A role for apical membrane antigen 1 during invasion of hepatocytes
|
|
|
- Cameron Walsh
- 5 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on December 15, 2003 as Manuscript M A role for apical membrane antigen 1 during invasion of hepatocytes by Plasmodium falciparum sporozoites. Olivier Silvie a, Jean-François Franetich a, Stéphanie Charrin b, Markus S. Mueller c, Anthony Siau a, Myriam Bodescot a, Eric Rubinstein b, Laurent Hannoun d, Yupin Charoenvit e, Clemens H. Kocken f, Alan W. Thomas f, Geert-Jan van Gemert g, Robert W. Sauerwein g, Michael J. Blackman h, Robin F. Anders i, Gerd Pluschke c, and Dominique Mazier a. a Inserm U511, Immunobiologie Cellulaire et Moléculaire des Infections Parasitaires, Centre Hospitalo-Universitaire Pitié-Salpêtrière, Université Pierre et Marie Curie, Paris, France; b Inserm U268, Institut André-Lwoff, Université Paris XI, Hôpital Paul-Brousse, Villejuif, France; c Molecular Immunology, Swiss Tropica l Institute, CH-4002 Basel, Switzerland; d Service de Chirurgie Digestive, Hépato -Bilio -Pancréatique et Transplantation Hépatique, Hôpital Pitié-Salpêtrière, Paris, France; e Malaria Program, Naval Medical Research Center, Silver Spring, MA20910, USA; f Biomedical Primate Research Centre, Department of Parasitology, 2280 GH Rijswijk, The Netherlands; g Department of Medical Microbiology, University Medical Centre St. Radboud, 6500 HB Nijmegen, The Netherlands; h Division of Parasitology, National Institute for Medical Research, Mill Hill, London NW7 1AA, UK; i Department of Biochemistry and Cooperative Research Centre for Vaccine Technology, La Trobe University, Victoria 3086, Australia. Running title : Proteolytic processing in Plasmodium sporozoites. Key words: Plasmodium, hepatocyte, AMA-1, TRAP, proteolytic processing. All correspondence to: Dr Olivier Silvie, Inserm U511, Faculté de Médecine Pitié- Salpêtrière, 91 Bvd de l Hôpital, Paris, France. Phone: Fax: E -mail: silvie@ext.jussieu.fr 1 Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
2 SUMMARY Plasmodium sporozoites are transmitted through the bite of infected mosquitoes and invade hepatocytes, as a first and obligatory step of the parasite life cycle in man. Hepatocyte invasion involves proteins secreted from parasite vesicles called micronemes, the most characterized being the thrombospondin -related adhesive protein (TRAP). Here we investigated the expression and function of another microneme protein recently identified in Plasmodium falciparum sporozoites, apical membrane antigen 1 (AMA-1). P. falciparum AMA-1 is expressed in sporozoites and is lost after invasion of hepatocytes, and anti-ama-1 antibodies inhibit sporozoite invasion, suggesting that the protein is involved during invasion of hepatocytes. As observed with TRAP, AMA -1 is initially mostly sequestered within the sporozoite. Upon microneme exocytosis, AMA -1 and TRAP relocate to the sporozoite surface, where they are proteolytically cleaved, resulting in the shedding of soluble fragments. A subset of serine protease inhibitors blocks the processing and shedding of both AMA-1 and TRAP, and inhibits sporozoite infectivity, suggesting that interfering with sporozoite proteolytic processing may constitute a valuable strategy to prevent hepatocyte infection. 2
3 INTRODUCTION The apicomplexan parasite Plasmodium falciparum is a causative agent of malaria, one of the major human infectious diseases, responsible for more than 1 million deaths per year worldwide. Plasmodium sporozoites are transmitted through the bite of infected mosquitoes and invade the liver of the mammalian host, where they undergo schizogony and differentiate into merozoites that subsequently invade erythrocytes and cause disease. Blocking sporozoite invasion of hepatocytes represents an attractive anti-malarial strategy since it would prevent malaria symptoms and parasite transmission to the mosquito, which both occur at erythrocytic stages. Sporozoites migrate through tissues and invade target cells using gliding motility (1), a process involving secretor y vesicles called micronemes. Upon microneme exocytosis, proteins are delivered onto the parasite surface, and then redistributed from the anterior to the posterior end of the parasite, leading to its forward movement and penetration into a host cell (2). Only two sporozoite microneme proteins, namely circumsporozoite protein (CSP) 1 and thrombospondin - related adhesive protein (TRAP), have been extensively studied (3). It is quite likely however that other microneme proteins are involved during the invasion process. Recently, Florens et al. reported a comprehensive view of the proteome of P. falciparum, obtained with high-resolution liquid chromatography and tandem mass spectrometry (4). Interestingly, some of the proteins detected in sporozoites had not been reported in this stage before, including apical membrane antigen 1 (AMA-1), a microneme protein involved in merozoite invasion of erythrocytes and considered a leading candidate for inclusion in a vaccine against erythrocytic stages of P. falciparum (5-7). Here, we have analyzed AMA-1 expression in P. falciparum sporozoites and investigated its potential role during invasion of hepatocytes. 3
4 EXPERIMENTAL PROCEDURES Antibodies and inhibitors. We used previously characterized anti- P. falciparum AMA-1 (PfAMA-1) domain III monoclonal antibody (mab) DV5 (8), anti-pfama-1 prodomain mab 5G8 (9), anti-pfama-1 domain I mab 1F9 (9), anti- 3D7 strain PfAMA-1 polyclonal rabbit IgG (10), anti-fvo strain PfAMA-1 polyclonal rabbit IgG (11), anti-pftrap mab SSP2.2 (12), and anti-pfcsp mab E9 (13). For liver schizont counting, we used anti-plasmodium HSP-70 sera obtained from mice immunized with i72 recombinant protein (gift from D. Mattei, Institut Pasteur, Paris, France). Secondary antibodies were fluorescein isothiocyanate conjugated goat anti-mouse IgG (GAM-FITC, Sigma), Alexa Fluor 594 goat antirabbit IgG (GAR- Alexa Fluor 594, Molecular Probes), peroxidase goat anti-mouse and peroxidase goat anti-rabbit IgG (Becton Dickinson). Cytochalasin D (500µM stock in DMSO) and protease inhibitors chymostatin (2 mg/ml stock in DMSO), pepstatin A (1 mg/ml stock in DMSO), leupeptin (1 mg/ml stock in water), phenylmethylsulfonyl fluoride (PMSF, 100 mm stock in ethanol) and N-tosyl-Llysine chloromethyl ketone (TLCK, 20 mm stock in water) were from Sigma. The peptidyl chloromethylketone N -benzyloxycarbonyl-gly-met-leu-ch 2 Cl (Z-GML- CH 2 Cl, 10 mm stock in DMSO) was synthesized as described (14). Isolation of P. falciparum sporozoites. Anopheles stephensi adult females were infected with the NF54 strain of P. falciparum, using a membrane -based feeder system (15). After 14 to 21 days, mosquitoes were killed and their salivary glands were aseptically dissected and disrupted by trituration in a glass tissue grinder, and the sporozoites were counted in a KovaSlide chamber. Isolation and culture of human hepatocytes. Primary human hepatocyte cultures were prepared as described (16), with minor modifications. We used either 4
5 fresh or cryopreserved hepatocytes, both susceptible to infection with P. falciparum sporozoites (17,18). In our hands, P. falciparum sporozoite infectivity is similar in fresh and cryopreserved hepatocytes 2. Briefly, cells were isolated by collagenase (PAA Laboratories) perfusion of human liver fragments, used in agreement with the French ethical regulations, and were further purified over a 40% Percoll gradient. Hepatocytes were cryopreserved in liquid nitrogen in fetal calf serum (FCS) with 10% DMSO; after thawing, viable hepatocytes were purified over a 40% Percoll gradient. Hepatocytes were seeded in eight-chamber plastic Lab-Tek slides or in 24-well culture plates (Nalge Nunc International), coated with rat tail collagen I (Becton Dickinson), at a density of 18 x 10 4 cells per cm 2 (fresh hepatocytes) or 25 x 10 4 cells per cm 2 (cryopreserved hepatocytes). Hepatocytes were cultured at 37 C in 4% CO2, in William s E medium (Gibco) supplemented to a final concentration of 10% FCS (Gibco), 2% penicillin-streptomycin (100X, stock solution, Gibco), 1% sodium pyruvate (100X, stock solution, Gibco), 1% L-glutamine (100X, stock solution, Gibco), 1% insulin-transferrin -selenium (ITS, 100X, stock solution, Gibco). After complete cell adherence (12-24 hours), the culture medium was supplemented with 10-7 M dexamethasone (Sigma) and 2% DMSO to maintain hepatocyte differentiation (19). Culture of P. falciparum liver stages. Human hepatocytes were cultured for at least 48 hours before inoculation with P. falciparum sporozoites (18). After removal of medium from the culture chambers, sporozoites in culture medium were added to the Lab-Tek wells (1 x 10 5 sporozoites per well) or plate wells (1 x 10 6 sporozoites per well), and incubated with the hepatocytes for 3 hours. Cultures were then washed and further incubated in complete culture medium supplemented with 0.25% DMSO. Culture medium was renewed every 2-3 days. 5
6 Reverse Transcriptase (RT)-PCR. Total RNA was purified from P. falciparum NF54 sporozoites using the QIAamp RNA Blood Mini Kit (Qiagen), including a DNase treatment step. Reverse transcription was performed using the ProSTAR First- Strand RT-PCR Kit (Stratagene). One µl of cdna was PCR amplified with the sense primer 5 -TTCAAATACTACTTTTTTAACACCG-3 and the antisense primer 5 - CTCTTTCGATTTCTTTCATTATTTC-3, by use of 40 cycles of 45 sec of denaturation at 94 C, 45 sec of annealing at 55 C, and 60 sec of elongation at 70 C. The resulting products (890 base pair amplicon) were stained with ethidium bromide and run on a 1% agarose gel in TAE buffer. Immunofluorescence assays. For detection of AMA-1 expression in P. falciparum sporozoites and liver stages, air dried sporozoites and methanol fixed infected hepatocyte cultures were blocked with 3% bovine serum albumin (BSA) in PBS for 30 min, then incubated with anti-pfama-1 mab DV5 for 1h at 37 C, followed by incubation with GAM-FITC and 1µg/ml DNA stain diamidinophenylindole (DAPI) for 30 minutes before examination by fluorescence confocal microscopy. Surface staining was performed on non-permeabilized sporozoites deposited on poly-l-lysine-coated multiwell glass slides, in a wet sporozoite assay (20); sporozoites were fixed in 2% paraformaldehyde (PFA) for 20 min at room temperature and endogenous fluorescence was quenched with 0.1 M glycine. Parasites were blocked with 3% BSA in PBS for 1h, and then incubated with primary antibodies (anti-pfama-1 mabs DV5, 1F9 or 5G8, or rabbit polyclonal anti-pfama - 1 IgG, as indicated in the figure legends) for 2h at 37 C, followed by incubation with secondary antibodies (GAM-FITC for mabs or GAR- Alexa Fluor 594 for rabbit IgG) and DAPI for 1h, before examination by fluorescence microscopy. 6
7 Western blotting and secretion assays. For detection of parasite proteins in sporozoite lysates and supernatants, parasites were dissolved in SDS-PAGE sample buffer and incubated at 70 C for 5 min. Protein samples were subjected to 10% SDS- PAGE (1x10 5 parasites or equivalent per lane for AMA-1 and TRAP detection, 1 x 10 3 parasites or equivalent per lane for CSP detection) and transferred onto polyvinylidene membranes. These were probed with primary antibodies, followed by a peroxidase -conjugated secondary antibody, and antigens were revealed by enhanced chemiluminescence. To analyze proteins secreted in the parasite supernatant, sporozoites were resuspended in medium without serum, and incubated for 2h at 37 C, in the presence or absence of hepatocytes. The parasite suspension was then dissolved by adding SDS-PAGE sample buffer. In some experiments, supernatants were separated from the sporozoites by centrifugation at 15000g for 5 min, and parasite pellets and supernatants were dissolved in SDS-PAGE sample buffer separately. Gliding assays. To analyze sporozoite motility, sporozoites were deposited on multispot glass slide wells precoated with anti-pfcsp mab E9 (100 µg/ml 1 h at 37 C), and incubated at 37 C for 1 h. Slides were then washed and the deposited CSP trails fixed with 4 % PFA for 15 min. The trails were then labeled using the anti-pfcsp mab E9 conjugated to Alexa Fluor 488 and visualized under a fluorescence microscope. Inhibition of sporozoite invasion assays. To analyze sporozoite invasion, triplicate hepatocyte cultures were inoculated with P. falciparum sporozoites (1 x 10 5 per Lab-Tek well). After 3 hours at 37 C, cultures were washed and further incubated in fresh medium for 3 days, then fixed in methanol. Liver schizonts were stained using an anti-hsp -70 mouse serum followed by goat anti-mouse FITC conjugate, and 7
8 counted under a fluorescence microscope. To determine the effects of anti-ama-1 antibodies on sporozoite infectivity, sporozoites were incubated with hepatocytes in the presence of increasing concentrations of anti-pfama-1 rabbit IgG. Percentage inhibition was determined in comparison to control rabbit IgG. To determine the effects of protease inhibitors and cytochalasin D on sporozoite infectivity, P. falciparum sporozoites were incubated with these inhibitors (each from a 100X stock solution diluted directly in the parasite suspension) for 5 min at room temperature, and then washed (except for cytochalasin D, the effects of which are reversible), before inoculation onto hepatocytes. Percentage inhibition was determined in comparison to control sporozoites treated with solvents alone. Inhibition results were analyzed for statistical significance using the one-way ANOVA followed by the Tukey multiple comparison test. 8
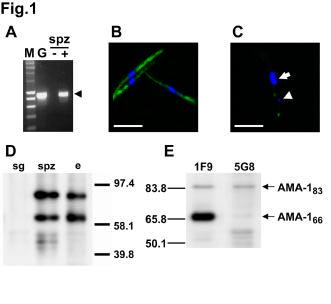
9 RESULTS AMA-1 is expressed in P. falciparum sporozoites. In a recent analysis of P. falciparum proteome, peptides corresponding to the merozoite antigen AMA-1 were identified in sporozoites. Since AMA-1 had never been reported in sporozoites before, we were interested in further characterizing its expression at this stage. Using RT-PCR, we could readily detect ama -1 transcripts in salivary gland s porozoites (Fig. 1A), thus confirming the expression of the gene at this stage. To test whether AMA -1 protein is also present in sporozoites, we performed immunofluorescence assays (IFA) using the anti-pfama-1 domain III mab DV5 (8). All permeabilized sporozoites displayed a strong fluorescence (Fig. 1B ), with a bipolar nucleus-sparing pattern similar to the immunofluorescence pattern observed with antibodies to TRAP, also known as sporozoite surface protein 2 (SSP -2) (12). In contrast, surface labeling with the anti-ama-1 mab showed no or very little surface fluorescence on sporozoites (Fig. 1C). When detected, AMA-1 surface fluorescence was restricted to the apical extremity of the sporozoites, on the same side of the nucleus as the apicoplast, a DNA-containing organelle visualized as a DAPI-positive spot forward of the nucleus, as described in Toxoplasma (21,22). A similar surface fluorescence pattern was observed using polyclonal anti-pfama-1 antibodies (see Fig. 5A). Western blotting of protein extracts from P. falciparum sporozoites probed with DV5 confirmed AMA- 1 expression in sporozoites. Two proteins of ~83 kda and ~66 kda were detected in both sporozoites and parasitized erythrocytes, and were absent from non-infected mosquito salivary glands (Fig. 1D). In P. falciparum erythrocytic stages, these two bands have been shown to correspond respectively to a precursor protein, that contains a prosequence, and to a mature form of AMA-1, that lacks the prosequence (23,24). To confirm that the two bands observed in the sporozoite lysates also 9
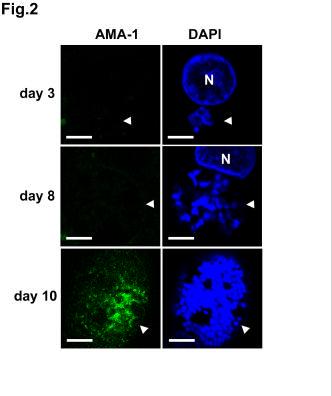
10 correspond to the precursor and mature forms, we performed western blot analysis using the anti-ama-1 mabs 1F9, specific for P. falciparum AMA-1 domain I, and 5G8, specific for AMA -1 prosequence. The mab 5G8 recognized only the upper ~83 kda band, while 1F9 recognized both ~83 kda and ~66 kda proteins (Fig. 1E), thus confirming that in P. falciparum sporozoites these two bands correspond to the precursor and the mature form of AMA-1, respectively. The relative amount of precursor and mature proteins detected by western blot varied between the different sporozoite preparations (Fig. 1 and see Fig. 4 ). AMA-1 is lost after invasion of hepatocytes, and is re-expressed in liver merozoites. To assess the fate of AMA-1 after sporozoite invasion of hepatocytes, we analysed AMA-1 expression in primary human hepatocytes infected with sporozoites (18). We consistently failed to detect the protein in early liver-stages by IFA (Fig. 2), suggesting that it was lost after invasion of hepatocytes. AMA-1 was only reexpressed in mature P. falciparum liver schizonts, with a punctate pattern likely corresponding to nascent liver merozoites (Fig. 2). Anti-AMA-1 antibodies inhibit the invasion of human hepatocytes by P. falciparum sporozoites. The expression of AMA-1 in P. falciparum sporozoites, and the fact that AMA-1 was not detected in liver schizonts until the differentiation of liver merozoites, suggested that AMA -1 could be involved in the process of invasion of hepatocytes by sporozoites. Therefore, we tested whether antibodies to AMA-1 could inhibit the invasion of hepatocytes by P. falciparum sporozoites. Polyclonal IgG raised in rabbits immunized with a recombinant AMA-1 protein from the P. falciparum 3D7 clone (10) inhibited invasion of human hepatocytes by NF54 10
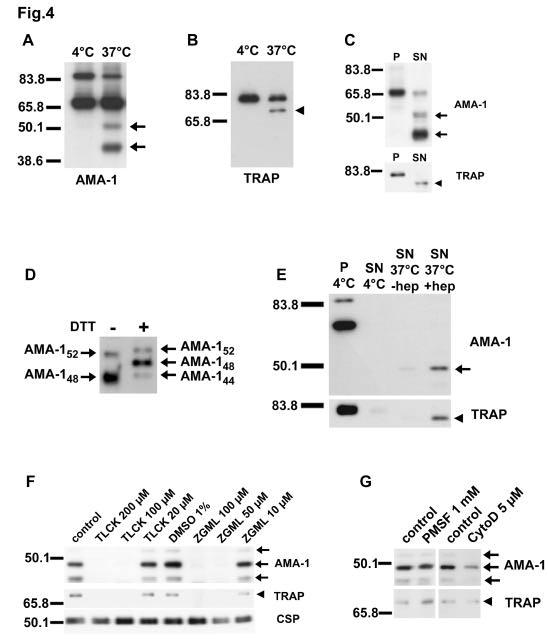
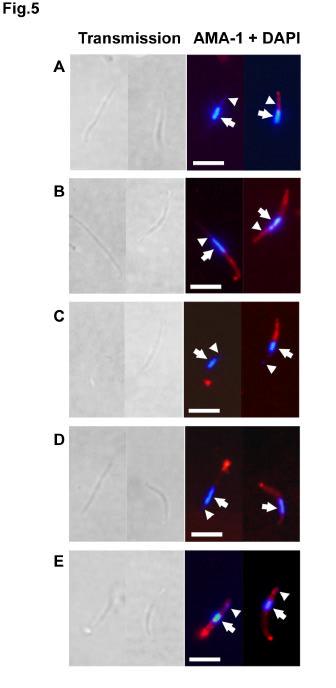
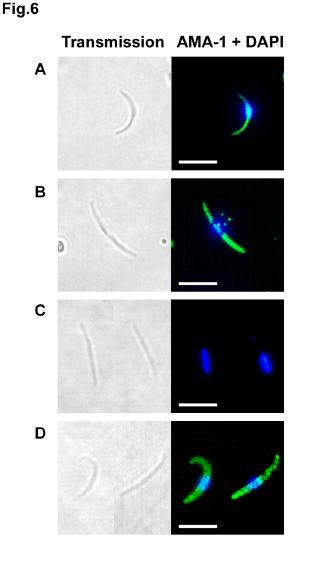
11 (homologous strain) P. falciparum sporozoites, in a concentration-dependent manner (Fig. 3). Relatively high concentrations were necessary to achieve significant inhibition, but equivalently high concentrations of IgG from rabbits immunized with a reduced and alkylated form of the recombinant 3D7 PfAMA -1 protein did not inhibit sporozoite invasion (Fig. 3), indicating that inhibition is mediated by antibodies directed to conformational epitopes. IgG from rabbits immunized with a recombinant AMA-1 from the P. falciparum heterologous strain FVO (11) also inhibited NF54 P. falciparum sporozoite invasion (Fig. 3). Upon microneme exocytosis, AMA-1 and TRAP are translocated to the sporozoite surface and shed as soluble forms after proteolytic processing. Only a minority of salivary gland sporozoites (~5-10%) displayed detectable amounts of AMA-1 on their apical surface (Fig. 1C and Fig. 5A), suggesting that most of the protein is sequestered within the sporozoite micronemes, as observed with TRAP (25). Upon incubation at 37 C, which induces microneme exocytosis (26,27), most of the sporozoites (~60-80%) expressed AMA -1 on their surface, distributed either over the whole surface or as a posterior cap (Fig. 5B). Sporozoite surface AMA-1 reacted with 1F9 but not with 5G8 mab, indicating that only the mature 66 kda protein translocates to the parasite surface (Fig. 6, C-D). As expected, both 5G8 and 1F9 mabs labeled air-dried permeabilized sporozoites (Fig. 6, A-B), with a fluorescent pattern similar to that observed with DV5. In P. falciparum merozoites, AMA-1 translocates to the surface upon microneme exocytosis, and is proteolytically cleaved and shed as soluble fragments (14,24). To determine if similar processing events also occur in P. falciparum sporozoites, we analysed by western blot lysates from sporozoites incubated at 37 C. Indeed, an AMA-1 cleavage product of ~48 kda, 11
12 associated with a less abundant product of ~52 kda, was found in lysates from sporozoites incubated at 37 C, but not in lysates from sporozoites kept at 4 C upon isolation from mosquito salivary glands (Fig. 4A). A TRAP cleavage product was also detected in the preparations from sporozoites incubated at 37 C, 5-10 kda smaller than the sporozoite TRAP, consistent with the size of the TRAP ectodomain (Fig. 4B). Importantly, when sporozoites and supernatants were analysed separately, the cleavage products of both AMA -1 and TRAP were found only in the supernatants, not in the sporozoite pellets (Fig. 4C), demonstrating that these products correspond to soluble proteins shed in the supernatant. When sporozoite supernatants were run under reducing conditions, the AMA-1 48 kda protein migrated as two distinct bands, a major ~48 kda and a minor ~44 kda product (Fig. 4D), as observed with merozoite AMA-1 (14). The shed TRAP product migrated as a single band under both reducing and non-reducing conditions (data not shown). AMA -1 and TRAP processing and shedding was observed in sporozoites incubated at 37 C in the absence of host cells, showing that the protease(s) involved in this processing is (are) encoded by the parasite. Nevertheless, incubation of sporozoites at 37 C in the presence of hepatocytes consistently enhanced the shedding of AMA-1 and TRAP in the sporozoite supernatants (Fig. 4E). AMA-1 and TRAP are processed by a sporozoite serine protease. To further characterize the protease activities mediating AMA-1 and TRAP processing, we used a set of protease inhibitors. Chymostatin, leupeptin and pepstatin A had no effect on AMA-1 and TRAP shedding (data not shown). In contrast, two serine protease inhibitors, TLCK and Z-GML-CH 2 Cl, had a profound dose-dependent inhibitory effect on both AMA-1 and TRAP processing and shedding, but had no effect on CSP 12
13 release into the supernatant (Fig. 4F). CSP was detected as a doublet in both sporozoite pellets and supernatants, without any additional cleavage product detectable in parasites incubated at 37 C (data not shown). Strikingly, PMSF, another serine protease inhibitor, had no significant activity on AMA -1 or TRAP processing in sporozoites (Fig. 4G). To further characterize the timing of processing events, we analyzed the effects of protease inhibitors on the surface exposure of AMA-1 and TRAP. Treatment with 200 µm TLCK or 100 µm Z-GML-CH 2 Cl, though profoundly inhibiting the shedding of AMA-1 and TRAP, did not prevent the translocation of AMA-1 to the sporozoite surface at 37 C. On the contrary, the surface exposure of AMA -1, which tended to accumulate at the posterior end of the parasites, was rather enhanced following treatment with these inhibitors (Fig. 5, C-D). These observations, together with the fact that AMA-1 and TRAP cleavage products were not found associated with the sporozoite pellets, indicate that the proteolytic processing and shedding of AMA-1 and TRAP follows the translocation of these proteins to the parasite surface upon microneme exocytosis, suggesting that the protease involved operates on the sporozoite surface. Surface translocation and proteolytic processing of AMA-1 and TRAP are not sensitive to cytochalasin D. Gliding motility and host cell invasion rely on a parasite actomyosin motor that operates beneath the parasite plasma membrane and drives the anterior-to-posterior translocation of microneme proteins once they are released onto the parasite surface (28). As this actomyosin motor is blocked by the actin polymerization inhibitor cytochalasin D (29), we investigated the effects of cytochalasin D on AMA-1 and TRAP processing. Treatment of sporozoites with 5 µm cytochalasin D completely inhibited both sporozoite gliding and invasion of 13
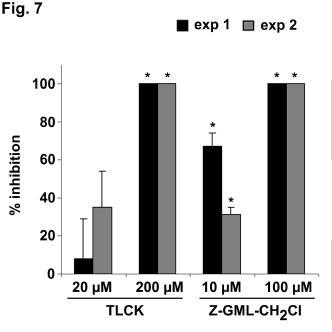
14 hepatocytes (data not shown), but did not prevent the translocation of AMA -1 and TRAP to the sporozoite surface nor their subs equent processing upon passage at 37 C (Fig. 4G and Fig. 5 E). However, a slight reduction of AMA -1 and TRAP shedding was observed when sporozoites were co-incubated with hepatocytes in the presence of cytochalasin D (Fig. 4G), probably because cytochalasin D, by suppressing sporozoite motility, prevents the up-regulation of microneme exocytosis induced by sporozoite migration through cells (30). Interfering with surface protein processing inhibits P. falciparum sporozoite infectivity. Finally, to determine if AMA -1 and TRAP processing is required during sporozoite invasion of hepatocytes, we analysed the effects of protease inhibitors on sporozoite infectivit y in vitro. Pretreatment of sporozoites with 200 µm TLCK and 100 µm Z-GML-CH 2 Cl, both inhibiting AMA-1 and TRAP shedding, abrogated sporozoite gliding on glass slides (data not shown). More importantly, treatment with TLCK and Z -GML-CH 2 Cl inhibited sporozoite invasion of hepatocytes, in a dosedependent manner (Fig. 6). In contrast, treatment of the parasites with chymostatin, leupeptin, pepstatin A and PMSF, which do not prevent AMA-1 and TRAP processing, had no significant effect on sporozoite motility and infectivity, and pretreatment of hepatocytes with the different protease inhibitors had no effect on sporozoite invasion (data not shown). 14
15 DISCUSSION Until recently, most of the studies on hepatocyte invasion by sporozoites focused on two sporozoite proteins, CSP and TRAP (3). Many other proteins are expressed at the sporozoite stage (4), some of them being potentially involved in the invasion process. Here we show that P. falciparum AMA-1, a microneme protein involved in merozoite invasion of erythrocytes (7), is also expressed in sporozoites, thus confirming mass spectrometry data (4). AMA-1 is lost after sporozoite invasion, and is only re-expressed in liver merozoites, suggesting that it could play a role during invasion of hepatocytes. Indeed, antibodies specific for the P. falciparum clone 3D7 AMA-1 inhibit invasion of human hepatocytes by sporozoites of the homologous strain NF54. This inhibitory activity is directed primarily to conformational epitopes, as indicated by the absence of neutralizing activity of antibodies raised against a reduced and alkylated form of recombinant AMA-1, as previously reported for blood stages (10). Antibodies raised against AMA-1 of the heterologous FVO strain also inhibit NF54 strain sporozoite invasion, suggesting that at least part of the targeted epitopes are conserved. Relatively high concentrations of antibodies induced only partial inhibition of invasion. The neutralizing anti-ama-1 antibodies presumably represent a relatively small proportion of the total rabbit serum IgG preparations used in these experiment s. However, AMA-1 is initially mostly sequestered within the sporozoites, and therefore may not be readily accessible to antibodies, as previously reported with TRAP (25). Much higher degrees of inhibition have been reported with P. falciparum merozoite invasion, using the same rabbit IgG preparations (10,11). Interestingly, AMA-1 relocates to the merozoite surface upon schizont rupture, so that most of the free merozoites express AMA-1 on their surface (10,14,31). These differences in AMA-1 surface exposure may explain the higher level of inhibition 15
16 seen with merozoite as compared to sporozoite invasion. Nevertheless, the inhibition of sporozoite invasion by anti-ama-1 antibodies, though partial, still indicates that AMA-1 could be considered as a potential candidate to be included in a multi-stage malaria vaccine, targeting both erythrocytic and pre-erythrocytic stages. In P. falciparum blood stages, the mature 66 kda AMA -1 relocates to the merozoite surface upon schizont rupture, and is proteolytically cleaved, resulting in the shedding of nearly the whole AMA-1 ectodomain as a 48 kda protein (24), associated with a 44 kda product resulting from an additional intra-domain III cleavage (14), and with a less abundant 52 kda protein consistent with AMA-1 intramembrane cleavage, as described for Toxoplasma microneme proteins (32). Here we show that similar processing events occur for AMA-1 in sporozoites upon incubation at 37 C, leading to the shedding of a major 48 kda protein and of less abundant products of 44 and 52 kda. Similarly, incubation of sporozoites at 37 C induces TRAP processing and shedding of a fragment 5-10 kda smaller than fulllength TRAP, consistent with the size of the TRAP ectodomain and confirming that TRAP is released after cleavage of the protein, as suggested previously (26,33). Both AMA-1 and TRAP processing are observed in the absence of host cells, clearly demonstrating the parasite origin of the protease(s) involved. Still, incubation of P. falciparum sporozoites at 37 C in the presence of human hepatocytes up-regulates the shedding of both AMA-1 and TRAP in the culture supernatants. This effect is likely due to enhanced microneme exocytosis induced by host cells (25,30). In both merozoites (14) and sporozoites (this study), AMA -1 translocation to the parasite surface and its subsequent processing are not sensitive to cytochalasin D, suggesting that microneme exocytosis in Plasmodium is independent of the actin-dependent gliding machinery. 16
17 Strikingly, using a set of protease inhibitors, we observed exactly the same profile of activity on both AMA-1 and TRAP processing. Chymostatin, leupeptin, pepstatin A and PMSF are ineffective, while TLCK inhibits in a dose-dependent fashion the processing of both proteins. Another chloromethyl ketone, Z-GML- CH 2 Cl, initially designed to inhibit merozoite surface protein 1 (MSP-1) processing in P. falciparum merozoites (14), also inhibits both AMA-1 and TRAP processing. In P. falciparum merozoites, a single protease is thought to process both AMA -1 and MSP-1 (14), this protease being extremely sensitive to PMSF but not to TLCK. In contrast, PMSF does not inhibit AMA -1 and TRAP processing in sporozoites while TLCK is very effective. This finding raises the interesting hypothesis that the protease mediating AMA-1 and TRAP processing in sporozoites may not be the same as the one mediating AMA -1 and MSP-1 processing in merozoites. Thus, stage -specific regulation of protein processing may occur in P. falciparum through the expression and/or activity of distinct proteases during the parasite life cycle. Treatment of P. falciparum sporozoites with the inhibitors TLCK and Z-GML- CH 2 Cl, which both irreversibly block AMA-1 and TRAP processing, inhibits the gliding motility of sporozoites, and more importantly, their ability to invade human hepatocytes. These results suggest that proteolytic processing of sporozoite proteins is essential for sporozoite motility and infectivity, but do not allow us to conclude that the loss of sporozoite motility and infectivity results solely from inhibition of TRAP and/or AMA-1 processing. It has been proposed that posterior translocation of sporozoite TRAP provides the force for gliding and host cell invasion via interaction with the parasite cytoskeleton (2). This critical role of TRAP has been demonstrated using gene knock-out (34) or amino acid substitution experiments, which have notably suggested that the TRAP cytoplasmic tail mediates anterior to posterior 17
18 redistribution and posterior shedding of the protein, both functions crucial for sporozoite gliding motility and host cell invasion (33). Our inhibition results are consistent with this proposed critical role of TRAP. Unlike TRAP, which is not expressed during erythrocytic stages, AMA -1 plays a central role during Plasmodium erythrocytic multiplication, so that ama-1 gene knock-out is not possible (7). Alternative strategies will be required, including conditional gene inactivation, to confirm that AMA-1 is required during sporozoite invasion of hepatocytes. Further studies will be needed to identify and characterize the sporozoite proteases involved in AMA-1 and TRAP processing. Even though other processing events may be affected by TLCK and Z-GML-CH 2 Cl, our results nevertheless demonstrate that interfering with proteolytic processing inhibits sporozoite infectivity, and suggest that sporozoite serine proteases may constitute potential drug targets for preventive antimalarial strategies. 18
19 ACKNOWLEDGEMENTS We thank Laurent Rénia and Claude Boucheix for critically reviewing the manuscript. 19
20 REFERENCES 1. Kappe, S. H., Kaiser, K., and Matuschewski, K. (2003) Trends Parasitol 19, Menard, R. (2001) Cell Microbiol 3, Menard, R. (2000) Microbes Infect 2, Florens, L., Washburn, M. P., Raine, J. D., Anthony, R. M., Grainger, M., Haynes, J. D., Moch, J. K., Muster, N., Sacci, J. B., Tabb, D. L., Witney, A. A., Wolters, D., Wu, Y., Gardner, M. J., Holder, A. A., Sinden, R. E., Yates, J. R., and Carucci, D. J. (2002) Nature 419, Healer, J., Crawford, S., Ralph, S., McFadden, G., and Cowman, A. F. (2002) Infect Immun 70, Bannister, L. H., Hopkins, J. M., Dluzewski, A. R., Margos, G., Williams, I. T., Blackman, M. J., Kocken, C. H., Thomas, A. W., and Mitchell, G. H. (2003) J Cell Sci 116, Triglia, T., Healer, J., Caruana, S. R., Hodder, A. N., Anders, R. F., Crabb, B. S., and Cowman, A. F. (2000) Mol Microbiol 38, Mueller, M. S., Renard, A., Boato, F., Vogel, D., Naegeli, M., Zurbriggen, R., Robinson, J. A., and Pluschke, G. (2003) Infect Immun 71, Coley, A. M., Campanale, N. V., Casey, J. L., Hodder, A. N., Crewther, P. E., Anders, R. F., Tilley, L. M., and Foley, M. (2001) Protein Eng 14, Hodder, A. N., Crewther, P. E., and Anders, R. F. (2001) Infect Immun 69, Kocken, C. H., Withers-Martinez, C., Dubbeld, M. A., van der Wel, A., Hackett, F., Valderrama, A., Blackman, M. J., and Thomas, A. W. (2002) Infect Immun 70,
21 12. Charoenvit, Y., Fallarme, V., Rogers, W. O., Sacci, J. B., Jr., Kaur, M., Aguiar, J. C., Yuan, L. F., Corradin, G., Andersen, E., Wizel, B., Houghten, R. A., Oloo, A., De la Vega, P., and Hoffman, S. L. (1997) Infect Immun 65, Stuber, D., Bannwarth, W., Pink, J. R., Meloen, R. H., and Matile, H. (1990) Eur J Immunol 20, Howell, S. A., Wells, I., Fleck, S. L., Kettleborough, C., Collins, C., and Blackman, M. J. (2003) J Biol Chem 278, Ponnudurai, T., Meuwissen, J. H., Leeuwenberg, A. D., Verhave, J. P., and Lensen, A. H. (1982) Trans R Soc Trop Med Hyg 76, Guguen-Guillouzo, C., Campion, J. P., Brissot, P., Glaise, D., Launois, B., Bourel, M., and Guillouzo, A. (1982) Cell Biol Int Rep 6, Meis, J. F., Rijntjes, P. J., Verhave, J. P., Ponnudurai, T., Hollingdale, M. R., and Yap, S. H. (1985) Cell Biol Int Rep 9, Mazier, D., Beaudoin, R. L., Mellouk, S., Druilhe, P., Texier, B., Trosper, J., Miltgen, F., Landau, I., Paul, C., Brandicourt, O., and et al. (1985) Science 227, Isom, H. C., Secott, T., Georgoff, I., Woodworth, C., and Mummaw, J. (1985) Proc Natl Acad Sci U S A 82, Druilhe, P., Pradier, O., Marc, J. P., Miltgen, F., Mazier, D., and Parent, G. (1986) Infect Immun 53, Fichera, M. E., and Roos, D. S. (1997) Nature 390, Kohler, S., Delwiche, C. F., Denny, P. W., Tilney, L. G., Webster, P., Wilson, R. J., Palmer, J. D., and Roos, D. S. (1997) Science 275,
22 23. Crewther, P. E., Culvenor, J. G., Silva, A., Cooper, J. A., and Anders, R. F. (1990) Exp Parasitol 70, Howell, S. A., Withers -Martinez, C., Kocken, C. H., Thomas, A. W., and Blackman, M. J. (2001) J Biol Chem 276, Gantt, S., Persson, C., Rose, K., Birkett, A. J., Abagyan, R., and Nussenzweig, V. (2000) Infect Immun 68, Bhanot, P., Frevert, U., Nussenzweig, V., and Persson, C. (2003) Mol Biochem Parasitol 126, Carruthers, V. B., Giddings, O. K., and Sibley, L. D. (1999) Cell Microbiol 1, Bergman, L. W., Kaiser, K., Fujioka, H., Coppens, I., Daly, T. M., Fox, S., Matuschewski, K., Nussenzweig, V., and Kappe, S. H. (2003) J Cell Sci 116, Stewart, M. J., and Vanderberg, J. P. (1991) J Protozool 38, Mota, M. M., Hafalla, J. C., and Rodriguez, A. (2002) Nat Med 8, Narum, D. L., and Thomas, A. W. (1994) Mol Biochem Parasitol 67, Opitz, C., Di Cristina, M., Reiss, M., Ruppert, T., Crisanti, A., and Soldati, D. (2002) Embo J 21, Kappe, S., Bruderer, T., Gantt, S., Fujioka, H., Nussenzweig, V., and Menard, R. (1999) J Cell Biol 147, Sultan, A. A., Thathy, V., Frevert, U., Robson, K. J., Crisanti, A., Nussenzweig, V., Nussenzweig, R. S., and Menard, R. (1997) Cell 90,
23 FOOTNOTES 1 The abbreviations used are: AMA-1, apical membrane antigen 1; BSA, bovine serum albumin; CSP, circumsporozoite protein; DAPI, 4,6-diamidino-2-phenylindole; DMSO, dimethyl sulfoxide; DTT, dithiothreitol; FITC, fluorescein isothiocyanate; HSP-70, heat shock protein-70; IFA, immunofluorescence assay; mab, monoclonal antibody; MSP-1, merozoite surface protein-1; PAGE, polyacrylamide gel electrophoresis; PBS, phosphate buffer saline; PFA, paraformaldehyde; PMSF, phenylmethylsulfonyl fluoride; RT-PCR, reverse transcriptase-polymerase chain reaction; SDS, sodium dodecyl sulfate; FCS, fetal calf serum; TLCK, N-tosyl-Llysine chloromethyl ketone; TRAP, thrombospondin -related anonymous protein; Z - GML-CH2Cl, N-benzyloxycarbonyl-Gly-Met-Leu-chloromethyl ketone. 2 Silvie, O., unpublished observations. 23
24 FIGURE LEGENDS Fig. 1. AMA-1 is expressed in P. falciparum sporozoites. A. RT-PCR analysis of ama -1 expression in P. falciparum NF54 sporozoites (spz), with (+) or without (-) reverse transcriptase; M = size marker; G = 3D7 P. falciparum genomic DNA. The arrowhead indicates the 890 base pair amplicon. B. Immunofluorescence microscopy of air-dried permeabilized P. falciparum sporozoites stained with anti-ama-1 mab DV5 (green) and DAPI (blue). Bar = 5µm. C. Surface immunofluorescence of P. falciparum sporozoites stained with anti-ama- 1 mab DV5 (green) and DAPI (blue). The sporozoite nucleus is indicated with an arrow and the apicoplast with an arrowhead. Bar = 5 µm. D. Western blotting of lysates from non infected mosquito salivary glands (sg), P. falciparum sporozoite infected salivary glands (spz) and P. falciparum infected erythrocytes (e), using the anti-ama-1 mab DV5. E. Western blotting of lysates from P. falciparum sporozoites using the anti-ama-1 mabs 1F9 and 5G8. Fig. 2. AMA-1 is lost after invasion of hepato cytes and re -expressed in liver merozoites. P. falciparum liver schizonts (arrowheads) stained with the anti-ama-1 mab DV5 (green) and DAPI (blue), 3 days, 8 days or 10 days after infection of human hepatocyte cultures with sporozoites. N = hepatocyte nucleus. Bars = 5 µm. Fig. 3. Anti-AMA-1 polyclonal antibodies inhibit P. falciparum sporozoite invasion of human hepatocytes in vitro. Human hepatocytes were inoculated with P. falciparum NF54 sporozoites in the presence of IgG from rabbits immunized with a 3D7 clone PfAMA-1 recombinant protein (grey bars), with a reduced and alkylated 24
25 form of this 3D7 PfAMA-1 recombinant protein (white bar), or with a FVO strain PfAMA -1 recombinant protein (black bar). Results are expressed as the mean of % inhibition of at least two independent experiments, each done in triplicate wells, +/- standard deviation. Fig. 4. Upon incubation of sporozoites at 37 C, P. falciparum AMA-1 and TRAP undergo proteolytic processing and shedding, which is sensitive to a subset of serine protease inhibitors but not to cytochalasin D. A. Western blotting of sporozoite lysates probed with anti-ama-1 polyclonal rabbit IgG. The arrows indicate the 48 and 52 kda AMA-1 cleavage products, respectively. B. Western blotting of sporozoite lysates probed with anti-trap mab SSP2.2. The arrowhead indicates the TRAP cleavage product. C. Western blotting of pellet (P) and supernatant (SN) from sporozoites incubated at 37 C, probed with anti-ama-1 polyclonal rabbit IgG or with anti-trap mab SSP2.2. The arrows and the arrowhead indicate the cleavage products of AMA-1 and TRAP, respectively. D. Western blotting of supernatants from sporozoites incubated at 37 C, run under reducing (DTT +) or non reducing (DTT -) conditions and probed with anti-ama-1 polyclonal rabbit IgG. E. Western blotting of pellet (P) and supernatant (SN) from sporozoites kept at 4 C or incubated at 37 C, in the absence (-hep) or presence (+hep) of hepatocytes, run under reducing conditions and probed with anti-ama-1 polyclonal rabbit IgG or with anti- TRAP mab SSP2.2. The arrow and the arrowhead indicate the 48 kda AMA-1 and the TRAP cleavage products, respectively. 25
26 F. Western blotting of supernatants from sporozoites treated with protease inhibitors TLCK and Z -GML-CH 2 Cl before incubation at 37 C. Samples were run under reducing conditions and probed with anti-ama-1 polyclonal rabbit IgG, anti-trap mab SSP2.2 or anti-csp mab E9. The arrows and the arrowhead indicate the cleavage products of AMA-1 and TRAP, respectively. G. Western blotting of supernatants from sporozoites treated with PMSF or cytochalasin D and incubated at 37 C. Samples were run under reducing conditions and probed with anti-ama-1 polyclonal rabbit IgG or anti-trap mab SSP2.2. The arrows and the arrowhead indicate the cleava ge products of AMA-1 and TRAP, respectively. Fig. 5. Protease inhibitors and cytochalasin D do not prevent AMA-1 translocation to the sporozoite surface upon incubation at 37 C. Immunofluorescence analysis of AMA-1 surface exposure using anti-ama-1 polyclonal rabbit IgG (red) and DAPI (blue). (A) untreated sporozoites kept at 4 C after isolation from salivary glands ; (B) untreated sporozoites incubated at 37 C for 1h; (C) sporozoites treated with 200 µm TLCK and incubated at 37 C for 1h; (D) sporozoites treated with 100 µm Z-GML-CH 2 Cl and incubated at 37 C for 1h; (E) sporozoites treated with 5µM cytochalasin D and incubated at 37 C for 1h. The arrow and the arrowhead indicate the sporozoite nucleus and the apicoplast, respectively. Bar = 5µm. Fig. 6. Only the mature AMA-1 protein translocates to the sporozoite surface upon incubation at 37 C. Immunofluorescence analysis with anti-ama-1 mabs 5G8 (A and C, green) or 1F9 (B and D, green) and DAPI (blue), in air-dried 26
27 permeabilized sporozoites (A and B ) or non-permeabilized sporozoites incubated at 37 C for 1h (C and D). Bar = 5µm. Fig. 7. Serine protease inhibitors TLCK and Z-GML-CH 2 Cl inhibit P. falciparum sporozoite infectivity. Human hepatocytes were inoculated with P. falciparum NF54 sporozoites pretreated with the serine protease inhibitors TLCK and Z-GML-CH 2 Cl. For each inhibitor, the results of two independent experiments are shown and are expressed as the mean percentage of inhibition of invasion +/- standard deviation. *P < 0.05, as determined using the one-way ANOVA followed by the Tukey multiple comparison test. 27
28
29
30
31
32
33
34
35 A role for apical membrane antigen 1 during invasion of hepatocytes by Plasmodium falciparum sporozoites Olivier Silvie, Jean-François Franetich, Stéphanie Charrin, Markus S. Mueller, Anthony Siau, Myriam Bodescot, Eric Rubinstein, Laurent Hannoun, Yupin Charoenvit, Clemens Kocken, Alan W. Thomas, Geert-Jan van Gemert, Robert W. Sauerwein, Michael J. Blackman, Robin F. Anders, Gerd Pluschke and Dominique Mazier J. Biol. Chem. published online December 15, 2003 Access the most updated version of this article at doi: /jbc.M Alerts: When this article is cited When a correction for this article is posted Click here to choose from all of JBC's alerts
A Role for Apical Membrane Antigen 1 during Invasion of Hepatocytes by Plasmodium falciparum Sporozoites*
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 10, Issue of March 5, pp. 9490 9496, 2004 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. A Role for Apical
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 10, Issue of March 5, pp. 9490 9496, 2004 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. A Role for Apical
Gliding Motility Assay for P. berghei Sporozoites
 Gliding Motility Assay for P. berghei Sporozoites Important Notes: 1. For all dilutions (including antibodies and sporozoites), always make slightly more than needed. For instance, if you need 200 µl sporozoites
Gliding Motility Assay for P. berghei Sporozoites Important Notes: 1. For all dilutions (including antibodies and sporozoites), always make slightly more than needed. For instance, if you need 200 µl sporozoites
INVESTIGATING THE MOTILITY OF PLASMODIUM
 INVESTIGATING THE MOTILITY OF PLASMODIUM by Natasha Vartak A thesis submitted to Johns Hopkins University in conformity with the requirements for the degree of Master of Science Baltimore, Maryland April,
INVESTIGATING THE MOTILITY OF PLASMODIUM by Natasha Vartak A thesis submitted to Johns Hopkins University in conformity with the requirements for the degree of Master of Science Baltimore, Maryland April,
CelTOS, a novel malarial protein that mediates transmission to mosquito and vertebrate hosts
 Blackwell Publishing LtdOxford, UKMMIMolecular Microbiology0950-382X 2005 The Authors; Journal compilation 2005 Blackwell Publishing Ltd? 200559513691379Original ArticleA protein that mediates malarial
Blackwell Publishing LtdOxford, UKMMIMolecular Microbiology0950-382X 2005 The Authors; Journal compilation 2005 Blackwell Publishing Ltd? 200559513691379Original ArticleA protein that mediates malarial
PCR detection of Leptospira in. stray cat and
 PCR detection of Leptospira in 1 Department of Pathology, School of Veterinary Medicine, Islamic Azad University, Shahrekord Branch, Shahrekord, Iran 2 Department of Microbiology, School of Veterinary
PCR detection of Leptospira in 1 Department of Pathology, School of Veterinary Medicine, Islamic Azad University, Shahrekord Branch, Shahrekord, Iran 2 Department of Microbiology, School of Veterinary
PLASMODIUM MODULE 39.1 INTRODUCTION OBJECTIVES 39.2 MALARIAL PARASITE. Notes
 Plasmodium MODULE 39 PLASMODIUM 39.1 INTRODUCTION Malaria is characterized by intermittent fever associated with chills and rigors in the patient. There may be enlargement of the liver and spleen in the
Plasmodium MODULE 39 PLASMODIUM 39.1 INTRODUCTION Malaria is characterized by intermittent fever associated with chills and rigors in the patient. There may be enlargement of the liver and spleen in the
Identification of an AP2-family Protein That Is Critical for Malaria Liver Stage Development
 Identification of an AP2-family Protein That Is Critical for Malaria Liver Stage Development Shiroh Iwanaga, Izumi Kaneko, Tomomi Kato, Masao Yuda* Department of Medical Zoology, Mie University School
Identification of an AP2-family Protein That Is Critical for Malaria Liver Stage Development Shiroh Iwanaga, Izumi Kaneko, Tomomi Kato, Masao Yuda* Department of Medical Zoology, Mie University School
A Cysteine Protease Inhibitor of Plasmodium berghei Is Essential for Exo-erythrocytic Development
 A Cysteine Protease Inhibitor of Plasmodium berghei Is Essential for Exo-erythrocytic Development Christine Lehmann 1, Anna Heitmann 1, Satish Mishra 2, Paul-Christian Burda 3, Mirko Singer 4, Monica Prado
A Cysteine Protease Inhibitor of Plasmodium berghei Is Essential for Exo-erythrocytic Development Christine Lehmann 1, Anna Heitmann 1, Satish Mishra 2, Paul-Christian Burda 3, Mirko Singer 4, Monica Prado
Consuelo Pinzon-Ortiz, Jennifer Friedman, Jeffrey Esko, and Photini Sinnis
 THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 276, No. 29, Issue of July 20, pp. 26784 26791, 2001 2001 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The Binding of
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 276, No. 29, Issue of July 20, pp. 26784 26791, 2001 2001 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. The Binding of
The Transmembrane Isoform of Plasmodium falciparum MAEBL Is Essential for the Invasion of Anopheles Salivary Glands
 The Transmembrane Isoform of Plasmodium falciparum MAEBL Is Essential for the Invasion of Anopheles Salivary Glands Fabian E. Saenz 1,2, Bharath Balu 1, Jonah Smith 2, Sarita R. Mendonca 1,2, John H. Adams
The Transmembrane Isoform of Plasmodium falciparum MAEBL Is Essential for the Invasion of Anopheles Salivary Glands Fabian E. Saenz 1,2, Bharath Balu 1, Jonah Smith 2, Sarita R. Mendonca 1,2, John H. Adams
ACCEPTED. Parasitology Unit, Max Planck Institute for Infection Biology, Berlin, Germany
 EC Accepts, published online ahead of print on 30 January 2009 Eukaryotic Cell doi:10.1128/ec.00347-08 Copyright 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights
EC Accepts, published online ahead of print on 30 January 2009 Eukaryotic Cell doi:10.1128/ec.00347-08 Copyright 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights
Received 6 December 2000/Returned for modification 29 January 2001/Accepted 26 March 2001
 INFECTION AND IMMUNITY, June 2001, p. 3845 3852 Vol. 69, No. 6 0019-9567/01/$04.00 0 DOI: 10.1128/IAI.69.6.3845 3952.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Human Antibodies
INFECTION AND IMMUNITY, June 2001, p. 3845 3852 Vol. 69, No. 6 0019-9567/01/$04.00 0 DOI: 10.1128/IAI.69.6.3845 3952.2001 Copyright 2001, American Society for Microbiology. All Rights Reserved. Human Antibodies
Infecting Anopheles stephensi With Rodent Malaria Parasites Alida Coppi & Photini Sinnis
 Infecting Anopheles stephensi With Rodent Malaria Parasites Alida Coppi & Photini Sinnis A. Reagents: 1. DMEM or RPMI DMEM (4.5g/L glucose) RPMI 1640 Cellgro #MT-10-017-CM Cellgro #MT-10-040-CM 2. Giemsa
Infecting Anopheles stephensi With Rodent Malaria Parasites Alida Coppi & Photini Sinnis A. Reagents: 1. DMEM or RPMI DMEM (4.5g/L glucose) RPMI 1640 Cellgro #MT-10-017-CM Cellgro #MT-10-040-CM 2. Giemsa
Quantitative Dynamics of Plasmodium yoelii Sporozoite Transmission by Infected Anopheline Mosquitoes
 INFECTION AND IMMUNITY, July 2005, p. 4363 4369 Vol. 73, No. 7 0019-9567/05/$08.00 0 doi:10.1128/iai.73.7.4363 4369.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Quantitative
INFECTION AND IMMUNITY, July 2005, p. 4363 4369 Vol. 73, No. 7 0019-9567/05/$08.00 0 doi:10.1128/iai.73.7.4363 4369.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Quantitative
Marissa Vignali*, Cate Speake* and Patrick E Duffy*
 Minireview Malaria sporozoite proteome leaves a trail Marissa Vignali*, Cate Speake* and Patrick E Duffy* Addresses: *Malaria Program, Seattle Biomedical Research Institute, Seattle, Washington 98109,
Minireview Malaria sporozoite proteome leaves a trail Marissa Vignali*, Cate Speake* and Patrick E Duffy* Addresses: *Malaria Program, Seattle Biomedical Research Institute, Seattle, Washington 98109,
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12234 Supplementary Figure 1. Embryonic naked mole-rat fibroblasts do not undergo ECI. Embryonic naked mole-rat fibroblasts ( EF) were isolated from eight mid-gestation embryos. All the
doi:10.1038/nature12234 Supplementary Figure 1. Embryonic naked mole-rat fibroblasts do not undergo ECI. Embryonic naked mole-rat fibroblasts ( EF) were isolated from eight mid-gestation embryos. All the
Plasmodium yoelii Sporozoites with Simultaneous Deletion of P52 and P36 Are Completely Attenuated and Confer Sterile Immunity against Infection
 INFECTION AND IMMUNITY, Aug. 2007, p. 3758 3768 Vol. 75, No. 8 0019-9567/07/$08.00 0 doi:10.1128/iai.00225-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Plasmodium yoelii Sporozoites
INFECTION AND IMMUNITY, Aug. 2007, p. 3758 3768 Vol. 75, No. 8 0019-9567/07/$08.00 0 doi:10.1128/iai.00225-07 Copyright 2007, American Society for Microbiology. All Rights Reserved. Plasmodium yoelii Sporozoites
Parasitology Departement Medical Faculty of USU
 Malaria Mechanism of infection Parasitology Departement Medical Faculty of USU Introduction Malaria parasites Phylum Order Suborder Family Genus Species : : Apicomplexa : Eucoccidiida : Haemosporida :
Malaria Mechanism of infection Parasitology Departement Medical Faculty of USU Introduction Malaria parasites Phylum Order Suborder Family Genus Species : : Apicomplexa : Eucoccidiida : Haemosporida :
PDF hosted at the Radboud Repository of the Radboud University Nijmegen
 PDF hosted at the Radboud Repository of the Radboud University Nijmegen The following full text is a publisher's version. For additional information about this publication click this link. http://hdl.handle.net/2066/70169
PDF hosted at the Radboud Repository of the Radboud University Nijmegen The following full text is a publisher's version. For additional information about this publication click this link. http://hdl.handle.net/2066/70169
Medical Genetics and Diagnosis Lab #3. Gel electrophoresis
 Medical Genetics and Diagnosis Lab #3 Gel electrophoresis Background Information Gel electrophoresis is the standard lab procedure for separating DNA by size (e.g. length in base pairs) for visualization
Medical Genetics and Diagnosis Lab #3 Gel electrophoresis Background Information Gel electrophoresis is the standard lab procedure for separating DNA by size (e.g. length in base pairs) for visualization
PRINCIPAL INVESTIGATOR: Dr. Jetsumon (Sattabongkot) Prachumsri
 Prachumsri") AD (Leave blank) Award Number: W81XWH-07-2-0090 TITLE: Proteomic Study of Human Malaria Parasite Plasmodium Vivax Liver Stages for Development of Vaccines and Drugs PRINCIPAL INVESTIGATOR: Dr. Jetsumon
AD (Leave blank) Award Number: W81XWH-07-2-0090 TITLE: Proteomic Study of Human Malaria Parasite Plasmodium Vivax Liver Stages for Development of Vaccines and Drugs PRINCIPAL INVESTIGATOR: Dr. Jetsumon
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
Malaria in the Mosquito Dr. Peter Billingsley
 Malaria in the Mosquito Senior Director Quality Systems and Entomology Research Sanaria Inc. Rockville MD. 1 Malaria: one of the world s foremost killers Every year 1 million children die of malaria 250
Malaria in the Mosquito Senior Director Quality Systems and Entomology Research Sanaria Inc. Rockville MD. 1 Malaria: one of the world s foremost killers Every year 1 million children die of malaria 250
Malaria parasite exit from the host erythrocyte: A two-step process requiring extraerythrocytic proteolysis
 Malaria parasite exit from the host erythrocyte: A two-step process requiring extraerythrocytic proteolysis Brandy L. Salmon, Anna Oksman, and Daniel E. Goldberg* Howard Hughes Medical Institute, Departments
Malaria parasite exit from the host erythrocyte: A two-step process requiring extraerythrocytic proteolysis Brandy L. Salmon, Anna Oksman, and Daniel E. Goldberg* Howard Hughes Medical Institute, Departments
Developmental Biology of Sporozoite-Host. Malaria: Implications for Vaccine Design. Javier E. Garcia, Alvaro Puentes and Manuel E.
 Developmental Biology of Sporozoite-Host Interactions in Plasmodium falciparum Malaria: Implications for Vaccine Design Javier E. Garcia, Alvaro Puentes and Manuel E. Patarroyo Clin. Microbiol. Rev. 2006,
Developmental Biology of Sporozoite-Host Interactions in Plasmodium falciparum Malaria: Implications for Vaccine Design Javier E. Garcia, Alvaro Puentes and Manuel E. Patarroyo Clin. Microbiol. Rev. 2006,
The silent path to thousands of merozoites: the Plasmodium liver stage
 The silent path to thousands of merozoites: the Plasmodium liver stage Miguel Prudêncio*, Ana Rodriguez and Maria M. Mota* Abstract Plasmodium sporozoites are deposited in the skin of their vertebrate
The silent path to thousands of merozoites: the Plasmodium liver stage Miguel Prudêncio*, Ana Rodriguez and Maria M. Mota* Abstract Plasmodium sporozoites are deposited in the skin of their vertebrate
Evaluation of Different Antigens in Western Blotting Technique for the Diagnosis of Sheep Haemonchosis
 Original Article Evaluation of Different Antigens in Western Blotting Technique for the Diagnosis of Sheep Haemonchosis *B Meshgi, SH Hosseini Dept. of Parasitology, Faculty of Veterinary Medicine, University
Original Article Evaluation of Different Antigens in Western Blotting Technique for the Diagnosis of Sheep Haemonchosis *B Meshgi, SH Hosseini Dept. of Parasitology, Faculty of Veterinary Medicine, University
Diurnal variation in microfilaremia in cats experimentally infected with larvae of
 Hayasaki et al., Page 1 Short Communication Diurnal variation in microfilaremia in cats experimentally infected with larvae of Dirofilaria immitis M. Hayasaki a,*, J. Okajima b, K.H. Song a, K. Shiramizu
Hayasaki et al., Page 1 Short Communication Diurnal variation in microfilaremia in cats experimentally infected with larvae of Dirofilaria immitis M. Hayasaki a,*, J. Okajima b, K.H. Song a, K. Shiramizu
Malaria. This sheet is from both sections recording and includes all slides and diagrams.
 Malaria This sheet is from both sections recording and includes all slides and diagrams. Malaria is caused by protozoa family called plasmodium (Genus) mainly affect blood system specially RBCs and each
Malaria This sheet is from both sections recording and includes all slides and diagrams. Malaria is caused by protozoa family called plasmodium (Genus) mainly affect blood system specially RBCs and each
A. Effect upon human culture 1. Control of malaria has contributed to world=s population explosion 2. Africans brought to U.S.
 VI. Malaria A. Effect upon human culture 1. Control of malaria has contributed to world=s population explosion 2. Africans brought to U.S. because they were resistant to malaria & other diseases 3. Many
VI. Malaria A. Effect upon human culture 1. Control of malaria has contributed to world=s population explosion 2. Africans brought to U.S. because they were resistant to malaria & other diseases 3. Many
NOTES. Received 28 August 2001/Returned for modification 17 October 2001/Accepted 3 December 2001
 INFECTION AND IMMUNITY, Mar. 2002, p. 1599 1603 Vol. 70, No. 3 0019-9567/02/$04.00 0 DOI: 10.1128/IAI.70.3.1599 1603.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. NOTES Babesia
INFECTION AND IMMUNITY, Mar. 2002, p. 1599 1603 Vol. 70, No. 3 0019-9567/02/$04.00 0 DOI: 10.1128/IAI.70.3.1599 1603.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. NOTES Babesia
Chimeric Plasmodium falciparum parasites expressing Plasmodium vivax circumsporozoite protein fail to produce salivary gland sporozoites
 https://doi.org/10.1186/s12936-018-2431-1 Malaria Journal RESEARCH Open Access Chimeric Plasmodium falciparum parasites expressing Plasmodium vivax circumsporozoite protein fail to produce salivary gland
https://doi.org/10.1186/s12936-018-2431-1 Malaria Journal RESEARCH Open Access Chimeric Plasmodium falciparum parasites expressing Plasmodium vivax circumsporozoite protein fail to produce salivary gland
Arrested oocyst maturation in Plasmodium parasites. lacking type II NADH:ubiquinone dehydrogenase
 Supplemental Information for: Arrested oocyst maturation in Plasmodium parasites lacking type II NADH:ubiquinone dehydrogenase Katja E. Boysen and Kai Matuschewski Contents: - Supplemental Movies 1 and
Supplemental Information for: Arrested oocyst maturation in Plasmodium parasites lacking type II NADH:ubiquinone dehydrogenase Katja E. Boysen and Kai Matuschewski Contents: - Supplemental Movies 1 and
Developmentally Regulated!nfectivity of Malaria Sporozoites for Mosquito Salivary Glands and the Vertebrate Host
 Developmentally Regulated!nfectivity of Malaria Sporozoites for Mosquito Salivary Glands and the Vertebrate Host By Musa G. Touray, Alon Warburg, Andre Laughinghouse, Antoniana U. Krettli,* and Louis H.
Developmentally Regulated!nfectivity of Malaria Sporozoites for Mosquito Salivary Glands and the Vertebrate Host By Musa G. Touray, Alon Warburg, Andre Laughinghouse, Antoniana U. Krettli,* and Louis H.
alaria Parasite Bank Collection sites of P. falciparum isolates PARASITE BIOLOGY
 M alaria Parasite Bank established in 1992 is a supporting unit for research activities on different aspects of malaria. The main objective of establishing this facility is to strengthen researches at
M alaria Parasite Bank established in 1992 is a supporting unit for research activities on different aspects of malaria. The main objective of establishing this facility is to strengthen researches at
Motility precedes egress of malaria parasites from oocysts
 RESEARCH ARTICLE Motility precedes egress of malaria parasites from oocysts Dennis Klug*, Friedrich Frischknecht* Integrative Parasitology, Center for Infectious Diseases, Heidelberg University Medical
RESEARCH ARTICLE Motility precedes egress of malaria parasites from oocysts Dennis Klug*, Friedrich Frischknecht* Integrative Parasitology, Center for Infectious Diseases, Heidelberg University Medical
Antimalarial Activity of Allicin, a Biologically Active Compound from Garlic Cloves
 ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, May 2006, p. 1731 1737 Vol. 50, No. 5 0066-4804/06/$08.00 0 doi:10.1128/aac.50.5.1731 1737.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved.
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, May 2006, p. 1731 1737 Vol. 50, No. 5 0066-4804/06/$08.00 0 doi:10.1128/aac.50.5.1731 1737.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved.
Plasmodium vivax: A Monoclonal Antibody Recognizes a Circumsporozoite Protein Precursor on the Sporozoite Surface
 University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln US Army Research U.S. Department of Defense 1998 Plasmodium vivax: A Monoclonal Antibody Recognizes a Circumsporozoite Protein
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln US Army Research U.S. Department of Defense 1998 Plasmodium vivax: A Monoclonal Antibody Recognizes a Circumsporozoite Protein
Exposure of Plasmodium sporozoites to the intracellular concentration of potassium enhances infectivity and reduces cell passage activity
 Molecular & Biochemical Parasitology 156 (2007) 32 40 Exposure of Plasmodium sporozoites to the intracellular concentration of potassium enhances infectivity and reduces cell passage activity Kota Arun
Molecular & Biochemical Parasitology 156 (2007) 32 40 Exposure of Plasmodium sporozoites to the intracellular concentration of potassium enhances infectivity and reduces cell passage activity Kota Arun
CIRCUMSPOROZOITE PROTEINS OF HUMAN MALARIA PARASITES PLASMODIUM FALCIPARUM AND PLASMODIUM VIVA,F*
 CIRCUMSPOROZOITE PROTEINS OF HUMAN MALARIA PARASITES PLASMODIUM FALCIPARUM AND PLASMODIUM VIVA,F* BY ELIZABETH H. NARDIN, VICTOR NUSSENZWEIG, RUTH S. NUSSENZWEIG, WILLIAM E. COLLINS, K. TRANAKCHIT HARINASUTA,
CIRCUMSPOROZOITE PROTEINS OF HUMAN MALARIA PARASITES PLASMODIUM FALCIPARUM AND PLASMODIUM VIVA,F* BY ELIZABETH H. NARDIN, VICTOR NUSSENZWEIG, RUTH S. NUSSENZWEIG, WILLIAM E. COLLINS, K. TRANAKCHIT HARINASUTA,
Protozoan parasites of the genus Plasmodium are the causative
 Exploring the transcriptome of the malaria sporozoite stage Stefan H. I. Kappe*, Malcolm J. Gardner, Stuart M. Brown, Jessica Ross*, Kai Matuschewski*, Jose M. Ribeiro, John H. Adams, John Quackenbush,
Exploring the transcriptome of the malaria sporozoite stage Stefan H. I. Kappe*, Malcolm J. Gardner, Stuart M. Brown, Jessica Ross*, Kai Matuschewski*, Jose M. Ribeiro, John H. Adams, John Quackenbush,
Novel ELISA method as exploratory tool to assess immunity induced by radiated attenuated sporozoites to decipher protective immunity
 DOI 10.1186/s12936-017-2129-9 Malaria Journal METHODOLOGY Open Access Novel ELISA method as exploratory tool to assess immunity induced by radiated attenuated sporozoites to decipher protective immunity
DOI 10.1186/s12936-017-2129-9 Malaria Journal METHODOLOGY Open Access Novel ELISA method as exploratory tool to assess immunity induced by radiated attenuated sporozoites to decipher protective immunity
ELECTROPHORETIC ANALYSIS OF SERUM PROTEINS OF BIRDS AND MAMMALS
 ELECTROPHORETIC ANALYSIS OF SERUM PROTEINS OF BIRDS AND MAMMALS Emanuel G. E. HELAL 1, Samir A. M. ZAHKOUK 1, Hamdy A. MEKKAWY 2 1 Zoology Department, Faculty of Science, Al-Azhar University for Girls,
ELECTROPHORETIC ANALYSIS OF SERUM PROTEINS OF BIRDS AND MAMMALS Emanuel G. E. HELAL 1, Samir A. M. ZAHKOUK 1, Hamdy A. MEKKAWY 2 1 Zoology Department, Faculty of Science, Al-Azhar University for Girls,
Enzootic Bovine Leukosis: Milk Screening and Verification ELISA: VF-P02210 & VF-P02220
 Enzootic Bovine Leukosis: Milk Screening and Verification ELISA: VF-P02210 & VF-P02220 Introduction Enzootic Bovine Leukosis is a transmissible disease caused by the Enzootic Bovine Leukosis Virus (BLV)
Enzootic Bovine Leukosis: Milk Screening and Verification ELISA: VF-P02210 & VF-P02220 Introduction Enzootic Bovine Leukosis is a transmissible disease caused by the Enzootic Bovine Leukosis Virus (BLV)
II. MATERIALS AND METHODS
 e- ISSN: 2394-5532 p- ISSN: 2394-823X General Impact Factor (GIF): 0.875 Scientific Journal Impact Factor: 1.205 International Journal of Applied And Pure Science and Agriculture www.ijapsa.com Evaluation
e- ISSN: 2394-5532 p- ISSN: 2394-823X General Impact Factor (GIF): 0.875 Scientific Journal Impact Factor: 1.205 International Journal of Applied And Pure Science and Agriculture www.ijapsa.com Evaluation
Comparative Plasmodium gene overexpression reveals distinct perturbation of sporozoite transmission by profilin
 MBoC ARTICLE Comparative Plasmodium gene overexpression reveals distinct perturbation of sporozoite transmission by profilin Yuko Sato a,b, *, Marion Hliscs a,c, Josefine Dunst a,d, Christian Goosmann
MBoC ARTICLE Comparative Plasmodium gene overexpression reveals distinct perturbation of sporozoite transmission by profilin Yuko Sato a,b, *, Marion Hliscs a,c, Josefine Dunst a,d, Christian Goosmann
Blood protozoan: Plasmodium
 Blood protozoan: Plasmodium Dr. Hala Al Daghistani The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans: four species are associated The Plasmodium spp.
Blood protozoan: Plasmodium Dr. Hala Al Daghistani The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans: four species are associated The Plasmodium spp.
Understanding Epidemics Section 3: Malaria & Modelling
 Understanding Epidemics Section 3: Malaria & Modelling PART B: Biology Contents: Vector and parasite Biology of the malaria parasite Biology of the anopheles mosquito life cycle Vector and parasite Malaria
Understanding Epidemics Section 3: Malaria & Modelling PART B: Biology Contents: Vector and parasite Biology of the malaria parasite Biology of the anopheles mosquito life cycle Vector and parasite Malaria
Visit ABLE on the Web at:
 This article reprinted from: Lessem, P. B. 2008. The antibiotic resistance phenomenon: Use of minimal inhibitory concentration (MIC) determination for inquiry based experimentation. Pages 357-362, in Tested
This article reprinted from: Lessem, P. B. 2008. The antibiotic resistance phenomenon: Use of minimal inhibitory concentration (MIC) determination for inquiry based experimentation. Pages 357-362, in Tested
Blood protozoan: Plasmodium
 Blood protozoan: Plasmodium The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans:four species are associated The Plasmodium spp. life cycle can be divided
Blood protozoan: Plasmodium The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans:four species are associated The Plasmodium spp. life cycle can be divided
Diagnosis of Heartworm (Dirofilaria immitis) Infection in Dogs and Cats by Using Western Blot Technique
 Infection in Dogs and Cats by Using Western Blot Technique") 284 Kasetsart J. (Nat. Sci.) 40 : 284-289 (2006) Kasetsart J. (Nat. Sci.) 40(5) Diagnosis of Heartworm (Dirofilaria immitis) Infection in Dogs and Cats by Using Western Blot Technique Tawin Inpankaew*,
284 Kasetsart J. (Nat. Sci.) 40 : 284-289 (2006) Kasetsart J. (Nat. Sci.) 40(5) Diagnosis of Heartworm (Dirofilaria immitis) Infection in Dogs and Cats by Using Western Blot Technique Tawin Inpankaew*,
FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT
![FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT](/thumbs/93/113164093.jpg "FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT") FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT INSTRUCTION MANUAL Sufficient for 12/120 assays 22 APR 2018 Biogal Galed Laboratories Acs Ltd. tel: 972-4-9898605. fax: 972-4-9898690 e-mail:info@biogal.co.il
FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT INSTRUCTION MANUAL Sufficient for 12/120 assays 22 APR 2018 Biogal Galed Laboratories Acs Ltd. tel: 972-4-9898605. fax: 972-4-9898690 e-mail:info@biogal.co.il
Detection of UV-Induced Thymine Dimers in Individual Cryptosporidium parvum and Cryptosporidium hominis Oocysts by Immunofluorescence Microscopy
 APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Feb. 2007, p. 947 955 Vol. 73, No. 3 0099-2240/07/$08.00 0 doi:10.1128/aem.01251-06 Copyright 2007, American Society for Microbiology. All Rights Reserved. Detection
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Feb. 2007, p. 947 955 Vol. 73, No. 3 0099-2240/07/$08.00 0 doi:10.1128/aem.01251-06 Copyright 2007, American Society for Microbiology. All Rights Reserved. Detection
Running title: Model to down-select human malaria vaccines
 CVI Accepts, published online ahead of print on 27 March 2013 Clin. Vaccine Immunol. doi:10.1128/cvi.00066-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10
CVI Accepts, published online ahead of print on 27 March 2013 Clin. Vaccine Immunol. doi:10.1128/cvi.00066-13 Copyright 2013, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10
Epigenetic regulation of Plasmodium falciparum clonally. variant gene expression during development in An. gambiae
 Epigenetic regulation of Plasmodium falciparum clonally variant gene expression during development in An. gambiae Elena Gómez-Díaz, Rakiswendé S. Yerbanga, Thierry Lefèvre, Anna Cohuet, M. Jordan Rowley,
Epigenetic regulation of Plasmodium falciparum clonally variant gene expression during development in An. gambiae Elena Gómez-Díaz, Rakiswendé S. Yerbanga, Thierry Lefèvre, Anna Cohuet, M. Jordan Rowley,
Malaria Parasite Pre-Erythrocytic Stage Infection: Gliding and Hiding
 Malaria Parasite Pre-Erythrocytic Stage Infection: Gliding and Hiding Ashley M. Vaughan, 1 Ahmed S.I. Aly, 1 and Stefan H.I. Kappe 1,2, * 1 Seattle Biomedical Research Institute, Seattle, WA 98109, USA
Malaria Parasite Pre-Erythrocytic Stage Infection: Gliding and Hiding Ashley M. Vaughan, 1 Ahmed S.I. Aly, 1 and Stefan H.I. Kappe 1,2, * 1 Seattle Biomedical Research Institute, Seattle, WA 98109, USA
Proteasome Inhibitors Block Development of Plasmodium spp.
 ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Oct. 1998, p. 2731 2738 Vol. 42, No. 10 0066-4804/98/$04.00 0 Copyright 1998, American Society for Microbiology. All Rights Reserved. Proteasome Inhibitors Block
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Oct. 1998, p. 2731 2738 Vol. 42, No. 10 0066-4804/98/$04.00 0 Copyright 1998, American Society for Microbiology. All Rights Reserved. Proteasome Inhibitors Block
Sera from 2,500 animals from three different groups were analysed:
 FIELD TRIAL OF A BRUCELLOSIS COMPETITIVE ENZYME LINKED IMMUNOABSORBENT ASSAY (ELISA) L.E. SAMARTINO, R.J. GREGORET, G. SIGAL INTA-CICV Instituto Patobiología Area Bacteriología, Buenos Aires, Argentina
FIELD TRIAL OF A BRUCELLOSIS COMPETITIVE ENZYME LINKED IMMUNOABSORBENT ASSAY (ELISA) L.E. SAMARTINO, R.J. GREGORET, G. SIGAL INTA-CICV Instituto Patobiología Area Bacteriología, Buenos Aires, Argentina
Malaria parasites of rodents of the Congo (Brazzaville) :
 :") Annales de Parasitologie (Paris), 1976, t. 51, n 6, pp. 637 à 646 Malaria parasites of rodents of the Congo (Brazzaville) : Plasmodium cbabaudi adami subsp. nov. and Plasmodium vinckei lentum Landau, Michel,
Annales de Parasitologie (Paris), 1976, t. 51, n 6, pp. 637 à 646 Malaria parasites of rodents of the Congo (Brazzaville) : Plasmodium cbabaudi adami subsp. nov. and Plasmodium vinckei lentum Landau, Michel,
BIO Parasitology Spring 2009
 BIO 475 - Parasitology Spring 2009 Stephen M. Shuster Northern Arizona University http://www4.nau.edu/isopod Lecture 10 Malaria-Life Cycle a. Micro and macrogametocytes in mosquito stomach. b. Ookinete
BIO 475 - Parasitology Spring 2009 Stephen M. Shuster Northern Arizona University http://www4.nau.edu/isopod Lecture 10 Malaria-Life Cycle a. Micro and macrogametocytes in mosquito stomach. b. Ookinete
How to load and run an Agarose gel PSR
 How to load and run an Agarose gel PSR Agarose gel electrophoresis is the most effective way of separating DNA fragments of varying sizes ranging from100 bp to 25 kb. This protocol divided into three stages:
How to load and run an Agarose gel PSR Agarose gel electrophoresis is the most effective way of separating DNA fragments of varying sizes ranging from100 bp to 25 kb. This protocol divided into three stages:
11111L A _W ' I III! MICROCOPY RESOLUTION TEST CHART NATIONAL BUREAU OF STANDARDS 1963-A 2,1
 RD-AI?2 464 CELL PNYSIOLOOY OF THE NRARIAX PRRRSITE(U) NEN VOR 1/1 UNIV NEDICRI. CENTER N V J YANOERDERO AUG 64 DADA7-73-C-3027 UNCLSSIFIED F/0 615 NL MNNE / 4r 11111L A _W '18 2.5 11111-2 2.2I 11111125
RD-AI?2 464 CELL PNYSIOLOOY OF THE NRARIAX PRRRSITE(U) NEN VOR 1/1 UNIV NEDICRI. CENTER N V J YANOERDERO AUG 64 DADA7-73-C-3027 UNCLSSIFIED F/0 615 NL MNNE / 4r 11111L A _W '18 2.5 11111-2 2.2I 11111125
Gliding Motility Leads to Active Cellular Invasion by Cryptosporidium parvum Sporozoites
 INFECTION AND IMMUNITY, Sept. 2005, p. 5379 5387 Vol. 73, No. 9 0019-9567/05/$08.00 0 doi:10.1128/iai.73.9.5379 5387.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Gliding
INFECTION AND IMMUNITY, Sept. 2005, p. 5379 5387 Vol. 73, No. 9 0019-9567/05/$08.00 0 doi:10.1128/iai.73.9.5379 5387.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Gliding
Int.J.Curr.Microbiol.App.Sci (2018) 7(8):
 7(8):") International Journal of Current Microbiology and Applied Sciences ISSN: 2319-7706 Volume 7 Number 08 (2018) Journal homepage: http://www.ijcmas.com Original Research Article https://doi.org/10.20546/ijcmas.2018.708.378
International Journal of Current Microbiology and Applied Sciences ISSN: 2319-7706 Volume 7 Number 08 (2018) Journal homepage: http://www.ijcmas.com Original Research Article https://doi.org/10.20546/ijcmas.2018.708.378
THE ROLE OF RHOMBOID PROTEASES AND A OOCYST CAPSULE PROTEIN IN MALARIA PATHOGENESIS AND PARASITE DEVELOPMENT PRAKASH SRINIVASAN
 THE ROLE OF RHOMBOID PROTEASES AND A OOCYST CAPSULE PROTEIN IN MALARIA PATHOGENESIS AND PARASITE DEVELOPMENT BY PRAKASH SRINIVASAN Submitted in partial fulfillment of the requirements For the degree of
THE ROLE OF RHOMBOID PROTEASES AND A OOCYST CAPSULE PROTEIN IN MALARIA PATHOGENESIS AND PARASITE DEVELOPMENT BY PRAKASH SRINIVASAN Submitted in partial fulfillment of the requirements For the degree of
Review Article Immune Evasion Strategies of Pre-Erythrocytic Malaria Parasites
 Mediators of Inflammation, Article ID 362605, 6 pages http://dx.doi.org/10.1155/2014/362605 Review Article Immune Evasion Strategies of Pre-Erythrocytic Malaria Parasites Hong Zheng, Zhangping Tan, and
Mediators of Inflammation, Article ID 362605, 6 pages http://dx.doi.org/10.1155/2014/362605 Review Article Immune Evasion Strategies of Pre-Erythrocytic Malaria Parasites Hong Zheng, Zhangping Tan, and
23 Plasmodium coatneyi Eyles, Fong, Warren, Guinn, Sandosham, and Wharton, 1962
 23 Plasmodium coatneyi Eyles, Fong, Warren, Guinn, Sandosham, and Wharton, 1962 IN the course of studies on simian malaria begun by the late Dr. Don Eyles in Malaya, he and his co-workers isolated a new
23 Plasmodium coatneyi Eyles, Fong, Warren, Guinn, Sandosham, and Wharton, 1962 IN the course of studies on simian malaria begun by the late Dr. Don Eyles in Malaya, he and his co-workers isolated a new
CERTIFIED REFERENCE MATERIAL IRMM 313
 EUROPEAN COMMISSION JOINT RESEARCH CENTRE Institute for Reference Materials and Measurements (Geel) CERTIFIED REFERENCE MATERIAL IRMM 313 CERTIFICATE OF ANALYSIS PFGE AGAROSE PLUGS Certified value 2) SmaI
EUROPEAN COMMISSION JOINT RESEARCH CENTRE Institute for Reference Materials and Measurements (Geel) CERTIFIED REFERENCE MATERIAL IRMM 313 CERTIFICATE OF ANALYSIS PFGE AGAROSE PLUGS Certified value 2) SmaI
A:Malaria (Plasmodium species) Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign
 Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign") A:Malaria (Plasmodium species) Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign tertian malaria P. ovale: causes benign tertian malaria
A:Malaria (Plasmodium species) Plasmodium falciparum causes malignant tertian malaria P. malariae: causes Quartan malaria P. vivax: causes benign tertian malaria P. ovale: causes benign tertian malaria
Cattle Serologically Positive for Brucella abortus Have Antibodies
 CLINICAL AND DIAGNOSTIC LABORATORY IMMUNOLOGY, Sept. 1994, p. 506-510 Vol. 1, No. 5 1071-412X/94/$04.00+0 Copyright X) 1994, American Society for Microbiology Cattle Serologically Positive for Brucella
CLINICAL AND DIAGNOSTIC LABORATORY IMMUNOLOGY, Sept. 1994, p. 506-510 Vol. 1, No. 5 1071-412X/94/$04.00+0 Copyright X) 1994, American Society for Microbiology Cattle Serologically Positive for Brucella
Antigenic Cross-reactivity among Haemonchus contortus, Oesophagostomum columbianum and Trichuris ovis of Goat
 Iran J Parasitol Tehran University of Medical Sciences Publication http:// tums.ac.ir Open access Journal at http:// ijpa.tums.ac.ir Iranian Society of Parasitology http:// isp.tums.ac.ir Original Article
Iran J Parasitol Tehran University of Medical Sciences Publication http:// tums.ac.ir Open access Journal at http:// ijpa.tums.ac.ir Iranian Society of Parasitology http:// isp.tums.ac.ir Original Article
MRSA surveillance 2014: Poultry
 Vicky Jasson MRSA surveillance 2014: Poultry 1. Introduction In the framework of the FASFC surveillance, a surveillance of MRSA in poultry has been executed in order to determine the prevalence and diversity
Vicky Jasson MRSA surveillance 2014: Poultry 1. Introduction In the framework of the FASFC surveillance, a surveillance of MRSA in poultry has been executed in order to determine the prevalence and diversity
Plasmodium Pre-Erythrocytic Stages: Biology, Whole Parasite Vaccines and Transgenic Models
 American Journal of Immunology, 2012, 8 (3), 88-100 ISSN 1553-619X 2012 Science Publication doi:10.3844/ajisp.2012.88.100 Published Online 8 (3) 2012 (http://www.thescipub.com/aji.toc) Plasmodium Pre-Erythrocytic
American Journal of Immunology, 2012, 8 (3), 88-100 ISSN 1553-619X 2012 Science Publication doi:10.3844/ajisp.2012.88.100 Published Online 8 (3) 2012 (http://www.thescipub.com/aji.toc) Plasmodium Pre-Erythrocytic
Influence of ph on Adaptive Resistance of Pseudomonas aeruginosa to Aminoglycosides and Their Postantibiotic Effects
 ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Jan. 1996, p. 35 39 Vol. 40, No. 1 0066-4804/96/$04.00 0 Copyright 1996, American Society for Microbiology Influence of ph on Adaptive Resistance of Pseudomonas aeruginosa
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Jan. 1996, p. 35 39 Vol. 40, No. 1 0066-4804/96/$04.00 0 Copyright 1996, American Society for Microbiology Influence of ph on Adaptive Resistance of Pseudomonas aeruginosa
Automated classification of Plasmodium sporozoite movement patterns reveals a shift towards productive motility during salivary gland infection
 Automated classification of Plasmodium sporozoite movement patterns reveals a shift towards productive motility during salivary gland infection Stephan Josef Hegge, Mikhail Kudryashev, Ashley Smith, Friedrich
Automated classification of Plasmodium sporozoite movement patterns reveals a shift towards productive motility during salivary gland infection Stephan Josef Hegge, Mikhail Kudryashev, Ashley Smith, Friedrich
RICKETTSIA SPECIES AMONG TICKS IN AN AREA OF JAPAN ENDEMIC FOR JAPANESE SPOTTED FEVER
 RICKETTSIA SPECIES AMONG TICKS IN AN AREA OF JAPAN ENDEMIC FOR JAPANESE SPOTTED FEVER Makoto Kondo 1, Katsuhiko Ando 2, Keiichi Yamanaka 1 and Hitoshi Mizutani 1 1 Department of Dermatology, 2 Department
RICKETTSIA SPECIES AMONG TICKS IN AN AREA OF JAPAN ENDEMIC FOR JAPANESE SPOTTED FEVER Makoto Kondo 1, Katsuhiko Ando 2, Keiichi Yamanaka 1 and Hitoshi Mizutani 1 1 Department of Dermatology, 2 Department
Malaria parasites: virulence and transmission as a basis for intervention strategies
 Malaria parasites: virulence and transmission as a basis for intervention strategies Matthias Marti Department of Immunology and Infectious Diseases Harvard School of Public Health The global malaria burden
Malaria parasites: virulence and transmission as a basis for intervention strategies Matthias Marti Department of Immunology and Infectious Diseases Harvard School of Public Health The global malaria burden
THE SPOROZOITE ENZYME-LINKED IMMUNOSORBENT ASSAY : APPLICATION IN MALARIA EPIDEMIOLOGY
 THE SPOROZOITE ENZYME-LINKED IMMUNOSORBENT ASSAY : APPLICATION IN MALARIA EPIDEMIOLOGY Michael J. Bangs* ABSTRACT Recent biotechnological breakthroughs have led to the development of various methods for
THE SPOROZOITE ENZYME-LINKED IMMUNOSORBENT ASSAY : APPLICATION IN MALARIA EPIDEMIOLOGY Michael J. Bangs* ABSTRACT Recent biotechnological breakthroughs have led to the development of various methods for
EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS WORK-PROGRAMME PROPOSAL Version 2 VISAVET. Universidad Complutense de Madrid
 FOR BOVINE TUBERCULOSIS WORK-PROGRAMME PROPOSAL Version 2 VISAVET. Universidad Complutense de Madrid") EUROPEAN COMMISSION HEALTH & CONSUMERS DIRECTORATE-GENERAL Directorate D Animal Health and Welfare Unit D1- Animal health and Standing Committees EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS
EUROPEAN COMMISSION HEALTH & CONSUMERS DIRECTORATE-GENERAL Directorate D Animal Health and Welfare Unit D1- Animal health and Standing Committees EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and
 Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
antibody test Voller (1963) have shown both in vitro and in vivo that the third stage of Ascaris larvae of man and
 have shown both in vitro and in vivo that the third stage of Ascaris larvae of man and") J. clin. Path. (1968), 1, 449-4 The detection of circulating antibody in human toxocara infections using the indirect fluorescent antibody test B. BISSERU AND A. W. WOODRUFF From the Department of Clinical
J. clin. Path. (1968), 1, 449-4 The detection of circulating antibody in human toxocara infections using the indirect fluorescent antibody test B. BISSERU AND A. W. WOODRUFF From the Department of Clinical
Evaluation of the hair growth and retention activity of two solutions on human hair explants
 activity of two solutions on human hair explants Study Directed by Dr E. Lati of Laboratoire Bio-EC, Centre de Recherches Biologiques et d Experimentations Cutanees, on behalf of Pangaea Laboratories Ltd.
activity of two solutions on human hair explants Study Directed by Dr E. Lati of Laboratoire Bio-EC, Centre de Recherches Biologiques et d Experimentations Cutanees, on behalf of Pangaea Laboratories Ltd.
A Unique Approach to Managing the Problem of Antibiotic Resistance
 A Unique Approach to Managing the Problem of Antibiotic Resistance By: Heather Storteboom and Sung-Chul Kim Department of Civil and Environmental Engineering Colorado State University A Quick Review The
A Unique Approach to Managing the Problem of Antibiotic Resistance By: Heather Storteboom and Sung-Chul Kim Department of Civil and Environmental Engineering Colorado State University A Quick Review The
Opsonization and Phagocytosis of Plasmodium falciparum Merozoites Measured by Flow Cytometry
 CLINICAL AND DIAGNOSTIC LABORATORY IMMUNOLOGY, Jan. 2000, p. 9 13 Vol. 7, No. 1 1071-412X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Opsonization and Phagocytosis
CLINICAL AND DIAGNOSTIC LABORATORY IMMUNOLOGY, Jan. 2000, p. 9 13 Vol. 7, No. 1 1071-412X/00/$04.00 0 Copyright 2000, American Society for Microbiology. All Rights Reserved. Opsonization and Phagocytosis
Malaria remains the most important parasitic disease. Review Article
 Review Article Pre-erythrocytic Stage of Malaria Infection and the Molecular Targets Available for Interventions Dickson Adah 1,2, Yi Jun Yang 1,2, Quan Liu 1,2, Limei Qin 1, Li Qin 1, Xiaoping Chen 1
Review Article Pre-erythrocytic Stage of Malaria Infection and the Molecular Targets Available for Interventions Dickson Adah 1,2, Yi Jun Yang 1,2, Quan Liu 1,2, Limei Qin 1, Li Qin 1, Xiaoping Chen 1
POST SCREENING METHODS FOR THE DETECTION OF BETA-LACTAM RESIDUES IN PIGS.
 POST SCREENING METHODS FOR THE DETECTION OF BETA-LACTAM RESIDUES IN PIGS. Lorraine Lynas, Deborah Currie and John D.G. McEvoy. Department of Agriculture and Rural Development for Northern Ireland, Veterinary
POST SCREENING METHODS FOR THE DETECTION OF BETA-LACTAM RESIDUES IN PIGS. Lorraine Lynas, Deborah Currie and John D.G. McEvoy. Department of Agriculture and Rural Development for Northern Ireland, Veterinary
of Nebraska - Lincoln
 University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln US Army Research U.S. Department of Defense 2013 Transgenic Parasites Stably Expressing Full-Length Plasmodium falciparum
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln US Army Research U.S. Department of Defense 2013 Transgenic Parasites Stably Expressing Full-Length Plasmodium falciparum
A rapid and scalable density gradient purification method for Plasmodium sporozoites
 Kennedy et al. Malaria Journal 2012, 11:421 METHODOLOGY Open Access A rapid and scalable density gradient purification method for Plasmodium sporozoites Mark Kennedy 1, Matthew E Fishbaugher 1, Ashley
Kennedy et al. Malaria Journal 2012, 11:421 METHODOLOGY Open Access A rapid and scalable density gradient purification method for Plasmodium sporozoites Mark Kennedy 1, Matthew E Fishbaugher 1, Ashley
Effect of amikacin, cephalothin, clindamycin and vancomycin on in vitro fibroblast growth
 Research Article Genetics and Molecular Biology, 27, 3, 454-459 (2004) Copyright by the Brazilian Society of Genetics. Printed in Brazil www.sbg.org.br Effect of amikacin, cephalothin, clindamycin and
Research Article Genetics and Molecular Biology, 27, 3, 454-459 (2004) Copyright by the Brazilian Society of Genetics. Printed in Brazil www.sbg.org.br Effect of amikacin, cephalothin, clindamycin and
Venom Research at Natural Toxins Research Center (NTRC)
") Venom Research at Natural Toxins Research Center (NTRC) Dr. John C. Pérez Regents Professor and Director of the NTRC Texas A&M University-Kingsville Snake Venom Research is Important for Numerous Reasons
Venom Research at Natural Toxins Research Center (NTRC) Dr. John C. Pérez Regents Professor and Director of the NTRC Texas A&M University-Kingsville Snake Venom Research is Important for Numerous Reasons
DETECTION OF INHIBITORY SUBSTANCES IN MILK
 DETECTION OF INHIBITORY SUBSTANCES IN MILK DELVOTEST P 5 PACK/Visual & DelvoScan Reader (raw commingled cow milk, raw commingled goat milk and NCIMS accepted pasteurized cow and goat milk products) [Unless
DETECTION OF INHIBITORY SUBSTANCES IN MILK DELVOTEST P 5 PACK/Visual & DelvoScan Reader (raw commingled cow milk, raw commingled goat milk and NCIMS accepted pasteurized cow and goat milk products) [Unless
9 Parasitology 9 EXERCISE EQA. Objectives EXERCISE
 0696T_c09_81-90.qxd 07/01/2004 23:19 Page 81 EXERCISE 9 Parasitology Exercise Pre-Test Attempt to answer the following questions before starting this exercise. They will serve as a guide to important concepts.
0696T_c09_81-90.qxd 07/01/2004 23:19 Page 81 EXERCISE 9 Parasitology Exercise Pre-Test Attempt to answer the following questions before starting this exercise. They will serve as a guide to important concepts.
Downloaded from:
 Al-Khattaf, FS; Tremp, AZ; El-Houderi, A; Dessens, JT (2016) The Plasmodium alveolin IMC1a is stabilised by its terminal cysteine motifs and facilitates sporozoite morphogenesis and infectivity in a dosedependent
Al-Khattaf, FS; Tremp, AZ; El-Houderi, A; Dessens, JT (2016) The Plasmodium alveolin IMC1a is stabilised by its terminal cysteine motifs and facilitates sporozoite morphogenesis and infectivity in a dosedependent
EVALUATION OF THE SENSITIVITY AND SPECIFICITY OF THE EHRLICHIA CANIS DIAGNOSTIC TEST: Anigen Rapid E.canis Ab Test Kit
 EVALUATION OF THE SENSITIVITY AND SPECIFICITY OF THE EHRLICHIA CANIS DIAGNOSTIC TEST: Anigen Rapid E.canis Ab Test Kit FINAL REPORT Research contract (art. 83 of the L.O.U) between the Ehrlichiosis Diagnostic
EVALUATION OF THE SENSITIVITY AND SPECIFICITY OF THE EHRLICHIA CANIS DIAGNOSTIC TEST: Anigen Rapid E.canis Ab Test Kit FINAL REPORT Research contract (art. 83 of the L.O.U) between the Ehrlichiosis Diagnostic
ANNEX I SUMMARY OF PRODUCT CHARACTERISTICS
 ANNEX I SUMMARY OF PRODUCT CHARACTERISTICS 1 1. NAME OF THE VETERINARY MEDICINAL PRODUCT BLUEVAC BTV8 suspension for injection for cattle and sheep 2. QUALITATIVE AND QUANTITATIVE COMPOSITION Each ml of
ANNEX I SUMMARY OF PRODUCT CHARACTERISTICS 1 1. NAME OF THE VETERINARY MEDICINAL PRODUCT BLUEVAC BTV8 suspension for injection for cattle and sheep 2. QUALITATIVE AND QUANTITATIVE COMPOSITION Each ml of
Biology 120 Lab Exam 2 Review
 Biology 120 Lab Exam 2 Review Student Learning Services and Biology 120 Peer Mentors Sunday, November 26 th, 2017 4:00 pm Arts 263 Important note: This review was written by your Biology Peer Mentors (not
Biology 120 Lab Exam 2 Review Student Learning Services and Biology 120 Peer Mentors Sunday, November 26 th, 2017 4:00 pm Arts 263 Important note: This review was written by your Biology Peer Mentors (not
Overview. There are commonly found arrangements of bacteria based on their division. Spheres, Rods, Spirals
 Bacteria Overview Bacteria live almost everywhere. Most are microscopic ranging from 0.5 5 m in size, and unicellular. They have a variety of shapes when viewed under a microscope, most commonly: Spheres,
Bacteria Overview Bacteria live almost everywhere. Most are microscopic ranging from 0.5 5 m in size, and unicellular. They have a variety of shapes when viewed under a microscope, most commonly: Spheres,
Inactivation of Burkholderia mallei in equine serum for laboratory use.
 JCM Accepted Manuscript Posted Online 11 February 2015 J. Clin. Microbiol. doi:10.1128/jcm.03141-14 Copyright 2015, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10 11 12 13
JCM Accepted Manuscript Posted Online 11 February 2015 J. Clin. Microbiol. doi:10.1128/jcm.03141-14 Copyright 2015, American Society for Microbiology. All Rights Reserved. 1 2 3 4 5 6 7 8 9 10 11 12 13
Fluoroquinolones ELISA KIT
 Fluoroquinolones ELISA KIT Cat. No.:DEIA6883 Pkg.Size:96T Intended use The Fluoroquinolones ELISA KIT is an immunoassay for the detection of Fluoroquinolones in contaminated samples including water, fish
Fluoroquinolones ELISA KIT Cat. No.:DEIA6883 Pkg.Size:96T Intended use The Fluoroquinolones ELISA KIT is an immunoassay for the detection of Fluoroquinolones in contaminated samples including water, fish
Malaria Sporozoites Traverse Host Cells within Transient Vacuoles
 Malaria Sporozoites Traverse Host Cells within Transient Vacuoles Veronica Risco-Castillo, Selma Topçu, Carine Marinach, Giulia Manzoni, Améliee. Bigorgne, Sylvie Briquet, Xavier Baudin, Maryse Lebrun,
Malaria Sporozoites Traverse Host Cells within Transient Vacuoles Veronica Risco-Castillo, Selma Topçu, Carine Marinach, Giulia Manzoni, Améliee. Bigorgne, Sylvie Briquet, Xavier Baudin, Maryse Lebrun,