ANDRZEJ ELZANOWSKI. (plate 3)
|
|
|
- Janice Willis
- 5 years ago
- Views:
Transcription
1 ANDRZEJ ELZANOWSKI SKULLS OF GOBIPTER YX (AVES) FROM THE UPPER CRETACEOUS OF MONGOLIA (plate 3) Abstract. - The discovery of a second skull of Gobipteryx minuta (ElZANOWSKI 1974) establishes this to be a palaeo. gnathous bird, showing the typical pattern of the palate as well as the features directly related to ratite rhynchokinesis Some palatal characters 01 Gobipteryx are closely similar to those of the cassowary. Gobipteryx skulls are described and new partial reconstructions are given. Previously erected taxons are rediagnosed. INTRODUCTION After the description of the first Gobipteryx skull had been published (ELzANowsKI 1974), a second skull was found among other small fossils collected by the 1971 Polish-Mongolian Palaeontological Expedition to the Gobi Desert (KIELAN-JAWOROWSKA & BARSBOLD 1972). Both Gobipteryx specimens were found in the same locality (Khulsan), in beds of the Barun Goyot Formation, recognized by KrELAN-JAWOROWSKA (1974) as? Middle Campanian. Both specimens are housed in the In stitute of Paleobiology of the Polish Academy of Sciences in Warsaw, abbreviated as ZPAL. Although examination of the new skull has fully confirmed the palaeognathine affinities of Gobipteryx, there are many misinterpretations in the author's preliminary description that are corrected in the present paper. Gobipteryx is the oldest form which can be assigned to the Palaeognathae, other earliest records (Opisthodactylus patagonicus Ameghino, Eleutherornis helveticus SCHAUB and Eremopezus eocaenus ANDREWS) being from Eocene (FISHER 1967). The only ratite fossils hitherto known from Asia are Neogene and Quaternary struthious remains (BURCHAK-ABRAMOVICH 1962) and the Miocene and, possibly, Eocene egg-shells of a separate "aepyornithid type" (SAUER 1972). ACKNOWLEDGEMENTS The comments and corrections done by Dr. P. V. RICH (Texas Tech University) and Prof. J. H. OSTROM (Yale University) proved to be of greatest importance and I am extremely grateful for them. I wish to acknowledge my debt to Dr. Z. BocHENsKI (Institute of Systematic and Experimental Zoology, Polish Academy of Sciences, Cracow), Dr. O. L. Rosscusro (Zoological Museum, Moscow University, USSR) and Dr. B. STEPHAN (Museum of Natural History, Humboldt University, Berlin, GDR) who lent me specimens for comparative study. I especially want to thank Dr. A. SULIMSKI (Institute of Paleobiology, Polish Academy of Sciences, Warsaw) who made the drawings. My thanks are also due to Mrs. J. SKARZYNSKA of the same Institute, for the expert preparation of the second Gobipteryx skull.
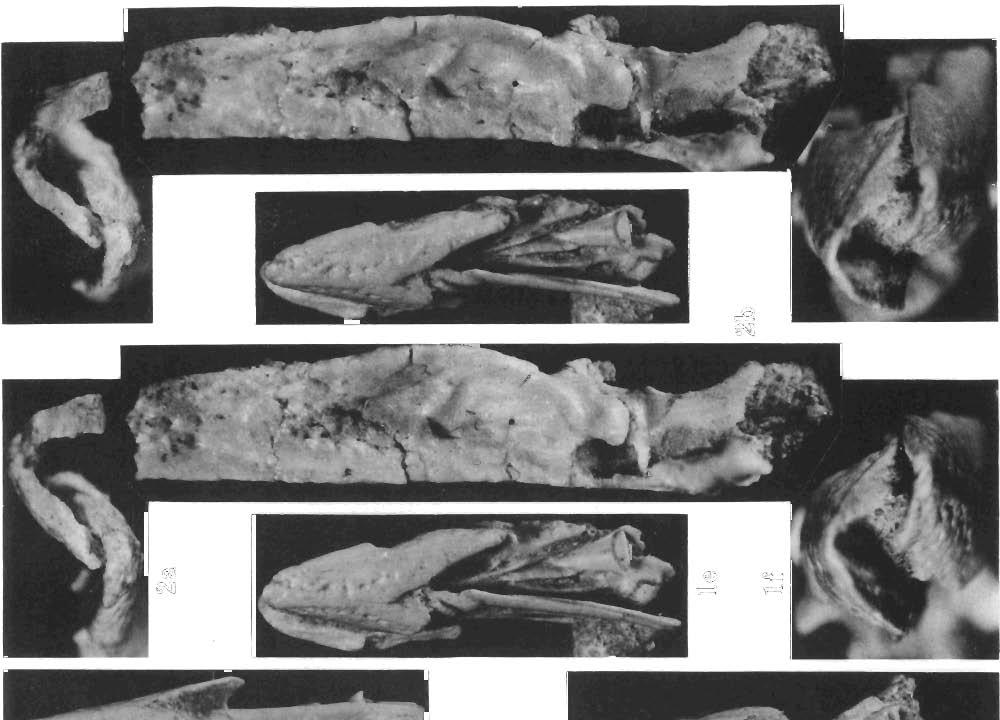
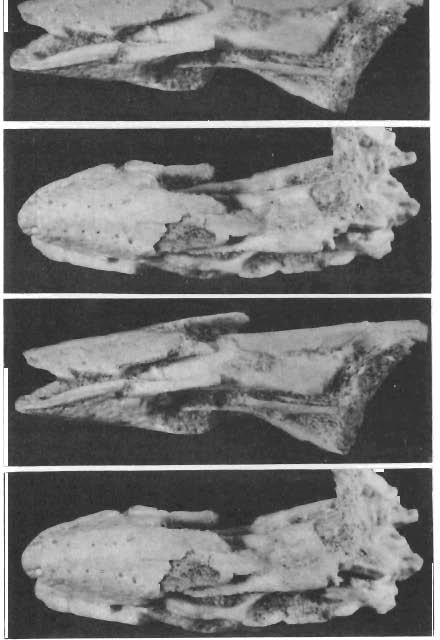
2 154 ANDRZEJ ELZANOWSKI DESCRIPTIONS Order Gobipterygiformes, ELZANOWSKI 1974 Revised diagnosis. - Mandibular articulation of the quadrate bipartite. Skull of palaeognathous type, approaching the cassowary conditions: (a) vomers fused far anteriorly, gradually diverging backw ards to articulate suturally with pterygoids, (b) pterygoids widens posteriorly, (c) palatine supported mostly by the pterygoid, (d) ascending process ofthe maxilla present. Basal part of the palatine enclosed in the lateral sinus of the pterygoid. Family Gobipterygidae, ELZANOWSKI 1974 Genus Gobipteryx, ELZANOWSKI 1974 Gobipteryx minuta, ELZANOWSKI 1974 (pi. 37, text-fig. 1) 1974 Gobipteryx minuta n.g., n.sp.; A. ELZANOWSKI, pp , pls Material. - Two specimens, both from Khul san, Gobi Desert, Mongolia, Baron Goyot Formation: the holotype ZPAL MgR 1/12 and the second skull (ZPAL MgR-I/32), with the calvarium and the mandible, in the second specimen preserved in the anterior two thirds only and somewhat compressed laterally. Revised diagnosis. - Culmen straight near its rostral end. No distinct grooves on the outer surface of the terminal part of the beak. Lateral contacting margins of the premaxilla and dentary flat in the terminal part of the beak (but behind the very tip) and well delimited from the middle depression. Mandible in postarticular reg'on smooth vent rally (with no cristae). Upper mandibular margin well elevated above the level of the external mandibular process. Dimensions. - See table I. Holotype skull (ZPAL MgR-I/1 2). See ELZANOWSKI (1974). Two pairs of rows of nutrient foramina -two medial and two iateral --can be distinguished on the dorsal side ofthe premaxilla, although the bone surface is much weathered and not all the foramina preserved. The maxillary processes of the prem axilla begin some 2 mm anterior to the rostral extremity of the nasal opening. Despite previous interpretation, the transverse slit across the premaxilla appears now to be a simple fracture. In ventral view, many details of the palate have to be reinterpreted. Anteriorly, on the right side of the skull, a poor vestige of the bony ridge, converging anteromesiad, which was described as the vomer, appears to be a remnant ofthe medial maxillopalatine margin. Its spurious symmetrical left counterpart seems to be a true vestige of the left vomer, lying medial to the left maxillo-palatine. Medial to the right maxillopalatine there is an obscure elevation probably representing the right vomer. The left pterygoid, or rather its out er edge, is represented by a tongue-shaped fragment extending caud ally from the left vomer vestige. Th e piece was formerly described as a part ofthe palatine. Posteriorly, the external pterygoid margin curves laterad and then forward forming the lateral sinus. Here the pterygoid becomes much wider. The right pterygoid begins as an elevation covered by a bony lamina, nearly at the same level as the left counterpart. The lateral sinus surrounding the palatine base is very obscure and immediately posteriorly there is a secondarily refilled break which supposedly represents the longitudinal overlap with the pterygoid bar. It follows that the prequadrate part of the pterygoid was probably longer than preserved. On the posterior part of the right pterygoid, behind the break, two articular sockets are exposed, the outer for the quadrate, the inner for the basipterygoid process. Just behind, or even across the sockets, the posteriormost part of the pterygoid is broken off. Some pieces of this seem to be crushed against the orbital process of the quadrate. A small terminal fragment of some
3 SKULLS OF GOBIPTERYX 155 longer bone was obviously dislocated above the left pterygoid. It was previously supposed to be a part of the quadratojugal, certainly false. The piece is boot-shaped and consists of two parts that meet at right angle. In the corner, the edges of both perpendicular parts are flat and so their contact is precisely rectangular. The elongated "sole" is regularly rounded on the "underside" and suggestive of some articular connection. A minute, circular excavation is present on the very tip. On one side of the other part, directly broken off from the whole, there is a (pneumatic?) foramen and a slight ridge descending to the sole. The opposite side is not preserved. The fragment is supposed to be the posterior end of the pterygoid, the sole possibly abuted the quadrate medially, well above the internal condyle. The pneumatic foramen near the comparable ridge is present in the cassowary. Although hardly identifiable, the right palatine seems to be preserved. Its lateral, thickened edge can be observed prolonging the outer pterygoid margin rostrally. The anterior part of the left palatine, fused with the maxillopalatine, may also be present. On the left side, the palatine can be clearly seen or.ly near to anterior fusion with the maxillopalatine, where the vertically oriented bar passes anteriorly into the horizontal blade of the maxillopalatine. The disarticulated quadrate is preserved in a quite artificial position, its mandibular articular surface facing forwards and the otic process projecting ventrally. Only the basal part of the orbital process is fairly distinct and can be seen to descend from the otic process up to a point a little above the internal condyle. The remaining part of the orbital process is crushed against the braincase. The process was formerly called "the pterygoid process of the quadrate" (see discussion). Between the ventral base of the orbital process and the inner mandibular condyle of the quadrate, there is a minute tubercle, obviously for articulation with the pterygoid. Unfortunately, further details of the pterygoid-quadrate articulation can not be reconstructed with sufficient reliability, since both the posterior end of the pterygoid and the orbital process are incomplete. The mandibular articulation of the quadrate, as described before, is bicondylar. The lateral edge of the quadrate, in particular just above the outer condyle, is somewhat damaged, and the quadratojugal articulation is gone. In dorsal view of the right mandible, two articular surfaces can be recognized. The outer surface, roughly circular (2.5 mm in diameter) extends laterally on the external process of the mandible. Anteromedially there is a small, somewhat excavated area where the posterior adductor commonly inserts. Posteromedial to the outer surface there is a large pit, whose anterolateral wall, the only preserved part of the inner articular surface, evidently articulated with the inner quadrate condyle. The latter slanted some 2 mm below the outer surface of the mandible. Just behind the external mandibular process there is a small tubercle, behind which the upper edge of the mandible curves gently dcwnwards and then, still further backwards, rises in the elevated upper edge of the retroarticular proce ss. In external view of the right mandible, some 12 mm from the rostral tip, the dentary bifurcates into the dorsolateral and ventrolateral process es with the longitudinal groove in between, extending about 8 mm posteriorly. No definable suture between the dorsolateral process of the dentary and the supraangular can be recogniz ed. Beginning from the pcsterior end of the longitudinal groove, the upper margin of the ventrolateral process of the dentary curves downwards, and the process ends as a narrow, acute tongue, wedged between the supraangular and the angular. The supraangular, forming the whole coronoid elevation, is convex below and seems to continue in the groove between the dentary processes. Posteriorly, the bone extends the whole length of the retroarticular process forming the upper half of its height at the caudal end. The lower half is formed by the angular, which tapers gradually anteriorly and disappears to be replaced by the dentary on the lower edge of the mandible. The whole postdentary region of the left side is gone and no new details were noticed on the preserved part except that at the point ofdivergence of the lateral dentary processes there is a particularly well marked depression corresponding to the anterior mandibular fenestra. In internal view of the mandible, the splenials seem to be absent (see discussion) and the



4 156 ANDRZEJ ELZANOWSKI SA la D 3 AN EXT > < Imm FP lcm MXP PH
5 SKULLS OF GOBIBTERYX 157 Table 1 Comparison of some cranial dimensions of two Gobipteryx specimens values are of fragments (in mm). Paranthetic I First specimen Second specimen ZPAL MgR-T/I2 I ZPAL MgR-1/32 Skull lengt h est. 45 (27+) Length of the rostral part of the lower edge of premaxilla, to the level of the anteriormost margin of the nasal opening 8 8 Width of premaxilla at the base of the maxillary processes 7 7 Premaxillary height at the base of the maxillary processes 4 4 Vomer length (in ventral view): laterally - 9 medially - 7 Pterygoid length (in ventral view) est. 12 (7+6) Mandible length 38 (29+) Mandibular symphysis length 9 9 Mandible height at the level of the anterior foramen 4 4 anterior, ascending part of the angular is exposed. On the only complete right mandible, the sutures are secondarily obliterated and their courses very difficult to establish. The coronoid elevation, inflected inwards, is composed mostly of the supraangular which also closes the posterior fossa. The diagonal ridge dividing the fossa is met by a small pseudotemporal process (MULLER, 1963)projecting upwards and a little backwards from a rod of bone extending from the articular region. This rod of bone appears to be the ossified part of the MECKEL'S cartilage or Knochenstab (MULLER, 1963). Only the upper edge of the Knochenstab is exposed for it is covered below by the narrow prearticular lamina running forwards and upwards from the articular. Below the prearticular, the angular appears in the medial side of the mandible and forms its lower edge up to the level of the anterior end of the fossa, where the angular is replaced by the dentary. Then the angular ascends and tapers gradually reaching the dorsomedial process of the dentary above. In the anteriormost region of the right mandible there is a loose fragment of the bony lamina which may be a remnant of the splenial. In medial view of the only well preserved anterior part of the left mandible, the ascending part of the angular is very distinct. A break occurs at the point where the angular has just left the lower margin of the mandible. Some very poor fragments of the posterior part of the left mandible were dislocated forwards and attached to the outer side of the anterior part. Only the pseudotemporal process could be distinguished clearly among these pieces Fig. 1 Gobipteryx minuta ELZANOWSKI 1974: 1. Partial reconstructions of the mand ible, a - in lateral view, b - in medial view, c - in dorsal view. 2. Skull ofthe type specimen (ZPAL MgR-I/I2) in ventral view. 3. Fragment ofthe type specimen, unidentified, possibly posteriormost ending of the pterygoid. 4. Partial reconstruction of the quadrate in medial view. 5. Skull of the second specimen (ZPAL MgR-I/32), a - in dorsal view, b - in ventral view, C - in left lateral view. Abbreviations: AN - angular, AR - articular, CH - internal naris, CV - cervical vertebra, D - dentary, EXTexternal process of the mandible, FP - pneumatic foramen, INT - intern al process of the mandible, KS - Knochenstab, LPT- left pterygoid, LV - left vomer, MXA - ascending process of the maxillopalatine, M XP - maxillopalatine, NPM - premaxillary process of the nasal, N - nasal, PAL - palatine, PL - pleurapophysis of the cervical vertebra, PAR - prearticular, PST- pseudotemporal process, PMM - maxillary process of the premaxilla, PMN - nasal process of the premaxilla, PT- pterygoid, PTB - basipterygoid articulation of the pterygoid, PTQ - quadrate articu lation of the pterygoid, PIT- outer tine of the pterygoid, RPT- right pterygoid, RV - right vomer, QOR - orbital process of the quadrate, QOT - otic process of the quadrate, RET- retroarticular process of the mandible, SA - surangular.
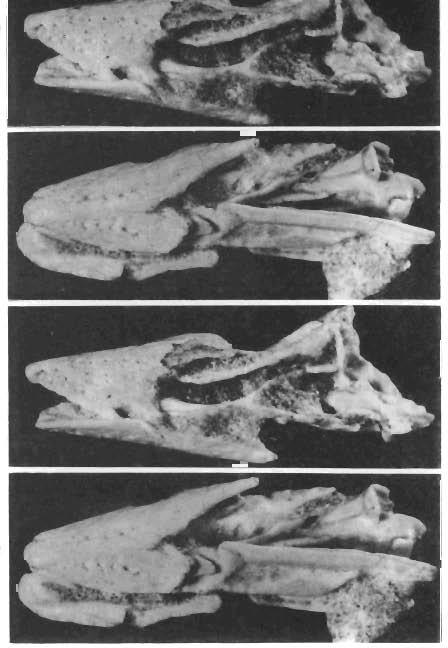
6 158 ANDRZEJ ELZANOWSKI Second skull (ZPAL MgR-Ij32). Only the rostral part ofthe skull is preserved. In the region of the anterior orbital r'm, the bones are broken and partially dislocated, the fracture crossing the nasals and pterygoids. The skull is compressed laterally to some extent. The premaxilla is wide, its rostral tip is rounded and the lateral edges converge, their prolongations meet at the relatively great angle of about 40. This all suggests a short, robust beak. Dorsally, the premaxilla is strongly domed with many nutrient foramina, the majority of them arranged in four rows: two medial and two lateral, the last running just above the premaxillary edges. There are about 7 foramina in each medial and about 5 in each lateral row. At the rostral tip of the premaxilla, the medial rows begin with the vestigial grooves. Nasal openings extend far anteriorly. Below and, unexpectedly, a little (approximately 2 mm) forwards, the maxillary processes of the premaxilla originate as separate projections. The broken anterior part of the maxillary process is preserved only on the right side. The nasal processes, strongly tapering posteriorly, extend 22 mm from the rostral end of the premaxilla, and fit between the premaxillary processes of the nasals. The last extend to the anterior rim of the nasal opening and are overlapped, as the thin laminae, by the nasal processes of the premaxillae. On the better preserved, left side of the specimen, the corresponding nasal and premaxillary processes are disarticulated, the nasal process ofthe premaxilla having been lifted off the premaxillary process of the nasal. Such a condition suggests a loose and movable connection between these bones. Beneath the right nasal, part of the jugal bar is preserved. It appears to consist mostly of the caudal prolongationofthe maxillary process ofthe premaxilla. The lateral edge ofthe maxilla is represented by only a small medial fragment, other maxillary parts were identified are the maxillopalatines (see below). Neither the jugal nor the quadratojugal could be identified. The middle part of the palate, with vomers, palatines and pterygoids arranged in typical palaeognathous pattern, is fairly well preserved in ventral view, with some dislocation resulting in slight asymmetry in the hind part of the specimen. The vomers are fused anteriorly and begin mm from the premaxillary rostral end. About 4 mm posteriorly they diverge (at 20 to the lateral edges) and run 5 mm further caudally to be suturally overlapped by the pterygoids. The separate parts of the vomers are inclined in such a way that the lateral edges are more ventral than the medial edges. The vomeropterygoid suture is oblique (about 25 to the sagittal plane) and 3 mm long, while the vomeropterygoid bar is little more than 1 mm wide at this level. From the vomeropterygoid suture the pterygoids widen gradually toward the palatinal suture posteriorly. The pterygoid laminae are inclined in accordance with the vomers position. In the palatine region the right pterygoid becomes horizontal and, just behind this, an outer wing projects laterally. The lateral pterygoid margin curves outwards and then forwards to project as a terminal outer tine and thus encloses the posterior edge of the palatine in the rostrally open lateral sinus. The outer tine of the pterygoid projects slightly beyond the adjacent external palatine margin. The pterygoid achieves the greatest width of nearly 3.5 mm at the level of the outer tine, where the medial pterygoid margin curves gently posterolaterated. The right pterygoid is broken just behind the palatine base and the remaining part is gone. The left pterygoid is dislocated to the midline so that the shape and size of the interpterygoid vacuity cannot be established directly. The bone is broken at a more anterior position than its right counterpart; only a part of the lateral sinus remains. The remaining, posterior part of the left pterygoid is dislocated on the dorsal side so that its posterior end, appearing to be boot-shaped, touches the left nasal. The right palatine is based in the pterygoid sinus posteriorly. Medially, the contact with the pterygoid extends to the vomeropterygoid suture anteriorly. The lateral margin of the palatine is much thicker than the inner lamina and projects somewhat ventrally as a ridge which continues forwards as the lateral rim of the internal naris. The indistinct maxillopalatine-palatine suture seems to cross the choanal rim at the level of the vomeropterygoid suture. Anteriorlythe ridge disappears and the medial margin of the right maxillopalatine curves to the vomer enclosing the internal naris anteriorly. A small part of the
7 SKULLS OF GOBIPTERYX 159 left maxillopalatine is visible in dorsal view. Posteriorly it sends out the ascending or nasal process with the free end projecting dorsoposteriorly and partially separating the nasal and the ant orbital openings. In ventral view, only the posterior part of the left maxillopalatine is visible as a shelf fused medially with the palatine without any definable suture and facing somewhat outwards, as in the cassowary. The mandible has a long smooth symphysis. The rounded outline ofthe rostral tip is exactly the same as that of the premaxilla. In dorsal view the rostral parts of the lateral edges are also equally wide but quite flat with no concavity. In ventral view there is a very similar pattern of nutrient foramina as occurs on the upper jaw: a pair of medial rows beginning with the slight grooves on the very tip, and a pair of lateral rows. There are 9-10 foramina in each medial and 6 or more in each lateral row. Posteriorly, the mandible edges are damaged, so that some lateral foramina are probably gone. In the right mandible, the whole lower part of the dentary is present. The upper part is broken across the dorsolateral process. The whole postdentary region of the mandible is missing. The preserved part of the right mandible is badly fractured, the rostral piece being overlaid by the posterior one - so, exact length measurements are not possible. From the external side, the posterior bifurcation ofthe dentary into upper and lower processes is fairly distinct. The posterior part of the dorsolateral process is absent. The ventral process tapers backwards, its upper margin descending gradually, to end as a narrow tongue. Behind this the lower edge of the mandible is formed by the angular. The dorsolateral process of the dentary forms a part of the dorsal edge of the mandible, the middle part of which is composed of the surangular. What I interpret to be also the surangular closes the space between the lateral dentary processes and, in this way, forms a bottom of the 8 mm long external depression, occurring at mid-height of the mandible. Behind this depression, the surangular becomes convex, just in front of the broken end of the mandible. In internal view the anterior part of the angular rises gradually exposing the ventral process of the dentary beneath. The splenial seems to be absent on the specimen. In the left mandible, only the anterior part of the dentary is preserved and no additional details were noticed. Remarks. - An uncertain point on the reconstructed ventral view (text-fig. 2, 1a) is the pterygoid-quadrate articulation, which is complex and quite variable in palaeognathous birds. A posterodorsal process of the pterygoid occurs adjacent to the medial side of the quadrate. in Ratitae (Rhea, Struthio, Casuarius, Dromaius - pers.. obs.) and perhaps in Gobipteryx as the puzzling, boot-shaped fragment, that I suppose to be a caudal end of the pterygoid. The lack of splenials on both specimens appears to be natural, for in all Ratitae the splenial is quite loosely connected with other bones. In view of the poor preservation of both skulls, it is highly probable that the splenials became disarticulated. In the holotype skull, the orbital process, previously called "the pterygoid process of the quadrate", projects into the articular socket of the pterygoid. However, this position appears to be accidental and, thus, the commonly used term "the orbital process" is introduced here, although both names are sometimes used as synonyms. Although the jaws of the two specimens are almost the only parts that can be compared, their detailed similarity together with identical measurements (table I) indicates both Gobipteryx specimens to be conspecific. Striking similarity is conveyed by the terminal parts of the jaws, with their flat margins, slightly concave on the premaxilla, converging at the same angle of about 40. The rostral part of the culmen is rectilinear, ascending from the tip at the angle of 32 to the rostrolateral premaxillary edges (measured in the sagittal plane). A possible difference between the specimens is the number of nutrient foramina which seem to be fewer on the premaxilla of the first skull, although the bone surface is weathered, and smaller foramina could have been destroyed. The symphysial surface of the holotype mandible is completely ground off, and no relevant details are observable. Differences in the kind of food are commonly reflected in the bill structure, especially in ground-feeding birds (CODY, 1974: 35-39, and references the rein). Therefore, the high degree
8 160 ANDRZEJ ELZANOWSKI of similarity of the bill shape and dimensions of both Gobipteryx specimens would suggest closely similar food requirements. As far as the Gause's principle is still valid (CODY, 1974, p. 54), a persistent coexistence of two species with common food resources is less probable. DISCUSSION Gobipteryx provides new evidence in favour of the classic hypothesis of PYCRFAT (1900), who argued for the morphological and phylogenetic unity of palaeognathous birds. PYCRAFT'S views have been challenged many times and therefore require some comments in the light of contemporary knowledge. The main criticism was furnished by McDoWELL (1948), who claimed the impossibility of defining the palaeognathous palate. The princip al characters used in the definition (the vomeropterygoid and the palatinopterygoid sutures as well as the lack of pterygoparasphenoidal contact) were shown not to be universal. In Struthio the small vomer does not contact the pterygoid, the latter touching the parasphenoid; the occurrence of the pterygoparasphenoidal contact is also mentioned for Apteryx, although it is not indicated in the description of the "apterygiform type"; lastly, the lack of contact between the palatine and the pterygoid in Dromaius is emphasized. Moreover, McDoWELL argued that many features thought to specify the palaeognathous condition occur in various neognathine groups, for example the backward position of basipterygoid processes "can be matched by the Musophagidae and Turnicidae among the Neognathae". Meanwhile, a great deal of evidence (BOCK, 1963) in favour of monophyly of the palaeognathous birds accumulated since the time of publication of the McDoWELL'S paper. Even if such a rigorous requirement of universality for every morphological criterion is justified, the factual basis of the McDoWELL'S argument was shown to be dubious. In the case of struthious palate, vomers are extremely variable in shape and size and reach the pterygoid in some specimens (WEBB, 1957). In the kiwi, the pterygoparasphenoidal contact does not exist on the specimen of Apteryx mantelli examined by the present author. In most specimens of the emu, the palatine does articulate with the pterygoid (SIMONETTA, 1960). PARKES and CLARK (1966) noticed anew feature linking the palaeognathine groups, which is a specific pattern of division of the rhamphotheca, distinct from that occurring in some other birds with a complex horn-sheath of the beak. Among other kinds of research, MEISE (1963) compared behavioral features and came to the conclusion that the Ratitae on one hand and tinamous on the other are sister groups. The arguments for palaeognathine monophyly have been recently summarized by CRACRAFT (1974). The most convincing argument, especially in the present context, for the unity of the palaeognathous condition was given by BOCK (1963). If the whole complex of features, and not the single ones taken independently, is included in the definition, the palaeognathous pattern can be precisely defined even if one or another of these characters happens to occur in some neognathine group. Following (with some modifications) BOCK'S (1963) important paper, the palaeognathous complex includes: (1) Relatively large vomer articulating with the premaxillae and maxillopalatines anteriorly and (Struthio excepted in most cases) with the pterygoids posteriorly; (2) Palatine articulating suturally with the vomeropterygoid bar, never with the parasphenoid (this criterion is universal for palaeognathous birds); (3) Large basipterygoid processes articulating with the pterygoid close to the quadrate articulation; (4) Complex articulation between the pterygoid and quadrate, with a well developed articular surface on the orbital process and/or the posterodorsal process of the pterygoid ascending along and adhering medially to the base of the orbital process. This complex ofcharacters is not simply an arbitrary assemblage chosen from many other possibilities. It is substantiated by the biomechanical interpretations (HOFER, 1945) of the palaeognathous skulls, further elaborated by BOCK (1963). HOFER showed, that the primary importance of the parasphenoid contact of the palatopterygoid
9 SKULLS OF GOBIPTER YX ]61 joint is for transmission of longitudinal forces to the basicranium, what is especially important in the case of shocks coming from the beak. Palaeognathous birds have neither such a middle palate joint nor a mesial support, so there is almost full transmission ofthe forc es from the beak to the quadrate. It follows that shocks can not be reduced in this way and reach the quadrate, its articulations being the most critical points (HoFER, 1945). In palaeognath ous bird s, the shocks are possibly absorbed, instead, by another automat ic device (ELZANOWSKI, in press): when the pt erygoid is pu shed backward s, the quadrate rotates downwards and somewhat inwards, and at the same time, its orbital process presses the pt erygoid against the basipterygoid process. The entire mechanism would fail if the quadrat e were not prevent ed from moving outwards by the zygomatic process of th e squamosa l. Thu s, th e general morphology of the anterior part of the palaeognath ous palate can be correlated with the particular structure ofthe quadratepterygoid articulation and the peculiar zygomatic pr ocess. All palaeognathous birds are primarily rhynchokin etic, although th eir rhynchokinesis is of a special kind. The uniformity of their kinetic mechanism is the second major reason, by which the unity of this group should be recog nized. The structural features relat ed directly with the rhynchokinesis are : (1) Orbital septum continuous with the nasal septum; (2) Nasal openings extending far anteriorly; (3) Lack of cont inuous bony connection between the nasal and the maxilla. Moreover, ratite rhynchokin esis is not associated with any definable zone of bending in the bones roofing the beak, and in some ratit es at least (Casuariformes, Dinornithiform es, Apterygiformes) the nasals are lifted off the premaxillae during the upward movement of the upp er jaw (SIMONETTA 1960). In any case, in all the ratites and some tinamous at least (Crypturus rufescens and Tinamotis elegans) th ere is a loose pr emaxillary-nasal connection (pers. obs.). From the preserved remains, the skulls of Gobipteryx appear to be palaeognathous in nearly all relevant features: (1) In the palate, the pt erygoids articulate suturally with the vomers anteriorly and with the palatines laterally. The posteriorl y diverging vomers are relatively large and are approached by the maxillo-palatines anteriorly. The only obvious interpretation for the medial socket of the posterior part of the pterygoid in the first specimen is the existence ofa basipterygoid joint. Its caudal position would be also indicative of a palaeognathous condition. (2) Among the features directly relat ed with ratite rhynchokinesis, the forward extension of the nasal opening is clearly seen on both specimens. The lack of a bony connection between the nasal and the maxilla is evident on th e second specimen in the free ending nasal process ofthe maxilla. Among birds such a process occurs only in some ratit es (GADOW & SELENKA, 1891). The loose connection between th e nasal and the premaxilla concurs with the other ratite rhynchokinetic charact ers. Hence, Gobipteryx gives a new piece of evidence of the correlation between palatal and directly rhynchokin etic features - all together they can be viewed as the "functional unit of characters" (BOCK, 1963). One of the features that has been thought typical of the palaeognathous skull (PARKES & CLARK, 1966) is lacking in Gobipteryx: on the outer surfaces of the premaxilla and dentary there are no distinct grooves, indicative of rhamphotheca1 division, only some dubious vestiges being present on th e very tips of the bones. It must be noted that the grooves are also lacking in some recent tinam ous (PARKES & CLARK, 1966). The unusual structure of the mand ibular articulation of the Gobipteryx quadrate, not known in the other Palaeognathae, raises a more serious problem. In all other Palaeognathae, as well as in the majority of the Neognath ae, the mandibul ar articulation of the quadrate is roughly triradiate with one part projecting backward s, well behind a straight line between the inner condyle and the quadratojugal articulation. In Apteryx and tinamous, the mandibular articulation of the quadrat e is rath er bipartite with th e outer part moved well backwards, leaving in front a bony shelf, seen in rostral view as a ridge extending from the base of the orbital process and supp orting the quadrat ojugal articulation medially. In the rem aining ratites, the part of the articulation projecting backwards is separated as the posterior condyle, the whole 11 - Palaeontologla Polonica No. 37




. 2.")
10 la Iffi\, I \, I \ \ \ \ \, \ ' \ ' \ I \ I \ I \ " 11XP \ tern 2a Il~ 3 1Je m - ~ I '.\.: '~ -.~-... I \ \\ I ~-, _._.: f \ \ I, ',,:: \ \ I /.1 \, I I \, ' I \ \ I I \ \ I I PAL ", 1/ \\ I, PTT \\ I ", \, \ \ I, \, I ' 1\ ", ::, \'!L x': ;"..:-\~ ' : QOR -i! l '<> ~, I,??/ "<,? >- 8 ~ ltj r No > Z o ~ en ES lb QOT NPM NiIl. I ~ l e m _---t ,, ~--- f, C ~3l...:r="' :!f!"f} "!>liu -~~-- :! ~t-----,-:-=i MXP ~ I cm Fig Gobipteryx minuta E LZANOWSKI, 1974, partial reconstructions' of the skull, a - in ventral view, b - in lateral view (rostral part). 2. Casuarius casuarius (L.), palate.. 3. Apteryx mantelli, BARTL., palate. Abbreviations as in text-fig. 1 II 2b
11 SKULLS OF GOBIPTERYX 163 being strictly comparable with the bipartite condition. In contrast, in the bipartite mandibular articulation of the Gobipteryx quadrate, no part is retracted behind the straight line connecting the inner condyle with the quadratojugal articulation. The lower part of the Gobipteryx quadrate is rather pigeon-like. Against the overwhelming evidence of the palaeognatine affinities of Gobipteryx, the difference in the structure of the mandibular articulation of the quadrate is quite isolated and seems to be of secondary importance for the relations between the major avian taxa. In Platalea the mandibular articulation of the quadrate lacks the posterior part, which occurs in other Ciconiiformes. Within the Palaeognathae, however, this feature appears to be fairly important (although its phylogenetic significance can not be understood until its biomechanical role is ascertained). Among the known palaeognathous birds, Gobipteryx displays the closest similarities to Casuariformes and especially Casuarius: (l ) vomers diverging gradually with (2) the ventral edges of their laminae deflected laterally; (3) pterygoids much widening in the region of the palatine articulation; (4) palatines articulating mostly with the pterygoid; (5) outer margin of the palatine distinctly thicker and lying more ventrally than the inner blade; (5) ascending process of the maxillopalatine present. The last cassowary character was indicated by BOCK (1963, see fig. 4) and is clearly seen on the specimen at the author's disposal (text-fig. 2), yet not referred to by GADOW & SELENKA (1891, p. 31) and SIMONETTA (1960, see fig. 5). In Casuarius, the ascending process is much weaker than in Rhea and, as in Dromaius, it is developed as the posterior, terminal process of the "dorsal, arched lamina" (McDoWELL 1948) of the maxillopalatine. The vestigial, anterior bifurcation of the pterygoid of Gobipteryx seems to be intermediate between Casuarius and Apteryx. In Casuarius the entire posterior margin of the palatine is supported by the pterygoid. In Gobipteryx, the posterior palatine margin is surrounded by the pterygoid, which bears the small outer tine projecting anterolaterally. A similar outer tine is visible on the ventral side of the Apteryx palate as figured by McDoWELL (1948, see text-fig. 5) and observed by the present author on a specimen of A. mantelli. In the figures given by BOCK (1963) and PARKER and HASWELL (1910), the outer tine is not indicated. In any case it does exist and can be seen in its entire extent in dorsal view of the palate, where it has been identified in the second specimen ofgobipteryx. The anterior pterygoid bifurcation in the kiwi was thought by McDoWELL (1948) to be unique among birds and provided one of the principal criterions to distinguish the "apterygiform type" of the palate. In the light of the Gobipteryx palatal structure, the "casuariform"and "apterygiform"types are clearly linked. Therefore, Gobipteryx would also provide a direct evidence for the morphological and phylogenetic unity of the palaeognathous birds and, unless the structure of pterygoid-palatine articulation is a plesiomorphic feature, for closer relations between the Casuariformes and Apterygiformes. Thus, the old views of MIVART (18771 and PARKER (1895), adhered to by SIMONETTA 1 (1957, 1960) and PARKES and CLARK (1966), postulating closer affinities between emus and cassowaries on the one hand and kiwis and moas on the other, would be further confirmed. These hypotheses have recently been denied by CRACRAFT (1974), who approaches emus and cassowaries to the ostriches and rheas on the basis of the synapomorphic features of the postcranial skeleton. Although the relationships between the birds assigned to Palaeognathae seem to be well substantiated, a taxonomic value of the sole palaeognathous condition of the palate appears to be questionable, this condition being "probably truly primitive in birds" (GINGERICH, 1973). In this light, the palaeognathous palate, which occurred in Hesperornis (GINGERICH, 1973), would designate only an early grade of the avian phylogeny. Thus, if the phylogenetic relationships are to be established, a great attention should be paid to differences rather than 1 It should be noted, that the allocation of Casuariformes and Apterygiformes (together with Dinornithiforrnes) into one of three distinct palaeognathous groups has been claimed by SlMONETIA on the basis of the skull anatomy and kinetism. 11
12 164 AND RZE J ELZANOWSKI similarities of the basic pattern, an d the abberrant structure of the Gobipteryx quad rat e may be mor e imp ortant th an assumed in the present study. Palaeogn athous birds are the prime avian example of southern dispersal and th eir present distribution is explaine d by the breakup of the G ondwanalan d (CRACRAFT, 1974; RICH, 1975). Sau er (1972) pointed out, however, that their pa st distribution could be much wider than the present and this opinion is well corroborated by the Gobipteryx discovery. Uniwersytet Warszawski Instyt ut Geologii Podstawowej al. Zw irki i Wigury Warszawa REFER EN CES BOCK, W. J The cra nial evidence for ratite affinities. - Proc. 13th Int. Orn ith. Congr., BVRCHAK-ABRAMOVICH, N. J. (EYP<IAI{-Ar;PAMoDll<I H. H. ) M atcpllajibi K 113)"IeII1lIO ncxonaeasrx CTpaYCOB craporo CBeTa. - Vertebrata Palasiatica 6, 3, CoDY, M. L Competition and the structure of bird communities. Monographs in popul ation biology , Princeton, Princeton Uni v. Press. CRACRAFT, J Conti nent al drift, palaeoclimatology, and the evolution and biogeography of birds. - J, Zool., 169, Phylogeny and evolution of the rati te birds. - Ibis, 116, ELZANOWSKI, A Preliminary note on the palaeognathous bird fro m the Upper Cretaceous of Mongolia. Results Pol.-M ong. Pal. Exp. V. - Palaeont. Pol., 30, (in press). On the role of basipterygoid processes in some birds. - Verh. Anat, Ges. FISCHER, J Aves. In : The fossil record. A symposi um with documentation, Chapter 29, , Geol. Soc. of London. GADOW, H. & SELENKA, E Vogel, 1. Anatomischer Theil. In: H. G. BRONN'S Klassen und Ordnungen des Thier Reichs, 6 (4), , Wintersche VeIL, Leipzig. GINGERICH, P. D Skull of Hesperornis and ea rly evolutio n of birds. - Nature, 243, HOFER, H Untersuchun gen Iiber den Bau des Vogelschadels, besonders ilber den der Spechte und Steisshiihner. Zool. Jahrb. (Ana t.), 69, KIELAN-JAWOROWSKA, Z Multituberculate succession in the late Cre taceo us of the Gobi Desert (Mongolia). Results Pol.-Mong. Pal. Exp. V. - Palaeont. Pol., 30, KIELAN-JAWOROWSKA, Z.& BARSBOLD, R Narra tive of the Polish-Mongolia n Palaeont ological Expeditions Result s Pol.-Mong. Pal. Exp. IV. - Ibidem, 27, McDoWELL, S. B The bony palate of birds. Part. 1. The Palaeogna thae. - Auk, 65, MEISE, W Verhalten der straussar tigen Vogel und Monophylie der Rat itae. - Proc. 13th Int. Ornith. Congr., MIVART, ST. G On the axial skeleton of the Struthionida e. - Trans. Zool. Soc. London, 10, MULLER, H. J Die Morphologie und Entwicklung des Craniu ms von Rhea americana LINNE. 2. Viszeralskelett, Mittelohr und Osteocranium. - Zeit. Wiss. Z ool., 168, PARKER, T. J On the cranial osteology, classification and phylogeny of the Dinornithidae. - Trans. Zool. Soc. London, 13, & HASWELL, W. A A text-book of zoo logy, 2, MacM illan & Company, London. PARKES, K. C. & CLARK, G. A. Jr An additiona l character link ing Rati tes an d Tinamous, and an interpretation of their monophyly, - Condor, 68, RICH, P. V Ant arctic dispersal routes, wanderin g continents, and the origin of Australia' s non-passeriforrn avifauna. - Man. Nat. Mus. Vic., 36, SAVER, E. G. F Ratit e eggshells and phylogenetic questions. - Bonner Z oo!' Beitr., 23, SIMONETTA, A. M Osservazioni sulla mecanica del cranio degli uccelli dro meognathi. - Att i Soc. Tosc. Sci. Nat., B, On the mechanical implications of the avian skull and their bearing on the evolution and classification of birds. - Quart. Rev. Bioi., 35, WEBB, M The ont ogeny of the cran ial bones, cranial perip heral and cra nial parasympathetic nerves, together with a study of the visceral muscles of Stru thio. - Acta Z ool., 38,
13 SKULLS OF GOBIPTERYX 165 EXPLA NATIO N OF PLATE PLATE 37 Gobipteryx minuta ELZANOWSKI, Page 154 Upper Cretaceous, Barun Goyot Formation, Khulsan, Gob i Desert, Mongolia la. Stereophotograph of the skull in dor salview. ZPAL MgR-I/32; x 2.5. lb. Stereophotograph of the same in ventral and somewhat lateral view; x 2.5. I c. Stereophotograph of the same in right, lateral view; x 2.5. Id. Stereophotograph of the same in left lateral view; x 2.5. l e. Stereophotograph of the same in ventral view; x 2.5. If. Stereophotograph of the same in rostral view; x 5. 2a. Stereophotograph of the skull in rostral view. Type specimen, ZPAL MgR-I/12; x 5. 2b. Stereophotograph of the mandi ble of the same specimen in medial view; x 6. Photo : E. Wyr zyko"ska

14 Palaeontologia Polonica. No. 37, 1977 PI. 37 A. ELZANOWSKI: SKULLS OF GOBJPTERYX
AMERICAN MUSEUM NOVITATES Published by
 AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
ONLINE APPENDIX 1. Morphological phylogenetic characters scored in this paper. See Poe (2004) for
 for") ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
SUPPLEMENTARY ONLINE MATERIAL FOR. Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor
 http://app.pan.pl/som/app61-ratsimbaholison_etal_som.pdf SUPPLEMENTARY ONLINE MATERIAL FOR Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor Ontogenetic changes in the craniomandibular
http://app.pan.pl/som/app61-ratsimbaholison_etal_som.pdf SUPPLEMENTARY ONLINE MATERIAL FOR Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor Ontogenetic changes in the craniomandibular
CHAPTER 6 CRANIAL KINESIS IN PALAEOGNATHOUS BIRDS. 6. Cranial Kinesis in Palaeognathous Birds
 6. Cranial Kinesis in Palaeognathous Birds CHAPTER 6 CRANIAL KINESIS IN PALAEOGNATHOUS BIRDS Summary In palaeognathous birds the morphology of the Pterygoid-Palatinum Complex (PPC) is remarkably different
6. Cranial Kinesis in Palaeognathous Birds CHAPTER 6 CRANIAL KINESIS IN PALAEOGNATHOUS BIRDS Summary In palaeognathous birds the morphology of the Pterygoid-Palatinum Complex (PPC) is remarkably different
New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia
 1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
complex in cusp pattern. (3) The bones of the coyote skull are thinner, crests sharper and the
 The bones of the coyote skull are thinner, crests sharper and the") DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
9. Summary & General Discussion CHAPTER 9 SUMMARY & GENERAL DISCUSSION
 9. Summary & General Discussion CHAPTER 9 SUMMARY & GENERAL DISCUSSION 143 The Evolution of the Paleognathous Birds 144 9. Summary & General Discussion General Summary The evolutionary history of the Palaeognathae
9. Summary & General Discussion CHAPTER 9 SUMMARY & GENERAL DISCUSSION 143 The Evolution of the Paleognathous Birds 144 9. Summary & General Discussion General Summary The evolutionary history of the Palaeognathae
HONR219D Due 3/29/16 Homework VI
 Part 1: Yet More Vertebrate Anatomy!!! HONR219D Due 3/29/16 Homework VI Part 1 builds on homework V by examining the skull in even greater detail. We start with the some of the important bones (thankfully
Part 1: Yet More Vertebrate Anatomy!!! HONR219D Due 3/29/16 Homework VI Part 1 builds on homework V by examining the skull in even greater detail. We start with the some of the important bones (thankfully
2. Skull, total length versus length of the presacral vertebral column: (0); extremely elongated neck (e.g. Tanystropheus longobardicus).
; extremely elongated neck (e.g. Tanystropheus longobardicus).") Character list of the taxon-character data set 1. Skull and lower jaws, interdental plates: absent (0); present, but restricted to the anterior end of the dentary (1); present along the entire alveolar
Character list of the taxon-character data set 1. Skull and lower jaws, interdental plates: absent (0); present, but restricted to the anterior end of the dentary (1); present along the entire alveolar
A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA
 A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA MYCTEROSAURUS LONGICEPS S. W. WILLISTON University of Chicago The past summer, Mr. Herman Douthitt, of the University of Chicago paleontological expedition,
A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA MYCTEROSAURUS LONGICEPS S. W. WILLISTON University of Chicago The past summer, Mr. Herman Douthitt, of the University of Chicago paleontological expedition,
SUPPLEMENTARY INFORMATION
 Character 155, interdental ridges. Absence of interdental ridge (0) shown in Parasaniwa wyomingensis (Platynota). Interdental ridges (1) shown in Coniophis precedens. WWW.NATURE.COM/NATURE 1 Character
Character 155, interdental ridges. Absence of interdental ridge (0) shown in Parasaniwa wyomingensis (Platynota). Interdental ridges (1) shown in Coniophis precedens. WWW.NATURE.COM/NATURE 1 Character
A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.)
") A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.) by Ouyang Hui Zigong Dinosaur Museum Newsletter Number 2 1989 pp. 10-14 Translated By Will Downs Bilby
A new sauropod from Dashanpu, Zigong Co. Sichuan Province (Abrosaurus dongpoensis gen. et sp. nov.) by Ouyang Hui Zigong Dinosaur Museum Newsletter Number 2 1989 pp. 10-14 Translated By Will Downs Bilby
YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
 Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE
 PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
v:ii-ixi, 'i':;iisimvi'\>!i-:: "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi L I E) R.ARY OF THE VERSITY U N I or ILLINOIS REMO
 R.ARY OF THE VERSITY U N I or ILLINOIS REMO") "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
"^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the
 Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the") Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
Mammalogy Laboratory 1 - Mammalian Anatomy
 Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province
 from Dashanpu, Zigong Municipality, Sichuan Province") A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
A skull without mandihle, from the Hunterian Collection (no.
 4 MR. G. A. BOULENGER ON CHELONIAN REMAINS. [Jan. 6, 2. On some Chelonian Remains preserved in the Museum of the Eojal College of Surgeons. By G. A. Boulenger. [Eeceived December 8, 1890.] In the course
4 MR. G. A. BOULENGER ON CHELONIAN REMAINS. [Jan. 6, 2. On some Chelonian Remains preserved in the Museum of the Eojal College of Surgeons. By G. A. Boulenger. [Eeceived December 8, 1890.] In the course
Vol. XIV, No. 1, March, The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S.
 By S.") Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Anatomy. Name Section. The Vertebrate Skeleton
 Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC
 FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
THE SKULLS OF ARAEOSCELIS AND CASEA, PERMIAN REPTILES
 THE SKULLS OF REOSCELIS ND CSE, PERMIN REPTILES University of Chicago There are few Permian reptiles of greater interest at the present time than the peculiar one I briefly described in this journal' three
THE SKULLS OF REOSCELIS ND CSE, PERMIN REPTILES University of Chicago There are few Permian reptiles of greater interest at the present time than the peculiar one I briefly described in this journal' three
Williston, and as there are many fairly good specimens in the American
 56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
The basal clades of modern birds
 The basal clades of modern birds Joel Cracraft Department of Ornithology, American Museum of Natural History Central Park West at 79th Street, New York, NY 10024 U.S.A. E-mail: JLC@amnh.org Julia Clarke
The basal clades of modern birds Joel Cracraft Department of Ornithology, American Museum of Natural History Central Park West at 79th Street, New York, NY 10024 U.S.A. E-mail: JLC@amnh.org Julia Clarke
SOME NEW AMERICAN PYCNODONT FISHES.
 SOME NEW AMERICAN PYCNODONT FISHES. By James Williams Gidley, Assistant Curator of Fossil Mammals, United States National Museum. In the United States National Museum are several specimens representing
SOME NEW AMERICAN PYCNODONT FISHES. By James Williams Gidley, Assistant Curator of Fossil Mammals, United States National Museum. In the United States National Museum are several specimens representing
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A.
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
THE PALAEOGNATHOUS PTERYGOID-PALATINUM COMPLEX. A TRUE CHARACTER?
 2. The Palaeognathous Pterygoid-Palatinum Complex. A True Character? CHAPTER 2 THE PALAEOGNATHOUS PTERYGOID-PALATINUM COMPLEX. A TRUE CHARACTER? Summary Molecular analyses show that modern birds can be
2. The Palaeognathous Pterygoid-Palatinum Complex. A True Character? CHAPTER 2 THE PALAEOGNATHOUS PTERYGOID-PALATINUM COMPLEX. A TRUE CHARACTER? Summary Molecular analyses show that modern birds can be
The cranial osteology of Belebey vegrandis (Parareptilia: Bolosauridae), from the Middle Permian of Russia, and its bearing on reptilian evolution
, from the Middle Permian of Russia, and its bearing on reptilian evolution") Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1511 191214 Original Articles RUSSIAN BOLOSAURID REPTILER. R. REISZ ET AL.
Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082 2007 The Linnean Society of London? 2007 1511 191214 Original Articles RUSSIAN BOLOSAURID REPTILER. R. REISZ ET AL.
The Discovery of a Tritylodont from the Xinjiang Autonomous Region
 The Discovery of a Tritylodont from the Xinjiang Autonomous Region Ailing Sun and Guihai Cui (Institute of Vertebrate Paleontology, Paleoanthropology, Academia Sinica) Vertebrata PalAsiatica Volume XXVII,
The Discovery of a Tritylodont from the Xinjiang Autonomous Region Ailing Sun and Guihai Cui (Institute of Vertebrate Paleontology, Paleoanthropology, Academia Sinica) Vertebrata PalAsiatica Volume XXVII,
Cranial osteology of the African gerrhosaurid Angolosaurus skoogi (Squamata; Gerrhosauridae) HOLLY A. NANCE
 HOLLY A. NANCE") African Journal of Herpetology, 2007 56(1): 39-75. Herpetological Association of Africa Original article Cranial osteology of the African gerrhosaurid Angolosaurus skoogi (Squamata; Gerrhosauridae) HOLLY
African Journal of Herpetology, 2007 56(1): 39-75. Herpetological Association of Africa Original article Cranial osteology of the African gerrhosaurid Angolosaurus skoogi (Squamata; Gerrhosauridae) HOLLY
.56 m. (22 in.). COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition
. COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition") Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Description of Cranial Elements and Ontogenetic Change within Tropidolaemus wagleri (Serpentes: Crotalinae).
.") East Tennessee State University Digital Commons @ East Tennessee State University Electronic Theses and Dissertations 5-2016 Description of Cranial Elements and Ontogenetic Change within Tropidolaemus
East Tennessee State University Digital Commons @ East Tennessee State University Electronic Theses and Dissertations 5-2016 Description of Cranial Elements and Ontogenetic Change within Tropidolaemus
Mar., 1963 RELATIONSHIPS BETWEEN THE BIRDS OF PARADISE AND THE BOWER BIRDS. By WALTER J. BOCK
 Mar., 1963 91 RELATIONSHIPS BETWEEN THE BIRDS OF PARADISE AND THE BOWER BIRDS By WALTER J. BOCK INTRODUCTION Ever since their discovery in the early days of world exploration, the birds of paradise and
Mar., 1963 91 RELATIONSHIPS BETWEEN THE BIRDS OF PARADISE AND THE BOWER BIRDS By WALTER J. BOCK INTRODUCTION Ever since their discovery in the early days of world exploration, the birds of paradise and
Mammalogy Lecture 8 - Evolution of Ear Ossicles
 Mammalogy Lecture 8 - Evolution of Ear Ossicles I. To begin, let s examine briefly the end point, that is, modern mammalian ears. Inner Ear The cochlea contains sensory cells for hearing and balance. -
Mammalogy Lecture 8 - Evolution of Ear Ossicles I. To begin, let s examine briefly the end point, that is, modern mammalian ears. Inner Ear The cochlea contains sensory cells for hearing and balance. -
CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN OF RUSSIA AND THE EVOLUTIONARY RELATIONSHIPS OF CASEIDAE
 FROM THE MIDDLE PERMIAN OF RUSSIA AND THE EVOLUTIONARY RELATIONSHIPS OF CASEIDAE") Journal of Vertebrate Paleontology 28(1):160 180, March 2008 2008 by the Society of Vertebrate Paleontology ARTICLE CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN
Journal of Vertebrate Paleontology 28(1):160 180, March 2008 2008 by the Society of Vertebrate Paleontology ARTICLE CRANIAL ANATOMY OF ENNATOSAURUS TECTON (SYNAPSIDA: CASEIDAE) FROM THE MIDDLE PERMIAN
BREVIORA LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1
, A NEW INDO-PACIFIC SAND CRAB. Ian E. Efford 1") ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
ac lc BREVIORA CAMBRIDGE, MASS. 30 APRIL, 1969 NUMBER 318 LEUCOLEPIDOPA SUNDA GEN. NOV., SP. NOV. (DECAPODA: ALBUNEIDAE), A NEW INDO-PACIFIC SAND CRAB Ian E. Efford 1 ABSTRACT. Leucolepidopa gen. nov.
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/329/5998/1481/dc1 Supporting Online Material for Tyrannosaur Paleobiology: New Research on Ancient Exemplar Organisms Stephen L. Brusatte,* Mark A. Norell, Thomas D.
www.sciencemag.org/cgi/content/full/329/5998/1481/dc1 Supporting Online Material for Tyrannosaur Paleobiology: New Research on Ancient Exemplar Organisms Stephen L. Brusatte,* Mark A. Norell, Thomas D.
The Lower Jaws of Baenid Turtles
 AMERICAN MUSEUM Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N.Y. 10024 Number 2749, pp. 1-10, figs. 1-4, table 1 September 27, 1982 The Lower
AMERICAN MUSEUM Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N.Y. 10024 Number 2749, pp. 1-10, figs. 1-4, table 1 September 27, 1982 The Lower
A Fossil Snake (Elaphe vulpina) From A Pliocene Ash Bed In Nebraska
 From A Pliocene Ash Bed In Nebraska") University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to
 1 Supplementary data CHARACTER LIST List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to characters used by Tchernov et al. (2000), Rieppel, et al. (2002), and Lee
1 Supplementary data CHARACTER LIST List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to characters used by Tchernov et al. (2000), Rieppel, et al. (2002), and Lee
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO
 A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
THREE-DIMENSIONAL KINEMATICS OF SKELETAL ELEMENTS IN AVIAN PROKINETIC AND RHYNCHOKINETIC SKULLS DETERMINED BY ROENTGEN STEREOPHOTOGRAMMETRY
 The Journal of Experimental Biology 4, 1735 1744 (01) Printed in Great Britain The Company of Biologists Limited 01 JEB3132 1735 THREE-DIMENSIONAL KINEMATICS OF SKELETAL ELEMENTS IN AVIAN PROKINETIC AND
The Journal of Experimental Biology 4, 1735 1744 (01) Printed in Great Britain The Company of Biologists Limited 01 JEB3132 1735 THREE-DIMENSIONAL KINEMATICS OF SKELETAL ELEMENTS IN AVIAN PROKINETIC AND
MACROCEPHALOSAURIDAE AND POLYGL YPHANODONTIDAE (SAURIA) FROM THE LATE CRETACEOUS OF MONGOLIA
 FROM THE LATE CRETACEOUS OF MONGOLIA") ANDRZEJ SULIMSKI MACROCEPHALOSAURIDAE AND POLYGL YPHANODONTIDAE (SAURIA) FROM THE LATE CRETACEOUS OF MONGOLIA (Plates VIII-XXVII) Abstract - Large Late Cretaceous lizards from the?santonian Djadokhta Formation,?
ANDRZEJ SULIMSKI MACROCEPHALOSAURIDAE AND POLYGL YPHANODONTIDAE (SAURIA) FROM THE LATE CRETACEOUS OF MONGOLIA (Plates VIII-XXVII) Abstract - Large Late Cretaceous lizards from the?santonian Djadokhta Formation,?
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at
 The Evolution of the Mammalian Jaw Author(s): A. W. Crompton Source: Evolution, Vol. 17, No. 4 (Dec., 1963), pp. 431-439 Published by: Society for the Study of Evolution Stable URL: http://www.jstor.org/stable/2407093
The Evolution of the Mammalian Jaw Author(s): A. W. Crompton Source: Evolution, Vol. 17, No. 4 (Dec., 1963), pp. 431-439 Published by: Society for the Study of Evolution Stable URL: http://www.jstor.org/stable/2407093
290 SHUFELDT, Remains of Hesperornis.
 290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
THE CONDOR THE JAWS OF THE CRETACEOUS TOOTHED BIRDS,
 THE CONDOR VOLUME 54 MARCH-APRIL, 1952 NUMBER 2 THE JAWS OF THE CRETACEOUS TOOTHED BIRDS, ICHTHYORNIS AND HESPERORNIS By JOSEPH T. GREGORY The remarkable Cretaceous birds with teeth, widel; cited in paleontological
THE CONDOR VOLUME 54 MARCH-APRIL, 1952 NUMBER 2 THE JAWS OF THE CRETACEOUS TOOTHED BIRDS, ICHTHYORNIS AND HESPERORNIS By JOSEPH T. GREGORY The remarkable Cretaceous birds with teeth, widel; cited in paleontological
A Complete Late Cretaceous Iguanian (Squamata, Reptilia) from the Gobi and Identification of a New Iguanian Clade
 from the Gobi and Identification of a New Iguanian Clade") PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3584, 47 pp., 19 figures September 6, 2007 A Complete Late Cretaceous Iguanian (Squamata,
PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3584, 47 pp., 19 figures September 6, 2007 A Complete Late Cretaceous Iguanian (Squamata,
New information on the palate and lower jaw of Massospondylus (Dinosauria: Sauropodomorpha)
") New information on the palate and lower jaw of Massospondylus (Dinosauria: Sauropodomorpha) Paul M. Barrett 1* & Adam M. Yates 2* 1 Department of Palaeontology, The Natural History Museum, Cromwell Road,
New information on the palate and lower jaw of Massospondylus (Dinosauria: Sauropodomorpha) Paul M. Barrett 1* & Adam M. Yates 2* 1 Department of Palaeontology, The Natural History Museum, Cromwell Road,
TRACHEMYS SCULPTA. A nearly complete articulated carapace and plastron of an Emjdd A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE,
 A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE
 OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT This is a report of measurements on the skeleton of a male se1 whale caught in the Antarctic. The skeleton of
OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT This is a report of measurements on the skeleton of a male se1 whale caught in the Antarctic. The skeleton of
The family Gnaphosidae is a large family
 Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
FIELDIANA GEOLOGY NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA
 FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL
 Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia
 from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia") First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia Tsogtbaatar Chinzorig¹, ³ *, Yoshitsugu Kobayashi², Khishigjav Tsogtbaatar³,
First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia Tsogtbaatar Chinzorig¹, ³ *, Yoshitsugu Kobayashi², Khishigjav Tsogtbaatar³,
A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province
 A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province by Hu Shaojin (Kunming Cultural Administrative Committee, Yunnan Province) Vertebrata PalAsiatica Vol. XXXI, No. 1
A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province by Hu Shaojin (Kunming Cultural Administrative Committee, Yunnan Province) Vertebrata PalAsiatica Vol. XXXI, No. 1
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE
 FROM THE") A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
VARIATION IN MONIEZIA EXPANSA RUDOLPHI
 VARIATION IN MONIEZIA EXPANSA RUDOLPHI STEPHEN R. WILLIAMS, Miami University, Oxford, Ohio In making a number of preparations of proglottids for class study at the stage when sex organs are mature and
VARIATION IN MONIEZIA EXPANSA RUDOLPHI STEPHEN R. WILLIAMS, Miami University, Oxford, Ohio In making a number of preparations of proglottids for class study at the stage when sex organs are mature and
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * Dr. L.D. Boonstra. Paleontologist, South African Museum, Cape Town
 THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
AMERICAN MUSEUM. Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET
 AMERICAN MUSEUM Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET NEW YORK, N.Y. 10024 U.S.A. NUMBER 2662 NOVEMBER 21, 1978 RONN W. COLDIRON Acroplous vorax
AMERICAN MUSEUM Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET NEW YORK, N.Y. 10024 U.S.A. NUMBER 2662 NOVEMBER 21, 1978 RONN W. COLDIRON Acroplous vorax
VERTEBRATA PALASIATICA
 41 2 2003 2 VERTEBRATA PALASIATICA pp. 147 156 figs. 1 5 1) ( 100044), ( Parakannemeyeria brevirostris),,, : ( Xiyukannemeyeria),,, Q915. 864 60 Turfania (,1973), Dicynodon (, 1973 ; Lucas, 1998), (Lystrosaurus)
41 2 2003 2 VERTEBRATA PALASIATICA pp. 147 156 figs. 1 5 1) ( 100044), ( Parakannemeyeria brevirostris),,, : ( Xiyukannemeyeria),,, Q915. 864 60 Turfania (,1973), Dicynodon (, 1973 ; Lucas, 1998), (Lystrosaurus)
CENE RUMINANTS OF THE GENERA OVIBOS AND
 DESCRIPTIONS OF TWO NEW SPECIES OF PLEISTO- CENE RUMINANTS OF THE GENERA OVIBOS AND BOOTHERIUM, WITH NOTES ON THE LATTER GENUS. By James Williams Gidley, Of the United States National Museum. Two interesting
DESCRIPTIONS OF TWO NEW SPECIES OF PLEISTO- CENE RUMINANTS OF THE GENERA OVIBOS AND BOOTHERIUM, WITH NOTES ON THE LATTER GENUS. By James Williams Gidley, Of the United States National Museum. Two interesting
SMITHSONIAN CONTRIBUTIONS TO PALEOBIOLOGY NUMBER 89
 SMITHSONIAN CONTRIBUTIONS TO PALEOBIOLOGY NUMBER 89 Avian Paleontology at the Close of the 20th Century: Proceedings of the 4th International Meeting of the Society of Avian Paleontology and Evolution,
SMITHSONIAN CONTRIBUTIONS TO PALEOBIOLOGY NUMBER 89 Avian Paleontology at the Close of the 20th Century: Proceedings of the 4th International Meeting of the Society of Avian Paleontology and Evolution,
Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved
 Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved This was a private report in 2003 on my thoughts on Platecarpus planifrons.
Bulletin of Big Bend Paleo-Geo An Open Access Publication from Mosasaur Ranch Museum, Terlingua and Lajitas, Texas All rights reserved This was a private report in 2003 on my thoughts on Platecarpus planifrons.
Marshall Digital Scholar. Marshall University. F. Robin O Keefe Marshall University,
 Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 2008 Cranial anatomy and taxonomy of Dolichorhynchops bonneri new combination, a polycotylid (Sauropterygia:
Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 2008 Cranial anatomy and taxonomy of Dolichorhynchops bonneri new combination, a polycotylid (Sauropterygia:
DISCOVERY OF GENUS PLATOLENES (COLEOP TERA : TENEBRIONIDAE) FROM INDIA WITH DESCRIPTION OF TWO NEW SPECIES G. N. SABA
 FROM INDIA WITH DESCRIPTION OF TWO NEW SPECIES G. N. SABA") Rec. zool. Surv. India, 85(3) : 433-437,1988 DISCOVERY OF GENUS PLATOLENES (COLEOP TERA : TENEBRIONIDAE) FROM INDIA WITH DESCRIPTION OF TWO NEW SPECIES By G. N. SABA Zoological Survey of India M-Block,
Rec. zool. Surv. India, 85(3) : 433-437,1988 DISCOVERY OF GENUS PLATOLENES (COLEOP TERA : TENEBRIONIDAE) FROM INDIA WITH DESCRIPTION OF TWO NEW SPECIES By G. N. SABA Zoological Survey of India M-Block,
PALEONTOLOGY AND BIOSTRATIGRAPHY OF MONGOLIA
 PALEONTOLOGY AND BIOSTRATIGRAPHY OF MONGOLIA THE JOINT SOVIET-MONGOLIAN PALEONTOLOGICAL EXPEDITION (Transactions, vol. 3) EDITORIAL BOARD: N. N. Kramarenko (editor-in-chief) B. Luvsandansan, Yu. I. Voronin,
PALEONTOLOGY AND BIOSTRATIGRAPHY OF MONGOLIA THE JOINT SOVIET-MONGOLIAN PALEONTOLOGICAL EXPEDITION (Transactions, vol. 3) EDITORIAL BOARD: N. N. Kramarenko (editor-in-chief) B. Luvsandansan, Yu. I. Voronin,
Department of Biology, Faculty of Science, Razi University, Kermanshah, Iran 2
 Iranian Journal of Animal Biosystematics (IJAB) Vol.13, No.2, 247-262, 2017 ISSN: 1735-434X (print); 2423-4222 (online) DOI: 10.22067/ijab.v13i2.64614 A comparative study of the skull between Trachylepis
Iranian Journal of Animal Biosystematics (IJAB) Vol.13, No.2, 247-262, 2017 ISSN: 1735-434X (print); 2423-4222 (online) DOI: 10.22067/ijab.v13i2.64614 A comparative study of the skull between Trachylepis
PEABODY MUSEUM OF NATURAL HISTORY, YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 85 September 21, 1964 A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA STANLEY J. RIEL
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 85 September 21, 1964 A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA STANLEY J. RIEL
A NEW SPECIES OF TROODONT DINOSAUR FROM THE
 A NEW SPECIES OF TROODONT DINOSAUR FROM THE LANCE FORMATION OF WYOMING By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION The intensive search to which
A NEW SPECIES OF TROODONT DINOSAUR FROM THE LANCE FORMATION OF WYOMING By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION The intensive search to which
A New Dromaeosaurid Theropod from Ukhaa Tolgod (Ömnögov, Mongolia)
") PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3545, 51 pp., 25 figures, 1 table December 7, 2006 A New Dromaeosaurid Theropod from Ukhaa
PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3545, 51 pp., 25 figures, 1 table December 7, 2006 A New Dromaeosaurid Theropod from Ukhaa
Notes on Ceratopsians and Ankylosaurs at the Royal Ontario Museum
 Notes on Ceratopsians and Ankylosaurs at the Royal Ontario Museum Andrew A. Farke, Ph.D. Raymond M. Alf Museum of Paleontology 1175 West Baseline Road Claremont, CA 91711 email: afarke@webb.org Introduction
Notes on Ceratopsians and Ankylosaurs at the Royal Ontario Museum Andrew A. Farke, Ph.D. Raymond M. Alf Museum of Paleontology 1175 West Baseline Road Claremont, CA 91711 email: afarke@webb.org Introduction
[Accepted 8th October CONTENTS INTRODUCTION
 183 THE CRANIAL MORPHOLOGY OF A NEW GENUS AND SPECIES OF ICTIDOSAURAN BY A. W. CROMPTON S. A. Museum, Cape Town [Accepted 8th October 19571 (With 7 figures in the text) CONTENTS lntroduction..............
183 THE CRANIAL MORPHOLOGY OF A NEW GENUS AND SPECIES OF ICTIDOSAURAN BY A. W. CROMPTON S. A. Museum, Cape Town [Accepted 8th October 19571 (With 7 figures in the text) CONTENTS lntroduction..............
Diurus, Pascoe. sp. 1). declivity of the elytra, but distinguished. Length (the rostrum and tails 26 included) mm. Deep. exception
. declivity of the elytra, but distinguished. Length (the rostrum and tails 26 included) mm. Deep. exception") 210 DIURUS ERYTIIROPUS. NOTE XXVI. Three new species of the Brenthid genus Diurus, Pascoe DESCRIBED BY C. Ritsema+Cz. 1. Diurus erythropus, n. sp. 1). Allied to D. furcillatus Gylh. ²) by the short head,
210 DIURUS ERYTIIROPUS. NOTE XXVI. Three new species of the Brenthid genus Diurus, Pascoe DESCRIBED BY C. Ritsema+Cz. 1. Diurus erythropus, n. sp. 1). Allied to D. furcillatus Gylh. ²) by the short head,
A New Ceratopsian Dinosaur from the Upper
 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 63. NUMBER 3 A New Ceratopsian Dinosaur from the Upper Cretaceous of Montana, with Note on Hypacrosaurus (With Two Plates) CHARLES W. GILMORE Assistant Curator
SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 63. NUMBER 3 A New Ceratopsian Dinosaur from the Upper Cretaceous of Montana, with Note on Hypacrosaurus (With Two Plates) CHARLES W. GILMORE Assistant Curator
Lower Cretaceous Kwanmon Group, Northern Kyushu
 Bull. Kitakyushu Mus. Nat. Hist., 11: 87-90. March 30, 1992 A New Genus and Species of Carnivorous Dinosaur from the Lower Cretaceous Kwanmon Group, Northern Kyushu Yoshihiko Okazaki Kitakyushu Museum
Bull. Kitakyushu Mus. Nat. Hist., 11: 87-90. March 30, 1992 A New Genus and Species of Carnivorous Dinosaur from the Lower Cretaceous Kwanmon Group, Northern Kyushu Yoshihiko Okazaki Kitakyushu Museum
OF THE TRIAS THE PHYTOSAURIA
 THE PHYTOSAURIA OF THE TRIAS MAURICE G. MEHL University of Wisconsin Some time ago the writer gave a brief notice of a new genus of phytosaurs of which Angistorhinus grandis Mehl was the type.' It is the
THE PHYTOSAURIA OF THE TRIAS MAURICE G. MEHL University of Wisconsin Some time ago the writer gave a brief notice of a new genus of phytosaurs of which Angistorhinus grandis Mehl was the type.' It is the
Cranial morphology of Sinornithosaurus millenii Xu et al (Dinosauria: Theropoda: Dromaeosauridae) from the Yixian Formation of Liaoning, China
 from the Yixian Formation of Liaoning, China") 1739 Cranial morphology of Sinornithosaurus millenii Xu et al. 1999 (Dinosauria: Theropoda: Dromaeosauridae) from the Yixian Formation of Liaoning, China Xing Xu and Xiao-Chun Wu Abstract: The recent discovery
1739 Cranial morphology of Sinornithosaurus millenii Xu et al. 1999 (Dinosauria: Theropoda: Dromaeosauridae) from the Yixian Formation of Liaoning, China Xing Xu and Xiao-Chun Wu Abstract: The recent discovery
THE OCCURRENCE OF CONTOGENYS-LIKE LIZARDS IN THE LATE CRETACEOUS AND EARLY TERTIARY OF THE WESTERN INTERIOR OF THE U.S.A.
 Journal of Vertebrate Paleontology 29(3):677 701, September 2009 # 2009 by the Society of Vertebrate Paleontology ARTICLE THE OCCURRENCE OF CONTOGENYS-LIKE LIZARDS IN THE LATE CRETACEOUS AND EARLY TERTIARY
Journal of Vertebrate Paleontology 29(3):677 701, September 2009 # 2009 by the Society of Vertebrate Paleontology ARTICLE THE OCCURRENCE OF CONTOGENYS-LIKE LIZARDS IN THE LATE CRETACEOUS AND EARLY TERTIARY
A new species of torrent toad (Genus Silent Valley, S. India
 Proc. Indian Acad. Sci. (Anirn. ScL), Vol. 90, Number 2, March 1981, pp. 203-208. Printed in India. A new species of torrent toad (Genus Silent Valley, S. India Allsollia) from R S PILLAI and R PATTABIRAMAN
Proc. Indian Acad. Sci. (Anirn. ScL), Vol. 90, Number 2, March 1981, pp. 203-208. Printed in India. A new species of torrent toad (Genus Silent Valley, S. India Allsollia) from R S PILLAI and R PATTABIRAMAN
 The following text is generated from uncorrected OCR. [Begin Page: Page 1] A NEW CERATOPSIAN DINOSAUR FROM THE UPPER CRETACEOUS OF MONTANA, WITH NOTE ON HYPACROSAURUS ' By CHARLES W. GILMORE assistant
The following text is generated from uncorrected OCR. [Begin Page: Page 1] A NEW CERATOPSIAN DINOSAUR FROM THE UPPER CRETACEOUS OF MONTANA, WITH NOTE ON HYPACROSAURUS ' By CHARLES W. GILMORE assistant
On the morphoplogy and taxonomic status of Xinpusaurus kohi JIANG et al., 2004 (Diapsida: Thalattosauria) from the Upper Triassic of China
 from the Upper Triassic of China") Palaeodiversity 7: 47 59; Stuttgart 30 December 2014. 47 On the morphoplogy and taxonomic status of Xinpusaurus kohi JIANG et al., 2004 (Diapsida: Thalattosauria) from the Upper Triassic of China MICHAEL
Palaeodiversity 7: 47 59; Stuttgart 30 December 2014. 47 On the morphoplogy and taxonomic status of Xinpusaurus kohi JIANG et al., 2004 (Diapsida: Thalattosauria) from the Upper Triassic of China MICHAEL
A NEW SPECIES OF A USTROLIBINIA FROM THE SOUTH CHINA SEA AND INDONESIA (CRUSTACEA: BRACHYURA: MAJIDAE)
") 69 C O a g r ^ j^a RAFFLES BULLETIN OF ZOOLOGY 1992 40(1): 69-73 A NEW SPECIES OF A USTROLIBINIA FROM THE SOUTH CHINA SEA AND INDONESIA (CRUSTACEA: BRACHYURA: MAJIDAE) H P Waener SMITHSONIAN INSTITUTE
69 C O a g r ^ j^a RAFFLES BULLETIN OF ZOOLOGY 1992 40(1): 69-73 A NEW SPECIES OF A USTROLIBINIA FROM THE SOUTH CHINA SEA AND INDONESIA (CRUSTACEA: BRACHYURA: MAJIDAE) H P Waener SMITHSONIAN INSTITUTE
Toothless Dinosaurs of Mongolia. R. Barsbold. Fossil Vertebrates of Mongolia Academy of Sciences of USSR Paleontological Institute
 Toothless Dinosaurs of Mongolia R. Barsbold Fossil Vertebrates of Mongolia Academy of Sciences of USSR Paleontological Institute Academy of Sciences of the MPR Geological Institute Nauka Moscow 1981 Trudy
Toothless Dinosaurs of Mongolia R. Barsbold Fossil Vertebrates of Mongolia Academy of Sciences of USSR Paleontological Institute Academy of Sciences of the MPR Geological Institute Nauka Moscow 1981 Trudy
NOTE XVII. Dr. A.A.W. Hubrecht. which should he in accordance with. of my predecessors. alive or in excellent. further
 further either EUROPEAN NEMERTEANS. 93 NOTE XVII. New Species of European Nemerteans. First Appendix to Note XLIV, Vol. I BY Dr. A.A.W. Hubrecht In the above-mentioned note, published six months ago, several
further either EUROPEAN NEMERTEANS. 93 NOTE XVII. New Species of European Nemerteans. First Appendix to Note XLIV, Vol. I BY Dr. A.A.W. Hubrecht In the above-mentioned note, published six months ago, several
University of Iowa Iowa Research Online
 University of Iowa Iowa Research Online Theses and Dissertations Spring 2016 A reassessment of the late Eocene - early Oligocene crocodylids Crocodylus megarhinus Andrews 1905 and Crocodylus articeps Andrews
University of Iowa Iowa Research Online Theses and Dissertations Spring 2016 A reassessment of the late Eocene - early Oligocene crocodylids Crocodylus megarhinus Andrews 1905 and Crocodylus articeps Andrews
A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae)
") Genus Vol. 14 (3): 413-418 Wroc³aw, 15 X 2003 A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae) JAROS AW KANIA Zoological Institute, University of Wroc³aw, Sienkiewicza
Genus Vol. 14 (3): 413-418 Wroc³aw, 15 X 2003 A new species of Antinia PASCOE from Burma (Coleoptera: Curculionidae: Entiminae) JAROS AW KANIA Zoological Institute, University of Wroc³aw, Sienkiewicza
Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons
 Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons 1. Head skeleton of lamprey Cyclostomes are highly specialized in both the construction of the chondrocranium and visceral skeleton.
Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons 1. Head skeleton of lamprey Cyclostomes are highly specialized in both the construction of the chondrocranium and visceral skeleton.
Supplementary Note 1. Additional osteological description
 Supplementary Note 1 Additional osteological description The text below provides additional details of Jianianhualong that were not pertinent to the salient osteological description provided in the main
Supplementary Note 1 Additional osteological description The text below provides additional details of Jianianhualong that were not pertinent to the salient osteological description provided in the main
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS
 AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1964. Merostomoidea (Arthropoda, Trilobitomorpha) from the Australian Middle Triassic. Records of the Australian Museum 26(13): 327 332, plate 35.
NEW INFORMATION ON THE CRANIUM OF BRACHYLOPHOSAURUS CANADENSIS (DINOSAURIA, HADROSAURIDAE), WITH A REVISION OF ITS PHYLOGENETIC POSITION
, WITH A REVISION OF ITS PHYLOGENETIC POSITION") Journal of Vertebrate Paleontology 25(1):144 156, March 2005 2005 by the Society of Vertebrate Paleontology NEW INFORMATION ON THE CRANIUM OF BRACHYLOPHOSAURUS CANADENSIS (DINOSAURIA, HADROSAURIDAE), WITH
Journal of Vertebrate Paleontology 25(1):144 156, March 2005 2005 by the Society of Vertebrate Paleontology NEW INFORMATION ON THE CRANIUM OF BRACHYLOPHOSAURUS CANADENSIS (DINOSAURIA, HADROSAURIDAE), WITH
Results of Prof. E. Stromer's Research Expedition in the Deserts of Egypt
 Proceedings of the Royal Bavarian Academy of Science Mathematical-physical Division Volume XXVIII, Paper 3 Results of Prof. E. Stromer's Research Expedition in the Deserts of Egypt II. Vertebrate Remains
Proceedings of the Royal Bavarian Academy of Science Mathematical-physical Division Volume XXVIII, Paper 3 Results of Prof. E. Stromer's Research Expedition in the Deserts of Egypt II. Vertebrate Remains
Temporal lines. More forwardfacing. tubular orbits than in the African forms 3. Orbits larger relative to skull size than in the other genera 2.
 Asian lorises More forwardfacing and tubular orbits than in the African forms 3. Characterized by a marked extension of the ectotympanic into a tubular meatus and a more angular auditory bulla than in
Asian lorises More forwardfacing and tubular orbits than in the African forms 3. Characterized by a marked extension of the ectotympanic into a tubular meatus and a more angular auditory bulla than in
A REDESCRIPTION OF THE HOLOTYPE OF CALLIANASSA MUCRONATA STRAHL, 1861 (DECAPODA, THALASSINIDEA)
") Crustaceana 52 (1) 1977, E. J. Brill, Leiden A REDESCRIPTION OF THE HOLOTYPE OF CALLIANASSA MUCRONATA STRAHL, 1861 (DECAPODA, THALASSINIDEA) BY NASIMA M. TIRMIZI Department of Zoology, University of Karachi,
Crustaceana 52 (1) 1977, E. J. Brill, Leiden A REDESCRIPTION OF THE HOLOTYPE OF CALLIANASSA MUCRONATA STRAHL, 1861 (DECAPODA, THALASSINIDEA) BY NASIMA M. TIRMIZI Department of Zoology, University of Karachi,
REVISION OF THE GENUS MARTINICHTHYS, MARINE FISH (TELESOSTEI, TSELFATIIFORMES) FROM THE LATE CRETACEOUS OF KANSAS (UNITED STATES)
 FROM THE LATE CRETACEOUS OF KANSAS (UNITED STATES)") 1 REVISION OF THE GENUS MARTINICHTHYS, MARINE FISH (TELESOSTEI, TSELFATIIFORMES) FROM THE LATE CRETACEOUS OF KANSAS (UNITED STATES) TAVERNE L., 2000. Revision of the genus Martinichthys, marine fish (Teleostei,
1 REVISION OF THE GENUS MARTINICHTHYS, MARINE FISH (TELESOSTEI, TSELFATIIFORMES) FROM THE LATE CRETACEOUS OF KANSAS (UNITED STATES) TAVERNE L., 2000. Revision of the genus Martinichthys, marine fish (Teleostei,
A M E G H I N I A N A. Revista de la Asociación Paleontológia Argentina. Volume XV September-December 1978 Nos. 3-4
 A M E G H I N I A N A Revista de la Asociación Paleontológia Argentina Volume XV September-December 1978 Nos. 3-4 COLORADIA BREVIS N. G. ET N. SP. (SAURISCHIA, PROSAUROPODA), A PLATEOSAURID DINOSAUR FROM
A M E G H I N I A N A Revista de la Asociación Paleontológia Argentina Volume XV September-December 1978 Nos. 3-4 COLORADIA BREVIS N. G. ET N. SP. (SAURISCHIA, PROSAUROPODA), A PLATEOSAURID DINOSAUR FROM
On the cranial anatomy of the polycotylid plesiosaurs, including new material of Polycotylus latipinnis, Cope, from Alabama
 Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 2004 On the cranial anatomy of the polycotylid plesiosaurs, including new material of Polycotylus latipinnis,
Marshall University Marshall Digital Scholar Biological Sciences Faculty Research Biological Sciences 2004 On the cranial anatomy of the polycotylid plesiosaurs, including new material of Polycotylus latipinnis,
UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA
 FROM JAVA, INDONESIA") NOTES AND NEWS UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA BY NGUYEN NGOC-HO i) Faculty of Science, University of Saigon, Vietnam Among material recently collected
NOTES AND NEWS UPOGEBIA LINCOLNI SP. NOV. (DECAPODA, THALASSINIDEA, UPOGEBIIDAE) FROM JAVA, INDONESIA BY NGUYEN NGOC-HO i) Faculty of Science, University of Saigon, Vietnam Among material recently collected
520 McDowaLL, Bony Palate of Birds
 520 McDowaLL, Bony Palate of Birds LOct. [Auk MIRANDA RIBEIRO, PAULO DE 1929. Da nidifica o de Chaetura ½inereiventris. Bol. Mus. Nac. [Rio], 4: 101-105. RIDGWAY, R. 1911. The birds of North and Middle
520 McDowaLL, Bony Palate of Birds LOct. [Auk MIRANDA RIBEIRO, PAULO DE 1929. Da nidifica o de Chaetura ½inereiventris. Bol. Mus. Nac. [Rio], 4: 101-105. RIDGWAY, R. 1911. The birds of North and Middle