OSTEOLOGY OF PYGMY BLUE WHALE WITH ADDITIONAL INFORMATION ON EXTERNAL AND OTHER CHARACTERISTICS
|
|
|
- Justin Peters
- 5 years ago
- Views:
Transcription
1 OSTEOLOGY OF PYGMY BLUE WHALE WITH ADDITIONAL INFORMATION ON EXTERNAL AND OTHER CHARACTERISTICS HIDEO OMURA, TADAYOSHI ICHIHARA* AND TOSHIO KASUYA** INTRODUCTION The name" Pygmy blue whale" was first given by Ichihara (1961) to blue whales inhabiting the waters around Kerguelen Island, recognizing differences in the external characteristics from the blue whales elsewhere in the Antarctic. Since then researches on this population have been carried out by various authors (lchihara, 1963, 1966; Gambell, 1964; Ichihara and Doi, 1964; Zemsky and Boronin, 1964). The subspecies Balaenoptera musculus brevicauda was proposed by lchihara in a paper read in 1963 at the First International Symposium on Cetacean Research. His paper was not published until 1966 (Ichihara, 1966). Meanwhile, Zemsky and Boronin (1964) published the name brevicauda without calling it a new subspecies and without crediting lchihara (Rice and Scheffer, 1968). The identification of the subspecies is mainly based upon the external characteristics. In 1966 the Whales Research Institute was granted a special permission to take three pygmy blue whales for scientific researches, and a complete skeleton of this subspecies has been secured. The present paper deals with mainly the osteological study of the skeleton. OSTEOLOGY A complete skeleton of the pygmy blue whale was secured in This whale (66 Pl), a male of 18.6 m in length, was taken on 25 December 1966 at a position of 42-08'S and 44-09'E. The skeleton was transported on board factory ship to Ja pan. It had been hurried in sand, at a corner of campus of the College of Marine Science and Technology, Takai University, in Shimizu city during a period of about one and a half year from April 1967 to September 1968, for extraction of oils contained in bones. In September 1968 the bones were dug out and we made investigation of the bones, after cleaning. The photographs contained in this paper were also taken at this occasion. This skeleton is now mounted and being kept in the exhibition hall of the Marine Science Museum of the University. The body length of this whale is only 18.6 m (61 feet), but it had already attained physical maturity, because all of the vertebral epiphyses are fused completely to their centra, though * Far Seas Fisheries Research Laboratory, Fisheries Agency. ** Ocean Research Institute, University of Tokyo. No. 22, 1970, 1-27.



2
3 "" ~~ ""::0 TABLE 1. SKULL MEASUREMENTS OF BLUE AND PYGMY BLUE WHALES ~~._ Tomilin 1957 Tomilin 1957 Miller 1924 True 1904 Present specimen ~~ ZMAN ZMMGU U.S. Nat. Mus. Philadelphia Mus. Takai Univ.. " Ad. F. Ad. F (?) Subad. M Juv. F. Ad.M ~ Measurements Ostend Arctic Newfoundland Ocean City Antarctic ~ Skeleton 25 m Body m Body m Body m Body 18.6 m ~ ~ ~ ~ cm % cm % ~ cm 0/,o cm % ""!. Condylo-premaxillary length Zygomatic width Orbital width Rostrum length Distance between orbital processes of maxillaries Rostrum width at base * 35.6* "d >-<: 7. Rostrum width at middle l 163* 28.2* 28.8** ~ 8. Length of maxillary >-<: 9. Length of premaxillary td!:"' 10. Condyle width c 11. Occipital condyle height Occipital foramen width :r: 13. Occipital foramen height :i>!:"' 21. Length of nasal bones t'1 22. Total anterior width of both nasals Occipital bone width Occipital length, foramen to vertex Lower jaw length (straight) Lower jaw length (on curve) Lower jaw height at middle Lower jaw height, incl. coronoid * Curved length. ** Estimated by True. See text. The same measurements number in Table 13 of Tomilin (1957) is used here. t'1 ::E
.")
4 4 OMURA, ICHIHARA AND KASUYA whale is in the range of male blue whale, and this coincides with the conclusion by Ichihara (1966) that no difference is noted in the head region between pygmy and ordinary blue whales, from a study of external body proportions. In the lateral aspect the skull has a very flat appearance (Pl. I, Fig. 1 ). The profile resembles very closely to that of the Newfoundland specimen (No , US National Museum) as reported by Miller (1924). The most striking point in general view of the skull of pygmy blue whale is in the dorsal aspect. The principal characteristic feature of the ordinary blue whale's cranium is its wide rostrum with convex margins. The width of the rostrum at the middle of its length is as wide, or almost as wide as the width of its base (Tomilin, 1957). This is not applied to the rostrum of the pygmy blue whale. As seen in Pl. I the rostrum is less curved at its outer margin, but tapering from its base. In Table 1 the skull measurements of blue and pygmy blue whales are compared. The width of the rostrum at base does not differ between blue and pygmy blue whales, showing % of the skull length. On the contrary, in the width at its middle there is a good difference between the blue and pygmy blue whales. In the former this value is about 29 % whereas in the latter about 25 %- To our regret, however, only few papers on the osteology of blue whale are available. The only reliable measurement of the width at middle of the rostrum is that given by Tomilin (1957, specimen ZMMGU) which shows 29.1 % of the skull length (Table 1 ). Miller (1924) presents measurements of the skeleton of the Newfoundland specimen, found among the MS. notes left by Doctor True. But in the Table breadths of rostrum at middle as well as at base were measured on curve. The Ocean City specimen lacks premaxillae, and maxillae were separated from the cranium (True, 1904 ), but True states " With a suitable allowance for the premaxillae and interspace, the breadth of the rostrum (at middle) is 28.8 % (of the skull length). He states nothing whether this is straight or curved, but probably the latter, judged from the measurements reported by Miller (1924). If so, this value agrees well to that of the Newfoundland specimen. In any case no great difference between measurements in straight and on curve is expected at this position of the rostrum. Accordingly we can safely conclude that the rostrum width of the pygmy blue whale is about 25 % whereas in blue whale % at its middle. The fact that the width of the rostrum at the middle of its length is as wide, or almost as wide as the width of its base in blue whale is well supported by a drawing by van Beneden and Gervais (1880) and also by two photographs (dorsal and ventral aspects of the same skull) presented by Miller (1924). To our knowledge no paper is available on the osteology of blue whale in the Antarctic and in the North Pacific. But from Fig. 1, which shows the rostrum of the blue whale in these oceans, no difference is suggested in this respect among blue whales from different oceans. Tomilin (1957) states that in the blue whale the age-determined variations of the cranium are quite markedly expressed, and they are associated with the relative elongation of the facial region (rostrum, maxillaries, and premaxillaries) and the lateral expansion of the posterior region of the cranium and rostrum. For the


5 -~ -


6
7 PYGMY BLUE WHALE 7 TABLE 2. SKULL MEASUREMENTS OF PYGMY BLUE WHALE Measurement Length of skull, straight,, beak,, premaxillary, straight,, maxillary along upper surface Tip ofpremaxillary to posterior end of maxillary Tip of premaxillary to vertex,, tip of nasals (mesial) Tip of premaxillary to anterior end of palatines (mesial) Tip of premaxillary to posterior end of palatines (mesial) Tip ofpremaxillary to posterior end ofpterygoid,, anterior end of maxillary,,,, vomer Length ofsupraoccipital from foramen magnum Greatest breadth of skull, squamosal Breadth of skull, frontal,,, maxillary,, beak at base,, middle across premaxillaries, greatest at base of beak,, middle of beak,, posterior ends maxillaries at posterior ends Breadth of pterygoids,, palatines between tympanic bullae, outer Length of nasals mesially Breadth of nasals at anterior ends,, posterior ends,, frontal plane posterior to premaxillary Breadth of orbit (frontal wing),, occiput between squamosal sutures across occipital condyle Height of occipital condyle Breadth of foramen magnum Height of foramen magnum Breadth across mastoid process, tip to tip, greatest Length of mandible, straight Actual length (mm) R L R L R L R. 445 L. 435 R. 290 L R. 275 L R % of skull length % of skull breadth Continued...

8 8 OMURA, ICHIHARA AND KASUYA TABLE 2. Continued. Measurement Actual % of % of length (mm) skull length skull breadth Length of mandible, straight L , along outer curve R L Height of mandible at coronoid R L ,, processus articularis R L ,, middle R L of the pygmy blue whale, and in these the total number is He also gives vertebral number of three blue whale fetuses in the North Pacific, and in these the number is From the above figures the amount of variation in the vertebral number of the pygmy blue whale is considerable, as True (1904) states for blue whale, and no difference between pygmy and ordinary blue whales is noted. Detailed measurements of each vertebra are shown in Table 3, as well as in Fig. 6. In this Table the most interesting feature is the fact that in the length of the centrum the 12th lumbar shows the largest value. As shown in Fig. 6 (the bottom figure) in the caudal region the length of the centra decrease gradually from the lst caudal to the 14th, thence very steeply until l 7th, and from there again less steep. A similar figure is also presented for the Bryde's whale, comparing with that of the sei whale (Omura 1959, Fig. 3). The curve for the pygmy blue whale is quite similar to that for the Bryde's whale, but it differs from that for the sei whale. In the sei whale the caudal vertebrae are more developed, showing a curve slightly rises from the lst caudal towards about 5th and then decreases gradually, and then steeply, and lastly again less steep towards the end. In the black right whale this curve is somewhat different from the above (Omura et al. 1969, Fig. 23), and the length of the centra decreases gradually from the lst caudal towards the last, showing no remarkable steep portion, but in this species too the longest centrum is in the lumbar region. These facts may possibly connected with the manner of swimming. In the species which swim fast or follow long distant migration the caudal vertebrae may develop well and hence longer than those of the slow swimmer. In this connection it is interesting to compare the pygmy blue whale and the ordinary blue whale, but to our regret no material is available for the latter. Tomilin (1957) states "In adult animals, the cervical region forms 5 %, the thoracic 23 %, the lumbar 34 %, and the caudal 38 % of the total length of the vertebral column (E. J. Slijper, 1936)." In our specimen of the pygmy blue whale the total length of the vertebral column is 12,054 mm, and the percentages of the respective regions are: cervical 5.13%, thoracic 24.46%, lumbar %, and caudal %- This specimen has attained already of its physical maturity, as
9 PYGMY BLUE WHALE 9 TABLE 3. MEASUREMENTS OF VERTEBRAE OF PYGMY BLUE WHALE (in mm) Serial No II Vertebral No. C I " D I " " L I " " " 12 " Ca I " " 10 Greatest breadth 755 1, ,030 1,120 1,115 1,125 1,130 1,155 1,180 1,205 1,210 1,205 1,205 1,200 1,215 1,195 1,180 1,155 1,100 1,130 1,070 1, Greatest height Centrum Breadth Height Length Neural canal ~ Breadth Height Continued...
10 10 OMURA, ICHIHARA AND KASUYA TABLE 3. Serial Vertebral Greatest Greatest No. No. breadth height Continued. Centrum Neural canal ~ Breadth Height Length Breadth Height 47 Ca II " " " " " " " " " TABLE 4. COMPARISON OF VERTEBRAE. OCEAN CITY AND PRESENT SPECIMENS ARE COMPARED Actual length in mm % of skull length % of skull breadth Measurements ~, _ ~ OCS PB OCS PB OCS PB Greatest breadth of axis 914 1, Height of centrum of axis Greatest breadth of lst dorsal Height of centrum of lst dorsal Greatest breadth of lst lumbar 1,194 1, Height of centrum of lst lumbar Greatest breadth of lst caudal Height of centrum of lst caudal OCS: Ocean City specimen (True, 1904). Skulllength 4,450 mm. Skull breadth 2,210 mm. PB: Present specimen of pygmy blue whale. Skull length 4,860 mm. Skull breadth 2,360 mm. stated already. In comparing these figures it is suggested that the pygmy blue whale has a somewhat shorter dorsal and lumbar region, both combined, and a longer caudal region than the blue whale, contrary to our expectation. This is probably due to the different methods of measurements. In our specimen each vertebra is measured and then they are added. Since from studies of the external body proportions the pygmy blue whale is concluded to have a shorter tail region than ordinary blue whale, this problem should be left to future study. True (1904) presents some vertebral measurements of the Ocean City specimen. In Table 4 these are compared with our specimen. As seen in this Table the
11 PYGMY BLUE WHALE 11 proportional height of the centrum is smaller in the pygmy blue whale than in the Ocean City specimen. Further the greatest breadth of axis and lst dorsal vertebrae of the pygmy blue whale are greater, but that of lst lumbar and lst caudal is smaller than in the Ocean City specimen. It is suggested, therefore, that in the pygmy blue whale the centrum of the vertebral bone is smaller in general than in the ordinary blue whale and it decreases more steeply its breadth in lumbar and caudal regions. This is clearly shown in the percentage figures of greatest breadth against the skull breadth. One problem which is needed for consideration in this matter is the size variation of vertebrae according to growth, as the Ocean City specimen is a juvenile one, but to our regret no material is available for further discussion. L L 200 z 100 (/) I- z w L w 1,200 0:: ::::J 1,000 (/) <( w L i HEi'HT,GREATEST I I HEIGHT, CENTRUM BREADTH,CENTRUM LENGTH,CENTRUM I Fig. 6. Vertebral measurements of pygmy blue whale. In our specimen of pygmy blue whale the first vertebra in which the transverse process is perforated is the 47th (llth caudal) and the neural arch is obsolete on.
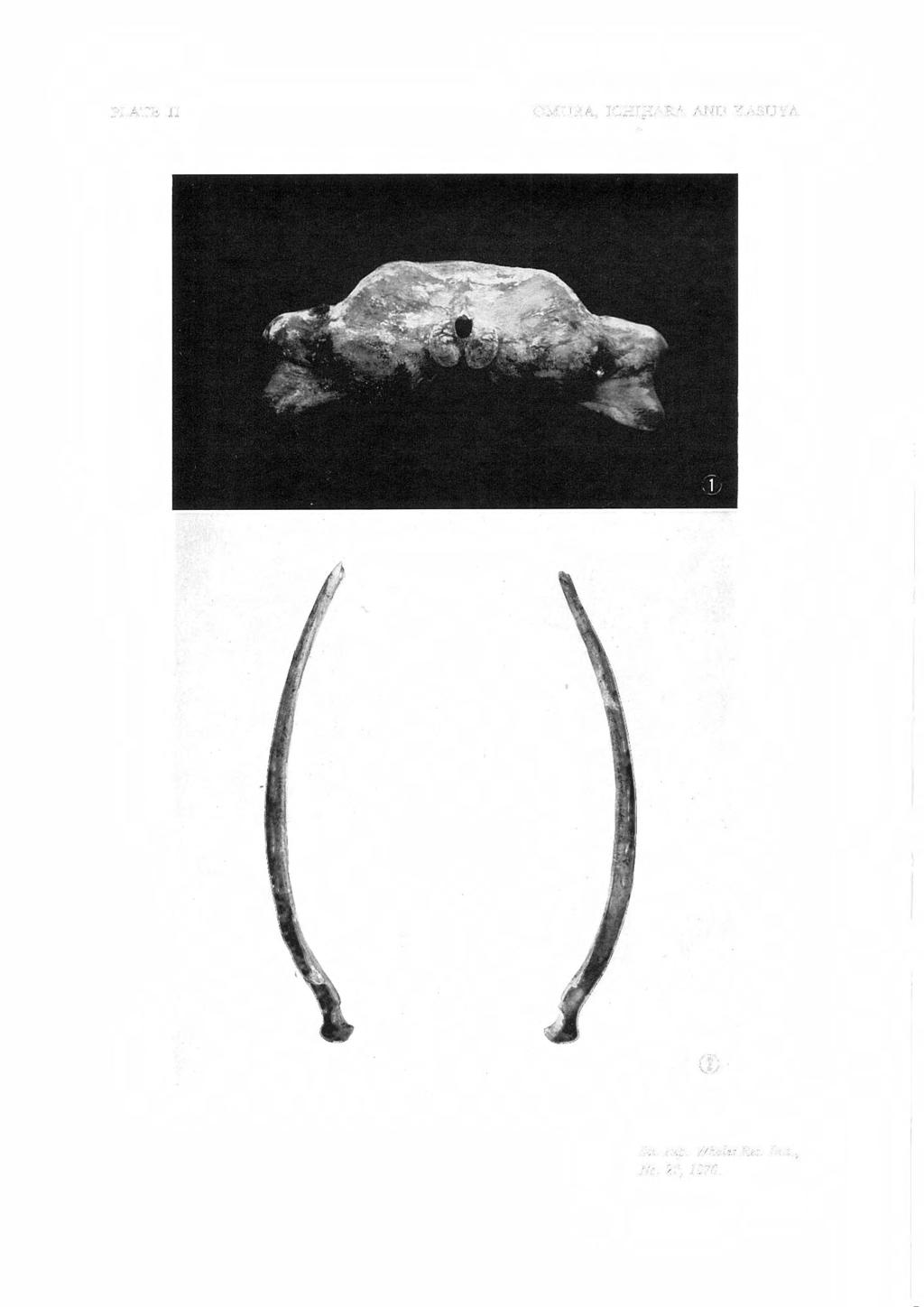
12 12 OMURA, ICHIHARA AND KASUYA Rib No TABLE 5. PB (right) MEASUREMENTS OF RIBS (in mm) NEWFOUNDLAND AND PRESENT SPECIMENS ARE COMPARED PB (left) Str. I. Curved 1. Str. 1. Curved 1. NFS Str. 1. % of skull length PB, right 1,345 1,605 1,315 1,575 1, ,850 2,080 1,825 2,060 1, ,170 2,460 2,145 2,405 2, ,245 2,605 2,245 2,595 2, ,250 2,675 2,220 2,670 2, ,260 2,725 2,310 2,705 2, ,215 2,655 broken 2, ,160 2,575 2,200 2,600 2, ,040 2,485 2,030 2,495 2, ,975 2,360 2,010 2,355 2, ,895 2,220 1,895 2,210 2, ,785 2,040 1,795 2,065 2, ,690 1,850 1,580 1,875 1, ,625 1,705 1,630 1,725 1, ,635 1,685 1,645 1,685 1, PB: Pygmy blue whale. Skull length 4,860 mm. NFS: Newfoundland specimen (Miller, 1924). Skull length 5, 790 mm. Str. I. : Straight length. Curved I.: Curved length. NFS I- (f) z <t: CJ:i: 50 PYGMY BLUE <t: I- C!l CJ -Z ~w,..x--x--x-- ll J x" X--x-- 0 J r' BLUE X--x- I I J, I I- :::J CJ::<'.'. z (f) w J '2R X RIB NUMBER Fig. 7. Length of ribs. Pygmy blue whale and Newfoundland specimen are compared. the 55th (19th caudal) vertebra. In the Ocean City specimen, as reported by True (1904), they are 46th and 55th respectively and show good agreement in these respects. Ribs (Pl. IV) Fifteen pairs of ribs are present in our specimen, but in addition one very small rib has been secured. Only the 3rd and 4th ribs are double headed. On the transverse processes of the dorsal vertebrae the articulating facet

13


14 r~l


15
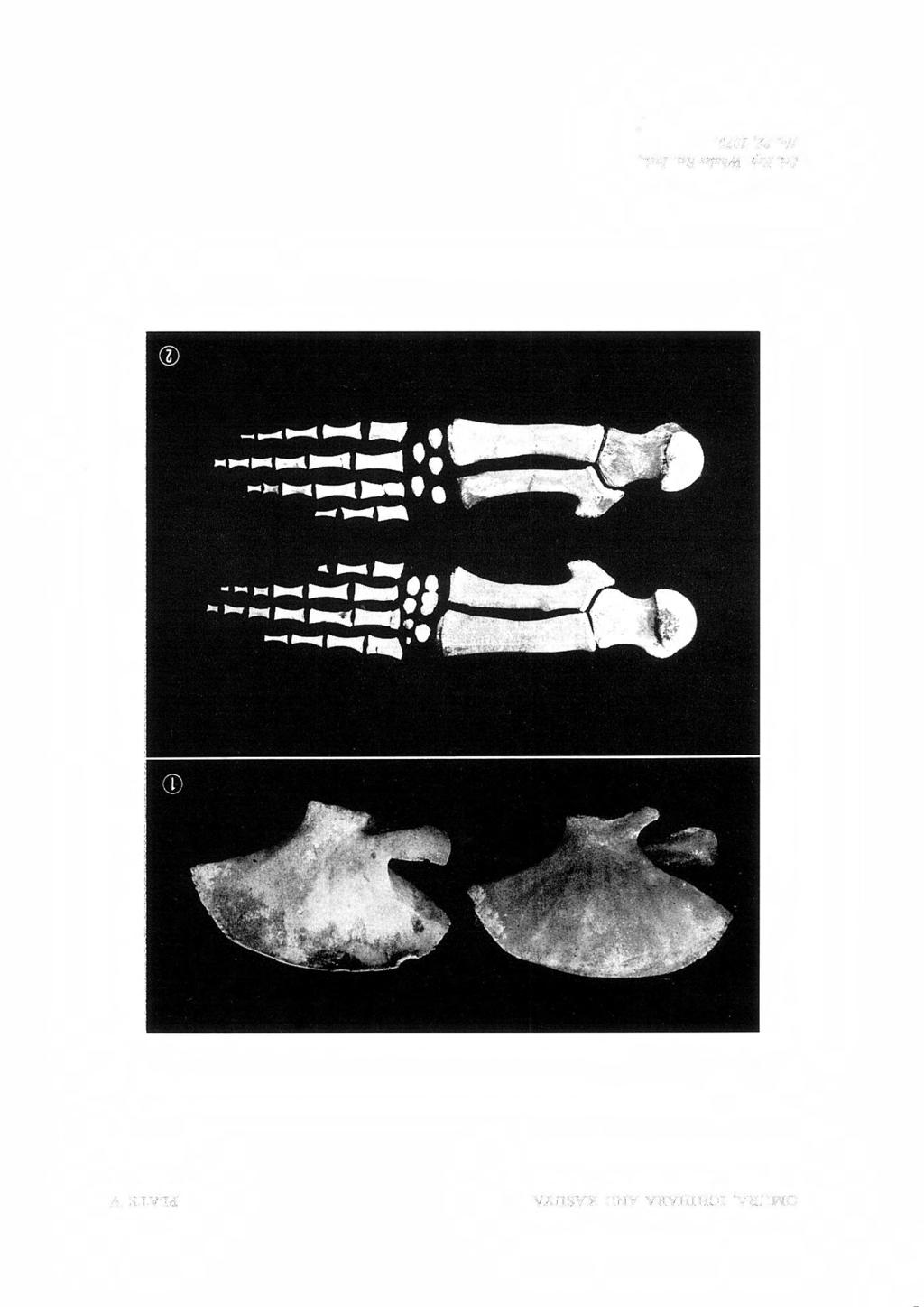
16 16 OMURA, ICHIHARA AND KASUYA TABLE 8. MEASUREMENTS OF SCAPULA OF PYGMY BLUE WHALE (in mm) Acromion Coracoid Glenoid fossa Breadth Height ~ ~ ~ Length Breadth Length Breadth Length Breadth Right 1, Left 1, TABLE 9. Measurements COMPARISON OF SCAPULA, HUMERUS, RADIUS, AND ULNA OF BLUE AND PYGMY BLUE WHALES Actual length in mm ocs NFS PB41 ocs Greatest breadth of scapula 1,257 1,450 1, Greatest height of scapula Length of humerus Length of radius , Length of ulna, greatest Breadth of radius at distal end Breadth of ulna at distal end % of skull length OCS: Ocean City specimen (True, 1904). Skull length 4,458 mm. NFS: Newfoundland specimen (Miller, 1924). Skull length 5,790 mm. PB: Present specimen of pygmy blue whale. Skull length 4,860 mm. 1) Without epiphyses. 2) Without inferior epiphysis. 3) Length above middle. Without inferior epiphysis. 4) Right side bones. NFS PB TABLE 10. MEASUREMENTS OF HUMERUS, RADIUS, AND ULNA OF PYGMY BLUE WHALE (in mm) Length, Breadth, Breadth, Breadth, Thickness, center prox. end dist. end at middle at middle Humerus Right Left Radius Right Left Ulna Right Left Pelvic bone (Fig. 11) Pelvic bones are different from those of the N ewfoundland specimen, judged from a photograph shown by Miller (1924). In our specimen they are more slender and the angle between the cranial and caudal processes is larger, hence less curved than in the Newfoundland specimen, and they resemble more to the pelvic bone of the ordinary blue whale in the Antarctic, as reported by Hosokawa (1951). In our specimen the lengths of right and left bones are 395 and 370 mm respectively and the right bone is somewhat larger than the left. Scapula (Pl. V, Fig. 1) Scapula is fan-shaped with a convex upper margin, and the acromion is well developed, as in the ordinary blue whale. Measurements
17 PYGMY BLUE WHALE 17 of scapula are shown in Table 8. True (1904) reports measurements of scapulae of blue whales from the North Atlantic Ocean, then known to him, and in these specimens the proportion of height to breadth is % He describes "The discrepancy in proportions, amounting to about 4 per cent, I am unable to account for. It affects both the American and European specimens and is not, apparently, due to difference in age and sex." Further Tomilin (1957) states "Scapula times as wide as high. In the males, the relative size of the scapula, and particularly its processes, is markedly greater than in the females." In the N ewfoundland specimen (Miller, 1924) the corresponding figure is 64.8 % (1.54 times as wide as high). In our specimen of pygmy blue whale these figures for the right and left scapulae are 62.2 % and 60.2 % (1.61 and 1.66 times as wide as high), respectively. In this respect, therefore, we find no difference between the pygmy and ordinary blue whales, but it is probable that even in a single specimen there is a slight difference between right and left scapulae. In the Miller's specimen the lengths of acromion and coracoid are 480 and 200 mm respectively. These are 33.1 % and 13.8 % of the breadth of scapula. In our specimen the corresponding figures are % and % respectively. Both specimens are males, but in our specimen of pygmy blue whale the processes are less developed than in the Newfoundland specimen. We can not conclude, however, whether this difference is of significant, due to limited data available. In Table 9 measurements of scapula, humerus, radius, and ulna of different TABLE II. MEASUREMENTS OF PHALANGES OF PYGMY BLUE WHALE (in mm) Right Left I II IV v I II IV v Length lst phalanx nd rd broken 4th th th Breadth at middle lst phalanx nd rd broken 4th th th Thickness at middle lst phalanx nd rd broken 4th th th
18 18 OMURA, ICHIHARA AND KASUYA specimens available are compared. In this Table measurements of each bone are also expressed as percentages of skull length. In this Table too our specimen of pygmy blue whale agrees well to the Newfoundland specimen. Humerus, Radius and Ulna (Pl. V Fig. 2) Comparison of these bones to those of the other specimens are included in Table 9. No special feature which separate the pygmy blue whale from the ordinary blue whale is observed from this Table as well as from a photograph presented by Miller (1924). Detailed measurements of these bones of our specimen are shown in Table 10. Carpals and Phalanges (Pl. V Fig. 2) Carpals are of no special feature. The phalangeal formula of our specimen is 1 5, 11 6, IV 5, V 3 Tomilin (1957) gives a formula of the ordinary blue whale, considering the variation in the number of phalanges in the specimens then available: 1 4 _ 5, 11 5 _ 8, IV 5 _ 7, V 3 _ 4 Our phalanx count is within the ranges of this formula. Measurements of phalanges in our specimen are shown in Table 11. Fig. 12. Measurement portion of tympanic bulla of blue whales Solid line for pygmy blue whales 130 Dotted line for blue whales from the North Pacific ' 100 ' 1: *,, E ' E 90 " 80 c.<::' 70 <f) :ii 60 E -~ 50 I 0 f-,~ 40 I :\,,,, \' 30 I ' I I ' \ \ \ 20 ~ 10 ~ \~ ' Measurement number Fig. 13. Comparison of size of tympanic bulla between pygmy blue whales from the Antarctic and blue whales from the North Pacific. Tympanic bulla. In relation to the rapid development of cetacean auditory sense, the tympanic bulla ceases to grow in the early stage of life, and its size is
19 PYGMY BLUE WHALE 19 TABLE 12. BIOLOGICAL DATA OF BLUE WHALES FROM WHICH TYMPANIC BULLA IS COLLECTED FOR COMPARISON OF SIZE No. Sex Body length Corpora number in ovaries Pygmy blue whales from the Antarctic, season Weight of testis Male 21.3 m 1.9, 1.9 kg Male , ll.7 Male , 4.2 Male , 16.7 Female Female Female Female Female Blue whales from the North Pacific, season 1965 Male , 1.2 Male , 1.1 Male , 20.5 Female Female 23.1 Female TABLE 13. BIOLOGICAL DATA ON THREE PYGMY BLUE WHALES TAKEN UNDER THE SPECIAL PERMISSION. No. Date of catch Position of catch Sex Laminae Body Weight number length of testis of ear plug 66Pl Dec. 25, 'S, 44-09'E Male 18.6m 13.4, 13.5 kg 46 66P2 Jan. 13, 'S, 73 -l5 1 E Male , P3 Jan. 17, 'S, 80-36'E Male , maintained throughout the life. Tympanic bullae were collected from 12 pygmy blue whales, of which 9 bullae were taken in the Antarctic season. For comparison, 6 tympanic bullae were collected from 6 blue whales taken by North Pacific expedition in Table 12 shows the biological data of these whales which are ranging from young to old. It is difficult to express accurately the form of bulla like renal and cowrie-shell. When the bulla is removed from the skull, a small fraction of bulla is usually broken down, however, 20 series as indicated in Fig. 12, can be measured for comparison. Measurement nos. 1, 2, 3, 12 and 14 are dimensions representing portions concerning the length of bulla. Nos. 4, 5, 6, 7, 10 and 11 show portions concerning the width of bulla. Nos. 8, 9, 13, 15, 16, 17 and 13 are related to the height of bulla. Mean value and range for each dimensions are indicated in Fig. 13. In the measurements except for no. 3, the value of the pygmy blue whale is smaller than that of blue whale from the North Pacific. When the mean of each measurement is connected by the line, the shape of bulla is supposed to resemble with each other, indicating a slight difference
20 20 OMURA, ICHIHARA AND KASUYA TABLE 14. Measurements Tip of upper jaw to notch of flukes Tip of upper jaw to blowholes Tip of upper jaw to eye (center) Tip of upper jaw to angle of gape Tip of upper jaw to tip of flipper Center of eye to ear hole Notch of flukes to tip of dorsal fin Notch of flukes to umbilicus Notch of flukes to end of ventral grooves Notch of flukes to anus Notch of flukes to anterior insertion of tail flukes Reproductive aperture to anus Dorsal fin, anterior insertion to tip Dorsal fin, height Flipper, anterior insertion to tip Flipper, greatest breadth BODY PROPORTIONS OF PYGMY BLUE WHALE (MALE) 66P Actual length (cm) 66Pl P P % of total length 66Pl P g 50 g_ 40 ~ 30 1j 20 (.) ~ 10 I + I --' ;\ / \ / \," \, \ I \ + \. ' ' ',_ The length from notch of flukes to centre of anus as a percentage of total length Fig. 14. Comparison of tail length between pygmy blue whales (solid line) and ordinary blue whales (broken line) in the Antarctic. which the bulla of the pygmy whale is proportionally smaller m measurement nos. 12 and 14, and proportionally larger in measurement no. 8. Yamada (1953) reports that the size of bulla from the Antarctic ordinary blue whale is cm long and cm wide. The Antarctic ordinary blue whale probably has a slightly larger tympanic bulla than the North Pacific blue whale, but unfortunately we have now no bulla from the former for the purpose of comparison. The mean weight of bulla in dry condition is g for the pygmy and g for the North Pacific blue whale. BODY PROPORTION Under a special permission three male pygmy blue whales were taken in the Antarctic in the season. Date and position of the catch are listed in Table 13
21 PYGMY BLUE WHALE 21 Items TABLE 15. Blubber Body, except ventral grooves Head Lower jaw Tail flukes Ventral grooves Total Meat Internal organs Tongue Lung Heart Liver Kidneys Pancreas Spleen Stomach Small intestine Large intestine Testes BODY WEIGHT OF PYGMY BLUE WHALES TAKEN IN THE ANTARCTIC SEASON 66P2 2, ,700 6,300 10, } ,496 Actual weight (kg) 66Pl 4,500 1,800 1, ,600 13,325 15,500 2, } , ,559 Bladder Fats Others Total Bones* Skull Lower jaws Vertebrae Ribs Chevron bones Scapulae Flippers Hyoids Total Baleen plates Total weight 1, , , ,737 1,710 1,230 2, ,404 1,200 42,988 * Weight in dried condition are shown in Appendix Table. 66P3 5,950 1,450 3, ,400 17,950 21,355 2, , } , ,674 2, , ,914 1,000 56,893 66P Percentage of total 66Pl P with biological data for each whale. The position of catch indicates that three whales were taken in the Subantarctic area, south of the Indian Ocean. From the histological observation on the tissue of testes, 66 Pl and 66 P3 whales were mature sexually, and 66 P2 immature. Laminae counting of ear plug suggests that 66 Pl and 66 P3 whales are 46 and 43 years old respectively, if we assume one lamina is accumulated per year. No ear plug was collected from young 66 P2
22 ~ 8 22 OMURA, ICHIHARA AND KASUYA TABLE 16. PERCENT WEIGHT OF EACH ORGAN AGAINST THE TOTAL WEIGHT OF INTERNAL ORGANS FOR PYGMY AND ORDINARY BLUE WHALES FROM THE ANTARCTIC Item Pygmy (A) Ordinary (B) Tongue Lung Heart Liver Kidney Stomach Small intestine 12.02* 8.29 Large intestine 4.01 * 2.64 Others Total Individuals examined 8 39 * from" 3 whales A/B lol_~~~ 1 ~ 6 ~~ 1 ~ 7 ~~,~8~~l9,--~2~0~~21~~22,--~23,--~24,--T.25;---,2~6-2n7---?;28, Bady length(m} Fig. 15. Length-weight relationship for both pygmy (closed circle) and ordinary (open circle) blue whales in the Antarctic. 20.~ ;J-r ~ ). ""'",.,...Q'" ;i Body length (m) Fig. 16. Length-weight relationship of internal organs for both pygmy (closed circle) and ordinary (open circle) blue whales in the Antarctic.

23 PYGMY BLUE WHALE 23 whale. External measurements of whales were carried out on the deck of the factory ship, and the results are shown in actual length and as the percentages of total body length in Table 14. The length from notch of flukes to center of anus, expressed as a percentage of total length, supports a trend which the tail portion of baleen whale becomes relatively short with the increment of body length. It is clear that 66 Pl whale, 18.6 metre in length, had already attained the physical maturity, as described in the foregoing chapter. lchihara (1963, 1966) concludes that the tail length of pygmy blue whale differs remarkably from that of ordinary blue whale in the Antarctic, examining the body proportion from the same length of whale. Since the form of whale body changes with the increment of age, it is desirable to compare the external measurements in full grown whales. As the ordinary male blue whale from the Antarctic attains the physical maturity at 79 feet (24.0 m) in length (Nishiwaki and Hayashi, 1950), we selected the data on the external measurements of larger male than 24 m from the Appendix III of Mackintosh and Wheeler (1929). These data were rearranged for the present purpose. Though the body length of physical maturity is not accurately known yet for the pygmy blue whale, we can safely assume that male pygmy of 69 feet (20.9 m) and over are physically matured. On the basis of selected samples, comparison of the tail portion of the pygmy blue with that of the ordinary blue is made in Fig. 14. The size of sample is 109 pygmy and 105 ordinary blue whales. The length from notch of flukes to center of anus as a percentage of total length is 25.68±0.19 (mean and two standard errors) for the pygmy, and ±0.22 for the ordinary blue whale. In the full grown male, the tail region is significantly smaller than that of the ordinary blue whale. BODY WEIGHT Body weights of three pygmy blue whales caught in the Antarctic season were measured on the deck of the factory ship. Actual weight and percentages weight are listed in Table 15, for each part of body. As the body weight of five pygmy blue whales was listed in Table 7 of lchihara's paper (1966), weights of a total of 8 pygmy blue can be compared with those of ordinary blue whales. In the Whales Research Institute are kept data on body weights of 14 male and 24 female ordinary blue whales which were measured in the Antarctic season, most of which were summarized by Nishiwaki (1950). Fig. 15 is obtained, when the mean body weight is plotted against the each meter range of whale length. The logarithmm is used for both body weight and length. As far as pygmy blue whale concerns, the length-weight relationship from the young to the old is not fully drawn for lack of data. The body weight of pygmy blue whale is heavier than that of ordinary blue whale of the same length, however, it is estimated that the rate of weight increment is almost similar between the two subspecies. In comparison of bones, meats and blubber weight, there is no difference between the two. Fig. 16 suggests that the difference of weight between the two

24 24 OMURA, ICHIHARA AND KASUYA is derived from the growth rate of the internal organs. The weight of internal organs in the pygmy blue whale at 21 m in length is about equal to that in the ordinary blue whale at 24 m in length. Table 16 shows the percent weight of each organ against the total weight of internal organs, for pygmy and ordinary blue whales. The item of others in Table 16 includes pancreas, spleen, reproductive organ, bladder, fat and etc. Except for the lung, liver and others, the percent weight of each organ from pygmy blue whales is greater than that for the ordinary blue whale. It is noticeable that the pygmy has heavier digestive organs than in the ordinary blue whale, but it is not known yet whether or not this is derived from the divergence of feeding habit. SUMMARY AND CONCLUSION From study of a skeleton of pygmy blue whale, mainly comparing with skeletons of blue whale in the North Atlantic reported by various authors, the followings are noted: 1. The body length of this specimen is 18.6 m, but it had attained physical maturity. The corresponding figure of ordinary blue whale in the Antarctic is 24.0m. 2. The width of the rostrum at middle of its length is as wide, or almost as wide as the width of its base in ordinary blue whale (29-31 % of skull length), but shorter by about 4 % of skull length in pygmy blue whale, though the width at base does not differ from the former. 3. Nasals are concave anteriorly, and the inner and outer border end at nearly the same level in ordinary blue whale, but rather convex and inner borders project more anteriorly than outer borders in pygmy blue whale. In the latter the length of nasals is shorter than in the former. 4. Mandibles are shorter, especially in curved length, than in ordinary blue whale. 5. The number of vertebrae does not differ from that of ordinary blue whale, but it is suggested that the centrum is smaller in general and it decreases more steeply its breadth in lumbar and caudal regions than in ordinary blue whale. 6. Straight lengths of ribs in the anterior portion of thorax are greater than in ordinary blue whale. 7. Tympanic bulla is smaller than that of blue whale in the North Pacific and slight difference in shape is also noted. 8. Malars of this specimen are congenital bipartite. From study of external and other characteristics the followings are noted: 9. Body proportions are reexamined with additional data and it is confirmed that in the full grown male the tail region is significantly smaller than that of ordinary blue whale. 10. The body weight of pygmy blue whale is heavier than that of ordinary blue whale of the same length in the Antarctic, and this difference is mainly due to the fact that the former has more heavier digestive organs than the latter.
25 PYGMY BLUE WHALE 25 In conclusion above we think there is a good additional reason pygmy blue whale from the ordinary blue whale as a subspecies, i.e. musculus brevicauda. to separate Balaenoptera ACKNOWLEDGEMENTS Our sincere thanks are due to Fisheries Agency of Japanese government who granted special permit to take three pygmy blue whales in the Antarctic season for scientific researches, and to crew of the Kyokuyo Maru No. 3 expedition who cooperated in the field works. We are much indebted to Dr. M. Nishiwaki of the Ocean Research Institute, University of Tokyo, and Dr. S. Ohsumi and other members of the Far Seas Fisheries Research Laboratory who cooperated in treating, including measurements and taking photographs, of the skeleton. Our grateful thanks are also extended to the staff of the Tokai University as well as Mr. K. Kawamura and Mr. S. Machida of the Whales Research Institute who also helped us in this study. REFERENCES FRASER, F. C. and A.J. E. CAVE, Congenital jugal bipartism in mysticetes. J.,Z,ool. Lond., 157: GAMBELL, R., A pygmy blue whale at Durban. Norsk Hvalfangst-Tid., 53 (3): HosoKAWA, H., On the pelvic cartilages of the Balaenoptera-foetuses, with remarks on the specifical and sexual difference. 5: lchihara, T., Blue whales in the waters around Kerguelen Island. Norsk Hvalfangst-Tid., 50 (!): lchihara, T., Identification of the pigmy blue whales in the Antarctic. Norsk Hvalfangst-Tid., 52 (5): lchihara, T., The pygmy blue whale, Balaenoptera musculus brevicauda, a new subspecies from the Antarctic. In Whales, dolphins, and porpoises. Ed. K. S. Norris, Univ. Calif. Press lchihara, T. and T. Dm, Stock assessment of pigmy blue whales in the Antarctic. Norsk Hval Jangst-Tid., 53 (6): MACKINTOSH, N. A. and J. F. G. WHEELER, Southern blue and fin whales. Discovery Rep., 1 : MILLER, G. S., Jr., Some hitherto unpublished photographs and measurements of the blue whale. Proc. U. S. N. Mus. 66 (7): 1-4. NISHIWAKI, M., On the body weight of whales. 4: NrsHIWAKI, M. and K. HAYASHI, Biological survey of fin and blue whales taken in the Antarctic season by the Japanese fleet. 3: OMURA, H., Bryde's whale from the coast of Japan. 14: OMURA, H A systematic study of the hyoid bones in the baleen whales. 18: OMURA, H., S. 0HSUMI, T. NEMOTO, K. NAsu, and T. KASUYA, Black right whales in the North Pacific. 21: RICE, D. W. and V. B. SCHEFFER, A list of the marine mammals of the world. U.S. Fish Wild!. Serv. Spee. Sci. Rep. Fish. 579, 16 pp. ToMILIN, A. G., Cetacea. Mammals of the U.S.S.R. and adjacent countries (Ed. S. I. Ognev) IX. English translation by Omry Ronen, J ersalem pp. TRUE, E. W., The whalebone whales of the western North Atlantic compared with those occuring in
26 26 OMURA, ICHIHARA AND KASUYA European waters, with some observations on the species of the North Pacific. Smithsonian Contr. Know!., 33: VAN BENEDEN, and P. GERVAIS, Ostt!ographie des ct!tact!s vivants etfossiles. Bertrand, Paris. 643 pp. YAMADA, M., Contribution to the anatomy of the organ of hearing of whales. Sci. Rep. Whales Res. Inst., 8: ZEMSKY, V. A. and V. A. BoRONIN, On the question of the pygmy blue whale taxonomic position. Norsk Hvaifangst-Tid., 53 ( 11) : APPENDIX TABLE. WEIGHT OF BONES OF PYGMY BLUE WHALE, IN DRIED CONDITION Skull Mandibles Vertebrae Cervical Dorsal Lumbar Caudal Ribs Scapulae Humerus Ulnae Radii Carpals and phalanges Hyoid bones Sternum Chevron bones Total weight 700 kg JO.I ,438.3 Note: The bones were weighed in October 1969 before mounting.
27 PYGMY BLUE WHALE 27 EXPLANATION OF PLATES Fig. I. Skull of pygmy blue whale. Fig. 2. The same. Ventral view. Fig. 3. The same. Dorsal view. Lateral view. PLATE I PLATE II Fig. I. Skull of pygmy blue whale. Posterior view. Fig. 2. Mandibles of pygmy blue whale. Dorsal view. Fig. 1. Fig. 2. Fig. 3. Fig. 4. Fig. 1. Fig. 2. PLATE III Atlas, axis, and 3rd cervical of pygmy blue whale. 4th-7th cervicals of the same specimen. Dorsal vertebrae of the same specimen. Lumbar vertebrae of the same specimen. PLATE IV Caudal vertebrae of pygmy blue whale. Ribs of the same specimen. Fig. I. Scapulae of pygmy blue whale. Fig. 2. Bones in flipper of the same specimen. PLATE V

 l")
28 sa1m1a1 if u!'s V~V HJH :) l v~nwo

29

 I")

30 ra7m1111 rt~u ~l.lv1d V~VH JH :) I 'v ~ 日 WO


31

32
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC
 FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
FURTHER STUDIES ON TWO SKELETONS OF THE BLACK RIGHT WHALE IN THE NORTH PACIFIC HIDEO OMURA, MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT Two skeletons of the black right whale were studied, supplementing
OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE
 OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT This is a report of measurements on the skeleton of a male se1 whale caught in the Antarctic. The skeleton of
OSTEOLOGICAL NOTE OF AN ANTARCTIC SEI WHALE MASAHARU NISHIWAKI* AND TOSHIO KASUYA* ABSTRACT This is a report of measurements on the skeleton of a male se1 whale caught in the Antarctic. The skeleton of
DISCOVERY AND CAPTURE
 46 NSHWAK AND KASUYA Hitherto, this species has occurred no other place in the world on 33 28' N. around, San Diego and Casabranca for example. Our news was not only the evidence of this occurrence. When
46 NSHWAK AND KASUYA Hitherto, this species has occurred no other place in the world on 33 28' N. around, San Diego and Casabranca for example. Our news was not only the evidence of this occurrence. When
AN OSTEOLOGICAL STUDY OF THE CUVIER'S BEAKED WHALE,,ZIPHIUS CAVIROSTRIS, IN THE NORTHWEST PACIFIC
 AN OSTEOLOGICAL STUDY OF THE CUVIER'S BEAKED WHALE,,ZIPHIUS CAVIROSTRIS, IN THE NORTHWEST PACIFIC HIDEO OMURA ABSTRACT Skulls and postcranial skeletons of :(,iphius cavirostris were studied for eleven
AN OSTEOLOGICAL STUDY OF THE CUVIER'S BEAKED WHALE,,ZIPHIUS CAVIROSTRIS, IN THE NORTHWEST PACIFIC HIDEO OMURA ABSTRACT Skulls and postcranial skeletons of :(,iphius cavirostris were studied for eleven
FIRST RECORD OF MESOPLODON DENSIROSTRIS FROM FORMOSA
 FIRST RECORD OF MESOPLODON DENSIROSTRIS FROM FORMOSA TOSHIO KASUYA* AND MASAHARU NISHIWAKI* ABSTRACT Two records of female Mesoplodon densirostris are reported. Comments on the external character, skull
FIRST RECORD OF MESOPLODON DENSIROSTRIS FROM FORMOSA TOSHIO KASUYA* AND MASAHARU NISHIWAKI* ABSTRACT Two records of female Mesoplodon densirostris are reported. Comments on the external character, skull
Mammalogy Laboratory 1 - Mammalian Anatomy
 Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
-Cl No. of baleen plates. ..c KASUYA AND RICE E ~20 Q. 10. Sci. Rep. Whales Res. Inst., No. 22, 1970.
 4 KASUYA AND RICE plate along the lateral edge. As seen in this figure, the length of the baleen plates in the anterior part of the series is not bilaterally symmetrical. The plates on the right side are
4 KASUYA AND RICE plate along the lateral edge. As seen in this figure, the length of the baleen plates in the anterior part of the series is not bilaterally symmetrical. The plates on the right side are
FERESA ATTENUATA CAPTURED
 FERESA ATTENUATA CAPTURED AT THE PACIFIC COAST OF JAPAN IN 1963 MASAHARU NISHIWAKI, TOSHIO KASUYA, TOSHIRO KAMIYA*, TERVO TOBAYAMA**, AND MASAYUKI NAKAJIMA*** INTRODUCTION The fishermen at some fishing
FERESA ATTENUATA CAPTURED AT THE PACIFIC COAST OF JAPAN IN 1963 MASAHARU NISHIWAKI, TOSHIO KASUYA, TOSHIRO KAMIYA*, TERVO TOBAYAMA**, AND MASAYUKI NAKAJIMA*** INTRODUCTION The fishermen at some fishing
New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia
 1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
1955 Doklady, Academy of Sciences USSR 104 (5):779-783 New Carnivorous Dinosaurs from the Upper Cretaceous of Mongolia E. A. Maleev (translated by F. J. Alcock) The present article is a summary containing
NECROPSY FORM STRAND LOCATION: FLOATING IN VAQUITA REFUGE BY MX TIME: 10 AM
 NECROPSY FORM FIELD #: Ps 9 NECROPSY DATE: April 4 2018 SPECIES: PHOCOENA SINUS STRAND DATE: March 28 2018 AGE CLASS: ADULT STRAND LOCATION: FLOATING IN VAQUITA REFUGE BY MX NAVY, BAJA CALIFORNIA, MX SEX:
NECROPSY FORM FIELD #: Ps 9 NECROPSY DATE: April 4 2018 SPECIES: PHOCOENA SINUS STRAND DATE: March 28 2018 AGE CLASS: ADULT STRAND LOCATION: FLOATING IN VAQUITA REFUGE BY MX NAVY, BAJA CALIFORNIA, MX SEX:
.56 m. (22 in.). COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition
. COMPSOGNATHOID DINOSAUR FROM THE. Medicine Bow, Wyoming, by the American Museum Expedition") Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Article XII.-ORNITHOLESTES HERMANNI, A NEW COMPSOGNATHOID DINOSAUR FROM THE UPPER JURASSIC. By HENRY FAIRFIELD OSBORN. The type skeleton (Amer. Mus. Coll. No. 6I9) of this remarkable animal was discovered
Fish 475: Marine Mammalogy
 Fish 475: Marine Mammalogy Taxonomy (continued) Friday, 3 April 2009 Amanda Bradford Course website: http://faculty.washington.edu/glennvb/fish475 Mysticeti: The baleen whales About 10-12 species; Formerly
Fish 475: Marine Mammalogy Taxonomy (continued) Friday, 3 April 2009 Amanda Bradford Course website: http://faculty.washington.edu/glennvb/fish475 Mysticeti: The baleen whales About 10-12 species; Formerly
Anatomy. Name Section. The Vertebrate Skeleton
 Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
A skull without mandihle, from the Hunterian Collection (no.
 4 MR. G. A. BOULENGER ON CHELONIAN REMAINS. [Jan. 6, 2. On some Chelonian Remains preserved in the Museum of the Eojal College of Surgeons. By G. A. Boulenger. [Eeceived December 8, 1890.] In the course
4 MR. G. A. BOULENGER ON CHELONIAN REMAINS. [Jan. 6, 2. On some Chelonian Remains preserved in the Museum of the Eojal College of Surgeons. By G. A. Boulenger. [Eeceived December 8, 1890.] In the course
( M amenchisaurus youngi Pi, Ouyang et Ye, 1996)
") 39 4 2001 10 V ERTEBRATA PALASIATICA pp. 266 271 fig. 1,pl. I ( 643013), ( M amenchisaurus hochuanensis),,, Q915. 864 1995 12 31 (ZDM0126) ( M amenchisau rus hochuanensis Young et Chao, 1972),,, ZDM0126
39 4 2001 10 V ERTEBRATA PALASIATICA pp. 266 271 fig. 1,pl. I ( 643013), ( M amenchisaurus hochuanensis),,, Q915. 864 1995 12 31 (ZDM0126) ( M amenchisau rus hochuanensis Young et Chao, 1972),,, ZDM0126
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
AMERICAN MUSEUM NOVITATES Published by
 AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
AMERICAN MUSEUM NOVITATES Published by Number 782 THE AmzRICAN MUSEUM OF NATURAL HISTORY Feb. 20, 1935 New York City 56.81, 7 G (68) A NOTE ON THE CYNODONT, GLOCHINODONTOIDES GRACILIS HAUGHTON BY LIEUWE
complex in cusp pattern. (3) The bones of the coyote skull are thinner, crests sharper and the
 The bones of the coyote skull are thinner, crests sharper and the") DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
Williston, and as there are many fairly good specimens in the American
 56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE
 PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
Mammalogy Lab 1: Skull, Teeth, and Terms
 Mammalogy Lab 1: Skull, Teeth, and Terms Be able to: Goals of today s lab Locate all structures listed on handout Define all terms on handout what they are or what they look like Give examples of mammals
Mammalogy Lab 1: Skull, Teeth, and Terms Be able to: Goals of today s lab Locate all structures listed on handout Define all terms on handout what they are or what they look like Give examples of mammals
SECOND RECORD OF SPECTACLED PORPOISE FROM SUBANTARCTIC SOUTHWEST PACIFIC
 SECOND RECORD OF SPECTACLED PORPOISE FROM SUBANTARCTIC SOUTHWEST PACIFIC R. EWAN FORDYCE Department of Geology, University of Otago, P, 0. Box 56, Dunedin, New Zealand ROBERT H. MATTLIN Fisheries Research
SECOND RECORD OF SPECTACLED PORPOISE FROM SUBANTARCTIC SOUTHWEST PACIFIC R. EWAN FORDYCE Department of Geology, University of Otago, P, 0. Box 56, Dunedin, New Zealand ROBERT H. MATTLIN Fisheries Research
A SYSTEMATIC STUDY OF THE HYOID BONES IN THE BALEEN WHALES
 A SYSTEMATIC STUDY OF THE HYOID BONES IN THE BALEEN WHALES HIDEO OMURA The hyoid bone is accepted as having characteristics which can be used as a taxonomic criterion in the terrestrial mammals (Sprague,
A SYSTEMATIC STUDY OF THE HYOID BONES IN THE BALEEN WHALES HIDEO OMURA The hyoid bone is accepted as having characteristics which can be used as a taxonomic criterion in the terrestrial mammals (Sprague,
SQUIDS FOUND IN THE STOMACH OF SPERM WHALES IN THE NORTHWESTERN PACIFIC*
 SQUIDS FOUND IN THE STOMACH OF SPERM WHALES IN THE NORTHWESTERN PACIFIC* T AKEHIKO KAWAKAMI Tokai Regional Fisheries Research Laboratory, Tokyo ABSTRACT The squids obtained from the stomachs of sperm whales
SQUIDS FOUND IN THE STOMACH OF SPERM WHALES IN THE NORTHWESTERN PACIFIC* T AKEHIKO KAWAKAMI Tokai Regional Fisheries Research Laboratory, Tokyo ABSTRACT The squids obtained from the stomachs of sperm whales
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO
 A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
A NEW SPECIES OF EXTINCT TURTLE FROM THE UPPER PLIOCENE OF IDAHO By Charles W. Gilmore Curator, Division of Vertebrate Paleontology United States National Museum Among the fossils obtained bj^ the Smithsonian
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL
 Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
Reprinted from: CRUSTACEANA, Vol. 32, Part 2, 1977 LEIDEN E. J. BRILL NOTES AND NEWS 207 ALPHE0PS1S SHEARMII (ALCOCK & ANDERSON): A NEW COMBINATION WITH A REDESCRIPTION OF THE HOLOTYPE (DECAPODA, ALPHEIDAE)
Color: body is dark brown to dark gray; belly and area near mouth and head are often light gray or white
 Sperm Whale Type: toothed Length: 33-60 feet Color: body is dark brown to dark gray; belly and area near mouth and head are often light gray or white Features/Markings: skin often appears wrinkled; enormous
Sperm Whale Type: toothed Length: 33-60 feet Color: body is dark brown to dark gray; belly and area near mouth and head are often light gray or white Features/Markings: skin often appears wrinkled; enormous
click for previous page SEA TURTLES
 click for previous page SEA TURTLES FAO Sheets Fishing Area 51 TECHNICAL TERMS AND PRINCIPAL MEASUREMENTS USED head width (Straight-line distances) head prefrontal precentral carapace central (or neural)
click for previous page SEA TURTLES FAO Sheets Fishing Area 51 TECHNICAL TERMS AND PRINCIPAL MEASUREMENTS USED head width (Straight-line distances) head prefrontal precentral carapace central (or neural)
AMERICAN NATURALIST. Vol. IX. -DECEMBER, No. 12. OR BIRDS WITH TEETH.1 OI)ONTORNITHES,
ONTORNITHES,") AMERICAN NATURALIST. Vol. IX. -DECEMBER, 1875.-No. 12. OI)ONTORNITHES, OR BIRDS WITH TEETH.1 BY PROFESSOR 0. C. MARSH. REMAINS of birds are amono the rarest of fossils, and few have been discovered except
AMERICAN NATURALIST. Vol. IX. -DECEMBER, 1875.-No. 12. OI)ONTORNITHES, OR BIRDS WITH TEETH.1 BY PROFESSOR 0. C. MARSH. REMAINS of birds are amono the rarest of fossils, and few have been discovered except
Vol. XIV, No. 1, March, The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S.
 By S.") Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
Vol. XIV, No. 1, March, 1950 167 The Larva and Pupa of Brontispa namorikia Maulik (Coleoptera: Chrysomelidae: Hispinae) By S. MAULIK BRITISH MUSEUM (NATURAL HISTORY) (Presented by Mr. Van Zwaluwenburg
ONLINE APPENDIX 1. Morphological phylogenetic characters scored in this paper. See Poe (2004) for
 for") ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
ONLINE APPENDIX Morphological phylogenetic characters scored in this paper. See Poe () for detailed character descriptions, citations, and justifications for states. Note that codes are changed from a
A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA
 A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA MYCTEROSAURUS LONGICEPS S. W. WILLISTON University of Chicago The past summer, Mr. Herman Douthitt, of the University of Chicago paleontological expedition,
A NEW GENUS AND SPECIES OF AMERICAN THEROMORPHA MYCTEROSAURUS LONGICEPS S. W. WILLISTON University of Chicago The past summer, Mr. Herman Douthitt, of the University of Chicago paleontological expedition,
A NEARLY COMPLETE TURTLE SKELETON FROM THE UPPER CRETACEOUS OF MONTANA
 CONTRIBUTIONS PBOM THE MUSEUM OF PALEONTOLOGY UNIVERSITY OF MICHIGAN VOL VI, No. 1. pp. 1-19 (18 figs.) D~c~arrrm 1, 1989 A NEARLY COMPLETE TURTLE SKELETON FROM THE UPPER CRETACEOUS OF MONTANA BY E. C.
CONTRIBUTIONS PBOM THE MUSEUM OF PALEONTOLOGY UNIVERSITY OF MICHIGAN VOL VI, No. 1. pp. 1-19 (18 figs.) D~c~arrrm 1, 1989 A NEARLY COMPLETE TURTLE SKELETON FROM THE UPPER CRETACEOUS OF MONTANA BY E. C.
YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
 Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
Vol. 30, No. 4 VERTEBRATA PALASIATICA pp. 313-324 October 1992 [SICHUAN ZIGONG ROUSHILONG YI XIN ZHONG] figs. 1-5, pl. I-III YANGCHUANOSAURUS HEPINGENSIS - A NEW SPECIES OF CARNOSAUR FROM ZIGONG, SICHUAN
A Fossil Snake (Elaphe vulpina) From A Pliocene Ash Bed In Nebraska
 From A Pliocene Ash Bed In Nebraska") University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Transactions of the Nebraska Academy of Sciences and Affiliated Societies Nebraska Academy of Sciences 198 A Fossil Snake
Central Marine Fisheries Research Institute, Mandapam Camp
 w«r n Mar. biol. Ass. India, 1961, 3 (1 & 2): 92-95 ON A NEW GENUS OF PORCELLANIDAE (CRUSTACEA-ANOMURA) * By C. SANKARANKUTTY Central Marine Fisheries Research Institute, Mandapam Camp The specimen described
w«r n Mar. biol. Ass. India, 1961, 3 (1 & 2): 92-95 ON A NEW GENUS OF PORCELLANIDAE (CRUSTACEA-ANOMURA) * By C. SANKARANKUTTY Central Marine Fisheries Research Institute, Mandapam Camp The specimen described
Macro-anatomical studies of the African giant pouched rat (Cricetomys gambianus) axial skeleton
 axial skeleton") Standard Scientific Research and Essays Vol1 (10): 221-227, October 2013 http://www.standresjournals.org/journals/ssre Research Article Macro-anatomical studies of the African giant pouched rat (Cricetomys
Standard Scientific Research and Essays Vol1 (10): 221-227, October 2013 http://www.standresjournals.org/journals/ssre Research Article Macro-anatomical studies of the African giant pouched rat (Cricetomys
A new species of sauropod, Mamenchisaurus anyuensis sp. nov.
 A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
A new species of sauropod, Mamenchisaurus anyuensis sp. nov. by Xinlu He, Suihua Yang, Kaiji Cai, Kui Li, and Zongwen Liu Chengdu University of Technology Papers on Geosciences Contributed to the 30th
Skulls & Evolution. 14,000 ya cro-magnon. 300,000 ya Homo sapiens. 2 Ma Homo habilis A. boisei A. robustus A. africanus
 Skulls & Evolution Purpose To illustrate trends in the evolution of humans. To demonstrate what you can learn from bones & fossils. To show the adaptations of various mammals to different habitats and
Skulls & Evolution Purpose To illustrate trends in the evolution of humans. To demonstrate what you can learn from bones & fossils. To show the adaptations of various mammals to different habitats and
CENE RUMINANTS OF THE GENERA OVIBOS AND
 DESCRIPTIONS OF TWO NEW SPECIES OF PLEISTO- CENE RUMINANTS OF THE GENERA OVIBOS AND BOOTHERIUM, WITH NOTES ON THE LATTER GENUS. By James Williams Gidley, Of the United States National Museum. Two interesting
DESCRIPTIONS OF TWO NEW SPECIES OF PLEISTO- CENE RUMINANTS OF THE GENERA OVIBOS AND BOOTHERIUM, WITH NOTES ON THE LATTER GENUS. By James Williams Gidley, Of the United States National Museum. Two interesting
THE SKULLS OF ARAEOSCELIS AND CASEA, PERMIAN REPTILES
 THE SKULLS OF REOSCELIS ND CSE, PERMIN REPTILES University of Chicago There are few Permian reptiles of greater interest at the present time than the peculiar one I briefly described in this journal' three
THE SKULLS OF REOSCELIS ND CSE, PERMIN REPTILES University of Chicago There are few Permian reptiles of greater interest at the present time than the peculiar one I briefly described in this journal' three
v:ii-ixi, 'i':;iisimvi'\>!i-:: "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi L I E) R.ARY OF THE VERSITY U N I or ILLINOIS REMO
 R.ARY OF THE VERSITY U N I or ILLINOIS REMO") "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
"^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
Comparative Vertebrate Anatomy
 Comparative Vertebrate Anatomy Presented by BIOBUGS: Biology Inquiry and Outreach with Boston University Graduate Students In association with LERNet and The BU Biology Teaching Laboratory Designed and
Comparative Vertebrate Anatomy Presented by BIOBUGS: Biology Inquiry and Outreach with Boston University Graduate Students In association with LERNet and The BU Biology Teaching Laboratory Designed and
FSS OPEN SHOW PROCEDURAL EXAM
 Judging Operations Department PO Box 900062 Raleigh, NC 27675-9062 (919) 816-3570 judgingops@akc.org www.akc.org Revised Sept 2013 FSS OPEN SHOW PROCEDURAL EXAM Refer to Rules, Policies and Guidelines
Judging Operations Department PO Box 900062 Raleigh, NC 27675-9062 (919) 816-3570 judgingops@akc.org www.akc.org Revised Sept 2013 FSS OPEN SHOW PROCEDURAL EXAM Refer to Rules, Policies and Guidelines
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the
 Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the") Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
On the morphoplogy and taxonomic status of Xinpusaurus kohi JIANG et al., 2004 (Diapsida: Thalattosauria) from the Upper Triassic of China
 from the Upper Triassic of China") Palaeodiversity 7: 47 59; Stuttgart 30 December 2014. 47 On the morphoplogy and taxonomic status of Xinpusaurus kohi JIANG et al., 2004 (Diapsida: Thalattosauria) from the Upper Triassic of China MICHAEL
Palaeodiversity 7: 47 59; Stuttgart 30 December 2014. 47 On the morphoplogy and taxonomic status of Xinpusaurus kohi JIANG et al., 2004 (Diapsida: Thalattosauria) from the Upper Triassic of China MICHAEL
There has bdeno complete account published of the osteology of this bird, and certainly no good figures of its skeleton, though I
 358 SHUFELDT, Osteology of the Passenger Pigeon. [ Auk I_July cornfield." About this same time, James Stuart in a journey from Montgomery to Mobile finds the "wild turkey abounds in these (Chattahoochee
358 SHUFELDT, Osteology of the Passenger Pigeon. [ Auk I_July cornfield." About this same time, James Stuart in a journey from Montgomery to Mobile finds the "wild turkey abounds in these (Chattahoochee
A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev Doklady Akademii Nauk, SSSR 87:
 translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
translated by Dr. Tamara and F. Jeletzky, 1956 A NEW ANKYLOSAUR FROM THE UPPER CRETACEOUS OF MONGOLIA E.A. Maleev 1952. Doklady Akademii Nauk, SSSR 87:273-276 Armored dinosaurs make a considerable part
A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China
 SUPPLEMENTARY INFORMATION A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China Ya-Ming Wang 1, Hai-Lu You 2,3 *, Tao Wang 4 1 School of Earth Sciences and Resources, China
SUPPLEMENTARY INFORMATION A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China Ya-Ming Wang 1, Hai-Lu You 2,3 *, Tao Wang 4 1 School of Earth Sciences and Resources, China
TRACHEMYS SCULPTA. A nearly complete articulated carapace and plastron of an Emjdd A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE,
 A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
A NEAKLY COMPLETE SHELL OF THE EXTINCT TURTLE, TRACHEMYS SCULPTA By Charles W. Gilmore Curator of Vertebrate Paleontology, United States National Museum INTRODUCTION A nearly complete articulated carapace
SUPPLEMENTARY ONLINE MATERIAL FOR. Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor
 http://app.pan.pl/som/app61-ratsimbaholison_etal_som.pdf SUPPLEMENTARY ONLINE MATERIAL FOR Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor Ontogenetic changes in the craniomandibular
http://app.pan.pl/som/app61-ratsimbaholison_etal_som.pdf SUPPLEMENTARY ONLINE MATERIAL FOR Nirina O. Ratsimbaholison, Ryan N. Felice, and Patrick M. O connor Ontogenetic changes in the craniomandibular
Fish 2/26/13. Chordates 2. Sharks and Rays (about 470 species) Sharks etc Bony fish. Tetrapods. Osteichthans Lobe fins and lungfish
 Sharks etc Bony fish. Tetrapods. Osteichthans Lobe fins and lungfish") Chordates 2 Sharks etc Bony fish Osteichthans Lobe fins and lungfish Tetrapods ns Reptiles Birds Feb 27, 2013 Chordates ANCESTRAL DEUTEROSTOME Notochord Common ancestor of chordates Head Vertebral column
Chordates 2 Sharks etc Bony fish Osteichthans Lobe fins and lungfish Tetrapods ns Reptiles Birds Feb 27, 2013 Chordates ANCESTRAL DEUTEROSTOME Notochord Common ancestor of chordates Head Vertebral column
ZOOLOGISCHE MEDEDELINGEN
 MINISTERIE VAN ONDERWIJS, KUNSTEN EN WETENSCHAPPEN ZOOLOGISCHE MEDEDELINGEN UITGEGEVEN DOOR HET RIJKSMUSEUM VAN NATUURLIJKE HISTORIE TE LEIDEN DEEL XXXVII, No. 10 10 juli 1961 THE FOSSIL HIPPOPOTAMUS FROM
MINISTERIE VAN ONDERWIJS, KUNSTEN EN WETENSCHAPPEN ZOOLOGISCHE MEDEDELINGEN UITGEGEVEN DOOR HET RIJKSMUSEUM VAN NATUURLIJKE HISTORIE TE LEIDEN DEEL XXXVII, No. 10 10 juli 1961 THE FOSSIL HIPPOPOTAMUS FROM
Sustainable Resources 11. Poultry Unit: Chicken Anatomy
 Sustainable Resources 11 Poultry Unit: Chicken Anatomy The Chicken Birds: Class AVES are winged, bipedal, endothermic (warm-blooded), egg-laying, vertebrates. Chicken: Gallus gallus are a domesticated
Sustainable Resources 11 Poultry Unit: Chicken Anatomy The Chicken Birds: Class AVES are winged, bipedal, endothermic (warm-blooded), egg-laying, vertebrates. Chicken: Gallus gallus are a domesticated
'Rain' of dead birds on central NJ lawns explained; Federal culling program killed up to 5,000 Associated Press, January 27, 2009
 'Rain' of dead birds on central NJ lawns explained; Federal culling program killed up to 5,000 Associated Press, January 27, 2009 Study May Give Hope That Ivory-billed Woodpeckers Still Around Science
'Rain' of dead birds on central NJ lawns explained; Federal culling program killed up to 5,000 Associated Press, January 27, 2009 Study May Give Hope That Ivory-billed Woodpeckers Still Around Science
Lab 2 Skeletons and Locomotion
 Lab 2 Skeletons and Locomotion Objectives The objectives of this and next week's labs are to introduce you to the comparative skeletal anatomy of vertebrates. As you examine the skeleton of each lineage,
Lab 2 Skeletons and Locomotion Objectives The objectives of this and next week's labs are to introduce you to the comparative skeletal anatomy of vertebrates. As you examine the skeleton of each lineage,
The family Gnaphosidae is a large family
 Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
Pakistan J. Zool., vol. 36(4), pp. 307-312, 2004. New Species of Zelotus Spider (Araneae: Gnaphosidae) from Pakistan ABIDA BUTT AND M.A. BEG Department of Zoology, University of Agriculture, Faisalabad,
Characteristics of Tetrapods
 Marine Tetrapods Characteristics of Tetrapods Tetrapod = four-footed Reptiles, Birds, & Mammals No marine species of amphibian Air-breathing lungs Class Reptilia Saltwater Crocodiles, Sea turtles, sea
Marine Tetrapods Characteristics of Tetrapods Tetrapod = four-footed Reptiles, Birds, & Mammals No marine species of amphibian Air-breathing lungs Class Reptilia Saltwater Crocodiles, Sea turtles, sea
Monodon monoceros Linnaeus, 1758 MONO Mono 1 NAR
 click for previous page 74 Marine Mammals of the World Monodon monoceros Linnaeus, 1758 MONO Mono 1 NAR FAO Names: En - Narwhal; Fr - Narval; Sp - Narval. Fig. 205 Monodon monoceros Distinctive Characteristics:
click for previous page 74 Marine Mammals of the World Monodon monoceros Linnaeus, 1758 MONO Mono 1 NAR FAO Names: En - Narwhal; Fr - Narval; Sp - Narval. Fig. 205 Monodon monoceros Distinctive Characteristics:
List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to
 1 Supplementary data CHARACTER LIST List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to characters used by Tchernov et al. (2000), Rieppel, et al. (2002), and Lee
1 Supplementary data CHARACTER LIST List of characters used in the phylogenetic analysis. Capital letters T, R, and L, refer to characters used by Tchernov et al. (2000), Rieppel, et al. (2002), and Lee
Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons
 Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons 1. Head skeleton of lamprey Cyclostomes are highly specialized in both the construction of the chondrocranium and visceral skeleton.
Biology 3315 Comparative Vertebrate Morphology Skulls and Visceral Skeletons 1. Head skeleton of lamprey Cyclostomes are highly specialized in both the construction of the chondrocranium and visceral skeleton.
A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province
 A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province by Hu Shaojin (Kunming Cultural Administrative Committee, Yunnan Province) Vertebrata PalAsiatica Vol. XXXI, No. 1
A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province by Hu Shaojin (Kunming Cultural Administrative Committee, Yunnan Province) Vertebrata PalAsiatica Vol. XXXI, No. 1
Frog Dissection Information Manuel
 Frog Dissection Information Manuel Anatomical Terms: Used to explain directions and orientation of a organism Directions or Positions: Anterior (cranial)- toward the head Posterior (caudal)- towards the
Frog Dissection Information Manuel Anatomical Terms: Used to explain directions and orientation of a organism Directions or Positions: Anterior (cranial)- toward the head Posterior (caudal)- towards the
Vertebrates. skull ribs vertebral column
 Vertebrates skull ribs vertebral column endoskeleton in cells working together tissues tissues working together organs working together organs systems Blood carries oxygen to the cells carries nutrients
Vertebrates skull ribs vertebral column endoskeleton in cells working together tissues tissues working together organs working together organs systems Blood carries oxygen to the cells carries nutrients
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province
 from Dashanpu, Zigong Municipality, Sichuan Province") A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
A new species of Hsisosuchus (Mesoeucrocodylia) from Dashanpu, Zigong Municipality, Sichuan Province Yuhui Gao (Zigong Dinosaur Museum) Vertebrata PalAsiatica Volume 39, No. 3 July, 2001 pp. 177-184 Translated
Temporal lines. More forwardfacing. tubular orbits than in the African forms 3. Orbits larger relative to skull size than in the other genera 2.
 Asian lorises More forwardfacing and tubular orbits than in the African forms 3. Characterized by a marked extension of the ectotympanic into a tubular meatus and a more angular auditory bulla than in
Asian lorises More forwardfacing and tubular orbits than in the African forms 3. Characterized by a marked extension of the ectotympanic into a tubular meatus and a more angular auditory bulla than in
A M E G H I N I A N A. Revista de la Asociación Paleontológia Argentina. Volume XV September-December 1978 Nos. 3-4
 A M E G H I N I A N A Revista de la Asociación Paleontológia Argentina Volume XV September-December 1978 Nos. 3-4 COLORADIA BREVIS N. G. ET N. SP. (SAURISCHIA, PROSAUROPODA), A PLATEOSAURID DINOSAUR FROM
A M E G H I N I A N A Revista de la Asociación Paleontológia Argentina Volume XV September-December 1978 Nos. 3-4 COLORADIA BREVIS N. G. ET N. SP. (SAURISCHIA, PROSAUROPODA), A PLATEOSAURID DINOSAUR FROM
Oribatid Mites of the Family Otocepheidae from Tian-mu Mountain in China (Acari: Oribatida)1'
1'") Acta arachnol,, 42 (1): 1-6, August 30, 1993 Oribatid Mites of the Family Otocepheidae from Tian-mu Mountain in China (Acari: Oribatida)1' Jun-ichi AoKI2' and Sheng-hao Hu3' Abstract Dolicheremaeus wangi
Acta arachnol,, 42 (1): 1-6, August 30, 1993 Oribatid Mites of the Family Otocepheidae from Tian-mu Mountain in China (Acari: Oribatida)1' Jun-ichi AoKI2' and Sheng-hao Hu3' Abstract Dolicheremaeus wangi
1/9/2013. Divisions of the Skeleton: Topic 8: Appendicular Skeleton. Appendicular Components. Appendicular Components
 /9/203 Topic 8: Appendicular Skeleton Divisions of the Skeleton: Cranial Postcranial What makes up the appendicular skeleton? What is the pattern of serial homology of the limbs? Tetrapod front limb morphology
/9/203 Topic 8: Appendicular Skeleton Divisions of the Skeleton: Cranial Postcranial What makes up the appendicular skeleton? What is the pattern of serial homology of the limbs? Tetrapod front limb morphology
1) Explain why the skeleton plays an important role in the overall shape of animal and human being.
 Explain why the skeleton plays an important role in the overall shape of animal and human being.") 1) Explain why the skeleton plays an important role in the overall shape of animal and human being. 2) Substantiate the differences in animal and human skeleton, with the human skeleton built in such a
1) Explain why the skeleton plays an important role in the overall shape of animal and human being. 2) Substantiate the differences in animal and human skeleton, with the human skeleton built in such a
Comparative Osteology of the Genus Pachytriton (Caudata: Salamandridae) from Southeastern China
 from Southeastern China") Asian Herpetological Research 2012, 3(2): 83 102 DOI: 10.3724/SP.J.1245.2012.00083 Comparative Osteology of the Genus Pachytriton (Caudata: Salamandridae) from Southeastern China Yunke WU 1, Yuezhao WANG
Asian Herpetological Research 2012, 3(2): 83 102 DOI: 10.3724/SP.J.1245.2012.00083 Comparative Osteology of the Genus Pachytriton (Caudata: Salamandridae) from Southeastern China Yunke WU 1, Yuezhao WANG
290 SHUFELDT, Remains of Hesperornis.
 290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
290 SHUFELDT, Remains of Hesperornis. [ Auk [July THE FOSSIL REMAINS OF A SPECIES OF HESPERORNIS FOUND IN MONTANA. BY R. W. SHUFELD% M.D. Plate XI7III. ExR,¾ in November, 1914, Mr. Charles W. Gihnore,
WHxVLEBONE WHALE FROM THE CALVERT CLIFFS, MARYLAND.
 DESCRIPTION OF A NEW GENUS AND SPECIES OF WHxVLEBONE WHALE FROM THE CALVERT CLIFFS, MARYLAND. By Remington Kellogg, Of the Bureau of Biological Survey, United States Departm'ent of Agriculture. In the
DESCRIPTION OF A NEW GENUS AND SPECIES OF WHxVLEBONE WHALE FROM THE CALVERT CLIFFS, MARYLAND. By Remington Kellogg, Of the Bureau of Biological Survey, United States Departm'ent of Agriculture. In the
MIOCENE DIATOMACEOUS EARTH OF LOMPOC, CALI- FORNIA.i
 DESCRIPTION OF THE SKULL OF MEGAPTERA MIO- CAENA, A FOSSIL HUMPBACK WHALE FROM THE MIOCENE DIATOMACEOUS EARTH OF LOMPOC, CALI- FORNIA.i By Remington Kellogg. Of the Bureau of Biological Survey, Department
DESCRIPTION OF THE SKULL OF MEGAPTERA MIO- CAENA, A FOSSIL HUMPBACK WHALE FROM THE MIOCENE DIATOMACEOUS EARTH OF LOMPOC, CALI- FORNIA.i By Remington Kellogg. Of the Bureau of Biological Survey, Department
HONR219D Due 3/29/16 Homework VI
 Part 1: Yet More Vertebrate Anatomy!!! HONR219D Due 3/29/16 Homework VI Part 1 builds on homework V by examining the skull in even greater detail. We start with the some of the important bones (thankfully
Part 1: Yet More Vertebrate Anatomy!!! HONR219D Due 3/29/16 Homework VI Part 1 builds on homework V by examining the skull in even greater detail. We start with the some of the important bones (thankfully
XXI.- ON TWO NEW SPECIES OI"~ EAGLE RAYS (MYLIOBATIDlE), WITH NOTES ON THE SKULL OF THE GENUS CERATOPTERA.
, WITH NOTES ON THE SKULL OF THE GENUS CERATOPTERA.") XXI.- ON TWO NEW SPECIES OI~ EAGLE RAYS (MYLIOBATIDlE), WITH NOTES ON THE SKULL OF THE GENUS CERATOPTERA. By R. E. LLOYD, M.B., B.Sc., Capt., I.M.S., formerly Surgeon Naturalist, Marine Survey of India.
XXI.- ON TWO NEW SPECIES OI~ EAGLE RAYS (MYLIOBATIDlE), WITH NOTES ON THE SKULL OF THE GENUS CERATOPTERA. By R. E. LLOYD, M.B., B.Sc., Capt., I.M.S., formerly Surgeon Naturalist, Marine Survey of India.
Letting rotting sharks lie: further evidence for shark identity of the Zuiyo-maru carcass
 Letting rotting sharks lie: further evidence for shark identity of the Zuiyo-maru carcass Pierre Jerlström and Bev Elliott Another basking shark carcass washed up on the New Zealand coast reveals features
Letting rotting sharks lie: further evidence for shark identity of the Zuiyo-maru carcass Pierre Jerlström and Bev Elliott Another basking shark carcass washed up on the New Zealand coast reveals features
A NEW SALTICID SPIDER FROM VICTORIA By R. A. Dunn
 Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
Dunn, R. A. 1947. A new salticid spider from Victoria. Memoirs of the National Museum of Victoria 15: 82 85. All text not included in the original document is highlighted in red. Mem. Nat. Mus. Vict.,
SUPPLEMENTARY OBSERVATIONS ON THE SKULL OF
 SUPPLEMENTARY OBSERVATIONS ON THE SKULL OF THE FOSSIL PORPOISE ZARHACHIS FLAGELLATOR COPE By Remington Kellogg Of the Bureau of Biological Survey, United States Department of Agriculture During the past
SUPPLEMENTARY OBSERVATIONS ON THE SKULL OF THE FOSSIL PORPOISE ZARHACHIS FLAGELLATOR COPE By Remington Kellogg Of the Bureau of Biological Survey, United States Department of Agriculture During the past
Name Class Date. After you read this section, you should be able to answer these questions:
 CHAPTER 14 4 Vertebrates SECTION Introduction to Animals BEFORE YOU READ After you read this section, you should be able to answer these questions: How are vertebrates different from invertebrates? How
CHAPTER 14 4 Vertebrates SECTION Introduction to Animals BEFORE YOU READ After you read this section, you should be able to answer these questions: How are vertebrates different from invertebrates? How
Endangered Species Origami
 Endangered Species Origami For most of the wild things on Earth, the future must depend upon the conscience of mankind ~ Dr. Archie Carr, father of modern marine turtle biology and conservation Humpback
Endangered Species Origami For most of the wild things on Earth, the future must depend upon the conscience of mankind ~ Dr. Archie Carr, father of modern marine turtle biology and conservation Humpback
2. Skull, total length versus length of the presacral vertebral column: (0); extremely elongated neck (e.g. Tanystropheus longobardicus).
; extremely elongated neck (e.g. Tanystropheus longobardicus).") Character list of the taxon-character data set 1. Skull and lower jaws, interdental plates: absent (0); present, but restricted to the anterior end of the dentary (1); present along the entire alveolar
Character list of the taxon-character data set 1. Skull and lower jaws, interdental plates: absent (0); present, but restricted to the anterior end of the dentary (1); present along the entire alveolar
Who Really Owns the Beach? The Competition Between Sea Turtles and the Coast Renee C. Cohen
 Who Really Owns the Beach? The Competition Between Sea Turtles and the Coast Renee C. Cohen Some Common Questions Microsoft Word Document This is an outline of the speaker s notes in Word What are some
Who Really Owns the Beach? The Competition Between Sea Turtles and the Coast Renee C. Cohen Some Common Questions Microsoft Word Document This is an outline of the speaker s notes in Word What are some
Madagascar, which entirely agree with one another. Rumph. specimens of. (1. c. pl. III, fig. 4). This species may be distinguished
. This species may be distinguished") UELA3IMUS MARIONJS. 67 NOTE XIII. On some species of Gelasimus Latr. and Macrophthalmus Latr. BY J.G. de Man March 1880. Gelasimus vocans Rumph. Milne Edwards, Observ. sur la classification des Crustacea,
UELA3IMUS MARIONJS. 67 NOTE XIII. On some species of Gelasimus Latr. and Macrophthalmus Latr. BY J.G. de Man March 1880. Gelasimus vocans Rumph. Milne Edwards, Observ. sur la classification des Crustacea,
UN? RSITYOF. ILLIiwiS LIBRARY AT URBANA-CHAMPAIGN NATURAL HIST. SURVEY
 UN? RSITYOF ILLIiwiS LIBRARY AT URBANA-CHAMPAIGN NATURAL HIST. SURVEY FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 July 29, 1954 No. 17 FAUNA OF THE VALE AND CHOZA: 7 PELYCOSAURIA:
UN? RSITYOF ILLIiwiS LIBRARY AT URBANA-CHAMPAIGN NATURAL HIST. SURVEY FIELDIANA GEOLOGY Published by CHICAGO NATURAL HISTORY MUSEUM Volume 10 July 29, 1954 No. 17 FAUNA OF THE VALE AND CHOZA: 7 PELYCOSAURIA:
A DESCRIPTION OF CALLIANASSA MARTENSI MIERS, 1884 (DECAPODA, THALASSINIDEA) AND ITS OCCURRENCE IN THE NORTHERN ARABIAN SEA
 AND ITS OCCURRENCE IN THE NORTHERN ARABIAN SEA") Crustaceana 26 (3), 1974- E. J. BiiU, Leide A DESCRIPTION OF CALLIANASSA MARTENSI MIERS, 1884 (DECAPODA, THALASSINIDEA) AND ITS OCCURRENCE IN THE NORTHERN ARABIAN SEA BY NASIMA M. TIRMIZI Invertebrate
Crustaceana 26 (3), 1974- E. J. BiiU, Leide A DESCRIPTION OF CALLIANASSA MARTENSI MIERS, 1884 (DECAPODA, THALASSINIDEA) AND ITS OCCURRENCE IN THE NORTHERN ARABIAN SEA BY NASIMA M. TIRMIZI Invertebrate
SUPPLEMENTARY INFORMATION
 Character 155, interdental ridges. Absence of interdental ridge (0) shown in Parasaniwa wyomingensis (Platynota). Interdental ridges (1) shown in Coniophis precedens. WWW.NATURE.COM/NATURE 1 Character
Character 155, interdental ridges. Absence of interdental ridge (0) shown in Parasaniwa wyomingensis (Platynota). Interdental ridges (1) shown in Coniophis precedens. WWW.NATURE.COM/NATURE 1 Character
WHALES. Marine Discovery Centre, Henley Beach, S.A.
 WHALES Marine Discovery Centre, Henley Beach, S.A. What is a whale? Whales are large, magnificent, intelligent, aquatic mammals. They breathe air through blowhole(s) into lungs (unlike fish who breathe
WHALES Marine Discovery Centre, Henley Beach, S.A. What is a whale? Whales are large, magnificent, intelligent, aquatic mammals. They breathe air through blowhole(s) into lungs (unlike fish who breathe
Recently Mr. Lawrence M. Lambe has described and figured in the
 56.81,9C(117:71.2) Article XXXV.-CORYTHOSAURUS CASUARIUS, A NEW CRESTED DINOSAUR FROM THE BELLY RIVER CRETA- CEOUS, WITH PROVISIONAL CLASSIFICATION OF THE FAMILY TRACHODONTIDA1X BY BARNUM BROWN. PLATE
56.81,9C(117:71.2) Article XXXV.-CORYTHOSAURUS CASUARIUS, A NEW CRESTED DINOSAUR FROM THE BELLY RIVER CRETA- CEOUS, WITH PROVISIONAL CLASSIFICATION OF THE FAMILY TRACHODONTIDA1X BY BARNUM BROWN. PLATE
Page # Diversity of Arthropoda Crustacea Morphology. Diversity of Arthropoda. Diversity of Arthropoda. Diversity of Arthropoda. Arthropods, from last
 Arthropods, from last time Crustacea are the dominant marine arthropods Crustacea are the dominant marine arthropods any terrestrial crustaceans? Should we call them shellfish? sowbugs 2 3 Crustacea Morphology
Arthropods, from last time Crustacea are the dominant marine arthropods Crustacea are the dominant marine arthropods any terrestrial crustaceans? Should we call them shellfish? sowbugs 2 3 Crustacea Morphology
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE
 FROM THE") A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
A NEW AUSTROSQUILLA (STOMATOPODA) FROM THE MARQUESAS ISLANDS BY ALAIN MICHEL Centre O.R.S.T.O.M., Noumea, New Caledonia and RAYMOND B. MANNING Smithsonian Institution, Washington, U.S.A. The At s,tstrosqzlilla
FROG DISSECTION. a. Why is there a difference in size proportion between the hind and fore limbs?
 FROG DISSECTION External Anatomy 1. The division of a frog s body includes the head, trunk and limbs. Examine the front and hind limbs of the frog. The hind limbs are the long, more muscular limbs of the
FROG DISSECTION External Anatomy 1. The division of a frog s body includes the head, trunk and limbs. Examine the front and hind limbs of the frog. The hind limbs are the long, more muscular limbs of the
OF THE TRIAS THE PHYTOSAURIA
 THE PHYTOSAURIA OF THE TRIAS MAURICE G. MEHL University of Wisconsin Some time ago the writer gave a brief notice of a new genus of phytosaurs of which Angistorhinus grandis Mehl was the type.' It is the
THE PHYTOSAURIA OF THE TRIAS MAURICE G. MEHL University of Wisconsin Some time ago the writer gave a brief notice of a new genus of phytosaurs of which Angistorhinus grandis Mehl was the type.' It is the
NIVOROUS DINOSAUR. (SECOND COMMUNICATION.) By HENRY FAIRFIELD OSBORN. PLATE XXXIX. This great carnivorous Dinosaur of the Laramie was contemporary
 By HENRY FAIRFIELD OSBORN. PLATE XXXIX. This great carnivorous Dinosaur of the Laramie was contemporary") 56, 8i, 9 T (I 7: 786) Article VI.-TYRANNOSAURUS, UPPER CRETACEOUS CAR- NIVOROUS DINOSAUR. (SECOND COMMUNICATION.) By HENRY FAIRFIELD OSBORN. PLATE I. This great carnivorous Dinosaur of the Laramie was
56, 8i, 9 T (I 7: 786) Article VI.-TYRANNOSAURUS, UPPER CRETACEOUS CAR- NIVOROUS DINOSAUR. (SECOND COMMUNICATION.) By HENRY FAIRFIELD OSBORN. PLATE I. This great carnivorous Dinosaur of the Laramie was
A NEW SPECIES OF BEAKED WHALE MESOPLODON PERRINI SP. N. (CETACEA: ZIPHIIDAE) DISCOVERED THROUGH PHYLOGENETIC ANALYSES OF MITOCHONDRIAL DNA SEQUENCES
 DISCOVERED THROUGH PHYLOGENETIC ANALYSES OF MITOCHONDRIAL DNA SEQUENCES") MARINE MAMMAL SCIENCE, 18(3):577 608 (July 2002) 2002 by the Society for Marine Mammalogy A NEW SPECIES OF BEAKED WHALE MESOPLODON PERRINI SP. N. (CETACEA: ZIPHIIDAE) DISCOVERED THROUGH PHYLOGENETIC ANALYSES
MARINE MAMMAL SCIENCE, 18(3):577 608 (July 2002) 2002 by the Society for Marine Mammalogy A NEW SPECIES OF BEAKED WHALE MESOPLODON PERRINI SP. N. (CETACEA: ZIPHIIDAE) DISCOVERED THROUGH PHYLOGENETIC ANALYSES
On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds
 and the origin of birds") On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds by Qiang Ji and Shu an Ji Chinese Geological Museum, Beijing Chinese Geology Volume 233 1996 pp.
On the Discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds by Qiang Ji and Shu an Ji Chinese Geological Museum, Beijing Chinese Geology Volume 233 1996 pp.
LEIDY, SHOWING THE BONES OF THE FEET 'AND LIMBS
 CQNTEUBUTIONS FBOM THE MUSEUM OF PALEONTOLOGY (Confindion of Con&&&m froin UB Muaercm of Gcologg) UNIVERSITY OF ' MICHIGAN VOL V, No. 6, pp. 6W3 (e ph.) DEAXMBER 31,1036 A SPECIMEN OF STYLEMYS NEBRASCENSIS
CQNTEUBUTIONS FBOM THE MUSEUM OF PALEONTOLOGY (Confindion of Con&&&m froin UB Muaercm of Gcologg) UNIVERSITY OF ' MICHIGAN VOL V, No. 6, pp. 6W3 (e ph.) DEAXMBER 31,1036 A SPECIMEN OF STYLEMYS NEBRASCENSIS
Field Trip: Harvard Museum of Natural History (HMNH)
") Field Trip: Harvard Museum of Natural History (HMNH) Objectives To observe the diversity of animals. To compare and contrast the various adaptations, body plans, etc. of the animals found at the HMNH.
Field Trip: Harvard Museum of Natural History (HMNH) Objectives To observe the diversity of animals. To compare and contrast the various adaptations, body plans, etc. of the animals found at the HMNH.
Appendix chapter 2: Description of Coloborhynchus spielbergi sp. nov. (Pterodactyloidea) from the Albian (Lower Cretaceous) of Brazil
 from the Albian (Lower Cretaceous) of Brazil") Appendix chapter 2: Description of Coloborhynchus spielbergi sp. nov. (Pterodactyloidea) from the Albian (Lower Cretaceous) of Brazil Appendix chapter 2 155 2.7. Appendix 2.7.1. Measurements Skull 15 12
Appendix chapter 2: Description of Coloborhynchus spielbergi sp. nov. (Pterodactyloidea) from the Albian (Lower Cretaceous) of Brazil Appendix chapter 2 155 2.7. Appendix 2.7.1. Measurements Skull 15 12
Diurus, Pascoe. sp. 1). declivity of the elytra, but distinguished. Length (the rostrum and tails 26 included) mm. Deep. exception
. declivity of the elytra, but distinguished. Length (the rostrum and tails 26 included) mm. Deep. exception") 210 DIURUS ERYTIIROPUS. NOTE XXVI. Three new species of the Brenthid genus Diurus, Pascoe DESCRIBED BY C. Ritsema+Cz. 1. Diurus erythropus, n. sp. 1). Allied to D. furcillatus Gylh. ²) by the short head,
210 DIURUS ERYTIIROPUS. NOTE XXVI. Three new species of the Brenthid genus Diurus, Pascoe DESCRIBED BY C. Ritsema+Cz. 1. Diurus erythropus, n. sp. 1). Allied to D. furcillatus Gylh. ²) by the short head,