Redpalh Museum, McGill University, Montreal, P.Q, Canada, HJA 2K6.
|
|
|
- Paul Charles
- 5 years ago
- Views:
Transcription

1 143 Palaeont. afr., 21, (1978) PERMO-TRIASSIC "LIZARDS" FROM THE KAROO SYSTEM PART II A GLIDING REPTILE FROM THE UPPER PERMIAN OF MADAGASCAR by Robert L. Carroll Redpalh Museum, McGill University, Montreal, P.Q, Canada, HJA 2K6. ABSTRACT DaedaLosaurus madagascariemis, gen. et sp. nov. from the Upper Permian of Madagascar is a small reptile in which the trunk ribs are greatly elongated to support a gliding "membrane" similar to those in the Upper Triassic lizards Kuehneosaurus and lcarosaurus, and the living agamid Draco. The membrane is supported by 21 pairs of ribs compared with five to seven in Draco, ten in l carmautus and II in Kuehneosaurus. The total body mass is estimated as 250 grams, the area of the membrane nearly 200 cm2, with a wing loading of approximately 1,25 glcm2. A second species in the fauna, belonging to the same family, Coelurosauravus elivemis Piveteau, has a very similar appendicular skeleton, but ribs of normal proportions. The maxillary dentition of Coelurosauravus is acrodont, that of DaedaLosaurus pleurodont. In neither genus is the temporal region of the skull adequately known, although the configuration of the jugal in Daedalosaurus suggests that the lower temporal bar may be reduced. The primitive nature of the appendicular skeleton, with little evidence or the specializations seen in contemporary lizards, suggests that these genera should not be classified among the Squamata, but among the Eosuchia. CONTENTS Page INTRODUCTION COELUROSAURAVUS ELIVENSIS PIVETEAU DAEDALOSAURUS MADAGASCARIENSIS, gen. et sp. nov Skull Vertebrae Ribs Appendicular skeleton Flight characteristics and comparison with other gliding genera THE TAXONOMIC POSITION OF COELUROSAURAVUS AND DAEDALOSAURUS ACKNOWLEDGEMENTS REFERENCES INTRODUCTION Knowledge of Late Permian and Early Triassic reptiles has come primarily from the Karoo age beds of South Africa and Russia. The fauna from these deposits consists almost entirely of therapsids, with diapsids as relatively rare elements. Reptiles of this age have also been described from Madagascar (Piveteau, 1926). Because of the difficulty of preparation and the lack of comparative material then available, the significance of this fauna has not been generally appreciated. Development of new techniques of preparation and casting reveals an extensive and varied fauna, quite different from that known elsewhere, consisting almost entirely of diapsids. Karoo age reptiles are known primarily from deposits exposed as a narrow band running north and south along the west side of the crystalline massif that makes up much of the eastern two-thirds of the island (fig. 1). Most of the material described by Piveteau, including all the specimens discussed in this paper, came from the upper course of the Sakamena River, near Mount Eliva in the southwestern part of the island. All the vertebrates described from this area occur in nodules, and are typically found weathered free from the surrounding matrix. In view of the significance of the fauna, made up of many genera as yet known only from Madagascar, it is important to establish the age of the beds with as much assurance as possible. Piveteau (1926) attri-

2 144 Figure I. Map showing approximate extent of the Lower Sakamena Formation in south-western Madagascar and localities /Tom which Upper Permian reptiles have been described. buted the reptilian fauna of Mount Eliva to the Upper Permian, primarily on the basis of assumed relationships to South African genera, and relative levels of evolution. Although concepts of the relationships of early diapsids have changed considerably since his determination, stratigraphic work and comparison of other fossils tend to support his estimate of the age of the beds. Stratigraphic work reported by Besairie (1972) places the beds which contain these vertebrates in the Lower Sakamena Formation. Farther north, this formation is overlain by beds containing the same genera as those of the extremely well-known Eotriassic fish fauna in the north of the island, e.g. Boreosomus and Birgeria (Lehman et ai., 1959). Recent palynological work by Goubin (1965) on numerous localities in Madagascar divides the Lower Sakamena into three zones, all of which are indicated as Upper Permian. The terrestrial vertebrates occur in the uppermost of the three zones recognized by Goubin, and so at the very top of the Permian. Hart (969) correlates the entire Lower Sakamena, typified by members of the Striatiti pollen zone, with the Russian Tatarian. In addition to the terrestrial vertebrates of the Mount Eliva fauna, Priem (924) described several fish specimens as members of a new species, Atherstonia colcanapi. According to recent study of this material by Gardner (pers. comm.) the fish accord well with other species of Atherstonia from the Upper Permian; members of this genus are not known in later horizons. Although perhaps less important as stratigraphic indicators, the mega-plant material from these beds should be cited as well. Beneath the vertebrate bed is a rich flora, dominated by Thinnjeldia and Glossopteris. Within the vertebrate bearing nodules are also impressions of Glossopteris indica (Carpentier, 1936). Most of the specimens from the area of Mount Eliva have been attributed to the aquatic eosuchian genera Tangasaurus and Hovasaurus. There are also three more or less complete skeletons of small lizard-like forms, well ossified, with slender limbs and without evident aquatic adaptations. Two of these were illustrated by Piveteau, and described as a new genus and species of coelurosaur, Coelurosauravus elivensis. The third is of the same general size and limb proportions, but has greatly elongate ribs, resembling those of the Upper Triassic gliding lizards Kuehneosaurus (Robinson, 1962) and Icarosaurus (Colbert, 1970). In none of the Madagascar specimens is the skull well preserved, but the dentition is very different in the two genera. Coelurosauravus has a distinctly acrodont attachment of the posterior maxillary teeth, which are few in number and widely spaced. The form with long ribs has a subpleurodont implantation, with a large number of slender teeth. Although the two forms clearly belong to distinct genera, the similarity of the appendicular skeleton suggests that they may belong to the same family. Both are clearly distinct from most other Late Permian or Early Triassic reptiles including the primitive lizards from the South African Karoo beds (Carroll, 1975 ). The apparent close relationship of the two Madagascan genera makes it useful to consider them together. COELUROSAURA VUS ELIVENSIS PIVETEAU The two specimens of Coelurosauravus described by Piveteau are numbered a (the type, fig. 2 and a, fig. 3) in the collection of the Institute of Palaeontology, National Museum of Natural History, Paris. He recognized as belonging to the same genus a disarticulated maxilla and other skull bones in the same block as a skeleton of the genus with long ribs ; fig. 5). The first two specimens show the remains of almost complete skeletons, preserved in nodules exhibiting a clearly

3 145 concentric structure in cross-section. The bone had entirely weathered out prior to collection. The surface has since been cleaned by washing and picking out mud from the deeper cavities with a needle. The matrix is rather coarse-grained and some areas badly weathered, so that surface detail tends to be poor. The specimens have been studied through the use of high fidelity latex casts. In none of the specimens is the cranium adequately preserved. All three show the maxilla, but the remainder of the skull appears to have been much less stoutly constructed, and much had disintegrated prior to preservation. The only bones of the skull roof to be seen clearly are the parietal and frontal, visible in the counterpart blocks of , adjacent to the maxilla. What appears at first glance to be a very elongate parietal is apparently the leti: frontal overlapping the right parietal, judging from the configuration of these bones in the primitive lizard Paliguana (Carroll, 1975). The frontal is an ob- 1cm /~./ :.::\.'. 11 Figure 2. Type of CoelUTOsauravUJ elivemis. Institut de Palt~ontologie, Paris, no a. Numbers indicate approximate position of presacral vertebrae and distal tarsals. Roman numerals indicate the number of metatarsals. a, astragalus; cal, calcaneum; cen, centrale; ec f, ectepicondylar foramen; en f, entepicondylar foramen; F, femur; Fi, fibula; H, humerus; m, maxilla; pm, premaxilla; R radius; T, tibia; U, ulna. X 2.

4 146 long bone, with a strong longitudinal ridge separating the orbital margin from the dorsolateral margin of the braincase. A groove on the anterolateral margin of the frontal may be interpreted as the position of prefrontal attachment. The area where the postfrontal might be expected to have attached is obscured where the frontal overlaps the anteromedial margin of the parietal. The parietal resembles those of both Youngina (Cow, 1975) and Paliguana in being short, with a prominent lateral process extending behind the dorsal temporal opening, and a large parietal foramen midway along the medial margin. The posterolateral margin is recessed in the area of the tabular and postparietal. The ventral surface is strongly concave. The configuration of the frontal and parietal in younginids and paliguanids is unfortunately so similar that it is not possible, on the basis of these bones, to identiry to which group Coelurosauravus might be allied. Much of the skull may originally have been present in a, but only the tooth-bearing elements are identifiable. The premaxilla shows a large margin for the external nares anteriorly (as the bone is oriented) with the posterior margin slanting posterodorsally. It is possible that this indicates an anteromedial position for the external nares as in Kuehneosaurus. Alternatively, this bone may have been reversed prior to fossilization. It shows three or four small, peg-like teeth, apparently quite unlike those of the maxilla. Maxillae are preserved in all three specimens. They appear to be very massive, and are peculiar in several respects. In most early lizard-like reptiles, the maxilla is very narrow posteriorly, but in these specimens, the posterior extremity is thick and ends abruptly in what appears to be a large surface for medial attachment, presumably with the ectopterygoid. The bones appear relatively short, although it is not certain whether any show the anterior margin. Each maxilla bears a small number of widely spaced triangular acrodont teeth. Seven is the maximum number preserved. The most anterior tooth in the type specimen appears to be set in a socket. The combination of peg-like premaxillary teeth, a socketed anterior maxillary tooth and acrodont teeth in the remainder of the maxilla is strikingly close to the pattern common among living agamid lizards. One apparent difference is that in agamids, the jugal extends far anteriorly, bracing the medial surface of the maxilla nearly to the front of the orbit. In Coelurosauravus, the maxilla appears very high posteriorly, and apparently forms most of the ventral margin of the orbit. This does not necessarily preclude a parallel anterior extension of the jugal, but there is no direct evidence for it. Oddly, no bone can assuredly be identified from the lower jaw. A splint-shaped element in a, associated with a bone which somewhat resembles a quadrate, might be an angular, sheathing a long retroarticular process. No further cranial elements can be recognized, and the specification of this form as a "lizard" or eosuchian cannot be based on evidence of the temporal region. In a most of the vertebral column is in place, extending from behind the skull in a tight circle, crossing over itself in the cervical region. Unfortunately, the most anterior vertebrae are obscured in the area of the occiput, precluding positive identification of the atlas or axis. Twenty-three vertebrae can be identified, almost certainly not including the sacrals. The first visible centrum is quite long, and so might be the axis but not the atlas, to judge by the relative length of these elements in most primitive reptiles. The last visible vertebra is hence probably the 24th presacral. In the type, the trunk vertebrae are less well displayed. The cervicals and anterior trunk vertebrae are too badly obscured to estimate their numbers. Several in the mid-region of the trunk are badly disarticulated. Only the posterior trunk, sacral and anterior caudal vertebrae are in articulation and relatively clearly exposed. Because of regional variation in the length of the centra, the relative position along the column may be correlated in the two specimens. The vertebrae which may thus be identified as the 25th in the type bears a short slender rib. Posterior to it are five or six additional vertebrae anterior to the first haemal arch. None are well enough preserved to determine which are the sacral vertebrae. There were apparently between 25 and 27 presacral vertebrae. Judging by the pattern in nearly all primitive reptiles, there were presumably two sacral vertebrae. Nine to eleven caudal vertebrae are preserved. The tail was almost certainly considerably longer in the living animal. Throughout the column, the centra are longer relative to their width than in most other primitive reptiles. Equally striking is the change in vertebral length throughout the column. In Saurostemon and Palaeagama from the Upper Permian and Lower Triassic of South Africa (Carroll, 1977 ) and the betterknown Jurassic lizards, the cervical vertebrae are relatively short, but the length remains essentially constant throughout the remainder of the column. In Coelurosauravus, the cervical vertebrae are not particularly short, but those in the posterior trunk region are considerably longer. This pattern may be seen in the gliding lizard lcarosaurus as well. The immediately presacral vertebrae are shorter, as are the sacrals and the most anterior caudals. More posteriorly, however, the caudals lengthen to approach the length of the longer trunk vertebrae. In the region of the carpus of the long-ribbed genus as preserved, are paired atlas arches and an axis which may pertain to Coelurosauravus. The paired nature of the atlas arch compares with eosuchians and sphenodontids, but contrasts with the median nature of this element in lizards, including the Lower Triassic genus Palaeagama (Carroll, 1975 ). Well-defined transverse processes suggest the presence of atlas ribs, absent in early lizards. The most posterior cervical and anterior trunk vertebrae have relatively high, rectangular neural spines; not elong-

5 147 Figure 3. CoeluTOsauravUJ elivensis, Institut de Paleonto10gie, Paris, no a. Numbers indicate the approximate position ot presacral vertebrae. ang, angular; Cl, clavicle; Clei, cleithrum; en f, entepicondylar foramen; H, humerus; m, maxilla; R, radius; S, scapulocoracoid; U, ulna. X 2. ate anteroposteriorly as are those of lcarojaurus, but clearly much more prominent than in most small primitive lizards. lbe spines become more triangular in shape posteriorly. The transverse processes of the fourth, fifth and sixth vertebrae extend laterally to an appreciable extent and have a long anteroventrally sloping articulating surface. As well as can be seen, those of the anterior seven or eight vertebrae are not significantly different from those of other early lizard-like forms. Most posteriorly, the articulating surfaces appear to be much shorter, and to shift posteriorly toward the middle of the elongate pedicle. Just anterior to the sacrum, they revert to a more anterior position. In the caudal series, the elongate anterior trans-

6 148 verse processes common to eosuchians and lizards are not visible. Normal haemal arches are evident in the anterior portion of the tail. Small crescentic intercentra can be seen anterior to vertebrae four and five in a. Ribs are present throughout most of the trunk region in a. Little can be seen of those that would have been attached to the cervical vertebrae, although the size of the transverse processes indicates that the heads would have been substantial. It is possible that the short ribs opposite vertebrae seven through nine are displaced from the cervical region. In a, ribs in the anterior trunk are much longer and appear wide. Ribs associated with vertebrae ten through 16 are more or less in position in a. Except for being somewhat expanded towards the distal end (perhaps as a result of crushing), they are of typical size and configuration seen in other small reptiles of this age. Ribs associated with the more posterior trunk vertebrae appear considerably smaller. In general, the ribs associated with this specimen can be said to be normal for a primitive reptile. One much longer rib accompanies a. It is approximately the length of the femur, it is not in articulation with any vertebra, and is therefore possibly from another specimen. The loss of several trunk vertebrae and most of the ribs in the central portion of the trunk makes it difficult, however, to rule out the presence of longer ribs in this region. The shoulder girdle is barely visible in the type. In a, however, much of the structure can be made out. The scapulocoracoid appears as a single ossification, with a large screw-shaped glenoid of generally primitive configuration. The posterior articulating surface is oriented vertically, suggesting that the humerus had more freedom to angle ventrally than in captorhinomorphs and pelycosaurs. The scapula appears tall and narrow, although the anterior margin cannot be determined. The interclavicle is not evident. The narrow blades of both clavicles can be seen, but not the extent of the stems. A bone adjacent to the blade of the scapula may be a remnant of the cleithrum. If so, this bone is relatively massive. There is no evidence of an ossified sternum. The humerus, exposed in both skeletons, is a long narrow bone, with the ends twisted in a primitive fashion. The proximal portion shows a well-defined narrow strap-shaped articulating surface. Except for proportional differences related to the relative size of the head of the humerus, it resembles the pattern seen in pelycosaurs and captorhinomorphs (Holmes, 1977) and does not show evidence of the inception of a lacertoid pattern, such as is noted in the contemporary South African lizard Saurostemon (Carroll, 1977). The articulating surface is somewhat bulbous, however, indicating less restriction of movement at this joint. The shaft is long and slim. The distal extremity is flattened. At the very posterior margin of the bone is a narrow slit of the entepicon- Figure 4. A :.... \.', :. :,, ~- \, ' \ \ : \, c ' : 1 '. ~ \ \ \~..' \ \.' Ra MC~~5 Lc,.JIo 5~~""<:",,j x\, \..\ '..:. B 1cm Limb elements or CoelurosauravUJ and DaedalosauTUJ. A, Ventral view of humerus o f Daedalosaurus madagalcariensij. B, Ventral view o f hand and associated ulna and radius or Daedalosaurus madof!,ascariensis. C, Ventral view or lower rear limb of CoelurosauravUJ elivensis. All X , distal carpals and tarsals ; a, astragal us; ca l, calcaneum; cap, capitellum ; cen, centrale; ec C ectepicondylar foramen ; en f, entepicondylar 10 r<ll11('n ; I, il1l C'rmedium ; Lc, lateral centrale; Mc, medial centrale ; Pi, pisiform; Ra, radiale; II', trochlea ; Ul, ulnarc'. dylar foramen. Anteriorly, there appears to be a small ectepicondylar foramen. Dorsally, approximately midway in the width of the bone is a slightly raised area for articulating with the olecranon. The surface of the humerus is distinctly recessed proximally and posterior to this area. The ventral surface of the distal extremity is not exposed. Pi

7 149 The radius is a narrow cylindrical bone, 62 per cent the length of the humerus. Distally, the bone is somewhat compressed in the plane of the carpus. The articulating surface is not well exposed. The total length of the ulna is 73 per cent that of the humerus, with a very well-developed olecranon, the tip of which shows a distinct notch for the insertion of the triceps. The proximal end of the bone is markedly compressed in an anteroposterior direction; the distal end of the shaft narrows considerably above the area of distal expansion. The distal end of the bone does not appear to be elongated as a specialized epiphyseal articulating surface as in Saurostemon. The carpus is not exposed. Metacarpals and phalanges are preserved in a but the hand is too incomplete to be restored without reference to other primitive reptiles. One digit shows three quite long phalanges. Other phalanges and possibly metacarpals can be seen in the type specimen, beyond the distal end of the humerus. angular. Proximally, it appears to be overriden by the lateral margin of the astragalus. Just beneath this area is apparently the margin of the perforating foramen. The astragalus has a prominent articulating surface on the lateral margin for the tibia. The centrale is a small bone largely obscured by overlying elements. The area of the fifth distal tarsal is poorly exposed, but there is no reason to think that this element is missing. Distal tarsals two, three and four are relatively well exposed, but the third and fourth appear broken, so that their original outline is difficult to restore exactly. The first distal tarsal is not exposed, but was probably bent under the other elements. Distal tarsals two and three are unusually long relative to their width. The metatarsals are all in place, although the proximal end of the first is obscured, and the second is underneath the third and fourth. The fifth, as in more primitive reptiles but in contrast with paliguanids, is nearly as long as the fourth, and shows no evidence of even incipient hooking. It does, however, articulate with the fourth TABLE 1 Dimension of appendicular skeleton of CoeluTOsauravus and Daedalosaurus (in cm) Humerus Ulna Ulna Radius Femur Tibia Fibula (total) (minus olecranon) C oelurosauravu.l Type a a 31, DaedaloJaurus 29, , ± 20± , In a, an ill-defined area probably representing the puboischiadic plate can be seen near the head of the femur, adjacent to the area of transition between the trunk and caudal vertebrae. No details can be seen. Most of the hind limb can be seen in this sp~cime n. The femur is approximately three per cent longer than the humerus, and has a somewhat thicker shaft. Details are obscured, but the bone can be interpreted as rather lizard-like in appearance, with a slightly sigmoidal curvature of the shaft and with the distal surface twisted anteroventrally. There appears to be a prominent internal trochanter arising a short distance distal to the head. There is certainly no trace of the adductor crest that distinguishes the ventral surface of the femur in many primitive tetrapods. The tibia and fibula are of nearly equal length, 66 per cent that of the femur. Both bones are lightly built with particularly narrow shafts. The distal end of the fibula is not greatly expanded. The foot (fig. 4) is exposed in ventral view, except for the third metatarsal which has rolled over to expose its dorsal surface. The definition of the individual elements is not especially good, the nature of the articulating surfaces poorly defined. The tarsus appears considerably more primitive than that of Sau _rostemon, with the ankle as a whole quite long relative to its width. The calcaneum is vaguely rect- as well as the fifth distal tarsal. One or more phalanges from each of the digits are preserved. There is only a single terminal phalanx, displaced but associated with the first digit because of its short distance from the tarsus. It is short, sharply pointed and recurved. Three phalanges of the fifth digit are in articulation and two from the second, the last of which are the longest. A phalangeal count of two, three, four, five, four can be confidently restored. Few, if any, ventral dermal scales are evident in either of the skeletons. DAEDALOSAURUS gen. nov. Type species, Daedalosaurus madagascariensis Diagnosis - Primitive reptile, apparently classifiable within the order Eosuchia, with trunk ribs nine through 26 greatly elongate to support a gliding membrane. Approximately 29 presacral vertebrae, transverse processes not greatly elongate. Ribs four through eight supporting thoracic cavity. Dentition pleurodont. Tall narrow scapula, not fenestrate anteriorly. Large cleithrum retaining dorsal blade. Sternum apparently unossified. Iliac blade expanded anteriorly to form a triangular plate. No evidence of ossified epiphyses.

8 150 Figure 5. Daedalosaurus madagascariensis, type, n. gen. et n. sp. Institut de Pah~ontologie, no Photograph of specimen in which the bone is represented largely as a natural cast. The portion including the skull and end of the tail is from the counterpart of the remainder of the specimen and is printed in reverse. Approximately X 1.
9

10 151 DAEDALOSAURUS MADAGASCARIENSIS sp. nov. Diagnosis - same as for genus. Type specimen - Nearly complete skeleton preserved in contiguous blocks numbered and and unnumbered, partial counterpart. Institut de Paleontologie, Museum National d'histoire Naturelle, Paris. HonLOn - Upper portion of the Lower Sakamena Formation, uppermost Permian. Locality - Vicinity of Mount Eliva, upper reaches of the Sakamena River Valley, South-western Madagascar. Etymology - Generic name in reference to Daedalus; according to Greek legend, the inventor of wings and the father of Icarus. The specific name refers to the island on which the fossil was found. Skull The skull can be seen on a small slab that was apparently the only portion to be collected of the counterpart of the block in which most of the skeleton is visible (figs. 5 and 7.) The orientation can be determined by the caudal vertebrae and the ends of three rit-s that are preserved in both blocks. Skull elements of Coelurosauravus are also preserved as counterparts in both blocks. The skull of Daedalosaurus appears to be displaced only slightly from its normal position relative to the cervical vertebrae. The posterior portion was unfortunately broken off, apparently at the time of collection to judge by the relative freshness of the break. The central and anterior part of the skull roof is visible in ventral view, together with disarticulated elements of the palate and cheek. Unfortunately the surface of the block was weathered so that details of the bone surface are not clear; identity of the disarticulated elements is thus uncertain. The right pre- and postfrontals, parietal, frontal and possibly a portion of the nasal are displayed. The upper temporal opening appears to be very lateral in position relative to the midline established by the frontal. This may be partially the result of flattening of the originally highly arched parietal into the plane of the rock, but probably also from lateral displacement of the bone relative to the frontal. The left maxilla is exposed medially. It resembles that of primitive captorhinomorph reptiles. The bone is incomplete anteriorly. The portion preserved bears 17 peg-like teeth, with space for two others. The teeth are set in a strongly developed medial ridge. Like those of most primitive reptiles, they are subpleurodont, with the lateral surface of the maxilla extending a short distance across their bases. Posteriorly the bone is notched for the jugal, which does not appear to have overlapped the bone extensively. As the skull is tentatively reassembled (fig. 8), there appears to be a considerable gap between the maxilla and prefrontal, indicating the presence of an extensive lachrymal. A triradiate bone in the area of the left cheek is almost certainly the postorbital, of the shape common to paliguanids and younginids. Beyond it is a thin bone which appears to be ~e right jugal in medial view and the adjacent portion of the pterygoid and ectopterygoid, reversed back to front. The jugal shows no posterior ramus, but the manner of preservation is such that it might originally have been present and yet not be exposed. Medially, the jugal shows a process for attachment to the ectopterygoid. The ectopterygoid is a stout, rounded bone, demonstrating the presence of a large suborbital fenestra in contrast to the plate-like ectopterygoid of captorhinomorphs. Farther away from the skull is a bone with limited exposure showing a small area of unfinished surface which may represent the region of the pterygoid that had articulated with the parasphenoid. Extending ventrally from the parietal is a thin plate of bone which is probably the dorsal portion of the prootic but not enough is exposed for description. Although the skull roof looks reasonably complete and some of the cheek elements are present, reliable reconstruction of the skull is not possible. There is nothing to compensate for the absence of the squamosal and quadrate in determining the nature of the temporal region. The pattern could resemble that of such taxonomically distinct forms as Youngina, Pro lacerta or Paliguana. Vertebrae Most of the vertebral column is obscured by the ribs as preserved (recalling that the specimen is preserved largely as a natural cast, these elements were lost with the missing portion of the counterpart block). In the restoration, the trunk vertebrae are drawn on the basis of those of Coelurosauravus since the features of the appendicular skeleton are very similar in the two genera. It may be significant that a similar elongation of the mid-trunk vertebrae also occurs in Icarosaurus and may be associated with flight in that genus. Only the cervical, thoracic, sacral and caudal vertebrae can be seen in Daedalosaurus. The main block is broken within the cervical series. Judging by its greater length, the first vertebra exposed is probably the axis. Nothing resembling an atlas is visible. The ribs of the first, second and third vertebra are short and single headed. This, together with their position anterior to the shoulder girdle, warrants designation of these vertebrae as cervicals. The next five vertebrae bear longer, presumably ventrally directed, ribs that supported the walls of the thoracic cavity. Behind them were at least 18 vertebrae bearing elongate ribs extending laterally from the column. The total presacral vertebral count appears to be 29. There are two sacral vertebrae, and 29 caudals are preserved, although the original length of the tail was probably considerably greater. The vertebrae are all amphicoelous. Vertebrae two through five are relatively well preserved in lateral view. Their general proportions resemble those of Coelurosauravus. The centra are relatively long and the neural spines tall and square BP - K

11 152 prf 1cm ~ ; po ~{., ~j'" -.~...., - pt ~.."....! r. /'''. art f / '. ~..' ".. ~.~: "~ " ~..:.~J pt A Figure 7. B Daedalosaurus TlUUiagascariensis. A, Drawing of latex cast of the counterpart of the block illustrated in Figure 6. B, End of tail illustrated in Figure 6. Numbers indicate position of caudal vertebrae; art f, facet of pterygoid for articulation with parasphenoid; ect, ectopterygoid; f, frontal; j, jugal (medial view); m, maxilla; n, nasal; p, parietal; pf, postfrontal; po, postorbital; prf, prefrontal; pro, prootic; pt, pterygoid. Maxilla with acrodont dentition, frontal and parietal below long ribs attributed to CoeluTosauravus. X 2.

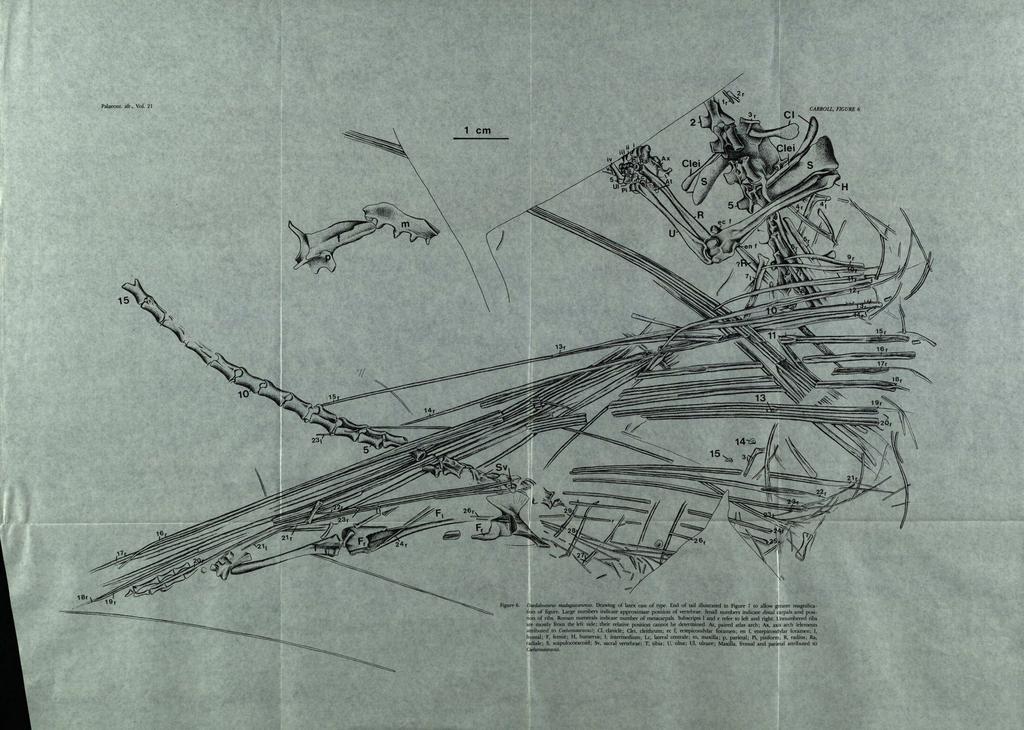
12 153 in outline. The transverse process of the second vertebra is not well preserved. The dorsal margin of the articulating surface of the third is oriented horizontally, at the end of a narrow, laterally directed process that extends beyond the level of the zygapophyses. The zygapophyses of this and the other anterior vertebrae are large, with the articulating surfaces flat and tilted at approximately 15 degrees from the horizontal. Vertebrae four through seven show a pattern of buttressing and excavation which can be attributed to selective forces acting to maximize the strength and minimize the mass. The zygapophyses are connected by a lateral ridge above the level of the transverse processes. The processes themselves are supported by three converging ridges; two extend horizontally beneath the ridge connecting the zygapophyses, and a third extends obliquely anteroventrally. The articulating surface is restricted to a sma1l oval at the end of the process. The transverse process of the eighth vertebra has a more normal configuration for a primitive reptile. The articulating surface is more extensive, and angles anteroventrally. It is thickest posterodorsally, and thins anteriorly. All that is visible of the more posterior trunk vertebrae is a series of transverse processes protruding between the ribs. Those that may be attributed to vertebrae 13, 14 and 15 show an articulating surface that appears horizontal dorsally. It is thick anteriorly, and tapers to the rear. Apparently the transverse processes of the trunk vertebrae were no longer than those more anterior in position, rather than being elongate as are those of Kuehneosaurus and [carosaurus. If the transverse processes were long, it is likely that they would have been oriented in the same plane as the remainder of the skeleton, rather than at right angles to it, and hence would have been preserved. Three poorly-exposed vertebrae can be seen just anterior to the sacrum. Their centra are short, as are those of the vertebrae just anterior to the sacrum of Coelurosauravus and [carosaurus. The two sacral vertebrae are incompletely exposed at the margin of the ilium, partially obscured by trunk ribs. The centra are short, and the neural spines low and rounded. Twenty-nine caudal vertebrae can be seen in sequence behind the sacrum. The first three are short, as are the sacrals, the centrum of the second approx- \ I I " Figure 8. Restoration of DaedaLosauruJ madagascariensis skull 1I1 lateral view, based on disarticulated elements. X 2. imately 3,7 mm in length., The length increases over the next three, after which the length is nearly constant (centra approximately 5 mm in length) to the end of the preserved series. The neural arches gradually decrease in height and width. The rreserved portion of the tail is nearly the length 0 the presacral column, but its original length may have been much greater. A striking feature of the tail is the apparent absence of long ribs fused to the transverse processes of the anterior vertebrae, a feature of both eosuchians and most lizards. This may be a result of the relatively poor preservation and difficulty of preparation in this area, but if these h4d been prominent features, it would be expected that they would be preserved in the plane of the block. The vertebrae in this area are, however, visible in lateral view, rather than dorsoventrally. Caudal vertebrae two and three show small, ventrolaterally directed articulating surfaces, low down on the anterior surface of the centrum, which may have accommodated unfused ribs, although none are visible. More posteriorly, there appear to be neither transverse processes nor ribs, although the vertebrae are well exposed. Unfused caudal ribs are characteristic of captorhinomorphs but not eosuchians, suggesting that Daedalosaurus is primitive in this regard. This feature may, in contrast, be a specialization, for the anterior transverse processes in Draco are extremely short. This may be associated with the great flexibility of the tail in relationship with flight control (Colbert, 1967). [carosaurus, however, has more normal transverse processes in this area. TABLE 2 Length of ribs supporting gliding membrane in DaedaLosauruJ (in em) Rib no. Length Rib no. Length Rib no. Length 9 1, ,4 23 9,1 10 2, ,0 24 7,5 II 2, ,2 25 3,9 (6,1 ) 12 4, ,8 26 4,7 13 6, ,7 27 1, , ,0 28 1, ,7 (II) 22 9,7 29 0,9 Numbers in parentheses indicate probable original length of incompletely preserved ribs. BP - L

13 154 For most of the column the presence or absence of trunk intercentra cannot be determined. It is possible that normal crescentic intercentra are present in the cervical region, but preservation is not good enough to determine this for certain. For much of the caudal region, intercentral ossifications are exposed. Only a few elongate haemal arches are observed. Like the apparent reduction of the caudal ribs this may contribute to a more efficient tail shape correlated with gliding. Ribs Ribs may be associated with all of the presacral vertebrae. Short, single-headed ribs are visible adjacent to the vertebrae recognized as the second and third cervicals, and one more anterior rib is probably from the missing atlas. These ribs are approximately three-fourths the length of the centra and were apparently oriented almost straight laterally to judge by the configuration of the rib head and the articulating surface of the transverse process. Ribs lying just behind the shoulder girdle have been identified as belonging with vertebrae four and five. They are much longer, have cylindrical or narrowly spatulate shafts, and the two heads are clearly separate. On the right side, ribs six, seven and eight are in articulation with their respective vertebrae. The lengths of the flattened shafts appear to increase progressively, although they are somewhat obscured by overlying elements. The width of the heads seems to narrow somewhat. It is probable that ribs four through eight extended ventrally to surround the chest cavity as in most primitive tetrapods. They are relatively long, to give a deep thoracic cavity. It is probable that they were continued ventrally in cartilage, and attached to a cartilagenous sternum, structures that are preserved in the contemporary (Permo-Triassic) paliguanid lizards. The orientation and configuration of the ribs changes dramatically behind the thoracic series, as in the Upper Triassic lizards Kuehneosaurus and Icarosaurus and the living agamid Draco; the trunk ribs were almost certainly elaborated to support a large gliding surface or "wing". From the ninth through 26th vertebra, the ribs of both left and right sides are, for the most part, extended laterally to the left of the column, those from the right side having been flipped over to expose their ventral surface. The ribs from the right side, for most of their length, overlie those from the left and appear to retain almost their normal position relative to one another, although the distance by which they are separated from each other in the fossil is almost certainly less than it was in life. Reconstruction of the original position and determination of the relative length of the ribs is extemely important in this form, but complicated by several factors. Where the ribs from both sides overlap one another, it is sometimes difficult to determine whether a particular segment is from the left or right side. Several of the upper ribs as seen in the specimen are incompletely exposed where they cross over the lower ribs and the sacral region, so that it is not always possible to associate proximal and distal portions of the same shaft. Ventral scales are plentiful in the specimen, and are of similar configuration to the smaller ribs and the terminal portions of the larger ribs. Nevertheless, the preservation of the right ribs is sufficiently complete that there was relatively little difficulty in assembling the "wing". Proximally, ribs nine through 26 from the right side all appear to be in their natural order, although a short gap separates numbers 20 and 21. The only serious difficulty in establishing the number and configuration lies with those most posterior in position. If it is assumed that all ribs lying more or less parallel to one another are from the right side, the count reaches 27, the most posterior of which is still very long. One of the posterior ribs, succeeding number 23, is angled slightly forward and beneath the others, suggesting that it may be from the left side. This suggests that the last long right rib present is the 26th. If three pairs of short presacral ribs (as seen on the left side) had originally been present, the presacral count would reach 29. That of Kuehneosaurus (Colbert, 1970, Table 1) is 28. In order to confirm the count on the right side, an attempt was made to determine the number present on the left, although they are much less well associated. None of the cervicals can be identified. Five disarticulated ribs, scattered through the trunk area (two of which have widely separated heads) may be from the thoracic region. None that are preserved can be identified with those making up the anterior margin of the right gliding surface: the ninth, tenth and 11 tho The 12th is only tentatively recognized. At least 15 other long ribs can be accounted for, although their specific location cannot be ascertained. One group of seven parallels the longest ribs from the right side, numbers 16 through 20. The proximal ends of these ribs are not exposed, however, so their total length cannot be established nor their original position specified. Two other ribs terminate with this group, but are displaced proximally. A second group of six originates at the level of the bases of ribs on the right side. They extend anteriorly at an angle of nearly 90 degrees to the first group. They appear complete proximally. The three longest ribs are only slightly shorter than the longest from the right side. From the established pattern of the anterior ribs on the right side, and the number of ribs accounted for on the left side, it would appear that the last long left rib is associated with the 26th vertebra. Three ribs, the last two of which are definitely short, and the first of which is incomplete distally, lie.iust anterior to the pubis, at right angles to the other trunk ribs. Since they underlie surrounding elements, they may be from the left side. This would account for a total of 29 ribs. Except for ribs 21, 22 and 26 from the right side, the proximal ends appear complete. There is little if
14

15 155 any differentiation of the head rrom the shaft and the articulating surfaces appear to be nearly flat. This surface appears to have been roughly oval in outline but this is difficult to ascertain due to crushing. The lateral extremities of the ribs are cylindrical and gradually diminish to a point. Proximally, the shaft is crushed, obscuring the original configuration. It is probable that this portion was originally a flattened oval, either hollow or filled with extremely cancellous bone. Crushing has resulted in the formation of a uniform linear groove, extending for much of the length of the shaft. Most of the ribs appear essentially straight. Ribs nine through 12 are curved slightly at the tip. It is difficult to determine whether the curvature was primarily posterior or ventral in the living animal. If ventral, it would have served to form a thick leading edge to the gliding surface, at least near the body. Similar curvature is seen in the 16th rib. Ribs 19 through 26 appear to show gentle curvature throughout their length, although the tip remains straight. The tips of ribs 15 and 25 on the right side, and 27 (from the left) are missing or obscured by other elements. When the ribs making up the wing are assembled in their natural order a plausible pattern is established, based on evidence from both sides. The length of ribs nine through 12 increases gradually. The length increases more rapidly from the 13th to 16th. The 18th is apparently the longest of all, with a length of approximately 16,2 cm. The 19th and 20th gradually diminish in length. The 21st is notably shorter, the remainder progressively so. The length of the 27th rib is uncertain. The last two are much shorter, but appear to be oriented laterally, extending beyond the expected limit of the trunk. The 29th is curved slightly posteriorly. Appendicular skeleton In contrast to the South African paliguanids, the shoulder girdle of Daedalosaurus shows no specifically lacertoid characteristics. It is surprisingly primitive in retaining a very large cleithrum. This bone is visible on both sides and shows a flattened dorsal portion extended posterodorsally over the margin of the scapula, as in primitive pelycosaurs. The shaft is a robust cylindrical rod, extending nearly to the level of the glenoid as preserved. The great length of the thoracic ribs, number four through eight, indicates the presence of a cartilagenous extrascapula, suggesting that the cleithrum had occupied a more dorsal position in life. The clavicle has a shorter stem, but with a somewhat widened blade that curved ventrally around the trunk. The interclavicle is not evident. The left half of the endochondral girdle is partially exposed in medial view beneath the anterior vertebrae. It appears to be ossified as a single unit. The scapular blade is very high and narrow, resembling that of primitive pelycosaurs. The coracoid area, seen 10 dorso-medial view on the left side, extends well behind the scapula, suggesting a relatively long glenoid, although little of this structure can be seen in this specimen. None of the foramina characteristic of the scapulocoracoid in other early reptiles can be recognized. The anterior margin of the left side of the pectoral girdle is at the level of the anterior end of the third centrum; that on the right, approximately one segment more posterior in position. Judging by the configuration of the "wing" and the relative position of these structures in Draco, the glenoid was probably at the level of the fifth and sixth centra. There is no evidence of an ossified sternum; a cartilaginous structure was probably present, however. The humerus is a long narrow bone, very similar to that of Coelurosauravus, with the ends twisted in a primitive fashion. The proximal articulating surface is long, narrow and clearly defined. It follows the pattern of primitive captorhinomorphs and pelycosaurs (Holmes, 1977) except for changes in proportions. There is no evidence of the lacertoid specialization seen in Paliguana (Carroll, 1977). The great width of the glenoid and humeral articulation suggests a retention of the sprawled posture of primitive reptiles, rather than the raised forelimb common to lizards. The shaft is long and slim. At the posterior margin is a narrow entepicondylar foramen. Anteriorly, the area of the ectepicondylar foramen is somewhat damaged, but this opening appears to be as in other early diapsids. Articulating surfaces for the ulna and radius are extremely well defined. Ventrally, the capitellum appears as a hemisphere, somewhat elongated in the direction of the long axis of the humerus. Dorsally, the distal surface resembles that of Coelurosauravus. In as far as proportions can be determined and details of structure seen, the ulna and radius of Daedalosaurus are identical with those of Coelurosauravus. The carpus of the right limb can be seen in ventral view. The elements are somewhat jumbled, but knowledge of the pattern common to captorhinids, eosuchians and primitive lizards allows identification of all the bones. Their specific orientation and configuration is more difficult to establish. The ulnare is especially poorly exposed. The intermedium, radiale and ulnare have considerable areas of finished bone. The other elements show primarily unfinished surface, making determination of the original orientation particularly difficult to determine. Judging from other primitive reptiles and modem lizards, the rounded appearance of the carpals is probably a result of incomplete ossification with the adult condition expected to show the pattern of a close fitting mosaic. The carpus appears much advanced from the configuration seen in the primitive diapsids Petrolacosaurus and Galesphyrus (Carroll, 1976), in its relatively limited extent, both proximodistally and mediolaterally, but the specific configuration of the bones does not approach the condition seen in lizards (Carroll, 1977). The proximal ends of

16 156 all four metacarpals are present. They overlap one another laterally when the carpus is restored to its natural width. The remainder of the hand is not preserved. In the restoration, it is shown with the phalanges of Coelurosauravus. The right half of the pelvic girdle is exposed in lateral view. Unlike that of most primitive lizards, but rather as in lcarosaurus, the iliac blade is broadly expanded above the acetabulum, especially anteriorly. This is surprising, in view of the limited degree of expansion of the sacral ribs. The pubis in lateral view appears very lacertoid, with the lateral expression of the pubic tubercle. It is tempting to restore a lizard-like thyroid fenestration, but this cannot be substantiated from the angle that the girdle is preserved. The pubic buttress of the acetabulum extends sharply laterally so as to limit the anterior swing of the femur. The ischium is largely obscured by the femur. The two femora are superposed and extend directly posteriorly from the girdle. Little can be seen, beyond the gross dimensions. Their length exceeds that of the humerus by only five per cent, in contrast to the relatively much longer femur in lcarosaurus and Kuehneosaurus. They are succeeded by slim tibiae 65 per cent the length of the femur. Bone is present in the area of the tarsus, but the individual elements cannot be distinguished. Numerous phalanges and?metatarsals can be seen, some in articulation with each other and others scattered about. They are similar to comparable elements in Coelurosauravus. Two articulated series compare with the fourth and fifth digits in that genus, although the individual elements are thinner. No impressions of squamate-like epidermal scales are evident on the block, although they are preserved with some of the eosuchian specimens from this horizon. Many ventral dermal scales are present. In comparison with those of most primitive reptiles, they appear very long. It is probable that they served to reinforce the abdominal wall in the absence of normal ventrally directed ribs. Here, the outdated term "ventral ribs" seems quite appropriate. Flight characteristics and comparison with other gliding genera A total of 21 pairs of ribs support the gliding surface in Daedalosaurus, a far greater number than that observed in other gliding lizards: five to seven in Draco, ten in Icarosaurus, eleven in Kuehneosaurus. The outline of the wing of Daedalosaurus appears intermediate between that of lcarosaurus and Draco. Its lateral extent, relative to other body proportions resembles that of lcarosaurus, but it has a greater anteroposterior extent, as does that of Draco. Ribs are present on all the presacral vertebrae in Daedalosaurus, but the posterior portion of the wing in Draco, Icarosaurus and Kuehneosaurus is without bony support. The ribs appear to differ markedly from those of Draco in their stiffness or lack of inherent curvature. In the modem genus, the ligaments attached to the ends of the ribs are very important in influencing the shape and overall configuration of the wing. In Daedalosaurus, as in Kuehneosaurus and lcarosaurus, the ribs appear to be the primary factor in defining the outline of the wing membrane. Restorations of the wing in all the gliding genera is shown in Figure 10. A major uncertainty in restoring the total area of the wing, and also the mass of the body, lies in the lack of direct information conceming the length of the posterior trunk vertebrae. Taking those of Coelurosauravus as a guide, the total snout-vent length is approximately 20 cm and the wing area is just less than 200 cm2. Mass can be estimated on the basis of a graph given by Pough (1973), comparing snoutvent length and body mass in modem lizards, assuming normal body proportions. On this basis, Daedalosaurus would have had a mass of approximately 250 grams. This does not take into consideration the mass of the wing itself, but this may have been compensated for by lightening of other parts of the skeleton, as is suggested by the configuration of the cervical vertebrae. As in Draco, the flight surface may have been augmented by other extensions of tissue, e.g. folds running along the side of the neck. Despite the much greater lateral extent of the membrane in Daedalosaurus, the wing load is very much higher, approximately 1,25 gr/cm2 compared with 0,23 gm/cm2 calculated for Icarosaurus and a range of 0,33 gm/cm2 to 0,76 gm/cm2 in Draco (Colbert, 1970). The greater wing load in Daedalosaurus does not in itself indicate that this genus was any less capable of gliding than Draco, since other factors greatly diminish the aerodynamic qualities in animals with very small wing surface. Colbert also cites an extreme amount of variability in relative wing size in Draco, suggesting that there appears to be no very critical mass to wing-area relationship. Wing load differences of up to 44 per cent are reported in animals of similar mass (Colbert, 1970). In Draco, the anterior margin of the wing is attached ventral to the general surface of the membrane, along the flank behind the forearm, thus providing a thick leading edge for the airfoil, at le~st near the body. The distal curvature of the first four ribs in Daedalosaurus, if ventrally oriented, may have served a similar purpose. Posteriorly, the wing in Draco attaches to the under surface of the thigh. The much longer posterior ribs in Daedalosaurus, if capable of folding or bending backwards, would have seriously interfered with movement of the rear limb if similarly attached. The posterior margin is here restored as ending on the dorsal surface of the thigh. For Draco, folding the wings is relatively simple since they are not very long, and the posterior margin is rounded so that none of the ribs are long enough to extend beyond the base of the rear limb. _ If the ribs of Daedalosaurus, or for that matter, Icarosaurus or Kuehneosaurus, are folded posteriorly, they
. All drawn to the same scale. pass above the rear limbs, well beyond the base of the tail.")
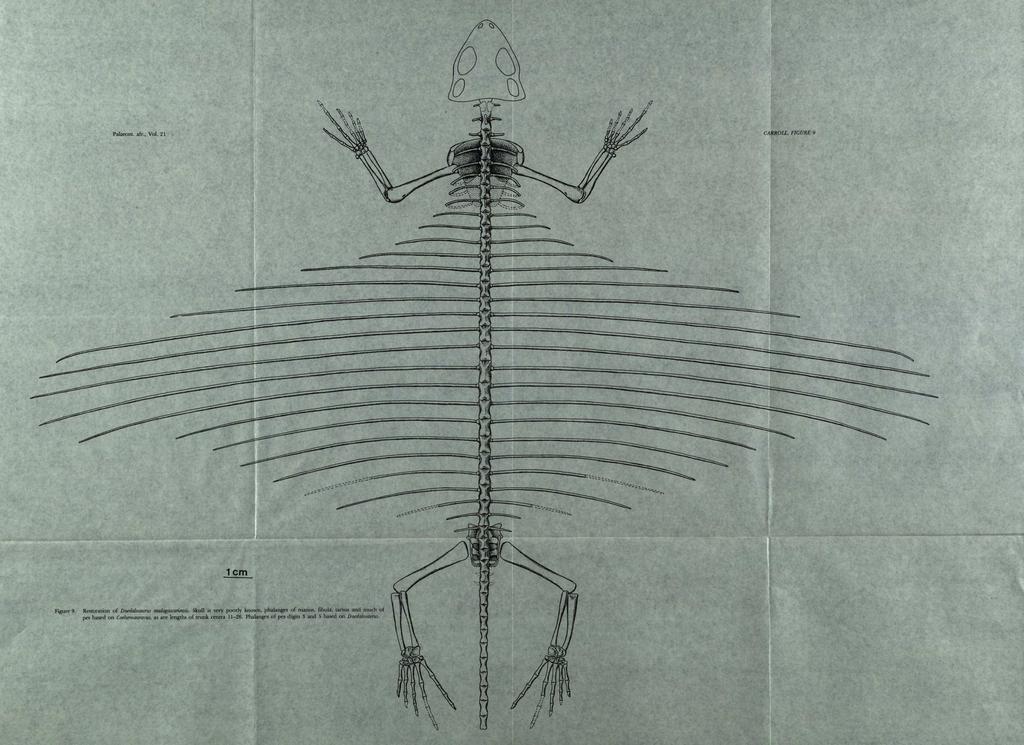
17 157 Figure 10. Restoration of gliding reptiles. A, Kuehneosaurus, Upper Triassic, modified!tom Robinson, printed in Romer (1966 ). B, lcaro.lauruj, Upper Triassic,!i'om Colbert (1970). C, Daeda!o.IGurus, Upper Permian. D, Draco, living genus, Iwm Colbert (1967). All drawn to the same scale. pass above the rear limbs, well beyond the base of the tail. Colbert has suggested that in Icarosaurus, the close association of the bases of the ribs would have restricted posterior folding and necessitated some degree of dorsal flexure. In Daedalosaurus, the transverse processes are all apparently short, as in Draco, and widely separated. The heads of the ribs are narrow but the nature of the articulating surfaces between the bones would have made folding in a posterior direction difficult. The transverse processes of vertebrae in the mid trunk region are flat dorsally, with the articulating surface thickest anteriorly and sloping to a rounded point posteriorly. This shape is only slightly modifiea from the pattern in other primitive reptiles and does not seem well suited to a posterior hinging of the ribs, if the wing is folded back in the manner of Draco. One might suggest from the nature of the transverse process and rib articulation in Daedalosaurus that the rib hinged dorsally. There is, however, no likely epaxial musculature that would have made such movement possible, and the antagonistic action of rib depressors would imply a totally improbable flapping flight. In Draco, the transverse processes, while not as long as those in Kuehneosaurus and lcarosaurus, extend well out from the arch and terminate in a rounded, evenly convex surface to which attached the end of the rib. Folding of the wings appears to

18 158 occur through both hinging at this joint and bending throughout the shaft of the ribs through the action of the ligaments. Because of the great width of the wing in Daedalosaurus, it is almost impossible to conceive of its being held rigidly from the body when not in use as a gliding membrane, but this genus may have had only limited ability to change its configuration. Although it is difficult to see beyond the flight characteristics of the "wings" in these genera, an extensive vascularized area of tissue such as this could not fail to have a striking effect on heat regulation as well. The living Draco is reported to use the brightly coloured surface of the wings in courtship displays (Colbert, 1970). THE TAXONOMIC POSITION OF COELUROSAURA VUS AND DAEDALOSAURUS Despite the clearly-defined differences in the dentition and the striking specialization of the ribs in Daedalosaurus, this genus and Coelurosauravus may be allocated to the same family on the basis of close similarities in the appendicular skeleton. In size and proportions, the elements of the forelimb are nearly indistinguishable and quite different from those described in other contemporary reptiles. To the extent that they are preserved in both genera, the pectoral girdle, vertebrae and rear limb are also similar. Romer (1968, p. 114) suggested the possibility that Coelurosauravus might be considered, along with Araeoscelis, as an early euryapsid, but admitted that adequate knowledge of the anatomy was lacking. Coelurosauravus differs from Araeoscelis in having a short rather than elongate neck, and in having acrodont rather than pleurodont teeth. There are otherwise no anatomical features of Coelurosauravus that suggest close affinity with nothosaurs or other, more specialized, euryapsids. A review of other members of the Permian Madagascar fauna demonstrates the presence of several genera that are plausible ancestors of nothosaurs and plesiosaurs. They are clearly diapsid derivatives, but show no important similarities to Coelurosauravus. The general configuration of the skeleton of Coelurosauravus and Daedalosaurus can be termed "lizard-like" in that they are small, with light bodies and long limbs. The skulls are poorly known but may well show the lacertoid pattern, with an upper temporal opening and a streptostylic quadrate. The pattern of the dentition in Coelurosauravus and the ribs in Daedalosaurus also emphasize lizard affinities, for both specializations may be seen in living members of the Lacertilia. Comparison with other contemporary Permo Triassic lizards, the paliguanids (Carroll, 1975 and 1977), demonstrates the absence of other structural and developmental aspects that appear basic to lizards as a group, however. Notably, Coelurosauravus and Daedalosaurus lack ossified epiphyses and specialized joint surfaces of the limbs. The sternum is unossified, and fenestration of the scapulocoracoid is not developed. The retention of a large c1eithrum also seems a very primitive feature. On the basis of our current understanding of primitive lizards and the possible origin of this group, these genera seem more conveniently placed among the Eosuchia - which remains an ill-defined assemblage of primitive lineages. It is reasonable to think that Coelurosauravus and Daedalosaurus are among the descendants of a more primitive assemblage that also gave rise to the Paliguanidae and hence to modem lizards. In a broad adaptive sense, the lizard habitus may be thought of as beginning with the eosuchian or even captorhinomorph level of structural sophistication. Hence we see in these forms an essentially modem type of dental and locomotor specialization together with very primitive features of the remainder of the skeleton. A further factor which should be considered in evaluatjng the taxonomic position of Daedalosaurus and Coelurosauravus is total body size. The estimated body mass for both is in the range of 200 to 300 grams. According to work by Pough (1973), primitive lizards rarely exceed a body mass of 150 grams unless they evolve the ability to feed primarily on plant material. Consequently most primitive lizards, fossil and living, are small insectivores. Members of more advanced lizard groups have developed structural or metabolic specializations that permit active predation on animals other than insects, and may reach a much larger body size. The relatively large body size of Daedalosaurus and Coelurosauravus is unexpected among primitive lizards, and suggests a different feeding and metabolic strategy. In common with most eosuchians which have a similar or even greater body size, they may have had a significantly lower metabolic rate, and correspondingly smaller food requirements. The living Tuatara (Farlow, 1975) may provide a useful analogy of a non-squamate lepidosaur, with low food requirements. On the basis of the known material of Daedalosaurus and Coelurosauravus, it may be assumed that the similarities of the appendicular skeleton are the inheritance from a close common ancestor. It seems probable that the gliding membrane evolved rapidly, with relatively little change in the remainder of the skeleton. All the trunk ribs are modified, in contrast with the greater regional specialization in the later genera, with fewer elongate ribs and the posterior trunk vertebrae without ribs at all. The vertebrae are excavated laterally, but the transverse processes are not elongate. It is possible that Coelurosauravus was adapted to an arboreal way of life, possibly including jumping from trees, that provided a structural pre-adaptation to the pattern of Daedalosaurus. The elongation of the vertebral column and the deep thoracic region may fall into this category.

19 159 It is natural to consider the possibility of relationship between DaedalosauTUS and the late Triassic kuehneosaurids. Unfortunately, this is made difficult by the lack of good cranial material of DaedalosauTUS and the limited information regarding the appendicular skeleton of Kuehneosaurus. On the basis of the available information, DaedalosaurUs seems more appropriately classified among the eosuchians. The skull of kuehneosaurids, on the other hand, unequivocally places them in the Lacertilia. The described elements of the appendicular skeleton, however, are generally more primitive than those of the Permo-Triassic paliguanid lizards. Some of the apparently primitive features, as the configuration of the endochondral shoulder girdle, may be specializations related to gliding. It is very difficult to evaluate whether such shared features are indicative of close relationship or to similarity in adaptation. Of the anatomical features known in all three genera, there are none that would, of themselves, preclude DaedalosauTUs having given rise to Kuehneosaurus and lcarosaurus. Such a derivation would imply, however, that most of the lacertoid features of the kuehneosaurids had evolved separately from those of the paliguanids and that the lacertoid grade of evolution had been achieved independently in the two families. It seems slightly more likely that the gliding adaptations were developed separately in the kuehneosaurids and the coelurosauravids in the light of the presence of both gliding and non-gliding genera in the latter family. Except for the extreme specialization of the ribs in DaedalosauTUs and the specialization of the dentition in Coelurosauravus, these genera may be included among the eosuchians on the basis of the generally primitive pattern of the limbs and pectoral girdle. If the paliguanids are classified among the Squamata, and the Prolactertiformes are no longer considered to be associated with the ancestry of lizards, the coelurosauravids may be the most lizard-like of the eosuchians. The presence of more specialized lacertoid features among the Upper Permian and Lower Triassic paliguanids (Carroll, 1977) indicates that this group must have diverged from primitive eosuchians by the early or middle Permian. Consequently, the known coelurosauravids are certainly too late to be considered ancestral to lizards. They appear, however, to retain primitive features which help us to visualize the nature of the ancestral group. ACKNOWLEDGEMENTS I wish to thank Professor Piveteau for encouraging me to study the Permian reptiles of Madagascar. His pioneering work in this fauna revealed an otherwise unexpected diversity among early diapsids. Professor Lehman has been very helpful in providing space and other facilities at the Institut de Paleontologie. Mr. D. Goujet deserves special thanks for locating specimens and arranging lor technical assistance. The photograph was taken by Mr. R. Kandarown and Mr. D. Serrette. Pamela Gaskill's painstaking illustration of the specimens is greatly appreciated. This study was an outgrowth of work on Karoo reptiles initiated in South Africa in The assistance of Drs. Cruickshank and Gow at the Bernard Price Institute and Drs. Barry and Cluver at the South African Museum is greatly appreciated. This work was supported by grants from the Faculty of Graduate Studies and Research, McGill University, the Merrill Trust, and the National Research Council of Canada. ADDENDUM Since completing this paper, my attention has been drawn to a paper by Schaumberg (1976 ) describing two newly discovered specimens from the Upper Permian of Germany. They are very poorly preserved, but clearly show the presence of ribs similar to those of DaedalosauTUl. Schaumberg suggests similarities with WeigeltisauTUl, originally described by Weigelt as Palaeochamaeleo. None of these specimens are sufficiently well-preserved or thoroughly enough described for detailed comparison, but all may have been closely related. BESAIRIE, H. (1972). Geologie de Madagascar I. Les terrains sedimentaires. Ann. Ceol. Mad. Fasc., 35, CARPENTIER, A. (1936). Additions a I'etude de la flore du Groupe de la Sakamena (Madagascar). Ann. Ceol. Servo des Mines (Madagascar), CARROLL, R. L. (1975 ). Permo-Triassic "lizards" from the Karroo. Palaeont. afr., 18, (1976). Calesphyrus capensis, a younginid eosuchian from the Cistecephalus zone of South Africa. Ann. S. Afr. Mus., 72, (1977 ). The origin of lizards, In: Miles, R. S., Andrews, S. M. and Walker, A., Eds., Problems in Vertebrate Evolution. Linnean Society Symposium Series No.4. COLBERT, E. H. (1967). Adaptations for gliding in the lizard Draco. Amer. Mus. Novitates, no. 2283, (1970). The Triassic gliding reptile Icarosaurus. Bull. Am. Mus. Nat. Hist., 143(2), FARLOW, J. O. (1975). Observations on a captive Tuatara '(Sphenodon punctatum). Jour. Herp., 9(4), GOUBIN, N. (1965). Description et repartition des principaux pollenites 'permiens, triassiques et jurassiques des sondages du bassin de Morondava (Madagascar). Rev. L'inst. Frarl.{ois Pitrole, 20(10), GOW, C. E. (1975). The morphology and relationships of Youngina capensis Broom and Prolacerta broorni. Parrington. Palaeon!. afr., 18, REFERENCES HART, G. F. (1969). The stratigraphic subdivision and equivalents of the Karroo sequence as suggested by palynology. Condwana Stratigraphy. lugs Symposium, UNESCO HOLMES, R. (1977). The osteology and musculature of the pectoral limb of small captorhinids. Jour. Morph., 152, LEHMAN, J.-P., CHATEAU, C., LAURAIN, M., AND NAUCH E, M. (1959). Paleontologie de Madagascar. XXVII : Les Poissons de la Sakamena moyene. Ann. Pal. (Paris), 45, PIVETEAU, J. (1926). Paleontologie de Madagascar, XIII. Amphibiens et reptiles permiens. Ann. Pal. (Paris), 15, POUGH, F. H. (1973). Lizard energetics and diet. Ecology, 54, PRIEM, F. (1924). Paleontologie de Madagascar, XII: Les poissons fossiles. Ann. Pal. (Paris), 13, 28 pp. ROBINSON, P. L. (1962). Gliding lizards from the Upper Keuper ot Great Britain. Proc. Ceol. Soc. London, no. 1601, ROMER, A. S. (1966). Vertebrate Paleontology, 3rd ed. Chicago, University of Chicago Press. 468 pp (1968). Notes and Comments on Vertebrate Paleontology. Chicago, University of Chicago Press. 304 pp. SCHAUMBERG, G. (1976). Zwei Reptilneufunde (Weigeltisau TUl KUHN [?), Lepidosauria [?J, Reptilia) aus dem Kupferschiefer von Richelsdorf (Perm, Hessen) Philippia III/1, 3-8.
Anatomy. Name Section. The Vertebrate Skeleton
 Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia
 1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
Mammalogy Laboratory 1 - Mammalian Anatomy
 Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A.
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
.56 m. (22 in.). COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition
. COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition") Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
A new species of sauropod, Mamenchisaurus anyuensis sp. nov.
 A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
AMERICAN MUSEUM NOVITATES Published by
 AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China
 SUPPLEMENTARY INFORMATION A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China Ya-Ming Wang 1, Hai-Lu You 2,3 *, Tao Wang 4 1 School of Earth Sciences and Resources, China
SUPPLEMENTARY INFORMATION A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China Ya-Ming Wang 1, Hai-Lu You 2,3 *, Tao Wang 4 1 School of Earth Sciences and Resources, China
1/9/2013. Divisions of the Skeleton: Topic 8: Appendicular Skeleton. Appendicular Components. Appendicular Components
 /9/203 Topic 8: Appendicular Skeleton Divisions of the Skeleton: Cranial Postcranial What makes up the appendicular skeleton? What is the pattern of serial homology of the limbs? Tetrapod front limb morphology
/9/203 Topic 8: Appendicular Skeleton Divisions of the Skeleton: Cranial Postcranial What makes up the appendicular skeleton? What is the pattern of serial homology of the limbs? Tetrapod front limb morphology
YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
 Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE
 PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province
 A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province by Hu Shaojin (Kunming Cultural Administrative Committee, Yunnan Province) Vertebrata PalAsiatica Vol. XXXI, No. 1
A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province by Hu Shaojin (Kunming Cultural Administrative Committee, Yunnan Province) Vertebrata PalAsiatica Vol. XXXI, No. 1
AMERICAN NATURALIST. Vol. IX. -DECEMBER, No. 12. OR BIRDS WITH TEETH.1 OI)ONTORNITHES,
ONTORNITHES,") AMERICAN NATURALIST. Vol. IX. -DECEMBER, 1875.-No. 12. OI)ONTORNITHES, OR BIRDS WITH TEETH.1 BY PROFESSOR 0. C. MARSH. REMAINS of birds are amono the rarest of fossils, and few have been discovered except
AMERICAN NATURALIST. Vol. IX. -DECEMBER, 1875.-No. 12. OI)ONTORNITHES, OR BIRDS WITH TEETH.1 BY PROFESSOR 0. C. MARSH. REMAINS of birds are amono the rarest of fossils, and few have been discovered except
ONLINE APPENDIX 1. Morphological phylogenetic characters scored in this paper. See Poe (2004) for
 for") ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC
 FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
UN? RSITYOF. ILLIiwiS LIBRARY AT URBANA-CHAMPAIGN NATURAL HIST. SURVEY
 UN? RSITYOF ILLIiwiS LIBRARY AT URBANA-CHAMPAIGN NATURAL HIST. SURVEY FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 July 29, 1954 No. 17 FAUNA OF THE VALE AND CHOZA: 7 PELYCOSAURIA:
UN? RSITYOF ILLIiwiS LIBRARY AT URBANA-CHAMPAIGN NATURAL HIST. SURVEY FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 July 29, 1954 No. 17 FAUNA OF THE VALE AND CHOZA: 7 PELYCOSAURIA:
2. Skull, total length versus length of the presacral vertebral column: (0); extremely elongated neck (e.g. Tanystropheus longobardicus).
; extremely elongated neck (e.g. Tanystropheus longobardicus).") Character list of the taxon-character data set 1. Skull and lower jaws, interdental plates: absent (0); present, but restricted to the anterior end of the dentary (1); present along the entire alveolar
Character list of the taxon-character data set 1. Skull and lower jaws, interdental plates: absent (0); present, but restricted to the anterior end of the dentary (1); present along the entire alveolar
The earliest reptiles
 J. Linn. SOC. (Zool), 45, no. 304, p. 61 With 14 tezt-figures Printed in Great Britain The earliest reptiles BY ROBERT L. CARROLL (Accepted for publication December 1963) Communicated by Errol I. White,
J. Linn. SOC. (Zool), 45, no. 304, p. 61 With 14 tezt-figures Printed in Great Britain The earliest reptiles BY ROBERT L. CARROLL (Accepted for publication December 1963) Communicated by Errol I. White,
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province
 from Dashanpu, Zigong Municipality, Sichuan Province") A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA
 A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA MYCTEROSAURUS LONGICEPS S. W. WILLISTON University of Chicago The past summer, Mr. Herman Douthitt, of the University of Chicago paleontological expedition,
A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA MYCTEROSAURUS LONGICEPS S. W. WILLISTON University of Chicago The past summer, Mr. Herman Douthitt, of the University of Chicago paleontological expedition,
THE SKULLS OF ARAEOSCELIS AND CASEA, PERMIAN REPTILES
 THE SKULLS OF REOSCELIS ND CSE, PERMIN REPTILES University of Chicago There are few Permian reptiles of greater interest at the present time than the peculiar one I briefly described in this journal' three
THE SKULLS OF REOSCELIS ND CSE, PERMIN REPTILES University of Chicago There are few Permian reptiles of greater interest at the present time than the peculiar one I briefly described in this journal' three
HONR219D Due 3/29/16 Homework VI
 Part 1: Yet More Vertebrate Anatomy!!! HONR219D Due 3/29/16 Homework VI Part 1 builds on homework V by examining the skull in even greater detail. We start with the some of the important bones (thankfully
Part 1: Yet More Vertebrate Anatomy!!! HONR219D Due 3/29/16 Homework VI Part 1 builds on homework V by examining the skull in even greater detail. We start with the some of the important bones (thankfully
Williston, and as there are many fairly good specimens in the American
 56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
SUPPLEMENTARY ONLINE MATERIAL FOR. Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor
 http://app.pan.pl/som/app61-ratsimbaholison_etal_som.pdf SUPPLEMENTARY ONLINE MATERIAL FOR Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor Ontogenetic changes in the craniomandibular
http://app.pan.pl/som/app61-ratsimbaholison_etal_som.pdf SUPPLEMENTARY ONLINE MATERIAL FOR Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor Ontogenetic changes in the craniomandibular
Lab 2 Skeletons and Locomotion
 Lab 2 Skeletons and Locomotion Objectives The objectives of this and next week's labs are to introduce you to the comparative skeletal anatomy of vertebrates. As you examine the skeleton of each lineage,
Lab 2 Skeletons and Locomotion Objectives The objectives of this and next week's labs are to introduce you to the comparative skeletal anatomy of vertebrates. As you examine the skeleton of each lineage,
LEIDY, SHOWING THE BONES OF THE FEET 'AND LIMBS
 CQNTEUBUTIONS FBOM THE MUSEUM OF PALEONTOLOGY (Confindion of Con&&&m froin UB Muaercm of Gcologg) UNIVERSITY OF ' MICHIGAN VOL V, No. 6, pp. 6W3 (e ph.) DEAXMBER 31,1036 A SPECIMEN OF STYLEMYS NEBRASCENSIS
CQNTEUBUTIONS FBOM THE MUSEUM OF PALEONTOLOGY (Confindion of Con&&&m froin UB Muaercm of Gcologg) UNIVERSITY OF ' MICHIGAN VOL V, No. 6, pp. 6W3 (e ph.) DEAXMBER 31,1036 A SPECIMEN OF STYLEMYS NEBRASCENSIS
On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds
 and the origin of birds") On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds by Qiang Ji and Shu an Ji Chinese Geological Museum, Beijing Chinese Geology Volume 233 1996 pp.
On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds by Qiang Ji and Shu an Ji Chinese Geological Museum, Beijing Chinese Geology Volume 233 1996 pp.
A NEW SALTICID SPIDER FROM VICTORIA By R. A. Dunn
 Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
PART FOUR: ANATOMY. Anatomy, Conformation and Movement of Dogs 41
 PART FOUR: ANATOMY Anatomy, Conformation and Movement of Dogs 41 ANATOMY The word anatomy is a scientific term that refers to the inner structure of the dog, comprising the muscles, skeleton and vital
PART FOUR: ANATOMY Anatomy, Conformation and Movement of Dogs 41 ANATOMY The word anatomy is a scientific term that refers to the inner structure of the dog, comprising the muscles, skeleton and vital
Recently Mr. Lawrence M. Lambe has described and figured in the
 56.81,9C(117:71.2) Article XXXV.-CORYTHOSAURUS CASUARIUS, A NEW CRESTED DINOSAUR FROM THE BELLY RIVER CRETA- CEOUS, WITH PROVISIONAL CLASSIFICATION OF THE FAMILY TRACHODONTIDA1X BY BARNUM BROWN. PLATE
56.81,9C(117:71.2) Article XXXV.-CORYTHOSAURUS CASUARIUS, A NEW CRESTED DINOSAUR FROM THE BELLY RIVER CRETA- CEOUS, WITH PROVISIONAL CLASSIFICATION OF THE FAMILY TRACHODONTIDA1X BY BARNUM BROWN. PLATE
A new carnosaur from Yongchuan County, Sichuan Province
 A new carnosaur from Yongchuan County, Sichuan Province by Dong Zhiming Institute of Vertebrate Palaeontology and Palaeoanthropology, Academia Sinica Zhang Yihong, Li Xuanmin, and Zhou Shiwu Chongqing
A new carnosaur from Yongchuan County, Sichuan Province by Dong Zhiming Institute of Vertebrate Palaeontology and Palaeoanthropology, Academia Sinica Zhang Yihong, Li Xuanmin, and Zhou Shiwu Chongqing
The cranial osteology of Belebey vegrandis (Parareptilia: Bolosauridae), from the Middle Permian of Russia, and its bearing on reptilian evolution
, from the Middle Permian of Russia, and its bearing on reptilian evolution") Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1511 191214 Original Articles RUSSIAN BOLOSAURID REPTILER. R. REISZ ET AL.
Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1511 191214 Original Articles RUSSIAN BOLOSAURID REPTILER. R. REISZ ET AL.
A Pterodactylus with Remains of Flight Membrane. by F. Broili (with 3 plates). Read at the Conference on 7th February 1925.
. Read at the Conference on 7th February 1925.") Broili, F. (1925) Ein Pterodactylus mit Resten der Flughaut. Sitzungsberichte der Bayerischen Königlichen Akademie der Wissenschaften, Mathematischen-Physicalischen Classe, 1925, 23-32. A Pterodactylus
Broili, F. (1925) Ein Pterodactylus mit Resten der Flughaut. Sitzungsberichte der Bayerischen Königlichen Akademie der Wissenschaften, Mathematischen-Physicalischen Classe, 1925, 23-32. A Pterodactylus
A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev Doklady Akademii Nauk, SSSR 87:
 translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * Dr. L.D. Boonstra. Paleontologist, South African Museum, Cape Town
 THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida. Evo-Devo Revisited. Development of the Tetrapod Limb
 Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida Evo-Devo Revisited Development of the Tetrapod Limb Limbs whether fins or arms/legs for only in particular regions or LIMB FIELDS. Primitively
Biology 340 Comparative Embryology Lecture 12 Dr. Stuart Sumida Evo-Devo Revisited Development of the Tetrapod Limb Limbs whether fins or arms/legs for only in particular regions or LIMB FIELDS. Primitively
v:ii-ixi, 'i':;iisimvi'\>!i-:: "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi L I E) R.ARY OF THE VERSITY U N I or ILLINOIS REMO
 R.ARY OF THE VERSITY U N I or ILLINOIS REMO") "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
"^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
SUPPLEMENTARY INFORMATION
 Character 155, interdental ridges. Absence of interdental ridge (0) shown in Parasaniwa wyomingensis (Platynota). Interdental ridges (1) shown in Coniophis precedens. WWW.NATURE.COM/NATURE 1 Character
Character 155, interdental ridges. Absence of interdental ridge (0) shown in Parasaniwa wyomingensis (Platynota). Interdental ridges (1) shown in Coniophis precedens. WWW.NATURE.COM/NATURE 1 Character
( M amenchisaurus youngi Pi, Ouyang et Ye, 1996)
") 39 4 2001 10 V ERTEBRATA PALASIATICA pp. 266 271 fig. 1,pl. I ( 643013), ( M amenchisaurus hochuanensis),,, Q915. 864 1995 12 31 (ZDM0126) ( M amenchisau rus hochuanensis Young et Chao, 1972),,, ZDM0126
39 4 2001 10 V ERTEBRATA PALASIATICA pp. 266 271 fig. 1,pl. I ( 643013), ( M amenchisaurus hochuanensis),,, Q915. 864 1995 12 31 (ZDM0126) ( M amenchisau rus hochuanensis Young et Chao, 1972),,, ZDM0126
MATERIAL The specimen of Glanosuchus macrops (SAM-PK K 7809) which forms the basis of this study was found in 1989 by
 which forms the basis of this study was found in 1989 by") The postcranial skeleton of the basal therocephalian Glanosuchus macrops (Scylacosauridae) and comparison of morphological and phylogenetic trends amongst the Theriodontia Heidi Fourie* & Bruce S. Rubidge
The postcranial skeleton of the basal therocephalian Glanosuchus macrops (Scylacosauridae) and comparison of morphological and phylogenetic trends amongst the Theriodontia Heidi Fourie* & Bruce S. Rubidge
complex in cusp pattern. (3) The bones of the coyote skull are thinner, crests sharper and the
 The bones of the coyote skull are thinner, crests sharper and the") DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
A Fossil Snake (Elaphe vulpina) From A Pliocene Ash Bed In Nebraska
 From A Pliocene Ash Bed In Nebraska") University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
Yimenosaurus, a new genus of Prosauropoda from Yimen County, Yunnan Province
 Yimenosaurus, a new genus of Prosauropoda from Yimen County, Yunnan Province by Ziqi Bai, Jie Yang, and Guohui Wang Yuxi Regional Administrative Academy of Yunnan Province Yuxiwenbo (Yuxi Culture and Scholarship)
Yimenosaurus, a new genus of Prosauropoda from Yimen County, Yunnan Province by Ziqi Bai, Jie Yang, and Guohui Wang Yuxi Regional Administrative Academy of Yunnan Province Yuxiwenbo (Yuxi Culture and Scholarship)
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL
 Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
FIELDIANA GEOLOGY NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA
 FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
A NEARLY COMPLETE TURTLE SKELETON FROM THE UPPER CRETACEOUS OF MONTANA
 CONTRIBUTIONS PBOM THE MUSEUM OF PALEONTOLOGY UNIVERSITY OF MICHIGAN VOL VI, No. 1. pp. 1-19 (18 figs.) D~c~arrrm 1, 1989 A NEARLY COMPLETE TURTLE SKELETON FROM THE UPPER CRETACEOUS OF MONTANA BY E. C.
CONTRIBUTIONS PBOM THE MUSEUM OF PALEONTOLOGY UNIVERSITY OF MICHIGAN VOL VI, No. 1. pp. 1-19 (18 figs.) D~c~arrrm 1, 1989 A NEARLY COMPLETE TURTLE SKELETON FROM THE UPPER CRETACEOUS OF MONTANA BY E. C.
List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to
 1 Supplementary data CHARACTER LIST List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to characters used by Tchernov et al. (2000), Rieppel, et al. (2002), and Lee
1 Supplementary data CHARACTER LIST List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to characters used by Tchernov et al. (2000), Rieppel, et al. (2002), and Lee
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO
 A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
ireican%mluseum A Gliding Reptile from the Triassic of New Jersey'
 A ireican%mluseum PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK 24, N.Y. NUMBER 2 246 MAY I9, I 966 A Gliding Reptile from the Triassic of New Jersey' BY
A ireican%mluseum PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK 24, N.Y. NUMBER 2 246 MAY I9, I 966 A Gliding Reptile from the Triassic of New Jersey' BY
UNIVERSITY OF ILLINOIS LIBRARY AT URBANA-CHAMPAIGN BIOLOGY. Hi 01^995
 UBRARY IttBMmXHALL f^bo 71995 UNIVERSITY OF ILLINOIS LIBRARY AT URBANA-CHAMPAIGN BIOLOGY Hi 01^995 590.5 FI n.s. No. 68 BIX.CM)I( ^v V > ' of Lacerta i Olivier Kic^'ikI Pubiitation 1437 PUBLJSHI-!)
UBRARY IttBMmXHALL f^bo 71995 UNIVERSITY OF ILLINOIS LIBRARY AT URBANA-CHAMPAIGN BIOLOGY Hi 01^995 590.5 FI n.s. No. 68 BIX.CM)I( ^v V > ' of Lacerta i Olivier Kic^'ikI Pubiitation 1437 PUBLJSHI-!)
SHEEPMEAT. Goatmeat primal preparation are the same specification and codes as Sheepmeat
 SHEEPMEAT Goatmeat primal preparation are the same specification and codes as Sheepmeat Item No. Page No. BONE-IN SHEEPMEAT Assorted Cuts... 5036...102 Breast and Flap... 5010...95 Breast and Flap Pieces...
SHEEPMEAT Goatmeat primal preparation are the same specification and codes as Sheepmeat Item No. Page No. BONE-IN SHEEPMEAT Assorted Cuts... 5036...102 Breast and Flap... 5010...95 Breast and Flap Pieces...
New Specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from Northeastern China
 from Northeastern China") PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3381, 44 pp., 31 figures, 2 tables August 16, 2002 New Specimens of Microraptor zhaoianus
PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3381, 44 pp., 31 figures, 2 tables August 16, 2002 New Specimens of Microraptor zhaoianus
BREVIORA LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1
, A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1") ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
Vol. XIV, No. 1, March, The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S.
 By S.") Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
On the morphoplogy and taxonomic status of Xinpusaurus kohi JIANG et al., 2004 (Diapsida: Thalattosauria) from the Upper Triassic of China
 from the Upper Triassic of China") Palaeodiversity 7: 47 59; Stuttgart 30 December 2014. 47 On the morphoplogy and taxonomic status of Xinpusaurus kohi JIANG et al., 2004 (Diapsida: Thalattosauria) from the Upper Triassic of China MICHAEL
Palaeodiversity 7: 47 59; Stuttgart 30 December 2014. 47 On the morphoplogy and taxonomic status of Xinpusaurus kohi JIANG et al., 2004 (Diapsida: Thalattosauria) from the Upper Triassic of China MICHAEL
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE
 FROM THE") A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
Animal Form and Function. Amphibians. United by several distinguishing apomorphies within the Vertebrata
 Animal Form and Function Kight Amphibians Class Amphibia (amphibia = living a double life) United by several distinguishing apomorphies within the Vertebrata 1. Skin Thought Question: For whom are integumentary
Animal Form and Function Kight Amphibians Class Amphibia (amphibia = living a double life) United by several distinguishing apomorphies within the Vertebrata 1. Skin Thought Question: For whom are integumentary
A new Middle Jurassic sauropod subfamily (Klamelisaurinae subfam. nov.) from Xinjiang Autonomous Region, China
 from Xinjiang Autonomous Region, China") A new Middle Jurassic sauropod subfamily (Klamelisaurinae subfam. nov.) from Xinjiang Autonomous Region, China by Xijing Zhao Institute of Vertebrate Paleontology and Paleoanthropology, Academia Sinica
A new Middle Jurassic sauropod subfamily (Klamelisaurinae subfam. nov.) from Xinjiang Autonomous Region, China by Xijing Zhao Institute of Vertebrate Paleontology and Paleoanthropology, Academia Sinica
AMERICAN MUSEUM NOVITATES
 AMERICAN MUSEUM NOVITATES Published by Number 89 THE AmERcAN Mueum OF NATuRAL HIsTORY October 11, 1923 New York City 56.81,9. PRELIMINARY NOTICES OF SKELETONS AND SKULLS OF DEINODONTIDE FROM THE CRETACEOUS
AMERICAN MUSEUM NOVITATES Published by Number 89 THE AmERcAN Mueum OF NATuRAL HIsTORY October 11, 1923 New York City 56.81,9. PRELIMINARY NOTICES OF SKELETONS AND SKULLS OF DEINODONTIDE FROM THE CRETACEOUS
APPENDIX. 344 Mni-s/i Restorations of Claosaurus and Geratosaurus.
 344 Mni-s/i Restorations of Claosaurus and Geratosaurus. Claosaurics, Marsh, 1890.* The most important feature in the restoration of Claosaurus annectens given on Plate VI is the skull, which will be fully
344 Mni-s/i Restorations of Claosaurus and Geratosaurus. Claosaurics, Marsh, 1890.* The most important feature in the restoration of Claosaurus annectens given on Plate VI is the skull, which will be fully
NOTES ON THE FIRST SKULL AND JAWS OF RIOJASAURUS INCERTUS (DINOSAURIA, PROSAUROPODA, MELANOROSAURIDAE) OF THE LATE TRIASSIC OF LA RIOJA, ARGENTINA
 OF THE LATE TRIASSIC OF LA RIOJA, ARGENTINA") NOTES ON THE FIRST SKULL AND JAWS OF RIOJASAURUS INCERTUS (DINOSAURIA, PROSAUROPODA, MELANOROSAURIDAE) OF THE LATE TRIASSIC OF LA RIOJA, ARGENTINA José F. Bonaparte and José A. Pumares translated by Jeffrey
NOTES ON THE FIRST SKULL AND JAWS OF RIOJASAURUS INCERTUS (DINOSAURIA, PROSAUROPODA, MELANOROSAURIDAE) OF THE LATE TRIASSIC OF LA RIOJA, ARGENTINA José F. Bonaparte and José A. Pumares translated by Jeffrey
Jurassic Ornithopod Agilisaurus louderbacki (Ornithopoda: Fabrosauridae) from Zigong, Sichuan, China
 from Zigong, Sichuan, China") Jurassic Ornithopod Agilisaurus louderbacki (Ornithopoda: Fabrosauridae) from Zigong, Sichuan, China Guangzhao Peng (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 30, No. 1 January, 1992 pp. 39-51
Jurassic Ornithopod Agilisaurus louderbacki (Ornithopoda: Fabrosauridae) from Zigong, Sichuan, China Guangzhao Peng (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 30, No. 1 January, 1992 pp. 39-51
TRACHEMYS SCULPTA. A nearly complete articulated carapace and plastron of an Emjdd A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE,
 A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
'Rain' of dead birds on central NJ lawns explained; Federal culling program killed up to 5,000 Associated Press, January 27, 2009
 'Rain' of dead birds on central NJ lawns explained; Federal culling program killed up to 5,000 Associated Press, January 27, 2009 Study May Give Hope That Ivory-billed Woodpeckers Still Around Science
'Rain' of dead birds on central NJ lawns explained; Federal culling program killed up to 5,000 Associated Press, January 27, 2009 Study May Give Hope That Ivory-billed Woodpeckers Still Around Science
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the
 Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the") Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
Description of Cranial Elements and Ontogenetic Change within Tropidolaemus wagleri (Serpentes: Crotalinae).
.") East Tennessee State University Digital Commons @ East Tennessee State University Electronic Theses and Dissertations 5-2016 Description of Cranial Elements and Ontogenetic Change within Tropidolaemus
East Tennessee State University Digital Commons @ East Tennessee State University Electronic Theses and Dissertations 5-2016 Description of Cranial Elements and Ontogenetic Change within Tropidolaemus
Comparative Osteology of the Genus Pachytriton (Caudata: Salamandridae) from Southeastern China
 from Southeastern China") Asian Herpetological Research 2012, 3(2): 83 102 DOI: 10.3724/SP.J.1245.2012.00083 Comparative Osteology of the Genus Pachytriton (Caudata: Salamandridae) from Southeastern China Yunke WU 1, Yuezhao WANG
Asian Herpetological Research 2012, 3(2): 83 102 DOI: 10.3724/SP.J.1245.2012.00083 Comparative Osteology of the Genus Pachytriton (Caudata: Salamandridae) from Southeastern China Yunke WU 1, Yuezhao WANG
A Complete Late Cretaceous Iguanian (Squamata, Reptilia) from the Gobi and Identification of a New Iguanian Clade
 from the Gobi and Identification of a New Iguanian Clade") PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3584, 47 pp., 19 figures September 6, 2007 A Complete Late Cretaceous Iguanian (Squamata,
PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3584, 47 pp., 19 figures September 6, 2007 A Complete Late Cretaceous Iguanian (Squamata,
NIVOROUS DINOSAUR. (SECOND COMMUNICATION.) By HENRY FAIRFIELD OSBORN. PLATE XXXIX. This great carnivorous Dinosaur of the Laramie was contemporary
 By HENRY FAIRFIELD OSBORN. PLATE XXXIX. This great carnivorous Dinosaur of the Laramie was contemporary") 56, 8i, 9 T (I 7: 786) Article VI.-TYRANNOSAURUS, UPPER CRETACEOUS CAR- NIVOROUS DINOSAUR. (SECOND COMMUNICATION.) By HENRY FAIRFIELD OSBORN. PLATE I. This great carnivorous Dinosaur of the Laramie was
56, 8i, 9 T (I 7: 786) Article VI.-TYRANNOSAURUS, UPPER CRETACEOUS CAR- NIVOROUS DINOSAUR. (SECOND COMMUNICATION.) By HENRY FAIRFIELD OSBORN. PLATE I. This great carnivorous Dinosaur of the Laramie was
Supplementary information to A new troodontid dinosaur from China with avian-like sleeping-posture. Xing Xu 1 and Mark Norell 2
 Supplementary information to A new troodontid dinosaur from China with avian-like sleeping-posture Xing Xu 1 and Mark Norell 2 1 Institute of Vertebrate Paleontology & Paleoanthropology, Chinese Academy
Supplementary information to A new troodontid dinosaur from China with avian-like sleeping-posture Xing Xu 1 and Mark Norell 2 1 Institute of Vertebrate Paleontology & Paleoanthropology, Chinese Academy
The Portuguese Podengo Pequeno
 The Portuguese Podengo Pequeno Presented by the Portuguese Podengo Pequenos of America, Inc For more information go to www.pppamerica.org HISTORY A primitive type dog, its probable origin lies in the ancient
The Portuguese Podengo Pequeno Presented by the Portuguese Podengo Pequenos of America, Inc For more information go to www.pppamerica.org HISTORY A primitive type dog, its probable origin lies in the ancient
Biology 204 Summer Session 2005
 Biology 204 Summer Session 2005 Mid-Term Exam 7 pages ANSWER KEY ***** This is exam is worth 10% of your final grade****** The class average was 54% Time to start studying for your final exam!!! The answer
Biology 204 Summer Session 2005 Mid-Term Exam 7 pages ANSWER KEY ***** This is exam is worth 10% of your final grade****** The class average was 54% Time to start studying for your final exam!!! The answer
Cretaceous, toothed pterosaurs from Brazil. A reappraisal
 5. Preliminary description of a skull and wing of a Brazilian Cretaceous (Santana Formation; Aptian Albian) pterosaur (Pterodactyloidea) in the collection of the AMNH 34 5.1. Introduction The collection
5. Preliminary description of a skull and wing of a Brazilian Cretaceous (Santana Formation; Aptian Albian) pterosaur (Pterodactyloidea) in the collection of the AMNH 34 5.1. Introduction The collection
Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved
 Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved This was a private report in 2003 on my thoughts on Platecarpus planifrons.
Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved This was a private report in 2003 on my thoughts on Platecarpus planifrons.
A NEW CROCODYLOMORPH ARCHOSAUR FROM THE UPPER TRIASSIC OF NORTH CAROLINA
 Journal of Vertebrate Paleontology 23(2):329 343, June 2003 2003 by the Society of Vertebrate Paleontology A NEW CROCODYLOMORPH ARCHOSAUR FROM THE UPPER TRIASSIC OF NORTH CAROLINA HANS-DIETER SUES 1 *,
Journal of Vertebrate Paleontology 23(2):329 343, June 2003 2003 by the Society of Vertebrate Paleontology A NEW CROCODYLOMORPH ARCHOSAUR FROM THE UPPER TRIASSIC OF NORTH CAROLINA HANS-DIETER SUES 1 *,
ALFRED GILLETT AND FOSSILS FROM STREET
 ALFRED GILLETT AND FOSSILS FROM STREET This collection of local fossils was formerly in the Crispin Hall, Street. Most of these fossils came from Alfred Gillett (1814-1904), a retired ironmonger who lived
ALFRED GILLETT AND FOSSILS FROM STREET This collection of local fossils was formerly in the Crispin Hall, Street. Most of these fossils came from Alfred Gillett (1814-1904), a retired ironmonger who lived
Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported
 Drawing of early ankle development of Alligator mississippiensis, as reported") Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported by a previous study 1. The intermedium is formed at
Supplementary Figure 1 Cartilaginous stages in non-avian amniotes. (a) Drawing of early ankle development of Alligator mississippiensis, as reported by a previous study 1. The intermedium is formed at
CHARACTER LIST: Nesbitt et al., 2011
 CHARACTER LIST: Nesbitt et al., 2011 1. Vaned feathers on forelimb symmetric (0) or asymmetric (1). The barbs on opposite sides of the rachis differ in length; in extant birds, the barbs on the leading
CHARACTER LIST: Nesbitt et al., 2011 1. Vaned feathers on forelimb symmetric (0) or asymmetric (1). The barbs on opposite sides of the rachis differ in length; in extant birds, the barbs on the leading
By HENRY FAIRFIELD OSBORN.
 Article XI.-FORE AND HINI) LIMBS OF CARNIVOR- OUS AND HERBIVOROUS DINOSAURS FROM THE JURASSIC OF WYOMING. DINOSAUR CONTRIBU- TIONS, NO. 3. By HENRY FAIRFIELD OSBORN. In the Bone Cabin Quarry, opened by
Article XI.-FORE AND HINI) LIMBS OF CARNIVOR- OUS AND HERBIVOROUS DINOSAURS FROM THE JURASSIC OF WYOMING. DINOSAUR CONTRIBU- TIONS, NO. 3. By HENRY FAIRFIELD OSBORN. In the Bone Cabin Quarry, opened by
OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE
 OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT This is a report of measurements on the skeleton of a male se1 whale caught in the Antarctic. The skeleton of
OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT This is a report of measurements on the skeleton of a male se1 whale caught in the Antarctic. The skeleton of
First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia
 from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia") First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia Tsogtbaatar Chinzorig¹, ³ *, Yoshitsugu Kobayashi², Khishigjav Tsogtbaatar³,
First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia Tsogtbaatar Chinzorig¹, ³ *, Yoshitsugu Kobayashi², Khishigjav Tsogtbaatar³,
Discovery of an Avialae bird from China, Shenzhouraptor sinensis gen. et sp. nov.
 Discovery of an Avialae bird from China, Shenzhouraptor sinensis gen. et sp. nov. by Qiang Ji 1, Shuan Ji 2, Hailu You 1, Jianping Zhang 3, Chongxi Yuan 3, Xinxin Ji 4, Jinglu Li 5, and Yinxian Li 5 1.
Discovery of an Avialae bird from China, Shenzhouraptor sinensis gen. et sp. nov. by Qiang Ji 1, Shuan Ji 2, Hailu You 1, Jianping Zhang 3, Chongxi Yuan 3, Xinxin Ji 4, Jinglu Li 5, and Yinxian Li 5 1.
TWO NEW PINE-FEEDING SPECIES OF COLEOTECHNITES ( GELECHIIDAE )
") Journal of the Lepidopterists' Society 32(2), 1978, 118-122 TWO NEW PINE-FEEDING SPECIES OF COLEOTECHNITES ( GELECHIIDAE ) RONALD W. HODGES l AND ROBERT E. STEVENS2 ABSTRACT. Two new species of moths,
Journal of the Lepidopterists' Society 32(2), 1978, 118-122 TWO NEW PINE-FEEDING SPECIES OF COLEOTECHNITES ( GELECHIIDAE ) RONALD W. HODGES l AND ROBERT E. STEVENS2 ABSTRACT. Two new species of moths,
THE MOUNTED SKELETONS OF CMIPTOSAURUS IN THE
 THE MOUNTED SKELETONS OF CMIPTOSAURUS IN THE UNITED STATES NATIONAL I^IUSEUM. By Charles W. Gilmore. Assistant Curator of Fossil Reptiles, United States National Museum. INTRODUCTION. Recently the exliibition
THE MOUNTED SKELETONS OF CMIPTOSAURUS IN THE UNITED STATES NATIONAL I^IUSEUM. By Charles W. Gilmore. Assistant Curator of Fossil Reptiles, United States National Museum. INTRODUCTION. Recently the exliibition
Comments on the Beauceron Standard By M. Maurice Hermel (Translated by C. Batson)
") Comments on the Beauceron Standard By M. Maurice Hermel (Translated by C. Batson) The following are comments written by M. Hermel for the FCI Standard #44 published on 10/25/06. They were approved by the
Comments on the Beauceron Standard By M. Maurice Hermel (Translated by C. Batson) The following are comments written by M. Hermel for the FCI Standard #44 published on 10/25/06. They were approved by the
CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN OF RUSSIA AND THE EVOLUTIONARY RELATIONSHIPS OF CASEIDAE
 FROM THE MIDDLE PERMIAN OF RUSSIA AND THE EVOLUTIONARY RELATIONSHIPS OF CASEIDAE") Journal of Vertebrate Paleontology 28(1):160 180, March 2008 2008 by the Society of Vertebrate Paleontology ARTICLE CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN
Journal of Vertebrate Paleontology 28(1):160 180, March 2008 2008 by the Society of Vertebrate Paleontology ARTICLE CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN
OCCASIONAL PAPERS OF THE MUSEUM OF ZOOLOGY UNIVERSITY OF MICHIGAN
 OCCASIONAL PAPERS OF THE MUSEUM OF ZOOLOGY ~- UNIVERSITY OF MICHIGAN A NEW FROG FROM BRITISH GUIANA A collection received by the IIuseum of Zoology froin British Gniana some time ago includes a single
OCCASIONAL PAPERS OF THE MUSEUM OF ZOOLOGY ~- UNIVERSITY OF MICHIGAN A NEW FROG FROM BRITISH GUIANA A collection received by the IIuseum of Zoology froin British Gniana some time ago includes a single
The family Gnaphosidae is a large family
 Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
Where have all the Shoulders gone?
 Where have all the Shoulders gone? Long time passing Where have all the shoulders gone Long time ago "Correct" fronts are the hardest structural trait to keep in dogs. Once correct fronts are lost from
Where have all the Shoulders gone? Long time passing Where have all the shoulders gone Long time ago "Correct" fronts are the hardest structural trait to keep in dogs. Once correct fronts are lost from
FSS OPEN SHOW PROCEDURAL EXAM
 Judging Operations Department PO Box 900062 Raleigh, NC 27675-9062 (919) 816-3570 judgingops@akc.org www.akc.org Revised Sept 2013 FSS OPEN SHOW PROCEDURAL EXAM Refer to Rules, Policies and Guidelines
Judging Operations Department PO Box 900062 Raleigh, NC 27675-9062 (919) 816-3570 judgingops@akc.org www.akc.org Revised Sept 2013 FSS OPEN SHOW PROCEDURAL EXAM Refer to Rules, Policies and Guidelines
A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.)
") A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.) by Ouyang Hui Zigong Dinosaur Museum Newsletter Number 2 1989 pp. 10-14 Translated By Will Downs Bilby
A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.) by Ouyang Hui Zigong Dinosaur Museum Newsletter Number 2 1989 pp. 10-14 Translated By Will Downs Bilby
UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA
 FROM JAVA, INDONESIA") NOTES AND NEWS UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA BY NGUYEN NGOC-HO i) Faculty of Science, University of Saigon, Vietnam Among material recently collected
NOTES AND NEWS UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA BY NGUYEN NGOC-HO i) Faculty of Science, University of Saigon, Vietnam Among material recently collected
A new species of torrent toad (Genus Silent Valley, S. India
 Proc. Indian Acad. Sci. (Anirn. ScL), Vol. 90, Number 2, March 1981, pp. 203-208. Printed in India. A new species of torrent toad (Genus Silent Valley, S. India Allsollia) from R S PILLAI and R PATTABIRAMAN
Proc. Indian Acad. Sci. (Anirn. ScL), Vol. 90, Number 2, March 1981, pp. 203-208. Printed in India. A new species of torrent toad (Genus Silent Valley, S. India Allsollia) from R S PILLAI and R PATTABIRAMAN
A Late Jurassic Protosuchian Sichuanosuchus huidongensis from Zigong, Sichuan Province. Guangzhao Peng. Zigong Dinosaur Museum, Zigong, Sichuan
 A Late Jurassic Protosuchian Sichuanosuchus huidongensis from Zigong, Sichuan Province Guangzhao Peng Zigong Dinosaur Museum, Zigong, Sichuan 643013 Vertebrata PalAsiatica Volume 34, Number 4 October,
A Late Jurassic Protosuchian Sichuanosuchus huidongensis from Zigong, Sichuan Province Guangzhao Peng Zigong Dinosaur Museum, Zigong, Sichuan 643013 Vertebrata PalAsiatica Volume 34, Number 4 October,
AMERICAN MUSEUM. Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET
 AMERICAN MUSEUM Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET NEW YORK, N.Y. 10024 U.S.A. NUMBER 2662 NOVEMBER 21, 1978 RONN W. COLDIRON Acroplous vorax
AMERICAN MUSEUM Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET NEW YORK, N.Y. 10024 U.S.A. NUMBER 2662 NOVEMBER 21, 1978 RONN W. COLDIRON Acroplous vorax
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS
 AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.
Appendix chapter 2: Description of Coloborhynchus spielbergi sp. nov. (Pterodactyloidea) from the Albian (Lower Cretaceous) of Brazil
 from the Albian (Lower Cretaceous) of Brazil") Appendix chapter 2: Description of Coloborhynchus spielbergi sp. nov. (Pterodactyloidea) from the Albian (Lower Cretaceous) of Brazil Appendix chapter 2 155 2.7. Appendix 2.7.1. Measurements Skull 15 12
Appendix chapter 2: Description of Coloborhynchus spielbergi sp. nov. (Pterodactyloidea) from the Albian (Lower Cretaceous) of Brazil Appendix chapter 2 155 2.7. Appendix 2.7.1. Measurements Skull 15 12
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature14472 1. Hypodigm for Pappochelys rosinae Holotype: SMNS 91360, partially articulated incomplete postcranial skeleton (Fig. 1a,b; EDFig. 5). Paratype: SMNS 90013, disarticulated skeleton
doi:10.1038/nature14472 1. Hypodigm for Pappochelys rosinae Holotype: SMNS 91360, partially articulated incomplete postcranial skeleton (Fig. 1a,b; EDFig. 5). Paratype: SMNS 90013, disarticulated skeleton
Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons
 Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons 1. Head skeleton of lamprey Cyclostomes are highly specialized in both the construction of the chondrocranium and visceral skeleton.
Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons 1. Head skeleton of lamprey Cyclostomes are highly specialized in both the construction of the chondrocranium and visceral skeleton.