Henipavirus V Protein Association with Polo-Like Kinase Reveals Functional Overlap with STAT1 Binding and Interferon Evasion
|
|
|
- Anna Floyd
- 5 years ago
- Views:
Transcription
1 JOURNAL OF VIROLOGY, July 2008, p Vol. 82, No X/08/$ doi: /jvi Copyright 2008, American Society for Microbiology. All Rights Reserved. Henipavirus V Protein Association with Polo-Like Kinase Reveals Functional Overlap with STAT1 Binding and Interferon Evasion Louise E. Ludlow, 1,2 Michael K. Lo, 3,4 Jason J. Rodriguez, 1,2 Paul A. Rota, 3 and Curt M. Horvath 1,2 * Department of Medicine, and Department of Biochemistry, Molecular Biology, and Cell Biology, Northwestern University, Evanston, Illinois ; Department of Medicine, Evanston Northwestern Healthcare, Evanston, Illinois ; Measles, Mumps, Rubella, and Herpes Laboratory Branch, Centers for Disease Control and Prevention, 1600 Clifton Road, MS C-22, Atlanta, Georgia ; and Department of Microbiology and Immunology, Emory University School of Medicine, 1510 Clifton Road, Atlanta, Georgia Received 25 February 2008/Accepted 7 April 2008 Emerging viruses in the paramyxovirus genus Henipavirus evade host antiviral responses via protein interactions between the viral V and W proteins and cellular STAT1 and STAT2 and the cytosolic RNA sensor MDA5. Polo-like kinase (PLK1) is identified as being an additional cellular partner that can bind to Nipah virus P, V, and W proteins. For both Nipah virus and Hendra virus, contact between the V protein and the PLK1 polo box domain is required for V protein phosphorylation. Results indicate that PLK1 is engaged by Nipah virus V protein amino acids 100 to 160, previously identified as being the STAT1 binding domain responsible for host interferon (IFN) signaling evasion, via a Thr-Ser-Ser-Pro motif surrounding residue 130. A distinct Ser-Thr-Pro motif surrounding residue 199 mediates the PLK1 interaction with Hendra virus V protein. Select mutations in the motif surrounding residue 130 also influenced STAT1 binding and innate immune interference, and data indicate that the V:PLK1 and V:STAT complexes are V mediated yet independent of one another. The effects of STAT1/PLK1 binding motif mutations on the function the Nipah virus P protein in directing RNA synthesis were tested. Remarkably, mutations that selectively disrupt the STAT or PLK1 interaction site have no effects on Nipah virus P protein-mediated viral RNA synthesis. Therefore, mutations targeting V protein-mediated IFN evasion will not alter the RNA synthetic capacity of the virus, supporting an attenuation strategy based on disrupting host protein interactions. The genus Henipavirus within the family Paramyxoviridae was created in 2002 to accommodate the recently emerging and closely related Nipah and Hendra viruses (10, 28, 46, 47). Nipah virus emerged in Peninsular Malaysia in 1998 as a result of human contact with infected swine, and recent outbreaks have been reported in Bangladesh and India between 2001 and 2005 (7, 17, 20, 27). Hendra virus emerged in Brisbane, Australia, in 1994 as a result of human contact with infected horses and last appeared in Queensland, Australia, in 2004 (3, 32). The viruses were responsible for zoonotic respiratory disease and severe acute encephalitis in humans and livestock and exhibit respiratory and neurological tropism (24). These human pathogens are set apart by their low homology to other paramyxoviruses, wide host range, and high level of virulence, which has limited investigations into the interaction of virally encoded Henipavirus proteins with their natural host and susceptible livestock. Furthermore, these viruses caused significant economic losses for the Malaysian swine industry (30), and their potential use in bioterrorist scenarios (23), due to a lack of therapeutic intervention, warrants further investigation. The cellular response to type I interferons (IFNs) (IFN- and IFN-, collectively referred to as IFN) results in the establishment of a potent antiviral state in addition to regulating components of the adaptive immune response (21, 22). This * Corresponding author. Mailing address: Pancoe-ENH Research Pavilion, Northwestern University, 2200 Campus Drive, Evanston, IL Phone: (847) Fax: (847) Published ahead of print on 16 April process is mediated by the signal transducer and activator of transcription (STAT) protein family. Following IFN exposure, STAT proteins translocate to the nucleus to directly regulate cellular gene transcription, creating a cellular antiviral state (1). Many viruses have evolved well-characterized adaptations to evade the IFN-induced antiviral responses of their hosts (15). The family Paramyxoviridae, comprised of enveloped negative-strand RNA viruses in several distinct genera, has developed unique molecular mechanisms of IFN signaling inhibition, several of which have been attributed to a virus-encoded protein derived from a polycistronic gene (10, 18, 19). As with other paramyxoviruses, the cotranscriptional insertion of nontemplated nucleotides generates alternative mrnas from the Henipavirus P locus, encoding the P, V, and W proteins (45). A fourth protein, C, is generated by alternate translation initiation site selection from all these mrnas and is unrelated to the other products. The P, V, and W proteins share 407 amino acids in their N termini but differ in their C termini as a result of alternate reading frames produced by the addition of nontemplated nucleotides at the editing site (Fig. 1A). The P protein contains an additional 301 amino acids to produce a 78-kDa polypeptide. The P protein is highly phosphorylated, albeit by unknown kinase(s) (43), and is an essential component of the RNA transcription and replication machinery (8). The V protein C-terminal domain (CTD) is 50 amino acids in length and encodes a zinc finger domain that is a highly conserved hallmark domain of paramyxovirus V proteins (26, 36). The Nipah virus and Hendra virus V proteins are well known to antagonize antiviral signaling by interaction and interference with STAT1 and STAT2 transcription factors (38, 6259

 illustrating the STAT1 binding domain, the cysteine-rich CTD, and")
 Nipah virus V and Hendra virus V (HeV) but not measles virus V (MeV) proteins")



 Expressed HA-tagged PLK1 copurifies with Nipah virus V and Hendra virus V")




 Purification of PLK1.")
 and elution with HA peptide.")
 Bacterial expression of recombinant Henipavirus GST-V proteins.")



.")



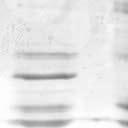

2 6260 LUDLOW ET AL. J. VIROL. Downloaded from FIG. 1. Henipavirus P, V, and W proteins associate with PLK1. (A) Schematic representation of the Nipah virus P, V, and W proteins (NiP, NiV, and NiW, respectively) illustrating the STAT1 binding domain, the cysteine-rich CTD, and the nuclear localization signal (NLS). (B) Nipah virus V and Hendra virus V (HeV) but not measles virus V (MeV) proteins copurify with endogenous PLK1. 293T cells were transfected to express FLAG-tagged measles virus V, Nipah virus V, Hendra virus V, or GFP, followed by immunoprecipitation with FLAG affinity gel and elution with FLAG peptide. The FLAG eluate and lysate samples were subjected to immunoblot analysis to detect endogenous PLK1 or FLAG-V protein. The asterisk indicates the immunoglobulin G (IgG) heavy chain, which migrates near PLK1. (C) Expressed HA-tagged PLK1 copurifies with Nipah virus V and Hendra virus V but not measles virus V. 293T cells were transfected to express HA-PLK1 together with either FLAG-tagged measles virus V, Nipah virus V, Hendra virus V, or GFP, followed by HA immunoprecipitation and elution with HA peptide. The HA immune complexes and lysate samples were subjected to immunoblotting with the FLAG antibody to detect V or the anti-ha antibody to detect HA-PLK1. (D) Nipah virus P and W proteins copurify with endogenous PLK1. 293T cells were transfected to express FLAG-tagged Nipah virus P, Nipah virus V, Nipah virus W, or GFP, followed by immunoprecipitation with FLAG affinity gel and elution with FLAG peptide. The FLAG eluate and lysate samples were subjected to immunoblot analysis to detect endogenous PLK1 or FLAG-V protein. (E) Purified HA-PLK1 phosphorylates recombinant Henipavirus V proteins in vitro. (i) Purification of PLK1. 293T cells were transfected to express HA-PLK1, followed by HA immunoprecipitation (I.P.) and elution with HA peptide. Purified PLK1 and the input samples were subjected to SDS-PAGE and silver staining. (ii) Bacterial expression of recombinant Henipavirus GST-V proteins. Recombinant GST-FLAG-Nipah virus V, GST-FLAG-Hendra virus V, and control GST-FLAG were purified from bacteria and analyzed by SDS-PAGE and Coomassie blue staining. (iii) Henipavirus V proteins are phosphorylated by purified HA-PLK1. Approximately 1, 5, or 10 ng of purified PLK1 was incubated with 5 g of bacterially expressed Henipavirus V protein or dephosphorylated bovine casein in the presence of [ 32 -P]ATP. Reaction mixtures were resolved by SDS-PAGE and visualized by autoradiography. on January 15, 2019 by guest 39, 41, 42). The Nipah virus V protein shuttles between the nucleus and cytoplasm, which is mediated by a chromosomal region maintenance 1-dependent nuclear export signal at amino acids 174 to 192 (37), and this behavior and sequence are conserved in the Hendra virus V protein. Henipavirus V proteins inhibit IFN responses by sequestering STAT1 and STAT2 in high-molecular-weight cytoplasmic complexes (38, 39). As a result, IFN-induced STAT tyrosine phosphorylation is prevented, resulting in the inhibition of an antiviral state. The primary IFN evasion target of the Nipah virus V protein is STAT1, which is absolutely required for the STAT2 association (37). Both IFN evasion and STAT1 binding were mapped to amino acids 100 to 160 in the N terminus of the Nipah virus V protein. In addition, the Henipavirus V protein CTD was found to interact with the RNA sensor MDA5, preventing double-stranded RNA signaling (2, 6). The W protein CTD is 44 amino acids long and contains a functional nuclear import signal (42). The Nipah virus W protein has been reported to modulate both STAT1 and Toll-like receptor 3-dependent IFN regulatory factor 3 signaling (41). All four products of the Nipah virus P locus have been demonstrated to antagonize cellular antiviral responses (34), and due to their identical
3 VOL. 82, 2008 HENIPAVIRUS V PROTEIN PHOSPHORYLATION BY PLK N-terminal domains, the P, V, and W proteins all share the 60-amino-acid STAT1 binding site between residues 100 and 160, which may account for much of their observed IFN signaling evasion capabilities. The formation of protein complexes is crucial for V-mediated IFN evasion, but more information regarding the interface with the host cell is essential for understanding Henipavirus pathogenesis and designing therapeutic or antiviral reagents. Affinity purification was used to identify additional host factors that might contribute to Henipavirus biology. This approach identified polo-like kinase 1 (PLK1) as being a Henipavirus P, V, and W protein-interacting partner. Data indicate that PLK1 is recruited by the 60-aminoacid STAT1 binding domain and phosphorylates the Nipah virus V protein. Molecular analysis demonstrates the importance of consensus phosphopeptide polo box domain (PBD) binding motifs with overlapping functions for both PLK1 and STAT1 interactions. MATERIALS AND METHODS Cell culture. Human embryonic kidney 293T, human fibrosarcoma 2fTGH, 2fTGH-derivative U3A:STAT1-deficient (29), and U6A:STAT2 deficient (25) cell lines were maintained in Dulbecco s modified Eagle medium supplemented with cosmic calf serum (10%; HyClone), penicillin (100 U/ml), and streptomycin (100 g/ml; Gibco-BRL). Plasmids, transfections, and luciferase assay. Expression plasmids pef- FLAG, pef-flag-measles V, pef-flag-nipah V, and pef-flag-hendra V were described previously (33, 38, 39). Nipah virus V truncation vectors and Rev1.4-green fluorescent protein (GFP) fusion vectors were previously described (37). FLAG-tagged Nipah virus and Hendra virus V cdnas were amplified by PCR and subcloned in frame into vector pgex-6p-1 (GE Healthcare) to allow the production of bacterially expressed glutathione S-transferase (GST) fusion proteins. Nipah virus P and W cdnas were subcloned in frame by PCR into vector pef-flag using templates ptm1-nip and pcaggs-niw (kindly provided by Michael K. Lo and Paul A. Rota, CDC, Atlanta, GA). Human PLK1 cdna (GenBank accession number NM_ ; OriGene) was amplified by PCR and cloned into pcdna3 (Invitrogen) with an N-terminal hemagglutinin (HA) epitope sequence. PLK1 fragments encoding the N terminus (bp 1 to 1034) and C terminus (bp 1035 to 1809) were generated by PCR and subcloned in frame into plasmid pcdna3-ha. Single point mutations in Nipah virus V, Nipah virus P, and PLK1 were introduced using the QuikChange II site-directed mutagenesis kit (Stratagene) according to the manufacturer s recommendations. All cdna constructs and point mutations were verified by DNA sequencing. High-efficiency transient transfection of 293T cells for affinity purification was carried out by use of a standard calcium phosphate method where 3 g of FLAG-tagged V protein was transfected per 100-mm plate. For cotransfection experiments, FLAG-V and HA-PLK1 were expressed at a 1:1 ratio. U3A and U6A cells were transfected using SuperFect (Qiagen). For luciferase assays, 2fTGH or 293T cells in six-well plates were transiently transfected with 2 g of reporter gene (5 IFN-stimulated response element (ISRE)-luciferase for IFN- and 4 M67 IFN- -activated site (GAS)-luciferase for IFN- ) and 6 g of FLAG-tagged V or plasmid pef-flag alone using SuperFect (Qiagen). IFN- (1,000 U/ml) or IFN- (5 ng/ml) was added for 10 h, and analysis was performed using the dual-luciferase assay system (Promega); as an internal control, the Renilla luciferase construct was used to normalize activity. Cell extracts, immunoprecipitation, and immunoblotting. Cells transfected with expression plasmids were lysed in whole-cell extract buffer (50 mm Tris-HCl [ph 8.0], 280 mm NaCl, 0.5% NP-40, 0.2 mm EDTA, 0.2 mm EGTA, 10% glycerol, 1 mm dithiothreitol supplemented with protease inhibitor cocktail [Complete; Boehringer Mannheim], and 1 mm Na 3 VO 4 ). The clarified cell lysate was precleared with Sepharose 6B (Sigma-Aldrich) and then incubated with anti-flag M2 affinity agarose (Sigma-Aldrich) or EZview Red anti-ha affinity gel (Sigma-Aldrich) to purify FLAG- or HA-tagged protein by immunoaffinity. Immunoprecipitation of the GFP fusion protein was previously described (37). Agarose beads with immune complexes were washed five times with whole-cell extract buffer and once with PEB:300 buffer (20 mm Tris-HCl [ph 7.5], 300 mm NaCl, 0.2 mm EDTA, 0.1% NP-40, 15% [vol/vol] glycerol), and bound proteins were competitively eluted in PEB:300 buffer using 150 g of3 FLAG peptide (Sigma-Aldrich) or influenza virus HA peptide (Sigma-Aldrich), boiled in protein loading buffer, and separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) for immunoblot analysis. Protein was transferred onto nitrocellulose, and antibodies were as follows: rabbit anti-stat1 (1:4,000) (sc-345; Santa Cruz), rabbit anti-stat2 (1:4,000) (sc-476; Santa Cruz), mouse anti-plk1 (1:2,000) (ab17056; AbCam), rabbit anti-flag tag (1:4,000) (F7425; Sigma-Aldrich), and rabbit anti-ha tag (1:4,000) (H6908; Sigma-Aldrich). Primary antibody incubations were done overnight at 4 C. Secondary antibodies used were goat anti-mouse and goat anti-rabbit conjugated to horseradish peroxide (VWR). Detection was performed with enhanced chemiluminescence reagent (Perkin-Elmer). Proteomics analysis. Following large-scale immunoprecipitation of FLAG- Nipah virus V and FLAG-Hendra virus V, copurified proteins were separated by SDS-PAGE, and large gels were stained using Coomassie brilliant blue R-250 (0.1% [wt/vol] R-250 in 40% [vol/vol] methanol 10% [vol/vol] acetic acid; Bio- Rad) and destained in 40% (vol/vol) and 10%(vol/vol) acetic acid. Selected protein bands were excised for identification using the thermo-ltq-ft mass spectrometer at the CBC-RRC Proteomics Core Laboratory (University of Illinois at Chicago). Peptides with multiple hits were assessed using the Mascot 2.1 software platform and Bioworks 3.2 Turbosequest. In vitro kinase assay. To obtain purified PLK1, 293T cells were transfected using the calcium phosphate transfection method with pcdna3.1-ha-plk1. Cells were lysed in TBSN buffer [20 mm Tris-Cl (ph 8.0), 150 mm NaCl, 0.5% NP-40, 5 mm EGTA, 1.5 mm EDTA supplemented with 0.5 mm Na 3 VO 4,20 mm p-nitrophenyl phosphate (Sigma-Aldrich), 1 mm 4-(2-aminoethyl)-benzenesulfonyl fluoride (AEBSF) (Pefabloc; Roche) supplemented with protease inhibitor cocktail (Complete; Boehringer Mannheim)]. Immunoprecipitation was carried out using HA affinity gel, and bound complexes were eluted using HA peptide in TBMD buffer (50 mm Tris-Cl [ph 8.0], 10 mm MgCl 2, 5 mm dithiothreitol, 2 mm EGTA, 0.5 mm Na 3 VO 4, and 20 nm p-nitrophenyl phosphate [Sigma-Aldrich]), and purified PLK1 was visualized using silver stain and quantified using Bradford estimation (4a). Kinase activity assays were carried out in TBMD buffer; approximately 5 g of bacterially produced V protein or 3 g of dephosphorylated bovine casein (Sigma-Aldrich) substrate was combined with either 1, 5, or 10 ng of purified HA-PLK1 in the presence of 50 M ATP and 5 Ci of [ - 32 P]ATP. Following incubation at 30 C for 30 min, the reaction was terminated by the addition of protein gel loading buffer to the reaction mixture, which was separated by SDS-PAGE and visualized by autoradiography. Immune complex kinase assay. Immunoprecipitation of FLAG-tagged V proteins was carried out as described above, and bound proteins were eluted in TBMD buffer supplemented as described above. Eluates were incubated in the presence of 50 M ATP and 5 Ci of [ - 32 P]ATP at 30 C for 30 min; the reaction was terminated by the addition of protein gel loading buffer to the reaction mixture, which was separated by SDS-PAGE and visualized by autoradiography. Indirect immunofluorescence. 2fTGH cells were grown to 60 to 80% confluence in Permanox chamber slides (Nalgene) and transfected with FLAG-tagged V protein using SuperFect (Qiagen). Indirect immunofluorescence was performed as described previously by Rodriguez et al. (38) using mouse anti-flag M2 monoclonal antibody (Sigma), followed by Alexa 546-conjugated anti-mouse secondary antibody (Invitrogen). The second stain for STAT1 and STAT2 was detected with anti-rabbit fluorescein isothiocyanate (Invitrogen). Images were obtained using a Leica confocal microscope at the Biological Imaging Facility, Northwestern University, Evanston, IL. Antiviral assay. Pools of 2fTGH cells stably expressing Nipah virus V protein, Hendra virus V protein, phosphopeptide binding motif mutations, and the pef- FLAG control were generated by the cotransfection of 10 g of V DNA and 1 g pbabe-puro (31) using SuperFect (Qiagen). Resistant pools were selected for 2 weeks using 1 g/ml puromycin (Sigma), and expression was verified using immunoblotting. Following an 8-h treatment with IFN- or medium alone, cells were infected with recombinant vesicular stomatitis virus (VSV) harboring a GFP transgene (VSV-GFP) (a gift of John Hiscott, Montreal, Canada). Infections were performed at a multiplicity of infection of 0.6 PFU/ml, and virus was diluted in serum-free medium and added to the cells for 1 h to allow adsorption. Virus was then removed, and Dulbecco s modified Eagle medium with 2% cosmic calf serum was added for the remaining time. After 22 h, GFP levels were analyzed by flow cytometry (BD FACSCalibur; BD Biosciences), and data were assessed using CELLquest (BD Biosciences). Percent GFP was calculated in comparison to uninfected 2fTGH cells. Cells were photographed using a Zeiss inverted fluorescence microscope. Minigenome assay and CAT ELISA. BHK/sr/T7 cells were transfected with 1.75 g of Nipah virus N, 0.8 g of Nipah virus P (or Nipah virus P point mutations), 1.2 g of Nipah virus L, and 3.5 g of Nipah virus chloramphenicol acetyltransferase (CAT) minigenome plasmid constructs in Opti-MEM medium (Life Technologies, Invitrogen) using LT-1 transfection reagent (Mirus) accord-









")





,")

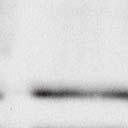






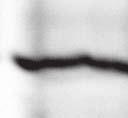
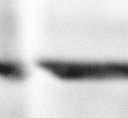















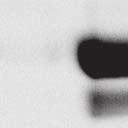













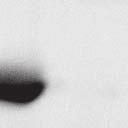

























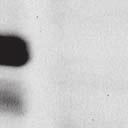









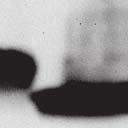












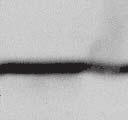
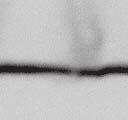





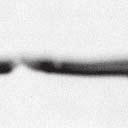
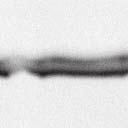
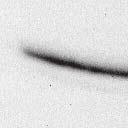
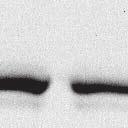
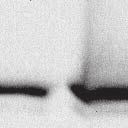
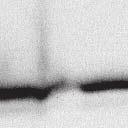
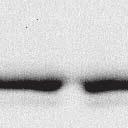
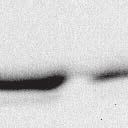
4 6262 LUDLOW ET AL. J. VIROL. Downloaded from FIG. 2. PLK1 binds Nipah virus V protein via the STAT1 binding domain. (A) Selective PLK1, STAT1, and STAT2 binding to Nipah virus V (NiV) fragments. 293T cells were transfected to express FLAG-tagged Nipah virus V, GFP control, and Nipah virus V truncation constructs (37), followed by FLAG immunoprecipitation and elution with FLAG peptide. The FLAG eluate and lysate samples were analyzed by immunoblotting using antibodies to detect endogenous PLK1, STAT1, STAT2, and FLAG-V proteins. (B) Interaction of Nipah virus V protein with PLK1, STAT1, and STAT2. 293T cells were transfected to express the Rev1.4-GFP fusion vectors as indicated (37), and lysates were immunoprecipitated (IP) with antisera to GFP. Immune complexes were probed for PLK1, STAT1, STAT2, and GFP. (C) Schematic representation of Nipah virus V protein illustrating the PLK1 and STAT1 interaction domains. The consensus S-[pS/pT]-P motifs for Nipah virus and Hendra virus V (HeV) proteins are indicated by boldface type and double underlining. ing to the manufacturer s instructions. Total amounts of transfected DNA were kept constant by the addition of ptm1 empty vector DNA where applicable. For the negative control, the Nipah virus L plasmid was replaced with an equivalent amount of ptm1 empty vector. Details of the Nipah virus minigenome replication assay were described elsewhere previously (16). Cytoplasmic extracts were prepared 42 h posttransfection in 500 l of lysis buffer from the CAT enzymelinked immunosorbent assay (ELISA) kit (Roche). To ensure comparable results, the amounts of harvested cytoplasmic extracts used per sample were adjusted for protein concentration using the BCA protein assay reagent (Pierce). The total protein concentration of the lysate supernatants and the concentration of CAT reporter protein in the samples were determined according to protein/ enzyme standards used for each respective assay. Twenty-five micrograms of total protein per sample lysate was used for the CAT ELISA (Roche), which was performed according to the manufacturer s protocols. The assay was performed three separate times, in which each sample was assayed in duplicate. RESULTS Henipavirus P, V, and W proteins copurify with PLK1. Large-scale immunoprecipitation and mass spectrometry identified PLK1 as being a Henipavirus V protein-interacting partner (Fig. 1). To confirm this association, FLAG-tagged V proteins expressed in human embryonic kidney 293T cells were immunoprecipitated using FLAG affinity agarose and assessed for endogenous PLK1 binding. Trace amounts of PLK1 were found to coprecipitate with control FLAG-GFP or FLAGtagged measles virus V protein, but the Nipah virus and Hendra virus V proteins strongly copurified endogenous PLK1 (Fig. 1B). The protein interaction was independently confirmed in human fibrosarcoma 2fTGH cells and 2fTGH cells stably expressing the FLAG-Nipah virus V protein (data not shown). In a complementary experiment, 293T cells expressing both HA-tagged PLK1 and FLAG-tagged V protein were analyzed by HA affinity purification (Fig. 1C). The expressed HA-PLK1 copurified with the Nipah virus and Hendra virus V proteins but not the measles virus V protein or control FLAG- GFP. To determine if the Nipah virus P and W proteins (Fig. on January 15, 2019 by guest

, or the C-terminal PBD (residues 345 to 603) together with either FLAG-tagged Nipah virus V")
 PBD contact residues mediate V protein interactions.")


.")
5 VOL. 82, 2008 HENIPAVIRUS V PROTEIN PHOSPHORYLATION BY PLK FIG. 3. The PLK1 PBD is necessary and sufficient to mediate Henipavirus V interactions. (A) Illustration of PLK1 domain structure. The PLK1 N-terminal kinase domain containing residue K82, which is required for ATP binding, and the C-terminal PBD containing residues W414, H538, and K540, which are required for contact with phosphopeptide substrates, are overlined. (B) The PBD mediates Henipavirus V protein interactions. 293T cells were cotransfected to express HA-PLK1, the HA-PLK1 N-terminal kinase domain (residues 1 to 344), or the C-terminal PBD (residues 345 to 603) together with either FLAG-tagged Nipah virus V (NiV) (i), Hendra virus V (HeV) (ii), or GFP (iii), followed by FLAG immunoprecipitation and elution. The FLAG immune complexes and lysate samples were subjected to immunoblot analysis to detect FLAG-V, HA-PLK1, or HA-PLK1 fragments. (C) PBD contact residues mediate V protein interactions. 293T cells were cotransfected to express HA-PLK1 or HA-PLK1 point mutations together with either FLAG-tagged Nipah virus V (i), Hendra virus V (ii), or GFP (iii), followed by FLAG immunoprecipitation and elution. The FLAG immune complexes and lysate samples were subjected to immunoblotting to detect FLAG-V, HA-PLK1, or HA-PLK1 point mutations. 1A) also copurified with endogenous PLK1, FLAG-tagged P and W proteins expressed in 293T cells were immunoprecipitated and assessed for endogenous PLK1 binding (Fig. 1D). In addition to the V protein, both the P and W proteins coprecipitated with PLK1. These results confirm PLK1 to be a Henipavirus protein partner. Purified PLK1 phosphorylates recombinant Henipavirus V proteins in vitro. To determine if PLK1 is able to phosphorylate the Henipavirus V proteins, an in vitro kinase assay was performed. PLK1 was purified by immunoprecipitation of HAtagged PLK1 expressed in human 293T cells, and silver staining revealed a highly enriched PLK1 preparation (Fig.1Ei). Recombinant GST-FLAG-Nipah V, GST-FLAG-Hendra V, and control GST-FLAG were expressed and purified from Escherichia coli cells (Fig. 1Eii). Approximately 1, 5, or 10 ng of purified PLK1 was incubated with 5 g of bacterially expressed Henipavirus V protein or a control, dephosphorylated bovine casein in the presence of [ 32 -P]ATP. Reaction mixtures were resolved by SDS-PAGE and visualized by autoradiography (Fig.1Eiii). Like the positive casein control, Henipavirus V proteins were phosphorylated by PLK1 in a dosedependent manner, but GST alone was not. This result suggests that the PLK1-V protein interaction can lead to V protein phosphorylation. PLK1 binds Nipah virus V protein via the STAT1 binding domain. To determine the V protein region required for the PLK1 association, two sets of previously described tagged Nipah virus V protein fragments were used for immunoprecipitation experiments (37). The immune complexes were tested for the presence of PLK1, STAT1, STAT2, and FLAG-V protein (Fig. 2A and B). In agreement with previously published results, STAT1 was found to interact with Nipah virus V amino acids 100 to 160, and STAT2 binding was confirmed for residues 100 to 300, with the fragment at residues 220 to 456 retaining a partial STAT2 association (37). Endogenous PLK1 was detected in the exact same pattern as STAT1, mapping its interaction to Nipah virus V protein residues 100 to 160 (Fig. 2A and B). This finding revealed a shared 60-amino-acid region at residues 100 to 160 that is necessary and sufficient for both PLK1 and STAT1 interactions with the Nipah virus V protein (Fig. 1A). The PLK1 PBD mediates interactions with both Henipavirus V proteins. PLK1 is a member of the polo kinase family characterized by a bipartite domain structure. The N-terminal 344



 Selected residues of the")








")



![32 -P]ATP, and reaction](/docs-images/93/113468441/images/6-35.jpg "mixtures were resolved by")












































6 6264 LUDLOW ET AL. J. VIROL. FIG. 4. Nipah virus V protein PLK1 interaction and phosphorylation require a consensus motif surrounding serine 130. (A) Selected residues of the consensus TSSP motif were mutated as indicated. (B) The TSSP motif is necessary for PLK1 binding to the Nipah virus V (NiV) protein. 293T cells were transfected to express FLAG-tagged Nipah virus V, the GFP control, or Nipah virus V point mutations, followed by FLAG immunoprecipitation and elution with FLAG peptide. The FLAG eluate and lysate samples were analyzed by immunoblotting using antibodies to detect associated PLK1 or FLAG-V protein. (C) Contact-dependent phosphorylation of Nipah virus V. 293T cells were transfected to express FLAG-tagged Nipah virus V, the GFP control, or Nipah virus V mutations, followed by FLAG immunoprecipitation and elution. Immune complexes were incubated in the presence of [ 32 -P]ATP, and reaction mixtures were resolved by SDS-PAGE and visualized by autoradiography. The FLAG eluate was further assessed by immunoblotting using antibodies detecting endogenous PLK1 and FLAG-V protein. *, immunoglobulin G heavy chain. amino acids form the catalytic kinase domain, while the C- terminal 259 amino acids form the PBD (Fig. 3A). This bipartite structure is involved in substrate selection and kinase activation. Current evidence supports a model that PLK1 is inactive in the steady state, but substrate proteins containing a phosphopeptide PBD binding motif trigger a kinase association. Proteins containing the consensus sequence Ser-[Ser/ Thr]-Pro, or SSP, are phosphorylated by priming kinase activity, converting the motif to Ser-[pSer/pThr]-Pro, which represents a high-affinity PBD recognition sequence. When the PBD binds the substrate, the kinase domain is activated to phosphorylate cis or trans targets (44). To test if the V protein association is mediated by the PBD, two PLK1 fragments representing the kinase domain (amino acids 1 to 344) or the PBD (amino acids 345 to 603) were expressed along with the Henipavirus V proteins (Fig. 3B). The V proteins effectively precipitated both full-length and PBD fragments but not the kinase domain fragment (Fig. 3B). These results confirm the importance of the PBD in directing the PLK1 association with the V proteins. Structural studies indicated that the specific coordination of the phosphopeptide motif relies on PBD residues W414, K540, and H538 (5, 12, 14). The W414F, K540M, and H538A mutations are known to abolish PLK1 binding to Cdc25C (12, 14). Immunoprecipitation was carried out as described above using the PBD W414F, K540M, and H538A mutations and the kinase-inactive N terminus K82R mutant as a control (40) (Fig. 3A). As expected, Henipavirus V proteins bound well to PLK1 and the K82R N-terminal kinase domain mutation. In contrast, all the PBD mutations resulted in defective PLK1 recognition (Fig. 3C). Together, these findings implicate the PBD as being the V protein interaction module. Distinct PLK1 binding motifs mediate Nipah virus and Hendra virus V protein association and phosphorylation. The PBD specifically binds to phosphopeptide motifs, and inspection of the 60-amino-acid STAT1 binding domain identified an SSP motif at residues 130 to 132 of both Henipavirus V proteins (Fig. 2C). To test the importance of this motif in PLK1 recruitment, selected amino acids were substituted with putative inactivating residues (Fig. 4Aand 5A) based on mutations described previously (11). The conserved serine at position 130 was mutated to both threonine, to preserve character, and alanine, and serine 131 was converted to tyrosine and to alanine. In addition, residue 129, which lies outside the SSP motif, was converted from threonine (in Nipah virus) or serine (in Hendra virus) to alanine. FLAG-tagged wild-type and Henipavirus V proteins containing mutations or control FLAG-GFP were expressed in 293T



 T200 is required for PLK1 binding to the Hendra virus V protein.")

. Therefore, the peptide 129 TSSP 132 is essential for PLK1 binding to the Nipah virus V protein.")




.")


7 VOL. 82, 2008 HENIPAVIRUS V PROTEIN PHOSPHORYLATION BY PLK FIG. 5. Hendra virus V protein PLK1 interaction and phosphorylation require an alternate consensus motif surrounding threonine 200. (A) Selected residues of the conserved SSSP motif were mutated as indicated. (B) Hendra virus V (HeV) protein mutations retain PLK1 binding. 293T cells were transfected to express FLAG-tagged Hendra virus V, the GFP control, or Hendra virus V point mutations and analyzed as described in the legend of Fig. 4. (C) Hendra virus V mutations retain phosphorylation. Immune complex kinase assays were carried out as described in the legend of Fig. 4. (D) An STP motif that is not conserved with Nipah virus surrounding T200 in the Hendra virus V protein mediates the PLK1 interaction. T200 was mutated to A as indicated. (E) T200 is required for PLK1 binding to the Hendra virus V protein. Proteins were expressed and analyzed for PLK1 associations and phosphorylation as described above. cells, followed by FLAG immunoprecipitation. As expected, wild-type Nipah virus V protein bound PLK1, but all mutations including threonine 129 strikingly abolished the ability of PLK1 to bind the Nipah virus V protein (Fig. 4B). Therefore, the peptide 129 TSSP 132 is essential for PLK1 binding to the Nipah virus V protein. The phosphorylation statuses of the substrate recognition motif mutations were tested by an immune complex kinase assay (Fig. 4C). Briefly, 293T cells were transfected with FLAG-tagged V, point mutations, and control GFP, followed by FLAG immunoprecipitation and elution with FLAG peptide. Immune complexes were incubated in the presence of [ 32 -P]ATP, and reaction mixtures were resolved by SDS-PAGE and visualized by autoradiography. The FLAG eluate was further assessed by immunoblotting using antibodies to detect endogenous PLK1 and FLAG-V protein. Wildtype Nipah virus V protein was phosphorylated, while all the TSSP motif mutations that failed to bind PLK1 were not phosphorylated (Fig. 4C). Therefore, the phosphorylation of Nipah virus V by PLK1 is contact dependent. The identical experiments carried out with the Hendra virus V protein mutations to the peptide 129 SSSP 132 (Fig. 5A) resulted in a dramatically different phenotype (Fig. 5B). None of the mutations in the Hendra virus V motif were deleterious for the PLK1 interaction (Fig. 5B). Therefore, the analogous SSPcontaining peptide is not the PLK1 recruitment motif for the Hendra virus V protein. In agreement with this, wild-type Hendra virus V protein and all mutations retained phosphorylation in the immune complex kinase assay (Fig. 5C). In conclusion, the two Henipavirus V proteins differ in their modes of PLK1 recruitment. Identification of the Hendra virus V protein PLK1 recruitment motif. Inspection of the Hendra virus V protein sequence identified a distinct consensus motif, 199 STP 201, that was not present in the Nipah virus V protein (Fig. 2C and 5D). To test this motif as a potential PLK1 recruitment site, mutagenesis was performed. Threonine 200 was converted to alanine, and PLK1 binding and phosphorylation were tested (Fig. 5E). As expected, wild-type Hendra virus V protein bound PLK1, but the mutation abolished the interaction. Thus, threonine 200 is required for PLK1 binding to the Hendra virus V protein. Accordingly, in immune complex kinase assays, wild-type Hendra virus V was phosphorylated, but the Hendra virus V T200A mutation was not (Fig. 5E). Separation of V-PLK1 and V-STAT complexes. The ability of the Nipah virus V protein to associate with both PLK1 and STAT1 via a single 60-amino-acid region invited inquiry into
 PLK1 does not associate with STAT1 and STAT2 in the absence of V protein.")
, and the supernatant following immunoprecipitation (I.")
 The PLK1 and V protein interactions are independent of STAT1 or STAT2.")

, Hendra virus V (HeV), or GFP, followed by")
.")



, and the supernatant following immunoprecipitation were evaluated by immunoblotting using")







8 6266 LUDLOW ET AL. J. VIROL. FIG. 6. Independence of STAT and PLK1 complexes. (A) PLK1 does not associate with STAT1 and STAT2 in the absence of V protein. 293T cells were transfected to express HA-tagged PLK1, followed by HA immunoprecipitation and elution. The lysate, sequential HA peptide eluates (E1 and E2), and the supernatant following immunoprecipitation (I.P) were assessed by immunoblotting to detect HA-PLK1, STAT1, or STAT2. (B) The PLK1 and V protein interactions are independent of STAT1 or STAT2. Parental 2fTGH cells, STAT1-deficient U3A, or STAT2-deficient U6A derivatives were transfected to express FLAG-tagged V proteins or GFP, followed by FLAG immunoprecipitation. Immune complexes and lysates were analyzed by immunoblotting to detect PLK1, STAT1, STAT2, and FLAG-V. (C) V:STAT and V:PLK1 exist as distinct complexes. 293T cells were transfected to express HA-PLK1 together with either FLAG-tagged Nipah virus V (NiV), Hendra virus V (HeV), or GFP, followed by FLAG immunoprecipitation and elution with FLAG peptide (FLAG eluate). The FLAG immune complexes were then reimmunoprecipitated with HA affinity gel and eluted using HA peptide (HA eluate). Lysate (input), FLAG, and HA eluates were assessed by immunoblot analysis to detect endogenous PLK1, STAT1, or STAT2 and then reprobed to detect HA-PLK1 or FLAG-V. the relationship between the two cellular proteins and their interactions with the V protein. First, the V protein could be coprecipitating an extant PLK1-STAT complex from the cell. Second, V could nucleate the formation of a higher-order complex including both PLK1 and STAT1. Third, the V protein could assemble with PLK1 and STAT1 independently. To test the possibility that PLK1 might be able to associate with STAT1 in the absence of the V protein, 293T cells were transfected with HA-tagged PLK1 followed by HA immunoprecipitation and peptide elution (Fig. 6A). The lysate, sequential HA eluates (E1 and E2), and the supernatant following immunoprecipitation were evaluated by immunoblotting using antibodies detecting HA-PLK1, STAT1, and STAT2. PLK1 did not copurify with STAT1 and STAT2 in the absence of V protein expression, suggesting that there is little or no preformed PLK1-STAT association. To confirm that the V-PLK1 interaction occurred independently of STAT1 and STAT2, parental human fibrosarcoma 2fTGH and derivative cells lines with single gene defects in STAT1, U3A (29), or STAT2, U6A (25), were utilized (Fig. 6B). Cells were transfected with FLAG-tagged Henipavirus V protein vectors or GFP followed by FLAG immunoprecipitation. Immune complexes and lysates were analyzed to detect PLK1, STAT1, and STAT2. In parental 2fTGH cells, Henipavirus V protein copurified with endogenous PLK1, and binding patterns for STAT1 and STAT2 matched previous findings; in particular, we confirmed that Henipavirus V proteins require cellular STAT1 to interact with STAT2 (37). Notably, in the absence of STAT1 or STAT2, V proteins retained the ability to copurify endogenous PLK1. These results indicate that PLK1:V interactions do not require STAT proteins. To establish if the V-induced association with STATs and PLK1 produces a single tripartite complex, a sequential immunoprecipitation assay was used. HA-tagged PLK1 was expressed together with FLAG-tagged Henipavirus V proteins or GFP followed by FLAG immunoprecipitation and elution with FLAG peptide. The FLAG eluates were then reimmunoprecipitated with HA affinity gel and eluted using HA peptide. Lysate, FLAG eluate, and HA eluate fractions were assessed by immunoblot analysis to detect STAT1, STAT2, or PLK1 (Fig. 6C). No background signals were observed with control FLAG-GFP. Henipavirus V proteins copurified with endogenous STAT1, STAT2, and HA-tagged PLK1, but reimmunoprecipitation with HA affinity gel separated HA-PLK1 from STAT1 and STAT2. This result demonstrates the segregation of the V:STAT and V:PLK1 complexes. Henipavirus SSP motifs are involved in STAT1 binding. The location of the PLK1 binding motif in the center of the STAT1 binding region of the Nipah virus V protein suggests a conserved function that might be related to IFN evasion. To evaluate the effects of the Henipavirus SSP motif mutations on STAT1 and STAT2 interactions, eluates from FLAG-V immunoprecipitations were analyzed by immunoblotting with antisera for STAT1 or STAT2 (Fig. 7A). As expected, wild-type Henipavirus V proteins bound STAT1 and STAT2, but some of the proteins containing mutations were defective for STAT interactions. The conservative substitution of serine with threonine at position 130 did not affect STAT binding, but the substitution to alanine significantly reduced the interaction. Changing Nipah virus V protein T129 or its Hendra virus counterpart S129 to alanine did not affect STAT binding. In contrast, the substitution of S131 with tyrosine or alanine dra-





 (i) or Hendra virus V (HeV)")



 Control analysis of the")




, point mutations, or an")

")


 the standard")








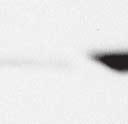



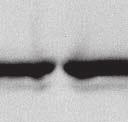
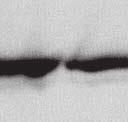








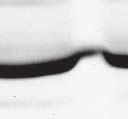

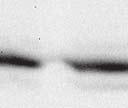
9 VOL. 82, 2008 HENIPAVIRUS V PROTEIN PHOSPHORYLATION BY PLK FIG. 7. The conserved T/S SSP motif is necessary for interaction with STAT1 and STAT2. (A) T/S SSP motifs are required for STAT1 and STAT2 V protein interactions. 293T cells were transfected to express FLAG-tagged Nipah virus V (NiV) (i) or Hendra virus V (HeV) (ii), followed by FLAG immunoprecipitation and elution. The FLAG eluate and lysate samples were analyzed by immunoblotting using antibodies to detect endogenous STAT1, STAT2, or FLAG-V protein. (iii) Control analysis of the FLAG-Hendra virus V T200A point mutation. (B) IFN signaling inhibition correlates with STAT1 and STAT2 binding abilities. 2fTGH cells were transfected with the GAS- or ISRE-luciferase (Luc) reporter gene and Nipah virus V (NiV) (i), Hendra virus V (HeV) (ii), point mutations, or an empty vector (CON), as indicated. Cells were stimulated with IFN- (5 ng/ml) or IFN- (1,000 U/ml) for 10 h or not stimulated ( ) prior to lysis and luciferase assays. Data were normalized to cotransfected Renilla luciferase, and the bars indicate the averages (n 3) the standard deviations. matically reduced the coprecipitation of STATs. Clearly, the serine residues at positions 130 and 131 in both Henipavirus V proteins contribute to the STAT association. In addition, the T200A substitution in Hendra virus V protein that eliminated the PLK1 association did not influence the STAT1 interaction (Fig. 7Aiii), a finding that is in agreement with the mapping of the core STAT binding region defined for the Nipah virus V protein (37) (Fig. 2C). Henipavirus SSP motifs are required for STAT1 relocalization and IFN signaling evasion. To verify these results in a biological context, the abilities of the mutated proteins to prevent IFN signal transduction were tested in luciferase assays (Fig. 7B). Wild-type Henipavirus V proteins prevented the induction of STAT-dependent IFN- -responsive and IFN- -responsive reporter genes. IFN signaling inhibition correlated exactly with STAT binding: only those V proteins that bound STATs were able to block IFN signaling. In addition, the Hendra virus V T200A substitution mutant that retained the STAT interaction also inhibited IFN signaling (not shown). The mechanism of V protein inhibition involves the retention of STATs in the cytoplasm. To confirm the consequences of the STAT interaction, indirect immunofluorescence was performed using 2fTGH cells transfected with FLAG-tagged Nipah virus V and the S130T, S130A, and S131Y point mutations followed by IFN- treatment to induce STAT1 translocation and accumulation in the nucleus (Fig. 8A). Both wildtype Nipah virus V and the S130T mutation retained STAT1 in the cytoplasm. In contrast, no STAT1 interference was detected in cells that expressed the S130A and S131Y mutations, as STAT1 still accumulated in the nucleus. To verify these results in the biologically meaningful context of virus infection, an antiviral assay was performed (Fig. 8B and C). Briefly, pools of 2fTGH cells expressing the control pef-flag plasmid, wild-type V protein, or point mutations were treated with IFN- or medium alone and then infected with a reporter virus, VSV-GFP. Approximately 90% of untreated cells became infected with VSV-GFP, but treatment with IFN- resulted in cellular resistance to VSV infection (Fig. 8B and C). Wild-type V proteins and all variants capable of binding to STAT1 prevented the antiviral state, but the S130A, S131Y, and S131A mutations that failed to inactivate the STAT proteins did not (Fig. 8B and Cii).
10 6268 LUDLOW ET AL. J. VIROL. FIG. 8. The conserved T/S SSP motif is essential for STAT1 relocalization and IFN signaling evasion. (A) A subset of mutated V proteins fail to retain STAT1 in the cytoplasm. 2fTGH cells were transfected with FLAG-tagged Nipah virus V (NiV) and point mutations, followed by indirect immunofluorescence to detect FLAG-V and STAT1. WT, wild type. (B) V proteins that fail to bind STAT1 do not disrupt the IFN antiviral state. Pools of 2fTGH cells stably expressing Nipah virus V protein, the Hendra virus V protein (HeV), phosphopeptide binding motif mutations, and the pef-flag control were treated with medium alone or IFN- for 8 h, followed by VSV-GFP infection. (C) After 22 h, GFP levels were analyzed by flow cytometry, and the percentage of cells expressing GFP was calculated in comparison to uninfected 2fTGH cells. N/A, not applicable.
11 VOL. 82, 2008 HENIPAVIRUS V PROTEIN PHOSPHORYLATION BY PLK FIG. 9. Interaction of STAT1 and PLK1 is dispensable for P protein-dependent Nipah virus RNA synthesis. BHK/sr/T7 cells were transfected to express Nipah virus N, Nipah virus P (or Nipah virus P point mutations), Nipah virus L, and the Nipah virus CAT minigenome plasmid. For the negative control (CON), the Nipah virus L plasmid was replaced with an equivalent amount of ptm1 empty vector. The assay was performed three separate times in which each sample was assayed in duplicate. Minigenome replication was assessed using a CAT ELISA system as described previously (16). WT, wild type. STAT1 and PLK1 interactions are dispensable for Nipah virus RNA transcription and replication. Our experimental procedures used Henipavirus V proteins to identify PLK1. However, the STAT1 and PLK1 binding region is shared among three proteins, P, V, and W. Both V and W have been referred to as accessory proteins required for host-suppressing actions and are not always required for virus replication in cell culture. In contrast, while expressed Nipah virus P protein can exhibit IFN signaling interference in vitro (42), it is better known to be an essential component of the viral RNA transcription and replication machinery (16). To study the effects of STAT1 or PLK1 binding deficiencies on the function of Nipah virus P protein, a Nipah virus minigenome replication assay that relies on the expression of the L, P, and NP proteins to replicate a CAT reporter gene was used (16). The assay was carried out with the wild type or TSSP mutations engineered in the context of the Nipah virus P protein. The substitution of mutated P proteins for the wild type did not alter replication activity, as all of the mutated proteins supported transcription and replication to the same extent as the wild-type P protein (Fig. 9). DISCUSSION PLK1 has essential roles in cell cycle regulation, controlling mitotic spindle formation and checkpoint responses to DNA damage. It has also been directly or indirectly linked to the replication of diverse viruses. For example, PLK1 interacts with and phosphorylates the human cytomegalovirus pp65 lower matrix protein and accompanies pp65 into budding particles (13). In addition, deregulated levels of PLK1 have been associated with simian immunodeficiency virus (4), human T- lymphotrophic virus type I p30 protein (9), and human papillomavirus type 16 E6 and E7 proteins (35). The identification of PLK1 as a Nipah virus and Hendra virus P, V, and W protein binding partner and the demonstration of interactiondependent V protein phosphorylation now connects this cellular Ser/Thr protein kinase to the emerging negative-strand RNA viruses in the paramyxovirus genus Henipavirus. As with other PLK targets, the mode of interaction with both Nipah virus and Hendra virus V proteins was found to involve the PBD-dependent recognition of Ser-[pSer/pThr]-Pro peptides. The identification of this motif is significant because the Henipavirus V proteins represent the first virally encoded proteins found to contain a functional PBD binding motif. However, several features that distinguish the two V proteins were identified. For Nipah virus, it was found that PLK1 interacts with a Ser-Ser-Pro motif surrounding serine 130. Despite the absolute conservation of the core PLK1 triad of Ser-Ser-Pro between Nipah virus and Hendra virus V proteins, the ability of this peptide to mediate the interaction with PLK1 was not conserved. Instead, the Hendra virus V protein interaction with PLK1 was conferred by another consensus site, Ser-Thr-Pro, surrounding threonine 200 (Table 1). In both cases, the integrity of the sequence motif is essential for PLK1-dependent V protein phosphorylation. To account for the different binding sites, we speculate that differences in flanking sequences may determine the association with a V protein-specific priming kinase(s) used to create the essential pser/pthr central to PBD recognition. One clue that may aid in the identification of the priming kinase is the observed critical importance, in the Nipah virus V protein, of Thr129 in the 2 position (with respect to the pser). When Thr129 is substituted, the mutated Nipah virus V protein no longer recruits or gets phosphorylated by PLK1. The potential involvement of multiple or diverse priming kinases may not be implausible, as the Henipavirus P and V proteins are well known to be highly phosphorylated (43). In fact, the entire V protein N terminus, comprising over 75% of the sequence, including the PLK1 binding sites, is shared with the viral phosphoprotein, P. Our molecular studies demonstrate interaction-dependent phosphorylation by PLK1, suggesting that the P, V, or W protein could represent a PLK1 substrate in vivo. However, the data indicate that the ability to associate with PLK1 does not alter the P protein s function in RNA synthesis, as Nipah virus P proteins lacking the TSSP consensus were competent for minigenome replication. No alteration in cell cycle regulation was observed following the overexpression and stable expression of the Henipavirus P, V, or W protein and mutations defective for PLK1 binding (L. E. Ludlow and C. M. Horvath, unpublished observations). Analysis of these mutations in the context of virus infection is crucial to dissect the contribution of this interaction in the host TABLE 1. Properties of proteins containing T/S SSP mutations Mutation Nipah virus V Consequence of mutation a PLK1 STAT1 Replicon Hendra virus V Nipah virus V Hendra virus V Nipah virus P Wild type T/S129A S130T S130A S131Y S131A T200A ND ND ND a ND, not determined.
12 6270 LUDLOW ET AL. J. VIROL. cells under natural conditions of infection. Determination of the precise role(s) for PLK1 during Henipavirus infections will require further studies using recombinant viruses grown under biosafety level 4 containment. Equally interesting implications of this study relate to the observation that the Nipah virus V protein PLK1 interaction motif lies in the center of the previously characterized STAT1 interaction site. The interaction with STAT1 causes a cellular defect in IFN-mediated transcription and antiviral effects and is therefore fundamental to innate immune interference mediated by both Nipah virus and Hendra virus. Evaluation of STAT binding to Hendra virus and Nipah virus V protein (T/S)SSP motif mutations revealed that Ser130 and Ser131 are both needed for binding to STAT1 and STAT2 and normal IFN signaling interference. Interestingly, the S130T mutant retained the ability to bind STATs, but the S130A mutant failed to bind STAT1 or block IFN signaling, revealing specific side-chain preferences for Henipavirus interference. Further insight was gained by examining PLK1 and STAT binding properties. PLK1 was not detected in association with STAT1 or STAT2, and the STATs were not required for V:PLK1 interactions. In agreement with this finding, the V:STAT and V:PLK1 subcomplexes exist independently within the cell. The ability to interfere with host defense pathways is paramount for virus pathogenesis, and we previously speculated that the ablation of Henipavirus IFN signaling evasion would produce an attenuated virus and therefore a plausible vaccine candidate (38, 39). However, the V protein region mediating interactions with STAT1 is also present in the P protein, which is essential for viral RNA synthesis and must be functional to create a recombinant virus. S130 and S131 mutations represent attenuating point mutations, as these mutations are capable of dissociating V:STAT interactions. The analysis of P proteins containing these STAT1 and PLK1 binding mutations revealed no difference in their abilities to support minigenome transcription and replication. These findings reveal that it is indeed possible to dissociate the IFN signaling evasion functions of the V protein from the RNA synthesis functions of the P protein and specifically indicate that mutations of S130 or S131 are likely to generate viable viruses. We predict that these viruses would be attenuated due to the inability of the V or W protein to disrupt host cell STAT-dependent IFN antiviral immune responses and would represent vaccine candidates. This study has improved our understanding of the virus-host interface and Henipavirus pathogenesis, providing information for the design of therapeutic or antiviral reagents. ACKNOWLEDGMENTS We are grateful to members of the Horvath laboratory for helpful discussion and technical advice. Special acknowledgment goes to Lily Garza for expert technical support. We also thank Eric Weiss and Jennifer Brace for discussions and reading of the manuscript. Mass spectrometry-based protein identification was carried out by Alexander Schilling and Bao-Shang Lee at the CBC-RRC Proteomics Core Laboratory (University of Illinois at Chicago). We acknowledge William Russin at the Biological Imaging Research Facility, Northwestern University. This research was supported by NIH grant R01 AI to C.M.H. REFERENCES 1. Aaronson, D. S., and C. M. Horvath A road map for those who don t know JAK-STAT. Science 296: Andrejeva, J., K. S. Childs, D. F. Young, T. S. Carlos, N. Stock, S. Goodbourn, and R. E. Randall The V proteins of paramyxoviruses bind the IFN-inducible RNA helicase, mda-5, and inhibit its activation of the IFNbeta promoter. Proc. Natl. Acad. Sci. USA 101: Anonymous Hendra virus Australia (Queensland). ProMED archive number International Society for Infectious Diseases, Brookline, MA. 4. Bostik, P., G. L. Dodd, F. Villinger, A. E. Mayne, and A. A. Ansari Dysregulation of the polo-like kinase pathway in CD4 T cells is characteristic of pathogenic simian immunodeficiency virus infection. J. Virol. 78: a.Bradford, M. M A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72: Cheng, K. Y., E. D. Lowe, J. Sinclair, E. A. Nigg, and L. N. Johnson The crystal structure of the human polo-like kinase-1 polo box domain and its phospho-peptide complex. EMBO J. 22: Childs, K., N. Stock, C. Ross, J. Andrejeva, L. Hilton, M. Skinner, R. Randall, and S. Goodbourn mda-5, but not RIG-I, is a common target for paramyxovirus V proteins. Virology 359: Chua, K. B., W. J. Bellini, P. A. Rota, B. H. Harcourt, A. Tamin, S. K. Lam, T. G. Ksiazek, P. E. Rollin, S. R. Zaki, W. Shieh, C. S. Goldsmith, D. J. Gubler, J. T. Roehrig, B. Eaton, A. R. Gould, J. Olson, H. Field, P. Daniels, A. E. Ling, C. J. Peters, L. J. Anderson, and B. W. Mahy Nipah virus: a recently emergent deadly paramyxovirus. Science 288: Curran, J., R. Boeck, and D. Kolakofsky The Sendai virus P gene expresses both an essential protein and an inhibitor of RNA synthesis by shuffling modules via mrna editing. EMBO J. 10: Datta, A., L. Silverman, A. J. Phipps, H. Hiraragi, L. Ratner, and M. D. Lairmore Human T-lymphotropic virus type-1 p30 alters cell cycle G2 regulation of T lymphocytes to enhance cell survival. Retrovirology 4: Eaton, B. T., C. C. Broder, D. Middleton, and L. F. Wang Hendra and Nipah viruses: different and dangerous. Nat. Rev. Microbiol. 4: Elia, A. E., L. C. Cantley, and M. B. Yaffe Proteomic screen finds pser/pthr-binding domain localizing Plk1 to mitotic substrates. Science 299: Elia, A. E., P. Rellos, L. F. Haire, J. W. Chao, F. J. Ivins, K. Hoepker, D. Mohammad, L. C. Cantley, S. J. Smerdon, and M. B. Yaffe The molecular basis for phosphodependent substrate targeting and regulation of Plks by the Polo-box domain. Cell 115: Gallina, A., L. Simoncini, S. Garbelli, E. Percivalle, G. Pedrali-Noy, K. S. Lee, R. L. Erikson, B. Plachter, G. Gerna, and G. Milanesi Polo-like kinase 1 as a target for human cytomegalovirus pp65 lower matrix protein. J. Virol. 73: Garcia-Alvarez, B., G. de Carcer, S. Ibanez, E. Bragado-Nilsson, and G. Montoya Molecular and structural basis of polo-like kinase 1 substrate recognition: implications in centrosomal localization. Proc. Natl. Acad. Sci. USA 104: Goodbourn, S., L. Didcock, and R. E. Randall Interferons: cell signalling, immune modulation, antiviral response and virus countermeasures. J. Gen. Virol. 81: Halpin, K., B. Bankamp, B. H. Harcourt, W. J. Bellini, and P. A. Rota Nipah virus conforms to the rule of six in a minigenome replication assay. J. Gen. Virol. 85: Harcourt, B. H., L. Lowe, A. Tamin, X. Liu, B. Bankamp, N. Bowden, P. E. Rollin, J. A. Comer, T. G. Ksiazek, M. J. Hossain, E. S. Gurley, R. F. Breiman, W. J. Bellini, and P. A. Rota Genetic characterization of Nipah virus, Bangladesh, Emerg. Infect. Dis. 11: Horvath, C. M Silencing STATs: lessons from paramyxovirus interferon evasion. Cytok. Growth Factor Rev. 15: Horvath, C. M Weapons of STAT destruction. Interferon evasion by paramyxovirus V protein. Eur. J. Biochem. 271: Hsu, V. P., M. J. Hossain, U. D. Parashar, M. M. Ali, T. G. Ksiazek, I. Kuzmin, M. Niezgoda, C. Rupprecht, J. Bresee, and R. F. Breiman Nipah virus encephalitis reemergence, Bangladesh. Emerg. Infect. Dis. 10: Isaacs, A., and J. Lindenmann Virus interference. I. The interferon. Proc. R. Soc. Lond. B Biol. Sci. 147: Kawai, T., and S. Akira TLR signaling. Cell Death Differ. 13: Lam, S. K Nipah virus a potential agent of bioterrorism? Antivir. Res. 57: Lee, B Envelope-receptor interactions in Nipah virus pathobiology. Ann. N. Y. Acad. Sci. 1102: Leung, S., S. A. Qureshi, I. M. Kerr, J. E. Darnell, Jr., and G. R. Stark Role of STAT2 in the alpha interferon signaling pathway. Mol. Cell. Biol. 15: Liston, P., and D. J. Briedis Measles virus V protein binds zinc. Virology 198: Luby, S. P., M. Rahman, M. J. Hossain, L. S. Blum, M. M. Husain, E. Gurley, R. Khan, B. N. Ahmed, S. Rahman, N. Nahar, E. Kenah, J. A. Comer, and T. G. Ksiazek Foodborne transmission of Nipah virus, Bangladesh. Emerg. Infect. Dis. 12:
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature12234 Supplementary Figure 1. Embryonic naked mole-rat fibroblasts do not undergo ECI. Embryonic naked mole-rat fibroblasts ( EF) were isolated from eight mid-gestation embryos. All the
doi:10.1038/nature12234 Supplementary Figure 1. Embryonic naked mole-rat fibroblasts do not undergo ECI. Embryonic naked mole-rat fibroblasts ( EF) were isolated from eight mid-gestation embryos. All the
PREPARED FOR: U.S. Army Medical Research and Materiel Command Fort Detrick, Maryland X Approved for public release; distribution unlimited
 Award Number: W8XWH--- TITLE: Defining the Role of Autophagy Kinase ULK Signaling in Therapeutic Response of Tuberous Sclerosis Complex to Inhibitors PRINCIPAL INVESTIGATOR: Reuben J. Shaw, Ph.D. CONTRACTING
Award Number: W8XWH--- TITLE: Defining the Role of Autophagy Kinase ULK Signaling in Therapeutic Response of Tuberous Sclerosis Complex to Inhibitors PRINCIPAL INVESTIGATOR: Reuben J. Shaw, Ph.D. CONTRACTING
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
The color and patterning of pigmentation in cats, dogs, mice horses and other mammals results from the interaction of several different genes
 The color and patterning of pigmentation in cats, dogs, mice horses and other mammals results from the interaction of several different genes 1 Gene Interactions: Specific alleles of one gene mask or modify
The color and patterning of pigmentation in cats, dogs, mice horses and other mammals results from the interaction of several different genes 1 Gene Interactions: Specific alleles of one gene mask or modify
Gliding Motility Assay for P. berghei Sporozoites
 Gliding Motility Assay for P. berghei Sporozoites Important Notes: 1. For all dilutions (including antibodies and sporozoites), always make slightly more than needed. For instance, if you need 200 µl sporozoites
Gliding Motility Assay for P. berghei Sporozoites Important Notes: 1. For all dilutions (including antibodies and sporozoites), always make slightly more than needed. For instance, if you need 200 µl sporozoites
Burton's Microbiology for the Health Sciences. Chapter 9. Controlling Microbial Growth in Vivo Using Antimicrobial Agents
 Burton's Microbiology for the Health Sciences Chapter 9. Controlling Microbial Growth in Vivo Using Antimicrobial Agents Chapter 9 Outline Introduction Characteristics of an Ideal Antimicrobial Agent How
Burton's Microbiology for the Health Sciences Chapter 9. Controlling Microbial Growth in Vivo Using Antimicrobial Agents Chapter 9 Outline Introduction Characteristics of an Ideal Antimicrobial Agent How
Medical Genetics and Diagnosis Lab #3. Gel electrophoresis
 Medical Genetics and Diagnosis Lab #3 Gel electrophoresis Background Information Gel electrophoresis is the standard lab procedure for separating DNA by size (e.g. length in base pairs) for visualization
Medical Genetics and Diagnosis Lab #3 Gel electrophoresis Background Information Gel electrophoresis is the standard lab procedure for separating DNA by size (e.g. length in base pairs) for visualization
Veterinary Diagnostics Portfolio Overview. Complete solutions for veterinary testing and pathogen research
 Veterinary Diagnostics Portfolio Overview Complete solutions for veterinary testing and pathogen research Sample preparation products Cat. no. (number of preps) Target analyte Product Short description
Veterinary Diagnostics Portfolio Overview Complete solutions for veterinary testing and pathogen research Sample preparation products Cat. no. (number of preps) Target analyte Product Short description
Was the Spotted Horse an Imaginary Creature? g.org/sciencenow/2011/11/was-the-spotted-horse-an-imagina.html
 Was the Spotted Horse an Imaginary Creature? http://news.sciencema g.org/sciencenow/2011/11/was-the-spotted-horse-an-imagina.html 1 Genotypes of predomestic horses match phenotypes painted in Paleolithic
Was the Spotted Horse an Imaginary Creature? http://news.sciencema g.org/sciencenow/2011/11/was-the-spotted-horse-an-imagina.html 1 Genotypes of predomestic horses match phenotypes painted in Paleolithic
Consequences of Antimicrobial Resistant Bacteria. Antimicrobial Resistance. Molecular Genetics of Antimicrobial Resistance. Topics to be Covered
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
MID 23. Antimicrobial Resistance. Consequences of Antimicrobial Resistant Bacteria. Molecular Genetics of Antimicrobial Resistance
 Antimicrobial Resistance Molecular Genetics of Antimicrobial Resistance Micro evolutionary change - point mutations Beta-lactamase mutation extends spectrum of the enzyme rpob gene (RNA polymerase) mutation
Antimicrobial Resistance Molecular Genetics of Antimicrobial Resistance Micro evolutionary change - point mutations Beta-lactamase mutation extends spectrum of the enzyme rpob gene (RNA polymerase) mutation
Antimicrobial Resistance
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Acquisition of Foreign DNA
 Antimicrobial Resistance Acquisition of Foreign DNA Levy, Scientific American Horizontal gene transfer is common, even between Gram positive and negative bacteria Plasmid - transfer of single or multiple
Antimicrobial Resistance Acquisition of Foreign DNA Levy, Scientific American Horizontal gene transfer is common, even between Gram positive and negative bacteria Plasmid - transfer of single or multiple
Antibiotic Resistance in Bacteria
 Antibiotic Resistance in Bacteria Electron Micrograph of E. Coli Diseases Caused by Bacteria 1928 1 2 Fleming 3 discovers penicillin the first antibiotic. Some Clinically Important Antibiotics Antibiotic
Antibiotic Resistance in Bacteria Electron Micrograph of E. Coli Diseases Caused by Bacteria 1928 1 2 Fleming 3 discovers penicillin the first antibiotic. Some Clinically Important Antibiotics Antibiotic
Enzootic Bovine Leukosis: Milk Screening and Verification ELISA: VF-P02210 & VF-P02220
 Enzootic Bovine Leukosis: Milk Screening and Verification ELISA: VF-P02210 & VF-P02220 Introduction Enzootic Bovine Leukosis is a transmissible disease caused by the Enzootic Bovine Leukosis Virus (BLV)
Enzootic Bovine Leukosis: Milk Screening and Verification ELISA: VF-P02210 & VF-P02220 Introduction Enzootic Bovine Leukosis is a transmissible disease caused by the Enzootic Bovine Leukosis Virus (BLV)
Selective toxicity. Antimicrobial Drugs. Alexander Fleming 10/17/2016
 Selective toxicity Antimicrobial Drugs Chapter 20 BIO 220 Drugs must work inside the host and harm the infective pathogens, but not the host Antibiotics are compounds produced by fungi or bacteria that
Selective toxicity Antimicrobial Drugs Chapter 20 BIO 220 Drugs must work inside the host and harm the infective pathogens, but not the host Antibiotics are compounds produced by fungi or bacteria that
EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS WORK-PROGRAMME PROPOSAL Version 2 VISAVET. Universidad Complutense de Madrid
 FOR BOVINE TUBERCULOSIS WORK-PROGRAMME PROPOSAL Version 2 VISAVET. Universidad Complutense de Madrid") EUROPEAN COMMISSION HEALTH & CONSUMERS DIRECTORATE-GENERAL Directorate D Animal Health and Welfare Unit D1- Animal health and Standing Committees EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS
EUROPEAN COMMISSION HEALTH & CONSUMERS DIRECTORATE-GENERAL Directorate D Animal Health and Welfare Unit D1- Animal health and Standing Committees EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS
Antibiotics & Resistance
 What are antibiotics? Antibiotics & esistance Antibiotics are molecules that stop bacteria from growing or kill them Antibiotics, agents against life - either natural or synthetic chemicals - designed
What are antibiotics? Antibiotics & esistance Antibiotics are molecules that stop bacteria from growing or kill them Antibiotics, agents against life - either natural or synthetic chemicals - designed
How to load and run an Agarose gel PSR
 How to load and run an Agarose gel PSR Agarose gel electrophoresis is the most effective way of separating DNA fragments of varying sizes ranging from100 bp to 25 kb. This protocol divided into three stages:
How to load and run an Agarose gel PSR Agarose gel electrophoresis is the most effective way of separating DNA fragments of varying sizes ranging from100 bp to 25 kb. This protocol divided into three stages:
Inhibiting Microbial Growth in vivo. CLS 212: Medical Microbiology Zeina Alkudmani
 Inhibiting Microbial Growth in vivo CLS 212: Medical Microbiology Zeina Alkudmani Chemotherapy Definitions The use of any chemical (drug) to treat any disease or condition. Chemotherapeutic Agent Any drug
Inhibiting Microbial Growth in vivo CLS 212: Medical Microbiology Zeina Alkudmani Chemotherapy Definitions The use of any chemical (drug) to treat any disease or condition. Chemotherapeutic Agent Any drug
Agarose Blenders. Code Description Size
 Agarose Blenders Code Description Size K669-100G Agarose I / TBE Blend 0.8% 100 grams K677-100G Agarose I / TBE Blend 1.5% 100 grams K678-100G Agarose I /TBE Blend 2.0% 100 grams K679-100G Agarose I /
Agarose Blenders Code Description Size K669-100G Agarose I / TBE Blend 0.8% 100 grams K677-100G Agarose I / TBE Blend 1.5% 100 grams K678-100G Agarose I /TBE Blend 2.0% 100 grams K679-100G Agarose I /
PCR detection of Leptospira in. stray cat and
 PCR detection of Leptospira in 1 Department of Pathology, School of Veterinary Medicine, Islamic Azad University, Shahrekord Branch, Shahrekord, Iran 2 Department of Microbiology, School of Veterinary
PCR detection of Leptospira in 1 Department of Pathology, School of Veterinary Medicine, Islamic Azad University, Shahrekord Branch, Shahrekord, Iran 2 Department of Microbiology, School of Veterinary
Lecture 6: Fungi, antibiotics and bacterial infections. Outline Eukaryotes and Prokaryotes Viruses Bacteria Antibiotics Antibiotic resistance
 Lecture 6: Fungi, antibiotics and bacterial infections Outline Eukaryotes and Prokaryotes Viruses Bacteria Antibiotics Antibiotic resistance Lecture 1 2 3 Lecture Outline Section 4 Willow and aspirin Opium
Lecture 6: Fungi, antibiotics and bacterial infections Outline Eukaryotes and Prokaryotes Viruses Bacteria Antibiotics Antibiotic resistance Lecture 1 2 3 Lecture Outline Section 4 Willow and aspirin Opium
Challenges Emerging resistance Fewer new drugs MRSA and other resistant pathogens are major problems
 Micro 301 Antimicrobial Drugs 11/7/12 Significance of antimicrobial drugs Challenges Emerging resistance Fewer new drugs MRSA and other resistant pathogens are major problems Definitions Antibiotic Selective
Micro 301 Antimicrobial Drugs 11/7/12 Significance of antimicrobial drugs Challenges Emerging resistance Fewer new drugs MRSA and other resistant pathogens are major problems Definitions Antibiotic Selective
Activation of the vrg6 Promoter of Bordetella pertussis by RisA
 JOURNAL OF BACTERIOLOGY, Mar. 2005, p. 1648 1658 Vol. 187, No. 5 0021-9193/05/$08.00 0 doi:10.1128/jb.187.5.1648 1658.2005 Activation of the vrg6 Promoter of Bordetella pertussis by RisA Tadhg Ó Cróinín,
JOURNAL OF BACTERIOLOGY, Mar. 2005, p. 1648 1658 Vol. 187, No. 5 0021-9193/05/$08.00 0 doi:10.1128/jb.187.5.1648 1658.2005 Activation of the vrg6 Promoter of Bordetella pertussis by RisA Tadhg Ó Cróinín,
Chapter concepts: What are antibiotics, the different types, and how do they work? Antibiotics
 Chapter concepts: Antibiotics What are antibiotics, the different types, and how do they work? How do we decided on the most appropriate antibiotic treatment? What are some of the ways that bacteria are
Chapter concepts: Antibiotics What are antibiotics, the different types, and how do they work? How do we decided on the most appropriate antibiotic treatment? What are some of the ways that bacteria are
Antimicrobial Therapy
 Chapter 12 The Elements of Chemotherapy Topics - Antimicrobial Therapy - Selective Toxicity - Survey of Antimicrobial Drug - Microbial Drug Resistance - Drug and Host Interaction Antimicrobial Therapy
Chapter 12 The Elements of Chemotherapy Topics - Antimicrobial Therapy - Selective Toxicity - Survey of Antimicrobial Drug - Microbial Drug Resistance - Drug and Host Interaction Antimicrobial Therapy
PRINCIPAL INVESTIGATOR: Dr. Jetsumon (Sattabongkot) Prachumsri
 Prachumsri") AD (Leave blank) Award Number: W81XWH-07-2-0090 TITLE: Proteomic Study of Human Malaria Parasite Plasmodium Vivax Liver Stages for Development of Vaccines and Drugs PRINCIPAL INVESTIGATOR: Dr. Jetsumon
AD (Leave blank) Award Number: W81XWH-07-2-0090 TITLE: Proteomic Study of Human Malaria Parasite Plasmodium Vivax Liver Stages for Development of Vaccines and Drugs PRINCIPAL INVESTIGATOR: Dr. Jetsumon
1. Division of Bacterial, Parasitic, and Allergenic Products, Center for Biologics Evaluation and
 JB Accepted Manuscript Posted Online 30 July 2018 J. Bacteriol. doi:10.1128/jb.00175-18 This is a work of the U.S. Government and is not subject to copyright protection in the United States. Foreign copyrights
JB Accepted Manuscript Posted Online 30 July 2018 J. Bacteriol. doi:10.1128/jb.00175-18 This is a work of the U.S. Government and is not subject to copyright protection in the United States. Foreign copyrights
Antimicrobial Resistance
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of Change in the approach to the administration of empiric antimicrobial therapy Increased
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of Change in the approach to the administration of empiric antimicrobial therapy Increased
BioSci 110, Fall 08 Exam 2
 1. is the cell division process that results in the production of a. mitosis; 2 gametes b. meiosis; 2 gametes c. meiosis; 2 somatic (body) cells d. mitosis; 4 somatic (body) cells e. *meiosis; 4 gametes
1. is the cell division process that results in the production of a. mitosis; 2 gametes b. meiosis; 2 gametes c. meiosis; 2 somatic (body) cells d. mitosis; 4 somatic (body) cells e. *meiosis; 4 gametes
A Unique Approach to Managing the Problem of Antibiotic Resistance
 A Unique Approach to Managing the Problem of Antibiotic Resistance By: Heather Storteboom and Sung-Chul Kim Department of Civil and Environmental Engineering Colorado State University A Quick Review The
A Unique Approach to Managing the Problem of Antibiotic Resistance By: Heather Storteboom and Sung-Chul Kim Department of Civil and Environmental Engineering Colorado State University A Quick Review The
How the eye sees. Properties of light. The light-gathering parts of the eye. 1. Properties of light. 2. The anatomy of the eye. 3.
 How the eye sees 1. Properties of light 2. The anatomy of the eye 3. Visual pigments 4. Color vision 1 Properties of light Light is made up of particles called photons Light travels as waves speed of light
How the eye sees 1. Properties of light 2. The anatomy of the eye 3. Visual pigments 4. Color vision 1 Properties of light Light is made up of particles called photons Light travels as waves speed of light
Vaccines for Cats. 2. Feline viral rhinotracheitis, FVR caused by FVR virus, also known as herpes virus type 1, FHV-1
 Vaccines for Cats Recent advances in veterinary medical science have resulted in an increase in the number and type of vaccines that are available for use in cats, and improvements are continuously being
Vaccines for Cats Recent advances in veterinary medical science have resulted in an increase in the number and type of vaccines that are available for use in cats, and improvements are continuously being
Mechanisms and Pathways of AMR in the environment
 FMM/RAS/298: Strengthening capacities, policies and national action plans on prudent and responsible use of antimicrobials in fisheries Final Workshop in cooperation with AVA Singapore and INFOFISH 12-14
FMM/RAS/298: Strengthening capacities, policies and national action plans on prudent and responsible use of antimicrobials in fisheries Final Workshop in cooperation with AVA Singapore and INFOFISH 12-14
Comparing DNA Sequences Cladogram Practice
 Name Period Assignment # See lecture questions 75, 122-123, 127, 137 Comparing DNA Sequences Cladogram Practice BACKGROUND Between 1990 2003, scientists working on an international research project known
Name Period Assignment # See lecture questions 75, 122-123, 127, 137 Comparing DNA Sequences Cladogram Practice BACKGROUND Between 1990 2003, scientists working on an international research project known
Approved by the Food Safety Commission on September 30, 2004
 Approved by the Food Safety Commission on September 30, 2004 Assessment guideline for the Effect of Food on Human Health Regarding Antimicrobial- Resistant Bacteria Selected by Antimicrobial Use in Food
Approved by the Food Safety Commission on September 30, 2004 Assessment guideline for the Effect of Food on Human Health Regarding Antimicrobial- Resistant Bacteria Selected by Antimicrobial Use in Food
WHY IS THIS IMPORTANT?
 CHAPTER 20 ANTIBIOTIC RESISTANCE WHY IS THIS IMPORTANT? The most important problem associated with infectious disease today is the rapid development of resistance to antibiotics It will force us to change
CHAPTER 20 ANTIBIOTIC RESISTANCE WHY IS THIS IMPORTANT? The most important problem associated with infectious disease today is the rapid development of resistance to antibiotics It will force us to change
Thiazole Antibiotic Thiostrepton Synergize with Bortezomib to Induce Apoptosis in Cancer Cells
 Thiazole Antibiotic Thiostrepton Synergize with Bortezomib to Induce Apoptosis in Cancer Cells Bulbul Pandit 1, Andrei L. Gartel 1,2,3 * 1 Department of Medicine, University of Illinois at Chicago, Chicago,
Thiazole Antibiotic Thiostrepton Synergize with Bortezomib to Induce Apoptosis in Cancer Cells Bulbul Pandit 1, Andrei L. Gartel 1,2,3 * 1 Department of Medicine, University of Illinois at Chicago, Chicago,
Mechanism of antibiotic resistance
 Mechanism of antibiotic resistance Dr.Siriwoot Sookkhee Ph.D (Biopharmaceutics) Department of Microbiology Faculty of Medicine, Chiang Mai University Antibiotic resistance Cross-resistance : resistance
Mechanism of antibiotic resistance Dr.Siriwoot Sookkhee Ph.D (Biopharmaceutics) Department of Microbiology Faculty of Medicine, Chiang Mai University Antibiotic resistance Cross-resistance : resistance
Cattle Serologically Positive for Brucella abortus Have Antibodies
 CLINICAL AND DIAGNOSTIC LABORATORY IMMUNOLOGY, Sept. 1994, p. 506-510 Vol. 1, No. 5 1071-412X/94/$04.00+0 Copyright X) 1994, American Society for Microbiology Cattle Serologically Positive for Brucella
CLINICAL AND DIAGNOSTIC LABORATORY IMMUNOLOGY, Sept. 1994, p. 506-510 Vol. 1, No. 5 1071-412X/94/$04.00+0 Copyright X) 1994, American Society for Microbiology Cattle Serologically Positive for Brucella
The Search For Antibiotics BY: ASLEY, ELIANA, ISABELLA AND LUNISCHA BSC1005 LAB 4/18/2018
 The Search For Antibiotics BY: ASLEY, ELIANA, ISABELLA AND LUNISCHA BSC1005 LAB 4/18/2018 The Need for New Antibiotics Antibiotic crisis An antibiotic is a chemical that kills bacteria. Since the 1980s,
The Search For Antibiotics BY: ASLEY, ELIANA, ISABELLA AND LUNISCHA BSC1005 LAB 4/18/2018 The Need for New Antibiotics Antibiotic crisis An antibiotic is a chemical that kills bacteria. Since the 1980s,
Centre for Public Health Research Laboratories
 2012 Centre for Public Health Research Laboratories Building 49 Ontario Veterinary College University of Guelph Guelph, Ontario N1G 2W1 Centre for Public Health and Zoonoses Last updated: 6/25/2012 Business
2012 Centre for Public Health Research Laboratories Building 49 Ontario Veterinary College University of Guelph Guelph, Ontario N1G 2W1 Centre for Public Health and Zoonoses Last updated: 6/25/2012 Business
Methicillin-Resistant Staphylococcus aureus
 Methicillin-Resistant Staphylococcus aureus By Karla Givens Means of Transmission and Usual Reservoirs Staphylococcus aureus is part of normal flora and can be found on the skin and in the noses of one
Methicillin-Resistant Staphylococcus aureus By Karla Givens Means of Transmission and Usual Reservoirs Staphylococcus aureus is part of normal flora and can be found on the skin and in the noses of one
Antimicrobial use in poultry: Emerging public health problem
 Antimicrobial use in poultry: Emerging public health problem Eric S. Mitema, BVM, MS, PhD CPD- Diagnosis and Treatment of Poultry Diseases FVM, CAVS, 6 th. August, 2014 AMR cont Antibiotics - Natural or
Antimicrobial use in poultry: Emerging public health problem Eric S. Mitema, BVM, MS, PhD CPD- Diagnosis and Treatment of Poultry Diseases FVM, CAVS, 6 th. August, 2014 AMR cont Antibiotics - Natural or
Antimicrobials & Resistance
 Antimicrobials & Resistance History 1908, Paul Ehrlich - Arsenic compound Arsphenamine 1929, Alexander Fleming - Discovery of Penicillin 1935, Gerhard Domag - Discovery of the red dye Prontosil (sulfonamide)
Antimicrobials & Resistance History 1908, Paul Ehrlich - Arsenic compound Arsphenamine 1929, Alexander Fleming - Discovery of Penicillin 1935, Gerhard Domag - Discovery of the red dye Prontosil (sulfonamide)
Evaluation of the hair growth and retention activity of two solutions on human hair explants
 activity of two solutions on human hair explants Study Directed by Dr E. Lati of Laboratoire Bio-EC, Centre de Recherches Biologiques et d Experimentations Cutanees, on behalf of Pangaea Laboratories Ltd.
activity of two solutions on human hair explants Study Directed by Dr E. Lati of Laboratoire Bio-EC, Centre de Recherches Biologiques et d Experimentations Cutanees, on behalf of Pangaea Laboratories Ltd.
POST SCREENING METHODS FOR THE DETECTION OF BETA-LACTAM RESIDUES IN PIGS.
 POST SCREENING METHODS FOR THE DETECTION OF BETA-LACTAM RESIDUES IN PIGS. Lorraine Lynas, Deborah Currie and John D.G. McEvoy. Department of Agriculture and Rural Development for Northern Ireland, Veterinary
POST SCREENING METHODS FOR THE DETECTION OF BETA-LACTAM RESIDUES IN PIGS. Lorraine Lynas, Deborah Currie and John D.G. McEvoy. Department of Agriculture and Rural Development for Northern Ireland, Veterinary
Visit ABLE on the Web at:
 This article reprinted from: Lessem, P. B. 2008. The antibiotic resistance phenomenon: Use of minimal inhibitory concentration (MIC) determination for inquiry based experimentation. Pages 357-362, in Tested
This article reprinted from: Lessem, P. B. 2008. The antibiotic resistance phenomenon: Use of minimal inhibitory concentration (MIC) determination for inquiry based experimentation. Pages 357-362, in Tested
INFECTIOUS HEPATITIS, PARVOVIRUS & DISTEMPER
 Canine VacciCheck INFECTIOUS HEPATITIS, PARVOVIRUS & DISTEMPER IgG ANTIBODY TEST KIT INSTRUCTION MANUAL Sufficient for 12/120 assays 13 JUL 2015 Biogal Galed Laboratories Acs. Ltd., tel: 972-4-9898605.
Canine VacciCheck INFECTIOUS HEPATITIS, PARVOVIRUS & DISTEMPER IgG ANTIBODY TEST KIT INSTRUCTION MANUAL Sufficient for 12/120 assays 13 JUL 2015 Biogal Galed Laboratories Acs. Ltd., tel: 972-4-9898605.
Author - Dr. Josie Traub-Dargatz
 Author - Dr. Josie Traub-Dargatz Dr. Josie Traub-Dargatz is a professor of equine medicine at Colorado State University (CSU) College of Veterinary Medicine and Biomedical Sciences. She began her veterinary
Author - Dr. Josie Traub-Dargatz Dr. Josie Traub-Dargatz is a professor of equine medicine at Colorado State University (CSU) College of Veterinary Medicine and Biomedical Sciences. She began her veterinary
ESBL Producers An Increasing Problem: An Overview Of An Underrated Threat
 ESBL Producers An Increasing Problem: An Overview Of An Underrated Threat Hicham Ezzat Professor of Microbiology and Immunology Cairo University Introduction 1 Since the 1980s there have been dramatic
ESBL Producers An Increasing Problem: An Overview Of An Underrated Threat Hicham Ezzat Professor of Microbiology and Immunology Cairo University Introduction 1 Since the 1980s there have been dramatic
 Tutorial 9 notes Super Bug: Antibiotics & Evolution Kristy J. Wilson Department of Pathology Emory University History of Antibiotics http://videos.howstuffworks.com/science-channel/29783-100-greatest-discoveries-penicillinvideo.htm
Tutorial 9 notes Super Bug: Antibiotics & Evolution Kristy J. Wilson Department of Pathology Emory University History of Antibiotics http://videos.howstuffworks.com/science-channel/29783-100-greatest-discoveries-penicillinvideo.htm
Fluoroquinolones ELISA KIT
 Fluoroquinolones ELISA KIT Cat. No.:DEIA6883 Pkg.Size:96T Intended use The Fluoroquinolones ELISA KIT is an immunoassay for the detection of Fluoroquinolones in contaminated samples including water, fish
Fluoroquinolones ELISA KIT Cat. No.:DEIA6883 Pkg.Size:96T Intended use The Fluoroquinolones ELISA KIT is an immunoassay for the detection of Fluoroquinolones in contaminated samples including water, fish
Genes What are they good for? STUDENT HANDOUT. Module 4
 Genes What are they good for? Module 4 Genetics for Kids: Module 4 Genes What are they good for? Part I: Introduction Genes are sequences of DNA that contain instructions that determine the physical traits
Genes What are they good for? Module 4 Genetics for Kids: Module 4 Genes What are they good for? Part I: Introduction Genes are sequences of DNA that contain instructions that determine the physical traits
Quantification of Chloramphenicol in Chicken Using Xevo TQD with RADAR Technology
 Quantification of Chloramphenicol in Chicken Using Xevo TQD with RADAR Technology Dimple Shah, Marian Twohig, and Jennifer A. Burgess Waters Corporation, Milford, MA, U.S.A. A P P L I C AT ION B E N E
Quantification of Chloramphenicol in Chicken Using Xevo TQD with RADAR Technology Dimple Shah, Marian Twohig, and Jennifer A. Burgess Waters Corporation, Milford, MA, U.S.A. A P P L I C AT ION B E N E
Genome 371; A 03 Berg/Brewer Practice Exam I; Wednesday, Oct 15, PRACTICE EXAM GENOME 371 Autumn 2003
 PRACTICE EXAM GENOME 371 Autumn 2003 These questions were part of the first exam from Autumn 2002. Take the exam in a quiet place and only when you are sure you will have time to complete the exam uninterrupted.
PRACTICE EXAM GENOME 371 Autumn 2003 These questions were part of the first exam from Autumn 2002. Take the exam in a quiet place and only when you are sure you will have time to complete the exam uninterrupted.
Epigenetic regulation of Plasmodium falciparum clonally. variant gene expression during development in An. gambiae
 Epigenetic regulation of Plasmodium falciparum clonally variant gene expression during development in An. gambiae Elena Gómez-Díaz, Rakiswendé S. Yerbanga, Thierry Lefèvre, Anna Cohuet, M. Jordan Rowley,
Epigenetic regulation of Plasmodium falciparum clonally variant gene expression during development in An. gambiae Elena Gómez-Díaz, Rakiswendé S. Yerbanga, Thierry Lefèvre, Anna Cohuet, M. Jordan Rowley,
10/15/08. Activity of an Antibiotic. Affinity for target. Permeability properties (ability to get to the target)
") Beta-lactam antibiotics Penicillins Target - Cell wall - interfere with cross linking Actively growing cells Bind to Penicillin Binding Proteins Enzymes involved in cell wall synthesis Activity of an Antibiotic
Beta-lactam antibiotics Penicillins Target - Cell wall - interfere with cross linking Actively growing cells Bind to Penicillin Binding Proteins Enzymes involved in cell wall synthesis Activity of an Antibiotic
of Conferences of OIE Regional Commissions organised since 1 June 2013 endorsed by the Assembly of the OIE on 29 May 2014
 of Conferences of OIE Regional Commissions organised since 1 June 2013 endorsed by the Assembly of the OIE on 29 May 2014 2 12 th Conference of the OIE Regional Commission for the Middle East Amman (Jordan),
of Conferences of OIE Regional Commissions organised since 1 June 2013 endorsed by the Assembly of the OIE on 29 May 2014 2 12 th Conference of the OIE Regional Commission for the Middle East Amman (Jordan),
An#bio#cs and challenges in the wake of superbugs
 An#bio#cs and challenges in the wake of superbugs www.biochemj.org/bj/330/0581/bj3300581.htm ciss.blog.olemiss.edu Dr. Vassie Ware Bioscience in the 21 st Century November 14, 2014 Who said this and what
An#bio#cs and challenges in the wake of superbugs www.biochemj.org/bj/330/0581/bj3300581.htm ciss.blog.olemiss.edu Dr. Vassie Ware Bioscience in the 21 st Century November 14, 2014 Who said this and what
Recommended for Implementation at Step 7 of the VICH Process on 15 December 2004 by the VICH Steering Committee
 VICH GL27 (ANTIMICROBIAL RESISTANCE: PRE-APPROVAL) December 2003 For implementation at Step 7 - Final GUIDANCE ON PRE-APPROVAL INFORMATION FOR REGISTRATION OF NEW VETERINARY MEDICINAL PRODUCTS FOR FOOD
VICH GL27 (ANTIMICROBIAL RESISTANCE: PRE-APPROVAL) December 2003 For implementation at Step 7 - Final GUIDANCE ON PRE-APPROVAL INFORMATION FOR REGISTRATION OF NEW VETERINARY MEDICINAL PRODUCTS FOR FOOD
Boosting Bacterial Metabolism to Combat Antibiotic Resistance
 Boosting Bacterial Metabolism to Combat Antibiotic Resistance The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published
Boosting Bacterial Metabolism to Combat Antibiotic Resistance The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published
Guidelines for Laboratory Verification of Performance of the FilmArray BCID System
 Guidelines for Laboratory Verification of Performance of the FilmArray BCID System Purpose The Clinical Laboratory Improvement Amendments (CLIA), passed in 1988, establishes quality standards for all laboratory
Guidelines for Laboratory Verification of Performance of the FilmArray BCID System Purpose The Clinical Laboratory Improvement Amendments (CLIA), passed in 1988, establishes quality standards for all laboratory
BIOLACTAM. Product Description. An innovative in vitro diagnostic for the rapid quantitative determination of ß-lactamase activity
 BIOLACTAM www.biolactam.eu An innovative in vitro diagnostic for the rapid quantitative determination of ß-lactamase activity 1.5-3h 20 Copyright 2014 VL-Diagnostics GmbH. All rights reserved. Product
BIOLACTAM www.biolactam.eu An innovative in vitro diagnostic for the rapid quantitative determination of ß-lactamase activity 1.5-3h 20 Copyright 2014 VL-Diagnostics GmbH. All rights reserved. Product
National Research Center
 National Research Center Update of immunodiagnosis of cystic echinococcosis cysts Global distribution of zoonotic strains of Echinococcus granulosus (Adapted from Eckert and Deplazes, 2004) Echinococcus
National Research Center Update of immunodiagnosis of cystic echinococcosis cysts Global distribution of zoonotic strains of Echinococcus granulosus (Adapted from Eckert and Deplazes, 2004) Echinococcus
Cell Wall Inhibitors. Assistant Professor Naza M. Ali. Lec 3 7 Nov 2017
 Cell Wall Inhibitors Assistant Professor Naza M. Ali Lec 3 7 Nov 2017 Cell wall The cell wall is a rigid outer layer, it completely surrounds the cytoplasmic membrane, maintaining the shape of the cell
Cell Wall Inhibitors Assistant Professor Naza M. Ali Lec 3 7 Nov 2017 Cell wall The cell wall is a rigid outer layer, it completely surrounds the cytoplasmic membrane, maintaining the shape of the cell
Diurnal variation in microfilaremia in cats experimentally infected with larvae of
 Hayasaki et al., Page 1 Short Communication Diurnal variation in microfilaremia in cats experimentally infected with larvae of Dirofilaria immitis M. Hayasaki a,*, J. Okajima b, K.H. Song a, K. Shiramizu
Hayasaki et al., Page 1 Short Communication Diurnal variation in microfilaremia in cats experimentally infected with larvae of Dirofilaria immitis M. Hayasaki a,*, J. Okajima b, K.H. Song a, K. Shiramizu
Antibiotic Resistance The Global Perspective
 Antibiotic Resistance The Global Perspective Scott A. McEwen Department of Population Medicine, University of Guelph, Guelph, ON N1G 2W1; Email: smcewen@uoguleph.ca Introduction Antibiotics have been used
Antibiotic Resistance The Global Perspective Scott A. McEwen Department of Population Medicine, University of Guelph, Guelph, ON N1G 2W1; Email: smcewen@uoguleph.ca Introduction Antibiotics have been used
Terrestrial and Aquatic Manuals and the mechanism of standard adoption
 Dr Patrick Bastiaensen Programme Officer OIE Sub-Regional Representation for Eastern Africa Terrestrial and Aquatic Manuals and the mechanism of standard adoption Presented during the Regional Workshop
Dr Patrick Bastiaensen Programme Officer OIE Sub-Regional Representation for Eastern Africa Terrestrial and Aquatic Manuals and the mechanism of standard adoption Presented during the Regional Workshop
AMOXICILLIN AND CLAVULANIC ACID TABLETS Draft proposal for The International Pharmacopoeia (February 2018)
") February 2018 Draft for comment 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 AMOXICILLIN AND CLAVULANIC ACID TABLETS Draft
February 2018 Draft for comment 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 AMOXICILLIN AND CLAVULANIC ACID TABLETS Draft
Wageningen Bioveterinary Research. Biomedical and veterinary research to safeguard animal and public health
 Wageningen Bioveterinary Research Biomedical and veterinary research to safeguard animal and public health Veterinary research to safeguard animal and public health Wageningen Bioveterinary Research (WBVR)
Wageningen Bioveterinary Research Biomedical and veterinary research to safeguard animal and public health Veterinary research to safeguard animal and public health Wageningen Bioveterinary Research (WBVR)
In the first half of the 20th century, Dr. Guido Fanconi published detailed clinical descriptions of several heritable human diseases.
 In the first half of the 20th century, Dr. Guido Fanconi published detailed clinical descriptions of several heritable human diseases. Two disease syndromes were named after him: Fanconi Anemia and Fanconi
In the first half of the 20th century, Dr. Guido Fanconi published detailed clinical descriptions of several heritable human diseases. Two disease syndromes were named after him: Fanconi Anemia and Fanconi
Detection of Mastitis
 Detection of Mastitis Changes in milk composition Changes in milk composition Physical examination Signs of inflammation Empty udder Differences in firmness Unbalanced quarters Taste Test 60% of salty
Detection of Mastitis Changes in milk composition Changes in milk composition Physical examination Signs of inflammation Empty udder Differences in firmness Unbalanced quarters Taste Test 60% of salty
Outcome of the Conference Towards the elimination of rabies in Eurasia Joint OIE/WHO/EU Conference
 Outcome of the Conference Towards the elimination of rabies in Eurasia Joint OIE/WHO/EU Conference WHO (HQ-MZCP) / OIE Inter-country Workshop on Dog and Wildlife Rabies Control in the Middle East 23-25
Outcome of the Conference Towards the elimination of rabies in Eurasia Joint OIE/WHO/EU Conference WHO (HQ-MZCP) / OIE Inter-country Workshop on Dog and Wildlife Rabies Control in the Middle East 23-25
NFI is an Essential Positive Transcription Factor for Human Papillomavirus Type 16 Early Gene Expression
 The Open Virology Journal, 2007, 1, 33-38 33 NFI is an Essential Positive Transcription Factor for Human Papillomavirus Type 16 Early Gene Expression Amy Baldwin 1, Melissa K. Hypes 2, Lucia Pirisi 3 and
The Open Virology Journal, 2007, 1, 33-38 33 NFI is an Essential Positive Transcription Factor for Human Papillomavirus Type 16 Early Gene Expression Amy Baldwin 1, Melissa K. Hypes 2, Lucia Pirisi 3 and
funded by Reducing antibiotics in pig farming
 funded by Reducing antibiotics in pig farming The widespread use of antibiotics (also known as antibacterials) in human and animal medicine increases the level of resistant bacteria. This makes it more
funded by Reducing antibiotics in pig farming The widespread use of antibiotics (also known as antibacterials) in human and animal medicine increases the level of resistant bacteria. This makes it more
Statistical Analysis of Nipah Virus Using R
 RESEARCH ARTICLE Statistical Analysis of Nipah Virus Using R Roopa Sri Paladugu 1, Sruthi Chilukuri 2, Dr.Sangeetha Yalamanchili 3 1(Department of IT, V.R.Siddhartha Engineering College, and Vijayawada)
RESEARCH ARTICLE Statistical Analysis of Nipah Virus Using R Roopa Sri Paladugu 1, Sruthi Chilukuri 2, Dr.Sangeetha Yalamanchili 3 1(Department of IT, V.R.Siddhartha Engineering College, and Vijayawada)
ENVIRACOR J-5 aids in the control of clinical signs associated with Escherichia coli (E. coli) mastitis
 mastitis") GDR11136 ENVIRACOR J-5 aids in the control of clinical signs associated with Escherichia coli (E. coli) mastitis February 2012 Summary The challenge data presented in this technical bulletin was completed
GDR11136 ENVIRACOR J-5 aids in the control of clinical signs associated with Escherichia coli (E. coli) mastitis February 2012 Summary The challenge data presented in this technical bulletin was completed
Antibiotics. Antimicrobial Drugs. Alexander Fleming 10/18/2017
 Antibiotics Antimicrobial Drugs Chapter 20 BIO 220 Antibiotics are compounds produced by fungi or bacteria that inhibit or kill competing microbial species Antimicrobial drugs must display selective toxicity,
Antibiotics Antimicrobial Drugs Chapter 20 BIO 220 Antibiotics are compounds produced by fungi or bacteria that inhibit or kill competing microbial species Antimicrobial drugs must display selective toxicity,
FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT
![FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT](/thumbs/93/113164093.jpg "FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT") FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT INSTRUCTION MANUAL Sufficient for 12/120 assays 22 APR 2018 Biogal Galed Laboratories Acs Ltd. tel: 972-4-9898605. fax: 972-4-9898690 e-mail:info@biogal.co.il
FELINE CORONAVIRUS (FCoV) [FIP] ANTIBODY TEST KIT INSTRUCTION MANUAL Sufficient for 12/120 assays 22 APR 2018 Biogal Galed Laboratories Acs Ltd. tel: 972-4-9898605. fax: 972-4-9898690 e-mail:info@biogal.co.il
Antimicrobial agents. are chemicals active against microorganisms
 Antimicrobial agents are chemicals active against microorganisms Antibacterial Agents Are chemicals active against bacteria Antimicrobials Antibacterial Antifungal Antiviral Antiparasitic: -anti protozoan
Antimicrobial agents are chemicals active against microorganisms Antibacterial Agents Are chemicals active against bacteria Antimicrobials Antibacterial Antifungal Antiviral Antiparasitic: -anti protozoan
2008 FELINE HEALTH GRANT AWARDS 10 projects funded for a total of $135,860
 2008 FELINE HEALTH GRANT AWARDS 10 projects funded for a total of $135,860 The Winn Feline Foundation receives proposals from veterinary researchers around the world who are interested in improving feline
2008 FELINE HEALTH GRANT AWARDS 10 projects funded for a total of $135,860 The Winn Feline Foundation receives proposals from veterinary researchers around the world who are interested in improving feline
Controlling Microbial Growth in the Body: Antimicrobial Drugs
 PowerPoint Lecture Presentations prepared by Mindy Miller-Kittrell, North Carolina State University C H A P T E R 10 Controlling Microbial Growth in the Body: Antimicrobial Drugs Interesting News https://www.technologyreview.com/s/601138/the-fridge-laserthat-detects-bacteria-crawling-all-over-food/
PowerPoint Lecture Presentations prepared by Mindy Miller-Kittrell, North Carolina State University C H A P T E R 10 Controlling Microbial Growth in the Body: Antimicrobial Drugs Interesting News https://www.technologyreview.com/s/601138/the-fridge-laserthat-detects-bacteria-crawling-all-over-food/
Bovine Brucellosis Control of indirect ELISA kits
 Bovine Brucellosis Control of indirect ELISA kits (Pooled milk samples) Standard Operating Procedure Control of Bovine brucellosis Milk ELISA kits SOP Page 1 / 6 02 February 2012 SAFETY PRECAUTIONS The
Bovine Brucellosis Control of indirect ELISA kits (Pooled milk samples) Standard Operating Procedure Control of Bovine brucellosis Milk ELISA kits SOP Page 1 / 6 02 February 2012 SAFETY PRECAUTIONS The
ANNEX I SUMMARY OF PRODUCT CHARACTERISTICS VIRBAGEN OMEGA - EN 1
 ANNEX I SUMMARY OF PRODUCT CHARACTERISTICS VIRBAGEN OMEGA - EN 1 1. NAME OF THE VETERINARY MEDICINAL PRODUCT Virbagen Omega 5 MU for dogs Virbagen Omega 10 MU for dogs 2. QUALITATIVE AND QUANTITATIVE COMPOSITION
ANNEX I SUMMARY OF PRODUCT CHARACTERISTICS VIRBAGEN OMEGA - EN 1 1. NAME OF THE VETERINARY MEDICINAL PRODUCT Virbagen Omega 5 MU for dogs Virbagen Omega 10 MU for dogs 2. QUALITATIVE AND QUANTITATIVE COMPOSITION
Antibiotics: mode of action and mechanisms of resistance. Slides made by Special consultant Henrik Hasman Statens Serum Institut
 Antibiotics: mode of action and mechanisms of resistance. Slides made by Special consultant Henrik Hasman Statens Serum Institut This presentation Definitions needed to discuss antimicrobial resistance
Antibiotics: mode of action and mechanisms of resistance. Slides made by Special consultant Henrik Hasman Statens Serum Institut This presentation Definitions needed to discuss antimicrobial resistance
Molecular Analysis of β-lactamase Genes in Antibiotic Resistant Bacteria
 Bowling Green State University ScholarWorks@BGSU Honors Projects Honors College Spring 5-1-2017 Molecular Analysis of β-lactamase Genes in Antibiotic Resistant Bacteria Neisha Medina Candelaria neisham@bgsu.edu
Bowling Green State University ScholarWorks@BGSU Honors Projects Honors College Spring 5-1-2017 Molecular Analysis of β-lactamase Genes in Antibiotic Resistant Bacteria Neisha Medina Candelaria neisham@bgsu.edu
Overview. There are commonly found arrangements of bacteria based on their division. Spheres, Rods, Spirals
 Bacteria Overview Bacteria live almost everywhere. Most are microscopic ranging from 0.5 5 m in size, and unicellular. They have a variety of shapes when viewed under a microscope, most commonly: Spheres,
Bacteria Overview Bacteria live almost everywhere. Most are microscopic ranging from 0.5 5 m in size, and unicellular. They have a variety of shapes when viewed under a microscope, most commonly: Spheres,
Development and improvement of diagnostics to improve use of antibiotics and alternatives to antibiotics
 Priority Topic B Diagnostics Development and improvement of diagnostics to improve use of antibiotics and alternatives to antibiotics The overarching goal of this priority topic is to stimulate the design,
Priority Topic B Diagnostics Development and improvement of diagnostics to improve use of antibiotics and alternatives to antibiotics The overarching goal of this priority topic is to stimulate the design,
Principles of Antimicrobial therapy
 Principles of Antimicrobial therapy Laith Mohammed Abbas Al-Huseini M.B.Ch.B., M.Sc, M.Res, Ph.D Department of Pharmacology and Therapeutics Antimicrobial agents are chemical substances that can kill or
Principles of Antimicrobial therapy Laith Mohammed Abbas Al-Huseini M.B.Ch.B., M.Sc, M.Res, Ph.D Department of Pharmacology and Therapeutics Antimicrobial agents are chemical substances that can kill or
VMP Focal point training Casablanca 6 8 December Dr Susanne Münstermann
 VMP Focal point training Casablanca 6 8 December 2011 Dr Susanne Münstermann The OIE Specialist Commissions and their mandate The Terrestrial Manual - overview Diagnostic Tests Vaccines The Aquatic Manual
VMP Focal point training Casablanca 6 8 December 2011 Dr Susanne Münstermann The OIE Specialist Commissions and their mandate The Terrestrial Manual - overview Diagnostic Tests Vaccines The Aquatic Manual
Arrested oocyst maturation in Plasmodium parasites. lacking type II NADH:ubiquinone dehydrogenase
 Supplemental Information for: Arrested oocyst maturation in Plasmodium parasites lacking type II NADH:ubiquinone dehydrogenase Katja E. Boysen and Kai Matuschewski Contents: - Supplemental Movies 1 and
Supplemental Information for: Arrested oocyst maturation in Plasmodium parasites lacking type II NADH:ubiquinone dehydrogenase Katja E. Boysen and Kai Matuschewski Contents: - Supplemental Movies 1 and
Sera from 2,500 animals from three different groups were analysed:
 FIELD TRIAL OF A BRUCELLOSIS COMPETITIVE ENZYME LINKED IMMUNOABSORBENT ASSAY (ELISA) L.E. SAMARTINO, R.J. GREGORET, G. SIGAL INTA-CICV Instituto Patobiología Area Bacteriología, Buenos Aires, Argentina
FIELD TRIAL OF A BRUCELLOSIS COMPETITIVE ENZYME LINKED IMMUNOABSORBENT ASSAY (ELISA) L.E. SAMARTINO, R.J. GREGORET, G. SIGAL INTA-CICV Instituto Patobiología Area Bacteriología, Buenos Aires, Argentina
OIE Collaborating Centres Reports Activities
 OIE Collaborating Centres Reports Activities Activities in 2015 This report has been submitted : 2016-03-24 20:54:12 Title of collaborating centre: Emerging and Re-Emerging Zoonotic Diseases Address of
OIE Collaborating Centres Reports Activities Activities in 2015 This report has been submitted : 2016-03-24 20:54:12 Title of collaborating centre: Emerging and Re-Emerging Zoonotic Diseases Address of
HEALTH REQUIREMENTS FOR ANIMALS EXHIBITED AT THE 2018 NEBRASKA STATE FAIR
 HEALTH REQUIREMENTS FOR ANIMALS EXHIBITED AT THE 2018 NEBRASKA STATE FAIR Nebraska Department of Agriculture State Veterinarian s Office 301 Centennial Mall South, Lincoln, Nebraska 68509-4787 Phone: (402)
HEALTH REQUIREMENTS FOR ANIMALS EXHIBITED AT THE 2018 NEBRASKA STATE FAIR Nebraska Department of Agriculture State Veterinarian s Office 301 Centennial Mall South, Lincoln, Nebraska 68509-4787 Phone: (402)
Amoxicillin trihydrate. Amoxicillin trihydrate. Amoxicillin trihydrate. Amoxicillin trihydrate. Amoxicillin trihydrate. Amoxicillin trihydrate
 Annex I List of the names, pharmaceutical form, strength of the veterinary medicinal product, animal species, route of administration, applicant in the Member States Member State EU/EEA Applicant Name
Annex I List of the names, pharmaceutical form, strength of the veterinary medicinal product, animal species, route of administration, applicant in the Member States Member State EU/EEA Applicant Name
Developmental expression of synthetic cis-regulatory systems composed of spatial control elements from two different genes
 Proc. Natl. Acad. Sci. USA Vol. 93, pp. 13849 13854, November 1996 Developmental Biology Developmental expression of synthetic cis-regulatory systems composed of spatial control elements from two different
Proc. Natl. Acad. Sci. USA Vol. 93, pp. 13849 13854, November 1996 Developmental Biology Developmental expression of synthetic cis-regulatory systems composed of spatial control elements from two different
number Done by Corrected by Doctor Dr Hamed Al-Zoubi
 number 8 Done by Corrected by Doctor Dr Hamed Al-Zoubi 25 10/10/2017 Antibacterial therapy 2 د. حامد الزعبي Dr Hamed Al-Zoubi Antibacterial therapy Figure 2/ Antibiotics target Inhibition of microbial
number 8 Done by Corrected by Doctor Dr Hamed Al-Zoubi 25 10/10/2017 Antibacterial therapy 2 د. حامد الزعبي Dr Hamed Al-Zoubi Antibacterial therapy Figure 2/ Antibiotics target Inhibition of microbial
Diseases of Concern: BVD and Trichomoniasis. Robert Mortimer, DVM Russell Daly, DVM Colorado State University South Dakota State University
 Diseases of Concern: BVD and Trichomoniasis Robert Mortimer, DVM Russell Daly, DVM Colorado State University South Dakota State University The Epidemiologic Triad Host Management Agent Environment Trichomoniasis
Diseases of Concern: BVD and Trichomoniasis Robert Mortimer, DVM Russell Daly, DVM Colorado State University South Dakota State University The Epidemiologic Triad Host Management Agent Environment Trichomoniasis