianichthys imparilis gen. et sp. nov., a new ionoscopiform (Halecomorphi) from the Middle Triassic of Guizhou Province, China XU Guang-Hui 1,2 SHEN
VERTEBRATA PALASIATICA ONLINE SUPPLEMENTARY MATERIAL Panxianichthys imparilis gen. et sp. nov., a new ionoscopiform (Halecomorphi) from the Middle Triassic of Guizhou Province, China XU Guang-Hui 1,2 SHEN
AMERICAN MUSEUM NOVITATES Published by
 AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province
 from Dashanpu, Zigong Municipality, Sichuan Province") A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
ONLINE APPENDIX 1. Morphological phylogenetic characters scored in this paper. See Poe (2004) for
 for") ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE
 PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
2. Skull, total length versus length of the presacral vertebral column: (0); extremely elongated neck (e.g. Tanystropheus longobardicus).
; extremely elongated neck (e.g. Tanystropheus longobardicus).") Character list of the taxon-character data set 1. Skull and lower jaws, interdental plates: absent (0); present, but restricted to the anterior end of the dentary (1); present along the entire alveolar
Character list of the taxon-character data set 1. Skull and lower jaws, interdental plates: absent (0); present, but restricted to the anterior end of the dentary (1); present along the entire alveolar
A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA
 A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA MYCTEROSAURUS LONGICEPS S. W. WILLISTON University of Chicago The past summer, Mr. Herman Douthitt, of the University of Chicago paleontological expedition,
A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA MYCTEROSAURUS LONGICEPS S. W. WILLISTON University of Chicago The past summer, Mr. Herman Douthitt, of the University of Chicago paleontological expedition,
PALEONTOLOGICAL CONTRIBUTIONS'
 THE UNIVERSITY OF KANSAS PALEONTOLOGICAL CONTRIBUTIONS' August, 1965 Paper 1 NEW UPPER CRETACEOUS TELEOST FISH FROM TEXAS DAVID BARDACK Department of Biological Sciences, University of Illinois at Chicago
THE UNIVERSITY OF KANSAS PALEONTOLOGICAL CONTRIBUTIONS' August, 1965 Paper 1 NEW UPPER CRETACEOUS TELEOST FISH FROM TEXAS DAVID BARDACK Department of Biological Sciences, University of Illinois at Chicago
SUPPLEMENTARY INFORMATION
 Character 155, interdental ridges. Absence of interdental ridge (0) shown in Parasaniwa wyomingensis (Platynota). Interdental ridges (1) shown in Coniophis precedens. WWW.NATURE.COM/NATURE 1 Character
Character 155, interdental ridges. Absence of interdental ridge (0) shown in Parasaniwa wyomingensis (Platynota). Interdental ridges (1) shown in Coniophis precedens. WWW.NATURE.COM/NATURE 1 Character
Anatomy. Name Section. The Vertebrate Skeleton
 Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Osteology and Relationships of the Eel Diastobranchus capensis (Pisces, Synaphobranchidae) I
 I") Pacific Science (1975), Vol. 29, No.2, p. 159-163 Printed in Great Britain Osteology and Relationships of the Eel Diastobranchus capensis (Pisces, Synaphobranchidae) I P. H. J. CASTLE2 ABSTRACT: An osteological
Pacific Science (1975), Vol. 29, No.2, p. 159-163 Printed in Great Britain Osteology and Relationships of the Eel Diastobranchus capensis (Pisces, Synaphobranchidae) I P. H. J. CASTLE2 ABSTRACT: An osteological
HONR219D Due 3/29/16 Homework VI
 Part 1: Yet More Vertebrate Anatomy!!! HONR219D Due 3/29/16 Homework VI Part 1 builds on homework V by examining the skull in even greater detail. We start with the some of the important bones (thankfully
Part 1: Yet More Vertebrate Anatomy!!! HONR219D Due 3/29/16 Homework VI Part 1 builds on homework V by examining the skull in even greater detail. We start with the some of the important bones (thankfully
SUPPLEMENTARY ONLINE MATERIAL FOR. Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor
 http://app.pan.pl/som/app61-ratsimbaholison_etal_som.pdf SUPPLEMENTARY ONLINE MATERIAL FOR Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor Ontogenetic changes in the craniomandibular
http://app.pan.pl/som/app61-ratsimbaholison_etal_som.pdf SUPPLEMENTARY ONLINE MATERIAL FOR Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor Ontogenetic changes in the craniomandibular
CRANIAL OSTEOLOGY OF SCHIZOTHORAICHTHYS NIGER (MECKEL) MISRA (CYPRINIDAE: SCHIZOTHORACINAE). L NEUROCRANIUM
 MISRA (CYPRINIDAE: SCHIZOTHORACINAE). L NEUROCRANIUM") CRANIAL OSTEOLOGY OF SCHIZOTHORAICHTHYS NIGER (MECKEL) MISRA (CYPRINIDAE: SCHIZOTHORACINAE). L NEUROCRANIUM A. R. YousuF, A. K. PANDIT AND A. R. KHAN Postgraduate Department of Zoology, University of Kashmir,
CRANIAL OSTEOLOGY OF SCHIZOTHORAICHTHYS NIGER (MECKEL) MISRA (CYPRINIDAE: SCHIZOTHORACINAE). L NEUROCRANIUM A. R. YousuF, A. K. PANDIT AND A. R. KHAN Postgraduate Department of Zoology, University of Kashmir,
YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
 Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
v:ii-ixi, 'i':;iisimvi'\>!i-:: "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi L I E) R.ARY OF THE VERSITY U N I or ILLINOIS REMO
 R.ARY OF THE VERSITY U N I or ILLINOIS REMO") "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
"^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to
 1 Supplementary data CHARACTER LIST List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to characters used by Tchernov et al. (2000), Rieppel, et al. (2002), and Lee
1 Supplementary data CHARACTER LIST List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to characters used by Tchernov et al. (2000), Rieppel, et al. (2002), and Lee
TWO THREE-DIMENSIONALLY PRESERVED TELEOST NEUROCRANIA FROM THE CORSICANA FORMATION (UPPER CRETACEOUS, MAESTRICHTIAN), BEXAR COUNTY, TEXAS, U.S.A.
, BEXAR COUNTY, TEXAS, U.S.A.") Paludicola 10(3):1.37-144 September 2015 by the Rochester Institute of Vertebrate Paleontology TWO THREE-DIMENSIONALLY PRESERVED TELEOST NEUROCRANIA FROM THE CORSICANA FORMATION (UPPER CRETACEOUS, MAESTRICHTIAN),
Paludicola 10(3):1.37-144 September 2015 by the Rochester Institute of Vertebrate Paleontology TWO THREE-DIMENSIONALLY PRESERVED TELEOST NEUROCRANIA FROM THE CORSICANA FORMATION (UPPER CRETACEOUS, MAESTRICHTIAN),
Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons
 Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons 1. Head skeleton of lamprey Cyclostomes are highly specialized in both the construction of the chondrocranium and visceral skeleton.
Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons 1. Head skeleton of lamprey Cyclostomes are highly specialized in both the construction of the chondrocranium and visceral skeleton.
THE SKULLS OF ARAEOSCELIS AND CASEA, PERMIAN REPTILES
 THE SKULLS OF REOSCELIS ND CSE, PERMIN REPTILES University of Chicago There are few Permian reptiles of greater interest at the present time than the peculiar one I briefly described in this journal' three
THE SKULLS OF REOSCELIS ND CSE, PERMIN REPTILES University of Chicago There are few Permian reptiles of greater interest at the present time than the peculiar one I briefly described in this journal' three
Published in "Zoological Journal of the Linnean Society 181 (3): , 2017" which should be cited to refer to this work.
: , 2017 which should be cited to refer to this work.") Published in "Zoological Journal of the Linnean Society 181 (3): 604 637, 2017" which should be cited to refer to this work. A new genus and species for the amiiform fishes previously assigned to Amiopsis
Published in "Zoological Journal of the Linnean Society 181 (3): 604 637, 2017" which should be cited to refer to this work. A new genus and species for the amiiform fishes previously assigned to Amiopsis
.56 m. (22 in.). COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition
. COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition") Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
THE family Carangidae (jacks, trevallies, pompanos,
 Copeia 2010, No. 2, 312 333 Osteology and Systematics of Parastromateus niger (Perciformes: Carangidae), with Comments on the Carangid Dorsal Gill-Arch Skeleton Eric J. Hilton 1, G. David Johnson 2, and
Copeia 2010, No. 2, 312 333 Osteology and Systematics of Parastromateus niger (Perciformes: Carangidae), with Comments on the Carangid Dorsal Gill-Arch Skeleton Eric J. Hilton 1, G. David Johnson 2, and
Mammalogy Laboratory 1 - Mammalian Anatomy
 Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia
 1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the
 Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the") Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO
 A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC
 FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
TRACHEMYS SCULPTA. A nearly complete articulated carapace and plastron of an Emjdd A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE,
 A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
FIELDIANA GEOLOGY NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA
 FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
by Louis TAVERNE Abstract Résumé Introduction
 BULLETIN DE L INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETENSCHAPPEN SCIENCES DE LA TERRE, 78: 209-228, 2008 AARDWETENSCHAPPEN, 78: 209-228,
BULLETIN DE L INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETENSCHAPPEN SCIENCES DE LA TERRE, 78: 209-228, 2008 AARDWETENSCHAPPEN, 78: 209-228,
A Fossil Snake (Elaphe vulpina) From A Pliocene Ash Bed In Nebraska
 From A Pliocene Ash Bed In Nebraska") University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
complex in cusp pattern. (3) The bones of the coyote skull are thinner, crests sharper and the
 The bones of the coyote skull are thinner, crests sharper and the") DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
Williston, and as there are many fairly good specimens in the American
 56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024
 PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3455, 21 pp., 10 figures, 1 table October 28, 2004 Redescription of Santanichthys diasii
PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3455, 21 pp., 10 figures, 1 table October 28, 2004 Redescription of Santanichthys diasii
Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved
 Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved This was a private report in 2003 on my thoughts on Platecarpus planifrons.
Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved This was a private report in 2003 on my thoughts on Platecarpus planifrons.
AKROMYSTAX TILMACHITON GEN. ET SP. NOV., A NEW PYCNODONTID FISH FROM THE LEBANESE LATE CRETACEOUS OF HAQEL AND EN NAMMOURA
 Journal of Vertebrate Paleontology 25(1):27 45, March 2005 2005 by the Society of Vertebrate Paleontology AKROMYSTAX TILMACHITON GEN. ET SP. NOV., A NEW PYCNODONTID FISH FROM THE LEBANESE LATE CRETACEOUS
Journal of Vertebrate Paleontology 25(1):27 45, March 2005 2005 by the Society of Vertebrate Paleontology AKROMYSTAX TILMACHITON GEN. ET SP. NOV., A NEW PYCNODONTID FISH FROM THE LEBANESE LATE CRETACEOUS
A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.)
") A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.) by Ouyang Hui Zigong Dinosaur Museum Newsletter Number 2 1989 pp. 10-14 Translated By Will Downs Bilby
A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.) by Ouyang Hui Zigong Dinosaur Museum Newsletter Number 2 1989 pp. 10-14 Translated By Will Downs Bilby
The cranial osteology of Belebey vegrandis (Parareptilia: Bolosauridae), from the Middle Permian of Russia, and its bearing on reptilian evolution
, from the Middle Permian of Russia, and its bearing on reptilian evolution") Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1511 191214 Original Articles RUSSIAN BOLOSAURID REPTILER. R. REISZ ET AL.
Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1511 191214 Original Articles RUSSIAN BOLOSAURID REPTILER. R. REISZ ET AL.
Cranial osteology of the African gerrhosaurid Angolosaurus skoogi (Squamata; Gerrhosauridae) HOLLY A. NANCE
 HOLLY A. NANCE") African Journal of Herpetology, 2007 56(1): 39-75. Herpetological Association of Africa Original article Cranial osteology of the African gerrhosaurid Angolosaurus skoogi (Squamata; Gerrhosauridae) HOLLY
African Journal of Herpetology, 2007 56(1): 39-75. Herpetological Association of Africa Original article Cranial osteology of the African gerrhosaurid Angolosaurus skoogi (Squamata; Gerrhosauridae) HOLLY
A new species of sauropod, Mamenchisaurus anyuensis sp. nov.
 A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
Description of Cranial Elements and Ontogenetic Change within Tropidolaemus wagleri (Serpentes: Crotalinae).
.") East Tennessee State University Digital Commons @ East Tennessee State University Electronic Theses and Dissertations 5-2016 Description of Cranial Elements and Ontogenetic Change within Tropidolaemus
East Tennessee State University Digital Commons @ East Tennessee State University Electronic Theses and Dissertations 5-2016 Description of Cranial Elements and Ontogenetic Change within Tropidolaemus
Osteological description of Barbus lacerta Heckel, 1843 (Cyprinidae) from Tigris basin of Iran
 from Tigris basin of Iran") 2016; 4(4): 473-477 E-ISSN: 2320-7078 P-ISSN: 2349-6800 JEZS 2016; 4(4): 473-477 2016 JEZS Received: 18-05-2016 Accepted: 19-05-2016 Nasrin Nikmehr Soheil Eagderi Pariya Jalili Osteological description
2016; 4(4): 473-477 E-ISSN: 2320-7078 P-ISSN: 2349-6800 JEZS 2016; 4(4): 473-477 2016 JEZS Received: 18-05-2016 Accepted: 19-05-2016 Nasrin Nikmehr Soheil Eagderi Pariya Jalili Osteological description
Comparative Osteology of the Genus Pachytriton (Caudata: Salamandridae) from Southeastern China
 from Southeastern China") Asian Herpetological Research 2012, 3(2): 83 102 DOI: 10.3724/SP.J.1245.2012.00083 Comparative Osteology of the Genus Pachytriton (Caudata: Salamandridae) from Southeastern China Yunke WU 1, Yuezhao WANG
Asian Herpetological Research 2012, 3(2): 83 102 DOI: 10.3724/SP.J.1245.2012.00083 Comparative Osteology of the Genus Pachytriton (Caudata: Salamandridae) from Southeastern China Yunke WU 1, Yuezhao WANG
Vol. XIV, No. 1, March, The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S.
 By S.") Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Descriptive anatomy of Iso rhothophilus (Ogilby), with a phylogenetic analysis of Iso and a redefinition of Isonidae (Atheriniformes)
, with a phylogenetic analysis of Iso and a redefinition of Isonidae (Atheriniformes)") aqua, Journal of Ichthyology and quatic Biology Descriptive anatomy of Iso rhothophilus (Ogilby), with a phylogenetic analysis of Iso and a redefinition of Isonidae (theriniformes) Basim Saeed, Walter
aqua, Journal of Ichthyology and quatic Biology Descriptive anatomy of Iso rhothophilus (Ogilby), with a phylogenetic analysis of Iso and a redefinition of Isonidae (theriniformes) Basim Saeed, Walter
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/329/5998/1481/dc1 Supporting Online Material for Tyrannosaur Paleobiology: New Research on Ancient Exemplar Organisms Stephen L. Brusatte,* Mark A. Norell, Thomas D.
www.sciencemag.org/cgi/content/full/329/5998/1481/dc1 Supporting Online Material for Tyrannosaur Paleobiology: New Research on Ancient Exemplar Organisms Stephen L. Brusatte,* Mark A. Norell, Thomas D.
CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN OF RUSSIA AND THE EVOLUTIONARY RELATIONSHIPS OF CASEIDAE
 FROM THE MIDDLE PERMIAN OF RUSSIA AND THE EVOLUTIONARY RELATIONSHIPS OF CASEIDAE") Journal of Vertebrate Paleontology 28(1):160 180, March 2008 2008 by the Society of Vertebrate Paleontology ARTICLE CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN
Journal of Vertebrate Paleontology 28(1):160 180, March 2008 2008 by the Society of Vertebrate Paleontology ARTICLE CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN
A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev Doklady Akademii Nauk, SSSR 87:
 translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province
 A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province by Hu Shaojin (Kunming Cultural Administrative Committee, Yunnan Province) Vertebrata PalAsiatica Vol. XXXI, No. 1
A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province by Hu Shaojin (Kunming Cultural Administrative Committee, Yunnan Province) Vertebrata PalAsiatica Vol. XXXI, No. 1
ABBREVIATIONS 2 AMERICAN MUSEUM NOVITATES
 2 AMERICAN MUSEUM NOVITATES NO. 3324 INTRODUCTION Recent discoveries of mid-cretaceous fossil fish assemblages in Venezuela (Moody and Maisey, 1993) and Colombia (Schultze and Stöhr, 1996) are of biogeographic
2 AMERICAN MUSEUM NOVITATES NO. 3324 INTRODUCTION Recent discoveries of mid-cretaceous fossil fish assemblages in Venezuela (Moody and Maisey, 1993) and Colombia (Schultze and Stöhr, 1996) are of biogeographic
SOME NEW AMERICAN PYCNODONT FISHES.
 SOME NEW AMERICAN PYCNODONT FISHES. By James Williams Gidley, Assistant Curator of Fossil Mammals, United States National Museum. In the United States National Museum are several specimens representing
SOME NEW AMERICAN PYCNODONT FISHES. By James Williams Gidley, Assistant Curator of Fossil Mammals, United States National Museum. In the United States National Museum are several specimens representing
A M E G H I N I A N A. Revista de la Asociación Paleontológia Argentina. Volume XV September-December 1978 Nos. 3-4
 A M E G H I N I A N A Revista de la Asociación Paleontológia Argentina Volume XV September-December 1978 Nos. 3-4 COLORADIA BREVIS N. G. ET N. SP. (SAURISCHIA, PROSAUROPODA), A PLATEOSAURID DINOSAUR FROM
A M E G H I N I A N A Revista de la Asociación Paleontológia Argentina Volume XV September-December 1978 Nos. 3-4 COLORADIA BREVIS N. G. ET N. SP. (SAURISCHIA, PROSAUROPODA), A PLATEOSAURID DINOSAUR FROM
Gladiopycnodontidae, a new family of pycnodontiform fishes from the Late Cretaceous of Lebanon, with the description of three genera
 European Journal of Taxonomy 57: 1-30 ISSN 2118-9773 http://dx.doi.org/10.5852/ejt.2013.57 www.europeanjournaloftaxonomy.eu 2013 Taverne L. & Capasso L. This work is licensed under a Creative Commons Attribution
European Journal of Taxonomy 57: 1-30 ISSN 2118-9773 http://dx.doi.org/10.5852/ejt.2013.57 www.europeanjournaloftaxonomy.eu 2013 Taverne L. & Capasso L. This work is licensed under a Creative Commons Attribution
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A.
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
PALEONTOLOGY AND BIOSTRATIGRAPHY OF MONGOLIA
 PALEONTOLOGY AND BIOSTRATIGRAPHY OF MONGOLIA THE JOINT SOVIET-MONGOLIAN PALEONTOLOGICAL EXPEDITION (Transactions, vol. 3) EDITORIAL BOARD: N. N. Kramarenko (editor-in-chief) B. Luvsandansan, Yu. I. Voronin,
PALEONTOLOGY AND BIOSTRATIGRAPHY OF MONGOLIA THE JOINT SOVIET-MONGOLIAN PALEONTOLOGICAL EXPEDITION (Transactions, vol. 3) EDITORIAL BOARD: N. N. Kramarenko (editor-in-chief) B. Luvsandansan, Yu. I. Voronin,
PARMOPS CORUSCANS, A NEW GENUS AND SPECIES OF FLASHLIGHT FISH (BERYCIFORMES: ANOMALOPIDAE) FROM THE SOUTH PACIFIC
 FROM THE SOUTH PACIFIC") 18 June 1991 PROC. BIOL. SOC. WASH. 104(2), 1991, pp. 328-334 PARMOPS CORUSCANS, A NEW GENUS AND SPECIES OF FLASHLIGHT FISH (BERYCIFORMES: ANOMALOPIDAE) FROM THE SOUTH PACIFIC Richard H. Rosenblatt and
18 June 1991 PROC. BIOL. SOC. WASH. 104(2), 1991, pp. 328-334 PARMOPS CORUSCANS, A NEW GENUS AND SPECIES OF FLASHLIGHT FISH (BERYCIFORMES: ANOMALOPIDAE) FROM THE SOUTH PACIFIC Richard H. Rosenblatt and
A NEW SPECIES OF TROODONT DINOSAUR FROM THE
 A NEW SPECIES OF TROODONT DINOSAUR FROM THE LANCE FORMATION OF WYOMING By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION The intensive search to which
A NEW SPECIES OF TROODONT DINOSAUR FROM THE LANCE FORMATION OF WYOMING By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION The intensive search to which
BREVIORA LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1
, A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1") ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
A skull without mandihle, from the Hunterian Collection (no.
 4 MR. G. A. BOULENGER ON CHELONIAN REMAINS. [Jan. 6, 2. On some Chelonian Remains preserved in the Museum of the Eojal College of Surgeons. By G. A. Boulenger. [Eeceived December 8, 1890.] In the course
4 MR. G. A. BOULENGER ON CHELONIAN REMAINS. [Jan. 6, 2. On some Chelonian Remains preserved in the Museum of the Eojal College of Surgeons. By G. A. Boulenger. [Eeceived December 8, 1890.] In the course
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS
 AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.
PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024
 PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3383, 43 pp., 21 figures, 3 color plates December 27, 2002 Paradox Lost : Skeletal Ontogeny
PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3383, 43 pp., 21 figures, 3 color plates December 27, 2002 Paradox Lost : Skeletal Ontogeny
Cranial osteology and phylogenetic relationships of Hamadasuchus rebouli (Crocodyliformes: Mesoeucrocodylia) from the Cretaceous of Morocco
 from the Cretaceous of Morocco") Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1494 533567 Original Articles HAMADASUCHUS REBOULIH. C. E. LARSSON and H.-D.
Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1494 533567 Original Articles HAMADASUCHUS REBOULIH. C. E. LARSSON and H.-D.
A New Ceratopsian Dinosaur from the Upper
 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 63. NUMBER 3 A New Ceratopsian Dinosaur from the Upper Cretaceous of Montana, with Note on Hypacrosaurus (With Two Plates) CHARLES W. GILMORE Assistant Curator
SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 63. NUMBER 3 A New Ceratopsian Dinosaur from the Upper Cretaceous of Montana, with Note on Hypacrosaurus (With Two Plates) CHARLES W. GILMORE Assistant Curator
Supplementary Information (ZHU and YU: A primitive fish close to the common ancestor of tetrapods and lungfish)
") 1 Supplementary Information (ZHU and YU: A primitive fish close to the common ancestor of tetrapods and lungfish) ------------------------------------------ I. List of 158 characters used for phylogenetic
1 Supplementary Information (ZHU and YU: A primitive fish close to the common ancestor of tetrapods and lungfish) ------------------------------------------ I. List of 158 characters used for phylogenetic
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL
 Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
Mammalogy Lab 1: Skull, Teeth, and Terms
 Mammalogy Lab 1: Skull, Teeth, and Terms Be able to: Goals of today s lab Locate all structures listed on handout Define all terms on handout what they are or what they look like Give examples of mammals
Mammalogy Lab 1: Skull, Teeth, and Terms Be able to: Goals of today s lab Locate all structures listed on handout Define all terms on handout what they are or what they look like Give examples of mammals
Friedman and Coates: Early morphological diversification of coelacanths
 Electronic Appendix A. Supplemental Morphological Data. Figure A1. Holopterygius nudus Jessen (P 7789a), latest Givetian-earliest Frasnian, Bergisch-Gladbach, Germany. Skull and pectoral girdle. (a) Photograph
Electronic Appendix A. Supplemental Morphological Data. Figure A1. Holopterygius nudus Jessen (P 7789a), latest Givetian-earliest Frasnian, Bergisch-Gladbach, Germany. Skull and pectoral girdle. (a) Photograph
OF THE TRIAS THE PHYTOSAURIA
 THE PHYTOSAURIA OF THE TRIAS MAURICE G. MEHL University of Wisconsin Some time ago the writer gave a brief notice of a new genus of phytosaurs of which Angistorhinus grandis Mehl was the type.' It is the
THE PHYTOSAURIA OF THE TRIAS MAURICE G. MEHL University of Wisconsin Some time ago the writer gave a brief notice of a new genus of phytosaurs of which Angistorhinus grandis Mehl was the type.' It is the
Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida. Evo-Devo Revisited. Development of the Tetrapod Limb
 Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida Evo-Devo Revisited Development of the Tetrapod Limb Limbs whether fins or arms/legs for only in particular regions or LIMB FIELDS. Primitively
Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida Evo-Devo Revisited Development of the Tetrapod Limb Limbs whether fins or arms/legs for only in particular regions or LIMB FIELDS. Primitively
Article. New semionotiform (Neopterygii) from the Tlayúa Quarry (Early Cretaceous, Albian), Mexico
 from the Tlayúa Quarry (Early Cretaceous, Albian), Mexico") Zootaxa 2749: 1 24 (2011) www.mapress.com/zootaxa/ Copyright 2011 Magnolia Press Article ISSN 1175-5326 (print edition) ZOOTAXA ISSN 1175-5334 (online edition) New semionotiform (Neopterygii) from the
Zootaxa 2749: 1 24 (2011) www.mapress.com/zootaxa/ Copyright 2011 Magnolia Press Article ISSN 1175-5326 (print edition) ZOOTAXA ISSN 1175-5334 (online edition) New semionotiform (Neopterygii) from the
 The following text is generated from uncorrected OCR. [Begin Page: Page 1] A NEW CERATOPSIAN DINOSAUR FROM THE UPPER CRETACEOUS OF MONTANA, WITH NOTE ON HYPACROSAURUS ' By CHARLES W. GILMORE assistant
The following text is generated from uncorrected OCR. [Begin Page: Page 1] A NEW CERATOPSIAN DINOSAUR FROM THE UPPER CRETACEOUS OF MONTANA, WITH NOTE ON HYPACROSAURUS ' By CHARLES W. GILMORE assistant
AMERICAN MUSEUM. Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET
 AMERICAN MUSEUM Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET NEW YORK, N.Y. 10024 U.S.A. NUMBER 2662 NOVEMBER 21, 1978 RONN W. COLDIRON Acroplous vorax
AMERICAN MUSEUM Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET NEW YORK, N.Y. 10024 U.S.A. NUMBER 2662 NOVEMBER 21, 1978 RONN W. COLDIRON Acroplous vorax
A NEW SALTICID SPIDER FROM VICTORIA By R. A. Dunn
 Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
Follow this and additional works at: Part of the Earth Sciences Commons
 Columbus State University CSU epress Faculty Bibliography 2012 The giant Cretaceous coelacanth (Actinistia, Sarcopterygii) Megalocoelacanthus dobiei Schwimmer, Stewart & Williams, 1994, and its bearing
Columbus State University CSU epress Faculty Bibliography 2012 The giant Cretaceous coelacanth (Actinistia, Sarcopterygii) Megalocoelacanthus dobiei Schwimmer, Stewart & Williams, 1994, and its bearing
AMERICAN NATURALIST. Vol. IX. -DECEMBER, No. 12. OR BIRDS WITH TEETH.1 OI)ONTORNITHES,
ONTORNITHES,") AMERICAN NATURALIST. Vol. IX. -DECEMBER, 1875.-No. 12. OI)ONTORNITHES, OR BIRDS WITH TEETH.1 BY PROFESSOR 0. C. MARSH. REMAINS of birds are amono the rarest of fossils, and few have been discovered except
AMERICAN NATURALIST. Vol. IX. -DECEMBER, 1875.-No. 12. OI)ONTORNITHES, OR BIRDS WITH TEETH.1 BY PROFESSOR 0. C. MARSH. REMAINS of birds are amono the rarest of fossils, and few have been discovered except
A preliminary note on Bobasatrania groenlandica.
 A preliminary note on Bobasatrania groenlandica. BY EIGIL NIELSEN. The third part of my monograph on the Triassic fishes from East Greenland is planned to deal Mvith Bobasatrania, a genus of deep-bodied
A preliminary note on Bobasatrania groenlandica. BY EIGIL NIELSEN. The third part of my monograph on the Triassic fishes from East Greenland is planned to deal Mvith Bobasatrania, a genus of deep-bodied
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE
 FROM THE") A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. GLYPTOLEPIS FROM THE MIDDLE DEVONIAN OF SCOTLAND
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 99 April 16, 1966 GLYPTOLEPIS FROM THE MIDDLE DEVONIAN OF SCOTLAND KEITH STEWART THOMSON 1 DEPARTMENT OF
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 99 April 16, 1966 GLYPTOLEPIS FROM THE MIDDLE DEVONIAN OF SCOTLAND KEITH STEWART THOMSON 1 DEPARTMENT OF
NEW INFORMATION ON THE CRANIUM OF BRACHYLOPHOSAURUS CANADENSIS (DINOSAURIA, HADROSAURIDAE), WITH A REVISION OF ITS PHYLOGENETIC POSITION
, WITH A REVISION OF ITS PHYLOGENETIC POSITION") Journal of Vertebrate Paleontology 25(1):144 156, March 2005 2005 by the Society of Vertebrate Paleontology NEW INFORMATION ON THE CRANIUM OF BRACHYLOPHOSAURUS CANADENSIS (DINOSAURIA, HADROSAURIDAE), WITH
Journal of Vertebrate Paleontology 25(1):144 156, March 2005 2005 by the Society of Vertebrate Paleontology NEW INFORMATION ON THE CRANIUM OF BRACHYLOPHOSAURUS CANADENSIS (DINOSAURIA, HADROSAURIDAE), WITH
The first record of Late Jurassic crossognathiform fishes from Europe and their phylogenetic importance for teleostean phylogeny
 Fossil Record 13 (2) 2010, 317 341 / DOI 10.1002/mmng.201000005 The first record of Late Jurassic crossognathiform fishes from Europe and their phylogenetic importance for teleostean phylogeny Gloria Arratia*,
Fossil Record 13 (2) 2010, 317 341 / DOI 10.1002/mmng.201000005 The first record of Late Jurassic crossognathiform fishes from Europe and their phylogenetic importance for teleostean phylogeny Gloria Arratia*,
CENE RUMINANTS OF THE GENERA OVIBOS AND
 DESCRIPTIONS OF TWO NEW SPECIES OF PLEISTO- CENE RUMINANTS OF THE GENERA OVIBOS AND BOOTHERIUM, WITH NOTES ON THE LATTER GENUS. By James Williams Gidley, Of the United States National Museum. Two interesting
DESCRIPTIONS OF TWO NEW SPECIES OF PLEISTO- CENE RUMINANTS OF THE GENERA OVIBOS AND BOOTHERIUM, WITH NOTES ON THE LATTER GENUS. By James Williams Gidley, Of the United States National Museum. Two interesting
UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA
 FROM JAVA, INDONESIA") NOTES AND NEWS UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA BY NGUYEN NGOC-HO i) Faculty of Science, University of Saigon, Vietnam Among material recently collected
NOTES AND NEWS UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA BY NGUYEN NGOC-HO i) Faculty of Science, University of Saigon, Vietnam Among material recently collected
LOWER CRETACEOUS OF SOUTH DAKOTA.
 A NEW DINOSAUR, STP^GOSAURUS MARSHl, FROM THE LOWER CRETACEOUS OF SOUTH DAKOTA. By Frederic A. Lucas, Curator, Divisioii of Coiiipnrative Anatomy, in charge, of Section of Vertebrate Fossils. The name
A NEW DINOSAUR, STP^GOSAURUS MARSHl, FROM THE LOWER CRETACEOUS OF SOUTH DAKOTA. By Frederic A. Lucas, Curator, Divisioii of Coiiipnrative Anatomy, in charge, of Section of Vertebrate Fossils. The name
On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds
 and the origin of birds") On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds by Qiang Ji and Shu an Ji Chinese Geological Museum, Beijing Chinese Geology Volume 233 1996 pp.
On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds by Qiang Ji and Shu an Ji Chinese Geological Museum, Beijing Chinese Geology Volume 233 1996 pp.
Beaufortia. (Rathke) ZOOLOGICAL MUSEUM - AMSTERDAM. July. Three new commensal Ostracods from Limnoria lignorum
 ZOOLOGICAL MUSEUM - AMSTERDAM. July. Three new commensal Ostracods from Limnoria lignorum") Beaufortia SERIES OF MISCELLANEOUS PUBLICATIONS ZOOLOGICAL MUSEUM - AMSTERDAM No. 34 Volume 4 July 30, 1953 Three new commensal Ostracods from Limnoria lignorum (Rathke) by A.P.C. de Vos (Zoological Museum,
Beaufortia SERIES OF MISCELLANEOUS PUBLICATIONS ZOOLOGICAL MUSEUM - AMSTERDAM No. 34 Volume 4 July 30, 1953 Three new commensal Ostracods from Limnoria lignorum (Rathke) by A.P.C. de Vos (Zoological Museum,
Phylogenetic Revision of the Fish Families Luvaridae and fkushlukiidae j&^j $it (Acanthuroidei), with a New Genus and rf^;'
, with a New Genus and rf^;'") *J J& ^m^ "*" Phylogenetic Revision of the Fish Families Luvaridae and fkushlukiidae j&^j $it (Acanthuroidei), with a New Genus and rf^;',j Two New Species of Eocene Luvarids PMl 4B3lRli» - I ^ V' SERIES
*J J& ^m^ "*" Phylogenetic Revision of the Fish Families Luvaridae and fkushlukiidae j&^j $it (Acanthuroidei), with a New Genus and rf^;',j Two New Species of Eocene Luvarids PMl 4B3lRli» - I ^ V' SERIES
Mammalogy Lecture 8 - Evolution of Ear Ossicles
 Mammalogy Lecture 8 - Evolution of Ear Ossicles I. To begin, let s examine briefly the end point, that is, modern mammalian ears. Inner Ear The cochlea contains sensory cells for hearing and balance. -
Mammalogy Lecture 8 - Evolution of Ear Ossicles I. To begin, let s examine briefly the end point, that is, modern mammalian ears. Inner Ear The cochlea contains sensory cells for hearing and balance. -
A New Pterosaur from the Middle Jurassic of Dashanpu, Zigong, Sichuan
 A New Pterosaur from the Middle Jurassic of Dashanpu, Zigong, Sichuan by Xinlu He (Chengdu College of Geology) Daihuan Yang (Chungking Natural History Museum, Sichuan Province) Chunkang Su (Zigong Historical
A New Pterosaur from the Middle Jurassic of Dashanpu, Zigong, Sichuan by Xinlu He (Chengdu College of Geology) Daihuan Yang (Chungking Natural History Museum, Sichuan Province) Chunkang Su (Zigong Historical
NORTH AMERICA. ON A NEW GENUS AND SPECIES OF COLUBRINE SNAKES FROM. The necessity of recognizing tlie two species treated of in this paper
 ON A NEW GENUS AND SPECIES OF COLUBRINE SNAKES FROM NORTH AMERICA. BY Leonhard Stejneger, and Batrachians. Curator of the Department of Reptiles The necessity of recognizing tlie two species treated of
ON A NEW GENUS AND SPECIES OF COLUBRINE SNAKES FROM NORTH AMERICA. BY Leonhard Stejneger, and Batrachians. Curator of the Department of Reptiles The necessity of recognizing tlie two species treated of
290 SHUFELDT, Remains of Hesperornis.
 290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
A Late Jurassic Protosuchian Sichuanosuchus huidongensis from Zigong, Sichuan Province. Guangzhao Peng. Zigong Dinosaur Museum, Zigong, Sichuan
 A Late Jurassic Protosuchian Sichuanosuchus huidongensis from Zigong, Sichuan Province Guangzhao Peng Zigong Dinosaur Museum, Zigong, Sichuan 643013 Vertebrata PalAsiatica Volume 34, Number 4 October,
A Late Jurassic Protosuchian Sichuanosuchus huidongensis from Zigong, Sichuan Province Guangzhao Peng Zigong Dinosaur Museum, Zigong, Sichuan 643013 Vertebrata PalAsiatica Volume 34, Number 4 October,
First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia
 from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia") First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia Tsogtbaatar Chinzorig¹, ³ *, Yoshitsugu Kobayashi², Khishigjav Tsogtbaatar³,
First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia Tsogtbaatar Chinzorig¹, ³ *, Yoshitsugu Kobayashi², Khishigjav Tsogtbaatar³,
Jesús Alvarado-Ortega 1,* and María del Pilar Melgarejo-Damián 2. REVISTA MEXICANA DE CIENCIAS GEOLÓGICAS v. 34, núm. 3, 2017, p.
 Alvarado-Ortega and Melgarejo-Damián REVISTA MEXICANA DE CIENCIAS GEOLÓGICAS v. 34, núm. 3, 2017, p. 234-249 Paraclupea seilacheri sp. nov., a double armored herring (Clupeomorpha, Ellimmichthyiformes)
Alvarado-Ortega and Melgarejo-Damián REVISTA MEXICANA DE CIENCIAS GEOLÓGICAS v. 34, núm. 3, 2017, p. 234-249 Paraclupea seilacheri sp. nov., a double armored herring (Clupeomorpha, Ellimmichthyiformes)
PALEONTOLOGICAL CONTRIBUTIONS
 THE UNIVERSITY OF KANSAS PALEONTOLOGICAL CONTRIBUTIONS August, 1965 Paper 2 A NEW WYOMING PHYTOSAUR By THEODORE H. EATON, JR. [Museum of Natural History, University of Kansas I ABSTRACT The skull of a
THE UNIVERSITY OF KANSAS PALEONTOLOGICAL CONTRIBUTIONS August, 1965 Paper 2 A NEW WYOMING PHYTOSAUR By THEODORE H. EATON, JR. [Museum of Natural History, University of Kansas I ABSTRACT The skull of a
Osteology of the telescopefishes of the genus Gigantura (Brauer, 1901), Teleostei: Aulopiformes
, Teleostei: Aulopiformes") Zoological Journal of the Linnean Society, 2017, 179, 338 353. With 8 figures Osteology of the telescopefishes of the genus Gigantura (Brauer, 1901), Teleostei: Aulopiformes PETER KONSTANTINIDIS 1 * and
Zoological Journal of the Linnean Society, 2017, 179, 338 353. With 8 figures Osteology of the telescopefishes of the genus Gigantura (Brauer, 1901), Teleostei: Aulopiformes PETER KONSTANTINIDIS 1 * and
NOTE XXXVIII. Three new species of the genus Helota DESCRIBED BY. C. Ritsema+Cz. is very. friend René Oberthür who received. Biet.
 Subshining; HELOTA MARIAE. 249 NOTE XXXVIII. Three new species of the genus Helota DESCRIBED BY C. Ritsema+Cz. The first of these species is very interesting as it belongs to the same section as the recently
Subshining; HELOTA MARIAE. 249 NOTE XXXVIII. Three new species of the genus Helota DESCRIBED BY C. Ritsema+Cz. The first of these species is very interesting as it belongs to the same section as the recently