Phenotypic modulation of the Bvg+ phase is not required for pathogenesis and. transmission of Bordetella bronchiseptica in swine
|
|
|
- Nathaniel Morris
- 5 years ago
- Views:
Transcription
1 IAI Accepts, published online ahead of print on 12 December 2011 Infect. Immun. doi: /iai Copyright 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. 1 2 Phenotypic modulation of the Bvg+ phase is not required for pathogenesis and transmission of Bordetella bronchiseptica in swine Tracy L. Nicholson 1*, Susan L. Brockmeier 1, Crystal L. Loving 1, Karen B. Register 1, Marcus E. Kehrli, Jr. 1, Scott E. Stibitz 2, and Sarah M. Shore 1 1 National Animal Disease Center, Agricultural Research Service, USDA, Ames, IA, 50010, USA. 2 Division of Bacterial, Parasitic, and Allergenic Products, Center for Biologics Evaluation and Research, Food and Drug Administration, Bethesda, MD, 20892, USA. * Correspondence author: Tracy L. Nicholson, Ph.D. National Animal Disease Center, ARS, USDA P.O. Box Dayton Ave. Ames, Iowa Telephone: Fax: tracy.nicholson@ars.usda.gov Running title: Bvg + phase is sufficient for infection and transmission in swine 23 1
2 24 Abstract The majority of virulence gene expression in Bordetella is regulated by a two-component sensory transduction system encoded by the bvg locus. In response to environmental cues, the BvgAS regulatory system controls expression of a spectrum of phenotypic phases transitioning between a virulent (Bvg + ) phase and a non-virulent (Bvg ) phase, a process referred to as phenotypic modulation. We hypothesized that the ability of B. bronchiseptica to undergo phenotypic modulation is required at one or more points during the infectious cycle in swine. To investigate the Bvg phase-dependent contribution to pathogenesis of B. bronchiseptica in swine, we constructed a series of isogenic mutants in a virulent B. bronchiseptica swine isolate and compared each mutant to the wild-type isolate, for its ability to colonize and cause disease. We additionally tested whether a BvgAS system capable of modulation is required for direct or indirect transmission. The Bvg phase-locked mutant was never recovered from any respiratory tract site at any time point examined. An intermediate phase-locked mutant (Bvg i ) was found in numbers lower than wild-type at all respiratory tract sites and time points examined and caused limited to no disease. In contrast, colonization of the respiratory tract and disease caused by the Bvg + phase-locked mutant and the wild-type strain were indistinguishable. The Bvg + phaselocked mutant transmitted to naïve pigs by both direct and indirect contact with equal efficiency as the wild-type isolate. These results indicate that while full activation of the BvgAS regulatory system is required for colonization and severe disease, it is not deleterious to direct and indirect transmission. Overall, our results demonstrate that the Bvg + phase is sufficient for respiratory infection and host-to-host transmission of B. bronchiseptica in swine. 45 2
3 46 Introduction Respiratory disease in pigs is a serious concern for swine producers today. The most recent survey conducted by the National Animal Health Monitoring System (NAHMS) found that respiratory problems are a major cause of mortality in swine herds, with 53.7% of nursery pig deaths and 60.1% of grower-finisher pig deaths attributed to respiratory problems (36). Bordetella bronchiseptica is widespread in swine populations and is an important contributor to respiratory disease in pigs. In young pigs, it is a primary cause of bronchopneumonia and in older pigs contributes to secondary pneumonia. It is the primary etiologic agent of nonprogressive atrophic rhinitis, a mild to moderately severe reversible condition, and it promotes colonization by toxigenic strains of Pasteurella multocida, which leads to severe progressive atrophic rhinitis (11, 22). In pigs with pneumonia, B. bronchiseptica is often isolated in combination with other pathogens (29). Numerous studies have demonstrated that co-infection with B. bronchiseptica increases colonization and exacerbates the severity of disease caused by both viral and bacterial pathogens, including swine influenza virus (SIV), porcine reproductive and respiratory syndrome virus (PRRSV), porcine respiratory coronavirus (PRCV), Haemophilus parasuis, Pasteurella multocida and Streptococcus suis (3, 5-8, 21, 37, 38). Additionally, there is a growing concern regarding the ability of B. bronchiseptica to inhibit the efficacy of SIV vaccines and to exacerbate enhanced pneumonia in pigs administered an inactivated SIV vaccine followed by challenge with a heterologous virus A universal underlying pathogenic mechanism is shared among Bordetella strains in that the majority of virulence gene expression is regulated by a two-component sensory transduction system encoded by the bvg locus. This locus comprises a histidine kinase sensor protein, BvgS, 3
4 and a DNA-binding response-regulator protein, BvgA. In response to environmental cues, such as temperature, MgSO4, or nicotinic acid concentrations, BvgAS controls expression of a spectrum of phenotypic phases transitioning between a virulent (Bvg + ) phase and a non-virulent (Bvg ) phase, a process referred to as phenotypic modulation. During the virulent Bvg + phase, the BvgAS system is fully active and many of the known virulence factors are expressed, such as filamentous hemagglutinin (FHA), pertactin, fimbriae, adenylate cyclase-hemolysin toxin, and dermonecrotic toxin (DNT), as well as a type III secretion system (TTSS) (12). Conversely, BvgAS is inactive during the Bvg phase, resulting in the maximal expression of motility loci, virulence-repressed genes (vrg genes), and genes required for the production of urease (1, 2, 25). Previous studies involving phase-locked and ectopic expression mutants demonstrated that the Bvg + phase promotes respiratory tract colonization by B. pertussis and B. bronchiseptica (1, 13, 14, 23, 26), while the Bvg phase of B. bronchiseptica promotes survival under conditions of nutrient deprivation, such as those potentially encountered in an environmental reservoir (13, 14). A third phenotypic phase, referred to as Bvg i or intermediate phase, can be induced by addition of MgSO 4 or nicotinic acid at concentrations lower than those needed to fully induce the Bvg phase (14). The Bvg i phase is characterized by the expression of a subset of the Bvg + phase genes (such as fhab and prn) and Bvg i -specific genes, such as bipa and bcfa (14, 33). Many respiratory pathogens are highly contagious and can be transmitted from host to host by direct and indirect means. B. bronchiseptica is highly contagious among mammals, including swine, and experimental direct and airborne transmission of B. bronchiseptica has been documented (4, 35). A proposed role for phenotypic modulation by the BvgAS signal transduction system is to coordinate regulation of gene sets required for survival in the different environments that may be encountered by B. bronchiseptica, including those encountered during 4
5 transmission between hosts. In this model, the Bvg i phase, or other phase intermediates between Bvg i and Bvg +, is proposed to be required for aerosol or indirect transmission (12, 14, 33). However, this frequently proposed hypothesis has never been experimentally addressed. In this report, we investigate the Bvg phase-dependent contribution to pathogenesis of B. bronchiseptica in swine by constructing a series of isogenic mutants in a virulent B. bronchiseptica swine isolate and comparing each mutant to the wild-type isolate for its ability to colonize and cause disease. Additionally, we test whether a BvgAS system capable of modulation is required for direct or indirect transmission. Our results demonstrate that in a natural host, phenotypic modulation of the Bvg + phase is not required for respiratory infection and host-to-host transmission of B. bronchiseptica in swine. Downloaded from on August 31, 2018 by guest 5
6 101 Materials and Methods 102 Bacterial strains and growth conditions B. bronchiseptica strain KM22, isolated from a swine herd with atrophic rhinitis, has been used in a number of studies by our laboratory in which we have shown it is capable of causing atrophic rhinitis and pneumonia in piglets (3-9, 21, 28). KM22 was used for construction of strains TN23, TN30, and TN31. All B. bronchiseptica strains were grown at 37 C on Bordet- Gengou (BG) agar (Difco, Sparks, MD) supplemented with 10% sheep s blood or, in some cases, in Stainer-Scholte (SS) broth. Escherichia coli One Shot TOP10 (Invitrogen, Carlsbad, CA) was used for all cloning steps, and E. coli SM10λpir was used to mobilize plasmids into B. bronchiseptica KM22. When appropriate, antibiotics were included at the following concentrations: carbenicillin, 100 µg/ml; chloramphenicol, 30 µg/ml; kanamycin, 100 µg/ml; and streptomycin, 40 µg/ml. Cloning and construction of B. bronchiseptica KM22 mutants. For construction of TN23, the Bvg+ phase-locked derivative of KM22, the bvgs gene from KM22 was amplified by PCR using primers bvgs-for and bvgs-rev (Table 1). The resulting amplicon was cloned into pcr2.1 (Invitrogen, Carlsbad, CA) to create psms3. The bvgs-c3 (13) mutation R570H was introduced into psms3 using primers R570Hfor and R570Hrev (Table 1) and the QuikChange Site-Directed Mutagenesis Kit (Agilent Technologies, Inc., Santa Clara, CA) according to manufacturer s instructions. The mutation was confirmed by sequencing and the resulting plasmid was named ptn15. Flanking attb sites were added by PCR using primers bvgs-5 -attb1 and bvgs-3 -attb2 (Table 1). The modified bvgs gene was cloned into pdonr201 (Invitrogen, Carlsbad, CA), using the Gateway cloning system (Invitrogen, 6
7 Carlsbad, CA), to obtain psms26. psms26 was recombined with pabb-crs2 (31), which confers sucrose sensitivity, and digested with NcoI and NotI, to obtain psms29 using the Gateway cloning system. psms29 was introduced into E. coli SM10λpir (32) and transconjugated into KM22, as previously described (17). Transconjugates were selected on medium containing streptomycin and chloramphenicol. Cells from a second recombination event, resulting in the excision of the plasmid, were then selected on medium containing sucrose. A single colony, designated TN23, was selected for subsequent use. Confirmation that the introduced mutation resulted in the specified phase-locked phenotype was obtained by growing wild-type KM22 and TN23 on BG agar and BG agar containing MgSO 4 and evaluating colony morphology and hemolytic activity. TN23 was further confirmed by PCR amplification of the bvgs gene and DNA sequence analysis. For TN30, the Bvg phase-locked mutant, a 1,046-bp DNA fragment covering the region immediately 5 of bvgs and extending to codon 7 was amplified by PCR from KM22 genomic DNA using primers bvgs-fa-for and bvgs-fa-rev (Table 1), which were designed such that SacI and XbaI sites would be generated at the 5 and 3 ends, respectively. The resulting PCR product was cloned into pcr2.1 (Invitrogen, Carlsbad, CA) to create ptn31. The insert of ptn31, containing the upstream region of bvgs, was cloned into puc19 (New England Biolabs, Ipswich, MA) via SacI and XbaI sites to obtain ptn39. A 1,097-bp DNA fragment extending from codon 1225 of bvgs into the adjacent downstream region was amplified from KM22 genomic DNA by PCR using primers bvgs-fb-for and bvgs-fb-rev (Table 1), which were designed such that XbaI and HindIII sites would be generated at the 5 and 3 ends, respectively. The resulting PCR product was cloned into pcr2.1 (Invitrogen, Carlsbad, CA) to obtain ptn32. The insert of ptn32, containing the downstream region of bvgs, was excised by XbaI and 7
8 HindIII digestion and cloned into XbaI-HindIII digested ptn39 to create ptn42. The 2,143-bp DNA fragment, containing the upstream and downstream regions of bvgs, was amplified from ptn42 by PCR using primers TN42-for and TN42-rev (Table 1), which were designed such that SacI and AvrII sites would be generated at the 5 and 3 ends, respectively. The resulting PCR product was cloned into pcr2.1 (Invitrogen, Carlsbad, CA) to create ptn46. The insert of ptn46 was excised by SacI and AvrII digestion and cloned into SacI-AvrII digested allelic exchange vector pss4245 (10, 19) to create ptn50. ptn50 was introduced into E. coli SM10λpir (32) and transconjugated into KM22 as previously described (10, 19). Briefly, Bvg phase B. bronchiseptica, obtained by growth on BG agar containing 50 mm MgSO 4, was coincubated with SM10λpir cells carrying ptn50 on BG agar containing 10 mm MgCl 2 and 50 mm MgSO 4 for 4 hours at 37 C. Cointegrants were selected on BG-Sm-Km-Carb agar containing 50 mm MgSO 4. Cointegrants were then streaked onto BG agar and incubated for 2 days at 37 C, allowing for the selection of B. bronchiseptica that had undergone a second recombination event resulting in a loss of the allelic exchange plasmid, ptn50. Colonies were screened for the presence of the mutant bvgs gene by growth on BG-Sm agar and lack of growth on BG-Sm-Km-Carb agar. A single colony, designated TN30, was selected for subsequent use. Confirmation that the introduced mutation resulted in the specified phase-locked phenotype was obtained by growing wild-type KM22 and TN30 on BG agar and BG agar containing MgSO 4 and evaluating colony morphology and hemolytic activity. TN30 was further confirmed by PCR amplification of a DNA fragment encoding upstream and downstream regions of the bvgs gene and DNA sequence analysis TN31, the Bvg i phase-locked derivative of KM22, was constructed by generating the bvgs-i1 (14) mutation T733M using the QuikChange Site-Directed Mutagenesis Kit (Agilent 8
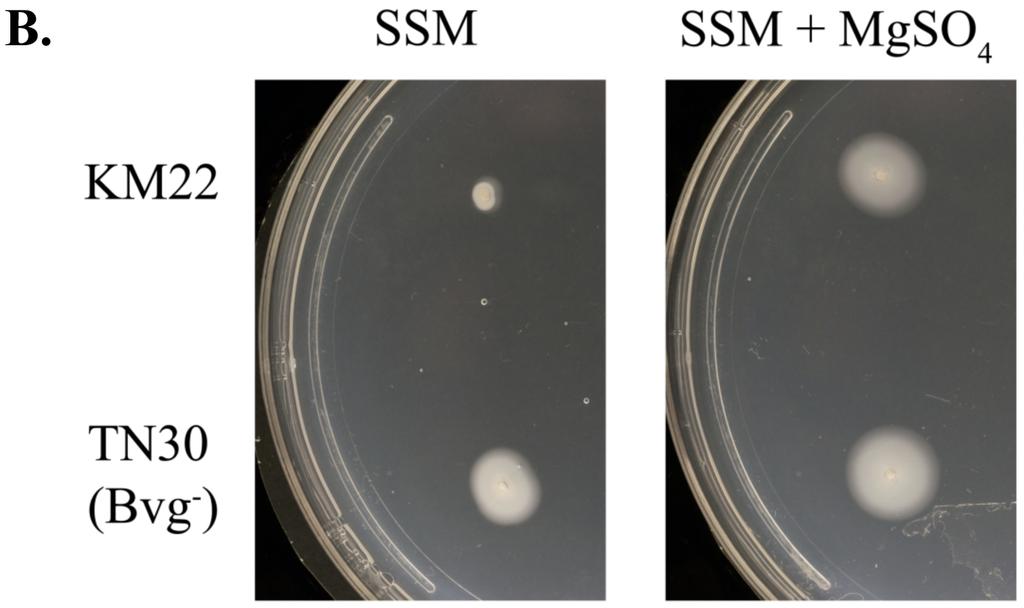
9 Technologies, Inc., Santa Clara, CA) according to manufacturer s instructions. ptn15, carrying the bvgs gene encoding the bvgs-c3 mutation R570H, served as the template for use with mutagenesis primers T733Mfor and T733Mrev (Table 1). The mutation was confirmed by sequencing and the resulting plasmid was named ptn18. The bvgs encoding both R570H and T733M mutations was then PCR amplified from ptn18 using primers TN18-for and TN18-rev (Table 1), which were designed such that AvrII and SpeI sites would be generated at the 5 and 3 ends respectively. The resulting PCR product was cloned into pcr-blunt II-TOPO (Invitrogen, Carlsbad, CA) to create psms87. The insert of psms87 was then excised by AvrII and SpeI digestion and cloned into AvrII-SpeI digested pss4245 to create psms95. psms95 was introduced into E. coli SM10λpir (32) and transconjugated into KM22 as described above for TN30. A single colony, designated TN31, was selected for subsequent use. Confirmation that the introduced mutation resulted in the specified phase-locked phenotype was obtained by growing wild-type KM22 and TN31 on BG agar and BG agar containing MgSO 4 and evaluating colony morphology and hemolytic activity. TN31 was further confirmed by PCR amplification of the bvgs gene and DNA sequence analysis. Motility Assay. Motility assays were performed as previously described (2). Briefly, plates prepared with Stainer-Scholte medium (SSM) containing 0.35% agarose, supplemented with 50 mm MgSO 4. A single KM22 or TN30 colony was stab inoculated into SSM or SSM+MgSO 4 and incubated at 37 C for at least 24 hours. Motile organisms display outward migration from the point of inoculation. 190 Quantitative real-time PCR. 9
10 Mid-log-phase liquid cultures of KM22, TN23, TN30, and TN31 were diluted in SS broth containing streptomycin to obtain a starting optical density at 600 nm (OD 600 ) of 0.02, in triplicate, and grown at 37 C with shaking at 275 rpm until an OD 600 of 0.8 was reached. Bacteria were harvested and total RNA was extracted with Trizol (Invitrogen, Carlsbad, CA), treated with RNase-free DNase I (Invitrogen, Carlsbad, CA) and purified using RNeasy columns (Qiagen, Valencia, CA) according to the manufacturer s instructions. DNase-treated total RNA (1µg) from each biological replicate was reverse transcribed using 300ng of random oligonucleotide hexamers and SuperScript III RTase (Invitrogen, Carlsbad, CA) according to the manufacturer s protocol. The resulting cdna was diluted 1:1000, and 1µL of this dilution was used in qpcr reactions containing 300 nm primers and 2XSYBR Green PCR Master Mix (Applied Biosystems, Biosystems, Foster City, CA) using an Applied Biosystems 7300 real-time PCR detection system (Applied Biosystems, Foster City, CA). All primers were designed using Primer Express software (Applied Biosystems, Foster City, CA) and listed in Table 1. To confirm the lack of DNA contamination, reactions without reverse transcriptase were performed. Dissociation curve analysis was performed for verification of product homogeneity. Threshold fluorescence was established within the geometric phase of exponential amplification and the Ct value for each reaction was determined. The Ct value from all three biological replicates for each strain was compiled and the 16S RNA amplicon was used as an internal control for data normalization. Fold-change in transcript level was determined using the relative quantitative method (ΔΔC T ) (20). Results were Log 2 transformed and analyzed for significance using a twoway mixed model ANOVA with a post-hoc comparison of fold-change in transcript level of each isogenic mutant to the wild-type isolate using the Bonferroni method. P value less than was considered significant. 10
11 214 Cytotoxicity assays These assays were carried out as previously described (24, 40). Briefly, J774 macrophages were cultured in DMEM broth supplemented with 10% fetal bovine serum (FBS), 1% penicillinstreptomycin, 1% non-essential amino acids and 1% sodium pyruvate to 85% confluency at 37 C with 5% CO 2. Warmed RMPI lacking phenol-red, with 5% FBS, 1% L-glutamine, 1% nonessential amino acids and 1% sodium pyruvate was then used to replace the DMEM. Bacterial infections using the indicated bacterial strains prepared from cultures collected in mid-log phase and were carried out using a multiplicity of infection (MOI) of 10. Bacterial suspensions were centrifuged onto the macrophages at 250 x g for 5 minutes and cell cultures were incubated at 37 C with 5% CO 2 for four hours. The cell culture supernatants were collected and percent lactate dehydrogenase (LDH) release was analyzed using the Cytotox96 kit (Promega, Madison, WI) according to the manufacturer s instructions. Results were analyzed for significance using the Student s t test and a P value less than 0.05 was considered significant. Bacterial adhesion assays. L2 rat lung epithelial cells were grown to ~80% confluency in a 24 well plate using 50/50 DMEM/F12 medium supplemented with 10% FBS, and then inoculated with suspensions of the indicated bacterial strains prepared from cultures collected in mid-log phase and diluted using growth medium, resulting in a MOI of 100. Dilutions of bacterial inocula were plated on BG plates containing streptomycin to determine CFU counts and verify the MOI. Adhesion assays were carried out in triplicate and performed by removing growth medium from L2 cells and then adding either medium alone or 1 ml of bacterial inoculum. Plates were then centrifuged for 5 minutes at 250 x g and incubated at 37 C with 5% CO 2 for 40 minutes. Wells were then washed 11
12 four times with 1 ml of the growth medium to remove nonadherent bacteria. L2 cells were then trypsinized using 0.5 ml of 0.125% trypsin and incubated for 10 minutes at 37 C. The total volume of each well was then brought up to 1 ml with growth medium and cells were homogenized by pipetting. Dilutions were then plated on BG plates containing streptomycin to determine CFU counts, which were then used to calculate the proportion of adherent bacteria, expressed as a percentage of the original inoculum. Results were analyzed for significance using the Student s t test and a P value less than 0.05 was considered significant. Pathogenesis experiments in swine. B. bronchiseptica strains KM22, TN23 (Bvg + ), TN31 (Bvg i ) and TN30 (Bvg ) were cultured on BG agar supplemented with 10% sheep s blood at 37 C for 40 hours. Suspensions of these cultures were prepared in phosphate buffered saline (PBS) to contain approximately 2 x 10 9 CFU/ml, and a 1:2000 dilution of this suspension was made in PBS for inoculation of the pigs. An aliquot of inoculum for each strain was plated on BG agar plates, with and without 50 mm MgSO 4, to determine CFU/mL and to confirm the expected colony morphology and hemolytic phenotype. All colonies from all inocula displayed the expected colony morphology and hemolytic characteristics. A single isolation room in BL2 barn facilities was used to house each experimental group. No B. bronchiseptica was isolated from nasal swabs prior to the start of either experiment. For the first pathogenesis trial, 56 pigs were divided into 3 experimental groups of 16 pigs each and 1 control group of 8. Pigs were inoculated intranasally at 1 week of age with 1 ml (0.5 ml/nostril) of a bacterial suspension containing 10 6 CFU of KM22, TN23 (Bvg + ), TN30 (Bvg - ) or with 1 ml of sterile PBS, respectively. For the second pathogenesis trial, 48 pigs were divided into 2 experimental groups of 16 pigs each and 1 control group of 16. Pigs were inoculated intranasally at 1 week of age with 1 ml (0.5 ml/nostril) of a bacterial suspension 12
13 of strain KM22, TN31 (Bvg i ) or with 1 ml of sterile PBS, respectively. For both trials, bacterial colonization of the nasal cavity was quantified by nasal swabs 1, 3, 5, and by nasal washes at indicated days post-inoculation. Animals were euthanized and bacterial load in the trachea and the lungs was quantified at indicated days post-inoculation. At necropsy snouts were transected and removed at the level of the first premolar tooth. A 1 cm cross-section was cut from the caudal portion of the snout and used for scoring turbinate atrophy. The trachea was then severed just below the larynx and the trachea and lung were removed. All housing, husbandry and experiments performed with pigs were in accordance with the law and approved by the Institutional Animal Care and Use Committee. Determination of colonization. Nasal swabs were placed into tubes containing 500 µl PBS and vortexed. Nasal washes were performed by injecting 5 ml of PBS into the nasal cavity through one nostril and collecting the effluent into a beaker. Tracheal washes were performed by placing a segment of trachea approximately 8 cm in length in a 15 ml centrifuge tube with 5 ml of PBS and shaking vigorously. Lung lavage was performed by filling the lungs with 100 ml of sterile PBS, gently massaging and aspirating; approximately 50 ml of the PBS was recovered. Serial 10-fold dilutions were made from nasal swab fluids, nasal and tracheal washes and lung lavages and the number of CFU of B. bronchiseptica per ml was determined by plating 100 µl of the dilutions on duplicate selective blood agar plates containing 20 µg/ml penicillin, 10 µg/ml amphotericin B, 10 µg/ml streptomycin and 10 µg/ml spectinomycin. The limit of detection was 10 CFU/ml. B. bronchiseptica was identified on the basis of colony morphology. Colonization levels in the TN31 (Bvg i ) inoculated pigs was confirmed using colony lifts hybridized with a probe for the Bordetella adenylate cyclase toxin gene, cyaa. Colonies from plates representing each 13
14 respiratory tract site (nasal cavity, trachea, and lung) from each experimental group were randomly screened by PCR amplification of the bvgs gene and DNA sequence analysis to confirm the expected bvgs genotype. Statistical analyses of the nasal colonization data were performed using a mixed linear model (SAS 9.2 for Windows XP, SAS Institute Inc., Cary, NC, USA) for repeated measures and a spatial spherical covariance structure to best account for unequal study day intervals. Linear combinations of the least squares means estimates for log10 colony forming units were used in a priori comparisons to test for a significant (P<0.05) effect of the bacterial challenge strains. Comparisons were made between challenge groups for each isolate at each time-point using a 5% level of significance (P<0.05) to assess statistical differences. Endpoint data for tracheal and lung bacterial loads were analyzed using analysis of variance using a general linear model for unbalanced data that included treatment group and study day, with CFU as the dependent variable. A 5% level of significance (P<0.05) was used to assess statistical differences. Pathological evaluation of the lung. Sections of lung taken at necropsy for microscopic evaluation were fixed in 10% neutral buffered formalin for 24 hours and then placed in 70% ethanol. All sections were routinely processed and embedded in paraffin, sectioned and stained with hematoxylin and eosin. At necropsy, an estimate of gross lung involvement was assigned based on the percentage of each lung lobe affected and the percentage of total lung volume represented by each lobe. Percentage of total lung volume of each lobe was estimated as 10% for the left cranial, 10% for the left middle, 25% for the left caudal, 10% for the right cranial, 10% for the right middle, 25% for the right caudal, and 10% for the intermediate lung lobes. 14
15 304 Turbinate atrophy scores Each of the four scrolls of the ventral turbinate in snout cross-sections was assigned an atrophy score that ranged from 0-4: 0 = normal, 1 = more than half of turbinate remaining, 2 = half or less of turbinate remaining, 3 = turbinate is straightened with only a small portion left, and 4 = total atrophy. The atrophic rhinitis score is the sum of the four turbinate atrophy scores and ranges from Transmission experiments in swine. B. bronchiseptica strains KM22 and TN23 (Bvg + ) were cultured on BG agar supplemented with 10% sheep s blood at 37 C for 40 hours. Suspensions of these cultures were prepared in phosphate buffered saline (PBS) to contain approximately 2 x 10 9 colony forming units (CFU)/ml, and a 1:2000 dilution of this suspension was made in PBS for inoculation of the pigs. An aliquot of inoculum for each strain was plated on BG agar plates, with and without 50 mm MgSO 4, to determine CFU/mL and to confirm the expected colony morphology and hemolytic phenotype. All colonies from both inocula displayed the expected characteristics. A single isolation room in BL2 barn facilities was used to house each experimental group. For the direct transmission study, 2 experimental groups of 16 one-week-old piglets and 1 control group of 8 were intranasally inoculated with 10 6 CFU of KM22 or TN23 (Bvg + ) or 1 ml sterile PBS. At 3 days post-inoculation, two naïve pigs were added to each room to serve as direct contacts. Bacterial colonization of the nasal cavity was assessed by nasal washes 2, 8, 18, 39, 46, 69, 76, and 84 days post-contact. Four primary challenged pigs were removed on days 10, 17, 34, and 60 days post-contact for necropsy, leaving only direct contacts in the room beyond day 60 postcontact. On day 84 post-contact necropsies were performed and pigs were euthanized with an 15
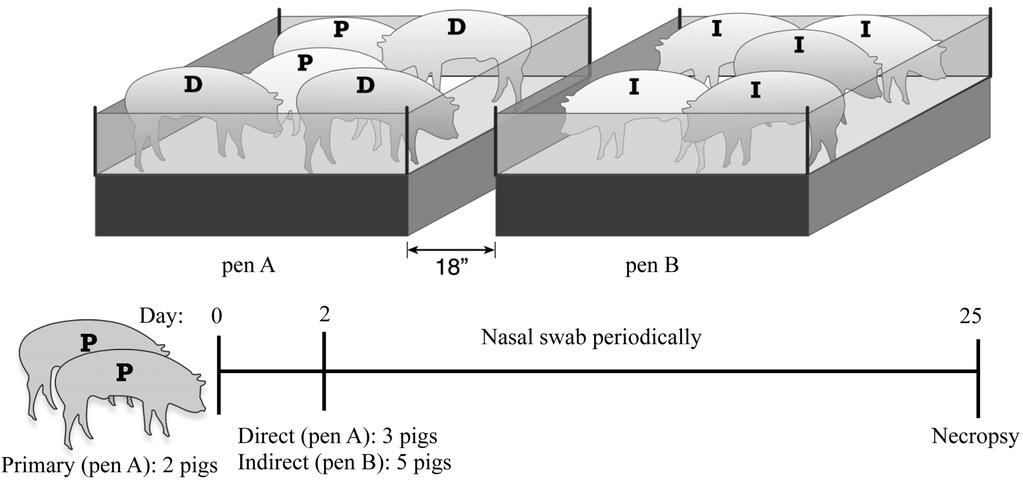
16 overdose of barbiturate. For the direct and indirect transmission study, three isolation rooms in BL2 barn facilities were used to house each experimental group. Two isolation rooms were identically set up with two pens placed 18 inches apart. Two pigs were intranasally inoculated with 10 6 CFU of KM22 and placed in a pen together in one isolation room, and two pigs were intranasally inoculated with 10 6 CFU of TN23 (Bvg + ) and placed in a pen together in a second isolation room. These pigs served as the primary-challenged pigs in the study. A control group of 8 pigs were intranasally inoculated with 1 ml sterile PBS and housed together in a third isolation room with no pens. Two days post-inoculation of primary-challenged pigs, 3 naïve pigs, serving as a direct-contacts, were placed in the pen containing the primary-challenged pigs and 5 naïve pigs, serving as indirect-contacts, were placed in the second pen. Nasal colonization of all pigs was monitored by periodical nasal swabbing 4, 7, 12, 18, and 25 days post-contact. Nasal swabs were obtained from indirect-contacts first, followed by direct-contacts, and then nasal swabs were obtained from primary-challenged pigs. Necropsies were performed on day 25 post-contact and all pigs were euthanized with an overdose of barbiturate. For all transmission experiments, the trachea was then severed just below the larynx and the trachea and lung were removed for determination of colonization. All housing, husbandry and experiments performed with pigs were in accordance with the law and approved by the Institutional Animal Care and Use Committee. 16
17 345 Results Phenotype and gene expression pattern of each KM22 isogenic mutant (Bvg, Bvg i, and Bvg + ). To investigate the role of phenotypic modulation by B. bronchiseptica during infection of swine, we constructed a series of isogenic phase-locked mutants in the virulent swine strain KM22. Since regulation of virulence gene expression by the bvg locus is highly conserved among Bordetella strains, we hypothesized that isogenic mutants (Bvg, Bvg i, and Bvg + ) of KM22 would have similar phenotypes as those reported for each corresponding isogenic mutant in the well-characterized laboratory strain, RB50. Gene expression in TN23 (Bvg + ), TN31 (Bvg i ), and TN30 (Bvg ) was analyzed by quantitative real-time PCR and compared to the wild-type KM22 strain. The phase-locked mutants and the wild-type KM22 were grown under non-modulating conditions at 37 C. 19 genes that have been shown in RB50 to be differentially expressed in response to phase modulation were chosen for analysis. These included genes for regulatory proteins (bvga, bvgs, bvgr), adhesins (prn, fima, fim2, fim3, fimn, fimx), toxins (cyaa, dnt), the type III secretion system (bopb, bopd, bopn), motility (flhc, chez, flgb) and the intermediatephase expressed genes bipa and bcfa (15, 27, 34). As shown in Figure 1A, the three isogenicmutant strains have unique patterns of gene expression. The expression of prn, fim3, fimn, fimx, and dnt were significantly increased in the Bvg + phase-locked mutant TN23, while the expression of flhc, chez, flgb, bipa, fima and fim2 were significantly decreased relative to the wild-type KM22 (Fig 1A). The expression levels of the other eight genes analyzed did not differ significantly from the wild-type KM22 (Fig 1A). In contrast, the expression of all the virulenceassociated genes analyzed was significantly decreased in the Bvg phase-locked mutant, TN30, relative to wild-type KM22 (Fig 1A). The expression of bcfa, prn, fim3, bipa, fima, and fimn 17
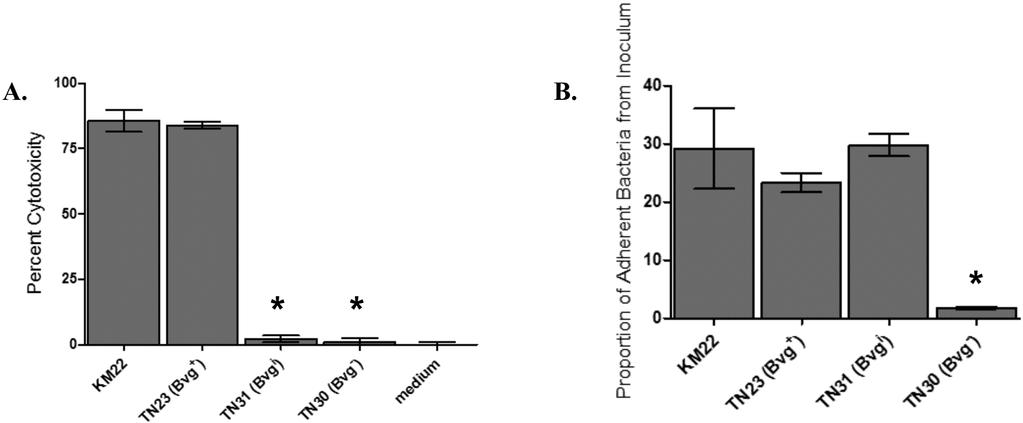
18 were significantly increased in the Bvg i phase-locked mutant TN31, while the expression of flhc, flgb, bopn, bopd, bopb, and fim2 were significantly decreased relative to the wild-type KM22 (Fig 1A). In summary, expression of all 19 genes analyzed in each isogenic mutant of KM22 was found to be similar to that reported for the corresponding isogenic mutants in the wellcharacterized laboratory strain, RB50. Perhaps the best characterized phenotype of the Bvg phase is motility and the expression of flagellar synthesis genes, which are tightly regulated through a complex hierarchy requiring the presence of the FrlAB regulatory locus, and the production of the flagellin monomer, FlaA (1, 2). Relative to B. bronchiseptica RB50, the expression of the flagella structural gene flaa has been reported to increase in RB54, the Bvg phase-locked derivative of RB50 (16, 33). To test whether TN30 is motile under both modulating and non-modulating conditions, a single colony of either wild-type KM22 or TN30 were stab inoculated into softagar plates containing SSM or SSM supplemented with 50 mm MgSO 4. As shown in Figure 1B, TN30 is motile in both non-modulating, SSM, and modulating, SSM supplemented with 50 mm MgSO 4, conditions, while KM22 is only motile in modulating conditions, SSM supplemented with 50 mm MgSO 4. These data demonstrate that the Bvg phase-locked mutant, TN30, is expressing the Bvg phase phenotype motility during both modulating and non-modulating conditions. A well-characterized function of the Bordetella Type III secretion system (TTSS) is killing of mammalian cells (30, 39, 40). Since the BvgAS system regulates the TTSS in B. bronchiseptica RB50, we hypothesized that the BvgAS system would also regulate TTSS in KM22. Therefore we assessed the ability of KM22, and the isogenic phase-locked mutants to mediate cytotoxicity in J774 murine macrophages in vitro. Both KM22 and the Bvg + -locked 18
19 mutant, TN23, displayed a high level of cytotoxicity (Fig. 2A). In contrast, the cytotoxic activity of the Bvg i -locked strain, TN31, and Bvg phase-locked strain, TN30 was statistically lower than KM22 and similar to medium alone (Fig 2A). These data correlate with the gene expression data and suggest that both KM22 and TN23 produce a functional TTSS, while the decreased expression of some TTSS genes in strains TN31 and TN30 may have resulted in the decreased cytotoxicity, a TTSS-dependent function. The ability of B. bronchiseptica to adhere to epithelial surfaces is required for the initiation and establishment of infection. The BvgAS system regulates a number of genes whose products have been postulated or, in some cases, directly shown to function as adhesins. To determine if the BvgAS system is involved in regulating cellular adherence by KM22, the ability of the phase-locked mutants to adhere to L2 rat lung epithelial cells in vitro was compared to that of the wild-type strain. KM22, TN23 (Bvg + ) and TN31 (Bvg i ) had a similar level of adherence to L2 cells, whereas TN30 (Bvg ) was unable to adhere (Fig. 2B). These results indicate that Bvg B. bronchiseptica does not express factors necessary for adhesion to L2 cells, supporting the hypothesis that Bvg -phase gene products are not involved in colonization and establishment of infection. Full activation of the BvgAS regulatory system is required for colonization and maximal disease severity. To investigate the Bvg phase-dependent contribution to pathogenesis of B. bronchiseptica in swine we assessed, in two separate trials, the ability of TN23 (Bvg + ), TN31 (Bvg i ) and TN30 (Bvg ) to colonize and cause disease as compared to the wild-type isolate. In the first trial, groups of one-week-old piglets were intranasally inoculated with KM22, TN23 (Bvg + ), TN30 (Bvg ), or 1 ml sterile PBS. Clinical signs were only noted in piglets from groups 19
20 inoculated with KM22 or TN23 (Bvg + ) and consisted of sneezing and coughing. No clinical signs were observed in piglets from groups inoculated with TN30 (Bvg ) or PBS Colonization of the nasal cavity was evaluated on days 1, 3, 5, 7, and weekly thereafter until day 57 post-infection (Fig. 3A). No B. bronchiseptica CFU were recovered from any site in the respiratory tract of piglets inoculated with TN30 (Bvg ) or PBS. The Bvg + phase-locked mutant TN23 was recovered from the nasal cavity in numbers similar to those for the wild-type KM22 at all days examined (Fig. 3A). Colonization of sites in the lower respiratory tract was also assessed at 7, 14, 31, and 57 days post-infection. Similar to nasal colonization, there was no difference in the number of CFUs recovered from TN23 (Bvg + ) as compared to KM22 in both the trachea and the lung (Fig. 3B and C). The degree of atrophic rhinitis was evaluated by determining the mean turbinate atrophy score from piglets inoculated with KM22, TN23 (Bvg + ), TN30 (Bvg ), or PBS on days 7, 14, 31, and 57 post-infection. No significant turbinate atrophy was seen in the pigs inoculated with either PBS or TN30 (Bvg ) on any of the days examined (Table 2). Only piglets infected with either KM22 or TN23 (Bvg + ) had any evidence of significant nasal turbinate atrophy, reflective of an atrophic rhinitis score greater than two (Table 2). Pigs infected with TN23 (Bvg + ) showed a similar degree of turbinate atrophy as pigs infected with the wild-type KM22 on all days except day 31, where an increased degree of turbinate atrophy was observed in KM22 infected pigs (Table 2) Lung pathology was assessed on days 7, 14, 31 and 57 days post-infection by determining the mean percentage of the lung affected by pneumonia. Overall, no pneumonia was observed in TN30 (Bvg ) or PBS infected piglets, where as a similar degree of pneumonia 20
21 was observed in TN23 (Bvg + ) and KM22 infected pigs on days 7, 14, and 31 (Table 2). On day 57 post-infection, the mean percentage of lung affected by pneumonia was slightly higher in TN23 (Bvg + ) challenged pigs, with 5.625% of lung affected compared to 2.75% of lung affected in KM22 infected pigs (Table 2). Additionally, only one out the four pigs infected with KM22 exhibited pneumonia whereas three out the four pigs infected with TN23 (Bvg + ) exhibited pneumonia. The pneumonia exhibited in TN23 (Bvg + ) and KM22 infected pigs was similar and consisted of areas of red, seen 7 and 14 days post-infection, to tan, seen 31 and 57 days postinfection, consolidation with well-demarcated borders and a cranial ventral distribution. Microscopically, the lesions in both TN23 (Bvg + ) and KM22 challenged pigs were typical of B. bronchiseptica pneumonia, characterized by suppurative bronchopneumonia that is gradually replaced by fibroplasia. In a second trial, we compared the ability of TN31 (Bvg i ) and the wild-type KM22 isolate to colonize and cause disease in swine following intranasal inoculation of groups of one-weekold piglets with KM22, TN31 (Bvg i ), or PBS. No clinical signs were observed in piglets inoculated with TN31 (Bvg i ) or in piglets receiving PBS. Clinical signs, consisting of sneezing and coughing, were noted only in piglets inoculated with KM22. Nasal cavity colonization was assessed at days 1, 3, 5, 7 and then weekly through day 56 post-infection. The Bvg i phase-locked mutant TN31 was recovered at levels significantly lower than those of wild-type KM22 on all days (Fig 4A). Throughout the course of the experiment, the number of CFU recovered from pigs infected with TN31 (Bvg i ) was approximately two logs less than from pigs infected with KM22. Additionally, the peak in the number of CFU recovered from the nasal cavity of TN31 (Bvg i ) infected pigs was delayed compared to KM22-infected pigs, with CFU levels barely rising above detectable limits through day 14 and reaching a peak at 21
22 day 28, and by day 56 CFU levels fell to barely detectable levels (Fig 4A). In contrast, the number of CFU recovered from the nasal cavity of KM22 infected pigs were significantly higher on day 1, reached a peak on day 21, and remained high throughout the course of the study (Fig 4A) Colonization of the trachea and lung was evaluated on days 7, 14, 28, and 56 postinfection (Fig. 4B and C). In general, significantly lower CFU levels were recovered from the trachea and lungs of TN31 (Bvg i )-inoculated pigs than from the trachea and lungs of pigs infected with KM22 (Fig 4B and C). Similar to nasal colonization, peak CFU recovered from the trachea and lungs was delayed until day 28 in pigs infected with TN31 compared to day 14 in KM22-inoculated pigs. Additionally, KM22 was able to persist in significantly high numbers through day 56 in both the trachea and lung, whereas the number of TN31 (Bvg i ) decreased more dramatically, approaching the lower limit of detection by day 56 (Fig 4B and C). No significant turbinate atrophy was seen in the pigs inoculated with either PBS or TN31 (Bvg i ) at any of the days examined (Table 3). Only piglets infected with KM22 had any evidence of significant nasal turbinate atrophy, reflective of an atrophic rhinitis score greater than two (Table 3). Pigs inoculated with TN31 (Bvg i ) displayed little to no pneumonia (Table 3), with the range of lung percentage affected as well as the total number of pigs displaying evidence of pneumonia similar to that of the PBS-inoculated control group. Despite isolation of significantly lower numbers of B. bronchiseptica from the lung, two TN31 (Bvg i )-infected pigs displayed mild lesions on days 7 and 56. Microscopically, the lesions on day 7 were consistent with Bordetella infection and included small areas of consolidated interstitial pneumonia with septal infiltrates consisting mainly of macrophages and neutrophils. Lesions detected on day 56 were suppurative bronchopneumonia, also consistent with Bordetella infection. The colonization 22
23 and pathology data from each of the two trials together suggest that the full activation of the BvgAS regulatory system is required for maximal colonization and disease severity in swine Phenotypic modulation of the Bvg+ phase is not required for direct or indirect transmission of B. bronchiseptica in swine. An attractive and frequently proposed hypothesis is that a BvgAS system capable of modulation between the Bvg + and Bvg phases is required for transmission (12, 14, 33). To experimentally address this hypothesis, we undertook two separate studies designed to test the ability of the Bvg + phase-locked mutant, TN23, to be transmitted, either directly or indirectly, from infected pigs to naïve pigs. To assess direct transmission, groups of 16 one-week-old piglets were intranasally inoculated with KM22, TN23 (Bvg + ) or 1 ml sterile PBS. At 3 days post-inoculation two naïve pigs were added to each room to serve as direct contacts. Nasal washes were performed on the direct contact pigs to test for colonization by each respective B. bronchiseptica strain. As shown in Table 4, the direct contact pigs in groups inoculated with KM22 or TN23 (Bvg + ) were rapidly colonized by each respective strain and remained colonized for the duration of the trial. No B. bronchiseptica was recovered from any site in the respiratory tract of piglets in the group inoculated with PBS. Colonization of the respiratory tract was evaluated in the direct contact pigs from each group 84 days post-contact with primary infected pigs. Both direct contact pigs were positive for nasal colonization from the group inoculated with KM22 and the group inoculated with TN23 (Bvg + ) (Table 4 and Table 5). B. bronchiseptica was recovered from the trachea in one direct contact pig from the group inoculated with KM22 and both direct contact pigs from the group inoculated with TN23 (Bvg + ) (Table 5). B. bronchiseptica was recovered 23
24 from the lung of one direct contact pig from the group inoculated with KM22, but neither of the direct contact pigs from the group inoculated with TN23 (Bvg + ) (Table 5). Together the data demonstrate that a Bvg + phase-locked mutant can transmit directly from pig-to-pig when animals are comingled and suggest that phenotypic modulation of the Bvg + phase is not required for direct transmission in swine. A second trial was designed to test whether the Bvg+ phase is sufficient for both direct and indirect transmission in swine. In this experiment, two pigs were intranasally inoculated with KM22 and placed in a pen within an isolator room (Fig 5). These two pigs served as primary-challenged pigs. After two days, three naïve pigs, serving as direct-contacts, were added to the same pen, and five naïve pigs were placed in a second pen placed at least 18 inches away from the first pen (Fig 5). These five pigs served as indirect-contact pigs. The exact experimental procedure was repeated in a second isolation room except that the two pigs serving as primary-challenged pigs were intranasally inoculated with TN23 (Bvg + ). In a third isolator room with no pens, pigs were intranasally inoculated with sterile PBS and allowed to comingle. Nasal colonization of all pigs was monitored by nasal swabbing. B. bronchiseptica was not recovered from any site in piglets inoculated with PBS. Similar to the first trial, direct transmission occurred rapidly for both KM22 and TN23 (Bvg + ) with the majority of directcontact pigs from each group becoming colonized by day 4 post-contact (Table 6). Transmission by indirect contact took longer; however, for both strains, an indirect-contact pig demonstrated nasal colonization twelve days after initial contact (Table 6). By day 25 post-contact, B. bronchiseptica was recovered from the nasal cavity of all pigs (primary-challenged, directcontact, and indirect-contact pigs) from both the KM22 and the TN23 (Bvg + ) groups. Colonization of the trachea and lungs was determined at necropsy 25 days post-contact. The B. 24
25 bronchiseptica strain given to the primary-challenged pigs from each group was isolated from the trachea and lungs in all pigs (primary, direct contact and indirect contact) from both the KM22 (wild-type) and TN23 (Bvg + ) groups (Table 7). These results show that both the wildtype and the Bvg + phase-locked mutant were equally capable of transmission in swine by both direct and indirect contact. Additionally, these data demonstrate that phenotypic modulation of Bvg + B. bronchiseptica is not required for direct or indirect transmission in swine. Downloaded from on August 31, 2018 by guest 25
26 534 Discussion Despite widespread use of B. bronchiseptica vaccines by swine producers throughout the world, Bordetella-associated respiratory diseases remains a significant problem for the industry (18, 36). The development of vaccines with improved efficacy is hindered by the absence of definitive data related to virulence mechanisms of B. bronchiseptica in swine. Evaluation of B. bronchiseptica-specific virulence factors involved in pathogenesis and transmission in swine provides data directly relevant and necessary for designing improved vaccines and therapeutic interventions. We hypothesized that the ability of B. bronchiseptica to undergo phenotypic modulation is required at some point during the infectious cycle in swine. To investigate the Bvg phase-dependent contribution to pathogenesis of B. bronchiseptica in swine, we constructed a series of isogenic mutants in a virulent B. bronchiseptica swine isolate and first evaluated the in vitro associated phenotypes of each KM22 isogenic mutant. We found the in vitro phenotypes associated with each KM22 isogenic mutant (Bvg, Bvg i, and Bvg + ) to be similar to those reported for each corresponding RB50 isogenic mutant. To investigate the Bvg phase-dependent contribution to pathogenesis of B. bronchiseptica in swine we compared TN23 (Bvg + ), TN31 (Bvg i ) and TN30 (Bvg ) to the wildtype swine isolate for their ability to colonize and cause disease. The Bvg phase-locked mutant, TN30, was never recovered from any respiratory tract site at any time point examined. Except for the degree of pneumonia observed late in the infection, the Bvg + phase-locked mutant, TN23, was indistinguishable from wild-type and caused similar disease severity. These results parallel previous studies involving phase-locked and ectopic expression mutants, which demonstrated that the Bvg + phase promotes respiratory tract colonization by B. pertussis and B. bronchiseptica (1, 13, 14, 23, 26), while the Bvg phase of B. bronchiseptica does not promote colonization or 26
27 disease in a host (13, 14). The increased pneumonia exhibited by the TN23 (Bvg+) infected pigs late in the infection suggests that the ability to modulate pathology during acute infection may be biologically beneficial for B. bronchiseptica. For example, while not evaluated here, the ability to modulate pathology could increase the possibility that a host remains more active and mobile, thereby increasing transmission dynamics within host populations. The Bvg phase of B. bronchiseptica has been demonstrated to promote bacterial survival under conditions of nutrient deprivation (13, 14). Therefore, while not tested here, the Bvg phase of B. bronchiseptica may promote increased survival in containment rooms, pens, and water sources in swine production facilities, thereby increasing the incidence of indirect transmission between animals. The inability to monitor such events illustrates a limitation of studying transmission in a confined experimental setting. As we followed colonization levels throughout the infection, the Bvg i phase-locked mutant, TN31, was recovered at significantly lower levels compared to wild-type KM22 at all respiratory tract sites and time points examined. This is in direct contrast to the in vitro data where we found no difference in the capacity of TN31 and KM22 to adhere to L2 cells. While it may be possible to identify a difference in the adherence capability between KM22 and TN31 using different cell types, we found significantly increased transcription levels for adhesion associated genes, bcfa, prn, fim3, bipa, fima, and fimn, in TN31 compared to KM22, and similar transcription levels for fimx in both TN31 and KM22. The decreased bacterial burdens throughout the respiratory tract of TN31-infected pigs observed late in the infection demonstrate the ability of swine to clear Bvg i -phase B. bronchiseptica more rapidly than wild-type B. bronchiseptica. In addition to the decreased in vivo adherence ability, this may be due to the lack of maximal expression and production of characterized immune suppressors and protective 27
28 proteins. One example includes the TTSS, which is capable of modulating the host immune response to aid in survival and persistence in vivo (30, 39, 40). The gene expression and cytotoxicity data correlate with the in vivo colonization data and suggest that TN31 does not produce a functional TTSS; however we cannot exclude the potential contribution of other factors as well. The decreased colonization of TN31-infected pigs compared to wild-type KM22 at all respiratory tract sites and time points examined demonstrates that in order to adhere, initiate infection, and persist to similar levels as wild-type in a natural host, Bvg + -phase factors are required. Further, pigs infected with TN31 displayed limited to no disease and few turbinate and lung lesions. Only pigs infected with KM22 or TN23 (Bvg+) displayed coughing and sneezing, which are mechanisms of transmission. These data suggest that a critical threshold of colonization is required to cause clinical signs of disease, such as sneezing and coughing, as well as degenerative pathology, such as turbinate atrophy, and are in agreement with previous findings (14). It is difficult to discern if pathological differences in the TN31-innoculated pigs were due to decreased colonization levels, lack of virulence factor expression or both. Given that the number of CFU recovered from the nasal cavity of TN31-infected pigs barely rose above detectable limits through the first two weeks of infection, we were unable to test the ability of this strain to transmit to naïve pigs. To test whether bacteria in the Bvg + phase are readily transmitted, we used the Bvg + phase-locked mutant, TN23, in direct and indirect transmission studies. Data from the first trial showed that a Bvg + phase-locked mutant can transmit directly from pig-to-pig when animals are comingled and demonstrates that phenotypic modulation of the Bvg+ phase is not required for direct transmission in swine. The experimental design of the second trial is set up to specifically address whether or not a Bvg+ phase-locked strain can transmit both directly and indirectly to 28
29 naïve pigs. Data from the second trial showed that a Bvg + phase-locked mutant can transmit directly to naïve pigs and indirectly to naïve pigs physically separated by pens. It is important to note that once B. bronchiseptica was recovered from one naïve pig from the pen containing indirect-contact pigs, it was no longer possible to discern the route of transmission, direct or indirect, that led to the B. bronchiseptica colonization of the other four naïve pigs housed in the same pen. In any case, the isolation of TN23 from the nasal cavity of an indirect-contact pig in a separate pen demonstrates that a Bvg+ phase-locked mutant is capable of transmission by an indirect route. We observed a difference in the timing of when B. bronchiseptica was recovered from the nasal cavity of direct-contact pigs from these studies. By day 2 post-contact B. bronchiseptica was recovered from direct-contact pigs in the first study, whereas in the second study B. bronchiseptica was not recovered until 7 days post-contact. The increase in the number of days for direct transmission to occur likely reflects the ratio of primary-infected pigs to naïve pigs, where in the first trial 16 primary-infected pigs to 3 naïve pigs were used and in the second trial there were 2 primary-infected pigs to 3 naïve pigs. These studies provide experimental data supporting conventional practices of decreasing the number of animals housed or comingled together in efforts to delay or decrease transmission. It was not unexpected to find no significant differences in colonization between KM22- inoculated and TN23 (Bvg + )-inoculated pigs. However, given that the hypothesis that some degree of phenotypic modulation is required for transmission of B. bronchiseptica between animals is frequently proposed (12, 14, 33), it was surprising to find the Bvg + phase-locked mutant to transmit to naïve pigs by both direct and indirect contact with equal efficiency as the wild-type isolate. Our results demonstrate that in a natural host, phenotypic modulation of the 29
30 fully active Bvg + phase is not required for respiratory infection and host-to-host transmission of B. bronchiseptica Downloaded from on August 31, 2018 by guest 30
31 631 Acknowledgements The authors would like to thank Kim Driftmier, William Boatwright, and Gwen Nordholm for excellent technical support, and Brian Pottebaum, Jason Huegel, Jason Crabtree and Dalene Whitney for animal care assistance. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. USDA is an equal opportunity provider and employer. Downloaded from on August 31, 2018 by guest 31
32 639 References Akerley, B. J., P. A. Cotter, and J. F. Miller Ectopic expression of the flagellar regulon alters development of the Bordetella-host interaction. Cell 80: Akerley, B. J., D. M. Monack, S. Falkow, and J. F. Miller The bvgas locus negatively controls motility and synthesis of flagella in Bordetella bronchiseptica. J. Bacteriol. 174: Brockmeier, S. L Prior infection with Bordetella bronchiseptica increases nasal colonization by Haemophilus parasuis in swine. Vet. Microbiol. 99: Brockmeier, S. L., and K. M. Lager Experimental airborne transmission of porcine reproductive and respiratory syndrome virus and Bordetella bronchiseptica. Vet. Microbiol. 89: Brockmeier, S. L., C. L. Loving, T. L. Nicholson, and M. V. Palmer Coinfection of pigs with porcine respiratory coronavirus and Bordetella bronchiseptica. Vet. Microbiol. 128: Brockmeier, S. L., M. V. Palmer, and S. R. Bolin Effects of intranasal inoculation of porcine reproductive and respiratory syndrome virus, Bordetella bronchiseptica, or a combination of both organisms in pigs. Am. J. Vet. Res. 61: Brockmeier, S. L., M. V. Palmer, S. R. Bolin, and R. B. Rimler Effects of intranasal inoculation with Bordetella bronchiseptica, porcine reproductive and respiratory syndrome virus, or a combination of both organisms on subsequent infection with Pasteurella multocida in pigs. Am. J. Vet. Res. 62: Brockmeier, S. L., and K. B. Register Expression of the dermonecrotic toxin by Bordetella bronchiseptica is not necessary for predisposing to infection with toxigenic Pasteurella multocida. Vet. Microbiol. 125: Brockmeier, S. L., K. B. Register, T. Magyar, A. J. Lax, G. D. Pullinger, and R. A. Kunkle Role of the dermonecrotic toxin of Bordetella bronchiseptica in the pathogenesis of respiratory disease in swine. Infect. Immun. 70: Buboltz, A. M., T. L. Nicholson, L. S. Weyrich, and E. T. Harvill Role of the type III secretion system in a hypervirulent lineage of Bordetella bronchiseptica. Infect. Immun. 77: Chanter, N., T. Magyar, and J. M. Rutter Interactions between Bordetella bronchiseptica and toxigenic Pasteurella multocida in atrophic rhinitis of pigs. Res. Vet. Sci. 47.: Cotter, P. A., and A. M. Jones Phosphorelay control of virulence gene expression in Bordetella. Trends Microbiol. 11: Cotter, P. A., and J. F. Miller BvgAS-mediated signal transduction: analysis of phase-locked regulatory mutants of Bordetella bronchiseptica in a rabbit model. Infect. Immun. 62: Cotter, P. A., and J. F. Miller A mutation in the Bordetella bronchiseptica bvgs gene results in reduced virulence and increased resistance to starvation, and identifies a new class of Bvg-regulated antigens. Mol. Microbiol. 24: Cummings, C. A., H. J. Bootsma, D. A. Relman, and J. F. Miller Species- and strain-specific control of a complex, flexible regulon by Bordetella BvgAS. J. Bacteriol. 188:
33 Deora, R., H. J. Bootsma, J. F. Miller, and P. A. Cotter Diversity in the Bordetella virulence regulon: transcriptional control of a Bvg-intermediate phase gene. Mol. Microbiol. 40: Donnenberg, M. S., and J. B. Kaper Construction of an eae deletion mutant of enteropathogenic Escherichia coli by using a positive-selection suicide vector. Infect. Immun. 59: Guerrero, R. J Respiratory disease: an important global problem in the swine industry, p. 98. Proc. 11th Int. Pig Vet. Soc., Lausanne, Switzerland. 19. Inatsuka, C. S., Q. Xu, I. Vujkovic-Cvijin, S. Wong, S. Stibitz, J. F. Miller, and P. A. Cotter Pertactin is required for Bordetella species to resist neutrophil-mediated clearance. Infect. Immun. 78: Livak, K. J., and T. D. Schmittgen Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔ C T Method. Methods 25: Loving, C. L., S. L. Brockmeier, A. L. Vincent, M. V. Palmer, R. E. Sacco, and T. L. Nicholson Influenza virus coinfection with Bordetella bronchiseptica enhances bacterial colonization and host responses exacerbating pulmonary lesions. Microb. Pathog. 49: Magyar, T., and A. J. Lax Atrophic Rhinitis, p In K. A. Brogden and J. Guthmiller (ed.), Polymicrobial Diseases. ASM Press, Washington, D.C. 23. Martinez de Tejada, G., P. A. Cotter, U. Heininger, A. Camilli, B. J. Akerley, J. J. Mekalanos, and J. F. Miller Neither the Bvg- phase nor the vrg6 locus of Bordetella pertussis is required for respiratory infection in mice. Infect. Immun. 66: Mattoo, S., M. H. Yuk, L. L. Huang, and J. F. Miller Regulation of type III secretion in Bordetella. Mol. Microbiol. 52: McMillan, D. J., M. Shojaei, G. S. Chhatwal, C. A. Guzman, and M. J. Walker Molecular analysis of the bvg-repressed urease of Bordetella bronchiseptica. Microb. Pathog. 21: Merkel, T. J., S. Stibitz, J. M. Keith, M. Leef, and R. Shahin Contribution of regulation by the bvg locus to respiratory infection of mice by Bordetella pertussis. Infect. Immun. 66: Nicholson, T. L Construction and validation of a first-generation Bordetella bronchiseptica long-oligonucleotide microarray by transcriptional profiling the Bvg regulon. BMC Genomics 8: Nicholson, T. L., S. L. Brockmeier, and C. L. Loving Contribution of Bordetella bronchiseptica filamentous hemagglutinin and pertactin to respiratory disease in swine. Infect. Immun. 77: Palzer, A., M. Ritzmann, G. Wolf, and K. Heinritzi Associations between pathogens in healthy pigs and pigs with pneumonia. Vet. Rec. 162: Pilione, M. R., and E. T. Harvill The Bordetella bronchiseptica type III secretion system inhibits gamma interferon production that is required for efficient antibodymediated bacterial clearance. Infect. Immun. 74: Sekiya, K., M. Ohishi, T. Ogino, K. Tamano, C. Sasakawa, and A. Abe Supermolecular structure of the enteropathogenic Escherichia coli type III secretion system and its direct interaction with the EspA-sheath-like structure. Proc. Natl. Acad. Sci. USA 98:
34 Simon, R., U. Priefer, and A. Puhler A broad host range mobilization system for in vivo genetic engineering: transposon mutagenesis in Gram negative bacteria. Bio/Technology 1: Stockbauer, K. E., B. Fuchslocher, J. F. Miller, and P. A. Cotter Identification and characterization of BipA, a Bordetella Bvg-intermediate phase protein. Mol. Microbiol. 39: Sukumar, N., M. Mishra, G. P. Sloan, T. Ogi, and R. Deora Differential Bvg phase-dependent regulation and combinatorial role in pathogenesis of two Bordetella paralogs, BipA and BcfA. J Bacteriol. 189: Trahan, C. J., E. H. Stephenson, J. W. Ezzell, and W. C. Mitchell Airborneinduced experimental Bordetella bronchiseptica pneumonia in strain 13 guineapigs. Lab Anim. 21: U. S. Department of Agriculture Swine III. Reference of swine health and health management in the United States. USDA/APHIS/VS CEAH N USDA, Fort Collins, CO. 37. Vecht, U., J. P. Arends, E. J. van der Molen, and L. A. van Leengoed Differences in virulence between two strains of Streptococcus suis type II after experimentally induced infection of newborn germ-free pigs. Am. J. Vet. Res. 50: Vecht, U., H. J. Wisselink, J. E. van Dijk, and H. E. Smith Virulence of Streptococcus suis type 2 strains in newborn germfree pigs depends on phenotype. Infect. Immun. 60: Yuk, M. H., E. T. Harvill, P. A. Cotter, and J. F. Miller Modulation of host immune responses, induction of apoptosis and inhibition of NF-kappaB activation by the Bordetella type III secretion system. Mol. Microbiol. 35: Yuk, M. H., E. T. Harvill, and J. F. Miller The BvgAS virulence control system regulates type III secretion in Bordetella bronchiseptica. Mol. Microbiol. 28:
35 757 Figure Legends FIG. 1A. Gene expression changes in B. bronchiseptica TN23 (Bvg+), TN31 (Bvg i ), and TN30 (Bvg ) evaluated by qrt-pcr. The X-axis indicates the genes analyzed. The Y-axis indicates the Log 2 fold-change in gene expression of each strain relative to KM22 using the relative quantitative method (ΔΔC T ). Values for bvgs in TN30 (Bvg ) were not attainable since the region targeted by the primers used in the assay is missing in this strain. Error bars represent +/- the standard error. An asterisk indicates that the P value is < FIG. 1B. Soft-agar motility of B. bronchiseptica KM22 and TN30 (Bvg ). A single colony of either KM22 or TN30, as indicated on left, was stab inoculated into SSM or SSM supplemented with 50 mm MgSO 4, as indicated above. Motile organisms display outward migration from the point of inoculation. FIG. 2A. TTSS-mediated cytotoxicity of B. bronchiseptica KM22, TN23 (Bvg+), TN31 (Bvg i ), and TN30 (Bvg ). Cytotoxicity as measured by percent LDH release in J774 macrophages following 4 hours of exposure to medium alone or to the indicated strain, using an MOI of 10. The error bars represent +/- the standard deviation. An asterisk indicates that the P value is < FIG. 2B. Adherence of B. bronchiseptica KM22, TN23 (Bvg+), TN31 (Bvg i ), and TN30 (Bvg ). Adherence in vitro to L2 cells is expressed as the proportion of bacteria in the original inoculum found to be adherent after centrifugation and a 40 min incubation period, using an MOI of 100. Adherence of bacteria to L2 cells is expressed as the proportion of adherent bacteria to the original inoculum. The error bars represent +/- the standard deviation. An asterisk indicates that the P value is <
36 FIG. 3. Colonization of the swine respiratory tract by wild-type B. bronchiseptica strain KM22 or TN23, a KM22 Bvg+ isogenic mutant. Groups of 16 pigs were inoculated intranasally with KM22 (open triangles) or TN23, a KM22 Bvg+ isogenic mutant (closed circles). Bacterial load in the nasal cavity (A) was quantified 1, 3, 5, 7, 14, 21, 31, 42, 49, and 57 days post-inoculation. Bacterial load in the trachea (B) and the lungs (C) was quantified 7, 14, 31, and 57 days postinoculation. The X-axis indicates days post-inoculation and the Y-axis indicates the mean CFU expressed as the log 10 mean +/- the standard error (error bars). The dashed line indicates the lower limit of detection (log 10 1). FIG. 4. Colonization of the swine respiratory tract by wild-type B. bronchiseptica strain KM22 or TN31, a KM22 Bvg i isogenic mutant. Groups of 16 pigs were inoculated intranasally with KM22 (open triangles) or TN31, a KM22 Bvg i isogenic mutant (open circles). Bacterial load in the nasal cavity (A) was quantified 1, 3, 5, 7, 14, 21, 28, 35, 42, 49, and 56 days post-inoculation. Bacterial load in the trachea (B) and the lungs (C) was quantified 7, 14, 28, and 56 days postinoculation. The X-axis indicates days post-inoculation and the Y-axis indicates the mean CFU expressed as the log 10 mean +/- the standard error (error bars). The dashed line indicates the lower limit of detection (log 10 1). Statistical difference is indicated as P< for TN31 (Bvg i ) compared to KM22 assessed as repeated measurements over all time points (a), P< for TN31 (Bvg i ) compared to KM22 assessed as repeated measurements over all time points (b), P< for TN31 (Bvg i ) compared to KM22 assessed as repeated measurements over all time points (c). A P value less than 0.05 was considered significant FIG. 5. Experimental design of direct and indirect contact transmission experiment. Two pens, indicated as pen A and pen B, were placed 18 inches apart in one isolator room in a BL2 barn facility. Two pigs were intranasally inoculated with the B. bronchiseptica strain tested, placed in 36
37 a pen A, and served as primary challenged pigs, indicated as P. After two days, three naïve pigs, serving as a direct contacts, indicated as D were added to pen A, and five naïve pigs were placed in a pen B, serving as indirect contacts, indicated as I. Nasal colonization of all pigs was monitored by periodical nasal swabbing and necropsy of all pigs was performed on day 25. Downloaded from on August 31, 2018 by guest 37
38 809 TABLE 1. Primers used for cloning and quantitative real-time PCR in this study. Primer Name or Gene Target Sequence (5 to 3 ) bvgs-for ACCTCGCCAAACGCAACAATCTCG bvgs-rev ATCTGCTGCCATGCCGGTATCTGC bvgs-fa-for GAGCTCTTAGGTTTTGAAATCCAGCGCAGTAGTCT bvgs-fa-rev TCTAGACCGCAGGCGGTGCGGGGCGGGCAT bvgs-fb-for TCTAGATGGCTACGCGCCTGGCTGGAGCAAC bvgs-fb-rev ACTTTTCCCAATCGTACTGCGTGAAGCTT TN42-for GAGCTCTTAGGTTTTGAAATCCAGCGCAGTA TN42-rev CCTAGGAAGCTTCACGCAGTACGATTGGGAAAAG R570Hfor CCGCTCGGCCCGCTTGTGCTGGCGGATCTGGCG R570Hrev CGCCAGATCCGCCAGCACAAGCGGGCCGAGCGG T733Mfor GCCGATGATCGCGTTCATCGGCATGCGGATCTCGTGGCTCATCG T733Mrev CGATGAGCCACGAGATCCGCATGCCGATGAACGCGATCATCGGC bvgs-5 -attb1 GGGGACAAGTTTGTACAAAAAAGCAGGCTACACCTCGCCAAACGCAACAATCTCG bvgs-3 -attb2 GGGGACCACTTTGTACAAGAAAGCTGGGTAATCTGCTGCCATGCCGGTATCTGC TN18-for CCTAGGTACTACCGCCGCGAGATGCCGCCCA TN18-rev ACTAGTGCAACAATCTCGCCTAGCGCCGCGCAT flhc CTCGGTGGACTGGTTCATGA CGTTGTAGAACAGCGACGAATG chez TGATGGCGCAGGATTTCC GCCCACCACGTCCATCAT flgb CTGATGTACCGCCTGCCTTAC GCTGTCCATGTCGACGGTATT bcfa GCTGCACGATCCAAATTATCG TGTTCATGGTGTCGAGATAGATGA cyaa CACTGAGCAGAACAATCCTTTCC CGTGAGCATCTGGCTTTCAC prn CAGCACGGCATCCACATC GCCTGACGACCGCTTACC bopn TGCCGAGGAAAAGCATCACT GCCAGAGCATCGGACGTT bopd CGGCTCGGTGAAGACATCTAC GCCTCCCGCATCTGTTGA bopb GCTCAATTCGACGAGGCCTAT TGTGCGTACTCGCCATATCG fim3 CATGGGCACCGACGAAA ACATCAGTGCCCGTGAAGGT bipa CGCGATCACGAACATGGA TTGCCGGAGACGGTAACG fima CAGACGAAAGCCGACAACGT TGGCGGAACCATCCAGAT bvga TTGACGATCACCCTGTACTGAGA 38
39 TCGAATCCTTCCTTTTCCATCA bvgs CGGCCGAGTCGATTCTTG GAATCAGTTCTTCGAGCAATTGC bvgr TGCTTACCGTCAGTACGTTCGA GCATACATGAGTTCTGGCATCAG fimn GCGACGCCGTTTTCCA CATTGCCCAGCGATTCG fimx fim2 dnt 16S CACGATCGAGGACCCGAG TTGGAGATCGTGGGCAGG TGCGATCATCGCTGCAA TTCTGGTTGGAGATGCGGAT GCAGAAAGTACGGCACTACAAGGT CCTGTTGTGATTTTCGATTCCA TCAGCATGTCGCGGTGAAT TGTGACGGGCGGTGTGTA Downloaded from on August 31, 2018 by guest 39
40 TABLE 2. Mean turbinate atrophy scores and percentages of the lungs affected by pneumonia in pigs inoculated with B. bronchiseptica KM22, TN23 (Bvg + isogenic mutant), or TN30 (Bvg - isogenic mutant) Day of Necropsy Group Mean turbinate atrophy Mean % pneumonia score (range) (range) 7 PBS Control 0(0) 0 (0) KM22 (Bvg WT ) 2.25 (1-4) 1.15 (0.1-2) TN23 (Bvg + ) 1.5 (0-4) (0.1-3) TN30 (Bvg - ) 0(0) 0 (0) 14 PBS Control 0 (0) 0 (0) KM22 (Bvg WT ) 2.25 (0-4) (0-9) TN23 (Bvg + ) 3.75 (2-5) 2.25 (0-9) TN30 (Bvg - ) 0 (0) 0 (0) 31 PBS Control 1.5 (1-2) 0 (0) KM22 (Bvg WT ) 4.25 (1-8) (0-1.5) TN23 (Bvg + ) 2.0 (0-3) (0-2.5) TN30 (Bvg - ) 0 (0) 0 (0) 57 PBS Control 1.5 (1-2) 0 (0) KM22 (Bvg WT ) 1.67 (0-3) 2.75 (0-11) TN23 (Bvg + ) 2.25 (2-3) (0-18) TN30 (Bvg - ) 0 (0) 0 (0) 40
41 TABLE 3. Mean turbinate atrophy scores and percentages of the lungs affected by pneumonia in pigs inoculated with B. bronchiseptica KM22 or TN31 (Bvg i isogenic mutant) Day of Necropsy Group Mean turbinate atrophy score (range) Mean % pneumonia (range) 7 PBS Control 1.5 (0-2) (0-2) KM22 (Bvg WT ) 3.75 (1-8) (0-10) TN31 (Bvg i ) 1.25 (0-2) 1.0 (0-3.5) 14 PBS Control 0.5 (0-1) 0.75 (0-3) KM22 (Bvg WT ) 4.5 (2-8) 7.5 (2-20) TN31 (Bvg i ) 1.0 (0-2) 0.25 (0-1) 28 PBS Control 0.25 (0-1) 0.25 (0-1) KM22 (Bvg WT ) 5.75 (3-8) 9.75 (0-17) TN31 (Bvg i ) 1.5 (1-4) 0 (0) 56 PBS Control 1.25 (1-2) 0 (0) KM22 (Bvg WT ) 4.75 (4-6) 5.0 (2-13) TN31 (Bvg i ) 1.5 (0-4) (0-3.5) 41
42 TABLE 4. Direct transmission to naïve pigs a. Contact strain Days b KM22 (Bvg WT ) TN23 (Bvg + ) 2 0/2 c 1/2 8 2/2 2/2 84 2/2 2/2 a. Three days after 16 primary pigs were challenged by the intranasal route with 10 6 CFU of the contact strain, two naïve pigs were added to each room. Four primary challenged pigs were removed on days 7, 14, 31, and 57 for necropsy, leaving only transmission pigs in the room beyond day 57. b. Days after initial contact with primary challenged pigs. c. Data are reported as the number of contact pigs positive out of a possible of two pigs for respective strain of B. bronchiseptica. TABLE 5. Respiratory tract colonization in direct transmission pigs 84 days after initial contact with primary challenged pigs a. Contact Strain KM22 (Bvg WT ) TN23 (Bvg + ) Sample Site Positive CFU/ml b Positive CFU/ml Nasal Wash 2/2 c 2.10/ /2 3.64/ 2.51 Tracheal Wash 1/2 2.24/ 0 2/2 2.88/ 1.74 Lung Lavage 1/2 2.04/ 0 0/2 0/ 0 a. Pigs were euthanized on day 87 of the experiment, 84-days after first contact with primary challenged pigs. b. CFU is reported as the log 10. c. Data are reported as the number of contact pigs positive out of a possible of two pigs for respective strain of B. bronchiseptica. 42
43 841 TABLE 6. Direct and indirect transmission to naïve pigs a Contact Strain KM22 (Bvg WT ) c TN23 (Bvg + ) Days b Primary Direct Indirect Primary Direct Indirect 4 2/2 d 2/3 0/5 2/2 3/3 0/5 7 2/2 2/3 0/5 2/2 3/3 0/5 12 2/2 2/3 1/5 2/2 3/3 1/5 18 2/2 3/3 2/5 2/2 3/3 3/5 25 2/2 3/3 5/5 2/2 3/3 5/5 a. Two days after 2 primary pigs were challenged by the intranasal route with 10 6 CFU of the contact strain, three naïve pigs were added to the same pin and represent direct contact pigs. On the same day five naïve pigs were added to a separate pin and represent indirect contact pigs. One isolator room was used per contact strain studied and the experimental procedure was duplicated in the two isolator rooms used. b. Days after initial contact with primary challenged pigs. c. Contact strain is reported as the name (phenotype). d. Data are reported as the number of contact pigs positive out of the total number possible for respective strain of B. bronchiseptica. 43
44 TABLE 7. Respiratory tract colonization in direct and indirect transmission pigs 25 days after initial contact with primary challenged pigs a Contact Strain KM22 b (Bvg WT ) TN23 (Bvg + ) Sample Site Primary Direct Indirect Primary Direct Indirect Nasal Wash 2/2 b 3/3 5/5 2/2 3/3 5/5 Tracheal Wash 2/2 3/3 5/5 2/2 3/3 5/5 Lung Lavage 2/2 3/3 5/5 2/2 3/3 5/5 a. Pigs were euthanized on day 27 of the experiment, 25-days after initial contact with primary challenged pigs. b. Data are reported as the number of contact pigs positive out of a possible of two pigs for respective strain of B. bronchiseptica. 44
45

46
47
48
49
50
Microarray and Functional Analysis of Growth Phase-Dependent Gene Regulation in Bordetella bronchiseptica
 INFECTION AND IMMUNITY, Oct. 2009, p. 4221 4231 Vol. 77, No. 10 0019-9567/09/$08.00 0 doi:10.1128/iai.00136-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Microarray and Functional
INFECTION AND IMMUNITY, Oct. 2009, p. 4221 4231 Vol. 77, No. 10 0019-9567/09/$08.00 0 doi:10.1128/iai.00136-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Microarray and Functional
Role of the Type III Secretion System in a Hypervirulent Lineage of Bordetella bronchiseptica
 INFECTION AND IMMUNITY, Sept. 2009, p. 3969 3977 Vol. 77, No. 9 0019-9567/09/$08.00 0 doi:10.1128/iai.01362-08 Copyright 2009, American Society for Microbiology. All Rights Reserved. Role of the Type III
INFECTION AND IMMUNITY, Sept. 2009, p. 3969 3977 Vol. 77, No. 9 0019-9567/09/$08.00 0 doi:10.1128/iai.01362-08 Copyright 2009, American Society for Microbiology. All Rights Reserved. Role of the Type III
Running title: Contribution of Bordetella Bps to Biofilm Formation and Respiratory Disease in
 IAI Accepted Manuscript Posted Online 30 May 2017 Infect. Immun. doi:10.1128/iai.00261-17 Copyright 2017 American Society for Microbiology. All Rights Reserved. 1 2 The Bordetella Bps Polysaccharide is
IAI Accepted Manuscript Posted Online 30 May 2017 Infect. Immun. doi:10.1128/iai.00261-17 Copyright 2017 American Society for Microbiology. All Rights Reserved. 1 2 The Bordetella Bps Polysaccharide is
Identification of a Locus Required for the Regulation of bvg- Repressed Genes in Bordetella pertussis
 JOURNAL OF BACTERIOLOGY, May 1995, p. 2727 2736 Vol. 177, No. 10 0021-9193/95/$04.00 0 Copyright 1995, American Society for Microbiology Identification of a Locus Required for the Regulation of bvg- Repressed
JOURNAL OF BACTERIOLOGY, May 1995, p. 2727 2736 Vol. 177, No. 10 0021-9193/95/$04.00 0 Copyright 1995, American Society for Microbiology Identification of a Locus Required for the Regulation of bvg- Repressed
Activation of the vrg6 Promoter of Bordetella pertussis by RisA
 JOURNAL OF BACTERIOLOGY, Mar. 2005, p. 1648 1658 Vol. 187, No. 5 0021-9193/05/$08.00 0 doi:10.1128/jb.187.5.1648 1658.2005 Activation of the vrg6 Promoter of Bordetella pertussis by RisA Tadhg Ó Cróinín,
JOURNAL OF BACTERIOLOGY, Mar. 2005, p. 1648 1658 Vol. 187, No. 5 0021-9193/05/$08.00 0 doi:10.1128/jb.187.5.1648 1658.2005 Activation of the vrg6 Promoter of Bordetella pertussis by RisA Tadhg Ó Cróinín,
1. Division of Bacterial, Parasitic, and Allergenic Products, Center for Biologics Evaluation and
 JB Accepted Manuscript Posted Online 30 July 2018 J. Bacteriol. doi:10.1128/jb.00175-18 This is a work of the U.S. Government and is not subject to copyright protection in the United States. Foreign copyrights
JB Accepted Manuscript Posted Online 30 July 2018 J. Bacteriol. doi:10.1128/jb.00175-18 This is a work of the U.S. Government and is not subject to copyright protection in the United States. Foreign copyrights
Bordetella bronchiseptica: A Candidate Mucosal Vaccine Vector
 University of Tennessee, Knoxville Trace: Tennessee Research and Creative Exchange Doctoral Dissertations Graduate School 5-2002 Bordetella bronchiseptica: A Candidate Mucosal Vaccine Vector Sreekumari
University of Tennessee, Knoxville Trace: Tennessee Research and Creative Exchange Doctoral Dissertations Graduate School 5-2002 Bordetella bronchiseptica: A Candidate Mucosal Vaccine Vector Sreekumari
Growth Phase- and Nutrient Limitation-Associated Transcript Abundance Regulation in Bordetella pertussis
 INFECTION AND IMMUNITY, Oct. 2006, p. 5537 5548 Vol. 74, No. 10 0019-9567/06/$08.00 0 doi:10.1128/iai.00781-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Growth Phase- and
INFECTION AND IMMUNITY, Oct. 2006, p. 5537 5548 Vol. 74, No. 10 0019-9567/06/$08.00 0 doi:10.1128/iai.00781-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Growth Phase- and
Methicillin-Resistant Staphylococcus aureus
 Methicillin-Resistant Staphylococcus aureus By Karla Givens Means of Transmission and Usual Reservoirs Staphylococcus aureus is part of normal flora and can be found on the skin and in the noses of one
Methicillin-Resistant Staphylococcus aureus By Karla Givens Means of Transmission and Usual Reservoirs Staphylococcus aureus is part of normal flora and can be found on the skin and in the noses of one
MRSA surveillance 2014: Poultry
 Vicky Jasson MRSA surveillance 2014: Poultry 1. Introduction In the framework of the FASFC surveillance, a surveillance of MRSA in poultry has been executed in order to determine the prevalence and diversity
Vicky Jasson MRSA surveillance 2014: Poultry 1. Introduction In the framework of the FASFC surveillance, a surveillance of MRSA in poultry has been executed in order to determine the prevalence and diversity
Guidelines for Laboratory Verification of Performance of the FilmArray BCID System
 Guidelines for Laboratory Verification of Performance of the FilmArray BCID System Purpose The Clinical Laboratory Improvement Amendments (CLIA), passed in 1988, establishes quality standards for all laboratory
Guidelines for Laboratory Verification of Performance of the FilmArray BCID System Purpose The Clinical Laboratory Improvement Amendments (CLIA), passed in 1988, establishes quality standards for all laboratory
SUMMARY OF PRODUCT CHARACTERISTICS
 SUMMARY OF PRODUCT CHARACTERISTICS 1. NAME OF THE VETERINARY MEDICINAL PRODUCT Oxycare 20 %w/v LA Solution for Injection 2. QUALITATIVE AND QUANTITATIVE COMPOSITION Active Substance: Oxytetracycline (Equivalent
SUMMARY OF PRODUCT CHARACTERISTICS 1. NAME OF THE VETERINARY MEDICINAL PRODUCT Oxycare 20 %w/v LA Solution for Injection 2. QUALITATIVE AND QUANTITATIVE COMPOSITION Active Substance: Oxytetracycline (Equivalent
Co-transfer of bla NDM-5 and mcr-1 by an IncX3 X4 hybrid plasmid in Escherichia coli 4
 SUPPLEMENTARY INFORMATION ARTICLE NUMBER: 16176 DOI: 10.1038/NMICROBIOL.2016.176 Co-transfer of bla NDM-5 and mcr-1 by an IncX3 X4 hybrid plasmid in Escherichia coli 4 5 6 7 8 9 10 11 12 13 14 15 16 17
SUPPLEMENTARY INFORMATION ARTICLE NUMBER: 16176 DOI: 10.1038/NMICROBIOL.2016.176 Co-transfer of bla NDM-5 and mcr-1 by an IncX3 X4 hybrid plasmid in Escherichia coli 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Interpretation of results from milk samples tested for mastitis bacteria with Mastit 4 qpcr test from DNA Diagnostic
 Mastit 4 Interpretation of results from milk samples tested for mastitis bacteria with Mastit 4 qpcr test from DNA Diagnostic The 40th ICAR Biennial Session Puerto Varas, Chile, 24-28 october 2016 Jorgen
Mastit 4 Interpretation of results from milk samples tested for mastitis bacteria with Mastit 4 qpcr test from DNA Diagnostic The 40th ICAR Biennial Session Puerto Varas, Chile, 24-28 october 2016 Jorgen
The Bvg Virulence Control System Regulates Biofilm Formation in Bordetella bronchiseptica
 JOURNAL OF BACTERIOLOGY, Sept. 2004, p. 5692 5698 Vol. 186, No. 17 0021-9193/04/$08.00 0 DOI: 10.1128/JB.186.17.5692 5698.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. The
JOURNAL OF BACTERIOLOGY, Sept. 2004, p. 5692 5698 Vol. 186, No. 17 0021-9193/04/$08.00 0 DOI: 10.1128/JB.186.17.5692 5698.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. The
Neither the Bvg Phase nor the vrg6 Locus of Bordetella pertussis Is Required for Respiratory Infection in Mice
 INFECTION AND IMMUNITY, June 1998, p. 2762 2768 Vol. 66, No. 6 0019-9567/98/$04.00 0 Copyright 1998, American Society for Microbiology Neither the Bvg Phase nor the vrg6 Locus of Bordetella pertussis Is
INFECTION AND IMMUNITY, June 1998, p. 2762 2768 Vol. 66, No. 6 0019-9567/98/$04.00 0 Copyright 1998, American Society for Microbiology Neither the Bvg Phase nor the vrg6 Locus of Bordetella pertussis Is
Role of Antibodies in Immunity to Bordetella Infections
 INFECTION AND IMMUNITY, Apr. 2003, p. 1719 1724 Vol. 71, No. 4 0019-9567/03/$08.00 0 DOI: 10.1128/IAI.71.4.1719 1724.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Role of
INFECTION AND IMMUNITY, Apr. 2003, p. 1719 1724 Vol. 71, No. 4 0019-9567/03/$08.00 0 DOI: 10.1128/IAI.71.4.1719 1724.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Role of
BIOLACTAM. Product Description. An innovative in vitro diagnostic for the rapid quantitative determination of ß-lactamase activity
 BIOLACTAM www.biolactam.eu An innovative in vitro diagnostic for the rapid quantitative determination of ß-lactamase activity 1.5-3h 20 Copyright 2014 VL-Diagnostics GmbH. All rights reserved. Product
BIOLACTAM www.biolactam.eu An innovative in vitro diagnostic for the rapid quantitative determination of ß-lactamase activity 1.5-3h 20 Copyright 2014 VL-Diagnostics GmbH. All rights reserved. Product
Summary of Product Characteristics
 Summary of Product Characteristics 1 NAME OF THE VETERINARY MEDICINAL PRODUCT Selectan 300 mg/ml solution for injection for cattle and swine. 2 QUALITATIVE AND QUANTITATIVE COMPOSITION Each ml contains:
Summary of Product Characteristics 1 NAME OF THE VETERINARY MEDICINAL PRODUCT Selectan 300 mg/ml solution for injection for cattle and swine. 2 QUALITATIVE AND QUANTITATIVE COMPOSITION Each ml contains:
Test Method Modified Association of Analytical Communities Test Method Modified Germicidal Spray Products as Disinfectants
 Study Title Antibacterial Activity and Efficacy of E-Mist Innovations' Electrostatic Sprayer Product with Multiple Disinfectants Method Modified Association of Analytical Communities Method 961.02 Modified
Study Title Antibacterial Activity and Efficacy of E-Mist Innovations' Electrostatic Sprayer Product with Multiple Disinfectants Method Modified Association of Analytical Communities Method 961.02 Modified
Consequences of Antimicrobial Resistant Bacteria. Antimicrobial Resistance. Molecular Genetics of Antimicrobial Resistance. Topics to be Covered
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Regulatory Mutants of Bordetella bronchiseptica in a
 INFCTION AND IMMUNITY, Aug. 1994, P. 3381-339 19-9567/94/$4.+ Copyright 3 1994, American Society for Microbiology Vol. 62, No. 8 BvgAS-Mediated Signal Transduction: Analysis of Phase-Locked Regulatory
INFCTION AND IMMUNITY, Aug. 1994, P. 3381-339 19-9567/94/$4.+ Copyright 3 1994, American Society for Microbiology Vol. 62, No. 8 BvgAS-Mediated Signal Transduction: Analysis of Phase-Locked Regulatory
MID 23. Antimicrobial Resistance. Consequences of Antimicrobial Resistant Bacteria. Molecular Genetics of Antimicrobial Resistance
 Antimicrobial Resistance Molecular Genetics of Antimicrobial Resistance Micro evolutionary change - point mutations Beta-lactamase mutation extends spectrum of the enzyme rpob gene (RNA polymerase) mutation
Antimicrobial Resistance Molecular Genetics of Antimicrobial Resistance Micro evolutionary change - point mutations Beta-lactamase mutation extends spectrum of the enzyme rpob gene (RNA polymerase) mutation
Antimicrobial Resistance
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Acquisition of Foreign DNA
 Antimicrobial Resistance Acquisition of Foreign DNA Levy, Scientific American Horizontal gene transfer is common, even between Gram positive and negative bacteria Plasmid - transfer of single or multiple
Antimicrobial Resistance Acquisition of Foreign DNA Levy, Scientific American Horizontal gene transfer is common, even between Gram positive and negative bacteria Plasmid - transfer of single or multiple
Burn Infection & Laboratory Diagnosis
 Burn Infection & Laboratory Diagnosis Introduction Burns are one the most common forms of trauma. 2 million fires each years 1.2 million people with burn injuries 100000 hospitalization 5000 patients die
Burn Infection & Laboratory Diagnosis Introduction Burns are one the most common forms of trauma. 2 million fires each years 1.2 million people with burn injuries 100000 hospitalization 5000 patients die
Enzootic Bovine Leukosis: Milk Screening and Verification ELISA: VF-P02210 & VF-P02220
 Enzootic Bovine Leukosis: Milk Screening and Verification ELISA: VF-P02210 & VF-P02220 Introduction Enzootic Bovine Leukosis is a transmissible disease caused by the Enzootic Bovine Leukosis Virus (BLV)
Enzootic Bovine Leukosis: Milk Screening and Verification ELISA: VF-P02210 & VF-P02220 Introduction Enzootic Bovine Leukosis is a transmissible disease caused by the Enzootic Bovine Leukosis Virus (BLV)
READER S DIGEST OVERVIEW: BIGHORN SHEEP. Peregrine Wolff, DVM
 READER S DIGEST OVERVIEW: RESPIRATORY DISEASE IN BIGHORN SHEEP Peregrine Wolff, DVM Nevada Department of Wildlife During the Lewis & Clark expedition (1804 1806) There may have been 2 million bighorn sheep
READER S DIGEST OVERVIEW: RESPIRATORY DISEASE IN BIGHORN SHEEP Peregrine Wolff, DVM Nevada Department of Wildlife During the Lewis & Clark expedition (1804 1806) There may have been 2 million bighorn sheep
Impact of Spores on the Comparative Efficacies of Five Antibiotics. Pharmacodynamic Model
 AAC Accepts, published online ahead of print on 12 December 2011 Antimicrob. Agents Chemother. doi:10.1128/aac.01109-10 Copyright 2011, American Society for Microbiology and/or the Listed Authors/Institutions.
AAC Accepts, published online ahead of print on 12 December 2011 Antimicrob. Agents Chemother. doi:10.1128/aac.01109-10 Copyright 2011, American Society for Microbiology and/or the Listed Authors/Institutions.
Evaluation of the Role of the Bvg Intermediate Phase in Bordetella pertussis during Experimental Respiratory Infection
 INFECTION AND IMMUNITY, Feb. 2005, p. 748 760 Vol. 73, No. 2 0019-9567/05/$08.00 0 doi:10.1128/iai.73.2.748 760.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Evaluation of
INFECTION AND IMMUNITY, Feb. 2005, p. 748 760 Vol. 73, No. 2 0019-9567/05/$08.00 0 doi:10.1128/iai.73.2.748 760.2005 Copyright 2005, American Society for Microbiology. All Rights Reserved. Evaluation of
Pipestone Veterinary Services
 NIAA: 2017 Antibiotic Symposium Oct 31 Nov 2, 2017 Joel Nerem, DVM Pipestone Veterinary Services 5 Locations Pipestone, MN Independence, IA Ottumwa, IA DeKalb, IL Rensselaer, IN Mixed Animal Practice.
NIAA: 2017 Antibiotic Symposium Oct 31 Nov 2, 2017 Joel Nerem, DVM Pipestone Veterinary Services 5 Locations Pipestone, MN Independence, IA Ottumwa, IA DeKalb, IL Rensselaer, IN Mixed Animal Practice.
Vaccines for Cats. 2. Feline viral rhinotracheitis, FVR caused by FVR virus, also known as herpes virus type 1, FHV-1
 Vaccines for Cats Recent advances in veterinary medical science have resulted in an increase in the number and type of vaccines that are available for use in cats, and improvements are continuously being
Vaccines for Cats Recent advances in veterinary medical science have resulted in an increase in the number and type of vaccines that are available for use in cats, and improvements are continuously being
Feeding Original XPC TM can help reduce Campylobacter in broilers and turkeys
 As published in RESEARCH UPDATE Campylobacter is one of the leading causes of foodborne illness. Traditional methods for controlling Campylobacter contamination have been focused within the processing
As published in RESEARCH UPDATE Campylobacter is one of the leading causes of foodborne illness. Traditional methods for controlling Campylobacter contamination have been focused within the processing
Filamentous Hemagglutinin of Bordetella bronchiseptica Is Required for Efficient Establishment of Tracheal Colonization
 INFECTION AND IMMUNITY, Dec. 1998, p. 5921 5929 Vol. 66, No. 12 0019-9567/98/$04.00 0 Copyright 1998, American Society for Microbiology. All Rights Reserved. Filamentous Hemagglutinin of Bordetella bronchiseptica
INFECTION AND IMMUNITY, Dec. 1998, p. 5921 5929 Vol. 66, No. 12 0019-9567/98/$04.00 0 Copyright 1998, American Society for Microbiology. All Rights Reserved. Filamentous Hemagglutinin of Bordetella bronchiseptica
R-factor mediated trimethoprim resistance: result of two three-month clinical surveys
 Journal of Clinical Pathology, 1978, 31, 850-854 R-factor mediated trimethoprim resistance: result of two three-month clinical surveys S. G. B. AMYES1, A. M. EMMERSON2, AND J. T. SMITH3 From the 'Department
Journal of Clinical Pathology, 1978, 31, 850-854 R-factor mediated trimethoprim resistance: result of two three-month clinical surveys S. G. B. AMYES1, A. M. EMMERSON2, AND J. T. SMITH3 From the 'Department
Attorneys for Plaintiffs Hells Canyon Preservation Council and The Wilderness Society UNITED STATES DISTRICT COURT FOR THE DISTRICT OF IDAHO
 Lauren M. Rule (ISB # 6863 ADVOCATES FOR THE WEST PO Box 1612 Boise ID 83701 (208 342-7024 lrule@advocateswest.org Attorney for Plaintiff Western Watersheds Project Jennifer R. Schemm (OSB #97008 602 O
Lauren M. Rule (ISB # 6863 ADVOCATES FOR THE WEST PO Box 1612 Boise ID 83701 (208 342-7024 lrule@advocateswest.org Attorney for Plaintiff Western Watersheds Project Jennifer R. Schemm (OSB #97008 602 O
PCR detection of Leptospira in. stray cat and
 PCR detection of Leptospira in 1 Department of Pathology, School of Veterinary Medicine, Islamic Azad University, Shahrekord Branch, Shahrekord, Iran 2 Department of Microbiology, School of Veterinary
PCR detection of Leptospira in 1 Department of Pathology, School of Veterinary Medicine, Islamic Azad University, Shahrekord Branch, Shahrekord, Iran 2 Department of Microbiology, School of Veterinary
Overview. There are commonly found arrangements of bacteria based on their division. Spheres, Rods, Spirals
 Bacteria Overview Bacteria live almost everywhere. Most are microscopic ranging from 0.5 5 m in size, and unicellular. They have a variety of shapes when viewed under a microscope, most commonly: Spheres,
Bacteria Overview Bacteria live almost everywhere. Most are microscopic ranging from 0.5 5 m in size, and unicellular. They have a variety of shapes when viewed under a microscope, most commonly: Spheres,
Veterinary Diagnostics Portfolio Overview. Complete solutions for veterinary testing and pathogen research
 Veterinary Diagnostics Portfolio Overview Complete solutions for veterinary testing and pathogen research Sample preparation products Cat. no. (number of preps) Target analyte Product Short description
Veterinary Diagnostics Portfolio Overview Complete solutions for veterinary testing and pathogen research Sample preparation products Cat. no. (number of preps) Target analyte Product Short description
Detection and Quantitation of the Etiologic Agents of Ventilator Associated Pneumonia in Endotracheal Tube Aspirates From Patients in Iran
 Letter to the Editor Detection and Quantitation of the Etiologic Agents of Ventilator Associated Pneumonia in Endotracheal Tube Aspirates From Patients in Iran Mohammad Rahbar, PhD; Massoud Hajia, PhD
Letter to the Editor Detection and Quantitation of the Etiologic Agents of Ventilator Associated Pneumonia in Endotracheal Tube Aspirates From Patients in Iran Mohammad Rahbar, PhD; Massoud Hajia, PhD
bvg Repression of Alcaligin Synthesis in Bordetella bronchiseptica Is Associated with Phylogenetic Lineage
 JOURNAL OF BACTERIOLOGY, Nov. 1995, p. 6058 6063 Vol. 177, No. 21 0021-9193/95/$04.00 0 Copyright 1995, American Society for Microbiology bvg Repression of Alcaligin Synthesis in Bordetella bronchiseptica
JOURNAL OF BACTERIOLOGY, Nov. 1995, p. 6058 6063 Vol. 177, No. 21 0021-9193/95/$04.00 0 Copyright 1995, American Society for Microbiology bvg Repression of Alcaligin Synthesis in Bordetella bronchiseptica
Virulence Factors of Bordetella bronchiseptica Associated with the
 INFECTION AND IMMUNITY, Jan. 1987, p. 217-222 0019-9567/87/010217-06$02.00/0 Copyright 1987, American Society for Microbiology Vol. 55, No. 1 Virulence Factors of Bordetella bronchiseptica Associated with
INFECTION AND IMMUNITY, Jan. 1987, p. 217-222 0019-9567/87/010217-06$02.00/0 Copyright 1987, American Society for Microbiology Vol. 55, No. 1 Virulence Factors of Bordetella bronchiseptica Associated with
GeNei TM. Antibiotic Sensitivity. Teaching Kit Manual KT Revision No.: Bangalore Genei, 2007 Bangalore Genei, 2007
 GeNei Bacterial Antibiotic Sensitivity Teaching Kit Manual Cat No. New Cat No. KT68 106333 Revision No.: 00180705 CONTENTS Page No. Objective 3 Principle 3 Kit Description 4 Materials Provided 5 Procedure
GeNei Bacterial Antibiotic Sensitivity Teaching Kit Manual Cat No. New Cat No. KT68 106333 Revision No.: 00180705 CONTENTS Page No. Objective 3 Principle 3 Kit Description 4 Materials Provided 5 Procedure
TEST REPORT. Client: M/s Ion Silver AB. Loddekopinge. Sverige / SWEDEN. Chandran. min and 30 min. 2. E. coli. 1. S. aureus
 TEST REPORT TEST TYPE: Liquid Suspension Time Kill Study -Quantitative Test Based On ASTM 2315 TEST METHOD of Colloidal Silver Product at Contact time points: 30 sec, 1 min, 2 min, 5 min, 10 min, 15 min
TEST REPORT TEST TYPE: Liquid Suspension Time Kill Study -Quantitative Test Based On ASTM 2315 TEST METHOD of Colloidal Silver Product at Contact time points: 30 sec, 1 min, 2 min, 5 min, 10 min, 15 min
Antimicrobial Resistance
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of Change in the approach to the administration of empiric antimicrobial therapy Increased
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of Change in the approach to the administration of empiric antimicrobial therapy Increased
Antibiotics & Resistance
 What are antibiotics? Antibiotics & esistance Antibiotics are molecules that stop bacteria from growing or kill them Antibiotics, agents against life - either natural or synthetic chemicals - designed
What are antibiotics? Antibiotics & esistance Antibiotics are molecules that stop bacteria from growing or kill them Antibiotics, agents against life - either natural or synthetic chemicals - designed
Isolation of antibiotic producing Actinomycetes from soil of Kathmandu valley and assessment of their antimicrobial activities
 International Journal of Microbiology and Allied Sciences (IJOMAS) ISSN: 2382-5537 May 2016, 2(4):22-26 IJOMAS, 2016 Research Article Page: 22-26 Isolation of antibiotic producing Actinomycetes from soil
International Journal of Microbiology and Allied Sciences (IJOMAS) ISSN: 2382-5537 May 2016, 2(4):22-26 IJOMAS, 2016 Research Article Page: 22-26 Isolation of antibiotic producing Actinomycetes from soil
Index. Note: Page numbers of article titles are in boldface type.
 Index Note: Page numbers of article titles are in boldface type. A Abdominal viscera, examination of, in investigation of emerging infectious diseases of food animals, 6 American Veterinary Medical Association,
Index Note: Page numbers of article titles are in boldface type. A Abdominal viscera, examination of, in investigation of emerging infectious diseases of food animals, 6 American Veterinary Medical Association,
Defining Resistance and Susceptibility: What S, I, and R Mean to You
 Defining Resistance and Susceptibility: What S, I, and R Mean to You Michael D. Apley, DVM, PhD, DACVCP Department of Clinical Sciences College of Veterinary Medicine Kansas State University Susceptible
Defining Resistance and Susceptibility: What S, I, and R Mean to You Michael D. Apley, DVM, PhD, DACVCP Department of Clinical Sciences College of Veterinary Medicine Kansas State University Susceptible
Burton's Microbiology for the Health Sciences. Chapter 9. Controlling Microbial Growth in Vivo Using Antimicrobial Agents
 Burton's Microbiology for the Health Sciences Chapter 9. Controlling Microbial Growth in Vivo Using Antimicrobial Agents Chapter 9 Outline Introduction Characteristics of an Ideal Antimicrobial Agent How
Burton's Microbiology for the Health Sciences Chapter 9. Controlling Microbial Growth in Vivo Using Antimicrobial Agents Chapter 9 Outline Introduction Characteristics of an Ideal Antimicrobial Agent How
6.0 ANTIBACTERIAL ACTIVITY OF CAROTENOID FROM HALOMONAS SPECIES AGAINST CHOSEN HUMAN BACTERIAL PATHOGENS
 6.0 ANTIBACTERIAL ACTIVITY OF CAROTENOID FROM HALOMONAS SPECIES AGAINST CHOSEN HUMAN BACTERIAL PATHOGENS 6.1 INTRODUCTION Microorganisms that cause infectious disease are called pathogenic microbes. Although
6.0 ANTIBACTERIAL ACTIVITY OF CAROTENOID FROM HALOMONAS SPECIES AGAINST CHOSEN HUMAN BACTERIAL PATHOGENS 6.1 INTRODUCTION Microorganisms that cause infectious disease are called pathogenic microbes. Although
MATTHEW S. CONOVER. A Dissertation Submitted to the Graduate Faculty of WAKE FOREST UNIVERSITY GRADUATE SCHOOL OF ARTS AND SCIENCES
 AN EXAMINATION OF THE FUNCTION AND THE TRANSCRIPTIONAL REGULATION OF THE BPS POLYSACCHARIDE IN BORDETELLA PERTUSSIS PATHOGENESIS AND BIOFILM DEVELOPMENT BY MATTHEW S. CONOVER A Dissertation Submitted to
AN EXAMINATION OF THE FUNCTION AND THE TRANSCRIPTIONAL REGULATION OF THE BPS POLYSACCHARIDE IN BORDETELLA PERTUSSIS PATHOGENESIS AND BIOFILM DEVELOPMENT BY MATTHEW S. CONOVER A Dissertation Submitted to
Association between teat skin colonization and intramammary infections with Staphylococcus aureus and Streptococcus agalactiae
 15/11/2017 1 Association between teat skin colonization and intramammary infections with Staphylococcus aureus and Streptococcus agalactiae Line Svennesen (PhD student) Yasser Mahmmod 1, Karl Pedersen
15/11/2017 1 Association between teat skin colonization and intramammary infections with Staphylococcus aureus and Streptococcus agalactiae Line Svennesen (PhD student) Yasser Mahmmod 1, Karl Pedersen
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
www.sciencemag.org/cgi/content/full/319/5870/1679/dc1 Supporting Online Material for Drosophila Egg-Laying Site Selection as a System to Study Simple Decision-Making Processes Chung-hui Yang, Priyanka
Microbiology: Practical Competence
 Microbiology: Practical Competence Introduction Infectious diseases in animals are caused by the invasion of tissues by bacteria, especially the epithelium, by microorganisms. This invasion have many effects
Microbiology: Practical Competence Introduction Infectious diseases in animals are caused by the invasion of tissues by bacteria, especially the epithelium, by microorganisms. This invasion have many effects
Diurnal variation in microfilaremia in cats experimentally infected with larvae of
 Hayasaki et al., Page 1 Short Communication Diurnal variation in microfilaremia in cats experimentally infected with larvae of Dirofilaria immitis M. Hayasaki a,*, J. Okajima b, K.H. Song a, K. Shiramizu
Hayasaki et al., Page 1 Short Communication Diurnal variation in microfilaremia in cats experimentally infected with larvae of Dirofilaria immitis M. Hayasaki a,*, J. Okajima b, K.H. Song a, K. Shiramizu
Regeneration of toxigenic Pasteurella multocida induced severe turbinate atrophy in pigs detected by computed tomography
 Magyar et al. BMC Veterinary Research 2013, 9:222 RESEARCH ARTICLE Open Access Regeneration of toxigenic Pasteurella multocida induced severe turbinate atrophy in pigs detected by computed tomography Tibor
Magyar et al. BMC Veterinary Research 2013, 9:222 RESEARCH ARTICLE Open Access Regeneration of toxigenic Pasteurella multocida induced severe turbinate atrophy in pigs detected by computed tomography Tibor
On- Farm Necropsies Who, What, Where, When and Why
 On- Farm Necropsies Who, What, Where, When and Why Thank you for par-cipa-ng in PorkBridge 2014. To start the presenta-on, advance one slide by pressing enter or the down arrow or right arrow key. Locke
On- Farm Necropsies Who, What, Where, When and Why Thank you for par-cipa-ng in PorkBridge 2014. To start the presenta-on, advance one slide by pressing enter or the down arrow or right arrow key. Locke
Probing the Function of Bordetella bronchiseptica Adenylate Cyclase Toxin by Manipulating Host Immunity
 INFECTION AND IMMUNITY, Mar. 1999, p. 1493 1500 Vol. 67, No. 3 0019-9567/99/$04.00 0 Copyright 1999, American Society for Microbiology. All Rights Reserved. Probing the Function of Bordetella bronchiseptica
INFECTION AND IMMUNITY, Mar. 1999, p. 1493 1500 Vol. 67, No. 3 0019-9567/99/$04.00 0 Copyright 1999, American Society for Microbiology. All Rights Reserved. Probing the Function of Bordetella bronchiseptica
Bovine Mastitis Products for Microbiological Analysis
 Bovine Mastitis Products for Microbiological Analysis 121917ss Hardy Diagnostics has everything for your laboratory! SAVE MONEY Now you have a choice for obtaining your supplies for mastitis testing. Hardy
Bovine Mastitis Products for Microbiological Analysis 121917ss Hardy Diagnostics has everything for your laboratory! SAVE MONEY Now you have a choice for obtaining your supplies for mastitis testing. Hardy
Int.J.Curr.Microbiol.App.Sci (2018) 7(8):
 7(8):") International Journal of Current Microbiology and Applied Sciences ISSN: 2319-7706 Volume 7 Number 08 (2018) Journal homepage: http://www.ijcmas.com Original Research Article https://doi.org/10.20546/ijcmas.2018.708.378
International Journal of Current Microbiology and Applied Sciences ISSN: 2319-7706 Volume 7 Number 08 (2018) Journal homepage: http://www.ijcmas.com Original Research Article https://doi.org/10.20546/ijcmas.2018.708.378
Introduction. Rabbit Respiratory Disease. Lecture Outline. Pre-consult. Initial presentation. History 26/01/2013
 Introduction Rabbit Respiratory Disease Joanna Hedley BVM&S DZooMed (Reptilian) MRCVS Rabbit, Exotic Animal and Wildlife Clinician at R(D)SVS Obligate nasal breathers Small thorax Tendency to mask symptoms
Introduction Rabbit Respiratory Disease Joanna Hedley BVM&S DZooMed (Reptilian) MRCVS Rabbit, Exotic Animal and Wildlife Clinician at R(D)SVS Obligate nasal breathers Small thorax Tendency to mask symptoms
SELECT NEWS. Florfenicol Monograph: Injectable & Oral Therapy for Swine
 SELECT NEWS Florfenicol Monograph: Injectable & Oral Therapy for Swine Did you know that? Florfenicol is one of the most powerful antibiotics currently available in veterinary medicine with one of the
SELECT NEWS Florfenicol Monograph: Injectable & Oral Therapy for Swine Did you know that? Florfenicol is one of the most powerful antibiotics currently available in veterinary medicine with one of the
Redefining Infection Management. Proven Clinical Outcomes
 Proven Clinical Outcomes Proof of Bacteria-Binding1 In the first 30 seconds, 1 square centimeter of Cutimed Sorbact binds wound bacteria - after 2 hours, the amount of bacteria bound are more than would
Proven Clinical Outcomes Proof of Bacteria-Binding1 In the first 30 seconds, 1 square centimeter of Cutimed Sorbact binds wound bacteria - after 2 hours, the amount of bacteria bound are more than would
Nicholas Schneider, DVM Schneider Veterinary Services, LLC. Milliken, CO
 Nicholas Schneider, DVM Schneider Veterinary Services, LLC. Milliken, CO Pipestone, MN Native Home of Pipestone Systems 2013 UMN CVM Graduate Schneider Veterinary Services, LLC. Solo Veterinary Practice
Nicholas Schneider, DVM Schneider Veterinary Services, LLC. Milliken, CO Pipestone, MN Native Home of Pipestone Systems 2013 UMN CVM Graduate Schneider Veterinary Services, LLC. Solo Veterinary Practice
SUMMARY OF PRODUCT CHARACTERISTICS
 SUMMARY OF PRODUCT CHARACTERISTICS 1. NAME OF THE VETERINARY MEDICINAL PRODUCT RONAXAN 20mg Tablet 2. QUALITATIVE AND QUANTITATIVE COMPOSITION Each tablet contains: Active substance : Doxycycline (as doxycycline
SUMMARY OF PRODUCT CHARACTERISTICS 1. NAME OF THE VETERINARY MEDICINAL PRODUCT RONAXAN 20mg Tablet 2. QUALITATIVE AND QUANTITATIVE COMPOSITION Each tablet contains: Active substance : Doxycycline (as doxycycline
SUMMARY OF PRODUCT CHARACTERISTICS. NUFLOR 300 mg/ml solution for injection for cattle and sheep
 SUMMARY OF PRODUCT CHARACTERISTICS 1. NAME OF THE VETERINARY MEDICINAL PRODUCT NUFLOR 300 mg/ml solution for injection for cattle and sheep 2. QUALITATIVE AND QUANTITATIVE COMPOSITION Each ml contains:
SUMMARY OF PRODUCT CHARACTERISTICS 1. NAME OF THE VETERINARY MEDICINAL PRODUCT NUFLOR 300 mg/ml solution for injection for cattle and sheep 2. QUALITATIVE AND QUANTITATIVE COMPOSITION Each ml contains:
Mechanisms and Pathways of AMR in the environment
 FMM/RAS/298: Strengthening capacities, policies and national action plans on prudent and responsible use of antimicrobials in fisheries Final Workshop in cooperation with AVA Singapore and INFOFISH 12-14
FMM/RAS/298: Strengthening capacities, policies and national action plans on prudent and responsible use of antimicrobials in fisheries Final Workshop in cooperation with AVA Singapore and INFOFISH 12-14
Presence of extended spectrum β-lactamase producing Escherichia coli in
 1 2 Presence of extended spectrum β-lactamase producing Escherichia coli in wild geese 3 4 5 A. Garmyn* 1, F. Haesebrouck 1, T. Hellebuyck 1, A. Smet 1, F. Pasmans 1, P. Butaye 2, A. Martel 1 6 7 8 9 10
1 2 Presence of extended spectrum β-lactamase producing Escherichia coli in wild geese 3 4 5 A. Garmyn* 1, F. Haesebrouck 1, T. Hellebuyck 1, A. Smet 1, F. Pasmans 1, P. Butaye 2, A. Martel 1 6 7 8 9 10
ZOETIS INC. 333 PORTAGE STREET, KALAMAZOO, MI, Telephone: Customer Service: Website: EXCEDE FOR SWINE
 ZOETIS INC. 333 PORTAGE STREET, KALAMAZOO, MI, 49007 Telephone: 269-359-4414 Customer Service: 888-963-8471 Website: www.zoetis.com Every effort has been made to ensure the accuracy of the information
ZOETIS INC. 333 PORTAGE STREET, KALAMAZOO, MI, 49007 Telephone: 269-359-4414 Customer Service: 888-963-8471 Website: www.zoetis.com Every effort has been made to ensure the accuracy of the information
Visit ABLE on the Web at:
 This article reprinted from: Lessem, P. B. 2008. The antibiotic resistance phenomenon: Use of minimal inhibitory concentration (MIC) determination for inquiry based experimentation. Pages 357-362, in Tested
This article reprinted from: Lessem, P. B. 2008. The antibiotic resistance phenomenon: Use of minimal inhibitory concentration (MIC) determination for inquiry based experimentation. Pages 357-362, in Tested
Author - Dr. Josie Traub-Dargatz
 Author - Dr. Josie Traub-Dargatz Dr. Josie Traub-Dargatz is a professor of equine medicine at Colorado State University (CSU) College of Veterinary Medicine and Biomedical Sciences. She began her veterinary
Author - Dr. Josie Traub-Dargatz Dr. Josie Traub-Dargatz is a professor of equine medicine at Colorado State University (CSU) College of Veterinary Medicine and Biomedical Sciences. She began her veterinary
BvgAS Is Sufficient for Activation of the Bordetella pertussis ptx Locus in Escherichia coli
 JOURNAL OF BACTERIOLOGY, Nov. 1995, p. 6477 6485 Vol. 177, No. 22 0021-9193/95/$04.00 0 Copyright 1995, American Society for Microbiology BvgAS Is Sufficient for Activation of the Bordetella pertussis
JOURNAL OF BACTERIOLOGY, Nov. 1995, p. 6477 6485 Vol. 177, No. 22 0021-9193/95/$04.00 0 Copyright 1995, American Society for Microbiology BvgAS Is Sufficient for Activation of the Bordetella pertussis
Field necropsy techniques in mammal and poultry
 Field necropsy techniques in mammal and poultry Kidsadagon Pringproa, DVM, MS, PhD Department of Veterinary Biosciences and Veterinary Public Health Faculty of Veterinary Medicine Chiang Mai University
Field necropsy techniques in mammal and poultry Kidsadagon Pringproa, DVM, MS, PhD Department of Veterinary Biosciences and Veterinary Public Health Faculty of Veterinary Medicine Chiang Mai University
BOVINE RESPIRATORY DISEASE COMPLEX. Kristen Mierzwiak LCS 630
 BOVINE RESPIRATORY DISEASE COMPLEX Kristen Mierzwiak LCS 630 Ring... You are called out to the farm of one of your regular dairy clients because some of the replacement heifers they bought at a public
BOVINE RESPIRATORY DISEASE COMPLEX Kristen Mierzwiak LCS 630 Ring... You are called out to the farm of one of your regular dairy clients because some of the replacement heifers they bought at a public
Evaluation of a computerized antimicrobial susceptibility system with bacteria isolated from animals
 J Vet Diagn Invest :164 168 (1998) Evaluation of a computerized antimicrobial susceptibility system with bacteria isolated from animals Susannah K. Hubert, Phouc Dinh Nguyen, Robert D. Walker Abstract.
J Vet Diagn Invest :164 168 (1998) Evaluation of a computerized antimicrobial susceptibility system with bacteria isolated from animals Susannah K. Hubert, Phouc Dinh Nguyen, Robert D. Walker Abstract.
The Disinfecting Effect of Electrolyzed Water Produced by GEN-X-3. Laboratory of Diagnostic Medicine, College of Medicine, Soonchunhyang University
 The Disinfecting Effect of Electrolyzed Water Produced by GEN-X-3 Laboratory of Diagnostic Medicine, College of Medicine, Soonchunhyang University Tae-yoon Choi ABSTRACT BACKGROUND: The use of disinfectants
The Disinfecting Effect of Electrolyzed Water Produced by GEN-X-3 Laboratory of Diagnostic Medicine, College of Medicine, Soonchunhyang University Tae-yoon Choi ABSTRACT BACKGROUND: The use of disinfectants
Dual Antibiotic Delivery from Chitosan Sponges Prevents In Vivo Polymicrobial Biofilm Infections
 Dual Antibiotic Delivery from Chitosan Sponges Prevents In Vivo Polymicrobial Biofilm Infections Ashley Parker, MS 1, James Smith, MS 1, Karen Beenken, PhD 2, Jessica Amber Jennings, PhD 3, Mark Smeltzer,
Dual Antibiotic Delivery from Chitosan Sponges Prevents In Vivo Polymicrobial Biofilm Infections Ashley Parker, MS 1, James Smith, MS 1, Karen Beenken, PhD 2, Jessica Amber Jennings, PhD 3, Mark Smeltzer,
Lecture 6: Fungi, antibiotics and bacterial infections. Outline Eukaryotes and Prokaryotes Viruses Bacteria Antibiotics Antibiotic resistance
 Lecture 6: Fungi, antibiotics and bacterial infections Outline Eukaryotes and Prokaryotes Viruses Bacteria Antibiotics Antibiotic resistance Lecture 1 2 3 Lecture Outline Section 4 Willow and aspirin Opium
Lecture 6: Fungi, antibiotics and bacterial infections Outline Eukaryotes and Prokaryotes Viruses Bacteria Antibiotics Antibiotic resistance Lecture 1 2 3 Lecture Outline Section 4 Willow and aspirin Opium
No-leaching. No-resistance. No-toxicity. >99.999% Introducing BIOGUARD. Best-in-class dressings for your infection control program
 Introducing BIOGUARD No-leaching. >99.999% No-resistance. No-toxicity. Just cost-efficient, broad-spectrum, rapid effectiveness you can rely on. Best-in-class dressings for your infection control program
Introducing BIOGUARD No-leaching. >99.999% No-resistance. No-toxicity. Just cost-efficient, broad-spectrum, rapid effectiveness you can rely on. Best-in-class dressings for your infection control program
MARBOCYL 10% SUMMARY OF PRODUCT CHARACTERISTICS
 MARBOCYL 10% SUMMARY OF PRODUCT CHARACTERISTICS 1. NAME OF THE VETERINARY MEDICINAL PRODUCT MARBOCYL 10%, solution for injection for cattle and swine 2. QUALITATIVE AND QUANTITATIVE COMPOSITION Marbofloxacin...100.0
MARBOCYL 10% SUMMARY OF PRODUCT CHARACTERISTICS 1. NAME OF THE VETERINARY MEDICINAL PRODUCT MARBOCYL 10%, solution for injection for cattle and swine 2. QUALITATIVE AND QUANTITATIVE COMPOSITION Marbofloxacin...100.0
THE COST OF COMPANIONSHIP
 THE COST OF COMPANIONSHIP Jared Gillingham and Robert Burlage Concordia University School of Pharmacy Mequon, WI Synopsis: Infectious diseases are always a concern, but when you are a person in an at-risk
THE COST OF COMPANIONSHIP Jared Gillingham and Robert Burlage Concordia University School of Pharmacy Mequon, WI Synopsis: Infectious diseases are always a concern, but when you are a person in an at-risk
SUMMARY OF PRODUCT CHARACTERISTICS
 SUMMARY OF PRODUCT CHARACTERISTICS 1. NAME OF THE VETERINARY MEDICINAL PRODUCT Tilmovet 250 mg/ml Concentrate for Oral Solution (BE, BG, CZ, EL, HU, IE, NL, PL, RO, UK) for pigs, chickens, turkeys and
SUMMARY OF PRODUCT CHARACTERISTICS 1. NAME OF THE VETERINARY MEDICINAL PRODUCT Tilmovet 250 mg/ml Concentrate for Oral Solution (BE, BG, CZ, EL, HU, IE, NL, PL, RO, UK) for pigs, chickens, turkeys and
Epigenetic regulation of Plasmodium falciparum clonally. variant gene expression during development in An. gambiae
 Epigenetic regulation of Plasmodium falciparum clonally variant gene expression during development in An. gambiae Elena Gómez-Díaz, Rakiswendé S. Yerbanga, Thierry Lefèvre, Anna Cohuet, M. Jordan Rowley,
Epigenetic regulation of Plasmodium falciparum clonally variant gene expression during development in An. gambiae Elena Gómez-Díaz, Rakiswendé S. Yerbanga, Thierry Lefèvre, Anna Cohuet, M. Jordan Rowley,
How to stop the snotty noses: Preventing feline upper respiratory infections. Staci Cannon, DVM, MPH, DACVPM, DABVP (Shelter Medicine Practice)
") How to stop the snotty noses: Preventing feline upper respiratory infections Staci Cannon, DVM, MPH, DACVPM, DABVP (Shelter Medicine Practice) Why is URI so hard to control? Multiple pathogens Chronic
How to stop the snotty noses: Preventing feline upper respiratory infections Staci Cannon, DVM, MPH, DACVPM, DABVP (Shelter Medicine Practice) Why is URI so hard to control? Multiple pathogens Chronic
Development of Resistant Bacteria Isolated from Dogs with Otitis Externa or Urinary Tract Infections after Exposure to Enrofloxacin In Vitro
 A. M. Brothers, P. S. Gibbs, and R. E. Wooley Development of Resistant Bacteria Isolated from Dogs with Otitis Externa or Urinary Tract Infections after Exposure to Enrofloxacin In Vitro Amy M. Brothers,
A. M. Brothers, P. S. Gibbs, and R. E. Wooley Development of Resistant Bacteria Isolated from Dogs with Otitis Externa or Urinary Tract Infections after Exposure to Enrofloxacin In Vitro Amy M. Brothers,
Presented at Central Veterinary Conference, Kansas City, MO, August 2013; Copyright 2013, P.L Ruegg, all rights reserved
 MILK MICROBIOLOGY: IMPROVING MICROBIOLOGICAL SERVICES FOR DAIRY FARMS Pamela L. Ruegg, DVM, MPVM, University of WI, Dept. of Dairy Science, Madison WI 53705 Introduction In spite of considerable progress
MILK MICROBIOLOGY: IMPROVING MICROBIOLOGICAL SERVICES FOR DAIRY FARMS Pamela L. Ruegg, DVM, MPVM, University of WI, Dept. of Dairy Science, Madison WI 53705 Introduction In spite of considerable progress
Alternatives to Antibiotics
 Alternatives to Antibiotics Cyril Gerard Gay, DVM, PhD Senior National Program Leader Animal Production and Protection Agricultural Research Service cyril.gay@ars.usda.gov Presentation Outline 1. Agricultural
Alternatives to Antibiotics Cyril Gerard Gay, DVM, PhD Senior National Program Leader Animal Production and Protection Agricultural Research Service cyril.gay@ars.usda.gov Presentation Outline 1. Agricultural
funded by Reducing antibiotics in pig farming
 funded by Reducing antibiotics in pig farming The widespread use of antibiotics (also known as antibacterials) in human and animal medicine increases the level of resistant bacteria. This makes it more
funded by Reducing antibiotics in pig farming The widespread use of antibiotics (also known as antibacterials) in human and animal medicine increases the level of resistant bacteria. This makes it more
Recommended for Implementation at Step 7 of the VICH Process on 15 December 2004 by the VICH Steering Committee
 VICH GL27 (ANTIMICROBIAL RESISTANCE: PRE-APPROVAL) December 2003 For implementation at Step 7 - Final GUIDANCE ON PRE-APPROVAL INFORMATION FOR REGISTRATION OF NEW VETERINARY MEDICINAL PRODUCTS FOR FOOD
VICH GL27 (ANTIMICROBIAL RESISTANCE: PRE-APPROVAL) December 2003 For implementation at Step 7 - Final GUIDANCE ON PRE-APPROVAL INFORMATION FOR REGISTRATION OF NEW VETERINARY MEDICINAL PRODUCTS FOR FOOD
Q1. (a) Clostridium difficile is a bacterium that is present in the gut of up to 3% of healthy adults and 66% of healthy infants.
 Clostridium difficile is a bacterium that is present in the gut of up to 3% of healthy adults and 66% of healthy infants.") Q1. (a) Clostridium difficile is a bacterium that is present in the gut of up to 3% of healthy adults and 66% of healthy infants. C. difficile rarely causes problems, either in healthy adults or in infants.
Q1. (a) Clostridium difficile is a bacterium that is present in the gut of up to 3% of healthy adults and 66% of healthy infants. C. difficile rarely causes problems, either in healthy adults or in infants.
towards a more responsible antibiotics use in asian animal production: supporting digestive health with essential oil compounds TECHNICAL PAPER
 TECHNICAL PAPER towards a more responsible antibiotics use in asian animal production: supporting digestive health with essential oil compounds www.provimi-asia.com Towards a more responsible use of antibiotics
TECHNICAL PAPER towards a more responsible antibiotics use in asian animal production: supporting digestive health with essential oil compounds www.provimi-asia.com Towards a more responsible use of antibiotics
SUMMARY OF PRODUCT CHARACTERISTICS. Lincomycin (as Lincomycin hydrochloride) Neomycin (as Neomycin sulphate) Excipients Disodium edetate
 Neomycin (as Neomycin sulphate) Excipients Disodium edetate") SUMMARY OF PRODUCT CHARACTERISTICS AN: 00221/2013 1. NAME OF THE VETERINARY MEDICINAL PRODUCT Lincocin Forte S Intramammary Solution 2. QUALITATIVE AND QUANTITATIVE COMPOSITION Active substances Lincomycin
SUMMARY OF PRODUCT CHARACTERISTICS AN: 00221/2013 1. NAME OF THE VETERINARY MEDICINAL PRODUCT Lincocin Forte S Intramammary Solution 2. QUALITATIVE AND QUANTITATIVE COMPOSITION Active substances Lincomycin
Lab Exercise: Antibiotics- Evaluation using Kirby Bauer method.
 Lab Exercise: Antibiotics- Evaluation using Kirby Bauer method. OBJECTIVES 1. Compare the antimicrobial capabilities of different antibiotics. 2. Compare effectiveness of with different types of bacteria.
Lab Exercise: Antibiotics- Evaluation using Kirby Bauer method. OBJECTIVES 1. Compare the antimicrobial capabilities of different antibiotics. 2. Compare effectiveness of with different types of bacteria.
BioSci 110, Fall 08 Exam 2
 1. is the cell division process that results in the production of a. mitosis; 2 gametes b. meiosis; 2 gametes c. meiosis; 2 somatic (body) cells d. mitosis; 4 somatic (body) cells e. *meiosis; 4 gametes
1. is the cell division process that results in the production of a. mitosis; 2 gametes b. meiosis; 2 gametes c. meiosis; 2 somatic (body) cells d. mitosis; 4 somatic (body) cells e. *meiosis; 4 gametes
Inhibiting Microbial Growth in vivo. CLS 212: Medical Microbiology Zeina Alkudmani
 Inhibiting Microbial Growth in vivo CLS 212: Medical Microbiology Zeina Alkudmani Chemotherapy Definitions The use of any chemical (drug) to treat any disease or condition. Chemotherapeutic Agent Any drug
Inhibiting Microbial Growth in vivo CLS 212: Medical Microbiology Zeina Alkudmani Chemotherapy Definitions The use of any chemical (drug) to treat any disease or condition. Chemotherapeutic Agent Any drug
SCIENTIFIC DISCUSSION
 SCIENTIFIC DISCUSSION I. SUMMARY OF THE DOSSIER Nobivac Bb is a lyophilised live vaccine containing Bordetella bronchiseptica strain B-C2. The lyophilised plug and the accompanying solvent for the vaccine
SCIENTIFIC DISCUSSION I. SUMMARY OF THE DOSSIER Nobivac Bb is a lyophilised live vaccine containing Bordetella bronchiseptica strain B-C2. The lyophilised plug and the accompanying solvent for the vaccine
EXCEDE Sterile Suspension
 VIAL LABEL MAIN PANEL PRESCRIPTION ANIMAL REMEDY KEEP OUT OF REACH OF CHILDREN READ SAFETY DIRECTIONS FOR ANIMAL TREATMENT ONLY EXCEDE Sterile Suspension 200 mg/ml CEFTIOFUR as Ceftiofur Crystalline Free
VIAL LABEL MAIN PANEL PRESCRIPTION ANIMAL REMEDY KEEP OUT OF REACH OF CHILDREN READ SAFETY DIRECTIONS FOR ANIMAL TREATMENT ONLY EXCEDE Sterile Suspension 200 mg/ml CEFTIOFUR as Ceftiofur Crystalline Free
ENVIRACOR J-5 aids in the control of clinical signs associated with Escherichia coli (E. coli) mastitis
 mastitis") GDR11136 ENVIRACOR J-5 aids in the control of clinical signs associated with Escherichia coli (E. coli) mastitis February 2012 Summary The challenge data presented in this technical bulletin was completed
GDR11136 ENVIRACOR J-5 aids in the control of clinical signs associated with Escherichia coli (E. coli) mastitis February 2012 Summary The challenge data presented in this technical bulletin was completed
SUMMARY OF PRODUCT CHARACTERISTICS
 SUMMARY OF PRODUCT CHARACTERISTICS 1. NAME OF THE VETERINARY MEDICINAL PRODUCT Marbocare 20 mg/ml solution for injection for cattle and pigs (UK, IE, FR) Odimar 20 mg/ml solution for injection for cattle
SUMMARY OF PRODUCT CHARACTERISTICS 1. NAME OF THE VETERINARY MEDICINAL PRODUCT Marbocare 20 mg/ml solution for injection for cattle and pigs (UK, IE, FR) Odimar 20 mg/ml solution for injection for cattle