Reproductive Success of Gulls in the Larus Glaucescens-occidentalis Complex on Protection Island, Washington
|
|
|
- Wendy Martin
- 5 years ago
- Views:
Transcription
1 Andrews University Digital Andrews University Master's Theses Graduate Research 2012 Reproductive Success of Gulls in the Larus Glaucescens-occidentalis Complex on Protection Island, Washington Libby C. Megna Andrews University This research is a product of the graduate program in Biology MS at Andrews University. Find out more about the program. Follow this and additional works at: Recommended Citation Megna, Libby C., "Reproductive Success of Gulls in the Larus Glaucescens-occidentalis Complex on Protection Island, Washington" (2012). Master's Theses This Thesis is brought to you for free and open access by the Graduate Research at Digital Andrews University. It has been accepted for inclusion in Master's Theses by an authorized administrator of Digital Andrews University. For more information, please contact repository@andrews.edu.
2 Thank you for your interest in the Andrews University Digital Library of Dissertations and Theses. Please honor the copyright of this document by not duplicating or distributing additional copies in any form without the author s express written permission. Thanks for your cooperation.
3 ABSTRACT REPRODUCTIVE SUCCESS OF GULLS IN THE LARUS GLAUCESCENS- OCCIDENTALIS COMPLEX ON PROTECTION ISLAND, WASHINGTON by Libby C. Megna Co-Chairs: James L. Hayward Shandelle M. Henson
4 ABSTRACT OF GRADUATE STUDENT RESEARCH Thesis Andrews University College of Arts and Sciences Title: REPRODUCTIVE SUCCESS OF GULLS IN THE LARUS GLAUCESCENS- OCCIDENTALIS COMPLEX ON PROTECTION ISLAND, WASHINGTON Name of researcher: Libby C. Megna Name and degree of faculty co-chairs: James L. Hayward, Ph.D. Shandelle M. Henson, Ph.D. Date completed: April 2012 Glaucous-winged Gulls (Larus glaucescens) and Western Gulls (Larus occidentalis) hybridize extensively where their ranges meet along the coasts of Washington and Oregon, producing a continuum of phenotypic intergrades between the two parental species. Previous work has shown that hybrids can experience greater reproductive success than parental types by combining adaptive behaviors, such as nest site selection, of both parental species, although success of hybrids may be affected by shifts in the ecotone between marine upwelling and non-upwelling environments. I investigated whether there is a correlation between phenotype, reproductive success, and nest site choice for gulls in Protection Island National Wildlife Refuge, Washington. I examined plumage melanism and bare part coloration in the field to determine a hybrid index for each bird sampled; indices for each member of a sample pair were summed to
5 produce a pair index. Nests were monitored until eggs hatched; nest habitat was recorded as sheltered or unsheltered. Sheltered nests contained larger clutches and exhibited better hatching success but choice of nest habitat was not associated with hybrid index. Pair index was correlated with mass of the third egg of the clutch, with more Western Gulllike pairs producing smaller eggs. However, hybrid index was not significantly correlated with clutch size or hatching success. The distance of an index to the mode of the distribution of indices also was not correlated significantly with clutch size or hatching success; that is, the most abundant phenotypes on the colony were not significantly more or less successful than any other phenotypes. Keywords: habitat choice, hybridization, Larus glaucescens, Larus occidentalis, hatching success
6 Andrews University College of Arts and Sciences REPRODUCTIVE SUCCESS OF GULLS IN THE LARUS GLAUCESCENS- OCCIDENTALIS COMPLEX ON PROTECTION ISLAND, WASHINGTON A Thesis Presented in Partial Fulfillment of the Requirements for the Degree Master of Science by Libby C. Megna April 2012
7 Copyright by Libby C. Megna 2012 All Rights Reserved
8 REPRODUCTIVE SUCCESS OF GULLS IN THE LARUS GLAUCESCENS- OCCIDENTALIS COMPLEX ON PROTECTION ISLAND, WASHINGTON A thesis presented in partial fulfillment of the requirements for the degree Master of Science by Libby C. Megna APPROVAL BY THE COMMITTEE: James L. Hayward, Ph.D. Shandelle M. Henson, Ph.D. Gordon J. Atkins, Ph.D. Date approved
9 For my parents iii
10 TABLE OF CONTENTS LIST OF FIGURES... v LIST OF TABLES... vi ACKNOWLEDGMENTS... vii Chapter 1. INTRODUCTION METHODS... 8 Study Site and Nest Monitoring... 8 Assignment of Hybrid Indices Analysis RESULTS Colony Composition and Assortative Mating Nest Site Choice and Reproductive Success DISCUSSION Appendix Methodology Colony Composition and Assortative Mating Nest Site Choice and Reproductive Success Limitations Conclusions A. DETAILED NEST HABITAT CATEGORIES B. HISTOGRAMS AND NORMAL PROBABILITY PLOTS C. ORDINAL LOGISTIC REGRESSION D. RAW DATA REFERENCE LIST iv
11 LIST OF FIGURES 1. Predictions of relative fitness by three hybrid zone models Frequency distribution of hybrid indices for all individuals observed Correlation of hybrid indices between males and females of pairs (n = 169) Schematic summary of relative fitness results from studies in the Larus glaucescensoccidentalis hybrid zone v
12 LIST OF TABLES 1. Scheme for conversion of raw scores to character scores (CS) Descriptive statistics for raw character scores of all males (n = 185) and females (n = 209) observed Average character scores and hybrid indexes for males (n = 185) and females (n = 209) and p-values for differences in these scores between males and females (twotailed Mann-Whitney U) Pearson s correlation coefficients and p-values for relationships between character scores of paired males and females (n = 169) Average value of hybrid indices for sheltered and unsheltered nests and p-values for differences in indices between the site types (two-tailed Mann-Whitney U) P-values for differences in index distance to mode (IDM) between sheltered and unsheltered nest sites (two-tailed Mann-Whitney U) Average egg masses from sheltered and unsheltered nests and p-values for differences in egg mass between the nest types (two-tailed independent t-test) Average clutch size and hatching success for sheltered and unsheltered nests and p- values for differences in clutch size or hatching success between the nest types (two-tailed Mann-Whitney U) Pearson correlation coefficients and p-values for relationships between hybrid indices and egg masses Pearson correlation coefficients and p-values for relationships between index distance to mode (IDM) and egg mass Coefficients (!) and their standard errors (SE) and p-values, odds ratios (OR), and 95% confidence intervals (CI) for the ORs for ordinal logistic regression of clutch size and number hatched with hybrid indices Coefficients (!) and their standard errors (SE) and p-values, odds ratios (OR), and 95% confidence intervals (CI) for the ORs for ordinal logistic regression of clutch size and number hatched with index distance to mode (IDM) vi
13 ACKNOWLEDGMENTS This project would not have been possible without fieldwork by several dedicated Seabird Ecology Team members. Shelley McLarty, Danielle Burton, Brianna Payne, Melissa McCormick, and André Moncrieff faithfully monitored nests and made working on the island sheer joy. Melissa in particular put in many early hours at the overlook counting pigeon guillemots so that I could spend time on the colony. The hybrid indices never would have been obtained with such rigor and precision without André, a.k.a. the only other Seabirder crazy (awesome) enough to spend countless hours staring at gulls with me. Jim Hayward and Shandelle Henson kept us all well fed and happy while on the island and gave me much-needed feedback and encouragement throughout the rest of the thesis process. Jim taught me that no field method is too crazy to try at least once. Shandelle taught me how to drive the boat. Thanks are due the U.S. Fish and Wildlife Service for granting the Seabird Team the great privilege of working at Protection Island NWR. I especially thank Kevin Ryan for kindly allowing us to carry out a variety of projects all over the island. The Rosario Beach Marine Laboratory (and particularly Dave Habenicht) provided critical logistical support, including but not limited to boat towing. I also am indebted to Dr. David Cheung for kindly advising me on my statistical methods. vii
14 CHAPTER 1 INTRODUCTION Glaucous-winged Gulls (Larus glaucescens) breed along the outer coast from northern Oregon to southwest Alaska and the Aleutians and within the Salish Sea; their nonbreeding range includes the breeding range and extends south along the Baja California Peninsula (Sibley 2000; Howell and Dunn 2007). Western Gulls (Larus occidentalis) breed along the outer coast of Washington south to Baja California Sur; their nonbreeding range extends north to the Washington portion of the Salish Sea and south to the tip of the Baja California Peninsula (Sibley 2000; Howell and Dunn 2007). These two species hybridize extensively where their breeding ranges overlap along the coasts of Washington and northern Oregon (specifically, L. glaucescens hybridizes with L. o. occidentalis); this region is also an ecotone between marine upwelling and nonupwelling, fjord environments (Hoffman et al. 1978; Bell 1996, 1997; Sibley 2000; Wahl 2005). Currently there is no precise definition of what constitutes a pure Glaucouswinged or Western Gull. Glaucous-winged Gulls are described as having pale gray backs and pale gray wingtips, pale yellow to bright yellow beaks, pink orbital rings, and typically dark irises, although the irises can be very pale (Bell 1997; Howell and Dunn 2007). A classic Glaucous-winged Gull has wingtips of approximately the same shade as the back (Sibley 2000); however, birds with wingtips some shades darker than the 1
15 back also are considered to be pure Glaucous-winged Gulls (Howell and Dunn 2007). Western Gulls have slaty gray backs and black wingtips although individuals that have blackish or dark gray wingtips also are considered to be pure Western Gulls bright yellow to orange beaks, yellow or yellow-orange orbital rings, and pale irises (Bell 1997; Howell and Dunn 2007). Bell (1996) collected gulls from 33 colonies along the coasts of British Columbia, Washington, and Oregon to determine whether hybrids could be identified by their intermediate coloration. Canonical discriminant function analysis of colorimetric characters, calibrated with presumably pure birds collected from sites far from the hybrid zone and hybrids from within the hybrid zone, allowed classification of individuals as Glaucous-winged, Western, or hybrid. Indeed, hybrids possessed colorimetric characters intermediate between those of parental types, although Bell also demonstrated (via analysis of 25 enzyme systems) that hybrids are genetically more similar to Glaucouswinged Gulls. Three main models have been applied to the Larus glaucescens-occidentalis hybrid zone: (1) the dynamic-equilibrium hypothesis, in which hybridization due to dispersal balances selection against less fit hybrids, and assortative mating is adaptive (Hewitt 1988; Gay et al. 2008); (2) geographically bounded hybrid superiority, where hybrids are more fit than parental types within an ecotone between the environments to which the parental species are adapted, and a preference for hybrid mates is adaptive (Moore 1977; Good et al. 2000); and (3) selection-hybridization balance, where hybrids are favored only during some breeding seasons due to changes in the environment (Grant and Grant 1992; Bell 1997). The predictions of these models regarding relative fitness are 2
16 summarized in Figure 1. Some support has been found for each of these models; it is not yet clear which model most accurately describes the L. glaucescens-occidentalis hybrid zone. Two studies have shown that hybrid individuals experience greater reproductive success within the hybrid zone. Hoffman et al. (1978) found that hybrid pairs on Destruction Island, Washington, in 1974 were more successful at hatching chicks than pure Glaucous-winged or Western pairs. Good et al. (2000) examined the breeding behavior of gulls at two colonies within the hybrid zone: one to the north (Tatoosh Island, WA), consisting of Glaucous-winged Gulls and hybrids, and one to the south (Grays Harbor, WA), consisting of Western Gulls and hybrids. Pairs that included one or two hybrid individuals experienced greater reproductive success as measured by clutch size, egg volume, hatching success, and fledging success. Good et al. (2000) ascribed the greater success of hybrids to their ability to combine adaptive behaviors of both parent species. Hybrids in the colony of mostly Western Gulls had significantly greater reproductive success due to their choice of nest sites sheltered by vegetation, which reduced predation of eggs. Hybrids in the colony of mostly Glaucous-winged Gulls had significantly greater hatching and fledging success; Good et al. (2000) postulated that the difference in success occurred because hybrids fed predominantly on fish rather than intertidal invertebrates, as did Glaucous-winged Gulls. Good et al. advanced the hypothesis that the model of geographically bounded hybrid superiority (see Moore 1977) best describes the L. glaucescens-occidentalis hybrid zone. It should be noted, however, that the results of previous work do not all concur; it is not yet clear how far (both metaphorically and geographically speaking) results from 3
17 one colony may be extrapolated to others. Bell (1997) found that Western Gulls had larger egg volume and clutch sizes than both hybrids and Glaucous-winged Gulls at eight study sites along the coasts of Washington and Oregon, including Destruction Island, Washington. The reasons for this shift to greater Western Gull reproductive success are unknown; Bell postulated that the inconsistent results were due to a change in the width of the ecotone between marine upwelling and non-upwelling environments. Western Gulls presumably are adapted best to marine upwelling environments and so would be favored during years when upwelling is predominant within the hybrid zone; hybrids would be favored other years. Bell proposed that this selection-hybridization balance, where greatest relative fitness of pure birds and hybrids changes in time, maintains the L. glaucescens-occidentalis hybrid zone. Evidence regarding the existence of assortative mating in this gull complex is also somewhat ambiguous: although both Hoffman et al. (1978) and Bell (1997) found a strong tendency for individuals to pair with mates having a similar phenotype (p-values < 0.001), Good et al. (2000) found only weak correlations between hybrid indices of males and females at one colony (0.05 < p < 0.10) and no correlation at another (p > 0.10). Classifying L. glaucescens and L. occidentalis as separate species has been based in part on the existence of assortative mating (Hoffman et al. 1978), and understanding mate choice is necessary to understand hybridization events in birds (Good et al. 2000). This study was conducted on Protection Island in Washington, located in the Strait of Juan de Fuca. The island hosts what is believed to be the largest breeding colony of Glaucous-winged/Western Gulls in Washington (Larsen 1982), hosting 30% of the Washington breeding population (Speich and Wahl 1989), and has been the site of 4
18 numerous studies of Glaucous-winged Gull behavior and ecology (see Henson and Hayward 2010). Describing these birds in terms of the Larus glaucescens-occidentalis spectrum will be valuable for future work. Previous studies of reproductive success in this hybrid complex have been limited to outer coastal islands; it is not yet known whether hybrids also fare better in the Salish Sea and, if so, by what mechanisms they experience increased reproductive success. If hybrid superiority is bounded, it is not known exactly where the boundary lies geographically, or what factors (e.g., habitat choice) are most important in increasing hybrid success across the zone. This study adds another data point to the investigation of hybrid success across geographic space. This study also provides recent information about the degree of hybridization of Protection Island gulls, which can be compared to the data obtained by Bell (1996) in the 1980s. Finally, I test the hypothesis that the most abundant phenotype on the colony experiences the greatest egg production and/or hatching success. 5
19 Fig. 1. Predictions of relative fitness by three hybrid zone models. The horizontal axis represents the hybrid index, placing phenotypes from Glaucous-winged Gull-like (G) to Western Gull-like (W). Predictions of relative fitness are indicated by the curves. For the selection-hybridization hypothesis, the double-headed arrows indicate temporal changes in relative fitness.
20 7!
21 CHAPTER 2 METHODS Study Site and Nest Monitoring Protection Island National Wildlife Refuge (48º07 40 N, 122º55 40 W) lies within Jefferson County, Washington, at the southeastern border of the Strait of Juan de Fuca. A low-lying gravel spit, Violet Point, extends approximately 1 km from the eastern side of the island. Violet Point typically hosts 2,500-3,000 breeding pairs of gulls; the colony is most dense around the marina, located at the western end of the spit, and along the south beach of the spit (Cowles and Galusha 2012). From 27 May through 15 July 2011, five assistants and I monitored gull nests every evening (approximately PST) within six sample plots around and north of the marina. We marked nests within the plots with a numbered wooden stake on the first evening they contained an egg; we searched plots thoroughly for new nests every evening. We labeled new eggs with a letter according to their order of appearance within the nest (a typical clutch consisted of A, B, and C eggs). We weighed each egg to the nearest 0.1 gram on the day that it appeared using an Ohaus Scout Pro SP401 Portable Scale. Every day we noted whether each egg was still present and intact or whether it had disappeared from the nest, hatched, was addled, did not complete pipping, or did not survive for some other reason. When possible, we recorded the fate of eggs that 8
22 disappeared or failed to hatch (e.g., eagle predation leaves telltale eggshells, some nests failed due to flood tides); when eggs disappeared singly from nests without leaving a trace, we assumed that gulls cannibalized them. We checked clutches for pipping eggs after the earliest surviving egg of the clutch had been present for 24 days. Once all chicks of the clutch hatched, we did not monitor the nest. I defined hatching success for a nest as the number of chicks hatched successfully from the nest regardless of the clutch size. I mapped locations of all nests in the plots with a Trimble GeoExplorer 6000 series GeoXH handheld with an external Tornado antenna. Horizontal accuracy after differential correction was cm for all GPS points. At the time of mapping, I designated each nest site as sheltered or unsheltered. I considered sheltered nests to be those that were placed next to a log taller than the nest, next to the cliff of the marina, or next to or in vegetation taller than the nest, often Gumweed (Grindelia integrifolia), American Dune Grass (Leymus mollis), or Silver Bur-weed (Ambrosia chamissonis). I considered unsheltered nests to be those that were not placed next to a structure taller than the nest, typically in short or sparse vegetation or on beach/marina cobblestones (see Appendix A for a more detailed explanation of the nest site categorization). I was not able to obtain hatching success data for all nests; monitoring of late nests became too risky for chicks past 15 July, once they were several days old and mobile and thus vulnerable to chick cannibalism. These unfinished nests were followed long enough to be certain of clutch size; hence, analyses involving clutch size included both finished and unfinished nests, except for three that had complications necessitating their elimination from analysis: clutch size was probably inaccurate for one nest because 9
23 we ignored an egg found just outside of the nest; eggs in another nest were probably mislabeled, leading to doubt about the true clutch size; and I eliminated a third nest from analysis because a labeled egg from a different nest appeared in the nest. Analyses involving hatching success included only finished nests, excepting the three mentioned previously and two for which there was ambiguity in number of chicks hatched (e.g., the chick probably hatched but I was unable to find it or eggshells indicating that it hatched). Assignment of Hybrid Indices An assistant and I viewed individual gulls 3-5 m distant through a Nikon Fieldscope ED82 and classified them within the L. glaucescens-occidentalis spectrum using a hybrid index patterned after that of Bell (1997) and Good et al. (2000). The index consisted of scores for back (mantle plus scapulars) shade, shade of primary tips, bill color, orbital ring color, and iris color. The assistant and I independently scored each character and averaged the scores if after discussion we could not agree. We obtained scores from 15 June through 14 July 2011 and only when cloud cover or fog rendered lighting conditions relatively consistent and good for observation of gray shades (Howell and Dunn 2007). We began data collection no earlier than 0700 PST and terminated it when the sunlight became bright enough to cause shadows (a solar radiation of roughly 300 W/m 2 ); most data collection occurred from PST. We found that sunny conditions washed out differences in shades and made the angle at which the bird was standing to the sun impact the apparent shade. Under diffuse light this variation was much reduced, although we still made an effort to score plumage characters when the bird was broadside to us. When oriented parallel to our line of sight, feathers (especially primaries) appeared darker. We obtained character scores for individuals only when we 10
24 were positive to which territory the bird belonged, either because it was interacting with the nest or with a bird known to belong to that nest. We determined the gender of individuals by head morphology, overall size, and behavior. When uncertain, we did not make a final decision until both members of the pair were present, making the determination simple. My assistant and I scored plumage characters according to a Kodak 20-step neutral gray scale (Color Separation Guide and Gray Scale [Small]; CAT ) because it is readily available and has been used by previous authors (Hoffman et al. 1978; Howell and Dunn 2007), although Bell (1996, 1997) and Good et al. (2000) used the Munsell scale. We scored bare part colors according to a 1-5 scale similar to that of Bell (1997) and Good et al. (2000); see Table 1. When scoring the primary tips, we compared the Kodak scale to those regions of the primary tips that were the least worn and bleached. I termed these original Kodak and bare part scores raw character scores. Raw character scores were obtained for all characters of 169 pairs and an additional 16 males and 40 females, a total of 394 birds. All analyses pertaining solely to hybrid indices were carried out on this dataset. I obtained complete hybrid indices and complete nesting data (egg masses, clutch size, hatching success) for 147 pairs and an additional 13 males and 29 females. Analysis I converted the raw scores into character scores by rescaling the observed range of the raw scores equally from 1-10 (Table 1). The observed range included raw character scores from a pilot study conducted in 2010 because I wished to make the scores comparable across years. Thus, the actual range of wingtip scores for the
25 data was 7 to 19, but I used a minimum value of 5 for wingtip score because that was the minimum value observed in I created a weighted index by adding the scaled (1-10) bare part scores to twice the sum of the scaled plumage scores: Hybrid Index = 2(back) + 2(wingtips) + beak + orbital ring + iris. All analyses reported here used these weighted hybrid indices, which are hereafter referred to as hybrid indices. I refer to the weighted hybrid index for the male of a pair as the male index and the weighted hybrid index for the female of a pair as the female index. Male and female indices were summed to produce the pair index. Note that, like the indices of Hoffman et al. (1978) but unlike those of Bell (1997) and Good et al. (2000), higher values of my index indicate darker or more Western Gull-like individuals. In order to evaluate the hypothesis that the most common phenotype exhibits the greatest reproductive success, I calculated the index distance to mode (IDM) for each male, female, and pair index. For male and female hybrid indices, the index distance to mode is the absolute value of the difference between the index and the mode of the distribution of all indices. The mode of all indices was 29.5, thus: IDM = index For pair indices, the IDM was calculated by: Pair IDM = pair index 2(29.5). Larger values of IDM represent individuals that are more like pure Glaucous-winged or Western gulls. Note that since the mode was closer to the Glaucous-winged end of the hybrid index range, individuals with the largest IDMs are Western-like individuals. 12
26 I used MATLAB with Statistics Toolbox (The MathWorks, Inc., Natick, Massachusetts) to conduct ordinal logistic regression and all statistical tests except Mann- Whitney U, which I performed in SPSS (IBM, Armonk, New York). I based my decision to use a parametric or nonparametric test for a given analysis on histograms and normal probability plots of the data. All tests were carried out at the! = 0.05 significance level. I used proportional odds model ordinal logistic regression (mnrfit in MATLAB) to test whether clutch size and hatching success were correlated with IDM or hybrid index (see Catry et al for an example of ordinal logistic regression used in a similar context). Proportional odds model ordinal logistic regression yields odds ratios (ORs). An OR quantifies the change in the odds of having a greater outcome (clutch size or hatching success) resulting from a change c in the independent variable (hybrid index; Hosmer and Lemeshow 2000). I set c = 10 index units for analyses of male and female indices and IDMs. Because the range of the pair index is twice that of the individual indices, I set c = 20 index units for analyses of pair index and IDM. 13
27 Table 1. Scheme for conversion of raw scores to character scores (CS). Back Wingtips Beak Orbital Ring Iris Kodak CS Kodak CS Color CS Color CS Color CS Pale yellow Entirely pink Very dark brown Medium yellow Mostly pink Darkish brown Bright yellow Pink and yellow Medium brown Yellow-orange Mostly yellow Light yellowish Orange Entirely yellow Very pale yellowish
28 CHAPTER 3 RESULTS Colony Composition and Assortative Mating Approximately 2,400 pairs of gulls nested on Violet Point in 2011 (Joe Galusha, personal communication); thus our sample of 394 birds contained 8% of the breeding gulls on Violet Point. Putatively pure individuals of both Glaucous-winged Gulls and Western Gulls occurred on the colony. However, Glaucous-winged Gulls and/or Glaucous-winged Gull-like hybrids dominated the colony, although relatively few exhibited the classic Glaucous-winged extremely pale wingtips (Fig. 2). Table 2 lists descriptive statistics for the raw character scores of all males and females observed. Plumage character scores and indices were not significantly different between genders, but beak and orbital ring scores were significantly higher for males and iris scores were significantly higher for females (Table 3). The hybrid indices of males and females from pairs were significantly directly correlated (r = 0.35, p = ); however, when the three pairs that exhibited the highest pair index were not included, the relationship was non-significant (r = 0.10, p = 0.21; Fig. 3). Individual character scores of males and females in pairs were significantly directly correlated, except for iris scores (Table 4). 15
29 Nest Site Choice and Reproductive Success Male index, female index, and pair index were not significantly different for sheltered and unsheltered nests; however, pair index was close to significantly different (Table 5). Male, female, and pair IDMs also were not significantly different for sheltered and unsheltered nests (Table 6). A, B, and C egg masses were not significantly different across habitats (Table 7). Fourth, fifth, and sixth (D, E, and F) eggs accounted for only 1% of the data and thus were not included in analyses of egg mass. Clutch size and hatching success were significantly different across habitats, with sheltered nests having larger clutch sizes and greater hatching success (Table 8). The main driver of decreased success for unsheltered nests was egg cannibalism: 14% of finished eggs in sheltered nests were cannibalized, whereas 27% of finished eggs in unsheltered nests were cannibalized. A and B egg masses were not significantly correlated with male index, female index, or pair index; however, C egg mass was significantly negatively correlated with pair index and trended with male index and female index, although these latter two relationships were not significant (Table 9). Thus, C eggs of Western Gull-like individuals tend to be smaller than C eggs of Glaucous-winged Gull-like individuals. A, B, and C egg masses were not significantly correlated with IDM, although the correlation for C egg mass with pair IDM was close to significant (Table 10). Clutch size and hatching success were not significantly correlated with male, female, or pair hybrid index, although p-values for pair index with clutch size and female index with hatching success approached significance at the 90% confidence level with Western Gull-like individuals tending to experience greater reproductive success (Table 16
30 11). Clutch size and hatching success were not significantly correlated to male, female, or pair IDM, although the p-value for female index with hatching success approached significance at the 90% confidence level with individuals most unlike the modal phenotype tending to experience greater reproductive success (Table 12). 17
31 Table 2. Descriptive statistics for raw character scores of all males (n = 185) and females (n = 209) observed. Average SD Males Back Wingtips Beak Orbital ring Iris Females Back Wingtips Beak Orbital ring Iris
32 Table 3. Average character scores and hybrid indexes for males (n = 185) and females (n = 209) and p-values for differences in these scores between males and females (two-tailed Mann-Whitney U).! Avg " Avg p Back Wingtips Beak < Orbital ring < Iris Index
33 Table 4. Pearson s correlation coefficients and p-values for relationships between character scores of paired males and females (n = 169). r p Back < Wingtips Beak Orbital ring < Iris
34 Table 5. Average value of hybrid indices for sheltered and unsheltered nests and p-values for differences in indices between the site types (two-tailed Mann-Whitney U). Average n p Males Sheltered Unsheltered Females Sheltered Unsheltered Pairs Sheltered Unsheltered
35 Table 6. P-values for differences in index distance to mode (IDM) between sheltered and unsheltered nest sites (two-tailed Mann-Whitney U). n p Males Sheltered 126 Unsheltered Females Sheltered 125 Unsheltered Pairs Sheltered 115 Unsheltered
36 Table 7. Average egg masses from sheltered and unsheltered nests and p-values for differences in egg mass between the nest types (two-tailed independent t-test). Average (g) n p A eggs Sheltered Unsheltered B eggs Sheltered Unsheltered C eggs Sheltered Unsheltered
37 Table 8. Average clutch size and hatching success for sheltered and unsheltered nests and p-values for differences in clutch size or hatching success between the nest types (two-tailed Mann-Whitney U). Average n p Clutch size Sheltered Unsheltered Hatching success Sheltered Unsheltered <
38 Table 9. Pearson correlation coefficients and p-values for relationships between hybrid indices and egg masses. A eggs B eggs C eggs r p n r p n r p n Male index Female index Pair index
39 Table 10. Pearson correlation coefficients and p-values for relationships between index distance to mode (IDM) and egg mass. A eggs B eggs C eggs r p n r p n r p n Male IDM Female IDM Pair IDM
40 Table 11. Coefficients (!) and their standard errors (SE) and p-values, odds ratios (OR), and 95% confidence intervals (CI) for the ORs for ordinal logistic regression of clutch size and number hatched with hybrid indices.! a SE b p c OR 95% CI n Clutch size Male index Female index Pair index Number hatched Male index Female index Pair index a Reversed signs on the betas outputted from MATLAB so that OR > 1 corresponds to increasing success as the index increases. b Dispersion estimated. 27
41 Table 12. Coefficients (!) and their standard errors (SE) and p-values, odds ratios (OR), and 95% confidence intervals (CI) for the ORs for ordinal logistic regression of clutch size and number hatched with index distance to mode (IDM).! a SE b p c OR 95% CI n Clutch size!! Male IDM Female IDM Pair IDM Number hatched Male IDM Female IDM Pair IDM a Reversed signs on the betas outputted from MATLAB so that OR > 1 corresponds to increasing success as the index increases. b Dispersion estimated. 28
.")
42 Fig. 2. Frequency distribution of hybrid indices for all individuals observed (n = 394). Higher values of the index represent more Western Gull-like individuals. 29
43 75! 65! 55! Female Index! 45! 35! 25! 15! 15! 25! 35! 45! 55! 65! 75! Male Index! Fig. 3. Correlation of hybrid indices between males and females of pairs (n = 169). This relationship is significant (r = 0.35, p < ). If the three pairs denoted by triangles are removed (those with the three highest pair indices), however, the relationship becomes non-significant (r = 0.10, p = 0.21). Higher values of the index represent more Western Gull-like individuals. Linear trendline for all data is shown. 30
44 CHAPTER 4 DISCUSSION Methodology My hybrid index values are not directly comparable to those obtained in previous work due to differences in methodology and in the gray scales used; even small differences in the methodology for obtaining hybrid indices can produce a noticeable effect. For example, in a pilot study for my thesis in 2010, my assistant and I did not use the spotting scope to find the least bleached and worn areas of the wingtips; this led to artificially lighter wingtip scores (A. E. Moncrieff unpubl. data). Previous authors gave no indication that their methodologies accounted for bleaching and wear of the feathers. I believe that my methodology accurately described relative differences in phenotypic characters of gulls in the Protection Island colony. Indices were made more rigorous by use of independent assessment by two observers; furthermore, my assistant and I were able to replicate results and identify birds based on raw character scores at later dates. Previous authors (Bell 1997; Good et al. 2000) did not scale their raw character scores; instead they simply added the bare part scores, which had smaller ranges, to the plumage scores, causing the plumage characters to have greater influence on the hybrid index. Hybrid indices have been constructed this way because plumage scores are more effective discriminators of pure and hybrid individuals, although inclusion of bare part 31
45 scores enhances the discrimination (Bell 1996). My method of weighting the hybrid index is not directly equivalent to that of previous authors, but I believe it is a more reasonable way of constructing the hybrid index because it weights the scores in a controllable way. Hybrid indices spanned a continuous range. Because no work has been done to define L glaucescens, L. occidentalis, and their hybrids in terms of Kodak scores, I chose analytic methods that did not require construction of artificial categories. Thus, instead of referring to individuals as Glaucous-winged Gulls I refer to individuals as Glaucouswinged Gull-like if their indices placed them towards the low end of my hybrid index; these individuals may in actuality be either pure L. glaucescens or hybrids. Colony Composition and Assortative Mating Bell (1996) collected 17 birds from Protection Island and used canonical discriminant function analysis of colorimetric characters to determine that eight were Glaucous-winged Gulls, one was a Western Gull, and eight were hybrids. The frequency distribution of my 394 birds roughly corresponds to this: the vast majority are Glaucouswinged Gulls and/or their hybrids potentially, there are about as many Glaucouswinged Gulls as there are hybrids, depending on how one defines the categories with relatively few Western Gull-like birds (Fig. 2). As did Hoffman et al. (1978) and Bell (1997), I found a strong tendency for individuals to pair with mates having a similar phenotype; however, the statistical significance of this relationship was driven entirely by a few Western Gull-like pairs. Individuals with intermediate and Glaucous-winged Gull-like hybrid indices did not necessarily pair with individuals very like themselves, although they did typically pair 32
46 with other intermediate or Glaucous-winged Gull-like individuals. I noticed only one pair in which the individuals looked very dissimilar: an intermediate male paired with a Western Gull-like female. Nest Site Choice and Reproductive Success Mass of the third egg of a clutch is used as a measure of success, because chicks from smaller eggs are less likely to survive and size of the third egg is dependent on the energy reserves of the female; if food is abundant, the third egg can be as large as the first and second eggs of the clutch (Pierotti and Bellrose 1986). Reported egg masses for Western and Glaucous-winged Gulls are similar (Verbeek 1986; Pierotti and Annett 1995), so I assume that trends in my data are due to female condition rather than species differences. Western Gull-like individuals on Protection Island had smaller third eggs, and thus by this indicator were less successful than hybrids and Glaucous-winged Gulls. Egg mass is a relatively minor measure of fitness, however, and this trend of decreased Western Gull success did not hold for more direct measures of success, namely clutch size and hatching success. I found that sheltered nests contained larger clutch sizes and exhibited better hatching success; this has been the case for the Protection Island colony during previous years (J. L. Hayward unpubl. data). Because hybrid indices and IDMs are not different for the two site types, however, there is no evidence that any particular phenotype is capitalizing on this. In Grays Harbor, Washington, hybrids were more successful than Western Gulls because they chose vegetated (sheltered) nest sites (Good et al. 2000; see also Good 2002). In contrast, Western Gull-like individuals on Protection Island did not tend to choose unsheltered nest sites. Of the 10 pairs with the greatest hybrid indices, 33
47 seven chose sheltered nest sites; six of these were sheltered by vegetation. If the nonsignificant trends identified by the odds ratios for relationships between hybrid indices and reproductive success actually are reflective of some biological difference, it is possible that the close-to-significant (p = 0.061) difference in hybrid index for sheltered and unsheltered nest sites explains the tendency for Western Gull-like birds to have greater success. It should be noted, however, that the average pair indices for the two nest site types differ by four index units; I hesitate to assign biological significance to such a small difference. Alternatively, it is possible that Western Gull-like individuals differed in diet choice, which increased clutch size and hatching success; Good et al. (2000) found evidence for this effect among hybrids on Tatoosh Island. Diet choice is known to affect reproductive success in both Western and Glaucous-winged Gulls (Murphy et al. 1984; Annett and Pierotti 1999). Limitations It is likely that my sample of gulls was not truly random. Although I tried to obtain character scores for all birds within our six study plots, I was not able to obtain scores for all birds: wary birds flew at my approach. Thus my sample is biased towards tamer birds. There also was bias against certain nest site types it was harder to obtain character scores for birds nesting in tall dune grass (Leymus mollis) because (1) they could not be observed while sitting on the nest and (2) once they were displaced from the nest they tended to be wary of approach. Furthermore, 57% of the nests for which I obtained character scores for one or both members of the pair occurred in two of six study plots. A six-year study of reproductive success on Protection Island has demonstrated that different plots, or sub-regions of the colony, experience different 34
48 pressures (e.g., amount of eagle predation or egg cannibalism) and have differential hatching success (J. L. Hayward unpubl. data). Because the gulls were not banded, I had to include only those that were actively nesting so I could identify individuals based on which nest they tended. I did not begin collecting character scores until most birds on the colony were well into egg-laying or beginning incubation, so early nesters that abandoned before my study began were not included in the sample. Thus, it is very possible that my sample is biased towards more successful gulls and did not capture the entire range of success outcomes experienced by the birds. I was unable to follow chicks through to fledging and thus cannot comment on whether the gulls might experience differential fledging success. Following chicks for the first week or two after hatching could yield interesting results, given that many chicks die in the first week or two when they are small enough to be easily grabbed by neighboring adult gulls. It is likely that chicks from unsheltered nests experience greater mortality in this manner than those from sheltered nests. It should be noted, however, that I found no strong correlation between hybrid index and habitat choice; thus, obtaining data on chick survival may not settle the question of hybrid superiority. Indeed, Good et al. (2000) found that when hybrids experienced greater hatching success, they also experienced greater fledging success. Conclusions I found no evidence of differential habitat choice according to phenotype, and reproductive success was not significantly greater for the most common phenotype on the colony or for pure Glaucous-winged or pure Western individuals. This result does not support the dynamic-equilibrium hypothesis, which postulates that hybrids always 35
49 experience the least success. It also does not support the bounded hybrid superiority hypothesis, which predicts that (1) if the colony is within the zone of hybrid superiority, hybrids experience the greatest success, and that (2) north of the hybrid zone, Glaucouswinged Gulls are most successful. It is possible that Protection Island is on or near the boundary between hybrid superiority and Glaucous-winged Gull superiority, and thus neither hybrids nor Glaucous-winged Gulls were most successful. This explanation fails to account, however, for the fact that Western Gull-like individuals experienced the same relative success as intermediates and Glaucous-winged Gull-like birds: There are no apparent fitness consequences for Western Gulls at the northern end of the hybrid zone. It is also possible that a selection-hybridization balance is operating; I do not have rigorous data for multiple years for the Protection Island colony, and so I cannot comment upon whether success varies for phenotypes across years. (However, limited data from a 2010 pilot study also showed no significant relationship between hybrid index and clutch size or hatching success.) It is known that Protection Island gulls experience decreased success during El Niño years (J. L. Hayward unpubl. data); determining whether success differs according to hybrid index under those conditions could be informative. Evidence to date regarding relative fitness of hybrids and parental types within the zone are summarized in Figure 4. Without data from multiple years for several colonies across the zone, it is unclear which model of the hybrid zone is most accurate. 36
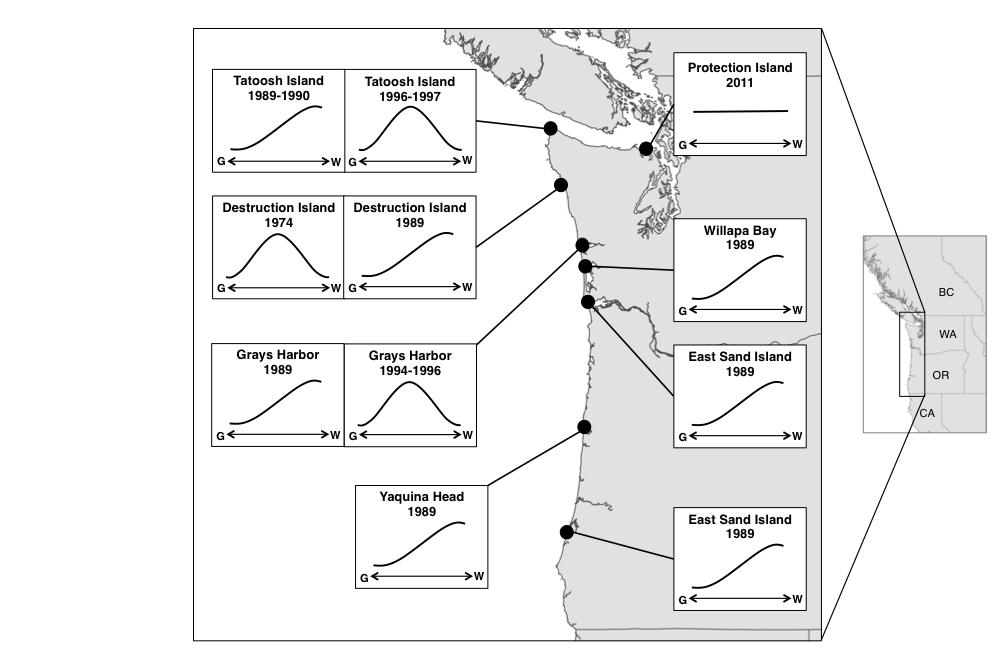
50 Fig. 4. Schematic summary of relative fitness results from studies in the Larus glaucescens-occidentalis hybrid zone. The horizontal axis represents the hybrid index, placing phenotypes from Glaucous-winged Gull-like (G) to Western Gull-like (W). Relative fitness results are indicated by the curves. The 1974 Destruction Island study was conducted by Hoffman et al. (1978), who determined that hybrids experienced greatest hatching success. The 1989 and 1990 studies were conducted by Bell (1997), who determined that Western Gulls had the smallest egg volume differences (largest relative C egg volume) at eight sites (two in Grays Harbor) in Washington and Oregon. The studies on Tatoosh Island and in Grays Harbor were conducted by Good et al. (2000), who determined that hybrids experienced greatest hatching and fledging success. Map credit: Esri, DeLorme Publishing Company, Inc.
51 38
52 APPENDIX A DETAILED NEST HABITAT CATEGORIES
53 Table 13. Acronyms for and descriptions of original nest site categories. Acronym Description Sheltered BS Beside shrub; often Gumweed (Grindelia integrifolia), Silver Bur-weed (Ambrosia chamissonis) or tall clump of (non-dune) grass CC LB LN TE TT On cobbles up against the gravel bank of the marina Next to a log (or logs) on the beach Next to a log (or logs) away from the beach At the edge of Dune Grass (Leymus mollis) At the end of a tunnel into Dune Grass Unsheltered CN SV On cobbles of the beach or marina but not up against a gravel bank In short vegetation, i.e. vegetation that was not much taller than the nest rim and didn t provide cover for the nest 40
54 APPENDIX B HISTOGRAMS AND NORMAL PROBABILITY PLOTS
55 A B Fig. 5. Histogram and normal probability plot for male index data (n = 185). I determined that male index data were not normally distributed by examining (A) a histogram and (B) a normal probability plot of the data. Normally distributed data should be symmetric about the mode in a histogram and should fall along the dashed line in a normal probability plot. 42
56 A B Fig. 6. Histogram and normal probability plot for female index data (n = 209). I determined that female index data were not normally distributed by examining (A) a histogram and (B) a normal probability plot of the data. Normally distributed data should be symmetrical about the mode in a histogram and should fall along the dashed line in a normal probability plot. 43
57 Fig. 7. Normal probability plots for A, B, and C egg masses (n = 225, 220, and 174, respectively). I determined that the (A) A egg masses, (B) B egg masses, and (C) C egg masses were normally distributed by examining normal probability plots. Normally distributed data should fall along the dashed line.
58 A B 45 C
 clutch size and (B) hatching success data were not normally distributed by examining histograms of the data.")
59 A B Fig. 8. Histograms for clutch size (n = 228) and hatching success (n = 194). I determined that (A) clutch size and (B) hatching success data were not normally distributed by examining histograms of the data. Normally distributed data should be symmetric around the mode. 46
60 APPENDIX C ORDINAL LOGISTIC REGRESSION
61 Logistic regression is employed for binary outcomes (0 or 1). In contrast, ordinal logistic regression admits multiple, rank-ordered outcomes. My data consisted of ordered reproductive success outcomes (i.e., clutch size or number of chicks hatched) that I wished to regress on hybrid index or index distance to mode (IDM). Following the notation of Hosmer and Lemeshow (2000), consider a system with K + 1 ordered outcomes denoted 0, 1, 2,, K. Let! k (x) denote the probability that outcome Y is equal to k given the hybrid index (or IDM) x. The log of the proportional odds, which compares the probability of an equal or smaller response to the probability of a larger response, is regressed on the hybrid index (or index distance) x: " P(Y! k x) % " ln$ ' = ln! (x)+! (x)+!+! (x) % 0 1 k $ ' = " 0 ( " 1 x, # P(Y > k x) & #! k+1 (x)+! k+2 (x)+!! K (x)& or equivalently, " P(Y > k x) % ln$ ' = (! 0 +! 1 x. # P(Y! k x) & (MATLAB estimates of " 0 and " 1 are based on the former equation, but I changed the signs of the estimated " 0 and " 1 in Tables 11 and 12 because the latter formulation is easier to interpret in my context.) The likelihood function is l(! 0,! 1 ) = n & i=1 "" 0 (x i ) z 0i " (x 1 i )z 1i!!!! K (x i ) z Ki # $ % (Hosmer and Lemeshow 2000) and the log-likelihood is n L(! 0,! 1 ) = %!" z 0i ln [! 0 (x i )] + z 1i ln [! 1 (x i )] +!+ z Ki ln [! K (x i )] # $, i=1 where z ji = 1 when y = j and z ji = 0 otherwise. I maximized L(" 0," 1 ) numerically using the mnrfit function, adjusted for overdispersion, in MATLAB. Overdispersion occurs when 48
Ovulation Synchrony as an Adaptive Response to Egg Cannibalism in a Seabird Colony
 Andrews University Digital Commons @ Andrews University Honors Theses Undergraduate Research 2015 Ovulation Synchrony as an Adaptive Response to Egg Cannibalism in a Seabird Colony Sumiko Weir This research
Andrews University Digital Commons @ Andrews University Honors Theses Undergraduate Research 2015 Ovulation Synchrony as an Adaptive Response to Egg Cannibalism in a Seabird Colony Sumiko Weir This research
INTERBREEDING OF GLAUCOUS-WINGED AND HERRING GULLS IN THE COOK INLET REGION, ALASKA. By FRANCIS S. L. WILLIAMSON and LEONARD J.
 24 Vol. 65 INTERBREEDING OF GLAUCOUS-WINGED AND HERRING GULLS IN THE COOK INLET REGION, ALASKA By FRANCIS S. L. WILLIAMSON and LEONARD J. PEYTON In the course of field studies of birds about the Cook Inlet
24 Vol. 65 INTERBREEDING OF GLAUCOUS-WINGED AND HERRING GULLS IN THE COOK INLET REGION, ALASKA By FRANCIS S. L. WILLIAMSON and LEONARD J. PEYTON In the course of field studies of birds about the Cook Inlet
The Behavior of Glaucous-winged Gull Egg Cannibals
 Andrews University Digital Commons @ Andrews University Honors Theses Undergraduate Research 2015 The Behavior of Glaucous-winged Gull Egg Cannibals Ashley A. Reichert This research is a product of the
Andrews University Digital Commons @ Andrews University Honors Theses Undergraduate Research 2015 The Behavior of Glaucous-winged Gull Egg Cannibals Ashley A. Reichert This research is a product of the
PROGRESS REPORT for COOPERATIVE BOBCAT RESEARCH PROJECT. Period Covered: 1 April 30 June Prepared by
 PROGRESS REPORT for COOPERATIVE BOBCAT RESEARCH PROJECT Period Covered: 1 April 30 June 2014 Prepared by John A. Litvaitis, Tyler Mahard, Rory Carroll, and Marian K. Litvaitis Department of Natural Resources
PROGRESS REPORT for COOPERATIVE BOBCAT RESEARCH PROJECT Period Covered: 1 April 30 June 2014 Prepared by John A. Litvaitis, Tyler Mahard, Rory Carroll, and Marian K. Litvaitis Department of Natural Resources
University of Canberra. This thesis is available in print format from the University of Canberra Library.
 University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
ABSTRACT. Ashmore Reef
 ABSTRACT The life cycle of sea turtles is complex and is not yet fully understood. For most species, it involves at least three habitats: the pelagic, the demersal foraging and the nesting habitats. This
ABSTRACT The life cycle of sea turtles is complex and is not yet fully understood. For most species, it involves at least three habitats: the pelagic, the demersal foraging and the nesting habitats. This
Dominance/Suppression Competitive Relationships in Loblolly Pine (Pinus taeda L.) Plantations
 Plantations") Dominance/Suppression Competitive Relationships in Loblolly Pine (Pinus taeda L.) Plantations by Michael E. Dyer Dissertation submitted to the Faculty of the Virginia Polytechnic Institute and Stand University
Dominance/Suppression Competitive Relationships in Loblolly Pine (Pinus taeda L.) Plantations by Michael E. Dyer Dissertation submitted to the Faculty of the Virginia Polytechnic Institute and Stand University
Bald Head Island Conservancy 2018 Sea Turtle Report Emily Goetz, Coastal Scientist
 Bald Head Island Conservancy 2018 Sea Turtle Report Emily Goetz, Coastal Scientist Program Overview The Bald Head Island Conservancy s (BHIC) Sea Turtle Protection Program (STPP) began in 1983 with the
Bald Head Island Conservancy 2018 Sea Turtle Report Emily Goetz, Coastal Scientist Program Overview The Bald Head Island Conservancy s (BHIC) Sea Turtle Protection Program (STPP) began in 1983 with the
Section: 101 (2pm-3pm) 102 (3pm-4pm)
 102 (3pm-4pm)") Stat 20 Midterm Exam Instructor: Tessa Childers-Day 12 July 2012 Please write your name and student ID below, and circle your section With your signature, you certify that you have not observed poor or
Stat 20 Midterm Exam Instructor: Tessa Childers-Day 12 July 2012 Please write your name and student ID below, and circle your section With your signature, you certify that you have not observed poor or
Reduced availability of refuse and breeding output in a herring gull (Larus argentatus) colony
 colony") Ann. Zool. Fennici 35: 37 42 ISSN 0003-455X Helsinki 4 June 1998 Finnish Zoological and Botanical Publishing Board 1998 Reduced availability of refuse and breeding output in a herring gull (Larus argentatus)
Ann. Zool. Fennici 35: 37 42 ISSN 0003-455X Helsinki 4 June 1998 Finnish Zoological and Botanical Publishing Board 1998 Reduced availability of refuse and breeding output in a herring gull (Larus argentatus)
Kodiak National Wildlife Refuge 2004 Bald Eagle Nesting and Productivity Survey
 Kodiak National Wildlife Refuge 2004 Bald Eagle Nesting and Productivity Survey ANNUAL REPORT by Denny Zwiefelhofer Key Words: Bald Eagle Nesting Productivity Kodiak Island Kodiak National Wildlife Refuge
Kodiak National Wildlife Refuge 2004 Bald Eagle Nesting and Productivity Survey ANNUAL REPORT by Denny Zwiefelhofer Key Words: Bald Eagle Nesting Productivity Kodiak Island Kodiak National Wildlife Refuge
A final programmatic report to: SAVE THE TIGER FUND. Scent Dog Monitoring of Amur Tigers-V ( ) March 1, March 1, 2006
 March 1, March 1, 2006") 1 A final programmatic report to: SAVE THE TIGER FUND Scent Dog Monitoring of Amur Tigers-V (2005-0013-017) March 1, 2005 - March 1, 2006 Linda Kerley and Galina Salkina PROJECT SUMMARY We used scent-matching
1 A final programmatic report to: SAVE THE TIGER FUND Scent Dog Monitoring of Amur Tigers-V (2005-0013-017) March 1, 2005 - March 1, 2006 Linda Kerley and Galina Salkina PROJECT SUMMARY We used scent-matching
AGE AT FIRST BREEDING AND CHANGE IN PLUMAGE OF KELP GULLS LARUS DOMINICANUS IN SOUTH AFRICA. R. J. M. CRAWFORD*, B. M. DYER* and L.
 S. Afr. J. mar. Sci. 22: 27 32 2000 27 AGE AT FIRST BREEDING AND CHANGE IN PLUMAGE OF KELP GULLS LARUS DOMINICANUS IN SOUTH AFRICA R. J. M. CRAWFORD*, B. M. DYER* and L. UPFOLD* In South Africa, kelp gulls
S. Afr. J. mar. Sci. 22: 27 32 2000 27 AGE AT FIRST BREEDING AND CHANGE IN PLUMAGE OF KELP GULLS LARUS DOMINICANUS IN SOUTH AFRICA R. J. M. CRAWFORD*, B. M. DYER* and L. UPFOLD* In South Africa, kelp gulls
Response to SERO sea turtle density analysis from 2007 aerial surveys of the eastern Gulf of Mexico: June 9, 2009
 Response to SERO sea turtle density analysis from 27 aerial surveys of the eastern Gulf of Mexico: June 9, 29 Lance P. Garrison Protected Species and Biodiversity Division Southeast Fisheries Science Center
Response to SERO sea turtle density analysis from 27 aerial surveys of the eastern Gulf of Mexico: June 9, 29 Lance P. Garrison Protected Species and Biodiversity Division Southeast Fisheries Science Center
Subdomain Entry Vocabulary Modules Evaluation
 Subdomain Entry Vocabulary Modules Evaluation Technical Report Vivien Petras August 11, 2000 Abstract: Subdomain entry vocabulary modules represent a way to provide a more specialized retrieval vocabulary
Subdomain Entry Vocabulary Modules Evaluation Technical Report Vivien Petras August 11, 2000 Abstract: Subdomain entry vocabulary modules represent a way to provide a more specialized retrieval vocabulary
STAT170 Exam Preparation Workshop Semester
 Study Information STAT Exam Preparation Workshop Semester Our sample is a randomly selected group of American adults. They were measured on a number of physical characteristics (some measurements were
Study Information STAT Exam Preparation Workshop Semester Our sample is a randomly selected group of American adults. They were measured on a number of physical characteristics (some measurements were
BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS
 Nov., 1965 505 BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS Lack ( 1954; 40-41) has pointed out that in species of birds which have asynchronous hatching, brood size may be adjusted
Nov., 1965 505 BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS Lack ( 1954; 40-41) has pointed out that in species of birds which have asynchronous hatching, brood size may be adjusted
Factors Influencing Egg Production
 June, 1930 Research Bulletin No. 129 Factors Influencing Egg Production II. The Influence of the Date of First Egg Upon Maturity and Production By C. W. KNOX AGRICULTURAL EXPERIMENT STATION IOWA STATE
June, 1930 Research Bulletin No. 129 Factors Influencing Egg Production II. The Influence of the Date of First Egg Upon Maturity and Production By C. W. KNOX AGRICULTURAL EXPERIMENT STATION IOWA STATE
Tree Swallows (Tachycineta bicolor) are breeding earlier at Creamer s Field Migratory Waterfowl Refuge, Fairbanks, AK
 are breeding earlier at Creamer s Field Migratory Waterfowl Refuge, Fairbanks, AK") Tree Swallows (Tachycineta bicolor) are breeding earlier at Creamer s Field Migratory Waterfowl Refuge, Fairbanks, AK Abstract: We examined the average annual lay, hatch, and fledge dates of tree swallows
Tree Swallows (Tachycineta bicolor) are breeding earlier at Creamer s Field Migratory Waterfowl Refuge, Fairbanks, AK Abstract: We examined the average annual lay, hatch, and fledge dates of tree swallows
Genetics and Probability
 Genetics and Probability Genetics and Probability The likelihood that a particular event will occur is called probability. The principles of probability can be used to predict the outcomes of genetic crosses.
Genetics and Probability Genetics and Probability The likelihood that a particular event will occur is called probability. The principles of probability can be used to predict the outcomes of genetic crosses.
Western Snowy Plover Recovery and Habitat Restoration at Eden Landing Ecological Reserve
 Western Snowy Plover Recovery and Habitat Restoration at Eden Landing Ecological Reserve Prepared by: Benjamin Pearl, Plover Program Director Yiwei Wang, Executive Director Anqi Chen, Plover Biologist
Western Snowy Plover Recovery and Habitat Restoration at Eden Landing Ecological Reserve Prepared by: Benjamin Pearl, Plover Program Director Yiwei Wang, Executive Director Anqi Chen, Plover Biologist
Research Summary: Evaluation of Northern Bobwhite and Scaled Quail in Western Oklahoma
 P-1054 Research Summary: Evaluation of Northern Bobwhite and Scaled Quail in Western Oklahoma Oklahoma Agricultural Experiment Station Division of Agricultural Sciences and Natural Resources Oklahoma State
P-1054 Research Summary: Evaluation of Northern Bobwhite and Scaled Quail in Western Oklahoma Oklahoma Agricultural Experiment Station Division of Agricultural Sciences and Natural Resources Oklahoma State
9-2 Probability and Punnett. Squares Probability and Punnett Squares. Slide 1 of 21. Copyright Pearson Prentice Hall
 9-2 Probability and Punnett 11-2 Probability and Punnett Squares Squares 1 of 21 11-2 Probability and Punnett Squares Genetics and Probability How do geneticists use the principles of probability? 2 of
9-2 Probability and Punnett 11-2 Probability and Punnett Squares Squares 1 of 21 11-2 Probability and Punnett Squares Genetics and Probability How do geneticists use the principles of probability? 2 of
Factors Influencing Local Recruitment in Tree Swallows, Tachycineta bicolor
 Grand Valley State University ScholarWorks@GVSU Honors Projects Undergraduate Research and Creative Practice 2013 Factors Influencing Local Recruitment in Tree Swallows, Tachycineta bicolor Danielle M.
Grand Valley State University ScholarWorks@GVSU Honors Projects Undergraduate Research and Creative Practice 2013 Factors Influencing Local Recruitment in Tree Swallows, Tachycineta bicolor Danielle M.
Summary Report of the Anatolian Shepherd Dog Health Survey. Data collected by ASDCA in partnership with OFA from December 1, 2009 to September 5, 2011
 Data collected by ASDCA in partnership with OFA from December 1, 2009 to September 5, 2011 Report Authors: Jessica Voss, DVM, MRCVS, ASDCA Health Coordinator Robert Owen, Ph.D. May 31, 2012 General Data:
Data collected by ASDCA in partnership with OFA from December 1, 2009 to September 5, 2011 Report Authors: Jessica Voss, DVM, MRCVS, ASDCA Health Coordinator Robert Owen, Ph.D. May 31, 2012 General Data:
Red-Tailed Hawk Buteo jamaicensis
 Red-Tailed Hawk Buteo jamaicensis This large, dark headed, broad-shouldered hawk is one of the most common and widespread hawks in North America. The Red-tailed hawk belongs to the genus (family) Buteo,
Red-Tailed Hawk Buteo jamaicensis This large, dark headed, broad-shouldered hawk is one of the most common and widespread hawks in North America. The Red-tailed hawk belongs to the genus (family) Buteo,
Adjustment Factors in NSIP 1
 Adjustment Factors in NSIP 1 David Notter and Daniel Brown Summary Multiplicative adjustment factors for effects of type of birth and rearing on weaning and postweaning lamb weights were systematically
Adjustment Factors in NSIP 1 David Notter and Daniel Brown Summary Multiplicative adjustment factors for effects of type of birth and rearing on weaning and postweaning lamb weights were systematically
Result Demonstration Report
 Result Demonstration Report Texas Quail Index Texas A&M AgriLife Extension Service Garza County Cooperator: Chimney Creek Ranch; Danny Robertson, Mgr Greg Jones, County Extension Agent-Ag for Garza County
Result Demonstration Report Texas Quail Index Texas A&M AgriLife Extension Service Garza County Cooperator: Chimney Creek Ranch; Danny Robertson, Mgr Greg Jones, County Extension Agent-Ag for Garza County
Bald Eagles in the Yukon. Wildlife in our backyard
 Bald Eagles in the Yukon Wildlife in our backyard The Bald Eagle at a glance Both male and female adult Bald Eagles have a dark brown body and wings with a white head, neck and tail. They have a yellow
Bald Eagles in the Yukon Wildlife in our backyard The Bald Eagle at a glance Both male and female adult Bald Eagles have a dark brown body and wings with a white head, neck and tail. They have a yellow
Result Demonstration Report
 Result Demonstration Report 2014 Texas Quail Index Texas A&M AgriLife Extension Service Archer County Cooperator: Brad Mitchell- Mitchell and Parkey Ranches Justin B Gilliam, County Extension Agent for
Result Demonstration Report 2014 Texas Quail Index Texas A&M AgriLife Extension Service Archer County Cooperator: Brad Mitchell- Mitchell and Parkey Ranches Justin B Gilliam, County Extension Agent for
Parameter: Productivity (black-legged and red-legged kittiwakes); populations (marine mammals)
; populations (marine mammals)") Wildlife Inventory Plan Alaska Maritime National Wildlife Refuge Protocol #27 Version 1.2 Parameter: Productivity (black-legged and red-legged kittiwakes); populations (marine mammals) Species: Black-legged
Wildlife Inventory Plan Alaska Maritime National Wildlife Refuge Protocol #27 Version 1.2 Parameter: Productivity (black-legged and red-legged kittiwakes); populations (marine mammals) Species: Black-legged
Longevity of the Australian Cattle Dog: Results of a 100-Dog Survey
 Longevity of the Australian Cattle Dog: Results of a 100-Dog Survey Pascal Lee, Ph.D. Owner of Ping Pong, an Australian Cattle Dog Santa Clara, CA, USA. E-mail: pascal.lee@yahoo.com Abstract There is anecdotal
Longevity of the Australian Cattle Dog: Results of a 100-Dog Survey Pascal Lee, Ph.D. Owner of Ping Pong, an Australian Cattle Dog Santa Clara, CA, USA. E-mail: pascal.lee@yahoo.com Abstract There is anecdotal
A SPATIAL ANALYSIS OF SEA TURTLE AND HUMAN INTERACTION IN KAHALU U BAY, HI. By Nathan D. Stewart
 A SPATIAL ANALYSIS OF SEA TURTLE AND HUMAN INTERACTION IN KAHALU U BAY, HI By Nathan D. Stewart USC/SSCI 586 Spring 2015 1. INTRODUCTION Currently, sea turtles are an endangered species. This project looks
A SPATIAL ANALYSIS OF SEA TURTLE AND HUMAN INTERACTION IN KAHALU U BAY, HI By Nathan D. Stewart USC/SSCI 586 Spring 2015 1. INTRODUCTION Currently, sea turtles are an endangered species. This project looks
Summary of 2017 Field Season
 Summary of 2017 Field Season Figure 1. The 2017 crew: L to R, Mark Baran, Collette Lauzau, Mark Dodds A stable and abundant food source throughout the chick provisioning period allowed for a successful
Summary of 2017 Field Season Figure 1. The 2017 crew: L to R, Mark Baran, Collette Lauzau, Mark Dodds A stable and abundant food source throughout the chick provisioning period allowed for a successful
SHEEP SIRE REFERENCING SCHEMES - NEW OPPORTUNITIES FOR PEDIGREE BREEDERS AND LAMB PRODUCERS a. G. Simm and N.R. Wray
 SHEEP SIRE REFERENCING SCHEMES - NEW OPPORTUNITIES FOR PEDIGREE BREEDERS AND LAMB PRODUCERS a G. Simm and N.R. Wray The Scottish Agricultural College Edinburgh, Scotland Summary Sire referencing schemes
SHEEP SIRE REFERENCING SCHEMES - NEW OPPORTUNITIES FOR PEDIGREE BREEDERS AND LAMB PRODUCERS a G. Simm and N.R. Wray The Scottish Agricultural College Edinburgh, Scotland Summary Sire referencing schemes
EXERCISE 14 Marine Birds at Sea World Name
 EXERCISE 14 Marine Birds at Sea World Name Section Polar and Equatorial Penguins Penguins Penguins are flightless birds that are mainly concentrated in the Southern Hemisphere. They were first discovered
EXERCISE 14 Marine Birds at Sea World Name Section Polar and Equatorial Penguins Penguins Penguins are flightless birds that are mainly concentrated in the Southern Hemisphere. They were first discovered
PROBABLE NON-BREEDERS AMONG FEMALE BLUE GROUSE
 Condor, 81:78-82 0 The Cooper Ornithological Society 1979 PROBABLE NON-BREEDERS AMONG FEMALE BLUE GROUSE SUSAN J. HANNON AND FRED C. ZWICKEL Parallel studies on increasing (Zwickel 1972) and decreasing
Condor, 81:78-82 0 The Cooper Ornithological Society 1979 PROBABLE NON-BREEDERS AMONG FEMALE BLUE GROUSE SUSAN J. HANNON AND FRED C. ZWICKEL Parallel studies on increasing (Zwickel 1972) and decreasing
Variation of Chicken Embryo Development by Temperature Influence. Anna Morgan Miller. Rockdale Magnet School for Science and Technology
 Variation of Chicken Embryo Development by Temperature Influence Anna Morgan Miller Rockdale Magnet School for Science and Technology Anna Morgan Miller Rockdale Magnet School 1174 Bulldog Circle Conyers,
Variation of Chicken Embryo Development by Temperature Influence Anna Morgan Miller Rockdale Magnet School for Science and Technology Anna Morgan Miller Rockdale Magnet School 1174 Bulldog Circle Conyers,
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/314/5802/1111/dc1 Supporting Online Material for Rapid Temporal Reversal in Predator-Driven Natural Selection Jonathan B. Losos,* Thomas W. Schoener, R. Brian Langerhans,
www.sciencemag.org/cgi/content/full/314/5802/1111/dc1 Supporting Online Material for Rapid Temporal Reversal in Predator-Driven Natural Selection Jonathan B. Losos,* Thomas W. Schoener, R. Brian Langerhans,
Lab 6: Energizer Turtles
 Lab 6: Energizer Turtles Screen capture showing the required components: 4 Sliders (as shown) 2 Buttons (as shown) 4 Monitors (as shown) min-pxcor = -50, max-pxcor = 50, min-pycor = -50, max-pycor = 50
Lab 6: Energizer Turtles Screen capture showing the required components: 4 Sliders (as shown) 2 Buttons (as shown) 4 Monitors (as shown) min-pxcor = -50, max-pxcor = 50, min-pycor = -50, max-pycor = 50
Coastal Birds of Haida Heritage Sites and Important Bird Areas.
 Coastal Birds of Haida Heritage Sites and Important Bird Areas www.ibacanada.ca Taadll Skaa anda Pacific Loon Photo : Tim Bowman, USFWS pale grey head, white vertical lines on neck, when in breeding plumage
Coastal Birds of Haida Heritage Sites and Important Bird Areas www.ibacanada.ca Taadll Skaa anda Pacific Loon Photo : Tim Bowman, USFWS pale grey head, white vertical lines on neck, when in breeding plumage
BIOL4. General Certificate of Education Advanced Level Examination June Unit 4 Populations and environment. Monday 13 June pm to 3.
 Centre Number Surname Candidate Number For Examiner s Use Other Names Candidate Signature Examiner s Initials General Certificate of Education Advanced Level Examination June 2011 Question 1 2 Mark Biology
Centre Number Surname Candidate Number For Examiner s Use Other Names Candidate Signature Examiner s Initials General Certificate of Education Advanced Level Examination June 2011 Question 1 2 Mark Biology
BLACK OYSTERCATCHER NEST MONITORING PROTOCOL
 BLACK OYSTERCATCHER NEST MONITORING PROTOCOL In addition to the mid-late May population survey (see Black Oystercatcher abundance survey protocol) we will attempt to continue monitoring at least 25 nests
BLACK OYSTERCATCHER NEST MONITORING PROTOCOL In addition to the mid-late May population survey (see Black Oystercatcher abundance survey protocol) we will attempt to continue monitoring at least 25 nests
Tristan Darwin Project. Monitoring Guide. A Guide to Monitoring Albatross, Penguin and Seal Plots on Tristan and Nightingale
 Tristan Darwin Project Monitoring Guide A Guide to Monitoring Albatross, Penguin and Seal Plots on Tristan and Nightingale Atlantic Yellow-nosed albatross Biology The yellow-nosed albatross or molly lays
Tristan Darwin Project Monitoring Guide A Guide to Monitoring Albatross, Penguin and Seal Plots on Tristan and Nightingale Atlantic Yellow-nosed albatross Biology The yellow-nosed albatross or molly lays
Ames, IA Ames, IA (515)
") BENEFITS OF A CONSERVATION BUFFER-BASED CONSERVATION MANAGEMENT SYSTEM FOR NORTHERN BOBWHITE AND GRASSLAND SONGBIRDS IN AN INTENSIVE PRODUCTION AGRICULTURAL LANDSCAPE IN THE LOWER MISSISSIPPI ALLUVIAL
BENEFITS OF A CONSERVATION BUFFER-BASED CONSERVATION MANAGEMENT SYSTEM FOR NORTHERN BOBWHITE AND GRASSLAND SONGBIRDS IN AN INTENSIVE PRODUCTION AGRICULTURAL LANDSCAPE IN THE LOWER MISSISSIPPI ALLUVIAL
GULLS (LARUS ARGENTATUS)
") TERRITORY SIZE DIFFERENCES IN RELATION TO REPRODUCTIVE STAGE AND TYPE OF INTRUDER IN HERRING GULLS (LARUS ARGENTATUS) JOANNA BURGER Department of Biology, Livingston College, Rutgers University, New Brunswick,
TERRITORY SIZE DIFFERENCES IN RELATION TO REPRODUCTIVE STAGE AND TYPE OF INTRUDER IN HERRING GULLS (LARUS ARGENTATUS) JOANNA BURGER Department of Biology, Livingston College, Rutgers University, New Brunswick,
Did you know that Snowy Plovers (Charadrius alexandrines char-ad-ree-us alex-an-dreen-us):
:") Did you know that Snowy Plovers (Charadrius alexandrines char-ad-ree-us alex-an-dreen-us): 2 - are listed as a threatened species in the state of Florida? As of 2006, Florida had only an estimated 225
Did you know that Snowy Plovers (Charadrius alexandrines char-ad-ree-us alex-an-dreen-us): 2 - are listed as a threatened species in the state of Florida? As of 2006, Florida had only an estimated 225
Texas Quail Index. Result Demonstration Report 2016
 Texas Quail Index Result Demonstration Report 2016 Cooperators: Josh Kouns, County Extension Agent for Baylor County Amanda Gobeli, Extension Associate Dr. Dale Rollins, Statewide Coordinator Bill Whitley,
Texas Quail Index Result Demonstration Report 2016 Cooperators: Josh Kouns, County Extension Agent for Baylor County Amanda Gobeli, Extension Associate Dr. Dale Rollins, Statewide Coordinator Bill Whitley,
Answers to Questions about Smarter Balanced 2017 Test Results. March 27, 2018
 Answers to Questions about Smarter Balanced Test Results March 27, 2018 Smarter Balanced Assessment Consortium, 2018 Table of Contents Table of Contents...1 Background...2 Jurisdictions included in Studies...2
Answers to Questions about Smarter Balanced Test Results March 27, 2018 Smarter Balanced Assessment Consortium, 2018 Table of Contents Table of Contents...1 Background...2 Jurisdictions included in Studies...2
Result Demonstration Report
 Result Demonstration Report 2014 Texas Quail Index Texas A&M AgriLife Extension Service Wichita County Cooperator: Waggoner Ranch David Graf, County Extension Agent for Wichita County Becky Ruzicka, Extension
Result Demonstration Report 2014 Texas Quail Index Texas A&M AgriLife Extension Service Wichita County Cooperator: Waggoner Ranch David Graf, County Extension Agent for Wichita County Becky Ruzicka, Extension
Blue-crowned Laughingthrush Dryonastes courtoisi Artificial Incubation and Hand Rearing Protocol At Waddesdon Manor, Buckinghamshire, UK
 Blue-crowned Laughingthrush Dryonastes courtoisi Artificial Incubation and Hand Rearing Protocol At Waddesdon Manor, Buckinghamshire, UK Andrew Owen & Ian Edmans Incubation Blue-crowned Laughingthrush
Blue-crowned Laughingthrush Dryonastes courtoisi Artificial Incubation and Hand Rearing Protocol At Waddesdon Manor, Buckinghamshire, UK Andrew Owen & Ian Edmans Incubation Blue-crowned Laughingthrush
American Samoa Sea Turtles
 American Samoa Sea Turtles Climate Change Vulnerability Assessment Summary An Important Note About this Document: This document represents an initial evaluation of vulnerability for sea turtles based on
American Samoa Sea Turtles Climate Change Vulnerability Assessment Summary An Important Note About this Document: This document represents an initial evaluation of vulnerability for sea turtles based on
ESTIMATING NEST SUCCESS: WHEN MAYFIELD WINS DOUGLAS H. JOHNSON AND TERRY L. SHAFFER
 ESTIMATING NEST SUCCESS: WHEN MAYFIELD WINS DOUGLAS H. JOHNSON AND TERRY L. SHAFFER U.S. Fish and Wildlife Service, Northern Prairie Wildlife Research Center, Jamestown, North Dakota 58402 USA ABSTRACT.--The
ESTIMATING NEST SUCCESS: WHEN MAYFIELD WINS DOUGLAS H. JOHNSON AND TERRY L. SHAFFER U.S. Fish and Wildlife Service, Northern Prairie Wildlife Research Center, Jamestown, North Dakota 58402 USA ABSTRACT.--The
The Hatching, Growth, and Fledging of Nestling Purple Martins
 Modified from: Hill, III, J. R. 1994. The growth of nestling Purple Martins. Purple Martin Update 5(3):1-9. The Hatching, Growth, and Fledging of Nestling Purple Martins Founder & Executive Director Emeritus
Modified from: Hill, III, J. R. 1994. The growth of nestling Purple Martins. Purple Martin Update 5(3):1-9. The Hatching, Growth, and Fledging of Nestling Purple Martins Founder & Executive Director Emeritus
Removal of Alaskan Bald Eagles for Translocation to Other States Michael J. Jacobson U.S Fish and Wildlife Service, Juneau, AK
 Removal of Alaskan Bald Eagles for Translocation to Other States Michael J. Jacobson U.S Fish and Wildlife Service, Juneau, AK Bald Eagles (Haliaeetus leucocephalus) were first captured and relocated from
Removal of Alaskan Bald Eagles for Translocation to Other States Michael J. Jacobson U.S Fish and Wildlife Service, Juneau, AK Bald Eagles (Haliaeetus leucocephalus) were first captured and relocated from
Below, we present the methods used to address these objectives, our preliminary results and next steps in this multi-year project.
 Background Final Report to the Nova Scotia Habitat Conservation Fund: Determining the role of food availability on swallow population declines Project Supervisor: Tara Imlay, tara.imlay@dal.ca In the past
Background Final Report to the Nova Scotia Habitat Conservation Fund: Determining the role of food availability on swallow population declines Project Supervisor: Tara Imlay, tara.imlay@dal.ca In the past
Breeding success of Greylag Geese on the Outer Hebrides, September 2016
 Breeding success of Greylag Geese on the Outer Hebrides, September 2016 Wildfowl & Wetlands Trust Report Author Carl Mitchell September 2016 The Wildfowl & Wetlands Trust All rights reserved. No part of
Breeding success of Greylag Geese on the Outer Hebrides, September 2016 Wildfowl & Wetlands Trust Report Author Carl Mitchell September 2016 The Wildfowl & Wetlands Trust All rights reserved. No part of
Snail Habitat Preference Following Relocation Throughout the Rocky Intertidal: Pretty in Pink Chapter 6. By Julianna Rick and Sara Pratt
 Snail Habitat Preference Following Relocation Throughout the Rocky Intertidal: Pretty in Pink Chapter 6 By Julianna Rick and Sara Pratt Abstract: This study tracks and recovers Common Periwinkles and Dog
Snail Habitat Preference Following Relocation Throughout the Rocky Intertidal: Pretty in Pink Chapter 6 By Julianna Rick and Sara Pratt Abstract: This study tracks and recovers Common Periwinkles and Dog
Female Persistency Post-Peak - Managing Fertility and Production
 May 2013 Female Persistency Post-Peak - Managing Fertility and Production Michael Longley, Global Technical Transfer Manager Summary Introduction Chick numbers are most often reduced during the period
May 2013 Female Persistency Post-Peak - Managing Fertility and Production Michael Longley, Global Technical Transfer Manager Summary Introduction Chick numbers are most often reduced during the period
Female Persistency Post-Peak - Managing Fertility and Production
 Female Persistency Post-Peak - Managing Fertility and Production Michael Longley, Global Technical Transfer Manager May 2013 SUMMARY Introduction Chick numbers are most often reduced during the period
Female Persistency Post-Peak - Managing Fertility and Production Michael Longley, Global Technical Transfer Manager May 2013 SUMMARY Introduction Chick numbers are most often reduced during the period
TECHNICAL BULLETIN Claude Toudic Broiler Specialist June 2006
 Evaluating uniformity in broilers factors affecting variation During a technical visit to a broiler farm the topic of uniformity is generally assessed visually and subjectively, as to do the job properly
Evaluating uniformity in broilers factors affecting variation During a technical visit to a broiler farm the topic of uniformity is generally assessed visually and subjectively, as to do the job properly
Chapter 35 Productivity of Marbled Murrelets in California from Observations of Young at Sea
 Chapter 35 Productivity of Marbled Murrelets in California from Observations of Young at Sea C. John Ralph Linda L. Long 1 Abstract: We designed and tested an intensive survey method in 1993 to identify
Chapter 35 Productivity of Marbled Murrelets in California from Observations of Young at Sea C. John Ralph Linda L. Long 1 Abstract: We designed and tested an intensive survey method in 1993 to identify
Short Report Key-site monitoring on Hornøya in Rob Barrett & Kjell Einar Erikstad
 Short Report 3-2011 Key-site monitoring on Hornøya in 2010 Rob Barrett & Kjell Einar Erikstad SEAPOP 2011 Key-site monitoring on Hornøya in 2010 Apart from the weather which was unusually wet, the 2010
Short Report 3-2011 Key-site monitoring on Hornøya in 2010 Rob Barrett & Kjell Einar Erikstad SEAPOP 2011 Key-site monitoring on Hornøya in 2010 Apart from the weather which was unusually wet, the 2010
Atlantic Puffins By Guy Belleranti
 Flying over my head are plump seabirds with brightly colored beaks and feet. Each bird's pigeonsized body looks a little like a football with wings. The wings are too small for gliding. However, by flapping
Flying over my head are plump seabirds with brightly colored beaks and feet. Each bird's pigeonsized body looks a little like a football with wings. The wings are too small for gliding. However, by flapping
DO DIFFERENT CLUTCH SIZES OF THE TREE SWALLOW (Tachycineta bicolor)
") DO DIFFERENT CLUTCH SIZES OF THE TREE SWALLOW (Tachycineta bicolor) HAVE VARYING FLEDGLING SUCCESS? Cassandra Walker August 25 th, 2017 Abstract Tachycineta bicolor (Tree Swallow) were surveyed over a
DO DIFFERENT CLUTCH SIZES OF THE TREE SWALLOW (Tachycineta bicolor) HAVE VARYING FLEDGLING SUCCESS? Cassandra Walker August 25 th, 2017 Abstract Tachycineta bicolor (Tree Swallow) were surveyed over a
Result Demonstration Report
 Result Demonstration Report 2014 Texas Quail Index Texas A&M AgriLife Extension Service Kent County Cooperator: Reserve Ranch Jay Kingston, County Extension Agent for Kent County Becky Ruzicka, Extension
Result Demonstration Report 2014 Texas Quail Index Texas A&M AgriLife Extension Service Kent County Cooperator: Reserve Ranch Jay Kingston, County Extension Agent for Kent County Becky Ruzicka, Extension
PROTECTING MANLY S PENGUINS
 PROTECTING MANLY S PENGUINS NATIONAL PARKS AND WILDLIFE SERVICES VOLUNTARY PENGUIN WARDEN PROGRAM CONFERENCE PRESENTATION WEDNESDAY JULY 23 RD 2008 BACKGROUND: LITTLE PENGUINS EUDYPTULA MINOR o Manly s
PROTECTING MANLY S PENGUINS NATIONAL PARKS AND WILDLIFE SERVICES VOLUNTARY PENGUIN WARDEN PROGRAM CONFERENCE PRESENTATION WEDNESDAY JULY 23 RD 2008 BACKGROUND: LITTLE PENGUINS EUDYPTULA MINOR o Manly s
Texas Quail Index. Result Demonstration Report 2016
 Texas Quail Index Result Demonstration Report 2016 Cooperators: Jerry Coplen, County Extension Agent for Knox County Amanda Gobeli, Extension Associate Dr. Dale Rollins, Statewide Coordinator Circle Bar
Texas Quail Index Result Demonstration Report 2016 Cooperators: Jerry Coplen, County Extension Agent for Knox County Amanda Gobeli, Extension Associate Dr. Dale Rollins, Statewide Coordinator Circle Bar
DISTRIBUTION AND RELATIVE ABUNDANCE OF THE ALLIGATOR IN LOUISIANA COASTAL MARSHES
 DISTRIBUTION AND RELATIVE ABUNDANCE OF THE ALLIGATOR IN LOUISIANA COASTAL MARSHES LARRY McNEASE, Louisiana Department of Wildlife and Fisheries, Grand Chenier, LA 70643 TED JOANEN, Louisiana Department
DISTRIBUTION AND RELATIVE ABUNDANCE OF THE ALLIGATOR IN LOUISIANA COASTAL MARSHES LARRY McNEASE, Louisiana Department of Wildlife and Fisheries, Grand Chenier, LA 70643 TED JOANEN, Louisiana Department
Field report to Belize Marine Program, Wildlife Conservation Society
 Field report to Belize Marine Program, Wildlife Conservation Society Cathi L. Campbell, Ph.D. Nicaragua Sea Turtle Conservation Program, Wildlife Conservation Society May 2007 Principal Objective Establish
Field report to Belize Marine Program, Wildlife Conservation Society Cathi L. Campbell, Ph.D. Nicaragua Sea Turtle Conservation Program, Wildlife Conservation Society May 2007 Principal Objective Establish
Adjustments In Parental Care By The European Starling (Sturnus Vulgaris): The Effect Of Female Condition
: The Effect Of Female Condition") Proceedings of The National Conference on Undergraduate Research (NCUR) 2003 University of Utah, Salt Lake City, Utah March 13-15, 2003 Adjustments In Parental Care By The European Starling (Sturnus Vulgaris):
Proceedings of The National Conference on Undergraduate Research (NCUR) 2003 University of Utah, Salt Lake City, Utah March 13-15, 2003 Adjustments In Parental Care By The European Starling (Sturnus Vulgaris):
Nathan A. Thompson, Ph.D. Adjunct Faculty, University of Cincinnati Vice President, Assessment Systems Corporation
 An Introduction to Computerized Adaptive Testing Nathan A. Thompson, Ph.D. Adjunct Faculty, University of Cincinnati Vice President, Assessment Systems Corporation Welcome! CAT: tests that adapt to each
An Introduction to Computerized Adaptive Testing Nathan A. Thompson, Ph.D. Adjunct Faculty, University of Cincinnati Vice President, Assessment Systems Corporation Welcome! CAT: tests that adapt to each
Aspect of Bobwhite Quail Mobility During Spring Through Fall Months
 National Quail Symposium Proceedings Volume 1 Article 24 1972 Aspect of Bobwhite Quail Mobility During Spring Through Fall Months David Urban Southern llinois University Follow this and additional works
National Quail Symposium Proceedings Volume 1 Article 24 1972 Aspect of Bobwhite Quail Mobility During Spring Through Fall Months David Urban Southern llinois University Follow this and additional works
Demography and breeding success of Falklands skua at Sea Lion Island, Falkland Islands
 Filippo Galimberti and Simona Sanvito Elephant Seal Research Group Demography and breeding success of Falklands skua at Sea Lion Island, Falkland Islands Field work report - Update 2018/2019 25/03/2019
Filippo Galimberti and Simona Sanvito Elephant Seal Research Group Demography and breeding success of Falklands skua at Sea Lion Island, Falkland Islands Field work report - Update 2018/2019 25/03/2019
Summary of 2016 Field Season
 Summary of 2016 Field Season (The first year of the transfer of responsibility for MSI seabird work from Tony Diamond to Heather Major) Figure 1. The 2016 crew: L to R, Angelika Aleksieva, Marla Koberstein,
Summary of 2016 Field Season (The first year of the transfer of responsibility for MSI seabird work from Tony Diamond to Heather Major) Figure 1. The 2016 crew: L to R, Angelika Aleksieva, Marla Koberstein,
The Development of Behavior
 The Development of Behavior 0 people liked this 0 discussions READING ASSIGNMENT Read this assignment. Though you've already read the textbook reading assignment that accompanies this assignment, you may
The Development of Behavior 0 people liked this 0 discussions READING ASSIGNMENT Read this assignment. Though you've already read the textbook reading assignment that accompanies this assignment, you may
Survivorship. Demography and Populations. Avian life history patterns. Extremes of avian life history patterns
 Demography and Populations Survivorship Demography is the study of fecundity and survival Four critical variables Age of first breeding Number of young fledged each year Juvenile survival Adult survival
Demography and Populations Survivorship Demography is the study of fecundity and survival Four critical variables Age of first breeding Number of young fledged each year Juvenile survival Adult survival
Conserving Birds in North America
 Conserving Birds in North America BY ALINA TUGEND Sanderlings Andrew Smith November 2017 www.aza.org 27 Throughout the country, from California to Maryland, zoos and aquariums are quietly working behind
Conserving Birds in North America BY ALINA TUGEND Sanderlings Andrew Smith November 2017 www.aza.org 27 Throughout the country, from California to Maryland, zoos and aquariums are quietly working behind
Effects of Cage Stocking Density on Feeding Behaviors of Group-Housed Laying Hens
 AS 651 ASL R2018 2005 Effects of Cage Stocking Density on Feeding Behaviors of Group-Housed Laying Hens R. N. Cook Iowa State University Hongwei Xin Iowa State University, hxin@iastate.edu Recommended
AS 651 ASL R2018 2005 Effects of Cage Stocking Density on Feeding Behaviors of Group-Housed Laying Hens R. N. Cook Iowa State University Hongwei Xin Iowa State University, hxin@iastate.edu Recommended
THE BLUE PENGUIN (Eudyptula minor) AT TAIAROA HEAD, OTAGO,
 AT TAIAROA HEAD, OTAGO,") SCIENCE & RESEARCH SERIES NO.86 THE BLUE PENGUIN (Eudyptula minor) AT TAIAROA HEAD, OTAGO, 1992-1993 by Lyndon Perriman and Bruce McKinlay Published by Head Office, Department of Conservation, P 0 Box
SCIENCE & RESEARCH SERIES NO.86 THE BLUE PENGUIN (Eudyptula minor) AT TAIAROA HEAD, OTAGO, 1992-1993 by Lyndon Perriman and Bruce McKinlay Published by Head Office, Department of Conservation, P 0 Box
Swan & Goose IDentification It s Important to Know
 Swan & Goose IDentification It s Important to Know Reports from wildlife watchers and sportsmen will help the biologists monitor the recovery of trumpeter swans (Cygnus buccinator). Positive identification
Swan & Goose IDentification It s Important to Know Reports from wildlife watchers and sportsmen will help the biologists monitor the recovery of trumpeter swans (Cygnus buccinator). Positive identification
Short Report Key-site monitoring on Hornøya in Rob Barrett & Kjell Einar Erikstad
 Short Report 2-2010 Key-site monitoring on Hornøya in 2009 Rob Barrett & Kjell Einar Erikstad SEAPOP 2010 Key-site monitoring on Hornøya in 2009 The 2009 breeding season was in general good for most species
Short Report 2-2010 Key-site monitoring on Hornøya in 2009 Rob Barrett & Kjell Einar Erikstad SEAPOP 2010 Key-site monitoring on Hornøya in 2009 The 2009 breeding season was in general good for most species
FEATURED PHOTO NOTES ON PLUMAGE MATURATION IN THE RED-TAILED TROPICBIRD
 FEATURED PHOTO NOTES ON PLUMAGE MATURATION IN THE RED-TAILED TROPICBIRD Ron Levalley, Mad River Biologists, 920 Samoa Blvd., Suite 210, Arcata, California 95521; ron@madriverbio.com PETER PYLE, The Institute
FEATURED PHOTO NOTES ON PLUMAGE MATURATION IN THE RED-TAILED TROPICBIRD Ron Levalley, Mad River Biologists, 920 Samoa Blvd., Suite 210, Arcata, California 95521; ron@madriverbio.com PETER PYLE, The Institute
Analysis of Sampling Technique Used to Investigate Matching of Dorsal Coloration of Pacific Tree Frogs Hyla regilla with Substrate Color
 Analysis of Sampling Technique Used to Investigate Matching of Dorsal Coloration of Pacific Tree Frogs Hyla regilla with Substrate Color Madeleine van der Heyden, Kimberly Debriansky, and Randall Clarke
Analysis of Sampling Technique Used to Investigate Matching of Dorsal Coloration of Pacific Tree Frogs Hyla regilla with Substrate Color Madeleine van der Heyden, Kimberly Debriansky, and Randall Clarke
Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve,
 at Sungei Buloh Wetland Reserve,") Author Title Institute Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve, Singapore Thesis (Ph.D.) National
Author Title Institute Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve, Singapore Thesis (Ph.D.) National
Living Planet Report 2018
 Living Planet Report 2018 Technical Supplement: Living Planet Index Prepared by the Zoological Society of London Contents The Living Planet Index at a glance... 2 What is the Living Planet Index?... 2
Living Planet Report 2018 Technical Supplement: Living Planet Index Prepared by the Zoological Society of London Contents The Living Planet Index at a glance... 2 What is the Living Planet Index?... 2
Unusual 2nd W Common Gull Larus canus at Helsingborg
 Unusual 2nd W Common Gull Larus canus at Helsingborg View PDF at high zoom for optimal picture resolution On 22 nd of March 2015, 3 rd CY Common Gull Larus canus with black markings in tail and to a lesser
Unusual 2nd W Common Gull Larus canus at Helsingborg View PDF at high zoom for optimal picture resolution On 22 nd of March 2015, 3 rd CY Common Gull Larus canus with black markings in tail and to a lesser
Steller Sea Lions at Cattle Point. Sarah Catherine Milligan. Pelagic Ecosystem Function Research Apprenticeship Fall 2014
 Pinniped Abundance and Distribution in the San Juan Channel, and Haulout Patterns of Steller Sea Lions at Cattle Point Sarah Catherine Milligan Pelagic Ecosystem Function Research Apprenticeship Fall 214
Pinniped Abundance and Distribution in the San Juan Channel, and Haulout Patterns of Steller Sea Lions at Cattle Point Sarah Catherine Milligan Pelagic Ecosystem Function Research Apprenticeship Fall 214
Piping Plover. Below: Note the color of the sand and the plover s back.
 Piping Plover Below: Note the color of the sand and the plover s back. Above: Chicks and one egg left in the nest. Once the eggs hatch the chicks leave the nest to forage for food on the sandbar. Plovers
Piping Plover Below: Note the color of the sand and the plover s back. Above: Chicks and one egg left in the nest. Once the eggs hatch the chicks leave the nest to forage for food on the sandbar. Plovers
Osprey Watch Osprey Monitoring Guidelines
 Osprey Watch Osprey Monitoring Guidelines Here are the guidelines for volunteering to be a member of Greenbelt s Osprey Watch! Below you will find methodology explained, tips, and other informational facts
Osprey Watch Osprey Monitoring Guidelines Here are the guidelines for volunteering to be a member of Greenbelt s Osprey Watch! Below you will find methodology explained, tips, and other informational facts
EIDER JOURNEY It s Summer Time for Eiders On the Breeding Ground
 The only location where Steller s eiders are still known to regularly nest in North America is in the vicinity of Barrow, Alaska (Figure 1). Figure 1. Current and historic Steller s eider nesting habitat.
The only location where Steller s eiders are still known to regularly nest in North America is in the vicinity of Barrow, Alaska (Figure 1). Figure 1. Current and historic Steller s eider nesting habitat.
Note: The following article is used with permission of Dr. Sonia Altizer.
 PROFESSIONAL BUTTERFLY FARMING PART I - By Nigel Venters (Contributing Author: Dr. Sonia Altizer) Note: The following article is used with permission of Dr. Sonia Altizer. Monarch Health Program, University
PROFESSIONAL BUTTERFLY FARMING PART I - By Nigel Venters (Contributing Author: Dr. Sonia Altizer) Note: The following article is used with permission of Dr. Sonia Altizer. Monarch Health Program, University
A practical field guide to the identification of Least Terns in various plumages
 A practical field guide to the identification of Least Terns in various plumages Edited by Marianne Korosy and Elizabeth A. Forys, PhD Photo: Charles Buhrman This is an adult Least Tern (Sternula antillarum)
A practical field guide to the identification of Least Terns in various plumages Edited by Marianne Korosy and Elizabeth A. Forys, PhD Photo: Charles Buhrman This is an adult Least Tern (Sternula antillarum)
Name. Period. Student Activity: Dichotomous Key. 1a. 1b. 2a. 2b. 3a. 3b. 4a. 4b. 5a. 5b. 6a. 6b. 7a. 7b. 8a.
 Name Period Student Activity: Dichotomous Key 1a. 1b. Question Identify/Go to 2a. 2b. 3a. 3b. 4a. 4b. 5a. 5b. 6a. 6b. 7a. 7b. 8a. 8b. Name Period CLASSIFICATION KEY FOR FISHES OF UTAH LAKE Examine the
Name Period Student Activity: Dichotomous Key 1a. 1b. Question Identify/Go to 2a. 2b. 3a. 3b. 4a. 4b. 5a. 5b. 6a. 6b. 7a. 7b. 8a. 8b. Name Period CLASSIFICATION KEY FOR FISHES OF UTAH LAKE Examine the
Mexican Gray Wolf Reintroduction
 Mexican Gray Wolf Reintroduction New Mexico Supercomputing Challenge Final Report April 2, 2014 Team Number 24 Centennial High School Team Members: Andrew Phillips Teacher: Ms. Hagaman Project Mentor:
Mexican Gray Wolf Reintroduction New Mexico Supercomputing Challenge Final Report April 2, 2014 Team Number 24 Centennial High School Team Members: Andrew Phillips Teacher: Ms. Hagaman Project Mentor:
Crotophaga major (Greater Ani)
") Crotophaga major (Greater Ani) Family: Cuculidae (Cuckoos and Anis) Order: Cuculiformes (Cuckoos, Anis and Turacos) Class: Aves (Birds) Fig. 1. Greater ani, Crotophaga major. [http://www.birdforum.net/opus/greater_ani,
Crotophaga major (Greater Ani) Family: Cuculidae (Cuckoos and Anis) Order: Cuculiformes (Cuckoos, Anis and Turacos) Class: Aves (Birds) Fig. 1. Greater ani, Crotophaga major. [http://www.birdforum.net/opus/greater_ani,
2009 Eagle Nest News from Duke Farms eagle nest Written by Larissa Smith, Assistant Biologist
 2009 Eagle Nest News from Duke Farms eagle nest Written by Larissa Smith, Assistant Biologist July 7 - The youngest chick was gone from the nest this morning but has returned to the nest several times
2009 Eagle Nest News from Duke Farms eagle nest Written by Larissa Smith, Assistant Biologist July 7 - The youngest chick was gone from the nest this morning but has returned to the nest several times
Pill Bug Environmental Preferences Based on Moisture
 Another lab report sample: This is Natasha and Stephanie s first attempt at a lab report so bear that in mind as you read this sample report. Natasha Duell Stephanie Moeller AP Bio/Sastry 10 September
Another lab report sample: This is Natasha and Stephanie s first attempt at a lab report so bear that in mind as you read this sample report. Natasha Duell Stephanie Moeller AP Bio/Sastry 10 September
Supplementary Fig. 1: Comparison of chase parameters for focal pack (a-f, n=1119) and for 4 dogs from 3 other packs (g-m, n=107).
 and for 4 dogs from 3 other packs (g-m, n=107).") Supplementary Fig. 1: Comparison of chase parameters for focal pack (a-f, n=1119) and for 4 dogs from 3 other packs (g-m, n=107). (a,g) Maximum stride speed, (b,h) maximum tangential acceleration, (c,i)
Supplementary Fig. 1: Comparison of chase parameters for focal pack (a-f, n=1119) and for 4 dogs from 3 other packs (g-m, n=107). (a,g) Maximum stride speed, (b,h) maximum tangential acceleration, (c,i)
SEASONAL PATTERNS OF NESTING IN THE RED-WINGED BLACKBIRD MORTALITY
 Condor, 80:290-294 0 The Cooper Ornithological Society 1978 SEASONAL PATTERNS OF NESTING IN THE RED-WINGED BLACKBIRD MORTALITY DONALD F. CACCAMISE It is likely that birds adjust their reproductive period
Condor, 80:290-294 0 The Cooper Ornithological Society 1978 SEASONAL PATTERNS OF NESTING IN THE RED-WINGED BLACKBIRD MORTALITY DONALD F. CACCAMISE It is likely that birds adjust their reproductive period