Europe s largest dinosaur? A giant brachiosaurid cervical vertebra from the Wessex Formation (Early Cretaceous) of southern England
|
|
|
- Georgia Cameron
- 5 years ago
- Views:
Transcription
1 Cretaceous Research 25 (2004) Europe s largest dinosaur? A giant brachiosaurid cervical vertebra from the Wessex Formation (Early Cretaceous) of southern England Darren Naish a, *, David M. Martill a, David Cooper b, Kent A. Stevens c a School of Earth and Environmental Sciences, University of Portsmouth, Portsmouth PO1 3QL, UK b Little Shate, Mill Lane, Brighstone, Newport, Isle of Wight PO30 4BU, UK c Department of Computer and Information Science, Deschutes Hall, University of Oregon, Eugene, OR 97403, USA Received 28 February 2002; accepted in revised form 14 July 2004 Abstract A single brachiosaurid sauropod cervical vertebra from the Wessex Formation (Barremian, Early Cretaceous) of the Isle of Wight is remarkable for its size. With a partial centrum length (i.e., excluding evidence of the anterior condyle) of 745 mm it represents the largest sauropod cervical reported from Europe and is close in size to cervical vertebrae of the giant brachiosaurid Brachiosaurus brancai from Late Jurassic Tanzania. The complete animal probably exceeded 20 m in total length. The specimen shares important morphological characters with Sauroposeidon proteles from Early Cretaceous USA, including extensive lateral fossae and well-developed posterior centroparapophyseal laminae, indicating that it is part of a Brachiosaurus Sauroposeidon clade, and in some characters is intermediate between the two. Owing to the complexities of Isle of Wight sauropod taxonomy the specimen is not attributed to a named taxon. Ó 2004 Elsevier Ltd. All rights reserved. Keywords: Dinosauria; Sauropoda; Brachiosauridae; Brachiosaurus; Sauroposeidon; Early Cretaceous; Isle of Wight; England 1. Introduction Sauropod dinosaurs, the long-necked giants of the Mesozoic, are well represented in the fossil record of Europe. Examples of most major sauropod groups are known from the European Triassic, Jurassic and Cretaceous, including basal sauropods, basal eusauropods, diplodocoids and titanosauriforms (McIntosh, 1990; Weishampel, 1990; Hunt et al., 1994; Upchurch, 1998; Godefroit and Knoll, 2003). The stratigraphic range of sauropods in Europe is extensive and ranges from the Rhaetian to the Maastrichtian (Hunt et al., 1994; Godefroit and Knoll, 2003). * Corresponding author. address: darren.naish@port.ac.uk (D. Naish). The Wessex Formation (Barremian, Early Cretaceous) of the Isle of Wight, southern England, is well known for its diverse dinosaur fauna (Benton and Spencer, 1995; Hutt et al., 2001; Martill and Naish, 2001), among which are several sauropod taxa (Naish and Martill, 2001). These include a possible diplodocoid and camarasaurid, several taxa traditionally regarded as brachiosaurids, a titanosaur and a number of indeterminate forms. Though as many as ten species-level names are attached to these remains, the holotypes are largely non-diagnostic and based on specimens that do not overlap with other UK Early Cretaceous sauropod holotypes (see Naish and Martill, 2001 for a review). Consequently, the majority of Isle of Wight sauropod genera and species are nomina dubia and none can presently be shown to be referable to the same species /$ - see front matter Ó 2004 Elsevier Ltd. All rights reserved. doi: /j.cretres
2 788 D. Naish et al. / Cretaceous Research 25 (2004) Isolated sauropod elements, though in some cases referable to higher clades, cannot be reliably referred to a genus or species. Here we report a large, isolated sauropod cervical vertebra collected from the foreshore in 1992 by Mr. Gavin Leng of the Isle of Wight. The specimen is notable for its size and also because it exhibits morphological features elsewhere seen only in the giant brachiosaurids Brachiosaurus brancai from the Upper Jurassic Tendaguru Formation of Tanzania (Janensch, 1950) and Sauroposeidon proteles from the Lower Cretaceous Antlers Formation of Oklahoma, USA (Wedel et al., 2000a,b). from the same locality and we suspect that a disarticulated skeleton is being eroded from the cliff over time. 3. Systematic palaeontology Dinosauria Owen, 1842 Saurischia Seeley, 1887 Sauropoda Marsh, 1878 Eusauropoda Upchurch, 1995 Titanosauriformes Salgado et al., 1997 Brachiosauridae Riggs, Locality and stratigraphy The new specimen, Isle of Wight Museum no. MIWG.7306, was found on the foreshore between Chilton Chine (Ordnance Survey grid reference ) and Sudmoor Point (OS grid ref ) (Fig. 1). The specimen was enclosed in matrix heavily overgrown by calcareous algae and other marine organisms. The matrix comprises a beige sideritic mudstone with fragments of plant debris and mud clasts and equates to plant debris bed L1 of Stewart (1978). This unit crops out in the cliffs at this locality directly above the Sudmoor Point Sandstone Member (SS2 of Stewart, 1978) of the Wessex Formation. A second, incomplete sauropod vertebra, IWCMS: , similar in overall form to MIWG.7306 but consisting only of an eroded centrum 640 mm long, has been collected Fig. 1. Sketch map of the Isle of Wight showing the locality at which the new specimen was discovered. Gen. et sp. indet. Figs. 2 4 Material. MIWG.7306, a cervical vertebra (possibly C6) 745 mm long. Relatively complete though crushed obliquely and missing most of the anterior condyle, the left prezygapophyis, the right diapophysis, most of the laminae associated with the neural arch and most of the left posterior centroparapophyseal lamina. A fragment of neural spine may be represented but cannot be attached to the rest of the specimen. The bone surface is highly brecciated and cemented by white calcite (Fig. 2A). The specimen has previously been figured by Martill and Naish (2001) and is currently on display at the Dinosaur Isle Visitor Centre, Sandown, Isle of Wight. Abbreviations and terminology. The description here follows Upchurch (1998) in use of the abbreviation EI. We employ Wilson s (1999) terminology for sauropod vertebral laminae and we also follow his system of abbreviations. Hence: EI, elongation index (total length of centrum divided by width of cotyle); Cpol, centropostzygapophyseal lamina; Pcdl, posterior centrodiapophyseal lamina; Pcpl, posterior centroparapophyseal lamina; Podl, postzygodiapophyseal lamina; Prdl, prezygodiapophyseal lamina; Tpol, intrapostzygapophyseal lamina. Description. MIWG.7306 is broken into two main pieces and examination of its interior shows that it is of camellate morphology. The internal cavities visible at the break are variable in size and shape and recall the cross-sectional view of a Brachiosaurus cervical centrum figured by Janensch (1947, fig. 71). The majority of cavities visible toward the dorsal surface of the centrum are taller than they are wide while the majority visible toward the ventral surface of the centrum are wider than they are tall. Most of the cavities are not measurable without ambiguity. Two measurable cavities located ventral to the level of the neural arch differ markedly in size. The smaller is 15 mm tall and 12 mm wide and the

3 D. Naish et al. / Cretaceous Research 25 (2004) Fig. 2. A, MIWG.7306 in right lateral view. B, right prezygapophysis in dorsal view. Note the reniform articulatory facet and the rounded depressions on the dorsal surface. For abbreviations, see text. Scale bar represents 100 mm. larger is 45 mm tall and 20 mm wide. The largest measurable cavity located close to the ventral surface of the centrum is 55 mm wide and 40 mm tall. The centrum is elongate with extensive, complex lateral fossae. The posterior articulatory face is concave and, although somewhat deformed, approximately 195 mm wide and 160 mm tall. Given that the total length is 745 mm, the EI sensu Upchurch (1998) is 3.8. In MIWG.7306, the centrum length divided by the height of the cotyle, a value used by Wilson and Sereno (1998), is 4.7. What remains of the anterior condyle in MIWG.7306 indicates that it was clearly bulbous and convex. The neural canal is subcircular, 50 mm wide and 40 mm tall. Posteriorly, and arising dorsolateral to the neural canal, distinct cpol extend to the ventromedial surface of the postzygapophyseal facet. A vertical strut extends dorsal to the neural canal to the base of the tpol which itself forms the ventral floor to the postspinous fossa (see Wilson, 1999, p. 647). On both sides, the cpol, tpol and vertical strut enclose deep, triangular peduncular fossae on the posterior surface of the postzygapophysis (Fig. 3). On the lateral side of the centrum there are a complex series of fossae. The most dorsal of these is a posteriorly located subtriangle pointing anteriorly (fossa 1 in
, bordered dorsally by the podl and ventrally by the pcdl.")
4 790 D. Naish et al. / Cretaceous Research 25 (2004) cpol 100 mm postzygapophysis triangular fossa tpol vertical strut neural canal Fig. 3. MIWG.7306 in posterior view. The arrows represent the original dorsoventral midline of the specimen. For abbreviations, see text. Fig. 2A; see also Fig. 4), bordered dorsally by the podl and ventrally by the pcdl. Similarly shaped and positioned dorsal fossae are seen in Sauroposeidon proteles and Brachiosaurus brancai (Janensch, 1950; Wedel et al., 2000a,b) (Fig. 5). This dorsal fossa is shallow compared to parts of the more ventral fossae and contains bony struts and smaller, shallow, rounded fossae on its medial bony wall. Ventral to the pcdl, another subtriangular fossa (fossa 2 in Fig. 2A) extends much further anteriorly than the dorsal fossa but does not extend as far posteriorly. Unlike the dorsal fossa, it widens anteriorly. An anteroventrally descending ridge ventral to this fossa then forms the dorsal border to a subrectangular ventral fossa. The ventral fossa (fossa 3 in Fig. 2A) is complex and descends ventrally towards its anterior end. On the left side, an oblique, posteroventrally directed bony strut (Fig. 4) divides the fossa into anterior and posterior portions and the fossa becomes shallower and pointed anteriorly. On the right side the dividing strut has an anteroventral inclination and the fossa is rectangular anteriorly. Ventral to the anterior part of the ventral fossa is a rectangular accessory fossa, bound dorsally and ventrally by prominent ridges. Within this fossa, ridges on the medial bony wall form a series of smaller rectangular fossae. There are two additional fossae on the anteroventral part of the centrum (labelled accessory ventral fossae in Fig. 2A), the most ventral of which excavates the lateral surface of the pcpl. As in B. brancai (Janensch, 1950), there is a small, shallow elliptical fossa on the anterior end of the ventral surface. There is no midline keel on the ventral surface of the centrum and the ventral surface of the centrum is gently concave. Beginning at a point ventral to the most ventral fossa on the lateral side of the centrum, the pcpl begin posteriorly as small ridges. On the right side, the pcpl originates further posteriorly than it does on the left side. This apparent asymmetry is a result of deformation due to compaction. Ventral to the anterior part of the ventral fossa, the pcpl expands ventrally to a marked degree, though on the left side the anteroventral part of the lamina is missing. The anteroventral extension of the pcpl of the right side is a very thin, sharp-edged plate of bone that is broken ventrally such that the parapophysis is missing (Fig. 2A). The right prezygapophysis is well preserved and broken from the rest of the vertebra. The prezygapophyseal facet is a flat oval, being 75 mm wide and 105 mm long, and with a rugose ridge marking the facet s ventral edge. On the medial surface just posterior to the prezygapophyseal facet there is a series of oval and rounded fossae. These do not appear to be features of the original osteology and may be of pathological origin or may represent bite marks or post-mortem borings (Fig. 2B). On the lateral side of the prezygapophysis, a prominent prdl extends posteriorly from the anterior end of the prezygapophysis. This lamina overhangs a concave area that is delimited ventrally by a sharp, posteroventrally curving ridge. The left prezygapophysis is missing. The postzygapophyseal facets are flat and reniform with the longest axis aligned mediolaterally (Fig. 4). There are several pneumatic fossae on the dorsal surface of the postzygapophyses, the number and position of which differ between the left and right postzygapophyses. What is interpreted here as the left diapophysis (it is unattached to the rest of the specimen) is missing its margins and is still attached to the proximal part of its cervical rib. The diapophysis is broad anteroposteriorly compared with those of B. brancai and S. proteles and, unlike the diapophyses of these species, possesses a rhomboidal fossa on the body of the diapophysis near its posterior margin. These features may be autapomorphies of whichever taxon MIWG.7306 belongs to. On its lateral side, the diapophysis is flat but on its medial side it bears a prominent median vertical lamina to which are connected a series of parallel subhorizontal bony bars. Cervical rib shafts are not preserved adjacent to the centrum but a fragment of cervical rib, 120 mm long, lies attached to the posterolateral surface of the centrum. This fragment has parallel dorsal and ventral margins

5 D. Naish et al. / Cretaceous Research 25 (2004) Fig. 4. Posterior part of MIWG.7306 in left lateral view showing (from top to bottom) fossa 1 and the posterior parts of fossae 2 and 3. The dividing strut of fossa 3 is visible at its anterior end. and is a mediolaterally compressed ovoid in crosssection. 4. Comparisons and phylogenetic affinities New phylogenetic schemes for Sauropoda have recently been proposed by Upchurch (1995, 1998), Calvo and Salgado (1995), Salgado et al. (1997), Wilson and Sereno (1998) and Wilson (2002). While the position of some sauropod taxa remains controversial, there is now a broad consensus on the affinities of all major sauropod clades. We examined MIWG.7306 within the larger context of sauropod phylogeny. The marked opisthocoely, high length:width ratio (3.8:1), extensive lateral fossae and complex system of bony laminae of MIWG.7306 demonstrate its sauropod identity. The presence of extensive invasive pneumatic depressions on the sides of the centrum, and of bony septa dividing some of these depressions, indicate that it is a member of the Omeisaurus C Neosauropoda clade of Wilson and Sereno (1998). Furthermore, because the laminae associated with the neural arch do not appear to be rudimentary in MIWG.7306, it is excluded from Wilson and Sereno s (1998) clade Somphospondyli (Euhelopus C Titanosauria). The presence in MIWG.7306 of camellate internal morphology invite comparison with Mamenchisaurus and members of Titanosauriformes, the only sauropods to exhibit this morphology (Upchurch, 1998; Wedel et al., 2000b; Wilson 2002). Furthermore, the centrum length/cotyle height value of 4.7 in MIWG.7306 is informative as values higher than 4 are only seen in Omeisaurus, Mamenchisaurus, some diplodocoids and
6
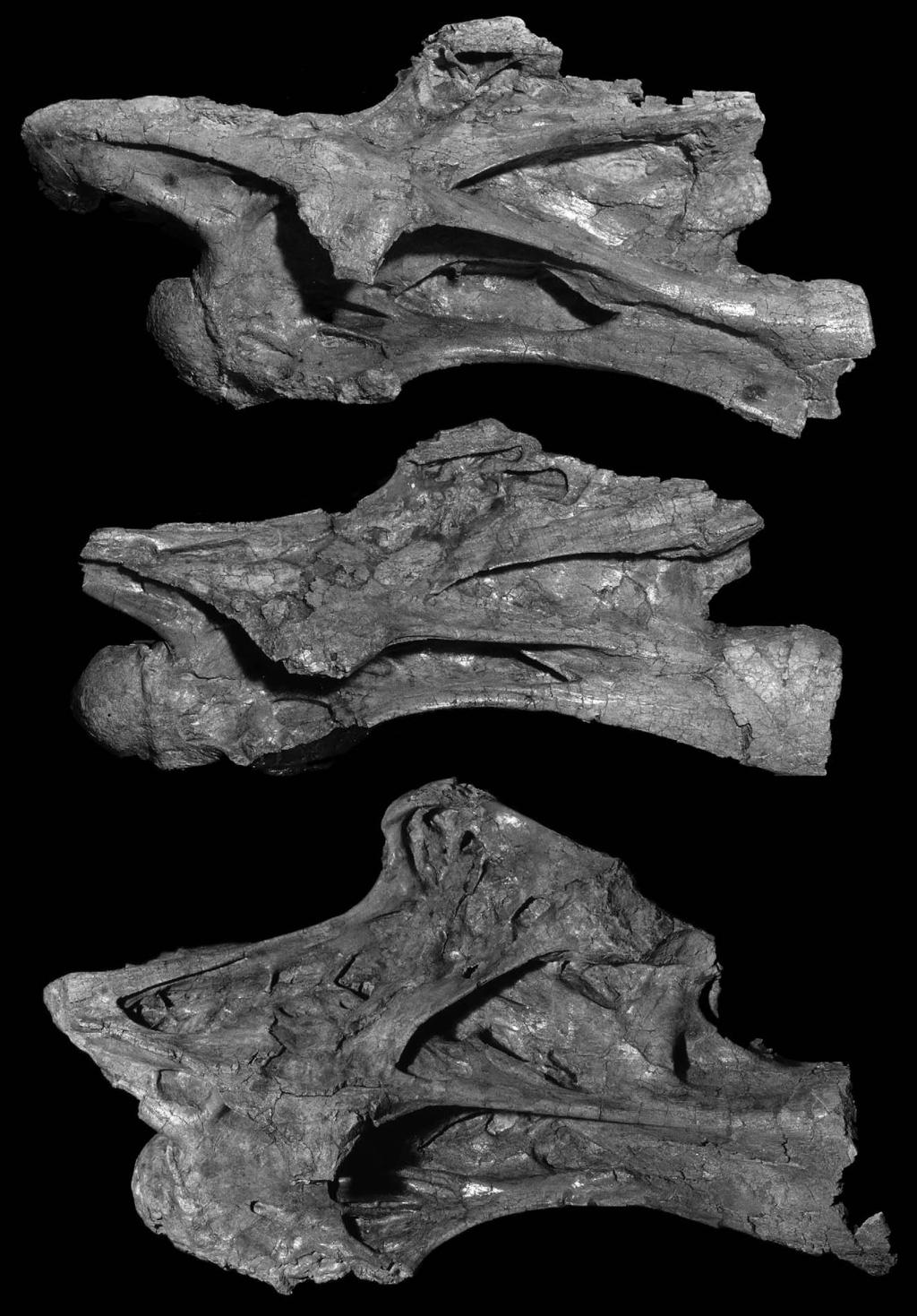
7 D. Naish et al. / Cretaceous Research 25 (2004) Brachiosauridae (Wilson, 2002). Combined with the distribution of characters outlined above, the presence of an elongate centrum and deep lateral depressions in MIWG.7306 indicate that it is from a brachiosaurid sensu Wilson and Sereno (1998). Possible affinities with the other taxa mentioned here can be excluded on the basis of the absence of camellate morphology (Omeisaurus and diplodocoids) or absence of deep lateral cavities (Mamenchisaurus). Though monophyly of Brachiosauridae is controversial (see below), Wilson and Sereno (1998) regarded the group as a clade characterised by a subrectangular muzzle, humerus subequal in length to the femur and with a prominent deltopectoral crest, and elongate cervical centra with deep accessory depressions. Salgado et al. (1997) argued that Brachiosauridae and Brachiosaurus were paraphyletic and that B. brancai was closer to Titanosauria than to B. altithorax. However, Brachiosaurus shares a number of characters with S. proteles including elongate cervical centra and ribs, prezygapophyses extensively overhanging the condyle, and a transition point in the neural spine height of the mid-cervical vertebrae (Wedel et al., 2000a,b). As these shared characters are not seen in other titanosauriforms we recognise a restricted monophyletic Brachiosauridae for Brachiosaurus and S. proteles. It may be that other taxa traditionally regarded as brachiosaurids, including Cedarosaurus, Eucamerotus, Ornithopsis, Pelorosaurus and Sonorasaurus, also belong to this clade (McIntosh, 1990; Blows, 1995; Tidwell et al., 1999; Naish and Martill, 2001). Notably, MIWG.7306 is reminiscent of S. proteles in exhibiting pcpl that begin in a relatively posteroventral position on the centrum, though they are not as hypertrophied as those of S. proteles. In this taxon the pcpl are evident as distinct ventrolateral laminae even at the posterior cotyle (Wedel et al., 2000b). These hypertrophied pcpl appear to be an autapomorphy of S. proteles as they are not present in other sauropods, including B. brancai (though Wilson, 2002 regarded S. proteles as referable to Brachiosaurus). In MIWG.7306, the pcpl begin ventral to the posterior half of the centrum and it therefore appears intermediate between other sauropods (including B. brancai) and S. proteles. Significantly, the lateral fossae of MIWG.7306 occupy more of the centrum s surface area than do those of B. brancai or other titanosauriforms (excepting S. proteles). We estimate that ca. 40% of the lateral surface of C6 in B. brancai is excavated by pneumatic depressions. In S. proteles, this value for C6 is ca. 80% or more and in MIWG.7306 it is between 50 and 60%. These values were obtained by drawing the vertebrae of these dinosaurs on graph paper and estimating the total area of the lateral side of the centrum occupied by pneumatic fossae. The calculation of these values is somewhat subjective. However, only in S. proteles is the area occupied by lateral fossae more extensive than that in MIWG We therefore interpret MIWG.7306 as intermediate in this character between B. brancai and S. proteles. We suggest that pcpl which originate ventral to the posterior half of the centrum and extensive lateral excavations that occupy more than ca. 50% of the lateral surface of the centrum are derived characters that unite MIWG.7306 and S. proteles within a restricted Brachiosauridae (Fig. 6). 5. Discussion Determining the position in the cervical series of MIWG.7306 is difficult because of its incompleteness but in shape and the position of the parapophyses it appears to correspond most closely with C6 of S. proteles and B. brancai. It therefore may not have been the longest vertebra in the series, or the centrum with the highest EI. The presence of possible autapomorphies in MIWG.7306 (an anteroposteriorly broad diapophysis and a rhomboidal fossa near the posterior margin of the diapophysis) indicates that it represents a diagnosable taxon. It is likely, however, that the specimen is referable to one of the named Isle of Wight sauropod taxa, though this cannot be demonstrated at present due to the absence of overlapping material. A cervical vertebra from the Lower Cretaceous of Croatia is similar in several important respects to MIWG.7306 and probably represent a closely related taxon (Dalla Vecchia, 1998), albeit one that did not reach an adult body size comparable with that probably achieved by MIWG.7306 (the centrum of the Croatian vertebra is 350 mm and closure of the neurocentral sutures indicates that its owner was adult). With a centrum length of 745 mm MIWG.7306 is the largest sauropod cervical vertebra reported from Europe, and one of the biggest known. In B. brancai the longest centra in the cervical series (C10 and C11 of HM SII) are 870 mm while C9 is 850 mm, C8 is 860 mm, C7 is 820 mm and C6 is 780 mm (Janensch, 1950). In S. proteles, the longest vertebra (C8) has a centrum length of 1250 mm while C6 has a centrum length of 1220 mm (Wedel et al., 2000b). B. brancai has been estimated to have obtained a total length of 25 m (Paul, 1988). S. proteles was presumably somewhat larger, though as noted by Wedel et al. (2000a,b) it is possible Fig. 5. Cervical vertebrae 5, 6 and 7 (from top to bottom) of Brachiosaurus brancai specimen SI for comparison. MIWG.7306 is not identical to any of these vertebrae but is closest in proportions and morphological details to C6. Combined centrum and anterior condyle lengths are as follows: C5, 560 mm, C6, 691 mm, C7, 705 mm. Photograph courtesy of Prof. Chris McGowan.
8 794 D. Naish et al. / Cretaceous Research 25 (2004) Camarasaurus Brachiosaurus brancai that it was similar in size, or perhaps smaller. Similarly, MIWG.7306 may have differed in proportions from B. brancai and thus any estimate of its total length is speculative. We suggest, however, that the live animal exceeded 20 m in total length and may have been comparable in size to B. brancai. Other titanosauriform elements from the Wessex Formation suggesting the presence of B. brancai-sized giants, though rare, are present. The tooth BMNH R964, the holotype of Oplosaurus armatus (see Naish and Martill, 2001), is 85 mm long (this length includes only the base of the root) and comparable in size to B. brancai teeth. However, Canudo et al. (2002) have recently suggested that this tooth may belong to a sauropod more closely related to Camarasaurus than Brachiosaurus. While the majority of Isle of Wight titanosauriforms were medium-sized, reaching approximately 15 m in length (Naish and Martill, 2001), MIWG.7306 is evidence that at least some of Europe s Early Cretaceous sauropods were giants and among the largest of known dinosaurs. Acknowledgements MIWG.7306 A Sauroposeidon proteles Fig. 6. Proposed phylogenetic relationships of MIWG.7306 to other sauropods. Group A is characterised by the presence of centrum length/cotyle height value O 4 and prezygapophyses extensively overhanging condyle. Group B is characterised by the presence of hypertrophied posterior centroparapophyseal laminae that originate ventral to the posterior half of the centrum and O ca. 50% of lateral surface of centrum excavated by pneumatic depressions. We thank Dr. Mike Bishop of Isle of Wight Leisure Services for allowing us to work on the material, Steve Hutt, Martin Munt and Dan Pemberton (Dinosaur Isle) for curatorial assistance, Bob Loveridge for assistance with photography and Gavin Leng for kindly donating the specimen to the Dinosaur Isle Visitor Centre. Comments by Dr. Paul Upchurch improved the manuscript, Prof. Chris McGowan is thanked for allowing use of Fig. 5, and Dr. Paul Barrett is thanked for discussions on Wealden sauropod taxonomy. We are especially grateful to Matthew Wedel for extensive discussion, encouragement, advice and data on Sauroposeidon, and for helpful comments on the manuscript. B Mike P. Taylor is also thanked for his encouragement on this much-delayed project. References Benton, M.J., Spencer, P.S., Fossil Reptiles of Great Britain. Chapman & Hall, London, 386 pp. Blows, W.T., The early Cretaceous brachiosaurid dinosaurs Ornithopsis and Eucamerotus from the Isle of Wight, England. Palaeontology 38, Calvo, J.O., Salgado, L., Rebbachisaurus tessonei sp. nov. A new Sauropoda from the Albian Cenomanian of Argentina; new evidence on the origin of the Diplodocidae. Gaia 11, Canudo, J.I., Ruiz-Omeñaca, J.I., Barco, J.L., Royo Torres, R., Sauro podos asiáticos en el Barremiense inferior (Cretácico Inferior) de España? Ameghiniana 39, Dalla Vecchia, F.M., Remains of Sauropoda (Reptilia, Saurischia) in the Lower Cretaceous (upper Hauterivian/lower Barremian) limestones of SW Istria (Croatia). Geologia Croatica 51, Godefroit, P., Knoll, F., Late Triassic dinosaur teeth from southern Belgium. Comptes Rendus Palevol 2, Hunt, A.P., Lockley, M.G., Lucas, S.G., Meyer, C.A., The global sauropod fossil record. Gaia 10, Hutt, S., Naish, D., Martill, D.M., Barker, M.J., Newbery, P., A preliminary account of a new tyrannosauroid theropod from the Wessex Formation (Early Cretaceous) of southern England. Cretaceous Research 22, Janensch, W., Pneumatizita t bei Wirbeln von Sauropoden und anderen Saurischiern. Palaeontographica 3 (Supplement 7), Janensch, W., Die Wirbelsaule von Brachiosaurus brancai. Palaeontographica 3 (Supplement 7), Martill, D.M., Naish, D. (Eds.), Dinosaurs of the Isle of Wight. In: Field Guides to Fossils, 10. The Palaeontological Association, London, 433 pp. McIntosh, J.S., Sauropoda. In: Weishampel, D.B., Dodson, P., Osmo lska, H. (Eds.), The Dinosauria. University of California Press, Berkeley, pp Naish, D., Martill, D.M., Saurischian dinosaurs 1: Sauropods. In: Martill, D.M., Naish, D. (Eds.), Dinosaurs of the Isle of Wight. Field Guides to Fossils 10, The Palaeontological Association, London, pp Paul, G.S., The brachiosaur giants of the Morrison and Tendaguru with a description of a new subgenus, Giraffatitan, and a comparison of the world s largest dinosaurs. Hunteria 2 (3), Salgado, L., Coria, R.A., Calvo, J.O., Evolution of the titanosaurid sauropods I: Phylogenetic analysis based on the postcranial evidence. Ameghiniana 34, Stewart, D.J., The sedimentology and palaeoenvironment of the Wealden Group of the Isle of Wight, southern England. Unpublished PhD thesis, University of Portsmouth, 347 pp. Tidwell, V., Carpenter, K., Brooks, W., New sauropod from the Lower Cretaceous of Utah. Oryctos 2, Upchurch, P., The evolutionary history of sauropod dinosaurs. Philosophical Transactions of the Royal Society of London, Series B 349, Upchurch, P., The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society 124, Wedel, M.J., Cifelli, R.L., Sanders, R.K., 2000a. Sauroposeidon proteles, a new sauropod from the early Cretaceous of Oklahoma. Journal of Vertebrate Paleontology 20, Wedel, M.J., Cifelli, R.L., Sanders, R.K., 2000b. Osteology, paleobiology, and relationships of the sauropod dinosaur Sauroposeidon proteles. Acta Palaeontologica Polonica 45,
9 D. Naish et al. / Cretaceous Research 25 (2004) Weishampel, D.B., Dinosaurian distribution. In: Weishampel, D.B., Dodson, P., Osmo lska, H. (Eds.), The Dinosauria. University of California Press, Berkeley, pp Wilson, J.A., A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology 19, Wilson, J.A., Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136, Wilson, J.A., Sereno, P.C., Early evolution and higher-level phylogeny of sauropod dinosaurs. Journal of Vertebrate Paleontology, Memoir 5, 1 68.
NEW INFORMATION ON A JUVENILE SAUROPOD SPECIMEN FROM THE MORRISON FORMATION AND THE REASSESSMENT OF ITS SYSTEMATIC POSITION
 [Palaeontology, Vol. 55, Part 3, 2012, pp. 567 582] NEW INFORMATION ON A JUVENILE SAUROPOD SPECIMEN FROM THE MORRISON FORMATION AND THE REASSESSMENT OF ITS SYSTEMATIC POSITION by JOSÉ L. CARBALLIDO 1,
[Palaeontology, Vol. 55, Part 3, 2012, pp. 567 582] NEW INFORMATION ON A JUVENILE SAUROPOD SPECIMEN FROM THE MORRISON FORMATION AND THE REASSESSMENT OF ITS SYSTEMATIC POSITION by JOSÉ L. CARBALLIDO 1,
A NEW TITANOSAURIFORM SAUROPOD (DINOSAURIA: SAURISCHIA) FROM THE EARLY CRETACEOUS OF CENTRAL TEXAS AND ITS PHYLOGENETIC RELATIONSHIPS. Peter J.
 FROM THE EARLY CRETACEOUS OF CENTRAL TEXAS AND ITS PHYLOGENETIC RELATIONSHIPS. Peter J.") Palaeontologia Electronica http://palaeo-electronica.org A NEW TITANOSAURIFORM SAUROPOD (DINOSAURIA: SAURISCHIA) FROM THE EARLY CRETACEOUS OF CENTRAL TEXAS AND ITS PHYLOGENETIC RELATIONSHIPS Peter J. Rose
Palaeontologia Electronica http://palaeo-electronica.org A NEW TITANOSAURIFORM SAUROPOD (DINOSAURIA: SAURISCHIA) FROM THE EARLY CRETACEOUS OF CENTRAL TEXAS AND ITS PHYLOGENETIC RELATIONSHIPS Peter J. Rose
NEW SAUROPOD FROM THE LOWER CRETACEOUS OF UTAH, USA
 ORYCTOS, Vol. 2 : 21-37, Décembre 1999 NEW SAUROPOD FROM THE LOWER CRETACEOUS OF UTAH, USA Virginia TIDWELL, Kenneth CARPENTER and William BROOKS Department of Earth and Space Sciences, Denver Museum of
ORYCTOS, Vol. 2 : 21-37, Décembre 1999 NEW SAUROPOD FROM THE LOWER CRETACEOUS OF UTAH, USA Virginia TIDWELL, Kenneth CARPENTER and William BROOKS Department of Earth and Space Sciences, Denver Museum of
A new species of sauropod, Mamenchisaurus anyuensis sp. nov.
 A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
Xenoposeidon is the earliest known rebbachisaurid sauropod dinosaur
 Xenoposeidon is the earliest known rebbachisaurid sauropod dinosaur Michael Taylor Corresp. 1 1 Department of Earth Sciences, University of Bristol Corresponding Author: Michael Taylor Email address: dino@miketaylor.org.uk
Xenoposeidon is the earliest known rebbachisaurid sauropod dinosaur Michael Taylor Corresp. 1 1 Department of Earth Sciences, University of Bristol Corresponding Author: Michael Taylor Email address: dino@miketaylor.org.uk
A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China
 SUPPLEMENTARY INFORMATION A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China Ya-Ming Wang 1, Hai-Lu You 2,3 *, Tao Wang 4 1 School of Earth Sciences and Resources, China
SUPPLEMENTARY INFORMATION A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China Ya-Ming Wang 1, Hai-Lu You 2,3 *, Tao Wang 4 1 School of Earth Sciences and Resources, China
FIELDIANA GEOLOGY NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA
 FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 Sbftember 22, 1968 No. 88 NEW SALAMANDERS OF THE FAMILY SIRENIDAE FROM THE CRETACEOUS OF NORTH AMERICA Coleman J. Coin AND Walter
AMERICAN MUSEUM NOVITATES
 AMERICAN MUSEUM NOVITATES Number 3700 October 22, 2010 The Illusory Evidence for Asian Brachiosauridae: New Material of Erketu ellisoni and a Phylogenetic Reappraisal of Basal Titanosauriformes DANIEL
AMERICAN MUSEUM NOVITATES Number 3700 October 22, 2010 The Illusory Evidence for Asian Brachiosauridae: New Material of Erketu ellisoni and a Phylogenetic Reappraisal of Basal Titanosauriformes DANIEL
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
The early evolution of titanosauriform sauropod dinosaurs
 bs_bs_banner Zoological Journal of the Linnean Society, 2012, 166, 624 671. With 8 figures The early evolution of titanosauriform sauropod dinosaurs MICHAEL D. D EMIC* Museum of Paleontology and Department
bs_bs_banner Zoological Journal of the Linnean Society, 2012, 166, 624 671. With 8 figures The early evolution of titanosauriform sauropod dinosaurs MICHAEL D. D EMIC* Museum of Paleontology and Department
6BT, UK b Museum für Naturkunde, Invalidenstrasse 43, 10115, Berlin, Germany
 This article was downloaded by: [University College London] On: 02 August 2012, At: 03:36 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office:
This article was downloaded by: [University College London] On: 02 August 2012, At: 03:36 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office:
ABSTRACT. Candice M. Stefanic and Sterling J. Nesbitt
 The axial skeleton of Poposaurus langstoni (Pseudosuchia: Poposauroidea) and its implications for accessory intervertebral articulation evolution in pseudosuchian archosaurs Candice M. Stefanic and Sterling
The axial skeleton of Poposaurus langstoni (Pseudosuchia: Poposauroidea) and its implications for accessory intervertebral articulation evolution in pseudosuchian archosaurs Candice M. Stefanic and Sterling
Journal of Systematic Palaeontology. ISSN: (Print) (Online) Journal homepage:
 (Online) Journal homepage:") Journal of Systematic Palaeontology ISSN: 1477-2019 (Print) 1478-0941 (Online) Journal homepage: http://www.tandfonline.com/loi/tjsp20 An articulated cervical series of Alamosaurus sanjuanensis Gilmore,
Journal of Systematic Palaeontology ISSN: 1477-2019 (Print) 1478-0941 (Online) Journal homepage: http://www.tandfonline.com/loi/tjsp20 An articulated cervical series of Alamosaurus sanjuanensis Gilmore,
Feruglio, Fontana 140, Trelew, Argentina Version of record first published: 25 Mar 2013.
 This article was downloaded by: [American Museum of Natural History] On: 25 March 2013, At: 05:07 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered
This article was downloaded by: [American Museum of Natural History] On: 25 March 2013, At: 05:07 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered
Osteology of the dorsal vertebrae of the giant titanosaurian sauropod dinosaur Dreadnoughtus schrani from the Late Cretaceous of Argentina
 Rowan University Rowan Digital Works School of Earth & Environment Faculty Scholarship School of Earth & Environment 1-1-2017 Osteology of the dorsal vertebrae of the giant titanosaurian sauropod dinosaur
Rowan University Rowan Digital Works School of Earth & Environment Faculty Scholarship School of Earth & Environment 1-1-2017 Osteology of the dorsal vertebrae of the giant titanosaurian sauropod dinosaur
A Nomenclature for Vertebral Fossae in Sauropods and Other Saurischian Dinosaurs
 A Nomenclature for Vertebral Fossae in Sauropods and Other Saurischian Dinosaurs Jeffrey A. Wilson*, Michael D. D Emic, Takehito Ikejiri, Emile M. Moacdieh, John A. Whitlock Museum of Paleontology and
A Nomenclature for Vertebral Fossae in Sauropods and Other Saurischian Dinosaurs Jeffrey A. Wilson*, Michael D. D Emic, Takehito Ikejiri, Emile M. Moacdieh, John A. Whitlock Museum of Paleontology and
Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms
 and the evolutionary history of basal titanosauriforms") bs_bs_banner Zoological Journal of the Linnean Society, 2013, 168, 98 206. With 30 figures Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary
bs_bs_banner Zoological Journal of the Linnean Society, 2013, 168, 98 206. With 30 figures Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary
Article. Universidade de Brasília - Faculdade UnB Planaltina, Brasília-DF, , Brazil. 2
 Zootaxa 3085: 1 33 (2011) www.mapress.com/zootaxa/ Copyright 2011 Magnolia Press Article ISSN 1175-5326 (print edition) ZOOTAXA ISSN 1175-5334 (online edition) A new sauropod (Macronaria, Titanosauria)
Zootaxa 3085: 1 33 (2011) www.mapress.com/zootaxa/ Copyright 2011 Magnolia Press Article ISSN 1175-5326 (print edition) ZOOTAXA ISSN 1175-5334 (online edition) A new sauropod (Macronaria, Titanosauria)
THE ANATOMY AND TAXONOMY OF CETIOSAURUS (SAURISCHIA, SAUROPODA) FROM THE MIDDLE JURASSIC OF ENGLAND
 FROM THE MIDDLE JURASSIC OF ENGLAND") Journal of Vertebrate Paleontology 23(1):208 231, March 2003 2003 by the Society of Vertebrate Paleontology THE ANATOMY AND TAXONOMY OF CETIOSAURUS (SAURISCHIA, SAUROPODA) FROM THE MIDDLE JURASSIC OF ENGLAND
Journal of Vertebrate Paleontology 23(1):208 231, March 2003 2003 by the Society of Vertebrate Paleontology THE ANATOMY AND TAXONOMY OF CETIOSAURUS (SAURISCHIA, SAUROPODA) FROM THE MIDDLE JURASSIC OF ENGLAND
A New Titanosaurian Sauropod from the Hekou Group (Lower Cretaceous) of the Lanzhou-Minhe Basin, Gansu Province, China
 of the Lanzhou-Minhe Basin, Gansu Province, China") A New Titanosaurian Sauropod from the Hekou Group (Lower Cretaceous) of the Lanzhou-Minhe Basin, Gansu Province, China Li-Guo Li 1,2 *, Da-Qing Li 3, Hai-Lu You 4, Peter Dodson 2 1 School of Earth Sciences
A New Titanosaurian Sauropod from the Hekou Group (Lower Cretaceous) of the Lanzhou-Minhe Basin, Gansu Province, China Li-Guo Li 1,2 *, Da-Qing Li 3, Hai-Lu You 4, Peter Dodson 2 1 School of Earth Sciences
A revision of Sanpasaurus yaoi Young, 1944 from the Early Jurassic of China, and its relevance to the early evolution of Sauropoda (Dinosauria)
") A revision of Sanpasaurus yaoi Young, 1944 from the Early Jurassic of China, and its relevance to the early evolution of Sauropoda (Dinosauria) Blair W. McPhee 1,2, Paul Upchurch 3, Philip D. Mannion 4,
A revision of Sanpasaurus yaoi Young, 1944 from the Early Jurassic of China, and its relevance to the early evolution of Sauropoda (Dinosauria) Blair W. McPhee 1,2, Paul Upchurch 3, Philip D. Mannion 4,
Nomenclature of Vertebral Laminae in Lizards, with Comments on Ontogenetic and Serial Variation in Lacertini (Squamata, Lacertidae)
") RESEARCH ARTICLE Nomenclature of Vertebral Laminae in Lizards, with Comments on Ontogenetic and Serial Variation in Lacertini (Squamata, Lacertidae) Emanuel Tschopp 1,2,3 * 1 Dipartimento di Scienze della
RESEARCH ARTICLE Nomenclature of Vertebral Laminae in Lizards, with Comments on Ontogenetic and Serial Variation in Lacertini (Squamata, Lacertidae) Emanuel Tschopp 1,2,3 * 1 Dipartimento di Scienze della
.56 m. (22 in.). COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition
. COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition") Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
SAUROPOD DINOSAURS FROM THE EARLY CRETACEOUS OF MALAWI, AFRICA. Elizabeth M. Gomani
 Palaeontologia Electronica http://palaeo-electronica.org SAUROPOD DINOSAURS FROM THE EARLY CRETACEOUS OF MALAWI, AFRICA Elizabeth M. Gomani ABSTRACT At least two titanosaurian sauropod taxa have been discovered
Palaeontologia Electronica http://palaeo-electronica.org SAUROPOD DINOSAURS FROM THE EARLY CRETACEOUS OF MALAWI, AFRICA Elizabeth M. Gomani ABSTRACT At least two titanosaurian sauropod taxa have been discovered
Cretaceous Research 34 (2012) 220e232. Contents lists available at SciVerse ScienceDirect. Cretaceous Research
 220e232. Contents lists available at SciVerse ScienceDirect. Cretaceous Research") Cretaceous Research 34 (2012) 220e232 Contents lists available at SciVerse ScienceDirect Cretaceous Research journal homepage: www.elsevier.com/locate/cretres The southernmost records of Rebbachisauridae
Cretaceous Research 34 (2012) 220e232 Contents lists available at SciVerse ScienceDirect Cretaceous Research journal homepage: www.elsevier.com/locate/cretres The southernmost records of Rebbachisauridae
Europatitan eastwoodi, a new sauropod from the lower Cretaceous of Iberia in the initial radiation of somphospondylans in Laurasia
 Europatitan eastwoodi, a new sauropod from the lower Cretaceous of Iberia in the initial radiation of somphospondylans in Laurasia Fidel Torcida Fernández-Baldor 1,2, José Ignacio Canudo 3,4, Pedro Huerta
Europatitan eastwoodi, a new sauropod from the lower Cretaceous of Iberia in the initial radiation of somphospondylans in Laurasia Fidel Torcida Fernández-Baldor 1,2, José Ignacio Canudo 3,4, Pedro Huerta
ON SOME REPTILIAN REMAINS FROM THE DINOSAUR BEDS OF NYASALAND. By S. H. HAUGHTON, D.Sc., F.G.S.
 ( 67 ) ON SOME REPTILIAN REMAINS FROM THE DINOSAUR BEDS OF NYASALAND. By S. H. HAUGHTON, D.Sc., F.G.S. (Published by permission of the Hon. the Minister for Mines and Industries.) (With Plates II-V and
( 67 ) ON SOME REPTILIAN REMAINS FROM THE DINOSAUR BEDS OF NYASALAND. By S. H. HAUGHTON, D.Sc., F.G.S. (Published by permission of the Hon. the Minister for Mines and Industries.) (With Plates II-V and
290 SHUFELDT, Remains of Hesperornis.
 290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
Postcranial Skeletal Pneumaticity in Sauropods and Its Implications for Mass Estimates
 SEVEN Postcranial Skeletal Pneumaticity in Sauropods and Its Implications for Mass Estimates Mathew J. Wedel O ne of the signal features of sauropods, and one of the cornerstones of our fascination with
SEVEN Postcranial Skeletal Pneumaticity in Sauropods and Its Implications for Mass Estimates Mathew J. Wedel O ne of the signal features of sauropods, and one of the cornerstones of our fascination with
Considerations of the neural laminae of sauropod dinosaurs and their morphofunctional meaning *
 Considerations of the neural laminae of sauropod dinosaurs and their morphofunctional meaning * Leonardo SALGADO, Rodolfo A. GARCÍA, & Juan D. DAZA Translated by Michael D. D Emic & Ariel Schepers; edited
Considerations of the neural laminae of sauropod dinosaurs and their morphofunctional meaning * Leonardo SALGADO, Rodolfo A. GARCÍA, & Juan D. DAZA Translated by Michael D. D Emic & Ariel Schepers; edited
New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia
 1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
A new Middle Jurassic sauropod subfamily (Klamelisaurinae subfam. nov.) from Xinjiang Autonomous Region, China
 from Xinjiang Autonomous Region, China") A new Middle Jurassic sauropod subfamily (Klamelisaurinae subfam. nov.) from Xinjiang Autonomous Region, China by Xijing Zhao Institute of Vertebrate Paleontology and Paleoanthropology, Academia Sinica
A new Middle Jurassic sauropod subfamily (Klamelisaurinae subfam. nov.) from Xinjiang Autonomous Region, China by Xijing Zhao Institute of Vertebrate Paleontology and Paleoanthropology, Academia Sinica
DINOSAUR DIVERSITY ANALYSED BY CLADE, AGE, PLACE AND YEAR OF DESCRIPTION
 DINOSAUR DIVERSITY ANALYSED BY CLADE, AGE, PLACE AND YEAR OF DESCRIPTION by MICHAEL P. TAYLOR School of Earth and Environmental Sciences, University of Portsmouth, Portsmouth PO1 3QL, UK (dino@miketaylor.org.uk)
DINOSAUR DIVERSITY ANALYSED BY CLADE, AGE, PLACE AND YEAR OF DESCRIPTION by MICHAEL P. TAYLOR School of Earth and Environmental Sciences, University of Portsmouth, Portsmouth PO1 3QL, UK (dino@miketaylor.org.uk)
The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda)
") PaleoBios 25(2):1 7, September 15, 2005 2005 University of California Museum of Paleontology The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda) MICHAEL P. TAYLOR and DARREN NAISH School
PaleoBios 25(2):1 7, September 15, 2005 2005 University of California Museum of Paleontology The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda) MICHAEL P. TAYLOR and DARREN NAISH School
Origin and Evolution of Birds. Read: Chapters 1-3 in Gill but limited review of systematics
 Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
NEW YUNNANOSAURID DINOSAUR (DINOSAURIA, PROSAUROPODA) FROM THE MIDDLE JURASSIC ZHANGHE FORMATION OF YUANMOU, YUNNAN PROVINCE OF CHINA
 FROM THE MIDDLE JURASSIC ZHANGHE FORMATION OF YUANMOU, YUNNAN PROVINCE OF CHINA") Memoir of the Fukui Prefectural Dinosaur Museum 6: 1 15 (2007) by the Fukui Prefectural Dinosaur Museum NEW YUNNANOSAURID DINOSAUR (DINOSAURIA, PROSAUROPODA) FROM THE MIDDLE JURASSIC ZHANGHE FORMATION
Memoir of the Fukui Prefectural Dinosaur Museum 6: 1 15 (2007) by the Fukui Prefectural Dinosaur Museum NEW YUNNANOSAURID DINOSAUR (DINOSAURIA, PROSAUROPODA) FROM THE MIDDLE JURASSIC ZHANGHE FORMATION
A New Sauropod Dinosaur From the Early Cretaceous of Oklahoma. Mathew J. Wedel. Oklahoma Museum of Natural History. and. Department of Zoology
 ..., ' A New Sauropod Dinosaur From the Early Cretaceous of Oklahoma by Mathew. Wedel Oklahoma Museum of Natural History " and Department of Zoology University of Oklahoma 1335 Asp Avenue Norman, OK 73019-0606
..., ' A New Sauropod Dinosaur From the Early Cretaceous of Oklahoma by Mathew. Wedel Oklahoma Museum of Natural History " and Department of Zoology University of Oklahoma 1335 Asp Avenue Norman, OK 73019-0606
Tel (0) David M. Martill. Corresponding Author
 David M. Martill. Corresponding Author") First occurrence of the pterosaur Coloborhynchus (Pterosauria, Ornithocheiridae) from the Wessex Formation (Lower Cretaceous) of the Isle of Wight, England Corresponding Author David M. Martill School
First occurrence of the pterosaur Coloborhynchus (Pterosauria, Ornithocheiridae) from the Wessex Formation (Lower Cretaceous) of the Isle of Wight, England Corresponding Author David M. Martill School
A turiasaurian sauropod dinosaur from the Early Cretaceous Wealden Supergroup of the United Kingdom
 A turiasaurian sauropod dinosaur from the Early Cretaceous Wealden Supergroup of the United Kingdom Philip D. Mannion Department of Earth Science and Engineering, Imperial College London, London, UK Submitted
A turiasaurian sauropod dinosaur from the Early Cretaceous Wealden Supergroup of the United Kingdom Philip D. Mannion Department of Earth Science and Engineering, Imperial College London, London, UK Submitted
Haplocanthosaurus (Saurischia: Sauropoda) from the lower Morrison Formation (Upper Jurassic) near Snowmass, Colorado
 from the lower Morrison Formation (Upper Jurassic) near Snowmass, Colorado") Volumina Jurassica, 2014, XII (2): 197 210 DOI: 10.5604/17313708.1130144 Haplocanthosaurus (Saurischia: Sauropoda) from the lower Morrison Formation (Upper Jurassic) near Snowmass, Colorado John R. FosteR
Volumina Jurassica, 2014, XII (2): 197 210 DOI: 10.5604/17313708.1130144 Haplocanthosaurus (Saurischia: Sauropoda) from the lower Morrison Formation (Upper Jurassic) near Snowmass, Colorado John R. FosteR
Demandasaurus darwini, a New Rebbachisaurid Sauropod from the Early Cretaceous of the Iberian Peninsula
 Demandasaurus darwini, a New Rebbachisaurid Sauropod from the Early Cretaceous of the Iberian Peninsula Author(s): Fidel Torcida Fernández-Baldor, José Ignacio Canudo, Pedro Huerta, Diego Montero, Xabier
Demandasaurus darwini, a New Rebbachisaurid Sauropod from the Early Cretaceous of the Iberian Peninsula Author(s): Fidel Torcida Fernández-Baldor, José Ignacio Canudo, Pedro Huerta, Diego Montero, Xabier
NOTES ON THE EVOLUTION OF VERTEBRAE IN THE SAUROPODOMORPHA. José F. BONAPARTE *
 NOTES ON THE EVOLUTION OF VERTEBRAE IN THE SAUROPODOMORPHA by José F. BONAPARTE * Museo Argentino de Ciencias Naturales Consejo Nacional de Investigaciones Científicas y Técnicas Avenida Angel Gallardo
NOTES ON THE EVOLUTION OF VERTEBRAE IN THE SAUROPODOMORPHA by José F. BONAPARTE * Museo Argentino de Ciencias Naturales Consejo Nacional de Investigaciones Científicas y Técnicas Avenida Angel Gallardo
Demandasaurus darwini, a new rebbachisaurid sauropod from the Early Cretaceous of the Iberian Peninsula
 Demandasaurus darwini, a new rebbachisaurid sauropod from the Early Cretaceous of the Iberian Peninsula FIDEL TORCIDA FERNÁNDEZ BALDOR, JOSÉ IGNACIO CANUDO, PEDRO HUERTA, DIEGO MONTERO, XABIER PEREDA SUBERBIOLA,
Demandasaurus darwini, a new rebbachisaurid sauropod from the Early Cretaceous of the Iberian Peninsula FIDEL TORCIDA FERNÁNDEZ BALDOR, JOSÉ IGNACIO CANUDO, PEDRO HUERTA, DIEGO MONTERO, XABIER PEREDA SUBERBIOLA,
Caudal Pneumaticity and Pneumatic Hiatuses in the Sauropod Dinosaurs Giraffatitan and Apatosaurus
 Caudal Pneumaticity and Pneumatic Hiatuses in the Sauropod Dinosaurs Giraffatitan and Apatosaurus Mathew J. Wedel 1 *, Michael P. Taylor 2 * 1 College of Osteopathic Medicine of the Pacific and College
Caudal Pneumaticity and Pneumatic Hiatuses in the Sauropod Dinosaurs Giraffatitan and Apatosaurus Mathew J. Wedel 1 *, Michael P. Taylor 2 * 1 College of Osteopathic Medicine of the Pacific and College
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province
 from Dashanpu, Zigong Municipality, Sichuan Province") A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
A new carnosaur from Yongchuan County, Sichuan Province
 A new carnosaur from Yongchuan County, Sichuan Province by Dong Zhiming Institute of Vertebrate Palaeontology and Palaeoanthropology, Academia Sinica Zhang Yihong, Li Xuanmin, and Zhou Shiwu Chongqing
A new carnosaur from Yongchuan County, Sichuan Province by Dong Zhiming Institute of Vertebrate Palaeontology and Palaeoanthropology, Academia Sinica Zhang Yihong, Li Xuanmin, and Zhou Shiwu Chongqing
Abstract RESEARCH ARTICLE
 RESEARCH ARTICLE Vertebral Pneumaticity in the Ornithomimosaur Archaeornithomimus (Dinosauria: Theropoda) Revealed by Computed Tomography Imaging and Reappraisal of Axial Pneumaticity in Ornithomimosauria
RESEARCH ARTICLE Vertebral Pneumaticity in the Ornithomimosaur Archaeornithomimus (Dinosauria: Theropoda) Revealed by Computed Tomography Imaging and Reappraisal of Axial Pneumaticity in Ornithomimosauria
A definite prosauropod dinosaur from the Lower Elliot Formation (Norian: Upper Triassic) of South Africa
 of South Africa") A definite prosauropod dinosaur from the Lower Elliot Formation (Norian: Upper Triassic) of South Africa Adam M. Yates Bernard Price Institute for Palaeontological Research, School of Geosciences, University
A definite prosauropod dinosaur from the Lower Elliot Formation (Norian: Upper Triassic) of South Africa Adam M. Yates Bernard Price Institute for Palaeontological Research, School of Geosciences, University
( M amenchisaurus youngi Pi, Ouyang et Ye, 1996)
") 39 4 2001 10 V ERTEBRATA PALASIATICA pp. 266 271 fig. 1,pl. I ( 643013), ( M amenchisaurus hochuanensis),,, Q915. 864 1995 12 31 (ZDM0126) ( M amenchisau rus hochuanensis Young et Chao, 1972),,, ZDM0126
39 4 2001 10 V ERTEBRATA PALASIATICA pp. 266 271 fig. 1,pl. I ( 643013), ( M amenchisaurus hochuanensis),,, Q915. 864 1995 12 31 (ZDM0126) ( M amenchisau rus hochuanensis Young et Chao, 1972),,, ZDM0126
A New Titanosaurian Sauropod from Late Cretaceous of Nei Mongol, China
 Vol. 80 No. 1 pp. 20 26 ACTA GEOLOGICA SINICA Feb. 2006 A New Titanosaurian Sauropod from Late Cretaceous of Nei Mongol, China XU Xing 1, *, ZHANG Xiaohong 2, TAN Qingwei 2, ZHAO Xijin 1 and TAN Lin 2
Vol. 80 No. 1 pp. 20 26 ACTA GEOLOGICA SINICA Feb. 2006 A New Titanosaurian Sauropod from Late Cretaceous of Nei Mongol, China XU Xing 1, *, ZHANG Xiaohong 2, TAN Qingwei 2, ZHAO Xijin 1 and TAN Lin 2
First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia
 from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia") First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia Tsogtbaatar Chinzorig¹, ³ *, Yoshitsugu Kobayashi², Khishigjav Tsogtbaatar³,
First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia Tsogtbaatar Chinzorig¹, ³ *, Yoshitsugu Kobayashi², Khishigjav Tsogtbaatar³,
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A.
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
REVISION OF REDONDASUCHUS (ARCHOSAURIA: AETOSAURIA) FROM THE UPPER TRIASSIC REDONDA FORMATION, NEW MEXICO, WITH DESCRIPTION OF A NEW SPECIES
 FROM THE UPPER TRIASSIC REDONDA FORMATION, NEW MEXICO, WITH DESCRIPTION OF A NEW SPECIES") Harris et al., eds., 2006, The Triassic-Jurassic Terrestrial Transition. New Mexico Museum of Natural History and Science Bulletin 37. REVISION OF REDONDASUCHUS (ARCHOSAURIA: AETOSAURIA) FROM THE UPPER
Harris et al., eds., 2006, The Triassic-Jurassic Terrestrial Transition. New Mexico Museum of Natural History and Science Bulletin 37. REVISION OF REDONDASUCHUS (ARCHOSAURIA: AETOSAURIA) FROM THE UPPER
Mathew John Wedel. B.S. (University of Oklahoma) A dissertation submitted in partial satisfaction of the. requirements for the degree of
 A dissertation submitted in partial satisfaction of the. requirements for the degree of") Postcranial Pneumaticity in Dinosaurs and the Origin of the Avian Lung by Mathew John Wedel B.S. (University of Oklahoma) 1997 A dissertation submitted in partial satisfaction of the requirements for the
Postcranial Pneumaticity in Dinosaurs and the Origin of the Avian Lung by Mathew John Wedel B.S. (University of Oklahoma) 1997 A dissertation submitted in partial satisfaction of the requirements for the
A NEW GIGANTIC SAUROPOD FROM THE MIDDLE JURASSIC OF SHANSHAN,
 A NEW GIGANTIC SAUROPOD FROM THE MIDDLE JURASSIC OF SHANSHAN, XINJIANG AUTONOMOUS REGION, CHINA Authors: Wu W.H., Zhou C.F, Wings O., Sekyia T.*, Dong Z.M. Abstract:A new gigantic sauropod dinosaur, Xinjiangtitan
A NEW GIGANTIC SAUROPOD FROM THE MIDDLE JURASSIC OF SHANSHAN, XINJIANG AUTONOMOUS REGION, CHINA Authors: Wu W.H., Zhou C.F, Wings O., Sekyia T.*, Dong Z.M. Abstract:A new gigantic sauropod dinosaur, Xinjiangtitan
YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
 Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
v:ii-ixi, 'i':;iisimvi'\>!i-:: "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi L I E) R.ARY OF THE VERSITY U N I or ILLINOIS REMO
 R.ARY OF THE VERSITY U N I or ILLINOIS REMO") "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
"^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
A Fossil Snake (Elaphe vulpina) From A Pliocene Ash Bed In Nebraska
 From A Pliocene Ash Bed In Nebraska") University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
Origin and Evolution of Birds. Read: Chapters 1-3 in Gill but limited review of systematics
 Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
Origin and Evolution of Birds Read: Chapters 1-3 in Gill but limited review of systematics Review of Taxonomy Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Aves Characteristics: wings,
TOPOTYPES OF TYPOTHORAX COCCINARUM, A LATE TRIASSIC AETOSAUR FROM THE AMERICAN SOUTHWEST
 Lucas, S.G. and Spielmann, J.A., eds., 2007, The Global Triassic. New Mexico Museum of Natural History and Science Bulletin 41. TOPOTYPES OF TYPOTHORAX COCCINARUM, A LATE TRIASSIC AETOSAUR FROM THE AMERICAN
Lucas, S.G. and Spielmann, J.A., eds., 2007, The Global Triassic. New Mexico Museum of Natural History and Science Bulletin 41. TOPOTYPES OF TYPOTHORAX COCCINARUM, A LATE TRIASSIC AETOSAUR FROM THE AMERICAN
A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev Doklady Akademii Nauk, SSSR 87:
 translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
Lower Cretaceous Kwanmon Group, Northern Kyushu
 Bull. Kitakyushu Mus. Nat. Hist., 11: 87-90. March 30, 1992 A New Genus and Species of Carnivorous Dinosaur from the Lower Cretaceous Kwanmon Group, Northern Kyushu Yoshihiko Okazaki Kitakyushu Museum
Bull. Kitakyushu Mus. Nat. Hist., 11: 87-90. March 30, 1992 A New Genus and Species of Carnivorous Dinosaur from the Lower Cretaceous Kwanmon Group, Northern Kyushu Yoshihiko Okazaki Kitakyushu Museum
Reassessment of the Evidence for Postcranial Skeletal Pneumaticity in Triassic Archosaurs, and the Early Evolution of the Avian Respiratory System
 Reassessment of the Evidence for Postcranial Skeletal Pneumaticity in Triassic Archosaurs, and the Early Evolution of the Avian Respiratory System Richard J. Butler 1,2 *, Paul M. Barrett 2, David J. Gower
Reassessment of the Evidence for Postcranial Skeletal Pneumaticity in Triassic Archosaurs, and the Early Evolution of the Avian Respiratory System Richard J. Butler 1,2 *, Paul M. Barrett 2, David J. Gower
Geo 302D: Age of Dinosaurs. LAB 7: Dinosaur diversity- Saurischians
 Geo 302D: Age of Dinosaurs LAB 7: Dinosaur diversity- Saurischians Last lab you were presented with a review of major ornithischian clades. You also were presented with some of the kinds of plants that
Geo 302D: Age of Dinosaurs LAB 7: Dinosaur diversity- Saurischians Last lab you were presented with a review of major ornithischian clades. You also were presented with some of the kinds of plants that
A Troodontid Dinosaur from Ukhaa Tolgod (Late Cretaceous Mongolia)
") PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3446, 9 pp., 4 figures June 2, 2004 A Troodontid Dinosaur from Ukhaa Tolgod (Late Cretaceous
PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3446, 9 pp., 4 figures June 2, 2004 A Troodontid Dinosaur from Ukhaa Tolgod (Late Cretaceous
4. Premaxilla: Foramen on the lateral surface of the premaxillary body (Yates 2007 ch. 4) 0 absent 1 present
 0 absent 1 present") The character matrix used as a basis for this study is that of Yates et al (2010) which is modified from the earlier matrix used by Yates (2007). This matrix includes characters acquired and/or modified
The character matrix used as a basis for this study is that of Yates et al (2010) which is modified from the earlier matrix used by Yates (2007). This matrix includes characters acquired and/or modified
PEABODY MUSEUM OF NATURAL HISTORY, YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 85 September 21, 1964 A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA STANLEY J. RIEL
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 85 September 21, 1964 A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA STANLEY J. RIEL
ONLINE APPENDIX 1. Morphological phylogenetic characters scored in this paper. See Poe (2004) for
 for") ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
NOTES ON THE FIRST SKULL AND JAWS OF RIOJASAURUS INCERTUS (DINOSAURIA, PROSAUROPODA, MELANOROSAURIDAE) OF THE LATE TRIASSIC OF LA RIOJA, ARGENTINA
 OF THE LATE TRIASSIC OF LA RIOJA, ARGENTINA") NOTES ON THE FIRST SKULL AND JAWS OF RIOJASAURUS INCERTUS (DINOSAURIA, PROSAUROPODA, MELANOROSAURIDAE) OF THE LATE TRIASSIC OF LA RIOJA, ARGENTINA José F. Bonaparte and José A. Pumares translated by Jeffrey
NOTES ON THE FIRST SKULL AND JAWS OF RIOJASAURUS INCERTUS (DINOSAURIA, PROSAUROPODA, MELANOROSAURIDAE) OF THE LATE TRIASSIC OF LA RIOJA, ARGENTINA José F. Bonaparte and José A. Pumares translated by Jeffrey
Erycine Boids from the Early Oligocene of the South Dakota Badlands
 Georgia Journal of Science Volume 67 No. 2 Scholarly Contributions from the Membership and Others Article 6 2009 Erycine Boids from the Early Oligocene of the South Dakota Badlands Dennis Parmley J. Alan
Georgia Journal of Science Volume 67 No. 2 Scholarly Contributions from the Membership and Others Article 6 2009 Erycine Boids from the Early Oligocene of the South Dakota Badlands Dennis Parmley J. Alan
Sauropod dinosaur remains from a new Early Jurassic locality in the Central High Atlas of Morocco
 Sauropod dinosaur remains from a new Early Jurassic locality in the Central High Atlas of Morocco CECILY S.C. NICHOLL, PHILIP D. MANNION, and PAUL M. BARRETT Nicholl, C.S.C., Mannion, P.D., and Barrett,
Sauropod dinosaur remains from a new Early Jurassic locality in the Central High Atlas of Morocco CECILY S.C. NICHOLL, PHILIP D. MANNION, and PAUL M. BARRETT Nicholl, C.S.C., Mannion, P.D., and Barrett,
Title: Phylogenetic Methods and Vertebrate Phylogeny
 Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Sauropoda from the Kelameili Region of the Junggar Basin, Xinjiang Autonomous Region
 Sauropoda from the Kelameili Region of the Junggar Basin, Xinjiang Autonomous Region Zhiming Dong (Institute of Vertebrate Paleontology and Paleoanthropology, Academia Sinica) Vertebrata PalAsiatica Volume
Sauropoda from the Kelameili Region of the Junggar Basin, Xinjiang Autonomous Region Zhiming Dong (Institute of Vertebrate Paleontology and Paleoanthropology, Academia Sinica) Vertebrata PalAsiatica Volume
GHBI /11/2013 MOHANRAJ.D Style 3. Juan Ignacio Canale a,b *, Fernando Emilio Novas a,c1 and Pol Diego a,d2
 Historical Biology, 2013 Vol. 00, No. 0, 1 32, http://dx.doi.org/10.1080/08912963.2013.861830 5 10 15 20 25 Osteology and phylogenetic relationships of Tyrannotitan chubutensis Novas, de Valais, Vickers-
Historical Biology, 2013 Vol. 00, No. 0, 1 32, http://dx.doi.org/10.1080/08912963.2013.861830 5 10 15 20 25 Osteology and phylogenetic relationships of Tyrannotitan chubutensis Novas, de Valais, Vickers-
On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds
 and the origin of birds") On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds by Qiang Ji and Shu an Ji Chinese Geological Museum, Beijing Chinese Geology Volume 233 1996 pp.
On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds by Qiang Ji and Shu an Ji Chinese Geological Museum, Beijing Chinese Geology Volume 233 1996 pp.
These small issues are easily addressed by small changes in wording, and should in no way delay publication of this first- rate paper.
 Reviewers' comments: Reviewer #1 (Remarks to the Author): This paper reports on a highly significant discovery and associated analysis that are likely to be of broad interest to the scientific community.
Reviewers' comments: Reviewer #1 (Remarks to the Author): This paper reports on a highly significant discovery and associated analysis that are likely to be of broad interest to the scientific community.
35. DATA REPORT: CRETACEOUS OSTRACODES FROM HOLES 865A AND 866A (MID-PACIFIC MOUNTAINS) 1. Renée Damotte 2
 1. Renée Damotte 2") Winterer, E.L., Sager, W.W., Firth, J.V., and Sinton, J.M. (Eds.), 1995 Proceedings of the Ocean Drilling Program, Scientific Results, Vol. 143 35. DATA REPORT: CRETACEOUS OSTRACODES FROM HOLES 865A AND
Winterer, E.L., Sager, W.W., Firth, J.V., and Sinton, J.M. (Eds.), 1995 Proceedings of the Ocean Drilling Program, Scientific Results, Vol. 143 35. DATA REPORT: CRETACEOUS OSTRACODES FROM HOLES 865A AND
Results of Prof. E. Stromer's Research Expedition in the Deserts of Egypt
 Proceedings of the Royal Bavarian Academy of Science Mathematical-physical Division Volume XXVIII, Paper 3 Results of Prof. E. Stromer's Research Expedition in the Deserts of Egypt II. Vertebrate Remains
Proceedings of the Royal Bavarian Academy of Science Mathematical-physical Division Volume XXVIII, Paper 3 Results of Prof. E. Stromer's Research Expedition in the Deserts of Egypt II. Vertebrate Remains
Overview of Sauropod Phylogeny and Evolution
 One Overview of Sauropod Phylogeny and Evolution Jeffrey A. Wilson SAUROPOD STUDIES FROM OWEN TO THE PRESENT This year marks the one hundred sixty-fourth anniversary of Richard Owen s (1841) description
One Overview of Sauropod Phylogeny and Evolution Jeffrey A. Wilson SAUROPOD STUDIES FROM OWEN TO THE PRESENT This year marks the one hundred sixty-fourth anniversary of Richard Owen s (1841) description
THE OCCURRENCE OF CONTOGENYS-LIKE LIZARDS IN THE LATE CRETACEOUS AND EARLY TERTIARY OF THE WESTERN INTERIOR OF THE U.S.A.
 Journal of Vertebrate Paleontology 29(3):677 701, September 2009 # 2009 by the Society of Vertebrate Paleontology ARTICLE THE OCCURRENCE OF CONTOGENYS-LIKE LIZARDS IN THE LATE CRETACEOUS AND EARLY TERTIARY
Journal of Vertebrate Paleontology 29(3):677 701, September 2009 # 2009 by the Society of Vertebrate Paleontology ARTICLE THE OCCURRENCE OF CONTOGENYS-LIKE LIZARDS IN THE LATE CRETACEOUS AND EARLY TERTIARY
THE LIGAMENT SYSTEM IN THE NECK OF RHEA AMERICANA AND ITS IMPLICATION FOR THE BIFURCATED NEURAL SPINES OF SAUROPOD DINOSAURS
 Journal of Vertebrate Paleontology 24(1):165 172, March 2004 2004 by the Society of Vertebrate Paleontology THE LIGAMENT SYSTEM IN THE NECK OF RHEA AMERICANA AND ITS IMPLICATION FOR THE BIFURCATED NEURAL
Journal of Vertebrate Paleontology 24(1):165 172, March 2004 2004 by the Society of Vertebrate Paleontology THE LIGAMENT SYSTEM IN THE NECK OF RHEA AMERICANA AND ITS IMPLICATION FOR THE BIFURCATED NEURAL
LOWER CRETACEOUS OF SOUTH DAKOTA.
 A NEW DINOSAUR, STP^GOSAURUS MARSHl, FROM THE LOWER CRETACEOUS OF SOUTH DAKOTA. By Frederic A. Lucas, Curator, Divisioii of Coiiipnrative Anatomy, in charge, of Section of Vertebrate Fossils. The name
A NEW DINOSAUR, STP^GOSAURUS MARSHl, FROM THE LOWER CRETACEOUS OF SOUTH DAKOTA. By Frederic A. Lucas, Curator, Divisioii of Coiiipnrative Anatomy, in charge, of Section of Vertebrate Fossils. The name
Vol. XIV, No. 1, March, The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S.
 By S.") Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Anatomy. Name Section. The Vertebrate Skeleton
 Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Appendix chapter 2: Description of Coloborhynchus spielbergi sp. nov. (Pterodactyloidea) from the Albian (Lower Cretaceous) of Brazil
 from the Albian (Lower Cretaceous) of Brazil") Appendix chapter 2: Description of Coloborhynchus spielbergi sp. nov. (Pterodactyloidea) from the Albian (Lower Cretaceous) of Brazil Appendix chapter 2 155 2.7. Appendix 2.7.1. Measurements Skull 15 12
Appendix chapter 2: Description of Coloborhynchus spielbergi sp. nov. (Pterodactyloidea) from the Albian (Lower Cretaceous) of Brazil Appendix chapter 2 155 2.7. Appendix 2.7.1. Measurements Skull 15 12
SNAKES OF THE SIWALIK GROUP (MIOCENE OF PAKISTAN): SYSTEMATICS AND RELATIONSHIP TO ENVIRONMENTAL CHANGE. Jason J. Head
: SYSTEMATICS AND RELATIONSHIP TO ENVIRONMENTAL CHANGE. Jason J. Head") Palaeontologia Electronica http://palaeo-electronica.org SNAKES OF THE SIWALIK GROUP (MIOCENE OF PAKISTAN): SYSTEMATICS AND RELATIONSHIP TO ENVIRONMENTAL CHANGE Jason J. Head ABSTRACT The lower and middle
Palaeontologia Electronica http://palaeo-electronica.org SNAKES OF THE SIWALIK GROUP (MIOCENE OF PAKISTAN): SYSTEMATICS AND RELATIONSHIP TO ENVIRONMENTAL CHANGE Jason J. Head ABSTRACT The lower and middle
PRELIMINARY REPORT ON A CLUTCH OF SIX DINOSAURIAN EGGS FROM THE UPPER TRIASSIC ELLIO T FORMATION, NORTHERN ORANGE FREE STATE. J. W.
 41 Pa/aeont. afr., 22, 41-45 (1979) PRELIMINARY REPORT ON A CLUTCH OF SIX DINOSAURIAN EGGS FROM THE UPPER TRIASSIC ELLIO T FORMATION, NORTHERN ORANGE FREE STATE b y J. W. Kitching ABSTRACT A clutch of
41 Pa/aeont. afr., 22, 41-45 (1979) PRELIMINARY REPORT ON A CLUTCH OF SIX DINOSAURIAN EGGS FROM THE UPPER TRIASSIC ELLIO T FORMATION, NORTHERN ORANGE FREE STATE b y J. W. Kitching ABSTRACT A clutch of
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1
 Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
Redescription of the Mongolian Sauropod NEMEGTOSAURUS MONGOLIENSIS Nowinski (Dinosauria:
 Journal of Systematic Palaeontology 3 (3): 283 318 Issued 24 August 2005 doi:10.1017/s1477201905001628 Printed in the United Kingdom C The Natural History Museum Redescription of the Mongolian Sauropod
Journal of Systematic Palaeontology 3 (3): 283 318 Issued 24 August 2005 doi:10.1017/s1477201905001628 Printed in the United Kingdom C The Natural History Museum Redescription of the Mongolian Sauropod
ZHAO XI-JIN, ROGER B. J. BENSON, STEPHEN L. BRUSATTE & PHILIP J. CURRIE
 Geol. Mag. 147 (1), 2010, pp. 13 27. c Cambridge University Press 2009 13 doi:10.1017/s0016756809990240 The postcranial skeleton of Monolophosaurus jiangi (Dinosauria: Theropoda) from the Middle Jurassic
Geol. Mag. 147 (1), 2010, pp. 13 27. c Cambridge University Press 2009 13 doi:10.1017/s0016756809990240 The postcranial skeleton of Monolophosaurus jiangi (Dinosauria: Theropoda) from the Middle Jurassic
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL
 Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
Cladistics (reading and making of cladograms)
") Cladistics (reading and making of cladograms) Definitions Systematics The branch of biological sciences concerned with classifying organisms Taxon (pl: taxa) Any unit of biological diversity (eg. Animalia,
Cladistics (reading and making of cladograms) Definitions Systematics The branch of biological sciences concerned with classifying organisms Taxon (pl: taxa) Any unit of biological diversity (eg. Animalia,
A R T I C L E S STRATIGRAPHIC DISTRIBUTION OF VERTEBRATE FOSSIL FOOTPRINTS COMPARED WITH BODY FOSSILS
 A R T I C L E S STRATIGRAPHIC DISTRIBUTION OF VERTEBRATE FOSSIL FOOTPRINTS COMPARED WITH BODY FOSSILS Leonard Brand & James Florence Department of Biology Loma Linda University WHAT THIS ARTICLE IS ABOUT
A R T I C L E S STRATIGRAPHIC DISTRIBUTION OF VERTEBRATE FOSSIL FOOTPRINTS COMPARED WITH BODY FOSSILS Leonard Brand & James Florence Department of Biology Loma Linda University WHAT THIS ARTICLE IS ABOUT
ALFRED GILLETT AND FOSSILS FROM STREET
 ALFRED GILLETT AND FOSSILS FROM STREET This collection of local fossils was formerly in the Crispin Hall, Street. Most of these fossils came from Alfred Gillett (1814-1904), a retired ironmonger who lived
ALFRED GILLETT AND FOSSILS FROM STREET This collection of local fossils was formerly in the Crispin Hall, Street. Most of these fossils came from Alfred Gillett (1814-1904), a retired ironmonger who lived
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC
 FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
A DINOSAUR FAUNA FROM THE LATE CRETACEOUS (CENOMANIAN) OF NORTHERN SUDAN. Oliver W. M. Rauhut
 OF NORTHERN SUDAN. Oliver W. M. Rauhut") Palaeont. afr., 35, 61-84 (1999) A DINOSAUR FAUNA FROM THE LATE CRETACEOUS (CENOMANIAN) OF NORTHERN SUDAN by Oliver W. M. Rauhut University o f Bristol, Department o f Geology, Wills Memorial Building,
Palaeont. afr., 35, 61-84 (1999) A DINOSAUR FAUNA FROM THE LATE CRETACEOUS (CENOMANIAN) OF NORTHERN SUDAN by Oliver W. M. Rauhut University o f Bristol, Department o f Geology, Wills Memorial Building,
POSTCRANIAL ANATOMY OF THE RAUISUCHIAN ARCHOSAUR BATRACHOTOMUS KUPFERZELLENSIS
 Journal of Vertebrate Paleontology 29(1):103 122, March 2009 # 2009 by the Society of Vertebrate Paleontology ARTICLE POSTCRANIAL ANATOMY OF THE RAUISUCHIAN ARCHOSAUR BATRACHOTOMUS KUPFERZELLENSIS DAVID
Journal of Vertebrate Paleontology 29(1):103 122, March 2009 # 2009 by the Society of Vertebrate Paleontology ARTICLE POSTCRANIAL ANATOMY OF THE RAUISUCHIAN ARCHOSAUR BATRACHOTOMUS KUPFERZELLENSIS DAVID
AMERICAN MUSEUM NOVITATES Published by
 AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
WHAT PNEUMATICITY TELLS US ABOUT PROSAUROPODS, AND VICE VERSA
 [Special Papers in Palaeontology 77, 2007, pp. 207 222] WHAT PNEUMATICITY TELLS US ABOUT PROSAUROPODS, AND VICE VERSA by MATHEW WEDEL University of California Museum of Paleontology and Department of Integrative
[Special Papers in Palaeontology 77, 2007, pp. 207 222] WHAT PNEUMATICITY TELLS US ABOUT PROSAUROPODS, AND VICE VERSA by MATHEW WEDEL University of California Museum of Paleontology and Department of Integrative
A NEW SAUROPOD DINOSAUR FROM THE LATE JURASSIC OF CHINA AND THE DIVERSITY, DISTRIBUTION, AND RELATIONSHIPS OF MAMENCHISAURIDS
 Journal of Vertebrate Paleontology e889701 (17 pages) Ó by the Society of Vertebrate Paleontology DOI: 10.1080/02724634.2014.889701 ARTICLE A NEW SAUROPOD DINOSAUR FROM THE LATE JURASSIC OF CHINA AND THE
Journal of Vertebrate Paleontology e889701 (17 pages) Ó by the Society of Vertebrate Paleontology DOI: 10.1080/02724634.2014.889701 ARTICLE A NEW SAUROPOD DINOSAUR FROM THE LATE JURASSIC OF CHINA AND THE