ECOLOGY OF AN URBAN POPULATION OF THE TEXAS HORNED LIZARD (PHRYNOSOMA CORNUTUM) IN CENTRAL OKLAHOMA DEBORA A. ENDRISS. Bachelor of Science
|
|
|
- Shavonne James
- 6 years ago
- Views:
Transcription
1 ECOLOGY OF AN URBAN POPULATION OF THE TEXAS HORNED LIZARD (PHRYNOSOMA CORNUTUM) IN CENTRAL OKLAHOMA By DEBORA A. ENDRISS Bachelor of Science State University of New York College of Environmental Science and Forestry Syracuse, New York 2003 Submitted to the Faculty of the Graduate College of the Oklahoma State University in partial fulfillment of the requirements for the Degree of MASTER OF SCIENCE July, 2006
2 ECOLOGY OF AN URBAN POPULATION OF THE TEXAS HORNED LIZARD (PHRYNOSOMA CORNUTUM) IN CENTRAL OKLAHOMA Thesis Approved: Dr. Stanley F. Fox Thesis Advisor Dr. Eric C. Hellgren Dr. Mahesh N. Rao Dr. A. Gordon Emslie Dean of the Graduate College ii
3 ACKNOWLEDGMENTS I would like to express my thanks to numerous individuals who supported my project and aided me in its completion. I wish to express my appreciation to my major advisors, Dr. Stanley Fox and Dr. Eric Hellgren, for their advice and guidance during data collection and in the preparation of my thesis. My appreciation extends to my third committee member, Dr. Mahesh Rao, for sharing his GIS expertise with me. I thank Ray Moody and John Krupovage at Tinker Air Force Base for their help and support during data collection. I also am grateful to Dr. Chip Leslie and the Oklahoma Cooperative Fish and Wildlife Unit for supporting this project and for providing me access to the Compos Analysis software. I also wish to acknowledge Tinker Air Force Base for funding this research and the Zoology Department of OSU for funding me when I was not in the field. I also wish to express my appreciation to individuals who aided with various aspects of this project. Joe Hackler collected data during the summer of 2003 and worked out many kinks in the project before I began. Corina Wack spent many hours in the field with me during the summer of 2005 while collecting data for her project and I enjoyed the time we spent together. I also thank her for collecting telemetry data for me while I was away from the study site and her aid in collecting data for the mark-recapture portion of my project. I am grateful to David Walter, who shared his GIS and statistical knowledge with me, sometimes numerous times a day. Dr. Richard Grantham aided me in identification of lizard stomach contents. Dr. Dan Shoup helped me with application iii
4 of Euler s equation. Finally, Verl Emrick, Jessica Dorr, as well as other individuals from Virginia Tech, created a vegetation map of the study site that was immensely valuable to this project. A special thanks to my mother, Cynthia Endriss, my sister, Karen Endriss, Linda Pratka, Timothy Periard, Gavin, and Zoology graduate students for supporting me morally during the completion of my thesis. iv
5 TABLE OF CONTENTS Chapter Page I. DEMOGRAPHY OF AN URBAN POPULATON OF THE TEXAS HORNED LIZARD IN CENTRAL OKLAHOMA Abstract Introduction.. 2 Study Area... 4 Methods... 4 Field Methods Morphometrics Demography Results Morphometrics.. 10 Demography.. 10 Discussion Morphometrics.. 13 Demography.. 15 Literature Cited II. SPATIAL ECOLOGY OF THE TEXAS HONED LIZARD ON AN URBAN AIR FORCE BASE Abstract Introduction Study Area Methods Field Methods Home Ranges Seasonal Movements Habitat Results Home Ranges Seasonal Movements Habitat Discussion Home Ranges Seasonal Movements Habitat v
6 Chapter Page Management Considerations.. 66 Literature Cited. 67 III. POPULATION STATUS, CAPTURE METHODS, AND TEMPORAL DIFFERENCES IN CAPTURE RATE OF THE TEXAS HORNED LIZARD. 105 Abstract Introduction Study Area Methods Field Methods. 108 Mark-Recapture and Density Estimate Transects Overall Captures Results Mark-Recapture and Density Estimate Transects. 112 Overall Captures Discussion Literature Cited APPENDICES Appendix A ESTIMATED OVERLAP BETWEEN 95% MINIMUM CONVEX POLYGON HOME RANGES Appendix B ESTIMATED OVERLAP BETWEEN 75% MINIMUM CONVEX POLYGON HOME RANGES Appendix C NOTES ON HIBERNATION OF PHRYNOSOMA CORNUTUM Appendix D NONTES ON THE DIET OF PHRYNOSOMA CORNUTUM IN THE ABSENCE OF POGONOMYRMEX SPP vi
7 LIST OF TABLES Table Page Chapter I. 1. Sex and age class of Texas horned lizards captured from June 2003 to October 2005 on Tinker Air Force Base, Oklahoma County, Oklahoma Characteristics of nesting sites for Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Chapter II. 1. The number of points, area of 95% minimum convex polygon (MCP), and 95% fixed kernel (FK) active-season home ranges of lizards tracked at Tinker Air Force Base, Oklahoma County, Oklahoma, May 2004 to October Correlation between body size and home-range size of adult male and female lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, Mean (± 1 SE) area (ha) and mean (± 1 SE) percentage of lizard s 95% MCP home ranges that overlapped between and within sexes of Texas horned lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, Mean (± 1 SE) area (ha) and mean (± 1 SE) percentage of lizard s 75% MCP home-ranges that overlapped between and within sexes of Texas horned lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, Mean (± 1 SE) daily distances (m) traveled per month by Texas horned lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, vii
8 Table Page Chapter II. cont. 6. Mean percentage of available and used macrohabitat categories in analysis of second- and third-order habitat selection of Texas horned lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, Ranking matrix for second-order selection, with zero values in used habitat replaced by 0.01, by Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Ranking matrix for second-order habitat selection, with zero values in used habitat replaced by 0.4, by Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Ranking matrix for third-order habitat selection, with zero values in used habitat replaced by 0.1, by Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Chapter III. 1. Summary of mark-recapture periods for Texas horned lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, Appendices Appendix A. Estimated areas of overlap between 95% minimum convex polygon home ranges for Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma Appendix B. Estimated areas of overlap between 75% minimum convex polygon home ranges for Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma Appendix C. Table 1. Characteristics of hibernation locations of Texas horned lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, 2004 and Appendix D. Table 1. Summary of invertebrates contained in digestive tracts of 3 adult Phrynosoma cornutum at Tinker Air Force Base, Oklahoma County, Oklahoma, viii
9 LIST OF FIGURES Figure Page Chapter I. 1. Study site (Reserve 3) at Tinker Air Force Base, Oklahoma County, Oklahoma Snout-vent length (mm) of age and sex cohorts of Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Total length (mm) of age and sex cohorts of Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Mass (g) of age and sex cohorts of Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Annual survival rates of Texas horned lizards for Category 1 and Category 2 at Tinker Air Force Base, Oklahoma County, Oklahoma, June 2003 to October Chapter II. 1. Study site (Reserve 3) at Tinker Air Force Base, Oklahoma County, Oklahoma Cumulative home-range size for radiolocation points collected for lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Extent of home-range overlap using the 95% minimum convex polygon estimate for adult male lizards during (A) 2004 and (B) 2005 at Tinker Air Force Base, Oklahoma County, Oklahoma Extent of home-range overlap using the 95% minimum convex polygon estimate for adult female lizards during (A) 2004 and (B) 2005 at Tinker Air Force Base, Oklahoma County, Oklahoma Home-range overlap of two pairs of male lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, ix
10 Figure Page Chapter II. cont. 6. Home-range overlap of a pair of adult male lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Home-range overlap of two pairs of female lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Extent of home-range overlap using the 75% minimum convex polygon estimate for adult male lizards during (A) 2004 and (B) 2005 at Tinker Air Force Base, Oklahoma County, Oklahoma Extent of home-range overlap using the 75% minimum convex polygon estimate for adult female lizards during (A) 2004 and (B) 2005 at Tinker Air Force Base, Oklahoma County, Oklahoma Mean percentage cover of grass, forb canopy, shrub/tree canopy, bare ground, rock, and litter within a 0.1-m 2 Daubenmire frame centered at used and random locations of Texas horned lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, Mean percentage of litter within a 0.1-m 2 Daubenmire frame centered at lizard locations for month and time categories of Texas horned lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, Chapter III. 1. The frequency of capture of adult females, adult males, juveniles, and hatchlings by time of day for May through August on Tinker Air Force Base, Oklahoma County, Oklahoma x
11 CHAPTER I DEMOGRAPHY OF AN URBAN POPULATION OF THE TEXAS HORNED LIZARD IN CENTRAL OKLAHOMA ABSTRACT The response of reptiles to urban development is poorly understood. I studied demography of an urban population of the Texas horned lizard (Phrynosoma cornutum), a species that apparently declines in abundance following urban development but may persist in localized areas. I located and captured lizards within an urban reserve at Tinker Air Force Base, Oklahoma, from June 2003 to October Lizards were monitored through radiotelemetry and data were collected on morphometrics, survival, and reproduction. These data were applied to a life-table analysis that estimated the hatchling survival rate necessary to maintain a stable population. Adult females were larger than adult males and juvenile females were larger than juvenile males. Annual rates of survival were estimated to be for adult lizards and 0.45 for juvenile lizards. Mean (± 1 SE) clutch size was 17.4 ± 0.8 eggs, incubation periods ranged from 49 to 68 days, and 60% of nests survived to hatching. Nest sites primarily occurred on S SW aspects with slopes ranging from < 5 to 30 o. From application of Euler s equation, the hatchling survival rate needed for population stability ranged from 0.19 to Relative to other, more southerly P. cornutum populations, my study population had higher survival but lower reproductive rates. Relative to other phrynosomatine lizards, Texas 1
12 horned lizards are characterized by high survival, moderate reproductive output, and delayed maturation. INTRODUCTION It is important to understand more about urban wildlife populations due to the high rate of development of natural areas. The response of reptile populations to urban development is probably the least understood of all vertebrates (Germaine and Wakeling 2001). An understanding of the unique management issues of urban populations is especially important for species, such as the Texas horned lizard (Phrynosoma cornutum), that apparently decline in abundance following urban and agricultural expansion (Carpenter et al. 1993). Urban development affects the physical structure and species composition of native vegetation in many ways, including the replacement of native vegetation by anthropogenic features and nonnative vegetation, soil compaction, decreased stability of microclimates, and isolation of undeveloped native habitat remnants (Germaine and Wakeling 2001). In turn, these vegetation, soil, and microclimate changes affect animal populations. Developed areas are frequently unable to support populations of many species of native wildlife. The Texas horned lizard occurs in a variety of habitats ranging from deserts and grasslands to shrublands (Carpenter et al. 1993, Fair and Henke 1998, Sherbrooke 2003). Numerous studies have provided evidence that populations of horned lizards are declining or have disappeared in portions of the species range (Carpenter et al. 1993, Donaldson et al. 1994, Henke 2003). Factors associated with urban areas thought to negatively affect populations of horned lizards include habitat alteration, use of insecticides, agricultural irrigation or tilling, lawn mowing and watering, over-collection 2
13 for the pet trade, increased predation by urban pests, and increased mortality on roads (Carpenter et al. 1993, Donaldson et al. 1994). In a survey conducted by Carpenter et al. (1993), many new landowners reported observing horned lizards when they first moved into a new housing development, followed by quick disappearance of the lizards. Insecticides are often used to kill ants, which are the main food of horned lizards, and insecticides also may be directly toxic to horned lizards, especially eggs and hatchlings (Carpenter et al. 1993, Donaldson et al. 1994). Tilling may directly kill or unearth hibernating lizards before the cold season has ended (Fair and Henke 1997). Urban development also may increase the density of potential adult and nest predators, such as feral cats, fire ants (Solenopsis invicta; Donaldson et al. 1994), and mesocarnivores. Lizards are frequently killed on roads while basking or traveling (Carpenter et al. 1993). Development may alter diversity of the prey base available to P. cornutum by enabling the spread of invasive ant species and displacement of sensitive species (Donaldson et al. 1994). Finally, development and habitat loss may alter the availability and quality of sites used by P. cornutum during critical periods, such as nesting or hibernation. Populations of Texas horned lizards often have a localized distribution and thus can survive in protected areas within an urban environment. Populations of Texas horned lizards are often found in small city parks and vacant lots (Carpenter et al. 1993, Donaldson et al. 1994, Stark 2000). Therefore, it is important to understand how urban development affects the ecology of P. cornutum to ensure the viability of populations in developed areas. My objectives were to describe the demography of an urban population of P. cornutum in central Oklahoma and to compare this population with P. cornutum at 3
14 different latitudes, as well as other lizard species. Data on morphometrics, survival, and reproduction were collected and applied towards a life-table analysis. STUDY AREA Tinker Air Force Base covered approximately 2,000 ha in Oklahoma County, Oklahoma (35 o 24' 58 N, 97 o 24' 41 W). I focused on Wildlife Reserve 3 (ca. 15 ha; Figure 1), which was located within the Urban Greenway (ca. 183 ha). The aerial photograph of the reserve was provided by Tinker Air Force Base and is a color digital orthoquad with 9-cm pixel resolution. This area was dominated by grassland and contained patches of forest and shrubs. The dominant vegetation included big bluestem (Andropogon gerardii), little bluestem (Schizachyrium scoparium), plains bluestem (Bothriochloa ischaemum), indiangrass (Sorghastrum nutans), side oats grama (Bouteloua curtipendula), Maximilian sunflower (Helianthus maximiliani), tall fescue (Lolium pratense), and eastern redcedar (Juniperus virginiana). Residential developments were located along the southern, western, and northwestern edges of the reserve; and military buildings were located beyond the northeastern and eastern boundaries. The northern section of the reserve was crossed east west by a road. The reserve contained paved and gravel trails used for jogging, bicycling, dog walking, and wildlife viewing. METHODS Field Methods Lizards were located and captured within the study area from June 2003 to October 2005 through fortuitous encounters on gravel and paved trails and vegetated areas throughout the study site. For each specimen, snout-vent length (SVL), total length 4
15 (TL), mass, and sex were recorded. Air temperature in the shade and cloacal temperature at time of capture also were recorded. Lizards > 5.0 g were implanted with a passive integrated transponder (PIT; Electronic ID, Inc., Cleburne, Texas), and all lizards were individually marked with a unique toe clip for future identification. Lizards were monitored through radiotelemetry using two sizes of transmitters (Holohil Inc., Model: BD-2): 1) 1.8 g with battery life of 14 weeks; and 2) 1.3 g with battery life of 9 weeks. As lizards were captured, they were released with transmitters if the mass of available transmitters was < 10% of the lizard s body mass. Transmitters were glued posterior to the lizard s head and secured with an elastic band around the neck. The elastic band ensured that lizards did not lose transmitters when lizards molted and it allowed reattachment of transmitters as they came unglued from lizards. Lizards were fitted with a new transmitter as battery life ended. Beginning 24 hours after release, lizards were relocated 1 5 times per week during summer months (May August) and once per week or every other week during the fall and spring. Lizards tracked to hibernation locations were periodically checked during the winter months. Once lizards were hibernating, a mesh cage (0.3 m x 0.3 m x 0.3 m) was placed over each location so that lizards could be recaptured and fitted with a new transmitter when they emerged in the spring. Morphometrics Mean SVL, TL, and mass measurements were calculated for adult, juvenile, and hatchling lizards. I considered lizards hatchlings until they emerged from their first hibernation at approximately 8 months of age. They do not breed during their first spring and summer, therefore, they are considered juveniles from 8 20 months of age. Male 5
16 and female lizards breed during their second spring; therefore, adult lizards were > 20 months of age. If lizards were captured more than once within an age class, means were calculated from the maximum measurement of lizards, although mass measurements of gravid females were not used in the mean. Hatchling measurements were reported from the initial capture. Juvenile and adult lizards were distinguished based on a combination of length and mass measurements. The first measurement collected for a lizard was used to determine age. In early summer, juvenile lizards were easily recognized due to their small size; however, by late summer juvenile lizards approached adult lizards in length and mass. Generally, the SVL and mass of adult females were > 63 mm and > 13 g, respectively. The SVL and mass of adult males was > 49 mm and > 9 g, respectively. Ballinger (1974) reported a minimum SVL at maturity for P. cornutum of mm and Henke and Montemayor (1997) classified lizards > 75 mm as adults, however, P. cornutum attains a larger body size in southern portions of its range. I conducted 2-way ANOVA with interaction to compare morphometrics (SVL, TL, mass) by age (adult, juvenile, hatchling) and sex (female, male; PROC MIXED procedure, SAS Institute 2001). The Kolmogorov-Smirnov test for normality indicated that the morphometric distributions by age and sex were normal (0.07< P >0.15), except for the SVL of hatchling males (P < 0.01; PROC UNIVARIATE, SAS Institute 2001). Levene s test revealed that variances of morphometric measurements were not homogenous (P < ; PROC GLM, SAS Institute 2001); however, the PROC MIXED procedure is robust to non-homogeneity of variance (Littell et al. 1996). If differences (P < 0.05) in morphometrics were found among age and sex of lizards, then an adjusted Tukey s test was conducted for all pairwise comparisons. 6
17 Demography Survival. Survival rates were calculated using the Kaplan-Meier estimator with staggered entry of individuals (Pollock et al. 1989) for all lizards from 2003 to 2005 that were tracked by telemetry for > 14 days. In addition to the annual survival rate, separate survival rates were calculated for the active (summer: 1 April 30 September) and inactive (hibernation: 1 October 31 March) seasons. I assumed that these seasons had different survival rates because of the decreased risk of predation and human-caused mortality while lizards are inactive and hibernating under the soil surface, but increased risk of death by freezing. The fate of 2 lizards during the active season and 1 lizard during the hibernation season was undetermined; therefore, two different survival estimates were calculated. The first, termed Category 1, assumed that lizards with undetermined fates were alive. Category 2 assumed that lizards with undetermined fates were dead. The causes of undetermined fates were loss of a transmitter signal, which could be due to either transmitter failure or predator removal, and discovery of a transmitter without a lizard, which could be due to either the lizard losing the transmitter or a predation event. The fate of the lizard was considered known if a live lizard s transmitter was removed, a carcass was found, a lizard with a lost signal was later recaptured, or a transmitter was found outside of the lizard s normal range and the transmitter was damaged beyond normal wear. Four lizards died because the antenna, transmitter, or elastic band around the neck became entangled in vegetation. These research-caused mortalities were not considered deaths in the survival estimate and these lizards were censored from the analysis at the last date at which they were known to be alive. 7
18 The log-rank test extension (Pollock et al. 1989) was used to determine if there was a difference in the estimated survival patterns between male and female lizards. A Z-test statistic (Pollock et al. 1989) was used to compare the endpoint of the survival curves between male and female lizards, which was the end of the hibernation season (31 March). Juvenile lizards were too small to carry transmitters. Therefore, juvenile survival was estimated by calculating the percentage of juveniles that were marked in 2003 and 2004 and recaptured the subsequent summer in 2004 and 2005, respectively. Reproduction. Radiotelemetry enabled the location of nest sites during 2004 and Gravid females with transmitters were located daily during the nesting season to increase the probability that they would be located while digging. Females frequently created trial nests before laying their eggs. If lizards exhibited nesting behavior, they were caught and weighed to detect if they had oviposited. At the height of the nesting season, females without transmitters were sometimes encountered digging trial and used nests. The date of nesting for 13 gravid females was determined and the nest site for 10 of the 13 gravid females was located. The 3 females whose nest sites were not known were found after depositing eggs, based on mass loss. The sample of 10 nests was used to determine clutch size, nest survival, and date of hatchling emergence. Habitat characteristics (slope, aspect, vegetation cover, substrate) of nest sites were determined at 7 nests monitored in Nests were monitored until hatching and cages (0.3 m x 0.3 m x 0.3 m) were placed over nests a few weeks before hatching to enable capture of hatchlings in After hatching, nests were uncovered, egg shells were counted, and length of incubation calculated. The 8
19 number of eggs in a clutch was compared to female SVL using least-squares regression (PROC REG procedure, SAS Institute 2001). The minimum number of eggs hatched by the population, or reproductive output (Germano and Williams 2005), was determined for each year by multiplying the total number of adult females found within the study site each year by average clutch size and percentage of nests that survived to hatching (pooled average from 2004 and 2005). Life-Table Analysis. I could not estimate hatchling survival directly, therefore, I used Euler s equation with estimated rates of adult and juvenile survival and estimated fecundity to solve for the hatchling survival rate needed for population stability (Hellgren et al. 2000). The equation used was: s h = 1/(m 1 + s j m 2 + s j s a m s j s n-1 a m n ), where s h was hatchling survival, s j was juvenile survival, s a was adult survival, and m x was the average fecundity of a female of age x. Hatchling survival is the survival rate for lizards from hatching in August (0.0 years old) to the following spring (0.5 years old). Juvenile survival was the survival rate of individuals from the first spring (0.5 years old) to the second spring (1.5 years old) and adult survival was the annual survival rate of reproductively active lizards (> 1.5 years old). Adult survival was estimated using the Kaplan-Meier staggered entry design and juvenile survival was estimated based on recaptures of first-year adults (see Methods, survival section). Fecundity (female hatchlings/female/year) was estimated as the product of average clutch size, nest survival rate, and 0.5 (assuming a 50:50 hatchling sex ratio). 9
20 RESULTS A total of 161 lizards was captured from June 2003 to October 2005 (Table 1). During these three years, 39 adult male and 41 adult female lizards (including lizards that were marked as juveniles and recaptured as adults the following summer) were uniquely marked, resulting in a sex ratio of 1 adult male to 1.05 adult females (49% males). The average percentage of the population composed of adult and juvenile lizards across the study period was 74% and 26%, respectively. Fifteen percent of the lizards that were captured in 2003 were recaptured in 2004 and 39% of the lizards captured in 2004 were recaptured in Morphometrics All 3 morphometric characteristics, SVL (P = ), TL (P = ), and mass (P = ), varied by an age-sex interaction (Figure 2, 3, and 4). This interaction was manifested in different effects of sex across age classes, as females grew larger than males, having begun life as hatchlings equal in size to males. Adult females were larger than adult males for all 3 morphometric measurements (P = < ), and juvenile females were larger than juvenile males for all 3 morphometric measurements (P = ). There were no differences between female and male hatchling lizards (P = 1.0). Demography Survival. Annual survival (S) for Category 1 was 0.70 (95% CI: ) and for Category 2 was 0.59 (95% CI: ; n = 45 lizards; Figure 5). Summer survival for Category 1 was 0.77 (95% CI: ) and for Category 2 was 0.70 (95% CI: ; n = 45 lizards). Hibernation survival for Category 1 was 0.91 (95% CI: ) 10
21 and for Category 2 was 0.83 (95% CI: ; n = 12 lizards). Three lizards were depredated: 2 by an avian predator and 1 by a grasshopper mouse (Onychomys leucogaster). One lizard died due to leg injuries that inhibited movement and resulted in emaciation, and another was buried in a fencepost hole during construction. Other known mortalities during summer for lizards that were either not tracked with telemetry or were not tracked for at least 14 days (and thus excluded from survival rate calculations) included 1 death due to being run over on a road bordering the study site, 1 death by a grass mower, and 1 death attributed to avian predation based on the finding of a lizard head on a trail. One known mortality occurred during hibernation. A lizard chose a hibernation location in a depression that became saturated during periods of high rainfall. It was found dead above ground and in the center of its hibernation cage. Most likely it was too cold for the lizard to have traveled to another location and re-burrow; therefore, this death was not included as a project-caused mortality. Male and female yearly survival patterns did not differ (P = 0.72) and male and female endpoint survival also did not differ (P = 0.97). Seven of 15 marked juvenile lizards were recaptured the subsequent summer, providing a juvenile survival estimate of Reproduction. I observed a pair of lizards copulating on 11 May In 2004 and 2005, 13 female lizards were monitored during the nesting season and each deposited a single clutch of eggs from 2 June to 23 June (Table 2). Mean (± 1 SE) clutch size (CS) was 17.4 ± 0.8 eggs (n = 8) and clutches ranged from 13 to 20 eggs. Neither hatchlings nor egg shells were recovered at 2 of the 10 nests. Clutch size (CS) was positively related to SVL with the regression equation: CS = 0.34 SVL 6.5 (R 2 = 0.64, P = 0.017). 11
22 In 2004 and 2005, all adult females with transmitters deposited a single clutch of eggs. Hatchling emergence occurred between 27 July and 30 August with incubation periods ranging from 49 to 68 days (n = 6). The nest of one female was located at the edge of a planted cedar and the attraction to the location may have been the ease of digging in the loose gardening soil and wood chips surrounding the cedar. This nest was in full shade for part of the day and had the longest incubation period (68 days). Six of 7 nests were on W-, SW-, or S-facing aspects and one nest (created by a first-year adult) had a NE aspect (Table 2). Other characteristics of nest sites included slopes that ranged from < 5 to 30 o and depths to eggs of 5 to 7.5 cm. Generally, grass or forb cover occurred above nest locations (Table 2). Four of 10 nests did not survive to hatching. Nest loss was determined by disturbance at the nest during the incubation period, if hatchlings were not captured in cages over nest sites, or egg shells were not found when the nests were excavated. One nest appeared to have been dug up by a predator during the incubation period, another was overturned during landscaping, and neither hatchlings nor egg shells were recovered at the remaining two nests. The rate of nest survival was 0.60; however, this estimate is based on a small sample size. Reproductive output (number of eggs hatched) by females on the study area (assuming all adult females oviposited) was estimated to be 167 in 2003, 146 in 2004, and 219 in Life-table analysis. The results presented for juvenile survivorship, adult survivorship, and fecundity were used to estimate the survival rate of hatchlings necessary to maintain a stable population. I made the following assumptions: first reproduction at > 1.5 years, 12
23 clutch frequency of 1.0/year, fecundity of 5.22 female hatchlings/female/year, 0.45 juvenile survival rate, and 0.7 adult survival rate (Category 1). The number of years a female is reproductive is unknown. Baur (1986) reported a life-span of at least 8 years for a captive female P. cornutum. Of 16 adult females marked in 2003 within my study site, only 3 (19%) were recaptured in 2004 and only 1 (6%) was recaptured in Of 14 adult females captured in 2004, 7 (50%) were recaptured in The estimated survival rate of 0.7 predicts that 34% of adults would survive for 3 years. Additional years of mark-recapture data are needed for this population to accurately estimate the number of years an adult female will reproduce. I assumed that adult females would reproduce for a minimum of 2 years to a maximum of 3 years at the estimated survival rate of 0.7. The hatchling survival rate needed to maintain a stable population was 0.25 if females reproduced for only two summers and 0.19 if females reproduced for three summers. DISCUSSION Morphometrics The size of lizards at Tinker Air Force Base was smaller than P. cornutum in the southern portion of its range and similar to P. cornutum in other northern parts of its range. For example, Stark (2000) reported mean adult SVL and mass measurements for a population of P. cornutum in central Oklahoma of about 60 mm and 18 g for adult females and 58 mm and 13 g for adult males and Montgomery and Mackessy (2003) reported mean SVL measurements of 70 mm for adult females and 61 mm for adult males for P. cornutum in Colorado. P. cornutum are larger in southern parts of the distribution. Moeller et al. (2005) reported SVL and mass measurements of a population 13
24 of P. cornutum in southern Texas to average 92 mm and 45 g for adult females and 85 mm and 39 g for adult males. Overall, Montgomery et al. (2003) found a trend of decreased size with increased latitude for 10 morphological characteristics in P. cornutum from central Mexico to southeastern Colorado and concluded that it was caused by the temperature gradient associated with latitude. Proximate mechanisms considered for the size gradient were the shortened activity season or decrease in net primary productivity in higher latitudes. This trend of a decrease in body size with increase in latitude is contrary to Bergmann s rule of an increase in body size with an increase in latitude (Ashton and Feldman 2003). Many researchers have debated if Bergmann s rule applies to ectotherms, which typically grow slower at low temperatures compared to high temperatures (Mousseau 1997, Belk and Houston 2002, Angilletta et al. 2004). Atkinson (1994) and Angilletta et al. (2004) show evidence that in ectotherms, a larger body size in colder environments may be a trade-off between growth and reproduction. Growth would be favored over reproduction when the benefit of expected gains in future reproduction exceeds the costs associated with loss of current reproduction. Throughout their range, P. cornutum are found to become reproductively active during their second season after hatching at > 1.5 years of age (Ballinger 1974, Howard 1974, this study). The unity in age at maturation and the negative correlation found between body size and latitude indicate that Phrynosoma cornutum does not follow Bergmann s rule. Ashton and Feldman (2003) found that most squamates do not follow Bergmann s rule, reporting that 61 of 83 species decrease in size with latitude and 40 of 56 species increase in size with temperature. Angilletta et al. (2004) examined life-history traits of Sceloporus 14
25 undulatus, a lizard that does follow Bergmann s rule. They found that juvenile survivorship, age at maturity, and size at maturity were related to latitude and environmental temperature, with juveniles at high latitudes having higher survival, delayed maturation, and larger body sizes at maturation. Demography Survival. The comparison of survival rates among populations of P. cornutum in different parts of its range is difficult because investigators used different methods to calculate survival, were able to track lizards only during part or all of the summer season, or have used different criteria for determining the fate of lizards when the cause of death or disappearance of the lizard was not clear. I used active-season survival estimates from the literature that were calculated similar to my categories to compare with estimate in my study. Overall, survival estimates calculated for southern populations of P. cornutum were similar to estimates in my study for Category 1 (0.77), but lower than estimates in my study for Category 2 (0.70; Munger 1986, Fair and Henke 1999, Burrow et al. 2002, Moeller 2004). Munger (1986) estimated summer survival rates for 27 P. cornutum tracked with radiotelemetry in southeastern Arizona of 0.86 for Category 1 and 0.48 for Category 2. Burrow et al. (2002) calculated survival (Category 1) for P. cornutum in southern Texas and found summer survival rates of 0.38 for unburned areas and 0.52 for burned areas, whereas Moeller (2004) found the survival rate of P. cornutum (n = 51) in the same area to be 0.74 for Category 1 and 0.44 for Category 2. Fair and Henke (1999) calculated daily survival rates for 9 P. cornutum in southern Texas that were tracked from 1 to 6 months. They converted the daily rates to survival estimates ranging from
26 (Category 2) to 0.54 (Category 1) based on an 8-month year, which excluded the hibernation season in that area. Lower survival rates (based on the assumption that missing lizards were dead) in southerly populations may be partially due to differences in predator populations between study sites. The number of potential predators found within my study site may be decreased by human-caused mortalities on typical predators of horned lizards. For example, snakes may be killed during military training. Also, large birds that may be hazardous to planes using Base runways are reduced in numbers by trapping of individuals and removal of nests and nesting sites. Conversely, other urban and wildland populations could have more diverse and abundant populations of predators than the present study site. It is unclear if there is latitudinal variation in mortality rates within these populations of P. cornutum. Predation pressure is thought to be greater on lower-latitude populations of various species (Pianka 1986, Tinker and Ballinger 1972, Wilson 1991 and references within). Lizards in warmer environments may have higher levels of activity and are therefore exposed to predators for longer periods of time. Also, there may be fewer predators at higher latitudes due to patterns in species diversity (Angilletta et al. 2004). However, Wilson (1991) found that predation on Uta stansburiana during the active season was not higher for populations at lower latitudes. A possible explanation was that the longer activity season of low-latitude populations may be balanced by shorter daily activity periods due to extreme temperatures, whereas cooler temperatures at higher latitudes may result in a shorter activity season with longer daily activity periods for the lizards and their predators. Also, in cooler environments, lizards 16
27 would need to spend more time basking and would be highly visible to predators at these times. More data are needed to determine the relationship between predation pressure on P. cornutum and latitude. Reproduction. The reproductive strategy of the Texas horned lizard has been classified as falling in the category of delayed maturation and one large clutch per year in contrast to early maturation and multiple small clutches per year (Tinkle et al. 1970, Pianka and Parker 1975). Generally, Phrynosoma lizards do not reproduce until months of age (Ballinger 1974, Pianka and Parker 1975), although reproductive strategies may vary with latitude. Latitudinal variation in reproductive and life-history strategies (i.e., age and size at maturity, clutch size and frequency, adult and juvenile survival) have been commonly observed in many other phrynosomatine lizards, such as Sceloporus spp. (Tinkle and Ballinger 1972, Ferguson et al. 1980, Ramirez-Bautista et al. 2004), Uta stansburiana (Parker and Pianka 1975), and Urosaurus ornatus (Dunham 1982). The genus Phrynosoma may contain a continuum of reproductive strategies (Howard 1974). Howard (1974) examined the internal reproductive state of preserved individuals of the Phrynosoma genus collected at various times of the year. He found evidence that in some portions of their ranges P. mcalli, P. modestum, P. platyrhinos, and P. solare may reproduce before their second hibernation at about 12 months of age, whereas P. cornutum and P. douglassi were not found to have early maturation (Howard 1974). Also, in southern portions of their ranges some Phrynosoma, including P. cornutum, may lay multiple clutches per year (Howard 1974, Burrow 2000). Latematuring species would likely have the trade-offs of high survival and/or high fecundity, 17
28 whereas early-maturing species would be characterized by low adult survival and/or small clutch or litter sizes (Tinkle et al. 1970, Adolph and Porter 1996). Species in the genus Phrynosoma, especially P. cornutum, produce large clutches compared to other phrynosomatine lizards. Fitch (1970) reported an average clutch size for P. cornutum of 22.5 eggs based on clutch sizes reported by researchers in the literature at that time. Howard (1974) counted yolked ovarian follicles and oviducal eggs and reported an average clutch size of 29.7 eggs, ranging from 13 to 45 for P. cornutum in southeastern Arizona, southwestern Texas, and northern Mexico. Ballinger (1974) reported a mean clutch size of 29 eggs, ranging from 18 to 49, for populations throughout Texas, New Mexico, and Arizona. For P. cornutum in southern Texas, Burrow (2000) reported an average clutch size of 21.7 eggs, ranging from 6 to 29. The comparatively small average clutch size of 17 eggs found in my study is likely due to the smaller body sizes and hence smaller reproductive capacities of northern populations of P. cornutum. Other researchers have reported a positive correlation between SVL of P. cornutum and clutch size (Ballinger 1974, Howard 1974). However, even this clutch size is large compared to other phrynosomatine lizards. Researchers have reported that average clutch sizes of other phrynosomatine lizards range from 2 to 15 eggs, with many species at the lower end of that range (Tinkle et al. 1970, Tinkle and Ballinger 1972, Parker and Pianka 1975, Ferguson et al. 1980, Dunham 1982, Stearns 1984, Pianka 1986). This trend remains when reproductive effort is measured by computing the ratio of clutch or litter mass to female body mass (Pianka 1986). Pianka (1986) showed that this ratio tended to be high in Phrynosoma lizards, whereas other phrynosomatine lizards had ratios equal or as low as half the ratio observed in Phrynosoma lizards. 18
29 Reproduction in P. cornutum fits with its life-history strategy. Vitt and Congdon (1978) demonstrated that lizard body shape can by used to predict relative clutch mass, especially when predator escape and foraging strategies are considered. P. cornutum has a unique life-history strategy that is characterized by a large body size to enable sufficient consumption of the highly chitinous ant species it consumes. Unlike most other lizard species that use sleek bodies and speed for predator avoidance, P. cornutum depends on crypsis and spines. Therefore, adult females can carry large clutches because they do not need to depend on speed to evade predation or capture prey (Pianka and Parker 1975, Vitt and Congdon 1978, Pianka 1986). Life-Table Analysis. The hatchling survival rates ( ) needed to maintain my study population at the estimated fecundity and survival rates were low compared to the juvenile (0.45) and adult ( ) survival estimates. From a life-history perspective, hatchling survival is expected to be low for P. cornutum due to its high adult survival and large clutch sizes (Pianka and Parker 1975). The anti-predator strategies of crypsis and spines may provide little defense for hatchlings, which have much shorter spikes and horns than adults. It has been shown in P. mcalli that individuals with shorter horns were more likely to be depredated by shrikes than individuals with longer horns (Young et al. 2004). Lower adult survival rates have been estimated for P. cornutum in southern areas of its range (Munger 1986, Fair and Henke 1999, Burrow 2000), but longer activity seasons and larger body sizes in these populations allow females the trade-off of larger clutches and possibly multiple clutches per year. This life-table analysis assumed the population size of P. cornutum at my study site was stable. The finite rate of increase (λ) was estimated for this population using 19
30 mark-recapture data collected from May August 2005 and the Jolly-Seber (Burnham) model to be 0.93 ± 0.07 (see Chapter 3). This indicates that this assumption may be valid for this population. Apparent monthly survival also was estimated from the markrecapture data for adult and juvenile lizards combined to be 0.86, which translates to a survival of 0.55 from May August. The apparent survival estimate is expected to be lower than the true survival because it does not differentiate between individuals that died and individuals that emigrated from the study area. The apparent survival estimate falls between the juvenile and adult survival estimates presented above. Life-table analyses have been conducted for some well-studied phrynosomatines such as S. undulatus, U. stansburiana, and Urosaurus ornatus (Parker and Pianka 1975, Ferguson et al. 1980, Dunham 1982). Comparatively, the adult survival estimates of P. cornutum from my study and others are at the higher end of the spectrum. Ferguson et al. (1980) summarized data on 10 populations of S. undulatus and reported a range in annual adult survival rates from 0.07 to In these populations, the age at which individuals first reproduced was 1 to 2 years, clutch frequency per year was 2 4, and average clutch size was eggs. Dunham (1982) reported adult survival rates of U. ornatus to range from , first reproduction occurred at 8 12 months of age, clutch size ranged from eggs, and 1 to 3 clutches were produced per year. Parker and Pianka (1975) reported average clutch sizes of U. stansburiana to range from 2.85 to 4.82 and 1 to 2 clutches were produced per year. Typically, the number of clutches produced a year is limited by latitudinal variation in the length of active season. In southern populations, females of some species may be able to produce as many eggs through multiple clutches as P. cornutum females produce in one clutch. Hatchling survival ( ; Tinkle and 20
31 Ballinger 1972) and juvenile survival [( ; Ferguson et al. 1980)( ; Dunham 1982) are also low in other phrynosomatine lizards. Overall, the life-history strategy of P. cornutum is characterized by delayed maturation, large clutch investment, and high adult survival. As latitude increases, P. cornutum tends toward smaller body sizes and smaller, fewer clutches per year. This decrease may be balanced by higher survival rates at higher latitudes. Compared to other phrynosomatines, adult survival in P. cornutum was higher, clutch size larger, clutch frequency lower, and maturation delayed. LITERATURE CITED Adolph, S. C., and W. P. Porter Growth, seasonality, and lizard life histories: age and size at maturity. Oikos 77: Angilletta, M. J., Jr., P. H. Niewiarowski, A. E. Dunham, A. D. Leache, and W. P. Porter Bergmann's clines in ectotherms: illustrating a life-history perspective with sceloporine lizards. The American Naturalist 164:E168-E183. Ashton, K. G., and C. R. Feldman Bergmann's rule in nonavian reptiles: turtles follow it, lizards and snakes reverse it. Evolution 57: Atkinson, D Temperature and organism size: a biological rule for ectotherms? Advances in Ecological Research 25:1-58. Ballinger, R. E Reproduction of the Texas horned lizard, Phrynosoma cornutum. Herpetologica 30: Baur, B Longevity of horned lizards of the genus Phrynosoma. Bulletin Maryland Herpetological Society 22:
32 Belk, M. C., and D. D. Houston Bergmann's rule in ectotherms: a test using freshwater fishes. The American Naturalist 160: Burrow, A. L The effect of prescribed burning and grazing on the threatened Texas horned lizard (Phrynosoma cornutum) in the western Rio Grande Plains. Unpubl. Thesis, Oklahoma State University, Stillwater. Burrow, A. L., R. T. Kazmaier, E. C. Hellgren, and D. C. Ruthven, III The effects of burning and grazing on survival, home range, and prey dynamics of the Texas horned lizard in a thornscrub ecosystem. In W. M. Ford, K. R. Russell, and C. E. Moorma (eds.), The role of fire in nongame wildlife management and community restoration: traditional uses and new directions: proceedings of a special workshop, pp USDA Forest Service Northeastern Research Station General Technical Report NE-288. Carpenter, C. C., R. St. Clair, and P. Gier Determination of the distribution and abundance of the Texas horned lizard (Phrynosoma cornutum) in Oklahoma. Final report, Federal Aid Project E-18, Oklahoma Department of Wildlife Conservation, Oklahoma City, OK. Donaldson, W., A. H. Price, and J. Morse The current status and future prospects of the Texas horned lizard (Phrynosoma cornutum) in Texas. The Texas Journal of Science 46: Dunham, A. E Demographic and life-history variation among populations of the iguanid lizard Urosaurus ornatus: implications for the study of life-history phenomena in lizards. Herpetologica 38:
33 Fair, W. S., and S. E. Henke Effects of habitat manipulation on Texas horned lizards and their prey. Journal of Wildlife Management 61: Fair, W. S., and S. E. Henke Habitat use of Texas horned lizards in southern Texas. Texas Journal of Agriculture and Natural Resources 11: Fair, W. S., and S. E. Henke Movements, home ranges, and survival of Texas horned lizards (Phrynosoma cornutum). Journal of Herpetology 33: Ferguson, G. W., C. H. Bohlen, and H. P. Woolley Sceloporus undulatus: comparative life history and regulation of a Kansas population. Ecology 61: Fitch, H. S Reproductive Cycles in Lizards and Snakes. Volume 52. The University of Kansas Printing Service, Lawrence. Germaine, S. S., and B. F. Wakeling Lizard species distributions and habitat occupation along an urban gradient in Tucson, Arizona, USA. Biological Conservation 97: Germano, D. J., and D. F. Williams Population ecology of blunt-nosed leopard lizards in high elevation foothill habitat. Journal of Herpetology 39:1-18. Hellgren, E. C., R. T. Kazmaier, D. C. Ruthven, and D. R. Synatzske Variation in tortoise life history: demography of Gopherus berlandieri. Ecology 81: Henke, S. E Baseline survey of Texas horned lizards, Phrynosoma cornutum, in Texas. The Southwestern Naturalist 48: Henke, S. E., and M. Montemayor Phrynosoma cornutum (Texas horned lizard). Growth. Herpetological Review 28:
34 Howard, C. W Comparative reproductive ecology of horned lizards (Genus Phrynosoma) in southwestern United States and northern Mexico. Journal of the Arizona Academy of Science 9: Littell, R. C., G. A. Milliken, W. W. Stroup, and R. D. Wolfinger SAS System for Mixed Models. SAS Institute Inc., Cary, North Carolina. Moeller, B. A Conservation ecology of the Texas horned lizard (Phrynosoma cornutum): comparative effects of summer and winter burning. Unpubl. Thesis, Oklahoma State University, Stillwater. Moeller, B. A., E. C. Hellgren, D. C. Ruthven III, R. T. Kazmaier, and D. R. Synatzske Temporal differences in activity patterns of male and female Texas horned lizards (Phrynosoma cornutum) in southern Texas. Journal of Herpetology 39: Montgomery, C. E., and S. P. Mackessy Natural history of the Texas horned lizard, Phrynosoma cornutum (Phrynosomatidae), in southeastern Colorado. The Southwestern Naturalist 48: Montgomery, C. E., S. P. Mackessy, and J. C. Moore Body size variation in the Texas horned lizard, Phrynosoma cornutum, from Central Mexico to Colorado. Journal of Herpetology 37: Mousseau, T. A Ectotherms follow the converse to Bergmann's rule. Evolution 51: Munger, J. C Rate of death due to predation for two species of horned lizard, Phrynosoma cornutum and P. modestum. Copeia 1986:
35 Parker, W. S., and E. R. Pianka Comparative ecology of populations of the lizard Uta stansburiana. Copeia 1974: Pianka, E. R Ecology and Natural History of Desert Lizards: Analyses of the Ecological Niche and Community Structure. Princeton University Press, Princeton. Pianka, E. R., and W. S. Parker Ecology of horned lizards: a review with special reference to Phrynosoma platyrhinos. Copeia 1975: Pollock, K. H., S. R. Winterstein, C. M. Bunck, and P. D. Curtis Survival analysis in telemetry studies: the staggered entry design. Journal of Wildlife Management 53:7-15. Raimrez-Bautista, A., E. Jimenez-Cruz, and J. C. Marshall Comparative life history for populations of the Sceloporus grammicus complex (Squamata: phrynosomatidae). Western North American Naturalist 64: SAS Institute SAS release 8.1. SAS Institute, Cary, North Carolina, USA. Sherbrooke, W. C Introduction to Horned Lizards of North America. Volume 64. University of California Press, Berkeley. Stark, R. C Habitat use, daily movements, and body size of the Texas horned lizard in an urban environment in north-central Oklahoma. Unpubl. Thesis, Oklahoma State University, Stillwater. Stearns, S. C The effects of size and phylogeny on patterns of covariation in the life history traits of lizards and snakes. The American Naturalist 123: Tinkle, D. W., and R. E. Ballinger Sceloporus undulatus: a study of the intraspecific comparative demography of a lizard. Ecology 53:
36 Tinkle, D. W., H. M. Wilbur, and S. G. Tilley Evolutionary strategies in lizard reproduction. Evolution 24: Vitt, L. J., and J. D. Congdon Body shape, reproductive effort, and relative clutch mass in lizards: resolution of a paradox. The American Naturalist 112: Wilson, B. S Latitudinal variation in activity season mortality rates of the lizard Uta stansburiana. Ecological Monographs 61: Young, K. V., E. D. Brodie, Jr., and E. D. Brodie, III How the horned lizard got its horns. Science 304:65. 26
37 Table 1. Sex and age class of Texas horned lizards captured from June 2003 to October 2005 on Tinker Air Force Base, Oklahoma County, Oklahoma. Year Adult females Adult males Juvenile females Juvenile males Hatchlings Total Recaptures from previous years Total
38 Table 2. Characteristics of nesting sites for Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Year Date laid Date hatched June #, eggs a Incubation period (days) Slope ( o ) Aspect Depth to eggs (cm) Canopy cover (% and type) Fate Unknown June a Unknown June 18 Depredated June 27 July % forb Hatched June % grass Human disturbance June a Unknown June b 25 W 15% grass Depredated June 4 August <5 SW 5 25% grass/forb Hatched June 7 August <5 W 5 30% grass Hatched June 4 August SW 7.5 < 5% grass Hatched June 7 August NE % grass Hatched June b 15 S 10% grass Depredated June 30 August SW 6 65% cedar Hatched a nest not located; b neither eggs shells or hatchlings were recovered from nest
39 Figure 1. Study site (Reserve 3) at Tinker Air Force Base, Oklahoma County, Oklahoma. A shows the location of Tinker Air Force Base within Oklahoma, B shows the extent of Tinker Air Force base and location of the study site within Tinker Air Force Base, and C is a close-up view of the study site. 29
40 30
41 Figure 2. Snout-vent length (mm) of age and sex cohorts of Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, (Adult females, n = 28; Adult males, n = 36; Juvenile females, n =14; Juvenile males, n = 11; Hatchling females, n = 10; Hatchling males, n = 17). Error bars represent ± 1SE around each mean. 31
42 Snout-vent length (mm) Females Males Hatchling Juvenile Adult Age Class
43 Figure 3. Total length (mm) of age and sex cohorts of Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, (Adult females, n = 28; Adult males, n = 36; Juvenile females, n =14; Juvenile males, n = 11; Hatchling females, n = 10; Hatchling males, n = 17). Error bars represent ± 1SE around each mean. 33
44 Total Length (mm) de Females Males Hatchling Juvenile Adult Age Class
45 Figure 4. Mass (g) of age and sex cohorts of Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, (Adult females, n = 28; Adult males, n = 36; Juvenile females, n =14; Juvenile males, n = 11; Hatchling females, n = 10; Hatchling males, n = 17). Error bars represent ± 1SE around each mean. 35
46 Mass (g) 10 Females Males 5 0 Hatchling Juvenile Adult Age Class
47 Figure 5. Annual survival rates of Texas horned lizards for Category 1 (S = 0.70, 95% CI = ) and Category 2 (S = 0.59, 95% CI = ; n = 45) at Tinker Air Force Base, Oklahoma County, Oklahoma, June 2003 to October Error bars represent ± 1SE around each mean. Calculations for Category 1 assumed that lizards with undetermined fates were alive and calculations for Category 2 assumed that lizards with undetermined fates were dead. 37
48 June Aug Sept March Feb Jan Dec Oct Nov July April May Month Category 1 Category 2 Survival de 38
49 CHAPTER II SPATIAL ECOLOGY OF THE TEXAS HORNED LIZARD ON AN URBAN AIR FORCE BASE ABSTRACT I studied home-range size, home-range overlap, movements, and macro- and microhabitat use of the Texas horned lizard at Tinker Air Force Base, Oklahoma, from May 2004 to October There was no difference between home-range sizes of male and female lizards. Home-range size (± 1 SE) averaged 0.43 ± 0.08 ha and 0.90 ± 0.16 ha for 95% minimum convex polygon (MCP) and 95% fixed kernel methods (FK), respectively. Body size was consistently and positively correlated to home-range size for male lizards, but not for female lizards. The amount or percentage of home-range overlap between or within the sexes did not vary for 75 or 95% MCP home-range methods. Daily movements of male lizards were greater than female lizards in May, but not June or July. The macrohabitat analysis suggested that due to suppression of natural disturbances, study animals appear to have positioned their home ranges around the paved and gravel trails within the study site. These areas probably provide the open habitat needed for basking activities. Lizards avoided redcedar shrubland areas within the study site, perhaps due to the increase in shaded areas within this habitat. The difference in the percentage of microhabitat categories at locations used by lizards and random locations indicates that lizards selected areas of less dense vegetation. Relationships 39
50 between habitat composition and home-range size were stronger for females than males. The positive relationship between female home-range size and the proportion of a home range containing either developed habitat or redcedar shrubland suggests that these two categories provide poor-quality habitat for P. cornutum. The differences in space use and movements of male and female P. cornutum may be explained by their different reproductive strategies. Male lizards benefit from encountering more females during the mating season, whereas female lizards may adjust areas of use and their movements based on resource requirements, such as food, shelter, and nesting sites. INTRODUCTION Home-range size, home-range overlap, and movements are related to the resource requirements and behavior of a species, and are influenced by habitat quality (Simon 1975, Stamps 1977, Ruby and Dunham 1987, Perry and Garland 2002). Factors such as the distribution of food and other resources, predator avoidance, thermoregulation, access to mates, and intraspecific competition affect daily movements and home-range dynamics (Huey and Pianka 1977, Waldschmidt 1983, Ims 1987, Hews 1993, Stapley 2004, Stark et al. 2005). Urban and agricultural development also may affect home-range size and movements via alteration of habitat characteristics. Urban development affects the physical structure and species composition of native vegetation by replacement of native vegetation by anthropogenic features and nonnative vegetation, soil compaction, decreased stability of microclimates, and isolation of undeveloped native habitat remnants (Germaine and Wakeling 2001). In turn, these vegetation, soil, and microclimate changes affect animal populations. In poor-quality habitat, an individual may have to increase its home-range size beyond the area required in a higher-quality 40
51 habitat to find necessary resource requirements (Stenger 1958, Simon 1975, Krekorian 1976). Home-range dynamics and daily movements also may be used as an indicator of the selective pressures on a species. Researchers have debated if typical iguanid behavior, such as territorial defense and social hierarchies, exists in Texas horned lizards (Phrynosoma cornutum; Fair and Henke 1999). Lack of territorial defense has been noted by some researchers (Lynn 1965), whereas others believe there is some evidence for low-level home-range defense (Munger 1984). Lynn (1965) hypothesized that because Phrynosoma lizards are ground dwellers in habitats with few perches and because individuals are widely scattered throughout available habitat, horned lizards rarely come into contact with conspecifics and would not be effective at home-range defense. Alternatively, home-range overlap may be reduced through mutual avoidance to reduce competition for ant prey species (Munger 1984, Fair and Henke 1999). Interactions between P. cornutum have been observed where two P. cornutum approached within 1 m, head bobbed, and then mutually retreated (Whitford and Whitford 1973, Endriss, personal observation). Combat between male P. cornutum also has been observed (Whitford and Whitford 1973). Zamudio (1998) developed the small-male advantage hypothesis, in which she proposed that horned lizard males were selected to be small and mobile to increase their chances of locating females during the mating season. Stark et al. (2005) found increased daily movements and daily activity areas of male P. cornutum during the mating season and hypothesized that this indicated a mate-searching strategy. Males who increased 41
52 their movements during the mating season would be more likely to encounter and mate with receptive females. Texas horned lizards have been found in a variety of habitats ranging from grasslands and shrublands to deserts (Fair and Henke 1998, Burrow et al. 2001). Microhabitat selection by P. cornutum has been reported in a few studies. Fair and Henke (1998) found that P. cornutum selected open areas in the morning and areas of high canopy cover in the afternoon. They believed that sites with intermediate canopy cover were not often used because these sites prevented basking during cool temperatures and did not provide sufficient cover from predators during hotter temperatures. Similarly, Burrow et al. (2001) found that although P. cornutum used bare ground and herbaceous vegetation similar to availability in the morning and evening, lizards avoided bare ground and selected for woody vegetation and litter for cover in the afternoon. Utilizing visual searches, Whiting et al. (1993) found that P. cornutum was clustered around 3 disturbed areas within their study site that contained less ground cover than the surrounding areas, which allowed ease of movement of lizards and made foraging ant workers visible. However, Stark et al. (2005) used fluorescent powder-tracking to show that lizards often traveled through areas of dense vegetation for extended distances. The authors concluded that dense vegetation provided cover from predators. Stark (2000) found that during a hotter summer (40 days > 38 o C), habitat use was nonrandom, and patches of densely vegetated habitat were used most frequently, followed by patches of open areas and then sand. However, during a cooler summer (16 days > 38 o C), lizards did not show similar selection for patches of habitat. Stark (2000) found that microhabitats of bare ground, grass, herbaceous vegetation, leaf litter, and gravel were 42
53 used the most, whereas cobble, embedded rock, sand, and woody vegetation were used the least. I used radiotelemetry and fluorescent powder-tracking to monitor habitat and space use of an urban population of the Texas horned lizard in central Oklahoma. My objectives were: 1) to determine average home-range size of P. cornutum within the study site; 2) to determine the extent of home-range overlap within and between sexes; 3) to describe movements of P. cornutum during and after the mating season; 4) to determine macro- and microhabitat selection by P. cornutum within the study site; and 5) to determine if home-range size was correlated to macrohabitat. STUDY AREA Tinker Air Force Base covered approximately 2,000 ha in Oklahoma County, Oklahoma (35 o 24' 58 N, 97 o 24' 41 W). I focused on Wildlife Reserve 3 (ca. 15 ha; Figure 1), which was located within the Urban Greenway (ca. 183 ha). The aerial photograph of the reserve was provided by Tinker Air Force Base and is a color digital orthoquad with 9-cm pixel resolution. This area was dominated by grassland and contained patches of forest and shrubs. The dominant vegetation included big bluestem (Andropogon gerardii), little bluestem (Schizachyrium scoparium), plains bluestem (Bothriochloa ischaemum), indiangrass (Sorghastrum nutans), side oats grama (Bouteloua curtipendula), Maximilian sunflower (Helianthus maximiliani), tall fescue (Lolium pratense), and eastern redcedar (Juniperus virginiana). Residential developments were located along the southern, western, and northwestern edges of the reserve; and military buildings were located beyond the northeastern and eastern boundaries. The northern section of the reserve was crossed east west by a road. The 43
54 reserve contained paved and gravel trails used for jogging, bicycling, dog walking, and wildlife viewing. METHODS Field Methods Lizards were located and captured within the study area from May 2004 to October 2005 through fortuitous encounters on gravel and paved trails and vegetated areas throughout the study site. For each specimen, snout-vent length (SVL), total length (TL), mass, and sex were recorded. Lizards > 5.0 g were implanted with a passive integrated transponder (PIT; Electronic ID, Inc., Cleburne, Texas), and all lizards were individually marked with a unique toe clip for future identification. Lizards were monitored through radiotelemetry using two sizes of transmitters (Holohil Inc., Model: BD-2): 1) 1.8 g with battery life of 14 weeks; and 2) 1.3 g with battery life of 9 weeks. As lizards were captured, they were released with transmitters if the mass of available transmitters was < 10% of the lizard s body mass. Transmitters were glued posterior to the lizard s head and secured with an elastic band around the neck. The elastic band ensured that lizards did not lose transmitters when lizards molted and it allowed reattachment of transmitters as they came unglued from lizards. Lizards were fitted with a new transmitter as battery life ended. Beginning 24 hours after release, lizards were relocated 1 5 times per week during summer months (May August) and once per week or every other week during fall and spring. Each location was marked with a small pin flag. I also recorded whether or not the lizard flushed from its original location before it was spotted. 44
55 Lizards tracked to hibernation locations were periodically checked during the winter months. Once lizards were hibernating, a mesh cage (0.3 m x 0.3 m x 0.3 m) was placed over each location so that lizards could be recaptured and fitted with a new transmitter when they emerged in the spring. Each observed lizard location was recorded with a GPS (Trimble GPS Pathfinder Pocket Receiver) in Universal Transverse Mercator (UTM) coordinates using the North American Datum 1983 (NAD 1983). UTM coordinates were imported into ArcView 3.3 and ArcGIS 9.0 software (ESRI 2005) and, when possible, were differentially corrected using Pathfinder software. This technique uses base stations with known coordinates to continuously monitor the distance and direction of error in GPS locations. Pathfinder software adjusted the coordinates of locations based on the calculated error at the time I used GPS to collect coordinates. Home Ranges Home-Range Size. Home-range size of lizards was calculated using 95% minimum convex polygons (MCP; Rose 1982, Stone and Baird 2002) and 95% fixed kernels (FK; Worton 1989) using the Animal Movement Analysis Program extension to ArcView 3.3 (Hooge et al. 1999). The smoothing factor (h) for the 95% FK estimate was determined using least-squares cross validation (Worton 1989). The minimum number of points needed to accurately estimate home-range size was determined by calculating cumulative 100% MCP home ranges (Rose 1982, Aebischer et al. 1993). Home-range size was calculated for the first 10 points and again for each additional 5 points (i.e, 10, 15, 20, etc.) collected for a lizard (Figure 2). Qualitatively, this analysis showed that home-range size usually leveled off after the first 20 points were collected; therefore, only lizards with > 20 points were included in the home range and habitat analyses. During the 45
56 summer, lizards were located as frequently as every day to every 5 days; therefore, 20 points corresponded to months. The one lizard whose home range continued to increase after 20 points (Figure 2) was a lizard that had moved to a previously unused area after its initial home range had been mowed. The 95% MCP and 95% FK home-range size were compared between female and male lizards (PROC TTEST procedure, SAS Institute 2001). Pearson s correlation was used to determine if home-range size of female and male lizards was related to lizard body size (SVL, TL, and mass; PROC CORR procedure, SAS Institute 2001). Male and female lizards were analyzed separately because previous analysis showed that females were larger than males (see Chapter 1). The Kolmogorov-Smirnov test for normality indicated that the distributions of the 95% MCP and 95% FK home ranges were normal (0.08 < P > 0.15) for females and males. Levene s test revealed that variances for the 95% MCP and 95% FK were homogeneous (P = 0.20 and P = 0.63, respectively). Home-Range Overlap. The area of home-range overlap was calculated for female:female pairs, male:male pairs, and female:male pairs using the 95% MCP estimates and ArcMap 9.0 software. I chose the 95% MCP home range for this analysis because MCP estimates do not extend the home-range area beyond the known points for a lizard. I compared the amount of area that females overlapped with females, males overlapped with males, or females overlapped with males using analyses of variance (PROC GLM procedure, SAS Institute 2001). For all lizard pairs, the percentage of each lizard s home range composed of overlap was calculated. I compared the percentage of their home range that females overlapped with females, males overlapped with males, females overlapped with males, and males overlapped with females using analyses of variance (PROC GLM procedure, 46
57 SAS Institute 2001). This method was repeated using 75% MCP home-range estimates to determine the amount of overlap among areas of home ranges that were more intensively used. Seasonal Movements The average distance traveled in separate months for male and female lizards was calculated by measuring the sum of straight-lines distance between consecutive points in a month divided by number of days using Hawth s analysis tools extension to ArcMap 9.0 (Beyer 2004). For each lizard, an average distance traveled per day was calculated for May, June, and July because these were the only months when lizards were located frequently enough to have an accurate estimate of the daily distance traveled by lizards. Furthermore, note that this distance represents a minimum distance traveled because the straight-line distance between points was measured whereas lizards most likely traveled in a circuitous path between points. The data were not normally distributed (P = 0.045); therefore, I used a Kruskal-Wallis test to test for differences between female and male lizards in the average daily distance traveled for each month (PROC NPAR1WAY procedure, SAS Institute 2001). I also used a Kruskal-Wallis test to test for differences among the average daily distances traveled by month within males and within females (PROC NPAR1WAY procedure, SAS Institute 2001). Adult male lizards that were too small to release with transmitters were tracked using fluorescent powder to obtain daily distances traveled during and after the breeding season. For this method, a small square (2.5 by 2.5 cm) of rabbit fur was glued to the ventral surface of the lizard and fluorescent powder was rubbed into the fur. Lizards were released between 1700 and 2000 hr and I returned the same day after sunset to track 47
58 the lizards with the use of a handheld ultraviolet light to their night location. This location was marked with a small pin flag and the fur was repowdered in the field. The subsequent night, I returned to this location and marked the lizard s trail every 5 m with a small pin flag. The exact route was measured with a 5-m flexible rope. The lizard s trail was not marked until the first full day to ensure that lizard movements were not affected due to the stress of capture. Total trail length was calculated and represents the actual distance traveled by a lizard in a day. The data were normally distributed (P > 0.15) and had equal variances (P = 0.16). I compared daily distances traveled by male lizards during the mating season (May) and post-mating season (June and July) using a t-test (PROC TTEST procedure, SAS Institute 2001). Habitat Macrohabitat. A vegetation map of the study site was created using GIS and aerial photography by Virginia Polytechnic Institute and State University (Dorr et al. 2005). The vegetation map was digitized from an aerial photograph, verified using ground truthing, and completed in Stands of vegetation were designated based on the dominant or combinations of dominant plant species. Vegetation stands were pooled and reclassified into 4 habitat categories: developed areas, redcedar shrubland, native prairie, and nonnative prairie. Developed areas included a parking lot located along the northwestern edge of the reserve and paved and gravel trails located throughout the reserve. Generally, redcedar shrubs coexisted with plains bluestem (Bothriochloa ischaemum), but also were found with other native and nonnative grass and forb species. Typical nonnative prairie species included plains bluestem (Bothriochloa ischaemum) and tall fescue (Lolium pratense). Typical native prairie species included big bluestem 48
59 (Andropogon gerardii), little bluestem (Schizachyrium scoparium), indiangrass (Sorghastrum nutans), side oats grama (Bouteloua curtipendula), and Maximilian sunflower (Helianthus maximiliani). I used compositional analysis to determine if lizards showed habitat selection among the four habitat categories (Aebischer et al. 1993). The vegetation map was imported into ArcMap 9.0 and individual lizard points and home ranges were overlaid on the map. Compositional analysis uses individual lizards, rather than points, as sample units and allows comparison of used and available habitat to determine if habitat use differs from the habitat s availability in the environment. Only lizards with > 20 points were used in the macrohabitat analysis (see Home Ranges methods). If there was a significant difference between the use and availability of habitat categories, they were ranked in order of relative preference and avoidance. Compos Analysis 6.2 Plus (Smith 2005) was used to analyze macrohabitat data. This program utilizes a randomization algorithm and 1000 iterations as recommended by Aebischer et al. (1993) to determine the significance of Wilk s lambda and t-values. It also implements the preferred option described by Aebischer et al. (1993) for zero values in availability data (Smith 2005). If zero values are present in used habitat data, the program recommends a suitable small percentage for substitution as recommended by Aebischer et al. (1993). For second-order selection (Johnson 1980), available habitat was defined as the percentage of each macrohabitat category contained in the area of the study site occupied by all telemetered lizards. This area was determined by creating a convex polygon around all lizard points used in this analysis. Used habitat was defined as the percentage of each macrohabitat category contained in each lizard s 95% MCP home range. I chose 49
60 the 95% MCP to represent used habitat because this estimate does not extend the home range of lizards beyond the locations that were known to be used by the lizard. For thirdorder selection (Johnson 1980), available habitat was defined as the percentage of each macrohabitat category contained in each lizard s 95% FK home range. Used habitat was defined as the percentage of each lizard s points that fell into the four macrohabitat categories. I chose the 95% FK home range over MCP estimates as the best representation of the habitat available to lizards because this home-range estimate places buffer of use around observed lizard points. The lizard points were not patchy; therefore, the kernel estimate did not result in isolated areas of activity that were centered on lizard points. Bingham and Brennan (2004) found that nonzero substitutions in used habitat categories resulted in inflated Type 1 error rates, especially for the smaller substitution vales of and In their analysis, substitution values between 0.3 and 0.7 had smaller, but still unacceptable, occurrences of Type 1 error rates. Substitution values < 0.3 resulted in incorrectly identifying habitats with small availabilities as significantly avoided, whereas substitution values > 0.7 resulted in incorrectly identifying habitats with small availabilities as significantly preferred. Therefore, to test if the substitution value affected my results, I ran the second- and third-order habitat selection analyses with the substitution value recommended by the Compos Analysis 6.2 Plus program (Smith 2005) and with a higher substitution value of 0.4. However, there are substantial differences between the structure of my data and the data used in the analysis of Bingham and Brennan (2004). The data they used in their analysis consisted of 5 habitat categories with availability percentages of 1, 3, 6, 40, and 50, and the 3 categories with the smallest 50
61 availability percentages contained all of the 0% use values. In my analysis for secondorder selection, there were 4 habitat categories (developed habitat, redcedar shrubland, native prairie, and nonnative prairie) with availability percentages of 7.5, 21.6, 34.9, and 36.0, respectively. The 0% use values were distributed throughout the 4 habitat categories and the count for each category was 4, 7, 5, and 3, respectively. For thirdorder selection, lizards had variable non-zero availability percentages ranging from 0.5 to 100%. However, only 7 of 24 lizards had small nonzero values (< 6%) for available habitat. Therefore, my data may not be affected by nonzero substitutions in the same manner described by Bingham and Brennan (2004). Microhabitat. Microhabitat was defined as vegetation structure in the immediate vicinity of a located lizard. Microhabitat data were collected within a 0.1-m 2 Daubenmire frame centered on flagged locations used by lizards and at paired locations 10 m from each lizard location in a random location. The location of each random point was paced in the direction of the second hand of a watch. The percent cover of grass, forb canopy, shrub or tree canopy, bare ground, rock, and litter was estimated in each frame. Percent of each vegetation type was recorded in the following categories: 0%, < 1%, 1-5%, 6-25%, 26-50%, 51-75%, 76-95%, and %. The midpoint of each range was used as the value for each vegetation variable in the analysis. Habitat use (lizard locations) was compared to habitat availability (random points) using multivariate analysis of variance (Burrow et al. 2001; PROC GLM, MANOVA, SAS Institute 2001). The main effects were type of location (used vs. random), time of day (morning, afternoon, evening), and month (May, June, July, August). Morning locations were defined as occurring during hr, afternoon locations occurred 51
62 during hr, and evening locations occurred during hr. The model included all main effects and all interactions. Because the 3-way interaction of location, time, and month was not significant (P < 0.05), I present results from a reduced model without the 3-way interaction. The 6 microhabitat factors described above were the dependent variables. If the overall MANOVA was significant for an interaction or main effect, individual ANOVAs for each microhabitat variable were examined to isolate the effect (Burrow et al. 2001). Habitat Quality. To determine the quality of each macrohabitat category, I correlated percentage of a lizard s home range in each macrohabitat category with size of the lizard s home range estimated by the 95% FK and 95% MCP methods (PROC CORR, SAS Institute 2001). The majority of patches were > 0.5 ha; therefore, these patches composed a large enough area of each lizard s home range to influence home-range size. RESULTS Home Ranges Home-Range Size. Active season home-range size (± 1 SE) averaged 0.43 ± 0.08 ha (range: 0.03 to 1.06 ha) by the 95% MCP method and 0.84 ± 0.15 ha (range: 0.07 to 1.70 ha; n = 13) by the 95% FK method for adult female lizards (Table 1). Three female lizards were tracked continuously from the active season of 2004 through 2005 and, therefore, were included twice in the home-range estimates. Active season home-range size (± 1 SE) averaged 0.57 ± 0.18 ha (range: 0.14 to 2.21 ha) by the 95% MCP method and 0.90 ± 0.16 ha (range: 0.15 to 1.71 ha; n = 11) by the 95% FK method for adult male lizards (Table 1). There was no difference between home-range sizes of male and female lizards for the 95% MCP (T 22 = -0.76, P = 0.46) or the 95% FK (T 22 = -0.30, P = 0.76) 52
63 methods. Home-range size (± 1 SE) pooled between both sexes averaged 0.50 ± 0.09 ha by the 95% MCP method and 0.87 ± 0.11 ha by the 95% FK method. Body size (SVL, TL, and mass) was positively correlated to home-range estimates based on both methods for male lizards (n = 11; Table 2). For female lizards, mass was positively correlated to the 95% MCP and 95% FK home ranges, and TL was positively correlated to the 95% FK (n = 13; Table 3). SVL was weakly correlated to the 95% FK, and SVL and TL were not correlated to the 95% MCP (Table 2). Home-Range Overlap. The amount of area within 95% MCP home ranges that overlapped did not vary between and within sexes (F 2 = 0.39, P = 0.68; Table 3; Figures 3 and 4). Similarly, the percentage overlap of 95% MCP home-ranges did not vary between and within sexes (F 3 = 0.45, P = 0.72; Table 3). There was large variation in the area and percentage of home ranges shared among the 3 types of lizard pairs (Appendix A). For example, the 95% MCP home ranges of one male:male pair of lizards lined up on their edges, whereas another male:male pair shared a considerable portion of their home ranges (Figure 5). Furthermore, the home ranges of another male:male pair of lizards overlapped at their edges, until the death of one of the lizards. Subsequently, the remaining lizard expanded its home range into the home range of the dead lizard (Figure 6). Similar patterns were observed among adult female lizards (Figure 7). Although the number of pairs of lizards whose home ranges overlapped was reduced when the 75% MCP method was used, there was still no variation within and between sexes in the amount of overlap (F 2 = 0.41, P = 0.67; Table 4; Figures 8 and 9). Similarly, the percentage overlap did not vary within and between sexes (F 3 = 1.52, P = 0.22; Table 4; Appendix B). 53
64 Seasonal Movements Daily movements of male lizards from telemetry data were larger than female lizards in May (χ 2 = 5.40, df = 1, P = 0.02), but there were no differences in June (χ 2 = 0.39, df = 1, P = 0.53) or July (χ 2 = 1.62, df = 1, P = 0.20; Table 5). Within each sex, daily movements did not vary across months (males: χ 2 = 3.96, df = 1, P = 0.14; females: χ 2 = 4.32, df = 2, P = 0.12). Qualitatively, movements were greatest in May for males and June for females. Mean (± 1 SE) length of fluorescent powder trails of adult male lizards during the mating season (103.6 ± 11.6 m; n = 6) was substantially longer than trail length of adult males post-mating season (43.5 ± 5.6 m; n = 4; T 8 = 3.95, P = 0.004). The average daily movements (mean ± 1 SE) for both methods and sexes were 25.0 ± 4.7 m (range = 2.5 to m). Habitat Macrohabitat. For second-order selection, when the value used for zero substitution in used habitat was 0.1, habitat use differed from habitat availability (Table 6; λ = 0.57, χ 2 = 13.53, P = 0.004, n = 24). The macrohabitat categories were ranked in the order: nonnative prairie > developed habitat > native prairie > redcedar shrubland (Table 7). The only statistically significant comparison was that developed habitat was selected more than redcedar shrubland (Table 7). When the value used for zero substitution in used habitat was 0.4, habitat use again differed from habitat availability (λ = 0.52, χ 2 = 15.83, P = ). The macrohabitat categories were ranked in the order: developed habitat > nonnative habitat > native prairie > redcedar shrubland (Table 8). Again, the only statistically significant result was that developed habitat was selected more than redcedar shrubland (Table 8). 54
65 For third-order selection, when the value used for zero substitution in used habitat was 0.01, habitat use differed from habitat availability (Table 6; λ = 0.69, χ 2 = 9.03, P = 0.03, n = 24). The macrohabitat categories were ranked in the order: nonnative prairie > native prairie > developed habitat > redcedar shrubland (Table 9). The only statistically significant comparison was that nonnative prairie was selected more than redcedar shrubland (Table 9). When the value used for zero substitution in used habitat was 0.4, habitat use did not differ from habitat availability (λ = 0.79, χ 2 = 5.66, P = 0.13). However, rank of the habitat categories remained in the same order as above (nonnative prairie > native prairie > developed habitat > redcedar shrubland), but there were no statistically significant differences between habitat categories. Microhabitat. I collected microhabitat data at 869 locations used by lizards and at 869 paired random locations. There was a difference between microhabitat categories at lizard and random locations (Wilk s λ = 0.93, F 6,1715 = 21.28, P < ), and location differences did not interact with month (Wilk s λ = 0.99, F 18, = 1.00, P = 0.46) or time (Wilk s λ = 0.34, F 12,3430 = 1.12, P = 0.34). Bonferroni adjustments were used for multiple comparisons and were considered statistically significant at α = (Tabachnick and Fidell 1989). There was less grass (F 1 = 15.12, P = 0.001) and litter (F 1 = 75.73, P < 0.001), more shrub/tree canopy (F 1 = 12.80, P = ), bare ground (F 1 = 18.66, P < 0.001), and rock (F 1 = 47.08, P < 0.001), and no difference in forb canopy (F 1 = 1.74, P = 0.19) at lizard locations compared to random locations (Figure 10). Microhabitat vegetation varied by a simple interaction between month and time, and this interaction remained when the analysis was run without the random locations (Wilk s λ = 0.93, F 36, = 1.84, P = ). After the Bonferroni adjustments, only 55
66 percentage of litter cover at lizard locations varied by a month and time interaction (F 6 = 4.24, P = ). Lizard locations contained less litter in the morning and evening than the afternoon in May, but not during June-August (Figure 11). The percentage of rock at lizard locations varied by time (Wilk s λ = 0.93, F 12,1704 = 2.78, P = ). More rock was used in the morning and evening than the afternoon (F 2 = 7.80, P = ). Habitat Quality. There was a positive relationship between the size of 95% FK home ranges and the percentage of developed habitat (r = 0.57, n = 13, P = 0.042) and redcedar shrubland (r = 0.72, P = 0.006) within the home ranges of female lizards. There was a negative relationship between 95% FK and the percentage of nonnative prairie within home ranges (r = -0.63, P = 0.022) and no relationship to the percentage of native prairie within 95% FK home ranges for female lizards (r = 0.40, P = 0.17). There was no relationship between the size of the 95% MCP home ranges and the percentage of the 4 macrohabitat categories within a female s home range (r < 0.43, P > 0.14). There was a positive relationship between the size of the 95% FK home-range estimate and the percentage of native prairie within the home range of male lizards (r = 0.61, n = 11, P = 0.046). There was no relationship between the size of the 95% FK home ranges and the percentage of developed habitat (r = -0.47, P = 0.15), redcedar shrubland (r = -0.43, P = 0.18), or nonnative prairie (r = -0.41, P = 0.22) within the home ranges of male lizards. Similarly, there was no relationship between the size of the 95% MCP home ranges and the percentage of the 4 macrohabitat categories within a male s home range (r < 0.38, P > 0.25). 56
67 DISCUSSION Home Ranges Home-range dynamics in P. cornutum may be determined by a complex interaction of habitat types, food resources, energetics, and behavior, as seen in the territorial lizard, Sceloporus merriami by Ruby and Dunham (1987) at 3 study sites along an elevational gradient. In that study, although male and female lizards had similar body sizes, male lizards had larger home-range sizes than needed based on food levels and energetic requirements. These home ranges may enable males to have exclusive access to females. Subordinate males have even larger home ranges that enable them to avoid dominant males by using areas between the home ranges of dominant males. They also found a correlation between home-range size and the length of daily activity time based on thermal constraints. In my study, many differences in home range use between males and females appear to be tied to their different reproductive strategies. My discussion of home-range dynamics in Texas horned lizards will focus on these links. Home-Range Size. Active season home-range size did not differ in a statistically significant way between adult male and female lizards for both methods of estimation; however, the smallest 95% MCP among female lizards was substantially less than the smallest 95% MCP among male lizards (0.03 ha and 0.14 ha, respectively). Male lizards may require a larger minimum area to acquire necessary resources (e.g., females) consistent with the hypothesis that male horned lizards are selected to be small and mobile, and increase movements during the mating season to find and mate with females (Zamudio 1998, Stark et al. 2005). A lack of difference in overall home-range size 57
68 between male and female Texas horned lizards also has been found for a Texas population (Burrow et al. 2002). Body size was consistently and positively correlated to home-range size for male lizards, but not for females. Larger males may simply need larger areas to procure adequate food resources. Males would benefit from large home-range sizes because it would enable them to encounter many females during the mating season. Older and, therefore, larger male lizards may be able to allocate more energy to travel and less to growth or may have more energy stores. Unfortunately, I did not have adequate data to test if daily distance traveled by male lizards during the mating season is correlated to body size. In addition, larger male horned lizards may be dominant over smaller males during encounters as in territorial phrynosomatine lizards (Stamps 1977, Ruby and Dunham 1987, Hews 1993). If true, then male body size may be a tradeoff between small size for mobility (Zamudio 1998) and large size for dominance and energy allocation for movement. Female body size was not consistently correlated to home-range size. Female lizards may use their home ranges for activities such as feeding, thermoregulation, predator avoidance, and nesting and, therefore, are more likely to adjust their home ranges based on resource density (Simon 1975, Ims 1987, Hews 1993). Hews (1993) found that females of another phrynosomatine lizard, Uta palmeri, shifted home ranges in response to food supplementation, whereas male lizards did not. In addition, increased variation in body mass among females may weaken body size to home-range correlations. For example, gravid females probably have reduced movements due to extra body weight and the importance of evading notice of predators while gravid. 58
69 However, they may increase movements and home-range size for a brief period as they search for an ideal nesting location (Burrow 2000). For the remainder of the year, if they can meet resource requirements within a small area, there is no need to increase their movements. The average home-range size of Texas horned lizards in this study (0.50 ± 0.09 ha by the 95% MCP method and 0.87 ± 0.11 ha by the 95% FK method) was smaller than home ranges reported by Moeller (2004) for P. cornutum in the Chaparral Wildlife Management Area of Texas, but similar to home-range size reported by Fair and Henke (1999) for a different Texas population. Moeller (2004) reported average home-range sizes (± 1 SE) of 2.22 ± 0.57 ha (range: 0.06 to ha) for the 95% MCP method and 2.71 ± 0.59 ha (range: 0.09 to ha) for the 95% FK method. One reason for this difference in home-range size is that the body size of P. cornutum in Texas is larger than the body sizes of lizards in Oklahoma (see Chapter 1). Also, the Chaparral WMA is a large area of contiguous habitat that is more xeric than Tinker AFB and thus may have lower density of lizard resources, especially food. Fair and Henke (1999) reported average home-range sizes (± SE) of 0.73 ± 0.21 ha (range: 0.03 to 1.47 ha) for the 100% MCP method. However, these home-range estimates were based on points collected over only 5 days per month for 1 to 6 months; therefore, the home-range estimate may have been an underestimate. Home-Range Overlap. The factors that affect home-range overlap in P. cornutum are unclear. Distribution of resources may affect whether lizards are able to share an area. For example, competition may be lax and lizards may allow home-range overlap with other lizards or do not participate in mutual avoidance in areas of high ant density. In 59
70 addition, male lizards may be more tolerant of other males during the post-mating season. Fair and Henke (1999) found minimal overlap among weekly home ranges of Texas horned lizards compared to overlap observed among total home-range areas. They believed their data on territoriality were inconclusive because they did not observe lizards defending an area and the lack of overlap in weekly home ranges could be due to mutual avoidance to reduce competition. Although my data did not show a statistically significant pattern in home-range overlap, the placement of lizard home ranges is striking (Figures 3 9). Many of the home ranges of pairs of male or female lizards were contiguous, which supports the theory that Texas horned lizards have low-level homerange defense or mutual avoidance of areas. Seasonal Movements Similar to Stark et al. (2005), I found evidence that adult male P. cornutum have longer daily movements during the mating season compared to adult females at that time and adult males post-mating season. I agree with the conclusions of Stark et al. (2005) that males have increased movements during this period to locate receptive females. The average overall daily movements of P. cornutum in my study (25 ± 4.7 m) are slightly shorter than those reported in other studies, which have reported means ranging from 36.5 to 46.8 m/day (Whitford and Bryant 1979, Fair and Henke 1999, Stark et al. 2005). Stark et al. (2005) used fluorescent powder-tracking and Whitford and Bryant (1979) used intensive observations to calculate total distances traveled, whereas Fair and Henke (1999) calculated minimum distances between telemetry points. I used both radiotelemetry and fluorescent powder-tracking methods in my analysis of daily movements. 60
71 Habitat Macrohabitat. The value used for zero substitution in used habitat categories affected the results, but it is not clear that the results were affected in the same manner demonstrated by Bingham and Brennan (2004). For second-order selection, increasing the zero substitution value for used habitat from 0.1 to 0.4 altered the rank of the habitat with the smallest availability percentage (developed habitat), but did not change the statistically significant differences between habitat types. In both cases, developed habitat was used significantly more than redcedar shrubland. For third-order selection, increasing the zero substitution value for used habitat from 0.01 to 0.4 affected whether or not significant differences were found between habitat categories, but did not change the rank of habitat categories. In this case, there was not a single habitat category that consistently had small values for availability percentages. Therefore, it is not clear if the difference in P values was a result of inflated Type 1 error. Second-order selection, which examined placement of lizard home ranges within the study area, showed that lizards were selecting for developed habitat and avoiding redcedar shrubland. Study animals positioned their home ranges around the paved and gravel trails within the study site. P. cornutum requires a mixture of open and vegetated habitats to meet thermoregulation requirements (Fair and Henke 1998, Burrow et al. 2001). Open areas are used for basking, whereas shaded areas are used as shelter from high temperatures. The gravel and paved trails within the study site are an important resource to P. cornutum because the surrounding vegetation is dense, due to suppression of burning and other natural disturbances, and lizards often use these trails for basking and traveling. Fair and Henke (1997) found the highest abundance of P. cornutum and 61
72 their scat on plots that were managed with burning compared to plots managed with disking, a combination of burning and disking, grazing, or plots within the Conservation Reserve Program (CRP). They believed that fire cleared ground litter and facilitated lizard movement. Without the use of these open trails at Tinker Air Force Base, lizards may not be able to locate enough areas of suitable habitat for thermoregulation and traveling. The redcedar shrubland habitat is probably too shaded and provides inadequate area for thermoregulation. It should be noted that selection for the developed trails in this analysis may be biased by the sample of lizards tracked with telemetry. Nine of 21 lizards that were tracked were initially found on the gravel or paved trails. I often used the trails while walking and it was easier to spot lizards on the trails than in dense vegetation. Thus, lizards initially found on trails would be more likely to be found subsequently on the trail system. Nevertheless, two of the telemetered lizards do not appear to have crossed a gravel or paved trail at any time and another 3 lizards only crossed a trail one time. A few lizards consistently stayed close to the trail system, but most lizards used areas away from developed trails more. While being tracked, all lizards that used the trails moved an average (± 1 SE) of 33 ± 4 m (range: 8 to 100 m) away from the trail system. Third-order selection, which examined lizard habitat use within a home range, showed that lizards were selecting for nonnative prairie and avoiding redcedar shrubland. There was not a statistically significant difference between the use of nonnative and native prairie. Native and nonnative prairie species provide the vegetation structure needed by P. cornutum to meet its cover and thermoregulation requirements. Without natural disturbances, both of these habitat types would become equally dense and P. 62
73 cornutum may not prefer one over the other. Developed habitat was ranked third, suggesting that although lizards may need trails within their home ranges, they spend more time in the vegetation around the trails. Again, redcedar shrubland was avoided, perhaps due to the increase in shaded areas within this habitat. McIntyre (2003) found that vegetation species alone did not predict the occurrence of P. cornutum, as the land-use history of a grassland area and presence of prey species also are very important. McIntyre (2003) found that the species of vegetation present may affect the abundance of the harvester ant, Pogonomyrmex spp., which is a preferred prey of P. cornutum. This ant species is not present within my study site, but extant ants may be affected similarly by vegetation species. McIntyre (2003) found the lowest density of ant mounds within CRP plots planted with Old World bluestem and the highest density of ant mounds within indigenous prairie. However, there was not a significant difference between the harvester ant density of CRP plots planted with exotic or native prairie grasses. Furthermore, P. cornutum were found only within plots where Pogonomyrmex spp. also were found and were absent when Pogonomyrmex spp. were absent. I did not collect data on prey species abundance; therefore, I do not know how this factor may have influenced macrohabitat selection. Microhabitat. The difference in the percentage of microhabitat categories at locations used by lizards and random locations indicates that lizards are selecting for areas of less dense vegetation. As discussed above, the vegetation within the study site is dense due to the suppression of natural disturbances. Although these lizards use and travel through areas of dense vegetation, they require more open areas for thermoregulation and other activities. There are few open patches within the study site and lizards appear to be 63
74 selecting for these areas. Gravel trails were often the source of rock microhabitat and it is logical that lizards would use this substrate more in the morning and evening when they would need to raise their body temperatures by basking. The percentage of shrub/tree canopy was higher at locations used by lizards compared to random locations; however, the percentage of shrub/tree was < 5% at both location types. During high temperatures, lizards would sometimes be found in the shade of redcedar shrubs, but the probability of the random point falling on another redcedar shrub was small. The shade produced by the redcedars was not the only source of shade available to the lizards, but probably was easier to access than burrowing under vegetation. Habitat Quality. Relationships between habitat composition and home-range size were stronger for female than male horned lizards. Assuming an inverse relationship between home-range size and habitat quality, the positive relationship between female home-range size and the proportion of a home range containing either developed habitat or redcedar shrubland suggests that these two categories provide poor-quality habitat for P. cornutum. In developed habitat, lizards may need to travel to vegetated areas to find shelter from predators and heat, whereas redcedar shrubland may be too shaded for the lizards. As the percentage of nonnative prairie increased in the home ranges of female lizards, the size of their home range decreased, whereas there was no relationship between the percentage of native prairie and home-range size. As in the macrohabitat analysis, it is possible that nonnative and native prairie provides similar habitat quality to P. cornutum. It is unclear why nonnative prairie seems to provide slightly higher quality habitat than native prairie. It also should be noted that macrohabitat categories in this analysis are dependent on one another because the habitat compositions in a home range 64
75 add to 1. Interestingly, an area of about 3 ha of nonnative prairie in the southern portion of the study site was mowed on 18 May 2005 and treated with herbicide in mid-june for native prairie restoration. These treatments opened up the vegetation and may have made travel and thermoregulation easier for lizards in this area. Two female lizards whose home ranges were composed of at least 98% nonnative prairie in this area had the smallest and third smallest home-range sizes of all females in the study. A relationship between female home-range size and habitat quality has been shown in other studies (Simon 1975, Ims 1987). Ferguson et al. (1983) found that supplementally fed juvenile Sceloporus undulatus had smaller home ranges, higher home-range overlap, and exhibited reduced territorial behavior compared to control juveniles. Waldschmidt (1983) found that supplemental feeding caused Uta stansburiana to use their home ranges at a slower rate; however, there was no difference between the home-range size of supplementally fed and unfed lizards, whereas Hews (1993) found that female, but not male, Uta palmeri, shifted the location of their home ranges in response to supplemental feeding. The lack of a relationship between home-range size and habitat composition of the home range of male Texas horned lizards may be explained by the different reproductive strategy of males. Because horned lizards are ground dwellers in areas with few perches and individuals are widely scattered, it has been proposed that male P. cornutum increase their movements during the breeding season to encounter more females (Lynn 1965, Zamudio 1998, Stark et al. 2005). Although the distribution of thermal and predator refuges and food resources undoubtedly affect male ranging behavior, the added variation associated with mate-searching means that size of a male 65
76 lizard s home range may be only indirectly related to habitat composition as it pertains to female distribution. Difference in space use and movements of male and female P. cornutum may be explained by sex-specific reproductive strategies. Whereas male lizards would benefit by encountering more females during the mating season, female lizards may benefit from adjusting areas of use and their movements based on resource requirements. These differences may have resulted in male lizards having a larger minimum home-range size than female lizards, longer daily movements than females during the mating season, and lack of a correlation between home-range size and habitat composition in males. Management Considerations My assessment of Texas horned lizard biology provides some insight into management of the species on Tinker Air Force Base. A better mixture of bare ground and vegetated areas in the grasslands of Wildlife Reserve 3 would likely expand usable space for horned lizards. Less dense vegetation composition is likely more important to P. cornutum than the species of vegetation present as lizards do not appear to select native prairie species over nonnative prairie species. However, it may be more efficient to manage native prairie species through treatments that would reduce the density of ground cover and vegetation. Generally, lizards appear to avoid areas containing redcedar shrubs; therefore, removal of redcedars or prevention of expansion of redcedar shrublands also will increase the area inhabited by P. cornutum. Herbicide, burning, or mowing treatments may provide the necessary habitat manipulation. Predation pressure on the population appears light and predator management is not warranted. Light human-related mortality occurs, but is difficult to predict and 66
77 minimize. This population appears to be stable based on the estimated value of λ of 0.93 ± 0.07 (see Chapter 1 and 3). Determination of density of horned lizards and assessment of population trends remain difficult questions to address due to the cryptic nature of the species. The best way to estimate abundance and density of P. cornutum is monthly mark-recapture periods. The method could be implemented every few years to assess long-term population trends. Four periods (May August) consisting of 20 person-hours of searching should yield enough data to determine current population size. Researchers should focus search periods to times of ideal weather (e.g., sunny, o C, light wind speed, see Chapter 3) and should cover the entire study site, but focus searching to areas of gravel and paved trails and patches of bare ground to increase chance of spotting lizards. LITERATURE CITED Aebischer, N. J., P. A. Robertson, and R. E. Kenward Compositional analysis of habitat use from animal radio-tracking data. Ecology 74: Beyer, H. L Hawth s Analysis Tools for ArcGIS. Available at Bingham, R. L., and L. A. Brennan Comparison of type I error rates for statistical analyses of resource selection. Journal of Wildlife Management 68: Burrow, A. L The effect of prescribed burning and grazing on the threatened Texas horned lizard (Phrynosoma cornutum) in the western Rio Grande Plains. Unpubl. Thesis, Oklahoma State University, Stillwater. 67
78 Burrow, A. L., R. T. Kazmaier, E. C. Hellgren, and D. C. Ruthven, III The effects of burning and grazing on survival, home range, and prey dynamics of the Texas horned lizard in a thornscrub ecosystem. In W. M. Ford, K. R. Russell, and C. E. Moorma (eds.), The role of fire in nongame wildlife management and community restoration: traditional uses and new directions: proceedings of a special workshop, pp USDA Forest Service Northeastern Research Station General Technical Report NE-288. Burrow, A. L., R. T. Kazmaier, E. C. Hellgren, and D. C. Ruthven Microhabitat selection by Texas horned lizards in southern Texas. Journal of Wildlife Management 65: Dorr, J. L., P. Swint, and V. R. Emrick Tinker Air Force Base vegetative communities map and classification system. Conservation Management Institute- Military Lands Division, College of Natural Resources, Virginia Polytechnic Institute and State University. CMI-MLD-2005-R34. Fair, W. S., and S. E. Henke Effects of habitat manipulation on Texas horned lizards and their prey. Journal of Wildlife Management 61: Fair, W. S., and S. E. Henke Habitat use of Texas horned lizards in southern Texas. Texas Journal of Agriculture and Natural Resources 11: Fair, W. S., and S. E. Henke Movements, home ranges, and survival of Texas horned lizards (Phrynosoma cornutum). Journal of Herpetology 33: Ferguson, G. W., J. L. Hughes, and K. L. Brown Food availability and territorial establishment of juvenile Sceloporus undulatus. Pages in R. B. Huey, E. 68
79 R. Pianka, and T. W. Schoener, editors. Lizard ecology: Studies of a model organism. Harvard University Press, Cambridge. Germaine, S. S., and B. F. Wakeling Lizard species distributions and habitat occupation along an urban gradient in Tucson, Arizona, USA. Biological Conservation 97: Hews, D. K Food resources affect female distribution and male mating opportunities in the iguanian lizard Uta palmeri. Animal Behaviour 46: Hooge, P. N., W. Eichenlaub, and E. Solomon The animal movement program. USGS, Alaska Biological Service Center. Huey, R. B., and E. R. Pianka Seasonal variation in thermoregulatory behavior and body temperature of diurnal Kalahari lizards. Ecology 58: Ims, R. A Responses in spatial organization and behaviour to manipulations of the food resource in the vole Clethrionomys rufocanus. Journal of Animal Ecology 56: Johnson, D. H The comparison of usage and availability measurements for evaluating resource preference. Ecology 61: Krekorian, C. O Home range size and overlap and their relationship to food abundance in the desert iguana, Dipsosaurs dorsalis. Herpetologica 32: Lynn, R. T A comparative study of display behavior in Phrynosoma (Iguanidae). The Southwestern Naturalist 10: McIntyre, N. E Effects of conservation reserve program seeding regime on harvester ants (Pogonomyrmex), with implications for the threatened Texas horned lizard (Phrynosoma cornutum). The Southwestern Naturalist 48:
80 Moeller, B. A Conservation ecology of the Texas horned lizard (Phrynosoma cornutum): comparative effects of summer and winter burning. Oklahoma State University, Stillwater. Munger, J. C Home ranges of horned lizards (Phrynosoma): circumscribed and exclusive? Oecologia 62: Perry, G., and T. J. Garland Lizard home ranges revisited: effects of sex, body sixe, diet, habitat, and phylogeny. Ecology 83: Rose, B Lizard home ranges: methodology and functions. Journal of Herpetology 16: Ruby, D. E., and A. E. Dunham Variation in home range size along an elevational gradient in the iguanid lizard Sceloporus merriami. Oecologia 71: SAS Institute SAS release 8.1. SAS Institute, Cary, North Carolina, USA. Simon, C. A The influence of food abundance on territory size in the iguanid lizard Sceloporus jarrovi. Ecology 56: Smith, P. G Compos Analysis, version 6.2 plus [software]. Smith Ecology Ltd., Abergavenny, UK. Stamps, J. A Social behavior and spacing patterns in lizards. Pages in C. Gans, and D. W. Tinkle, editors. Biology of the Reptilia Vol. 7. Academic Press, New York. Stapley, J Do mountain log skinks (Pseudemoia entrecasteauxii) modify their behaviour in the presence of two predators? Behavioral Ecology and Sociobiology 56:
81 Stark, R. C Habitat use, daily movements, and body size of the Texas horned lizard in an urban environment in north-central Oklahoma. Unpubl. Thesis, Oklahoma State University, Stillwater. Stark, R. C., S. F. Fox, and D. M. Leslie, Jr Male Texas horned lizards increase daily movements and area covered in spring: a mate searching strategy? Journal of Herpetology 39: Stenger, J Food habits and available food of ovenbirds in relation to territory size. Auk 75: Stone, P. A., and T. A. Baird Estimating lizard home range: the Rose model revisited. Journal of Herpetology 36: Tabachnick, B. G., and L. S. Fidell Using Multivariate Statistics. Harper Collins Publishers, New York, New York, USA. Waldschmidt, S The effect of supplemental feeding on home range size and activity patterns in the lizard Uta stansburiana. Oecologia 57:1-5. Whitford, W. B., and W. G. Whitford Combat in the horned lizard, Phrynosoma cornutum. Herpetologica 29: Whitford, W. G., and M. Bryant Behavior of a predator and its prey: the horned lizard (Phrynosoma cornutum) and harvester ants (Pogonomyrmex spp.). Ecology 60: Whiting, M. J., J. R. Dixon, and R. C. Murray Spatial distribution of a population of Texas horned lizards (Phrynosoma cornutum: Phrynosomatidae) relative to habitat and prey. The Southwestern Naturalist 38:
82 Worton, B. J Kernel methods for estimating the utilization distribution in homerange studies. Ecology 70: Zamudio, K. R The evolution of female-biased sexual size dimorphism: a population-level comparative study in horned lizards (Phrynosoma). Evolution 52:
83 Table 1. The number of points, area of 95% minimum convex polygon (MCP), and area of 95% fixed kernel (FK) active- season home ranges of lizards tracked at Tinker Air Force Base, Oklahoma County, Oklahoma, May 2004 to October Lizard ID Sex Number of points 95% MCP (ha) 95% FK (ha) 65 Female Female Female Female b Female a Female Female b Female Female Female a Female a Female b Female Male Male Male Male Male Male
84 Table 1. cont. 95 Male Male Male Male Male
85 Table 2. Correlation between body size and home-range size of adult male and female lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, Home-range method Body measurement Sex P-value Correlation coefficient (r) 95% MCP Snout-vent length Male % MCP Total length Male % MCP Mass Male % FK Snout-vent length Male % FK Total length Male % FK Mass Male % MCP Snout-vent length Female % MCP Total length Female % MCP Mass Female % FK Snout-vent length Female % FK Total length Female % FK Mass Female
86 Table 3. Mean (± 1 SE) area (ha) and mean (± 1 SE) percentage of lizard s 95% MCP home ranges that overlapped between and within sexes of Texas horned lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, Area of home-range overlap Percentage of home-range overlap Comparison n Mean (± 1 SE) n Mean (± 1 SE) Male:male Female:female Female:male Male:female Overlap of females territories 2 Overlap of males territories
87 Table 4. Mean (± 1 SE) area (ha) and mean (± 1 SE) percentage of lizard s 75% MCP home ranges that overlapped between and within sexes of Texas horned lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, Area of home-range overlap Percentage of home-range overlap Gender pairing N Mean (± 1 SE) n Mean (± 1 SE) Male:male Female:female Female:male Male:female Overlap of females territories 2 Overlap of males territories
88 Table 5. Mean (± 1 SE) daily distances (m) traveled per month by Texas horned lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, Sex Month n Mean SE Male May Male June Male July Female May Female June Female July
89 Table 6. Mean percentage of available and used macrohabitat categories in analysis of second- and third-order habitat selection by Texas horned lizards at Tinker Air Force Base, Oklahoma County, Oklahoma, Developed habitat Redcedar shrubland Native Prairie Nonnative Prairie Second-order selection Study site (Available) % MCP home ranges (Used) Third-order selection 79 95% FK home ranges (Available) Lizard locations (Used)
90 Table 7. Ranking matrix for second-order habitat selection, with zero values in used habitat replaced by 0.01, by Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Nonnative prairie Nonnative prairie Developed habitat Native prairie Redcedar shrubland Rank + a Developed habitat Native prairie Redcedar shrubland _ _ 0 a Three + s or three s indicate that a habitat category in the first column was used significantly more or less, respectively, than a habitat category in the top row. One + or one indicates that a habitat category was used more or less, respectively, but the difference was not statistically significant. 80
91 Table 8. Ranking matrix for second-order habitat selection, with zero values in used habitat replaced by 0.4, by Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Developed habitat Developed habitat Nonnative prairie Native prairie Redcedar shrubland Rank + a Nonnative prairie Native prairie Redcedar shrubland _ _ 0 a Three + s or three s indicate that a habitat category in the first column was used significantly more or less, respectively, than a habitat category in the top row. One + or one indicates that a habitat category was used more or less, respectively, but the difference was not statistically significant. 81
92 Table 9. Ranking matrix for third-order habitat selection, with zero values in used habitat replaced by 0.1, by Texas horned lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, Nonnative prairie Nonnative prairie Native prairie Developed habitat Redcedar shrubland Rank + a Native prairie Developed habitat Redcedar shrubland _ _ 0 a Three + s or three s indicate that a habitat category in the first column was used significantly more or less, respectively, than a habitat category in the top row. One + or one indicates that a habitat category was used more or less, respectively, but the difference was not statistically significant. 82
93 Figure 1. Study site (Reserve 3) at Tinker Air Force Base, Oklahoma County, Oklahoma. A shows the location of Tinker Air Force Base within Oklahoma, B shows the extent of Tinker Air Force base and location of the study site within Tinker Air Force Base, and C is a close-up view of the study site. 83
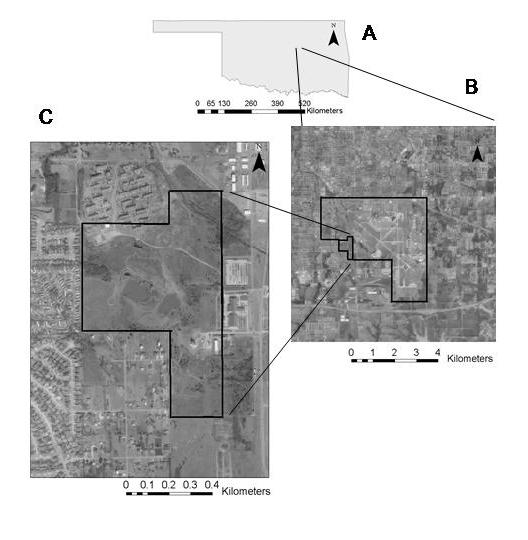
94 84
95 Figure 2. Cumulative home-range size for radiolocation points collected for lizards on Tinker Air Force Base, Oklahoma County, Oklahoma,
96 Number of points Cumulative home range size (ha)de 86
97 Figure 3. Extent of home-range overlap using the 95% minimum convex polygon estimate for adult male lizards during (A) 2004 and (B) 2005 at Tinker Air Force Base, Oklahoma County, Oklahoma. 87

98 A) B) 88
99 Figure 4. Extent of home-range overlap using the 95% minimum convex polygon estimate for adult female lizards during (A) 2004 and (B) 2005 at Tinker Air Force Base, Oklahoma County, Oklahoma 89
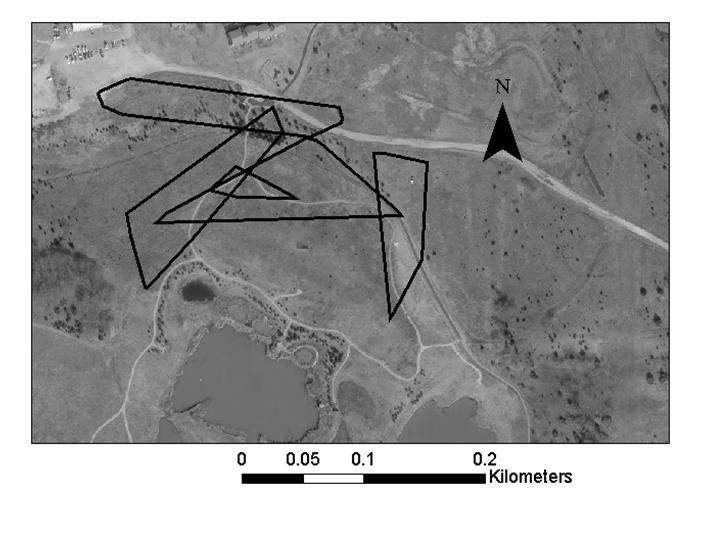

100 A) B) 90
101 Figure 5. Home-range overlap of two pairs of male lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, A shows two adult males whose home ranges marginally overlapped, while B shows two adult males with extensive home-range overlap. 91

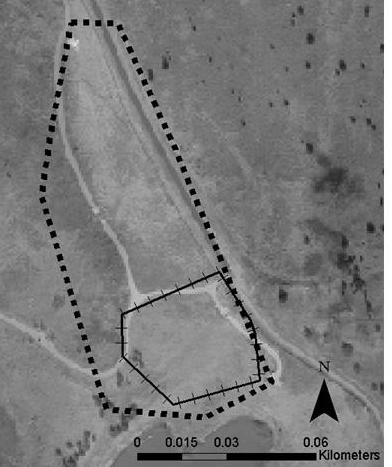
102 A) B) 92
103 Figure 6. Home-range overlap of a pair of adult male lizards on Tinker Air Force Base, Oklahoma County, Oklahoma. A shows both home ranges prior to the death of one lizard and B shows the expansion of the home range of the remaining lizard. 93


104 A) B) 94
105 Figure 7. Home-range overlap of two pairs of female lizards on Tinker Air Force Base, Oklahoma County, Oklahoma, A shows two adult females whose home ranges marginally overlapped, while B shows two adult females with extensive home-range overlap. 95


106 A) B) 96
107 Figure 8. Extent of home-range overlap using the 75% minimum convex polygon estimate for adult male lizards during (A) 2004 and (B) 2005 at Tinker Air Force Base, Oklahoma County, Oklahoma 97
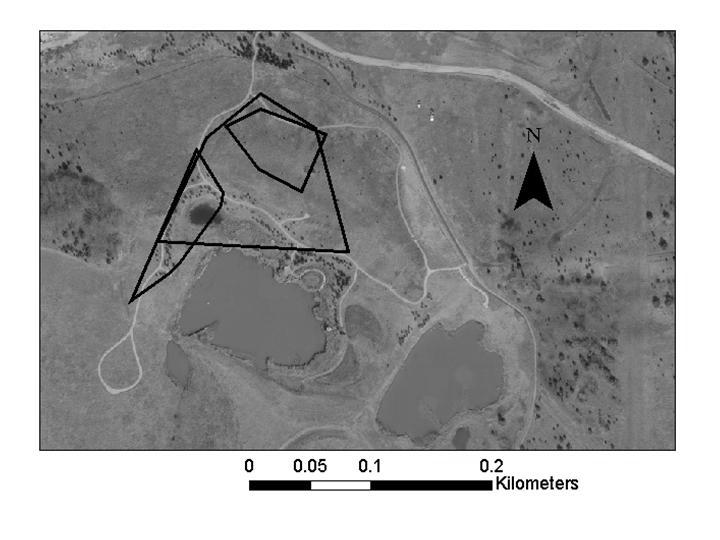
108 A) B)\ 98
109 Figure 9. Extent of home-range overlap using the 75% minimum convex polygon estimate for adult female lizards during (A) 2004 and (B) 2005 at Tinker Air Force Base, Oklahoma County, Oklahoma 99
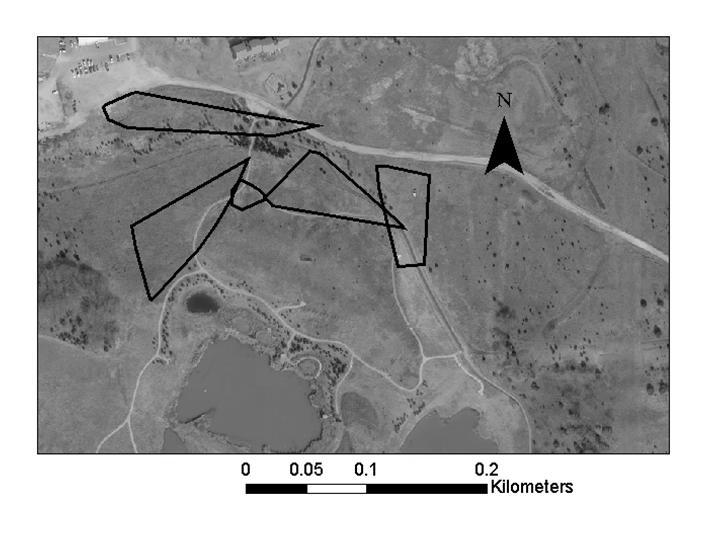

110 A) B) 100
Maturity and Other Reproductive Traits of the Kanahebi Lizard Takydromus tachydromoides (Sauria, Lacertidae) in Mito
 in Mito") Japanese Journal of Herpetology 9 (2): 46-53. 1981. Maturity and Other Reproductive Traits of the Kanahebi Lizard Takydromus tachydromoides (Sauria, Lacertidae) in Mito Sen TAKENAKA SUMMARY: Reproduction
Japanese Journal of Herpetology 9 (2): 46-53. 1981. Maturity and Other Reproductive Traits of the Kanahebi Lizard Takydromus tachydromoides (Sauria, Lacertidae) in Mito Sen TAKENAKA SUMMARY: Reproduction
Spatial and demographic ecology of Texas horned lizards within a conservation framework
 Southern Illinois University Carbondale OpenSIUC Theses Theses and Dissertations 8-1-2012 Spatial and demographic ecology of Texas horned lizards within a conservation framework Alexander J. Wolf Southern
Southern Illinois University Carbondale OpenSIUC Theses Theses and Dissertations 8-1-2012 Spatial and demographic ecology of Texas horned lizards within a conservation framework Alexander J. Wolf Southern
University of Canberra. This thesis is available in print format from the University of Canberra Library.
 University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve,
 at Sungei Buloh Wetland Reserve,") Author Title Institute Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve, Singapore Thesis (Ph.D.) National
Author Title Institute Sheikh Muhammad Abdur Rashid Population ecology and management of Water Monitors, Varanus salvator (Laurenti 1768) at Sungei Buloh Wetland Reserve, Singapore Thesis (Ph.D.) National
Managing Uplands with Keystone Species. The Case of the Gopher tortoise (Gopherus polyphemus)
") Managing Uplands with Keystone Species The Case of the Gopher tortoise (Gopherus polyphemus) Biology Question: Why consider the gopher tortoise for conservation to begin with? Answer: The gopher tortoise
Managing Uplands with Keystone Species The Case of the Gopher tortoise (Gopherus polyphemus) Biology Question: Why consider the gopher tortoise for conservation to begin with? Answer: The gopher tortoise
A.13 BLAINVILLE S HORNED LIZARD (PHRYNOSOMA BLAINVILLII)
") A. BLAINVILLE S HORNED LIZARD (PHRYNOSOMA BLAINVILLII) A.. Legal and Other Status Blainville s horned lizard is designated as a Department of Fish and Game (DFG) Species of Concern. A.. Species Distribution
A. BLAINVILLE S HORNED LIZARD (PHRYNOSOMA BLAINVILLII) A.. Legal and Other Status Blainville s horned lizard is designated as a Department of Fish and Game (DFG) Species of Concern. A.. Species Distribution
Seasonal Shifts in Reproductive Investment of Female Northern Grass Lizards ( Takydromus septentrionalis
 Seasonal Shifts in Reproductive Investment of Female Northern Grass Lizards (Takydromus septentrionalis) from a Field Population on Beiji Island, China Author(s): Wei-Guo Du and Lu Shou Source: Journal
Seasonal Shifts in Reproductive Investment of Female Northern Grass Lizards (Takydromus septentrionalis) from a Field Population on Beiji Island, China Author(s): Wei-Guo Du and Lu Shou Source: Journal
Title of Project: Distribution of the Collared Lizard, Crotophytus collaris, in the Arkansas River Valley and Ouachita Mountains
 Title of Project: Distribution of the Collared Lizard, Crotophytus collaris, in the Arkansas River Valley and Ouachita Mountains Project Summary: This project will seek to monitor the status of Collared
Title of Project: Distribution of the Collared Lizard, Crotophytus collaris, in the Arkansas River Valley and Ouachita Mountains Project Summary: This project will seek to monitor the status of Collared
Distribution, population dynamics, and habitat analyses of Collared Lizards
 Distribution, population dynamics, and habitat analyses of Collared Lizards The proposed project focuses on the distribution and population structure of the eastern collared lizards (Crotaphytus collaris
Distribution, population dynamics, and habitat analyses of Collared Lizards The proposed project focuses on the distribution and population structure of the eastern collared lizards (Crotaphytus collaris
Society for the Study of Amphibians and Reptiles
 Society for the Study of Amphibians and Reptiles Male Texas Horned Lizards Increase Daily Movements and Area Covered in Spring: A Mate Searching Strategy? Author(s): Richard C. Stark, Stanley F. Fox, David
Society for the Study of Amphibians and Reptiles Male Texas Horned Lizards Increase Daily Movements and Area Covered in Spring: A Mate Searching Strategy? Author(s): Richard C. Stark, Stanley F. Fox, David
Density, growth, and home range of the lizard Uta stansburiana stejnegeri in southern Dona Ana County, New Mexico
 Great Basin Naturalist Volume 33 Number 2 Article 8 6-30-1973 Density, growth, and home range of the lizard Uta stansburiana stejnegeri in southern Dona Ana County, New Mexico Richard D. Worthington University
Great Basin Naturalist Volume 33 Number 2 Article 8 6-30-1973 Density, growth, and home range of the lizard Uta stansburiana stejnegeri in southern Dona Ana County, New Mexico Richard D. Worthington University
ABSTRACT. Ashmore Reef
 ABSTRACT The life cycle of sea turtles is complex and is not yet fully understood. For most species, it involves at least three habitats: the pelagic, the demersal foraging and the nesting habitats. This
ABSTRACT The life cycle of sea turtles is complex and is not yet fully understood. For most species, it involves at least three habitats: the pelagic, the demersal foraging and the nesting habitats. This
ROGER IRWIN. 4 May/June 2014
 BASHFUL BLANDING S ROGER IRWIN 4 May/June 2014 4 May/June 2014 NEW HAMPSHIRE PROVIDES REGIONALLY IMPORTANT HABITAT FOR THE STATE- ENDANGERED BLANDING'S TURTLE BY MIKE MARCHAND A s a child, I loved to explore
BASHFUL BLANDING S ROGER IRWIN 4 May/June 2014 4 May/June 2014 NEW HAMPSHIRE PROVIDES REGIONALLY IMPORTANT HABITAT FOR THE STATE- ENDANGERED BLANDING'S TURTLE BY MIKE MARCHAND A s a child, I loved to explore
Rubber Boas in Radium Hot Springs: Habitat, Inventory, and Management Strategies
 : Habitat, Inventory, and Management Strategies ROBERT C. ST. CLAIR 1 AND ALAN DIBB 2 1 9809 92 Avenue, Edmonton, AB, T6E 2V4, Canada, email rstclair@telusplanet.net 2 Parks Canada, Box 220, Radium Hot
: Habitat, Inventory, and Management Strategies ROBERT C. ST. CLAIR 1 AND ALAN DIBB 2 1 9809 92 Avenue, Edmonton, AB, T6E 2V4, Canada, email rstclair@telusplanet.net 2 Parks Canada, Box 220, Radium Hot
Lizard malaria: cost to vertebrate host's reproductive success
 Parasilology (1983), 87, 1-6 1 With 2 figures in the text Lizard malaria: cost to vertebrate host's reproductive success J. J. SCHALL Department of Zoology, University of Vermont, Burlington, Vermont 05405,
Parasilology (1983), 87, 1-6 1 With 2 figures in the text Lizard malaria: cost to vertebrate host's reproductive success J. J. SCHALL Department of Zoology, University of Vermont, Burlington, Vermont 05405,
BOBWHITE QUAIL HABITAT EVALUATION
 BOBWHITE QUAIL HABITAT EVALUATION Introduction The Northern Bobwhite Quail (Colinus virginianus) is the most well known and popular upland game bird in Oklahoma. The bobwhite occurs statewide and its numbers
BOBWHITE QUAIL HABITAT EVALUATION Introduction The Northern Bobwhite Quail (Colinus virginianus) is the most well known and popular upland game bird in Oklahoma. The bobwhite occurs statewide and its numbers
MANAGEMENT OF TEXAS HORNED LIZARDS
 MANAGEMENT OF TEXAS HORNED LIZARDS Scott E. Henke and Wm. Scott Fair Research Scientists Caesar KIeberg Wildlife Research Institute Texas A&M University-Kingsville, Kingsville, Texas 78363 Abstract: Texas
MANAGEMENT OF TEXAS HORNED LIZARDS Scott E. Henke and Wm. Scott Fair Research Scientists Caesar KIeberg Wildlife Research Institute Texas A&M University-Kingsville, Kingsville, Texas 78363 Abstract: Texas
Consequences of Extended Egg Retention in the Eastern Fence Lizard (Sceloporus undulatus)
") Journal of Herpetology, Vol. 37, No. 2, pp. 309 314, 2003 Copyright 2003 Society for the Study of Amphibians and Reptiles Consequences of Extended Egg Retention in the Eastern Fence Lizard (Sceloporus
Journal of Herpetology, Vol. 37, No. 2, pp. 309 314, 2003 Copyright 2003 Society for the Study of Amphibians and Reptiles Consequences of Extended Egg Retention in the Eastern Fence Lizard (Sceloporus
A.13 BLAINVILLE S HORNED LIZARD (PHRYNOSOMA BLAINVILLII)
") A. BLAINVILLE S HORNED LIZARD (PHRYNOSOMA BLAINVILLII) A.. Legal and Other Status Blainville s horned lizard is designated as a Department of Fish and Game (DFG) Species of Concern. A.. Species Distribution
A. BLAINVILLE S HORNED LIZARD (PHRYNOSOMA BLAINVILLII) A.. Legal and Other Status Blainville s horned lizard is designated as a Department of Fish and Game (DFG) Species of Concern. A.. Species Distribution
Survivorship. Demography and Populations. Avian life history patterns. Extremes of avian life history patterns
 Demography and Populations Survivorship Demography is the study of fecundity and survival Four critical variables Age of first breeding Number of young fledged each year Juvenile survival Adult survival
Demography and Populations Survivorship Demography is the study of fecundity and survival Four critical variables Age of first breeding Number of young fledged each year Juvenile survival Adult survival
Research Summary: Evaluation of Northern Bobwhite and Scaled Quail in Western Oklahoma
 P-1054 Research Summary: Evaluation of Northern Bobwhite and Scaled Quail in Western Oklahoma Oklahoma Agricultural Experiment Station Division of Agricultural Sciences and Natural Resources Oklahoma State
P-1054 Research Summary: Evaluation of Northern Bobwhite and Scaled Quail in Western Oklahoma Oklahoma Agricultural Experiment Station Division of Agricultural Sciences and Natural Resources Oklahoma State
Lynx Update May 25, 2009 INTRODUCTION
 Lynx Update May 25, 2009 INTRODUCTION In an effort to establish a viable population of Canada lynx (Lynx canadensis) in Colorado, the Colorado Division of Wildlife (CDOW) initiated a reintroduction effort
Lynx Update May 25, 2009 INTRODUCTION In an effort to establish a viable population of Canada lynx (Lynx canadensis) in Colorado, the Colorado Division of Wildlife (CDOW) initiated a reintroduction effort
A Population Analysis of the Common Wall Lizard Podarcis muralis in Southwestern France
 - 513 - Studies in Herpetology, Rocek Z. (ed.) pp. 513-518 Prague 1986 A Population Analysis of the Common Wall Lizard Podarcis muralis in Southwestern France R. BARBAULT and Y. P. MOU Laboratoire d'ecologie
- 513 - Studies in Herpetology, Rocek Z. (ed.) pp. 513-518 Prague 1986 A Population Analysis of the Common Wall Lizard Podarcis muralis in Southwestern France R. BARBAULT and Y. P. MOU Laboratoire d'ecologie
Iguana Technical Assistance Workshop. Presented by: Florida Fish and Wildlife Conservation Commission
 Iguana Technical Assistance Workshop Presented by: Florida Fish and Wildlife Conservation Commission 1 Florida Fish and Wildlife Conservation Commission Protects and manages 575 species of wildlife 700
Iguana Technical Assistance Workshop Presented by: Florida Fish and Wildlife Conservation Commission 1 Florida Fish and Wildlife Conservation Commission Protects and manages 575 species of wildlife 700
FALL 2015 BLACK-FOOTED FERRET SURVEY LOGAN COUNTY, KANSAS DAN MULHERN; U.S. FISH AND WILDLIFE SERVICE
 INTRODUCTION FALL 2015 BLACK-FOOTED FERRET SURVEY LOGAN COUNTY, KANSAS DAN MULHERN; U.S. FISH AND WILDLIFE SERVICE As part of ongoing efforts to monitor the status of reintroduced endangered black-footed
INTRODUCTION FALL 2015 BLACK-FOOTED FERRET SURVEY LOGAN COUNTY, KANSAS DAN MULHERN; U.S. FISH AND WILDLIFE SERVICE As part of ongoing efforts to monitor the status of reintroduced endangered black-footed
Weaver Dunes, Minnesota
 Hatchling Orientation During Dispersal from Nests Experimental analyses of an early life stage comparing orientation and dispersal patterns of hatchlings that emerge from nests close to and far from wetlands
Hatchling Orientation During Dispersal from Nests Experimental analyses of an early life stage comparing orientation and dispersal patterns of hatchlings that emerge from nests close to and far from wetlands
Habitats and Field Methods. Friday May 12th 2017
 Habitats and Field Methods Friday May 12th 2017 Announcements Project consultations available today after class Project Proposal due today at 5pm Follow guidelines posted for lecture 4 Field notebooks
Habitats and Field Methods Friday May 12th 2017 Announcements Project consultations available today after class Project Proposal due today at 5pm Follow guidelines posted for lecture 4 Field notebooks
REPORT OF ACTIVITIES 2009 TURTLE ECOLOGY RESEARCH REPORT Crescent Lake National Wildlife Refuge 3 to 26 June 2009
 REPORT OF ACTIVITIES 2009 TURTLE ECOLOGY RESEARCH REPORT Crescent Lake National Wildlife Refuge 3 to 26 June 2009 A report submitted to Refuge Manager Mark Koepsel 17 July 2009 John B Iverson Dept. of
REPORT OF ACTIVITIES 2009 TURTLE ECOLOGY RESEARCH REPORT Crescent Lake National Wildlife Refuge 3 to 26 June 2009 A report submitted to Refuge Manager Mark Koepsel 17 July 2009 John B Iverson Dept. of
SURVIVAL, MOVEMENT, AND FINE-SCALE HABITAT SELECTION OF REINTRODUCED TEXAS HORNED LIZARDS (PHRYNOSOMA CORNUTUM). Alyssa A. Fink, B.S.
. Alyssa A. Fink, B.S.") SURVIVAL, MOVEMENT, AND FINE-SCALE HABITAT SELECTION OF REINTRODUCED TEXAS HORNED LIZARDS (PHRYNOSOMA CORNUTUM). by Alyssa A. Fink, B.S. A thesis submitted to the Graduate Council of Texas State University
SURVIVAL, MOVEMENT, AND FINE-SCALE HABITAT SELECTION OF REINTRODUCED TEXAS HORNED LIZARDS (PHRYNOSOMA CORNUTUM). by Alyssa A. Fink, B.S. A thesis submitted to the Graduate Council of Texas State University
Station 1 1. (3 points) Identification: Station 2 6. (3 points) Identification:
 Identification: Station 2 6. (3 points) Identification:") SOnerd s 2018-2019 Herpetology SSSS Test 1 SOnerd s SSSS 2018-2019 Herpetology Test Station 20 sounds found here: https://drive.google.com/drive/folders/1oqrmspti13qv_ytllk_yy_vrie42isqe?usp=sharing Station
SOnerd s 2018-2019 Herpetology SSSS Test 1 SOnerd s SSSS 2018-2019 Herpetology Test Station 20 sounds found here: https://drive.google.com/drive/folders/1oqrmspti13qv_ytllk_yy_vrie42isqe?usp=sharing Station
Use of Agent Based Modeling in an Ecological Conservation Context
 28 RIThink, 2012, Vol. 2 From: http://photos.turksandcaicostourism.com/nature/images/tctb_horz_033.jpg Use of Agent Based Modeling in an Ecological Conservation Context Scott B. WOLCOTT 1 *, Michael E.
28 RIThink, 2012, Vol. 2 From: http://photos.turksandcaicostourism.com/nature/images/tctb_horz_033.jpg Use of Agent Based Modeling in an Ecological Conservation Context Scott B. WOLCOTT 1 *, Michael E.
Post-Release Success of Captive Bred Louisiana Pine Snakes
 Post-Release Success of Captive Bred Louisiana Pine Snakes The Louisiana pine snake (Pituophis ruthveni) Most endangered reptile in the U.S. 1 st and only SSP for a U.S. reptile Only 6% of SSP s are for
Post-Release Success of Captive Bred Louisiana Pine Snakes The Louisiana pine snake (Pituophis ruthveni) Most endangered reptile in the U.S. 1 st and only SSP for a U.S. reptile Only 6% of SSP s are for
RADIOTELEMETRY AND POPULATION MONITORING OF SAND DUNE LIZARDS (SCELOPORUS ARENICOLUS) DURING THE NESTING SEASON, 2006
 DURING THE NESTING SEASON, 2006") Share With Wildlife Project Title: Dispersal and radio tracking of the Sand Dune Lizard, Sceloporus arenicolus RADIOTELEMETRY AND POPULATION MONITORING OF SAND DUNE LIZARDS (SCELOPORUS ARENICOLUS) DURING
Share With Wildlife Project Title: Dispersal and radio tracking of the Sand Dune Lizard, Sceloporus arenicolus RADIOTELEMETRY AND POPULATION MONITORING OF SAND DUNE LIZARDS (SCELOPORUS ARENICOLUS) DURING
Plestiodon (=Eumeces) fasciatus Family Scincidae
 fasciatus Family Scincidae") Plestiodon (=Eumeces) fasciatus Family Scincidae Living specimens: - Five distinct longitudinal light lines on dorsum - Juveniles have bright blue tail - Head of male reddish during breeding season - Old
Plestiodon (=Eumeces) fasciatus Family Scincidae Living specimens: - Five distinct longitudinal light lines on dorsum - Juveniles have bright blue tail - Head of male reddish during breeding season - Old
John Thompson June 09, 2016 Thompson Holdings, LLC P.O. Box 775 Springhouse, Pa
 John Thompson June 09, 2016 Thompson Holdings, LLC P.O. Box 775 Springhouse, Pa. 19477 Subject: Paraiso Springs Resort PLN040183 - Biological update Dear John, At your request I visited the Paraiso springs
John Thompson June 09, 2016 Thompson Holdings, LLC P.O. Box 775 Springhouse, Pa. 19477 Subject: Paraiso Springs Resort PLN040183 - Biological update Dear John, At your request I visited the Paraiso springs
Texas Quail Index. Result Demonstration Report 2016
 Texas Quail Index Result Demonstration Report 2016 Cooperators: Josh Kouns, County Extension Agent for Baylor County Amanda Gobeli, Extension Associate Dr. Dale Rollins, Statewide Coordinator Bill Whitley,
Texas Quail Index Result Demonstration Report 2016 Cooperators: Josh Kouns, County Extension Agent for Baylor County Amanda Gobeli, Extension Associate Dr. Dale Rollins, Statewide Coordinator Bill Whitley,
Habitats and Field Techniques
 Habitats and Field Techniques Keys to Understanding Habitat Shelter, Sunlight, Water, Food Habitats of Interest Rivers/Streams Lakes/Ponds Bogs/Marshes Forests Meadows Sandy Edge Habitat Rivers/Streams
Habitats and Field Techniques Keys to Understanding Habitat Shelter, Sunlight, Water, Food Habitats of Interest Rivers/Streams Lakes/Ponds Bogs/Marshes Forests Meadows Sandy Edge Habitat Rivers/Streams
GREATER SAGE-GROUSE BROOD-REARING HABITAT MANIPULATION IN MOUNTAIN BIG SAGEBRUSH, USE OF TREATMENTS, AND REPRODUCTIVE ECOLOGY ON PARKER MOUNTAIN, UTAH
 GREATER SAGE-GROUSE BROOD-REARING HABITAT MANIPULATION IN MOUNTAIN BIG SAGEBRUSH, USE OF TREATMENTS, AND REPRODUCTIVE ECOLOGY ON PARKER MOUNTAIN, UTAH Abstract We used an experimental design to treat greater
GREATER SAGE-GROUSE BROOD-REARING HABITAT MANIPULATION IN MOUNTAIN BIG SAGEBRUSH, USE OF TREATMENTS, AND REPRODUCTIVE ECOLOGY ON PARKER MOUNTAIN, UTAH Abstract We used an experimental design to treat greater
James Lowry*, Cheryl Nushardt Susan Reigler and Omar Attum** Dept. of Biology, Indiana University Southeast, 4201 Grant Line Rd, New Albany, IN 47150
 James Lowry*, Cheryl Nushardt Susan Reigler and Omar Attum** Dept. of Biology, Indiana University Southeast, 4201 Grant Line Rd, New Albany, IN 47150 * jamlowry@ius.edu ** FACULTY ADVISOR Outline Introduction
James Lowry*, Cheryl Nushardt Susan Reigler and Omar Attum** Dept. of Biology, Indiana University Southeast, 4201 Grant Line Rd, New Albany, IN 47150 * jamlowry@ius.edu ** FACULTY ADVISOR Outline Introduction
Gambel s Quail Callipepla gambelii
 Photo by Amy Leist Habitat Use Profile Habitats Used in Nevada Mesquite-Acacia Mojave Lowland Riparian Springs Agriculture Key Habitat Parameters Plant Composition Mesquite, acacia, salt cedar, willow,
Photo by Amy Leist Habitat Use Profile Habitats Used in Nevada Mesquite-Acacia Mojave Lowland Riparian Springs Agriculture Key Habitat Parameters Plant Composition Mesquite, acacia, salt cedar, willow,
Effects of prey availability and climate across a decade for a desert-dwelling, ectothermic mesopredator. R. Anderson Western Washington University
 Effects of prey availability and climate across a decade for a desert-dwelling, ectothermic mesopredator R. Anderson Western Washington University Trophic interactions in desert systems are presumed to
Effects of prey availability and climate across a decade for a desert-dwelling, ectothermic mesopredator R. Anderson Western Washington University Trophic interactions in desert systems are presumed to
Impacts of Prescribed Burning on Three Eastern Box Turtles (Terrapene carolina carolina) in Southwestern Virginia
 in Southwestern Virginia") Impacts of Prescribed Burning on Three Eastern Box Turtles (Terrapene carolina carolina) in Southwestern Virginia Todd S. Fredericksen, Gage Staton, Javin Metz Ferrum College P.O. Box 1000 Ferrum Virginia
Impacts of Prescribed Burning on Three Eastern Box Turtles (Terrapene carolina carolina) in Southwestern Virginia Todd S. Fredericksen, Gage Staton, Javin Metz Ferrum College P.O. Box 1000 Ferrum Virginia
Incubation temperature and phenotypic traits of Sceloporus undulatus: implications for the northern limits of distribution
 DOI 10.1007/s00442-006-0583-0 ECOPHYSIOLOGY Incubation temperature and phenotypic traits of Sceloporus undulatus: implications for the northern limits of distribution Scott L. Parker Æ Robin M. Andrews
DOI 10.1007/s00442-006-0583-0 ECOPHYSIOLOGY Incubation temperature and phenotypic traits of Sceloporus undulatus: implications for the northern limits of distribution Scott L. Parker Æ Robin M. Andrews
COMPARING BODY CONDITION ESTIMATES OF ZOO BROTHER S ISLAND TUATARA (SPHENODON GUNTHERI) TO THAT OF THE WILD, A CLINICAL CASE
 TO THAT OF THE WILD, A CLINICAL CASE") COMPARING BODY CONDITION ESTIMATES OF ZOO BROTHER S ISLAND TUATARA (SPHENODON GUNTHERI) TO THAT OF THE WILD, A CLINICAL CASE Kyle S. Thompson, BS,¹, ²* Michael L. Schlegel, PhD, PAS² ¹Oklahoma State University,
COMPARING BODY CONDITION ESTIMATES OF ZOO BROTHER S ISLAND TUATARA (SPHENODON GUNTHERI) TO THAT OF THE WILD, A CLINICAL CASE Kyle S. Thompson, BS,¹, ²* Michael L. Schlegel, PhD, PAS² ¹Oklahoma State University,
Commercial Collection. & Pit Fall Trap Updates. Jason L. Jones Herpetologist 23 June 2017 Commission Update
 Commercial Collection & Pit Fall Trap Updates Jason L. Jones Herpetologist 23 June 2017 Commission Update Everyone collects Everyone collects Some collections require permits Some are illegal. 16-17th
Commercial Collection & Pit Fall Trap Updates Jason L. Jones Herpetologist 23 June 2017 Commission Update Everyone collects Everyone collects Some collections require permits Some are illegal. 16-17th
Striped Skunk Updated: April 8, 2018
 Striped Skunk Updated: April 8, 2018 Interpretation Guide Status Danger Threats Population Distribution Habitat Diet Size Longevity Social Family Units Reproduction Our Animals Scientific Name Least Concern
Striped Skunk Updated: April 8, 2018 Interpretation Guide Status Danger Threats Population Distribution Habitat Diet Size Longevity Social Family Units Reproduction Our Animals Scientific Name Least Concern
Ames, IA Ames, IA (515)
") BENEFITS OF A CONSERVATION BUFFER-BASED CONSERVATION MANAGEMENT SYSTEM FOR NORTHERN BOBWHITE AND GRASSLAND SONGBIRDS IN AN INTENSIVE PRODUCTION AGRICULTURAL LANDSCAPE IN THE LOWER MISSISSIPPI ALLUVIAL
BENEFITS OF A CONSERVATION BUFFER-BASED CONSERVATION MANAGEMENT SYSTEM FOR NORTHERN BOBWHITE AND GRASSLAND SONGBIRDS IN AN INTENSIVE PRODUCTION AGRICULTURAL LANDSCAPE IN THE LOWER MISSISSIPPI ALLUVIAL
Turtle Research, Education, and Conservation Program
 Turtle Population Declines Turtle Research, Education, and Conservation Program Turtles are a remarkable group of animals. They ve existed on earth for over 200 million years; that s close to 100 times
Turtle Population Declines Turtle Research, Education, and Conservation Program Turtles are a remarkable group of animals. They ve existed on earth for over 200 million years; that s close to 100 times
NOTES ON THE ECOLOGY AND NATURAL HISTORY OF TWO SPECIES OF EGERNIA (SCINCIDAE) IN WESTERN AUSTRALIA
 IN WESTERN AUSTRALIA") NOTES ON THE ECOLOGY AND NATURAL HISTORY OF TWO SPECIES OF EGERNIA (SCINCIDAE) IN WESTERN AUSTRALIA By ERIC R. PIANKA Integrative Biology University of Texas at Austin Austin, Texas 78712 USA Email: erp@austin.utexas.edu
NOTES ON THE ECOLOGY AND NATURAL HISTORY OF TWO SPECIES OF EGERNIA (SCINCIDAE) IN WESTERN AUSTRALIA By ERIC R. PIANKA Integrative Biology University of Texas at Austin Austin, Texas 78712 USA Email: erp@austin.utexas.edu
Mexican Gray Wolf Reintroduction
 Mexican Gray Wolf Reintroduction New Mexico Supercomputing Challenge Final Report April 2, 2014 Team Number 24 Centennial High School Team Members: Andrew Phillips Teacher: Ms. Hagaman Project Mentor:
Mexican Gray Wolf Reintroduction New Mexico Supercomputing Challenge Final Report April 2, 2014 Team Number 24 Centennial High School Team Members: Andrew Phillips Teacher: Ms. Hagaman Project Mentor:
Texas Quail Index. Result Demonstration Report 2016
 Texas Quail Index Result Demonstration Report 2016 Cooperators: Jerry Coplen, County Extension Agent for Knox County Amanda Gobeli, Extension Associate Dr. Dale Rollins, Statewide Coordinator Circle Bar
Texas Quail Index Result Demonstration Report 2016 Cooperators: Jerry Coplen, County Extension Agent for Knox County Amanda Gobeli, Extension Associate Dr. Dale Rollins, Statewide Coordinator Circle Bar
Diet Selection by a Lizard Ant-Specialist in an Urban System Bereft of Preferred Prey
 Journal of Herpetology, Vol. 52, No. 1, 79 85, 2018 Copyright 2018 Society for the Study of Amphibians and Reptiles Diet Selection by a Lizard Ant-Specialist in an Urban System Bereft of Preferred Prey
Journal of Herpetology, Vol. 52, No. 1, 79 85, 2018 Copyright 2018 Society for the Study of Amphibians and Reptiles Diet Selection by a Lizard Ant-Specialist in an Urban System Bereft of Preferred Prey
A final programmatic report to: SAVE THE TIGER FUND. Scent Dog Monitoring of Amur Tigers-V ( ) March 1, March 1, 2006
 March 1, March 1, 2006") 1 A final programmatic report to: SAVE THE TIGER FUND Scent Dog Monitoring of Amur Tigers-V (2005-0013-017) March 1, 2005 - March 1, 2006 Linda Kerley and Galina Salkina PROJECT SUMMARY We used scent-matching
1 A final programmatic report to: SAVE THE TIGER FUND Scent Dog Monitoring of Amur Tigers-V (2005-0013-017) March 1, 2005 - March 1, 2006 Linda Kerley and Galina Salkina PROJECT SUMMARY We used scent-matching
Lizard Surveying and Monitoring in Biodiversity Sanctuaries
 Lizard Surveying and Monitoring in Biodiversity Sanctuaries Trent Bell (EcoGecko Consultants) Alison Pickett (DOC North Island Skink Recovery Group) First things first I am profoundly deaf I have a Deaf
Lizard Surveying and Monitoring in Biodiversity Sanctuaries Trent Bell (EcoGecko Consultants) Alison Pickett (DOC North Island Skink Recovery Group) First things first I am profoundly deaf I have a Deaf
Age structured models
 Age structured models Fibonacci s rabbit model not only considers the total number of rabbits, but also the ages of rabbit. We can reformat the model in this way: let M n be the number of adult pairs of
Age structured models Fibonacci s rabbit model not only considers the total number of rabbits, but also the ages of rabbit. We can reformat the model in this way: let M n be the number of adult pairs of
Minnesota_mammals_Info_9.doc 11/04/09 -- DRAFT Page 1 of 64. Minnesota mammals
 Minnesota_mammals_Info_9.doc 11/04/09 -- DRAFT Page 1 of 64 Minnesota mammals This is a short guide to Minnesota mammals, with information drawn from Hazard s Mammals of, Walker s Mammals of the World,
Minnesota_mammals_Info_9.doc 11/04/09 -- DRAFT Page 1 of 64 Minnesota mammals This is a short guide to Minnesota mammals, with information drawn from Hazard s Mammals of, Walker s Mammals of the World,
About Reptiles A Guide for Children. Cathryn Sill Illustrated by John Sill
 About Reptiles About Reptiles A Guide for Children Cathryn Sill Illustrated by John Sill For the One who created reptiles. Genesis 1:24 Published by PEACHTREE PUBLISHERS, LTD. 1700 Chattahoochee Avenue
About Reptiles About Reptiles A Guide for Children Cathryn Sill Illustrated by John Sill For the One who created reptiles. Genesis 1:24 Published by PEACHTREE PUBLISHERS, LTD. 1700 Chattahoochee Avenue
Desert Tortoise By Guy Belleranti
 Name: A turtle that lives on land is called a tortoise. One interesting tortoise is the desert tortoise. This reptile lives in the Sonoran and Mojave deserts of the southwestern United States and Mexico.
Name: A turtle that lives on land is called a tortoise. One interesting tortoise is the desert tortoise. This reptile lives in the Sonoran and Mojave deserts of the southwestern United States and Mexico.
The status of an introduced species (Phrynosoma cornutum) on barrier islands in South Carolina
 on barrier islands in South Carolina") Phrynosomatics The Newsletter of the Horned Lizard Conservation Society Our purpose is to document and publicize the values and conservation needs of horned lizards, to promote horned lizard conservation
Phrynosomatics The Newsletter of the Horned Lizard Conservation Society Our purpose is to document and publicize the values and conservation needs of horned lizards, to promote horned lizard conservation
reproductive life History and the effects of sex and season on morphology in CRoTALus oreganus (northern PaCifiC RATTLESNAKES)
") reproductive life History and the effects of sex and season on morphology in CRoTALus oreganus (northern PaCifiC RATTLESNAKES) Benjamin Kwittken, Student Author dr. emily n. taylor, research advisor abstract
reproductive life History and the effects of sex and season on morphology in CRoTALus oreganus (northern PaCifiC RATTLESNAKES) Benjamin Kwittken, Student Author dr. emily n. taylor, research advisor abstract
Reproduction in a Nebraska Sandhills Population of the Northern Prairie Lizard Sceloporus undulatus garmani
 University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Papers in Herpetology Papers in the Biological Sciences 7-1981 Reproduction in a Nebraska Sandhills Population of the Northern
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Papers in Herpetology Papers in the Biological Sciences 7-1981 Reproduction in a Nebraska Sandhills Population of the Northern
Observations on the response of four eastern box turtles (Terrapene carolina carolina) to clearcut logging and chipping in southern Virginia
 to clearcut logging and chipping in southern Virginia") Observations on the response of four eastern box turtles (Terrapene carolina carolina) to clearcut logging and chipping in southern Virginia Todd S. Fredericksen Joshua L. Bernard School of Natural Sciences
Observations on the response of four eastern box turtles (Terrapene carolina carolina) to clearcut logging and chipping in southern Virginia Todd S. Fredericksen Joshua L. Bernard School of Natural Sciences
Objectives: Outline: Idaho Amphibians and Reptiles. Characteristics of Amphibians. Types and Numbers of Amphibians
 Natural History of Idaho Amphibians and Reptiles Wildlife Ecology, University of Idaho Fall 2005 Charles R. Peterson Herpetology Laboratory Department of Biological Sciences, Idaho Museum of Natural History
Natural History of Idaho Amphibians and Reptiles Wildlife Ecology, University of Idaho Fall 2005 Charles R. Peterson Herpetology Laboratory Department of Biological Sciences, Idaho Museum of Natural History
PRELIMINARY EVALUATION OF THE IMPACT OF ROADS AND ASSOCIATED VEHICULAR TRAFFIC ON SNAKE POPULATIONS IN EASTERN TEXAS
 PRELIMINARY EVALUATION OF THE IMPACT OF ROADS AND ASSOCIATED VEHICULAR TRAFFIC ON SNAKE POPULATIONS IN EASTERN TEXAS D. Craig Rudolph, Shirley J. Burgdorf, Richard N. Conner, and Richard R. Schaefer, U.
PRELIMINARY EVALUATION OF THE IMPACT OF ROADS AND ASSOCIATED VEHICULAR TRAFFIC ON SNAKE POPULATIONS IN EASTERN TEXAS D. Craig Rudolph, Shirley J. Burgdorf, Richard N. Conner, and Richard R. Schaefer, U.
Status and Management of Amphibians on Montana Rangelands
 Status and Management of Amphibians on Montana Rangelands Society For Range Management Meeting February 9, 2011 - Billings, Montana Bryce A. Maxell Interim Director / Senior Zoologist Montana Natural Heritage
Status and Management of Amphibians on Montana Rangelands Society For Range Management Meeting February 9, 2011 - Billings, Montana Bryce A. Maxell Interim Director / Senior Zoologist Montana Natural Heritage
Home Range, Habitat Use, Feeding Ecology and Reproductive Biology of the Cuban Boa (Chilabothrus angulifer) at Naval Station Guantánamo Bay, Cuba
 at Naval Station Guantánamo Bay, Cuba") Home Range, Habitat Use, Feeding Ecology and Reproductive Biology of the Cuban Boa (Chilabothrus angulifer) at Naval Station Guantánamo Bay, Cuba Dr. Peter J. Tolson - Department of Conservation and Research,
Home Range, Habitat Use, Feeding Ecology and Reproductive Biology of the Cuban Boa (Chilabothrus angulifer) at Naval Station Guantánamo Bay, Cuba Dr. Peter J. Tolson - Department of Conservation and Research,
The Effects of Meso-mammal Removal on Northern Bobwhite Populations
 The Effects of Meso-mammal Removal on Northern Bobwhite Populations Alexander L. Jackson William E. Palmer D. Clay Sisson Theron M. Terhune II John M. Yeiser James A. Martin Predation Predation is the
The Effects of Meso-mammal Removal on Northern Bobwhite Populations Alexander L. Jackson William E. Palmer D. Clay Sisson Theron M. Terhune II John M. Yeiser James A. Martin Predation Predation is the
Red-Tailed Hawk Buteo jamaicensis
 Red-Tailed Hawk Buteo jamaicensis This large, dark headed, broad-shouldered hawk is one of the most common and widespread hawks in North America. The Red-tailed hawk belongs to the genus (family) Buteo,
Red-Tailed Hawk Buteo jamaicensis This large, dark headed, broad-shouldered hawk is one of the most common and widespread hawks in North America. The Red-tailed hawk belongs to the genus (family) Buteo,
ECOLOGY OF ISOLATED INHABITING THE WILDCAT KNOLLS AND HORN
 ECOLOGY OF ISOLATED GREATER SAGE GROUSE GROUSE POPULATIONS INHABITING THE WILDCAT KNOLLS AND HORN MOUNTAIN, SOUTHCENTRAL UTAH by Christopher J. Perkins Committee: Dr. Terry Messmer, Dr. Frank Howe, and
ECOLOGY OF ISOLATED GREATER SAGE GROUSE GROUSE POPULATIONS INHABITING THE WILDCAT KNOLLS AND HORN MOUNTAIN, SOUTHCENTRAL UTAH by Christopher J. Perkins Committee: Dr. Terry Messmer, Dr. Frank Howe, and
COLORADO LYNX DEN SITE HABITAT PROGRESS REPORT 2006
 COLORADO LYNX DEN SITE HABITAT PROGRESS REPORT 2006 by Grant Merrill Tanya Shenk U.S. Forest Service and Colorado Division of Wildlife Cooperative Effort September 30, 2006 INTRODUCTION Lynx (Lynx canadensis)
COLORADO LYNX DEN SITE HABITAT PROGRESS REPORT 2006 by Grant Merrill Tanya Shenk U.S. Forest Service and Colorado Division of Wildlife Cooperative Effort September 30, 2006 INTRODUCTION Lynx (Lynx canadensis)
Adjustments In Parental Care By The European Starling (Sturnus Vulgaris): The Effect Of Female Condition
: The Effect Of Female Condition") Proceedings of The National Conference on Undergraduate Research (NCUR) 2003 University of Utah, Salt Lake City, Utah March 13-15, 2003 Adjustments In Parental Care By The European Starling (Sturnus Vulgaris):
Proceedings of The National Conference on Undergraduate Research (NCUR) 2003 University of Utah, Salt Lake City, Utah March 13-15, 2003 Adjustments In Parental Care By The European Starling (Sturnus Vulgaris):
Northern Copperhead Updated: April 8, 2018
 Interpretation Guide Northern Copperhead Updated: April 8, 2018 Status Danger Threats Population Distribution Habitat Diet Size Longevity Social Family Units Reproduction Our Animals Scientific Name Least
Interpretation Guide Northern Copperhead Updated: April 8, 2018 Status Danger Threats Population Distribution Habitat Diet Size Longevity Social Family Units Reproduction Our Animals Scientific Name Least
Motuora island reptile monitoring report for common & Pacific gecko 2016
 Motuora island reptile monitoring report for common & Pacific gecko 6 Prepared by Su Sinclair August 7 Work on this monitoring project was carried out under a Wildlife Act Authority issued by the Department
Motuora island reptile monitoring report for common & Pacific gecko 6 Prepared by Su Sinclair August 7 Work on this monitoring project was carried out under a Wildlife Act Authority issued by the Department
Result Demonstration Report
 Result Demonstration Report 2014 Texas Quail Index Texas A&M AgriLife Extension Service Archer County Cooperator: Brad Mitchell- Mitchell and Parkey Ranches Justin B Gilliam, County Extension Agent for
Result Demonstration Report 2014 Texas Quail Index Texas A&M AgriLife Extension Service Archer County Cooperator: Brad Mitchell- Mitchell and Parkey Ranches Justin B Gilliam, County Extension Agent for
Great Horned Owl (Bubo virginianus) Productivity and Home Range Characteristics in a Shortgrass Prairie. Rosemary A. Frank and R.
 Productivity and Home Range Characteristics in a Shortgrass Prairie. Rosemary A. Frank and R.") Great Horned Owl (Bubo virginianus) Productivity and Home Range Characteristics in a Shortgrass Prairie Rosemary A. Frank and R. Scott Lutz 1 Abstract. We studied movements and breeding success of resident
Great Horned Owl (Bubo virginianus) Productivity and Home Range Characteristics in a Shortgrass Prairie Rosemary A. Frank and R. Scott Lutz 1 Abstract. We studied movements and breeding success of resident
REPORT OF ACTIVITIES TURTLE ECOLOGY RESEARCH REPORT Crescent Lake National Wildlife Refuge 31 May to 4 July 2017
 REPORT OF ACTIVITIES 2017 TURTLE ECOLOGY RESEARCH REPORT Crescent Lake National Wildlife Refuge 31 May to 4 July 2017 A report submitted to Refuge Biologist Marlin French 15 July 2017 John B Iverson Dept.
REPORT OF ACTIVITIES 2017 TURTLE ECOLOGY RESEARCH REPORT Crescent Lake National Wildlife Refuge 31 May to 4 July 2017 A report submitted to Refuge Biologist Marlin French 15 July 2017 John B Iverson Dept.
Photo by Drew Feldkirchner, WDNR
 Photo by Drew Feldkirchner, WDNR Wood Turtle in Wisconsin State listed Threatened Species Species of Greatest Conservation Need Species Description Medium sized (5 9.5 inches long) Carapace dark gray to
Photo by Drew Feldkirchner, WDNR Wood Turtle in Wisconsin State listed Threatened Species Species of Greatest Conservation Need Species Description Medium sized (5 9.5 inches long) Carapace dark gray to
This Coloring Book has been adapted for the Wildlife of the Table Rocks
 This Coloring Book has been adapted for the Wildlife of the Table Rocks All images and some writing belong to: Additional writing by: The Table Rocks Environmental Education Program I became the national
This Coloring Book has been adapted for the Wildlife of the Table Rocks All images and some writing belong to: Additional writing by: The Table Rocks Environmental Education Program I became the national
Investigations of Giant Garter Snakes in The Natomas Basin: 2002 Field Season
 Investigations of Giant Garter Snakes in The Natomas Basin: 2002 Field Season Investigations of Giant Garter Snakes in The Natomas Basin: 2002 Field Season By Glenn D. Wylie and Lisa L. Martin U.S. GEOLOGICAL
Investigations of Giant Garter Snakes in The Natomas Basin: 2002 Field Season Investigations of Giant Garter Snakes in The Natomas Basin: 2002 Field Season By Glenn D. Wylie and Lisa L. Martin U.S. GEOLOGICAL
BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS
 Nov., 1965 505 BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS Lack ( 1954; 40-41) has pointed out that in species of birds which have asynchronous hatching, brood size may be adjusted
Nov., 1965 505 BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS Lack ( 1954; 40-41) has pointed out that in species of birds which have asynchronous hatching, brood size may be adjusted
Bald Head Island Conservancy 2018 Sea Turtle Report Emily Goetz, Coastal Scientist
 Bald Head Island Conservancy 2018 Sea Turtle Report Emily Goetz, Coastal Scientist Program Overview The Bald Head Island Conservancy s (BHIC) Sea Turtle Protection Program (STPP) began in 1983 with the
Bald Head Island Conservancy 2018 Sea Turtle Report Emily Goetz, Coastal Scientist Program Overview The Bald Head Island Conservancy s (BHIC) Sea Turtle Protection Program (STPP) began in 1983 with the
Dominance/Suppression Competitive Relationships in Loblolly Pine (Pinus taeda L.) Plantations
 Plantations") Dominance/Suppression Competitive Relationships in Loblolly Pine (Pinus taeda L.) Plantations by Michael E. Dyer Dissertation submitted to the Faculty of the Virginia Polytechnic Institute and Stand University
Dominance/Suppression Competitive Relationships in Loblolly Pine (Pinus taeda L.) Plantations by Michael E. Dyer Dissertation submitted to the Faculty of the Virginia Polytechnic Institute and Stand University
Amphibians&Reptiles. MISSION READINESS While Protecting NAVY EARTH DAY POSTER. DoD PARC Program Sustains
 DoD PARC Program Sustains MISSION READINESS While Protecting Amphibians&Reptiles Program Promotes Species & Habitat Management & Conservation Navy s Environmental Restoration Program Boasts Successful
DoD PARC Program Sustains MISSION READINESS While Protecting Amphibians&Reptiles Program Promotes Species & Habitat Management & Conservation Navy s Environmental Restoration Program Boasts Successful
The Greater Sage-grouse: Life History, Distribution, Status and Conservation in Nevada. Governor s Stakeholder Update Meeting January 18 th, 2012
 The Greater Sage-grouse: Life History, Distribution, Status and Conservation in Nevada Governor s Stakeholder Update Meeting January 18 th, 2012 The Bird Largest grouse in North America and are dimorphic
The Greater Sage-grouse: Life History, Distribution, Status and Conservation in Nevada Governor s Stakeholder Update Meeting January 18 th, 2012 The Bird Largest grouse in North America and are dimorphic
SEASONAL CHANGES IN A POPULATION OF DESERT HARVESTMEN, TRACHYRHINUS MARMORATUS (ARACHNIDA: OPILIONES), FROM WESTERN TEXAS
, FROM WESTERN TEXAS") Reprinted from PSYCHE, Vol 99, No. 23, 1992 SEASONAL CHANGES IN A POPULATION OF DESERT HARVESTMEN, TRACHYRHINUS MARMORATUS (ARACHNIDA: OPILIONES), FROM WESTERN TEXAS BY WILLIAM P. MACKAY l, CHE'REE AND
Reprinted from PSYCHE, Vol 99, No. 23, 1992 SEASONAL CHANGES IN A POPULATION OF DESERT HARVESTMEN, TRACHYRHINUS MARMORATUS (ARACHNIDA: OPILIONES), FROM WESTERN TEXAS BY WILLIAM P. MACKAY l, CHE'REE AND
Scaled Quail (Callipepla squamata)
") Scaled Quail (Callipepla squamata) NMPIF level: Species Conservation Concern, Level 2 (SC2) NMPIF assessment score: 15 NM stewardship responsibility: Moderate National PIF status: Watch List, Stewardship
Scaled Quail (Callipepla squamata) NMPIF level: Species Conservation Concern, Level 2 (SC2) NMPIF assessment score: 15 NM stewardship responsibility: Moderate National PIF status: Watch List, Stewardship
Bio4009 : Projet de recherche/research project
 Bio4009 : Projet de recherche/research project Is emergence after hibernation of the black ratsnake (Elaphe obsoleta) triggered by a thermal gradient reversal? By Isabelle Ceillier 4522350 Supervisor :
Bio4009 : Projet de recherche/research project Is emergence after hibernation of the black ratsnake (Elaphe obsoleta) triggered by a thermal gradient reversal? By Isabelle Ceillier 4522350 Supervisor :
4 Many species of mammals, birds, reptiles, amphibians and fish 940L. Source 1 Habitats
 Source 1 Habitats 1 American Alligators can be found in fresh water environments like rivers, lakes, ponds, swamps and marshes. They also like to live in areas that are brackish, which means the water
Source 1 Habitats 1 American Alligators can be found in fresh water environments like rivers, lakes, ponds, swamps and marshes. They also like to live in areas that are brackish, which means the water
Writing: Lesson 31. Today the students will be learning how to write more advanced middle paragraphs using a variety of elaborative techniques.
 Top Score Writing Grade 4 Lesson 31 Writing: Lesson 31 Today the students will be learning how to write more advanced middle paragraphs using a variety of elaborative techniques. The following passages
Top Score Writing Grade 4 Lesson 31 Writing: Lesson 31 Today the students will be learning how to write more advanced middle paragraphs using a variety of elaborative techniques. The following passages
JoJoKeKe s Herpetology Exam
 ~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~~*~*~*~*~*~*~*~*~*~*~*~*~*~*~ JoJoKeKe s Herpetology Exam (SSSS) 2:30 to be given at each station- B/C Station 1: 1.) What is the family & genus of the shown
~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~*~~*~*~*~*~*~*~*~*~*~*~*~*~*~*~ JoJoKeKe s Herpetology Exam (SSSS) 2:30 to be given at each station- B/C Station 1: 1.) What is the family & genus of the shown
TURTLE PATROL VOLUNTEER REFERENCE GUIDE
 TURTLE PATROL VOLUNTEER REFERENCE GUIDE Intro to Loggerhead turtles and the Sunset Beach Turtle Watch Program This program is a private and non-profit program using volunteers to monitor the nesting of
TURTLE PATROL VOLUNTEER REFERENCE GUIDE Intro to Loggerhead turtles and the Sunset Beach Turtle Watch Program This program is a private and non-profit program using volunteers to monitor the nesting of
Lygosoma laterale. Breeding Cycle in the Ground Skink, HARVARD HENRY S. Museum of Natural History DEC S. University of Kansas Lawrence
 - i\jri - J- M^vcij mus. co i\..-. : LIBRARY University of Kansas Publications DEC S Museum of Natural History HARVARD Volume 15, No. 11, pp. 565-575, 3 figs. May 17, 1965 Breeding Cycle in the Ground
- i\jri - J- M^vcij mus. co i\..-. : LIBRARY University of Kansas Publications DEC S Museum of Natural History HARVARD Volume 15, No. 11, pp. 565-575, 3 figs. May 17, 1965 Breeding Cycle in the Ground
Using a Spatially Explicit Crocodile Population Model to Predict Potential Impacts of Sea Level Rise and Everglades Restoration Alternatives
 Using a Spatially Explicit Crocodile Population Model to Predict Potential Impacts of Sea Level Rise and Everglades Restoration Alternatives Tim Green, Daniel Slone, Michael Cherkiss, Frank Mazzotti, Eric
Using a Spatially Explicit Crocodile Population Model to Predict Potential Impacts of Sea Level Rise and Everglades Restoration Alternatives Tim Green, Daniel Slone, Michael Cherkiss, Frank Mazzotti, Eric
Effects of Cage Stocking Density on Feeding Behaviors of Group-Housed Laying Hens
 AS 651 ASL R2018 2005 Effects of Cage Stocking Density on Feeding Behaviors of Group-Housed Laying Hens R. N. Cook Iowa State University Hongwei Xin Iowa State University, hxin@iastate.edu Recommended
AS 651 ASL R2018 2005 Effects of Cage Stocking Density on Feeding Behaviors of Group-Housed Laying Hens R. N. Cook Iowa State University Hongwei Xin Iowa State University, hxin@iastate.edu Recommended
Reptiles and amphibian behaviour
 Reptiles and amphibian behaviour Understanding how a healthy reptile and amphibian should look and act takes a lot of observation and practice. Reptiles and amphibians have behaviour that relates to them
Reptiles and amphibian behaviour Understanding how a healthy reptile and amphibian should look and act takes a lot of observation and practice. Reptiles and amphibians have behaviour that relates to them
RED-EARED SLIDER TURTLES AND THREATENED NATIVE RED-BELLIED TURTLES IN THE UPPER DELAWARE ESTUARY. Steven H. Pearson and Harold W.
 RESOURCE OVERLAP AND POTENTIAL COMPETITION BETWEEN INVASIVE RED-EARED SLIDER TURTLES AND THREATENED NATIVE RED-BELLIED TURTLES IN THE UPPER DELAWARE ESTUARY Steven H. Pearson and Harold W. Avery Six Most
RESOURCE OVERLAP AND POTENTIAL COMPETITION BETWEEN INVASIVE RED-EARED SLIDER TURTLES AND THREATENED NATIVE RED-BELLIED TURTLES IN THE UPPER DELAWARE ESTUARY Steven H. Pearson and Harold W. Avery Six Most
Lacerta vivipara Jacquin
 Oecologia (Berl.) 19, 165--170 (1975) 9 by Springer-Verlag 1975 Clutch Size and Reproductive Effort in the Lizard Lacerta vivipara Jacquin R. A. Avery Department of Zoology, The University, Bristol Received
Oecologia (Berl.) 19, 165--170 (1975) 9 by Springer-Verlag 1975 Clutch Size and Reproductive Effort in the Lizard Lacerta vivipara Jacquin R. A. Avery Department of Zoology, The University, Bristol Received
NOTES ON THE ECOLOGY AND NATURAL HISTORY OF CTENOPHORUS CAUDICINCTUS (AGAMIDAE) IN WESTERN AUSTRALIA
 IN WESTERN AUSTRALIA") NOTES ON THE ECOLOGY AND NATURAL HISTORY OF CTENOPHORUS CAUDICINCTUS (AGAMIDAE) IN WESTERN AUSTRALIA By ERIC R. PIANKA Integrative Biology University of Texas at Austin Austin, Texas 78712 USA Email: erp@austin.utexas.edu
NOTES ON THE ECOLOGY AND NATURAL HISTORY OF CTENOPHORUS CAUDICINCTUS (AGAMIDAE) IN WESTERN AUSTRALIA By ERIC R. PIANKA Integrative Biology University of Texas at Austin Austin, Texas 78712 USA Email: erp@austin.utexas.edu
Twenty years of GuSG conservation efforts on Piñon Mesa: 1995 to Daniel J. Neubaum Wildlife Conservation Biologist Colorado Parks and Wildlife
 Twenty years of GuSG conservation efforts on Piñon Mesa: 1995 to 2015 Daniel J. Neubaum Wildlife Conservation Biologist Colorado Parks and Wildlife Early Efforts 1995 - Woods and Braun complete first study
Twenty years of GuSG conservation efforts on Piñon Mesa: 1995 to 2015 Daniel J. Neubaum Wildlife Conservation Biologist Colorado Parks and Wildlife Early Efforts 1995 - Woods and Braun complete first study
May Dear Blunt-nosed Leopard Lizard Surveyor,
 May 2004 Dear Blunt-nosed Leopard Lizard Surveyor, Attached is the revised survey methodology for the blunt-nosed leopard lizard (Gambelia sila). The protocol was developed by the San Joaquin Valley Southern
May 2004 Dear Blunt-nosed Leopard Lizard Surveyor, Attached is the revised survey methodology for the blunt-nosed leopard lizard (Gambelia sila). The protocol was developed by the San Joaquin Valley Southern