ABSTRACT. hemisphere. Cytauxzoonosis is caused by the tick-transmitted parasite Cytauxzoon felis, an
|
|
|
- Karin Hudson
- 5 years ago
- Views:
Transcription
1 ABSTRACT SCHREEG, MEGAN ELIZABETH. Cytauxzoon felis in a Post-Genomic Era: Taxonomy, Diagnosis, Treatment, and Prevention. (Under the direction of Dr. Adam Birkenheuer, Chair, and Dr. Michael Levy, Vice Chair.) Cytauxzoonosis is an emerging disease affecting felines throughout the Western hemisphere. Cytauxzoonosis is caused by the tick-transmitted parasite Cytauxzoon felis, an organism for which little is known. Since being discovered 40 years ago in Missouri, C. felis has spread across 1/3 of the United States, and is expected to continue spreading given the continental distribution of competent feline and tick hosts. C. felis is highly pathogenic to domestic cats. Disease progresses rapidly and even with the best treatment mortality remains high. No vaccine exists, so disease prevention relies on indoor confinement and acaricide prophylaxis. Given the rapid dispersal of this virulent parasite that lacks effective treatment or prevention options, further investigation is warranted. However, the inability to culture the parasite in vitro, ethical concerns with in vivo studies, and lack of funding for feline diseases has limited C. felis research. To counteract this, we have sequenced the parasite s mitochondrial and chromosomal genomes. Using these resources, we have identified useful genetic targets that resolve the taxonomy of C. felis and improve the treatment, diagnosis and prevention of cytauxzoonosis. The taxonomic placement of Cytauxzoon felis within Piroplasmida remains unsolved due to discrepancies between morphological and molecular data. We have clarified phylogeny of C. felis and other Piroplasmida using mitochondrial genome sequences and structures. Mitochondrial genome analysis supported the placement of C. felis within the Theileria clade, and indicated that T. equi, B. conradae, and B. microti organisms are
2 genetically distinct lineages. Characterization of additional mitochondrial genomes and subsequent reclassification of Piroplasmida genera are merited. Mortality of infected cats treated with atovaquone and azithromycin (A&A) is 40%. Atovaquone targets an electron transport protein encoded by mitochondrial cytb. Mutations in the cytb gene are associated with atovaquone resistance in related parasites. We hypothesized that C. felis cytb genotype would be associated with response to A&A treatment. After cytb-genotyping 69 samples, we identified a C. felis cytb genotype (cytb1) associated with increased survival of cats treated with A&A. Given this association, we hypothesized that cytb1 could aid in the prognosis of cytauxzoonosis. By developing a quantitative PCR panel that identifies cytb1-specific SNPs with high resolution melt (HRM) analysis, we distinguished C. felis cytb1 from all other C. felis cytb genotypes with 100% sensitivity and 98.2% specificity. This assay is cost-effective and can be completed in less than 3 hours, which is important given the high cost of A&A and rapid disease course. Diagnosis of C. felis is challenging during early infection when parasitemia is low and clinical signs remain vague. Mitochondrial genes are more sensitive molecular diagnostic targets for parasite detection than 18S in related parasites. We demonstrated that mitochondrial cox3 copy number is increased relative to 18S in blood and tissue samples from cats with acute cytauxzoonosis, and that cox3 is more sensitive for identifying early C. felis infection than 18S. This assay will aid in early detection of Cytauxzoon felis infection. Sequencing the C. felis genome has allowed for identification of 33 vaccine candidates. We have manufactured two DNA vaccines: an expression library vaccine including all candidates and a vaccine consisting of the recently described cf76. We describe
3 the immunization approach, serological response to vaccination, and subsequent clinical outcome following challenge with C. felis. All vaccinated cats became infected and developed disease, rendering both vaccines inadequate methods for prevention of cytauxzoonosis. However, the expression library vaccine showed evidence of aiding in disease control, providing a baseline for development of future subunit vaccines against cytauxzoonosis. In conclusion, the mitochondrial and chromosomal genomes have clarified the taxonomy of Cytauxzoon felis and further advanced our understanding of the treatment, diagnosis, and prevention of cytauxzoonosis.
4 Copyright 2015 by Megan Elizabeth Schreeg All Rights Reserved
5 Cytauxzoon felis in a Post-Genomic Era: Taxonomy, Diagnosis, Treatment, and Prevention by Megan Elizabeth Schreeg A dissertation submitted to the Graduate Faculty of North Carolina State University in partial fulfillment of the requirements for the Degree of Doctor of Philosophy Comparative Biomedical Sciences Raleigh, North Carolina 2015 APPROVED BY: Adam Birkenheuer Chair of Advisory Committee Michael Levy Vice Chair of Advisory Committee David Bird Luke Borst Jeffrey Yoder Committee Member Committee Member Committee Member

6 BIOGRAPHY Megan was born and raised in Kokomo, Indiana, where she was constantly surrounded by cats and consequently was inspired to become a veterinarian. Megan graduated summa cum laude with a Bachelor of Arts Degree in Biology and minors in Biochemistry and Latin from Hanover College in Hanover, Indiana in Megan is currently in the Combined DVM/PhD Program at North Carolina State University College of Veterinary Medicine. Megan has interests in feline medicine, immunology, comparative anatomy and Photo credit to Wendy Savage pathology, comparative molecular genomics, and parasitology. Following completion from the DVM/PhD program, Megan currently intends to apply for a residency in either anatomic or clinical pathology, and also hopes to pursue board certification as a veterinary parasitologist. Megan also has a passion for teaching, and her ultimate goal is to become a professor at a school of veterinary medicine. ii
7 ACKNOWLEDGEMENTS A researcher is only as good as her support system, and I ve been blessed enough to have a great team of lab mates, friends, and family helping me through every step of completing this work. First, I would like to thank my advisor Adam, who has mentored me not only in research, but also in life. None of this work would be possible without your passion for companion animal medicine, love for learning, or aptitude for training developing clinician scientists. The countless hours you have spent helping me both inside the lab and out have meant so much to me, and taught me what it truly means to be a mentor. You have taken a naive student and molded her into a slightly less naïve researcher and human being, and for that I cannot thank you enough! Next I would like to thank Henry, our laboratory technician, for teaching me the ropes of working at the bench, for helping me with my work and always patiently answering my often stupid questions, and for being a great friend. Together you and Adam have made me feel like part of a little lab family, which meant the world to a girl who left everything she knew behind and moved across the country to pursue an education. There are endless other mentors and supporters I have had in the world of science. First, a huge thank you to Dr. Sam Jones for accepting me in the DVM/PhD program you believed in my potential when I didn t, and have offered unending support throughout the program! A special thank you to my undergraduate advisor, Dr. Walter Bruyninckx, who showed me that a career as a researcher, veterinarian, teacher, and life-long learner was possible. An additional thank you to Dr. Leah Cohn, who I consider not only a key collaborator, but also a phenomenal clinician, a friend, and one of the most kind-hearted iii
8 people I know. Thanks to Dr. Brian Wiegmann for patiently teaching me about molecular systematics! A huge thanks as well to my committee members: Dr. Levy, Dr. Bird, Dr. Borst, and Dr. Yoder your support and collaboration have been critical for me on this journey. Last but certainly not least, I also would like to thank the rest of my office and lab members, including Kaye, Candace, Mitsu, Jingjing, Jaime, the folks of the VBDDL, and all the others that I am forgetting. You are like family to me and make coming to work a joy! A huge thanks as well to all of my friends both near and far, who have been with me through high school, college, and beyond. I would be lost without the Great 8 and my Hanover ladies, who have been some of my biggest cheerleaders in the pursuit of this degree. A special thanks to the newly minted Dr. Susan Grayden Shapiro, my DVM/PhD partner in crime I wouldn t have had the opportunity to do any of this work if you hadn t told me about the amazing veterinary school at NC State, and I can t wait to see how our destinies continue to cross in the future. My family deserves infinite praise for endlessly supporting and loving me. To Mom and Dad you imparted a love for problem-solving, nature, and animals in me from a very young age. You always nurtured my passions through the years, whether that meant getting me yet another cat, helping me tend to my science fair project, or pitching batting practice to me. You taught me the meaning of hard work and persistence, and instilled in me that anything was possible if I put my mind to it. I couldn t ask for better parents or role models, and I m forever grateful and proud to be your daughter. To my brothers Danny and Keagan whether you know it or not, you both have inspired me in your own way, and I wouldn t trade you for any other siblings on the planet! iv
9 More than any other person, I need to thank my husband Jacob. You are my anchor in all things I do! You moved across the country to come with me on this journey, and for some crazy reason along the way decided that I was worthy to be your wife. You have supported me every step of the way, whether that mean helping me study, coming into the lab with me, listening to me blather about my latest silly discovery, or taking care of our house and our animal family Murray, Pip, and Rue while I focused on school. You have the extraordinary ability to calm my nerves when I am discouraged, and your encouragement drives me to succeed you inspire me in all I do! I would never have been able to complete this work without your endless devotion and self-sacrifice. I love you more than you will ever know, and I m so happy that I get to spend eternity by your side as your wife and best friend. Most importantly, I would like to thank all of the cats, pet owners, and clinicians who have contributed to this work, particularly the cats who have succumbed to cytauxzoonosis. I know that this work can in no way undo the grief and suffering that this disease causes for cats and humans alike, but every sacrifice you have made gets us one step closer to helping more cats and ultimately winning the battle against Cytauxzoon felis. v
10 TABLE OF CONTENTS LIST OF TABLES... ix LIST OF FIGURES... xi CHAPTER 1: Literature Review...1 INTRODUCTION...1 TAXONOMY OF CYTAUXZOON FELIS...4 DIAGNOSIS OF CYTAUXZOON FELIS INFECTION AND COMMENTARY ON MITOCHONDRIAL BIOLOGY...10 TREATMENT AND PROGNOSIS OF CYTAUXZOONOSIS...14 PREVENTION OF CYTAUXZOONOSIS...18 REFERENCES...25 CHAPTER 2: Mitochondrial genome sequences and structures aid in the resolution of Piroplasmida phylogeny...40 ABSTRACT...40 INTRODUCTION...41 MATERIALS AND METHODS...49 RESULTS...56 DISCUSSION...64 ACKNOWLEDGEMENTS...72 REFERENCES...72 SUPPLEMENTAL MATERIAL...80 CHAPTER 3: Pharmacogenomics of Cytauxzoon felis cytochrome b: Implications for atovaquone and azithromycin therapy in domestic cats with cytauxzoonosis...95 vi
11 ABSTRACT...95 TEXT...95 ACKNOWLEDGEMENTS REFERENCES SUPPLEMENTAL MATERIAL CHAPTER 4: Rapid high resolution melt analysis of Cytauxzoon felis cytochrome b to aid in the prognosis of cytauxzoonosis ABSTRACT INTRODUCTION MATERIALS AND METHODS RESULTS DISCUSSION ACKNOWLEDGEMENTS REFERENCES SUPPLEMENTAL MATERIAL CHAPTER 5: PCR amplification of a multi-copy mitochondrial gene (cox3) for early detection of Cytauxzoon felis infection ABSTRACT INTRODUCTION MATERIALS AND METHODS RESULTS DISCUSSION ACKNOWLEDGEMENTS vii
12 References CHAPTER 6: DNA vaccination of domestic cats against Cytauxzoon felis: Approach, outcome, and future directions ABSTRACT INTRODUCTION MATERIALS AND METHODS RESULTS DISCUSSION ACKNOWLEDGEMENTS REFERENCES SUPPLEMENTAL MATERIAL CHAPTER 7: Conclusions APPENDIX CLINICAL DATA FOR CATS IN CYTAUXZOON FELIS DNA VACCINATION PILOT STUDY viii
13 LIST OF TABLES CHAPTER 2: Mitochondrial genome sequences and structures aid in the resolution of Piroplasmida phylogeny Table 1. Species and sequences utilized in phylogenetic analysis...50 Table 2. Primers utilized in PCR amplification of Piroplasmida mitochondrial genomes...51 CHAPTER 3: Pharmacogenomics of Cytauxzoon felis cytochrome b: Implications for atovaquone and azithromycin therapy in domestic cats with cytauxzoonosis Table 1. Correlation between survival rate of cats treated with A&A and C. felis cytb genotype CHAPTER 4: Rapid high resolution melt analysis of Cytauxzoon felis cytochrome b to aid in the prognosis of cytauxzoonosis Table 1. Nucleotide positions evaluated for the identification of cytb Table 2. Predicted and actual specificity of the PCR panel Table 3. Primers sequences for amplification of five SNP regions analyzed by HRM and full length C. felis cytb Table 4. Positive predictive value for identification of C. felis cytb1 is highest when all five nucleotide positions in the PCR panel are analyzed CHAPTER 5: PCR amplification of a multi-copy mitochondrial gene (cox3) for early detection of Cytauxzoon felis infection ix
14 Table 1. cox3 PCR detects C. felis with increased or equal sensitivity as 18S PCR for all samples CHAPTER 6: DNA vaccination of domestic cats against Cytauxzoon felis: Approach, outcome, and future directions Table 1. Summary of notable physical observations, laboratory findings, and clinical outcomes for individual cats x
15 LIST OF FIGURES CHAPTER 1: Literature Review Figure 1. Occlusion of pulmonary vessel with C. felis schizont-infected leukocytes..1 Figure 2. Geographic distribution of Cytauxzoon felis...3 Figure 3. Cytauxzoon felis schizonts appear to infect monocytes...4 Figure 4. Figure 5. Apicomplexan mitochondrial genome structural diversity...9 Identification of Cytauxzoon felis piroplasms and schizonts by light microscopy...11 Figure 6. Atovaquone inhibits electron transport within the parasite mitochondria...16 Figure 7. Identification of Cytauxzoon felis vaccine candidates...22 Figure 8. Conceptual depiction of expression library immunization against Cytauxzoon felis...24 CHAPTER 2: Mitochondrial genome sequences and structures aid in the resolution of Piroplasmida phylogeny Figure 1. 18S sequence alone is unable to resolve phylogeny of the Piroplasmida...43 Figure 2. Mitochondrial genome structures of Piroplasmida species characterized in this study...57 Figure 3. Phylogenetic analysis of concatenated mitochondrial genome and 18S nucleotide sequence identifies five distinct lineages within Piroplasmida.59 xi
16 Figure 4. Mitochondrial genome structures further support recognition of the five groups identified by phylogenetic analysis of concatenated mitochondrial and 18S sequences...63 Figure 5. Biology of Piroplasmida organisms is consistent with phylogeny inferred from analysis of concatenated mitochondrial and 18S sequences...65 Figure 6. Phylogenetic analysis of COX1 amino acid sequence recovers the same five Piroplasmida groups as concatenated mitochondrial and 18S nucleotide sequences...68 CHAPTER 3: Pharmacogenomics of Cytauxzoon felis cytochrome b: Implications for atovaquone and azithromycin therapy in domestic cats with cytauxzoonosis Figure 1. PCR amplification of C. felis cytb gene in three overlapping fragments..97 Figure 2. Presence of secondary peaks in C. felis cytb sequence as determined by Vector NTI...98 Figure 3. Characterization of 30 novel C. felis cytb genotypes...99 Figure 4. Evidence of missense mutations in or near putative atovaquone-binding sites of C. felis CYTB CHAPTER 4: Rapid high resolution melt analysis of Cytauxzoon felis cytochrome b to aid in the prognosis of cytauxzoonosis Figure 1. Identification of C. felis cytb1 by HRM analysis of five cytb nucleotides 117 xii
17 Figure 2. HRM analysis failed to detect relatively high A750G heteroplasmy yet was able to detect low G750A heteroplasmy at C. felis cytb nucleotide 750 in clinical samples Figure 3. Analysis of C. felis cytb clone mixtures confirms inability of HRM analysis to consistently discriminate heterogeneity in complex mixtures of genotypes CHAPTER 5: PCR amplification of a multi-copy mitochondrial gene (cox3) for early detection of Cytauxzoon felis infection Figure 1. Cytauxzoon felis mitochondrial genome copy number is increased in the blood of cats infected for less than 6 months but not cats infected for over a year Figure 2. In acutely infected cats, Cytauxzoon felis mitochondrial genome copy number in blood samples is equal to or higher than that in tissue samples Figure 3. Cox3 is more sensitive than 18S at detecting Cytauxzoon felis infection at 7 and 9 DPI Figure 4. Cytauxzoon felis piroplasms are highly pleiomorphic during acute infection CHAPTER 6: DNA vaccination of domestic cats against Cytauxzoon felis: Approach, outcome, and future directions Figure 1. Cytauxzoon felis DNA vaccination pilot study design xiii
18 Figure 2. Timeline of vaccination, infection, and sample collection for vaccinated cats Figure 3. Cats vaccinated with CF-Library have a higher survival rate (100%) than other infected cats Figure 4. Serological responses of individual vaccinated cats to candidates at different time points in study xiv
.")
19 CHAPTER 1: Literature Review INTRODUCTION Cytauxzoonosis is an emerging infectious disease that affects domestic and wild felids throughout North and South America. The causative agent of cytauxzoonosis is Cytauxzoon felis, a tick-transmitted hemoprotozoan parasite in the order Piroplasmida. C. felis can be extremely pathogenic to domestic cats, and the clinical disease caused by acute infection is typified by high morbidity and mortality (1, 2). Cats with clinical disease initially show vague clinical signs of lethargy, pyrexia, and anorexia (3). Without treatment, the disease rapidly progresses and within 2-5 days culminates in systemic inflammatory response syndrome, disseminated intravascular coagulation, ischemia, and multi-organ failure (3, 4). Much of the disease pathology is caused by the accumulation of parasiteinfected leukocytes that occlude the vasculature (Figure 1). The vast majority of cats presenting to veterinary hospitals succumb to the disease, and without antiprotozoal therapy, survival rates of affected cats are as low as 0.2-3% (2, 5). Figure 1. Occlusion of pulmonary vessel with C. felis schizont-infected leukocytes. Photo credit to Dr. Luke Borst 1
20 However, advances in treatment have greatly improved survival of cats with cytauxzoonosis. In a prospective randomized clinical trial, cats receiving atovaquone and azithromycin (A&A) had a significantly higher survival rate (60%) compared to cats treated with imidocarb dipropionate (26%), which was once considered the treatment of choice for acute cytauxzoonosis (3, 6, 7). While the advent of A&A treatment for cytauxzoonosis greatly increases a cat s chance at survival, this therapeutic approach still has limitations. Morbidity and mortality rates (40%) with A&A treatment still remains relatively high. Furthermore, A&A in conjunction with supportive care can cost thousands of dollars, which can prevent therapy from being a feasible option for many pet owners. Given the shortcomings of treating acute cytauxzoonosis, prevention of infection and/or clinical disease is the ideal method of control. However, no vaccine is currently available, so prevention is completely dependent on application of the appropriate prophylactic acaricides and keeping cats indoors (1, 8). In addition to being highly pathogenic and difficult to combat, Cytauxzoon felis is also undergoing a rapid geographic dissemination and evolution in epidemiology. Since its discovery in domestic cats in Missouri in 1976, C. felis has been recognized throughout the central and southeastern United States, and to date has been detected in domestic cats in 17 different states and in bobcats, the natural host, in at least two additional states (Figure 2, 9-12). Although domestic cats were originally considered an aberrant host, over the past twenty years a number cats have survived C. felis infection without antiprotozoal therapy and/or evidence of clinical disease (13-17). In some regions highly endemic for the disease as many as 30% of cats may be infected and subsequently serving as novel reservoirs for the spread of infection (17). The reason behind this change in disease epidemiology is unclear. 2
21 However, the growing population of subclinically infected cats combined with widespread bobcat reservoirs and competent tick vectors (Amblyomma americanum and Dermacentor variabilis) is likely to enhance the spread of C. felis infection in domestic cats throughout North and South America. The rapid spread and evolving epidemiology of this highly virulent parasite that lacks effective treatment or prevention options creates a pressing health concern for feline species in the Western hemisphere. However, in spite of this threat, a number of gaps still remain in our understanding of both the biology of Cytauxzoon felis as well as the clinical approach to overcoming the disease it causes. Due to the lack of funding for companion animal diseases, ethical concerns of extensive in vivo studies, and the inability to culture the parasite in vitro, our lab has utilized the Cytauxzoon felis genome as a tool to further study the parasite. Through the use of both the mitochondrial and chromosomal genome, this thesis addresses a number of gaps in the Cytauxzoon felis knowledge base, including the parasite s taxonomy as well as diagnosis, treatment, and prevention of cytauxzoonosis. Figure 2. Distribution of Cytauxzoon felis in the United States. States where C. felis has been detected in domestic cats (yellow), bobcats only (orange), or has yet to be detected (green) are indicated. 3
.")
22 TAXONOMY OF CYTAUXZOON FELIS The genus Cytauxzoon is taxonomically categorized within the order Piroplasmida, a collection of blood-borne protozoan parasites that also includes the genera Babesia and Theileria. Cytauxzoon was first described as a genus in 1948, when infection with a Theileria-like parasite was determined to be the cause of death in a duiker in South Africa (18). Cytauxzoon was differentiated from Theileria due to the presence of schizonts in monocytes as opposed to lymphocytes (Figure 3), although intraerythrocytic forms of the two genera were indistinguishable (18, 19). In the following years, similar Cytauxzoon species were identified in other African ruminants, including kudu, elands, and giraffes (20-22). However, some parasitologists opposed recognizing Cytauxzoon as a separate genus based on the criteria of schizont host cell preference alone (20). This opinion was Figure 3. Cytauxzoon felis schizonts appear to infect monocytes. A) Leukocytes that become infected with a C. felis schizont (S) have similar morphology to uninfected monocytes (indicated with arrow heads), but are easily distinguished by their large size, prominent nucleolus, increased cytoplasmic basophilia, and magenta-colored parasites throughout cytoplasm. B) Morphology of C. felis schizont-infected leukocytes (S) is more consistent with that of monocytes than that of neutrophils (N) or lymphocytes (L). However, the true identity of these cells has yet to be confirmed by molecular techniques (e.g. immunophenotyping). 4
23 strengthened when it was discovered that some so-called Cytauxzoon species as well as Theileria species could infect both lymphocytes and monocytes (23-25). Consequently, all ruminant Cytauxzoon species that could be reproducibly identified were reclassified as Theileria and to date are recognized as such (23, 26-30). Cytauxzoon felis was first described in 1976 during the midst of the debate over the legitimacy of the genus Cytauxzoon (31). C. felis was classified as such due to the development of schizonts in what appeared to be monocytic cells (Figure 3). However, despite the reclassification of the ruminant Cytauxzoon species as Theileria, the same has yet to be done for C. felis regardless of suggestions to do so (29). The advent of DNA sequencing has further muddled the taxonomy of C. felis as well as the genus Cytauxzoon. Phylogenetic analyses of the Piroplasmida using 18S rrna gene sequence have failed to pinpoint a definitive taxonomic placement for Cytauxzoon felis (32-36). Depending on the study and varying methods used, Cytauxzoon has been categorized as a sister group or ancestral to Theileria, and has occasionally been grouped with Theileria equi, another species whose taxonomy is strongly debated (26, 32-39). For decades, C. felis was the only organism recognized in the genus. More recently, a number of novel piroplasms have been identified that share >95% of 18S gene sequence with Cytauxzoon felis, and as such have been categorized within the genus (40-43). However, with the exception of Cytauxzoon felis, intraleukocytic schizonts have not been identified for any of these species (40-43). Thus, the defining characteristics of parasites classified as Cytauxzoon as well as its legitimacy as a genus remain in question. Defining the taxonomic placement of Cytauxzoon felis based only on the presumed cell type infected and/or the sequence of a single gene is a limited approach that fails to 5
24 consider other observations about the parasite. First, while intraerythrocytic forms cause disease for the majority Piroplasmida, the pathogenesis of cytauxzoonosis is caused by infection of leukocytes by schizonts. This is also the case for East Coast Fever and tropical theileriosis, which are caused by Theileria parva and Theileria annulata, respectively (44-47). Additionally, the grossly enlarged C. felis schizont-infected cells, which can reach 250 µm in diameter, bear a striking resemblance to schizont-infected cells of Theileria orientalis (48, 49). These mutual characteristics suggest a close relationship between Cytauxzoon felis and Theileria organisms. Furthermore, whole genome sequence indicates a close relationship between C. felis and T. parva, as the two organisms genomes have a similar size and structure and are highly syntenic (50). Last, it should be noted that C. felis schizontinfected cells have been identified as being monocytic solely on the basis of morphology and the presence of lysozyme, an enzyme commonly produced by both macrophages and neutrophils (31, 51). T. parva and T. annulata actually have been shown to hijack host cellular processes and transform host cells into a neoplastic-like state (44); if the same is true for C. felis, definitive phenotyping of the infected cell would likely require additional techniques (e.g. immunophenotyping) beyond morphological descriptions. Therefore, the cell type infected by C. felis remains uncertain, making its categorization within Cytauxzoon debatable even if Cytauxzoon is to be considered a separate genus at all. Due to this uncertainty in cell type infected, schizont-infected cells will be referred to as leukocytes throughout this thesis. Collectively, these physiological and molecular characteristics of C. felis indicate a close relationship with Theileria species and question its classification in a separate genus. 6
25 Unfortunately, Cytauxzoon felis is not the only Piroplasmida species with unresolved taxonomy. DNA sequence has revealed that the closely related genus Babesia is actually polyphyletic, and is comprised of those organisms meeting the traditional definition of Babesia (Babesia sensu stricto) as well as a collection of organisms that morphologically resemble Babesia but appear to be genetically ancestral to Babesia, Theileria, and Cytauxzoon species (Babesia sensu latu; 32, 33, 35, 52-56). However, morphological features thought to be characteristic of primitive Piroplasmida (infection of nucleated host cells) have not been identified for a number of Babesia sensu latu species (55, 56). Hence, it remains unclear whether this trait has simply not been discovered for these species or if it has been lost/gained multiple times throughout evolutionary history of the Piroplasmida. In addition to the obvious polyphyly of Babesia, a number of other species have unsolved taxonomy, including species proposed to be closely related to C. felis. One such species is Theileria equi, which was originally named Babesia equi but has since been shown to infect both lymphocytes and monocytes (38, 57). However, conflicting molecular analyses have categorized T. equi within, as a sister group to, or as ancestral to Theileria within Babesia sensu latu (26, 32-36, 38, 39). Thus, it is apparent that the phylogeny of the Piroplasmida needs to be reexamined to understand the taxonomic status of not only C. felis but also many other related organisms. The majority of Piroplasmida molecular phylogenetic analyses have relied on 18S sequence alone. However analysis of a single gene is a limited approach for estimating evolutionary relationships, and discrepancies in 18S alignment and analysis methods across studies have failed to conclusively resolve Piroplasmida phylogeny (26, 32-37, 58). Thus, alternative approaches are clearly needed for conducting molecular phylogenetic analyses of 7
26 these organisms. While comparisons of organisms whole genome sequence would be ideal and has been conducted for a limited number of Piroplasmida species (39, 59), this molecular dataset is not available for the majority of characterized Piroplasmida. Furthermore, whole genome sequence analysis may not be a practical taxonomic approach given the high number of emerging Piroplasmida. Therefore, new targets for molecular phylogenetic analysis of Piroplasmida should consist of relatively short sequences that are conserved enough to allow for ease of amplification, yet different enough to be useful for phylogenetic inference. For Piroplasmida, mitochondrial genomes fit these criteria. Apicomplexan parasites, including Piroplasmida, possess the smallest characterized mitochondrial genomes (~6-11 kb) of all eukaryotes (60-62). The genes encoded on the linear mitochondrial genome are conserved across apicomplexan species, and include fragmented large and small subunit rrna genes as well as three components of the electron transport chain: cytochrome c oxidase subunit I (cox1), cytochrome c oxidase subunit III (cox3), and cytochrome b (cytb; 60-62). Interestingly, the mitochondrial rrna sequences are extensively fragmented and scattered throughout the mitochondrial genome (60, 63). When combining different rrna fragmentation patterns with various arrangements of cox1, cox3, and cytb, a wide range of mitochondrial genome structures have emerged for apicomplexans (Figure 4; 60, 61, 64-67). These structural differences are valuable for deducing relationships between parasites, as each complex structure is highly unlikely to have evolved more than once (60, 68). In addition to the taxonomic value of mitochondrial gene order, the sequences of mitochondrial genes have also proven to be useful for inferring phylogenetic relationships (69-71). The high mutation rate of the mitochondrial genome makes its sequence particularly useful in determining relationships between recently diverged 8

 sequences (A-C), or in the case of B.")
27 species (69, 70), a fact that has been demonstrated through analysis of multiple protozoan organisms (67, 72-76). Figure 4. Apicomplexan mitochondrial genomes structures are diverse yet conserved between closely related species. Piroplasmida species (A-D) have linear mitochondrial genomes that exist as single monomeric DNA molecules. Mitochondrial genomes are either flanked with terminal inverted repeated (TIR) sequences (A-C), or in the case of B. microti complex species (D) include inverted repeats (IR-A, IR-B) that allow for recombination or flip-flopping of mitochondrial gene arrangement. Babesia sensu stricto and most Theileria species (A) have identical arrangement of mitochondrial genes. B) In contrast, the mitochondrial genome of Theileria orientalis has a ~3 kb inversion (borders indicated by red hash marks). Theileria equi has a unique mitochondrial genome structure that includes a cox3-like gene embedded within TIRs. In contrast to Piroplasmida, Haemosporida (E) and Coccidia (F) species have mitochondrial genomes that are tandemly repeated on a linear DNA molecule. Protein-coding genes (white) and large (light gray) and small (dark gray) rrna fragments that have been reported to GenBank are indicated. 9
28 The relative ease of sequence acquisition as well as their proven taxonomic value make mitochondrial genomes excellent candidates to aid in the resolution of Piroplasmida phylogeny, including solving the taxonomy of Cytauxzoon felis. This topic will be addressed in Chapter 2 of this thesis. DIAGNOSIS OF CYTAUXZOON FELIS INFECTION AND COMMENTARY ON MITOCHONDRIAL BIOLOGY Because of the high mortality, expense of treatment, and rapid progression of cytauxzoonosis, quick and accurate identification of C. felis is critical for the clinical decision-making progress. Unfortunately, diagnosis of the disease can be difficult given the vague clinical signs that cats initially present with, including lethargy, depression, anorexia, and pyrexia (1, 3, 77). Depending on how advanced disease is at the time of presentation, cats may be dehydrated, icteric, dyspneic, hypothermic, and have localized or diffuse lymphadenopathy (1, 3, 77). Hematological abnormalities often include leukopenia, thrombocytopenia, and non-regenerative anemia (1, 77). Clinical chemistry values are variable from case to case, but hyperbilirubinemia is common finding (1, 3, 77). While these symptoms and laboratory values are suggestive of cytauxzoonosis, identification of the parasite itself is required to make a definitive diagnosis. Schizontinfected leukocytes are best visualized by fine needle aspirate of affected organs (spleen, liver, or lymph node) and can occasionally be seen on the feathered edge of peripheral blood smears (Figure 5; 1, 5, 77, 78). Intraerythrocytic piroplasms can also be identified on a peripheral blood smear (Figure 5). However, during early infection when parasitemia remains low, this technique has a poor sensitivity, with a previous study reporting that less 10
 but is not commercially available as a diagnostic test. Additionally, it is unlikely that this would be a sensitive method of C.")
29 than 50% of infected cats had detectable parasites at death (2, 5, 77, 78). An indirect fluorescent antibody test for the detection of a humoral response against C. felis has been described (79) but is not commercially available as a diagnostic test. Additionally, it is unlikely that this would be a sensitive method of C. felis detection during the course of clinical illness, as titers often remain undetectable until post-mortem serum is tested (79). Figure 5. Identification of Cytauxzoon felis piroplasms and schizonts by light microscopy. A) Intraerythrocytic piroplasms can be visualized as the disease progresses. Organisms traditionally are circular with a nucleus identifiable at the periphery of light blue cytoplasm ( signet rings, arrowheads), but can take on a variety of other morphologies (*) and may be seen dividing (#). Multiple organisms (arrows) may be present in a single erythrocyte. Organisms can be differentiated from Howell-Jolly bodies (H) by the presence of cytoplasm. B) If present, schizont-infected cells (arrowheads) are often easily identified on peripheral blood smears at low magnification due to their large size and granular, basophilic cytoplasm. C) Suspected schizont-infected cells (S) visualized on the feathered edge of blood smears should be examined at high magnification to distinguish from platelet clumps. At this magnification, intracytoplasmic parasites and host cell nucleoli are prominent. D) Schizont-infected cells (S) are more prevalent in tissues (such as the spleen seen in this image) than in blood; this often makes fine needle aspiration of tissues a more sensitive early diagnostic tool than blood smear examination. 11
30 Alternatively, amplification of parasite DNA by polymerase chain reaction (PCR) has proven to be a diagnostic technique that is more sensitive than light microscopy for detection of hemoprotozoan parasites (80-85). Various PCR assays developed for the diagnosis of C. felis have been able to detect as low as 1 copy of parasite DNA target/reaction (78) and have identified infection in cats where parasites were undetected by light microscopy (80). Currently, parasite genes encoding ribosomal RNA, including 18S and ITS (internal transcribed spacer), are the primary diagnostic targets utilized for detection of C. felis in clinical samples (78, 80). However, it has been demonstrated for related apicomplexan parasites (Babesia, Theileria, and Plasmodium) that genes encoded on the mitochondrial genome are more sensitive PCR targets than ribosomal RNA genes (86-89). Mitochondrial genes are likely more sensitive diagnostic targets due to the increased copy number of the mitochondrial genome relative to parasite chromosomal genomes. Related apicomplexans (Babesia microti, Babesia rodhaini, Plasmodium, Eimeria) have anywhere from mitochondrial genome copies per non-dividing haploid organism (64, 66, 90-93). Mitochondrial genome copy number also can vary depending on the life stage of the parasite. The number of both the mitochondrial genome and mitochondria themselves are often increased in highly active life stages (trophozoites, gametocytes, and schizonts), which likely reflects the higher metabolic demand required of these forms (94-103). Unfortunately, none of this information is available for C. felis, as the mitochondrial genome sequence and copy number remain uncharacterized for all life stages. Furthermore, very little is known about the biology of C. felis mitochondria at all, which is surprising given that the parasite mitochondria is targeted by the only therapy demonstrated to have efficacy in treating 12
31 cytauxzoonosis (atovaquone and azithromycin; see Treatment and Prognosis of Cytauxzoonosis ). Reported information regarding C. felis mitochondria is limited to ultrastructural observations alone. C. felis mitochondria have been described as being acristate or nonplicated, a feature of some apicomplexan mitochondria that original was thought to indicate lack of functionality (46, 104, 105). However, the efficacy of A&A against C. felis proves otherwise. Furthermore, while most reported electron microscopy images of organisms show a single mitochondrion per cell, one study showed an intraerythrocytic piroplasm with two mitochondria (46). At the very least, this implies that multiple mitochondrial genome copies may exist in a single organism, even if each mitochondria only contains a single mitochondrial genome copy. While little is known about the mitochondrial biology and genome of C. felis, this topic has been studied more in depth for related parasites, particularly Plasmodium species. It is generally accepted that a single mitochondrion containing multiple genome copies is present in the Plasmodium merozoite, which is the asexual parasite stage that initially infects erythrocytes ( ). However, as merozoites develop into trophozoites and eventually schizonts, parasite DNA synthesis begins, which includes replication of the mitochondria and mitochondrial genome (91, 93, 109). Replication of the concatenated Plasmodium mitochondrial genome occurs in a unique recombination-dependent fashion that resembles that of Phage T4 (91, 93, 109), although it is unclear if the same is true for the monomeric Piroplasmida mitochondrial genomes (110). This extensive recombination as well as the inherently higher rate of sequence mutations results in a highly polymorphic mitochondrial genome (108, ). Furthermore, apicomplexan mitochondria are inherited at random 13
32 from the female parent (the macrogamete) as in other eukaryotes (114). Given the fact that multiple genomes will be contained within the inherited mitochondria, it is likely that each organism inherits a mixture of different mitochondrial genome sequences, a condition known at heteroplasmy (115, 116). However, there has been little investigation into the level of mitochondrial heteroplasmy present in apicomplexan species. Currently, no information regarding the mitochondrial genome of Cytauxzoon felis is available. However, if there are any similarities with related organisms, it is likely that 1) multiple copies of the C. felis mitochondrial genome exist in an organism and 2) that mitochondrial genes will be sensitive targets for the molecular detection of C. felis in clinical samples. This topic will be addressed in Chapter 5 of this thesis. Furthermore, although the inability to culture C. felis in vitro prevents extensive analyses of mitochondrial replication and inheritance, information about these processes could be inferred by investigating the level of mitochondrial heteroplasmy present in C. felis isolates. This topic will be addressed in Chapter 3 and 4 of this thesis. TREATMENT AND PROGNOSIS OF CYTAUXZOONOSIS For nearly 25 years after Cytauxzoon felis was first discovered in domestic cats, acute cytauxzoonosis was considered an untreatable disease. With the exception of a few isolated reports of cats surviving infection (2, 13, 117), the disease was thought to be universally fatal and euthanasia was considered the most humane option for infected cats (118). Early attempts at therapeutic intervention included a variety of different antibiotics as well as antiprotozoal agents used to treat bovine theileriosis (parvaquone and buparvaquone); however, these strategies showed little promise for being effective treatments against cytauxzoonosis (117, 119). The first reported therapies associated with the survival of 14
33 multiple cats were diminazene aceturate and imidocarb dipropionate (7, 14). However, diminazene aceturate is not approved for use in the United States and has been associated with adverse events in treated cats, and thus proved to be an impractical treatment option (120, 121). Imidocarb dipropionate, a more widely available drug commonly used to treat babesiosis, was claimed anecdotally to increase survival rates to around 50%, although little evidence of this efficacy was demonstrated in published work (7, 14). Imidocarb dipropionate was temporarily considered the therapy of choice (6) until atovaquone and azithromycin (A&A) therapy was introduced (3). In a prospective randomized clinical trial, the survival rate of cats treated with A&A (60%) was significantly higher (p<0.05) than that of cats treated with imidocarb dipropionate (26%, 3). Thus, A&A currently is the recommended treatment for acute cytauxzoonosis. Atovaquone and azithromycin have been used separately or together as treatments for a variety of protozoal infections in both humans and animals, including Plasmodium, Babesia, and Toxoplasma. Atovaquone, a ubiquinone analogue also known as 566C80, was developed as an antimalarial and antifungal chemotherapeutic (122). Atovaquone functions by disrupting parasite mitochondrial electron transport through multiple mechanisms, the most well described being through inhibition of the cytochrome bc1 complex in the electron transport chain (Figure 6; 110, ). Azithromycin is a relatively broad-spectrum macrolide that inhibits bacterial protein synthesis via binding to the 70S prokaryotic ribosome. In the treatment of eukaryotic infectious organisms, azithromycin is thought to bind to mitochondrial ribosomes (127). These drugs were first used in combination for the treatment of Babesia microti in hamsters and humans, and in the latter was shown to be better tolerated than alternative treatment options (128, 129). A&A has subsequently been 15
, and in addition to being used against Cytauxzoon felis, has also been used to successfully treat Babesia gibsoni and Babesia conradae infections in dogs (131, 132).")
34 recommended as the first treatment option for mild to moderate B. microti infections in humans (130), and in addition to being used against Cytauxzoon felis, has also been used to successfully treat Babesia gibsoni and Babesia conradae infections in dogs (131, 132). Figure 6. Atovaquone inhibits electron transport within the parasite mitochondria. As in other eukaryotes, the parasite electron transport chain is located within the inner mitochondrial membrane (I) and concentrates protons in the space between the inner and outer (O) mitochondrial membranes in order to create an electrochemical gradient to drive ATP production. Atovaquone binds to the ubiquinol oxidation site (Q o ) of the parasite cytochrome bc 1 complex, preventing the oxidation of electron carrier ubiquinol (QH 2 ) to ubiquinone (Q). This ultimately leads to inhibition of the electron transport chain, decreased ATP production, reduced metabolic output, and subsequent parasite death. However, atovaquone resistance has been documented for a number of these organisms, including many Plasmodium species, Babesia microti, and Babesia gibsoni (111, ). Resistance to atovaquone has been associated with missense mutations in the mitochondrial cytochrome b gene (cytb), which encodes the main subunit of the cytochrome bc1 complex; these mutations lead to subsequent protein conformational changes that inhibit atovaquone binding (111, 124, 133, 134, 136, 137). Resistance to A&A treatment has led to persistent parasitic infections and in some cases, patient death (135). In the case of 16
35 Cytauxzoon felis, 40% of cats treated with A&A died (3). While it is likely that other factors (e.g., severity of disease at clinical presentation to veterinary hospital) contributed to this relatively high mortality, we speculated that some of the cats that died may have been infected with C. felis strains that are inherently resistant to A&A therapy. We hypothesized that there would be an association between treatment response (survival) and C. felis cytb genotype, and that cats that died would be infected with C. felis strains with missense mutations in the cytb gene. This topic will be addressed in Chapter 3 of this thesis. Furthermore, we anticipate that C. felis cytb genotype may be able to predict response to A&A therapy and could be useful as a prognostic indicator for acutely infected cats. Currently, little work has been done in identifying molecular, epidemiological, or physiological parameters that predict survival of infected cats. Although it was originally believed that C. felis ITS1-ITS2 genotypes were markers of pathogenicity (138), it has since been demonstrated that this is not the case (16). Additionally, there is a high prevalence of infected yet asymptomatic domestic cats in the central United States (17), leading to the unconfirmed speculation that there are geographically distinct strains of C. felis that may vary in pathogenicity. Last, it has been shown that cats that die of acute cytauxzoonosis have more severe leukopenia and higher bilirubinemia (3). However, none of these potential prognostic indicators have been investigated in the context of A&A treatment. When considering that A&A treatment in conjunction with supportive care can cost thousands of dollars, the development of a cost-effective prognostic test that predicts response to A&A therapy would aid both pet owners and clinicians in deciding on treatment options. We hypothesize that C. felis cytb genotype will aid in the prognosis of 17
36 cytauxzoonosis in cats treated with A&A. This topic will be addressed in Chapter 4 of this thesis. PREVENTION OF CYTAUXZOONOSIS When considering the expanding geographic distribution of C. felis as well as the current shortcomings of treating cytauxzoonosis, it becomes clear that preventing infection and/or clinical disease would be the ideal method of control. Currently, the only method of preventing the spread of C. felis is to prevent tick attachment to domestic cats by environmental modification (i.e., restriction from outdoors) and/or the use of appropriate acaricides. However, there are a number of limitations to relying on these methods alone. While limiting outdoor exposure, especially in areas that promote propagation of the tick life cycle, will likely aid in curbing the spread of C. felis, this strategy alone is insufficient for preventing disease for a number of reasons. First, this strategy is unlikely to be adopted by all pet owners, as an estimated million pet cats are allowed to roam outdoors in the United States (139, 140). In these cases, it is unlikely that owners will be able to prevent cats from roaming in environments that support questing ticks. Second, even for those pet owners that do keep cats strictly indoors, infected ticks could still be carried inside the house by other pets or humans. A recent study indicated both indoor and outdoor cats living in areas surrounded by grassland, fragmented landscapes, or mixed forests were at equal risk for being infected with C. felis; in other words, restricting exposure to the outdoors did not significantly reduce exposure to C. felis (141). Furthermore, a number of factors may limit the effectiveness of acaricide prophylaxis in preventing cytauxzoonosis. First, in comparison to their canine counterparts, there are few safe acaricides available for cats (142). The majority of companion animal acaricides 18
37 currently marketed include permethrin or permethrin-derivatives, which are toxic to cats (143). The two primary acaricides available for feline species are fipronil and flumethrin. While flumethrin has been shown to effectively prevent the transmission of C. felis (8), no comparable studies have been conducted for fipronil. However, multiple studies have shown that fipronil works much more slowly than other acaricides, and can take as long as hours to kill ticks (144, 145). This could be important, as it may be possible for C. felis to be transmitted from tick to cat in less than 48 hours, although this hasn t been tested to date. Second, tick resistance to both of these compounds has been well documented ( ), although notably not for either of the tick species known to transmit C. felis. However, a recent study conducted in a region endemic for C. felis suggested that acaricide resistance is likely to emerge for Amblyomma americanum given the genetic diversity present in its expanding population (150). Last, a lack of compliance in application of acaricides by pet owners may prevent efficacy, as a recent study conducted at a veterinary teaching hospital indicated that as few as 38% of cats presenting for treatment received any form of tick prevention (151). Clearly, although helpful, these methods are not practical for completely preventing cats from contracting C. felis. Alternatively, the development of a vaccine against cytauxzoonosis has been proposed as an ideal mechanism of prevention. Previous studies have demonstrated that cats that mount an immune response against C. felis and survive are protected from clinical disease following subsequent infections, suggesting that an appropriately designed vaccine could also prevent illness (2, 117). Because so little is known about C. felis, investigation into vaccination strategies currently used against closely related 19
38 organisms (Theileria, Babesia, and Plasmodium species) may aid in choosing an effective approach for immunization against C. felis. In general, live vaccines offer the most potential for generating a protective immune response against disease, and currently live attenuated vaccines are commonly used for the prevention of bovine theilerioses and babesioses ( ). However, given the high virulence of C. felis, a live vaccine would need to be greatly attenuated in order to be safe. Although it has been speculated that naturally occurring strains of C. felis exist that are less pathogenic (17), this has not been confirmed. Thus, an attenuated strain of C. felis would need to be created. For related organisms, attenuation is typically achieved via prolonged in vitro culture of parasite-infected cells and/or direct genetic manipulation of the parasite (152, ). However, to date successful in vitro culture of C. felis has not been reported, making these attenuation strategies unfeasible. Even if laboratory production of an attenuated live vaccine was possible, there are a variety of safety and logistical concerns with this technique. If the attenuated strain administered as a vaccine were to infect ticks, reversion to virulence and spread to other felids could occur, as has happened for Theileria (154, 159). A live attenuated vaccine may also induce more severe clinical symptoms in younger animals (154), which may limit the use of the vaccine in small animal clinics where kittens would be the primary target of immunization. Last, requirements for storage (liquid nitrogen), preparation, and administration (must be given rapidly after thawing) of live attenuated vaccines are impractical for implementation in small animal clinics (152, 154). Other vaccination approaches utilizing whole organisms include killed vaccines (160) or heterologous vaccines (i.e., infecting the animal with an organism related to the pathogen of interest). Because these approaches don t require in vitro culture of C. felis they may be 20
39 more feasible, but still have a number of limitations. A killed C. felis vaccine could be produced from, for example, formalin-fixed tissues of a Cytauxzoon felis-infected cat. However, even if effective, this would require constantly infecting and sacrificing cats to produce the vaccine. Alternatively, Theileria annulata or parva, which are easily grown in vitro, could be used as a heterologous vaccine. However, in addition to the difficulties mentioned above for live attenuated vaccines, the threat of spread to cattle and potential risk to the beef and dairy industry would likely make this an unrealistic option. Most importantly, there is no data to support that either of these approaches would be effective or safe methods for the prevention of cytauxzoonosis. Consequently, because of the inability to study the parasite in vitro, initial pilot studies investigating efficacy and safety of any of these vaccination strategies would need to be conducted in cats. This would be less than ideal given both the financial restraints and the ethical concerns involved with conducting in vivo studies of cytauxzoonosis. For this reason, it would be preferable to create a vaccine with demonstrated ability to invoke an immune response prior to pursuing in vivo immunization studies. Fortunately, the rapid progression of genome sequencing technology has allowed for the creation of novel, genome-based approaches for developing vaccines, particularly against uncultivable and highly virulent pathogens such as C. felis. Known as reverse vaccinology, this strategy consists of mining the whole genome of the pathogen of interest for eligible candidates to incorporate into a subunit vaccine (161). Our lab has recently sequenced, assembled, and annotated the full Cytauxzoon felis genome, which includes a computationally predicted proteome that is searchable for vaccine candidates (50). We have used a multi-faceted approach to pinpoint C. felis vaccine candidates, including identifying 21
. Over 650 potential candidates were subsequently synthesized and probed with serum from C.")

40 orthologues to the leading vaccine candidates of related parasites (50), predicting highly antigenic peptides in silico, and identifying orthologous to Plasmodium falciparum proteins that are reactive against serum of C. felis-infected cats (Tarigo et al., unpublished data). Over 650 potential candidates were subsequently synthesized and probed with serum from C. felis-infected cats; from this pool, 33 candidates were differentially reactive against C. felisinfected serum (Figure 7; Schreeg et al., unpublished data). Using these candidates, we hope to manufacture a C. felis vaccine for assessment in a pilot study. However, it is unclear what strategy of antigen delivery will be most effective, safe, or feasible. Figure 7. Identification of Cytauxzoon felis vaccine candidates. Candidates were selected that were differentially reactive (p<0.05, denoted by black hash marks) against serum from C. felis-infected cats (red bars) compared to serum from uninfected cats (blue bars). P values for comparison between infected and uninfected cats are denoted by purple line. Candidates were also screened against serum from specific pathogen free (SPF) cats (green bars); reactivity of this serum was also compared to that against infected serum (blue line). Seroreactivity of 30 candidates is shown. In contrast to whole organism vaccines, subunit vaccines expose immunized individuals to specific portions of the pathogen that are highly antigenic, and rely on immune memory created from exposure to these antigens to combat the pathogen. Subunit vaccines have traditionally been composed of identified antigenic proteins combined with adjuvant to aid in triggering an immune reaction, and are currently being investigated as vaccine options for related parasites (162, 163). However, protein subunit vaccines are associated with an 22
41 increased risk of injection site sarcomas in cats (164), and additionally, synthesis of the amount of protein required for incorporation of all 33 candidates into a vaccine would be costly and labor-intensive. Alternatively, a number of novel systems have been developed for the delivery of antigens in a vaccine. One attractive option is creation of a recombinant virus expressing vaccine candidates, a strategy that is safely and effectively used in other feline vaccines and has shown promise in vaccination against Plasmodium (158, ). However, the viral vectors commonly used for production of feline vaccines are not commercially available and therefore unattainable for a pilot study. In contrast, a DNAbased vaccine is a strategy that is relatively straightforward and inexpensive to produce. DNA vaccines are currently being explored as viable immunization options against a variety of protozoal parasites (169). DNA vaccines deliver genes encoding vaccine candidates in intricately designed expression plasmids (170). Expression of vaccine candidates, which is under the control of promotors that induce high levels of transcription (e.g., cytomegalovirus promotor), and subsequent host immune response is theoretically achieved once injected into the animal. Furthermore, a large pool of vaccine candidates that otherwise may be too costly to synthesize as proteins or impractical for incorporation into a recombinant virus can be combined into a single DNA vaccine through a technique known as expression library immunization (ELI). ELI is a strategy in which individuals are vaccinated with a library of expression plasmids, often which contain a portion of or the entire genome of the pathogen of interest (Figure 8; 171, 172). Though frequently employed as a high-throughput approach for screening the pathogen genome for vaccine candidates, this technique has also been used to induce a broad immune response against multiple antigens. Expression library vaccines have been demonstrated to induce protection against a 23

42 variety of protozoal infections in mice, including Plasmodium, Leishmania, and Trypanosoma species (169, ). Similarly, we hypothesize that an ELI approach may be effective in preventing cytauxzoonosis in cats. This topic will be addressed in Chapter 6 of this thesis. Figure 8. Conceptual depiction of expression library immunization against Cytauxzoon felis. 24
43 REFERENCES 1. Meinkoth JH, Kocan AA Feline cytauxzoonosis. Vet Clin North Am Small Anim Pract 35:89-101, vi. 2. Ferris DH A progress report on the status of a new disease of American cats: cytauxzoonosis. Comp Immunol Microbiol Infect Dis 1: Cohn LA, Birkenheuer AJ, Brunker JD, Ratcliff ER, Craig AW Efficacy of atovaquone and azithromycin or imidocarb dipropionate in cats with acute cytauxzoonosis. J Vet Intern Med 25: Conner BJ, Hanel, R.M., Brooks, M., Cohn, L.A., Birkenheuer, A.J Coagulation abnormalities in 5 cats with naturally occurring cytauxzoonosis. Journal of Veterinary Emergency and Critical Care doi:doi: /vec Birkenheuer AJ, Le JA, Valenzisi AM, Tucker MD, Levy MG, Breitschwerdt EB Cytauxzoon felis infection in cats in the mid-atlantic states: 34 cases ( ). J Am Vet Med Assoc 228: Greene CE Infectious diseases of the dog and cat, 2nd ed. W.B. Saunders, Philadelphia. 7. Greene CE, Latimer K, Hopper E, Shoeffler G, Lower K, Cullens F Administration of diminazene aceturate or imidocarb dipropionate for treatment of cytauxzoonosis in cats. J Am Vet Med Assoc 215: , Reichard MV, Thomas JE, Arther RG, Hostetler JA, Raetzel KL, Meinkoth JH, Little SE Efficacy of an imidacloprid 10 % / flumethrin 4.5 % collar (Seresto(R), Bayer) for preventing the transmission of Cytauxzoon felis to domestic cats by Amblyomma americanum. Parasitol Res 112 Suppl 1: MacNeill AL, Barger AM, Skowronski MC, Lanka S, Maddox CW Identification of Cytauxzoon felis infection in domestic cats from southern Illinois. J Feline Med Surg doi: / x Miller J, Davis CD Increasing frequency of feline cytauxzoonosis cases diagnosed in western Kentucky from 2001 to Vet Parasitol 198: Birkenheuer AJ, Marr HS, Warren C, Acton AE, Mucker EM, Humphreys JG, Tucker MD Cytauxzoon felis infections are present in bobcats (Lynx rufus) in a region where cytauxzoonosis is not recognized in domestic cats. Vet Parasitol 153: Shock BC, Birkenheuer AJ, Patton LL, Olfenbuttel C, Beringer J, Grove DM, Peek M, Butfiloski JW, Hughes DW, Lockhart JM, Cunningham MW, Brown 25
44 HM, Peterson DS, Yabsley MJ Variation in the ITS-1 and ITS-2 rrna genomic regions of Cytauxzoon felis from bobcats and pumas in the eastern United States and comparison with sequences from domestic cats. Vet Parasitol 190: Walker DB, Cowell RL Survival of a domestic cat with naturally acquired cytauxzoonosis. J Am Vet Med Assoc 206: Meinkoth J, Kocan AA, Whitworth L, Murphy G, Fox JC, Woods JP Cats surviving natural infection with Cytauxzoon felis: 18 cases ( ). J Vet Intern Med 14: Haber MD, Tucker MD, Marr HS, Levy JK, Burgess J, Lappin MR, Birkenheuer AJ The detection of Cytauxzoon felis in apparently healthy freeroaming cats in the USA. Vet Parasitol 146: Brown HM, Lockhart JM, Latimer KS, Peterson DS Identification and genetic characterization of Cytauxzoon felis in asymptomatic domestic cats and bobcats. Vet Parasitol 172: Rizzi TE, Reichard MV, Cohn LA, Birkenheuer AJ, Taylor JD, Meinkoth JH Prevalence of Cytauxzoon felis infection in healthy cats from enzootic areas in Arkansas, Missouri, and Oklahoma. Parasit Vectors 8: Neitz WO, Thomas AD Cytauxzoon sylvicaprae gen. nov., spec. nov., a protozoon responsible for a hitherto undescribed disease in the duiker, Sylvicapra grimmia (Linne). Onderstepoort J Vet Sci Anim Ind 23: Nietz WO Theileriosis, gonderioses, and cytauxzoonoses. A review. Onderstepoort Journal of Veterinary Research 27: Levine ND Taxonomy of the Piroplasms. Transactions of the American Microscopal Society 90: Brocklesby DW Cytauxzoon taurotragi Martin & Brocklesby 1960, a piroplasm of the eland (Taurotragus oryx pattersonianus Lydekker, 1906). Research in Veterinary Science 3: McCully RM, Keep ME, Basson PA Cytauxzoonosis in a giraffe (Giraffa camelopardalis (Linnaeus, 1758)) in Zululand. Onderstepoort J Vet Res 37: Grootenhuis JG, Young AS, Dolan TT, Stagg DA Characteristics of Theileria species (eland) infections in eland and cattle. Res Vet Sci 27: Spooner RL, Innes EA, Glass EJ, Millar P, Brown CG Bovine mononuclear cell lines transformed by Theileria parva or Theileria annulata express different subpopulation markers. Parasite Immunol 10:
45 25. Spooner RL, Innes EA, Glass EJ, Brown CG Theileria annulata and T. parva infect and transform different bovine mononuclear cells. Immunology 66: Nijhof AM, Pillay V, Steyl J, Prozesky L, Stoltsz WH, Lawrence JA, Penzhorn BL, Jongejan F Molecular characterization of Theileria species associated with mortality in four species of African antelopes. J Clin Microbiol 43: Young AS, Grootenhuis JG, Leitch BL, Schein E The development of Theileria=Cytauxzoon taurotragi (Martin and Brocklesby, 1960) from eland in its tick vector Rhipicephalus appendiculatus. Parasitology 81: Thomas SE, Wilson DE, Mason TE Babesia, Theileria and Anaplasma spp. infecting sable antelope, Hippotragus niger (Harris, 1838), in southern Africa. Onderstepoort J Vet Res 49: Levine ND The protozoan phylum Apicomplexa. CRC Press, Boca Raton, Fla. 30. Uilenberg G, Franssen FF, Perie NM Relationships between Cytauxzoon felis and African piroplasmids. Vet Parasitol 26: Wagner JE A fatal cytauxzoonosis-like disease in cats. J Am Vet Med Assoc 168: Schnittger L, Rodriguez AE, Florin-Christensen M, Morrison DA Babesia: A world emerging. Infect Genet Evol 12: Allsopp MT, Allsopp BA Molecular sequence evidence for the reclassification of some Babesia species. Annals of the New York Academy of Sciences 1081: Criado-Fornelio A, Martinez-Marcos A, Buling-Sarana A, Barba-Carretero JC Molecular studies on Babesia, Theileria and Hepatozoon in southern Europe. Part II. Phylogenetic analysis and evolutionary history. Veterinary Parasitology 114: Lack JB, Reichard MV, Van Den Bussche RA Phylogeny and evolution of the Piroplasmida as inferred from 18S rrna sequences. International Journal for Parasitology 42: Reichard MV, Van Den Bussche RA, Meinkoth JH, Hoover JP, Kocan AA A new species of Cytauxzoon from Pallas' cats caught in Mongolia and comments on the systematics and taxonomy of piroplasmids. J Parasitol 91:
46 37. Allsopp MT, Cavalier-Smith T, De Waal DT, Allsopp BA Phylogeny and evolution of the piroplasms. Parasitology 108 ( Pt 2): Mehlhorn H, Schein E Redescription of Babesia equi Laveran, 1901 as Theileria equi Mehlhorn, Schein Parasitol Res 84: Kappmeyer LS, Thiagarajan M, Herndon DR, Ramsay JD, Caler E, Djikeng A, Gillespie JJ, Lau AO, Roalson EH, Silva JC, Silva MG, Suarez CE, Ueti MW, Nene VM, Mealey RH, Knowles DP, Brayton KA Comparative genomic analysis and phylogenetic position of Theileria equi. BMC Genomics 13: Ketz-Riley CJ, Reichard MV, Van den Bussche RA, Hoover JP, Meinkoth J, Kocan AA An intraerythrocytic small piroplasm in wild-caught Pallas's cats (Otocolobus manul) from Mongolia. J Wildl Dis 39: Millan J, Candela MG, Palomares F, Cubero MJ, Rodriguez A, Barral M, de la Fuente J, Almeria S, Leon-Vizcaino L Disease threats to the endangered Iberian lynx (Lynx pardinus). Vet J 182: Carli E, Trotta M, Chinelli R, Drigo M, Sinigoi L, Tosolini P, Furlanello T, Millotti A, Caldin M, Solano-Gallego L Cytauxzoon sp. infection in the first endemic focus described in domestic cats in Europe. Vet Parasitol 183: Leclaire S, Menard S, Berry A Molecular characterization of Babesia and Cytauxzoon species in wild South-African meerkats. Parasitology 142: Dobbelaere D, Heussler V Transformation of leukocytes by Theileria parva and T. annulata. Annu Rev Microbiol 53: Dobbelaere DA, Kuenzi P The strategies of the Theileria parasite: a new twist in host-pathogen interactions. Curr Opin Immunol 16: Kier AB, Wagner JE, Kinden DA The pathology of experimental cytauxzoonosis. J Comp Pathol 97: Snider TA, Confer AW, Payton ME Pulmonary histopathology of Cytauxzoon felis infections in the cat. Vet Pathol 47: Hagiwara K, Takahashi K, Taniyama H, Kawamoto S, Kurosawa T, Ikuta K, Ishihara C Detection of Theileria sergenti schizonts in bovine lymph node. Int J Parasitol 27: Sato M, Kamio T, Kawazu S, Taniguchi T, Minami T, Fujisaki K Histological observations on the schizonts in cattle infected with Japanese Theileria sergenti. J Vet Med Sci 55:
47 50. Tarigo JL, Scholl EH, Bird DM, Brown CC, Cohn LA, Dean GA, Levy MG, Doolan DL, Trieu A, Nordone SK, Felgner PL, Vigil A, Birkenheuer AJ A novel candidate vaccine for cytauxzoonosis inferred from comparative apicomplexan genomics. PLoS One 8:e Susta L, Torres-Velez F, Zhang J, Brown C An in situ hybridization and immunohistochemical study of cytauxzoonosis in domestic cats. Vet Pathol 46: Kjemtrup AM, Thomford J, Robinson T, Conrad PA Phylogenetic relationships of human and wildlife piroplasm isolates in the western United States inferred from the 18S nuclear small subunit RNA gene. Parasitology 120 ( Pt 5): Zahler M, Rinder H, Gothe R Genotypic status of Babesia microti within the piroplasms. Parasitol Res 86: Goethert HK, Telford SR, 3rd What is Babesia microti? Parasitology 127: Kjemtrup AM, Wainwright K, Miller M, Penzhorn BL, Carreno RA Babesia conradae, sp. Nov., a small canine Babesia identified in California. Vet Parasitol 138: Conrad PA, Kjemtrup AM, Carreno RA, Thomford J, Wainwright K, Eberhard M, Quick R, Telford SR, 3rd, Herwaldt BL Description of Babesia duncani n.sp. (Apicomplexa: Babesiidae) from humans and its differentiation from other piroplasms. Int J Parasitol 36: Ramsay JD, Ueti MW, Johnson WC, Scoles GA, Knowles DP, Mealey RH Lymphocytes and macrophages are infected by Theileria equi, but T cells and B cells are not required to establish infection in vivo. PLoS One 8:e Morrison DA, Ellis JT Effects of nucleotide sequence alignment on phylogeny estimation: a case study of 18S rdnas of apicomplexa. Mol Biol Evol 14: Cornillot E, Hadj-Kaddour K, Dassouli A, Noel B, Ranwez V, Vacherie B, Augagneur Y, Bres V, Duclos A, Randazzo S, Carcy B, Debierre-Grockiego F, Delbecq S, Moubri-Menage K, Shams-Eldin H, Usmani-Brown S, Bringaud F, Wincker P, Vivares CP, Schwarz RT, Schetters TP, Krause PJ, Gorenflot A, Berry V, Barbe V, Ben Mamoun C Sequencing of the smallest Apicomplexan genome from the human pathogen Babesia microti. Nucleic Acids Res 40:
48 60. Feagin JE Mitochondrial genome diversity in parasites. Int J Parasitol 30: Hikosaka K, Kita K, Tanabe K Diversity of mitochondrial genome structure in the phylum Apicomplexa. Mol Biochem Parasitol 188: Gray MW, Burger G, Lang BF Mitochondrial evolution. Science 283: Feagin JE, Harrell MI, Lee JC, Coe KJ, Sands BH, Cannone JJ, Tami G, Schnare MN, Gutell RR The fragmented mitochondrial ribosomal RNAs of Plasmodium falciparum. PLoS One 7:e Hikosaka K, Nakai Y, Watanabe Y, Tachibana S, Arisue N, Palacpac NM, Toyama T, Honma H, Horii T, Kita K, Tanabe K Concatenated mitochondrial DNA of the coccidian parasite Eimeria tenella. Mitochondrion 11: Hikosaka K, Watanabe Y, Kobayashi F, Waki S, Kita K, Tanabe K Highly conserved gene arrangement of the mitochondrial genomes of 23 Plasmodium species. Parasitol Int 60: Hikosaka K, Tsuji N, Watanabe YI, Kishine H, Horii T, Igarashi I, Kita K, Tanabe K Novel type of linear mitochondrial genomes with dual flip-flop inversion system in apicomplexan parasites, Babesia microti and Babesia rodhaini. BMC Genomics 13: Hikosaka K, Watanabe Y, Tsuji N, Kita K, Kishine H, Arisue N, Palacpac NM, Kawazu S, Sawai H, Horii T, Igarashi I, Tanabe K Divergence of the mitochondrial genome structure in the apicomplexan parasites, Babesia and Theileria. Mol Biol Evol 27: Sankoff D, Leduc G, Antoine N, Paquin B, Lang BF, Cedergren R Gene order comparisons for phylogenetic inference: evolution of the mitochondrial genome. Proc Natl Acad Sci U S A 89: Moritz C, Dowling, T. E., Brown, W. M Evolution of animal mitochondrial DNA: Relevance for population biology and systematics. Annual Review of Ecology and Systematics 18: Boore JL Animal mitochondrial genomes. Nucleic Acids Res 27: Le TH, Blair D, McManus DP Mitochondrial genomes of human helminths and their use as markers in population genetics and phylogeny. Acta Trop 77:
49 72. Gou H, Guan G, Liu A, Ma M, Xu Z, Liu Z, Ren Q, Li Y, Yang J, Chen Z, Yin H, Luo J A DNA barcode for Piroplasmea. Acta Trop 124: He L, Zhang Y, Zhang QL, Zhang WJ, Feng HH, Khan MK, Hu M, Zhou YQ, Zhao JL Mitochondrial genome of Babesia orientalis, apicomplexan parasite of water buffalo (Bubalus babalis, Linnaeus, 1758) endemic in China. Parasit Vectors 7: Kher CP, Doerder FP, Cooper J, Ikonomi P, Achilles-Day U, Kupper FC, Lynn DH Barcoding Tetrahymena: discriminating species and identifying unknowns using the cytochrome c oxidase subunit I (cox-1) barcode. Protist 162: Chantangsi C, Lynn DH Phylogenetic relationships within the genus Tetrahymena inferred from the cytochrome c oxidase subunit 1 and the small subunit ribosomal RNA genes. Mol Phylogenet Evol 49: Ogedengbe JD, Hanner RH, Barta JR DNA barcoding identifies Eimeria species and contributes to the phylogenetics of coccidian parasites (Eimeriorina, Apicomplexa, Alveolata). International Journal for Parasitology 41: Holman PJ, Snowden KF Canine hepatozoonosis and babesiosis, and feline cytauxzoonosis. Vet Clin North Am Small Anim Pract 39: , v. 78. Birkenheuer AJ, Marr H, Alleman AR, Levy MG, Breitschwerdt EB Development and evaluation of a PCR assay for the detection of Cytauxzoon felis DNA in feline blood samples. Vet Parasitol 137: Shindel N, Dardiri AH, Ferris DH An indirect fluorescent antibody test for the detection of Cytauxzoon-like organisms in experimentally infected cats. Can J Comp Med 42: Brown HM, Latimer KS, Erikson LE, Cashwell ME, Britt JO, Peterson DS Detection of persistent Cytauxzoon felis infection by polymerase chain reaction in three asymptomatic domestic cats. J Vet Diagn Invest 20: Birkenheuer AJ, Levy MG, Breitschwerdt EB Development and evaluation of a seminested PCR for detection and differentiation of Babesia gibsoni (Asian genotype) and B. canis DNA in canine blood samples. J Clin Microbiol 41: Wang G, Villafuerte P, Zhuge J, Visintainer P, Wormser GP Comparison of a quantitative PCR assay with peripheral blood smear examination for detection and quantitation of Babesia microti infection in humans. Diagn Microbiol Infect Dis 82:
50 83. Miro G, Checa R, Paparini A, Ortega N, Gonzalez-Fraga JL, Gofton A, Bartolome A, Montoya A, Galvez R, Mayo PP, Irwin P Theileria annae (syn. Babesia microti-like) infection in dogs in NW Spain detected using direct and indirect diagnostic techniques: clinical report of 75 cases. Parasit Vectors 8: Almeria S, Castella J, Ferrer D, Ortuno A, Estrada-Pena A, Gutierrez JF Bovine piroplasms in Minorca (Balearic Islands, Spain): a comparison of PCR-based and light microscopy detection. Vet Parasitol 99: Calder JA, Reddy GR, Chieves L, Courtney CH, Littell R, Livengood JR, Norval RA, Smith C, Dame JB Monitoring Babesia bovis infections in cattle by using PCR-based tests. J Clin Microbiol 34: Salem GH, Liu X, Johnsrude JD, Dame JB, Roman Reddy G Development and evaluation of an extra chromosomal DNA-based PCR test for diagnosing bovine babesiosis. Mol Cell Probes 13: Bilgic HB, Karagenc T, Shiels B, Tait A, Eren H, Weir W Evaluation of cytochrome b as a sensitive target for PCR based detection of T. annulata carrier animals. Vet Parasitol 174: Haanshuus CG, Mohn SC, Morch K, Langeland N, Blomberg B, Hanevik K A novel, single-amplification PCR targeting mitochondrial genome highly sensitive and specific in diagnosing malaria among returned travellers in Bergen, Norway. Malar J 12: Buling A, Criado-Fornelio A, Asenzo G, Benitez D, Barba-Carretero JC, Florin- Christensen M A quantitative PCR assay for the detection and quantification of Babesia bovis and B. bigemina. Vet Parasitol 147: Joseph JT, Aldritt SM, Unnasch T, Puijalon O, Wirth DF Characterization of a conserved extrachromosomal element isolated from the avian malarial parasite Plasmodium gallinaceum. Mol Cell Biol 9: Preiser PR, Wilson RJ, Moore PW, McCready S, Hajibagheri MA, Blight KJ, Strath M, Williamson DH Recombination associated with replication of malarial mitochondrial DNA. EMBO J 15: Vaidya AB, Arasu P Tandemly arranged gene clusters of malarial parasites that are highly conserved and transcribed. Mol Biochem Parasitol 22: Wilson RJ, Williamson DH Extrachromosomal DNA in the Apicomplexa. Microbiol Mol Biol Rev 61: Jura WG, Brown CG, Rowland AC Ultrastructural characteristics of in vitro parasite-lymphocyte behaviour in invasions with Theileria annulata and Theileria parva. Vet Parasitol 12:
51 95. Shaw MK, Tilney LG How individual cells develop from a syncytium: merogony in Theileria parva (Apicomplexa). J Cell Sci 101 ( Pt 1): Conrad PA, Kelly BG, Brown CG Intraerythrocytic schizogony of Theileria annulata. Parasitology 91 ( Pt 1): Conrad PA, Denham D, Brown CG Intraerythrocytic multiplication of Theileria parva in vitro: an ultrastructural study. Int J Parasitol 16: Fawcett DW, Conrad PA, Grootenhuis JG, Morzaria SP Ultrastructure of the intra-erythrocytic stage of Theileria species from cattle and waterbuck. Tissue Cell 19: Krungkrai J The multiple roles of the mitochondrion of the malarial parasite. Parasitology 129: Learngaramkul P, Petmitr S, Krungkrai SR, Prapunwattana P, Krungkrai J Molecular characterization of mitochondria in asexual and sexual blood stages of Plasmodium falciparum. Mol Cell Biol Res Commun 2: Petmitr S, Krungkrai J Mitochondrial cytochrome b gene in two developmental stages of human malarial parasite Plasmodium falciparum. Southeast Asian J Trop Med Public Health 26: Mehlhorn H, Schein, E, Ahmed, S. J Theileria, p In Kreier JP (ed), Parasitic Protozoa, 2 ed, vol 7. Academic Press, Inc., San Diego, CA Uchida TA, Yanagawa H, Mori T, Shiraishi S Ultrastructural observations of the intra-erythrocytic merozoite in Theileria sergenti. Trop Med Parasitol 36: Simpson CF, Harvey JW, Carlisle JW Ultrastructure of the intraerythrocytic stage of Cytauxzoon felis. Am J Vet Res 46: Aikawa M Variations in structure and function during the life cycle of malarial parasites. Bull World Health Organ 55: Hopkins J, Fowler R, Krishna S, Wilson I, Mitchell G, Bannister L The plastid in Plasmodium falciparum asexual blood stages: a three-dimensional ultrastructural analysis. Protist 150: Divo AA, Geary TG, Jensen JB, Ginsburg H The mitochondrion of Plasmodium falciparum visualized by rhodamine 123 fluorescence. J Protozool 32:
52 108. Jarvi SI, Farias ME, Lapointe DA, Belcaid M, Atkinson CT Nextgeneration sequencing reveals cryptic mtdna diversity of Plasmodium relictum in the Hawaiian Islands. Parasitology 140: Williamson DH, Preiser PR, Wilson RJ Organelle DNAs: The bit players in malaria parasite DNA replication. Parasitol Today 12: Mather MW, Henry KW, Vaidya AB Mitochondrial drug targets in apicomplexan parasites. Curr Drug Targets 8: Korsinczky M, Chen N, Kotecka B, Saul A, Rieckmann K, Cheng Q Mutations in Plasmodium falciparum cytochrome b that are associated with atovaquone resistance are located at a putative drug-binding site. Antimicrob Agents Chemother 44: Ekala MT, Khim N, Legrand E, Randrianarivelojosia M, Jambou R, Fandeur T, Menard D, Assi SB, Henry MC, Rogier C, Bouchier C, Mercereau-Puijalon O Sequence analysis of Plasmodium falciparum cytochrome b in multiple geographic sites. Malar J 6: Gassis S, Rathod PK Frequency of drug resistance in Plasmodium falciparum: a nonsynergistic combination of 5-fluoroorotate and atovaquone suppresses in vitro resistance. Antimicrob Agents Chemother 40: Creasey AM, Ranford-Cartwright LC, Moore DJ, Williamson DH, Wilson RJ, Walliker D, Carter R Uniparental inheritance of the mitochondrial gene cytochrome b in Plasmodium falciparum. Curr Genet 23: White DJ, Wolff JN, Pierson M, Gemmell NJ Revealing the hidden complexities of mtdna inheritance. Mol Ecol 17: Sobenin IA, Mitrofanov KY, Zhelankin AV, Sazonova MA, Postnov AY, Revin VV, Bobryshev YV, Orekhov AN Quantitative assessment of heteroplasmy of mitochondrial genome: perspectives in diagnostics and methodological pitfalls. Biomed Res Int 2014: Motzel SL, Wagner JE Treatment of experimentally induced cytauxzoonosis in cats with parvaquone and buparvaquone. Vet Parasitol 35: Wagner JE, Kier, A. B Cytauxzoonosis. In Barlough JE (ed), Manual of Small Animal Infectious Diseases. Churchill Livingstone, New York Hoover JP, Walker DB, Hedges JD Cytauxzoonosis in cats: eight cases ( ). J Am Vet Med Assoc 205: Lewis KM, Cohn LA, Marr HS, Birkenheuer AJ Failure of efficacy and adverse events associated with dose-intense diminazene diaceturate treatment of 34
53 chronic Cytauxzoon felis infection in five cats. J Feline Med Surg doi: / x Lewis KM, Cohn LA, Marr HS, Birkenheuer AJ Diminazene diaceturate for treatment of chronic Cytauxzoon felis parasitemia in naturally infected cats. J Vet Intern Med 26: Gutteridge WE Antimalarial drugs currently in development. J R Soc Med 82 Suppl 17:63-66; discussion Fry M, Pudney M Site of action of the antimalarial hydroxynaphthoquinone, 2-[trans-4-(4'-chlorophenyl) cyclohexyl]-3-hydroxy-1,4-naphthoquinone (566C80). Biochem Pharmacol 43: Vaidya AB, Mather MW Atovaquone resistance in malaria parasites. Drug Resist Updat 3: Kessl JJ, Lange BB, Merbitz-Zahradnik T, Zwicker K, Hill P, Meunier B, Palsdottir H, Hunte C, Meshnick S, Trumpower BL Molecular basis for atovaquone binding to the cytochrome bc1 complex. J Biol Chem 278: Birth D, Kao WC, Hunte C Structural analysis of atovaquone-inhibited cytochrome bc1 complex reveals the molecular basis of antimalarial drug action. Nat Commun 5: Gingras BA, Jensen JB Antimalarial activity of azithromycin and erythromycin against Plasmodium berghei. Am J Trop Med Hyg 49: Wittner M, Lederman J, Tanowitz HB, Rosenbaum GS, Weiss LM Atovaquone in the treatment of Babesia microti infections in hamsters. Am J Trop Med Hyg 55: Krause PJ, Lepore T, Sikand VK, Gadbaw J, Jr., Burke G, Telford SR, 3rd, Brassard P, Pearl D, Azlanzadeh J, Christianson D, McGrath D, Spielman A Atovaquone and azithromycin for the treatment of babesiosis. N Engl J Med 343: Vannier E, Krause PJ Update on babesiosis. Interdiscip Perspect Infect Dis 2009: Birkenheuer AJ, Levy MG, Breitschwerdt EB Efficacy of combined atovaquone and azithromycin for therapy of chronic Babesia gibsoni (Asian genotype) infections in dogs. J Vet Intern Med 18: Di Cicco MF, Downey ME, Beeler E, Marr H, Cyrog P, Kidd L, Diniz PP, Cohn LA, Birkenheuer AJ Re-emergence of Babesia conradae and effective 35
54 treatment of infected dogs with atovaquone and azithromycin. Vet Parasitol 187: Sakuma M, Setoguchi A, Endo Y Possible emergence of drug-resistant variants of Babesia gibsoni in clinical cases treated with atovaquone and azithromycin. J Vet Intern Med 23: Siregar JE, Syafruddin D, Matsuoka H, Kita K, Marzuki S Mutation underlying resistance of Plasmodium berghei to atovaquone in the quinone binding domain 2 (Qo(2)) of the cytochrome b gene. Parasitol Int 57: Wormser GP, Prasad A, Neuhaus E, Joshi S, Nowakowski J, Nelson J, Mittleman A, Aguero-Rosenfeld M, Topal J, Krause PJ Emergence of resistance to azithromycin-atovaquone in immunocompromised patients with Babesia microti infection. Clin Infect Dis 50: Srivastava IK, Morrisey JM, Darrouzet E, Daldal F, Vaidya AB Resistance mutations reveal the atovaquone-binding domain of cytochrome b in malaria parasites. Mol Microbiol 33: Siregar JE, Kurisu G, Kobayashi T, Matsuzaki M, Sakamoto K, Mi-ichi F, Watanabe Y, Hirai M, Matsuoka H, Syafruddin D, Marzuki S, Kita K Direct evidence for the atovaquone action on the Plasmodium cytochrome bc1 complex. Parasitol Int 64: Brown HM, Berghaus RD, Latimer KS, Britt JO, Rakich PM, Peterson DS Genetic variability of Cytauxzoon felis from 88 infected domestic cats in Arkansas and Georgia. J Vet Diagn Invest 21: Salo AL, Stone E A survey of the views of US veterinary teaching faculty to owned cat housing practices. J Feline Med Surg doi: / x Loyd KA, Hernandez SM, Abernathy KJ, Shock BC, Marshall GJ Risk behaviours exhibited by free-roaming cats in a suburban US town. Vet Rec 173: Raghavan RK, Almes K, Goodin DG, Harrington JA, Jr., Stackhouse PW, Jr Spatially heterogeneous land cover/land use and climatic risk factors of tickborne feline cytauxzoonosis. Vector Borne Zoonotic Dis 14: Blagburn BL, Dryden MW Biology, treatment, and control of flea and tick infestations. Vet Clin North Am Small Anim Pract 39: , viii Boland LA, Angles JM Feline permethrin toxicity: retrospective study of 42 cases. J Feline Med Surg 12:
55 144. White WH, Plummer PR, Kemper CJ, Miller RJ, Davey RB, Kemp DH, Hughes S, Smith CK, 2nd, Gutierrez JA An in vitro larval immersion microassay for identifying and characterizing candidate acaricides. J Med Entomol 41: Brianti E, Pennisi MG, Brucato G, Risitano AL, Gaglio G, Lombardo G, Malara D, Fogliazza A, Giannetto S Efficacy of the fipronil 10%+(S)-methoprene 9% combination against Rhipicephalus sanguineus in naturally infested dogs: speed of kill, persistent efficacy on immature and adult stages and effect of water. Vet Parasitol 170: Fernandez-Salas A, Rodriguez-Vivas RI, Alonso-Diaz MA First report of a Rhipicephalus microplus tick population multi-resistant to acaricides and ivermectin in the Mexican tropics. Vet Parasitol 183: Rodriguez-Vivas RI, Perez-Cogollo LC, Rosado-Aguilar JA, Ojeda-Chi MM, Trinidad-Martinez I, Miller RJ, Li AY, de Leon AP, Guerrero F, Klafke G Rhipicephalus (Boophilus) microplus resistant to acaricides and ivermectin in cattle farms of Mexico. Rev Bras Parasitol Vet 23: Guerrero FD, Lovis L, Martins JR Acaricide resistance mechanisms in Rhipicephalus (Boophilus) microplus. Rev Bras Parasitol Vet 21: Miller RJ, Almazan C, Ortiz-Estrada M, Davey RB, George JE, De Leon AP First report of fipronil resistance in Rhipicephalus (Boophilus) microplus of Mexico. Vet Parasitol 191: Trout RT, Steelman CD, Szalanski AL Population genetics of Amblyomma americanum (Acari: Ixodidae) collected from Arkansas. J Med Entomol 47: Gates MC, Nolan TJ Factors influencing heartworm, flea, and tick preventative use in patients presenting to a veterinary teaching hospital. Prev Vet Med 93: Shkap V, de Vos AJ, Zweygarth E, Jongejan F Attenuated vaccines for tropical theileriosis, babesiosis and heartwater: the continuing necessity. Trends Parasitol 23: Florin-Christensen M, Suarez CE, Rodriguez AE, Flores DA, Schnittger L Vaccines against bovine babesiosis: where we are now and possible roads ahead. Parasitology doi: /s : Darghouth MA Review on the experience with live attenuated vaccines against tropical theileriosis in Tunisia: considerations for the present and implications for the future. Vaccine 26 Suppl 6:G4-G10. 37
56 155. Pipano E, Shkap V Vaccination against tropical theileriosis. Ann N Y Acad Sci 916: Yunker CE, Kuttler KL, Johnson LW Attenuation of Babesia bovis by in vitro cultivation. Vet Parasitol 24: Hall R, Ilhan T, Kirvar E, Wilkie G, Preston PM, Darghouth M, Somerville R, Adamson R Mechanism(s) of attenuation of Theileria annulata vaccine cell lines. Trop Med Int Health 4:A Halbroth BR, Draper SJ Recent developments in malaria vaccinology. Adv Parasitol 88: Carson CA, Timms P, Cowman AF, Stewart NP Babesia bovis: evidence for selection of subpopulations during attenuation. Exp Parasitol 70: De Vos AJ, Bock RE Vaccination against bovine babesiosis. Ann N Y Acad Sci 916: Rappuoli R Reverse vaccinology, a genome-based approach to vaccine development. Vaccine 19: Darghouth MA, Boulter NR, Gharbi M, Sassi L, Tait A, Hall R Vaccination of calves with an attenuated cell line of Theileria annulata and the sporozoite antigen SPAG-1 produces a synergistic effect. Vet Parasitol 142: Morrison WI, Goddeeris BM New approaches to immunization of cattle against Theileria parva. Dev Biol Stand 62: Srivastav A, Kass PH, McGill LD, Farver TB, Kent MS Comparative vaccine-specific and other injectable-specific risks of injection-site sarcomas in cats. J Am Vet Med Assoc 241: Paoletti E Applications of pox virus vectors to vaccination: an update. Proc Natl Acad Sci U S A 93: Tine JA, Lanar DE, Smith DM, Wellde BT, Schultheiss P, Ware LA, Kauffman EB, Wirtz RA, De Taisne C, Hui GS, Chang SP, Church P, Hollingdale MR, Kaslow DC, Hoffman S, Guito KP, Ballou WR, Sadoff JC, Paoletti E NYVAC-Pf7: a poxvirus-vectored, multiantigen, multistage vaccine candidate for Plasmodium falciparum malaria. Infect Immun 64: Hebben M, Duquesne V, Cronier J, Rossi B, Aubert A Modified vaccinia virus Ankara as a vaccine against feline coronavirus: immunogenicity and efficacy. J Feline Med Surg 6:
57 168. Tartaglia J, Jarrett O, Neil JC, Desmettre P, Paoletti E Protection of cats against feline leukemia virus by vaccination with a canarypox virus recombinant, ALVAC-FL. J Virol 67: Smooker PM, Rainczuk A, Kennedy N, Spithill TW DNA vaccines and their application against parasites--promise, limitations and potential solutions. Biotechnol Annu Rev 10: Williams JA, Carnes AE, Hodgson CP Plasmid DNA vaccine vector design: impact on efficacy, safety and upstream production. Biotechnol Adv 27: Talaat AM, Stemke-Hale K Expression library immunization: a road map for discovery of vaccines against infectious diseases. Infect Immun 73: Barry MA, Howell DP, Andersson HA, Chen JL, Singh RA Expression library immunization to discover and improve vaccine antigens. Immunol Rev 199: Shibui A, Nakae S, Watanabe J, Sato Y, Tolba ME, Doi J, Shiibashi T, Nogami S, Sugano S, Hozumi N Screening of novel malaria DNA vaccine candidates using full-length cdna library. Exp Parasitol 135: Alberti E, Acosta A, Sarmiento ME, Hidalgo C, Vidal T, Fachado A, Fonte L, Izquierdo L, Infante JF, Finlay CM, Sierra G Specific cellular and humoral immune response in Balb/c mice immunised with an expression genomic library of Trypanosoma cruzi. Vaccine 16: Piedrafita D, Xu D, Hunter D, Harrison RA, Liew FY Protective immune responses induced by vaccination with an expression genomic library of Leishmania major. J Immunol 163: Melby PC, Ogden GB, Flores HA, Zhao W, Geldmacher C, Biediger NM, Ahuja SK, Uranga J, Melendez M Identification of vaccine candidates for experimental visceral leishmaniasis by immunization with sequential fractions of a cdna expression library. Infect Immun 68: Smooker PM, Setiady YY, Rainczuk A, Spithill TW Expression library immunization protects mice against a challenge with virulent rodent malaria. Vaccine 18:
58 CHAPTER 2: Mitochondrial genome sequences and structures aid in the resolution of Piroplasmida phylogeny Megan E. Schreeg* 1, Henry S. Marr 1, Jaime L. Tarigo 1,2, Leah A. Cohn 3, David M. Bird 4, Elizabeth H. Scholl 4, Michael G. Levy 1, Brian M. Wiegmann #4, Adam J. Birkenheuer #1 1. North Carolina State University, College of Veterinary Medicine, Raleigh, NC 2. University of Georgia, College of Veterinary Medicine, Athens, GA 3. University of Missouri, College of Veterinary Medicine, Columbia, MO 4. North Carolina State University, College of Agriculture and Life Sciences, Raleigh, NC #Co-Corresponding Authors: Adam J. Birkenheuer and Brian M. Wiegmann *Responsible for all bench work and majority of writing and aided in phylogenetic tree construction ABSTRACT The taxonomy of the order Piroplasmida, which includes a number of clinically and economically relevant organisms, is a hotly debated topic amongst parasitologists. Three genera (Babesia, Theileria, and Cytauxzoon) are recognized based on parasite life cycle characteristics, but molecular phylogenetic analyses of 18S sequences have suggested the presence of five or more distinct Piroplasmida lineages. However, these studies are plagued with low statistical support, and little consensus exists on the taxonomic placement of many organisms, including C. felis, T. equi, and B. conradae. We hypothesized that mitochondrial genome sequences and structures would aid in the inference of Piroplasmida phylogeny. In this study, we have PCR amplified, sequenced, and annotated seven previously uncharacterized mitochondrial genomes (Babesia canis canis, Babesia canis vogeli, Babesia 40
59 canis, Babesia sp. Coco, Babesia conradae, Babesia microti-like sp./t. annae, and Cytauxzoon felis) and identified additional ribosomal fragments in ten previously characterized mitochondrial genomes. Phylogenetic analysis of concatenated mitochondrial and 18S sequence as well as COX1 amino acid sequence identified five distinct Piroplasmida groups, each of which possesses a unique mitochondrial genome structure. Although known biological characteristics of Piroplasmida corroborates the proposed phylogeny, more investigation into parasite life cycles is warranted to further understand the evolution of the Piroplasmida. Results of this study question the validity of currently recognized genera, and future efforts in the reclassification of Piroplasmida should include characterization of additional mitochondrial genomes. INTRODUCTION Parasites in the order Piroplasmida, which includes Babesia, Theileria, and Cytauxzoon species, cause important diseases across the globe in humans, livestock, wildlife, and companion animals (1-7). Despite the clinical and economic importance of these ticktransmitted parasites, the taxonomic relationships between many Piroplasmida remain ambiguous, which is problematic when attempting to understand and treat the diseases they cause. Classical taxonomy of Piroplasmida has been based on mechanisms of transmission in the tick host, host cell type(s) infected, and to a certain extent parasite morphology and vertebrate host preference (7-13). Theileria and Cytauxzoon species are limited to transstadial transmission in the tick and initially infect nucleated cells within the vertebrate host (7, 13, 14). Alternatively, Babesia species have acquired character traits that presumably enhance their propagation, including transovarial transmission in the tick and exclusive infection of erythrocytes in the vertebrate host (15). However, as more molecular and 41
60 biological information has been discovered, it has become apparent that this classification scheme is limited and fails to reflect the diversity and evolution of the Piroplasmida. Molecular data (DNA sequence) indicates that the currently recognized genus Babesia is polyphyletic (16-19). Consequently, Babesia species have been informally divided into Babesia sensu stricto (s.s.) and Babesia sensu latu (s.l.; 7). Babesia s.s. refers to Babesia as classically defined, and likely represents a true genus, as species within this group share both genetic and morphologic characteristics. Babesia s.l., however, refers to all species that morphologically resemble Babesia but that have since been found to be molecularly distinct from Babesia (7, 9, 11, 17, 18, 20-22). Babesia s.l. likely includes at least two subgroups: the Archaeopiroplasmida/Microti group, which includes the extensive Babesia microti complex (10, 11, 20), and the Prototheilerids/Duncani/Western group, which includes multiple organisms identified in the Western United States (10, 11, 17, 21, 22). However, while molecular phylogenetic analysis of a variety of targets (18S, ITS, Beta tubulin) indicates that the Babesia microti complex represents a distinct lineage, the true taxonomic placement of the Western Babesia species in relation to other Piroplasmida remains unsolved (Figure 1; 7, 9-11, 20, 23). Additionally, the majority of molecular phylogenetic analyses suggest that Babesia s.l. organisms are more primitive than Babesia s.s. and Theileria (7, 9-11, 23). However, with the exception of B. microti (24), primitive morphological features (invasion of nucleated cells) have not been detected for these organisms (21, 22), which further calls their taxonomy into question. Likewise, discrepancies between molecular and morphological data has complicated the classification of Theileria and Cytauxzoon species. Cytauxzoon was originally established as a distinct genus from Theileria due to its invasion of monocytes rather than 42
61 Figure 1. 18S sequence alone is unable to resolve phylogeny of the Piroplasmida. Cartoons (branch lengths not to scale) depict topology of phylogenetic trees of Piroplasmida based on 18S rrna sequences in four previously reported studies (7, 9-11). Nomenclature for sub-groupings assigned in each study are maintained in each individual tree. Despite using the same gene for analysis, the taxonomic placement for C. felis, T. equi, and B. conradae varies from tree to tree and often does not have high statistical support. Stars denote nodes that had less than 70% bootstrap support (A-C) or less than 95% Bayesian posterior probability (C-D). Individual species included in each respective study whose mitochondrial genomes were utilized in this study are noted within each clade. Mitochondrial genomes first characterized in this study are underlined. +Low support for species included within clade *Although Lack et al. found strong support for a node uniting Clade I-III, positioning of Clade II with respect to Clades I and III within this node was unresolved **White clades in 1C indicate those for which no representative species were characterized in this study; Clade IV includes Babesia benneti, while Clade VII include Babesia poelea. 43

62 A) Allsopp and Allsopp,

63 B) Criado-Fornelio et al.,

64 C) Lack et al.,

65 D) Schnittger et al.,
66 lymphocytes (14). However, species in both genera were later shown to infect both lymphocytes and monocytes (25-27), and consequently the majority of Cytauxzoon species were reclassified as Theileria (25, 28-33). Despite the suggestion to completely eliminate the genus Cytauxzoon (32), Cytauxzoon felis was never reclassified as Theileria, and for decades was the lone species recognized within the genus. Unfortunately, molecular data has only confused this situation further, as a number of novel parasites have been classified as Cytauxzoon on the basis of molecular data (18S rrna sequence) alone, despite there being no evidence of invasion of nucleated cells for any of these species (34-37). Additionally, phylogenetic analyses of 18S have failed to agree on a definitive taxonomic placement for Cytauxzoon species (Figure 1; 7, 9-11, 23). One species occasionally categorized with Cytauxzoon is Theileria equi, a species whose own taxonomy has been strongly debated (7-11, 23, 29, 38, 39). Originally named Babesia equi, T. equi was reclassified upon discovery of lymphocyte invasion (38), and has since been shown to infect monocytes as well (40). However, molecular phylogenetic analysis of T. equi 18S has failed to clarify whether this species belongs within a currently recognized group or represents a unique lineage (Figure 1; 7-11, 23). The lack of consensus for Piroplasmida taxonomy underscores the need for novel approaches in conducting molecular phylogenetic analyses of these organisms. The majority of previous analyses have used 18S rrna sequence to estimate phylogenetic relationships. However, the complexity of 18S secondary structure has led to inconsistencies in gene alignment across studies (41). As a result, current phylogenetic analyses of Piroplasmida have often been plagued with low statistical support, and as noted above, fail to agree on the taxonomic placement of many species (Figure 1; 7, 9-11, 23, 29). As an alternative to 18S, 48
67 mitochondrial genome sequences and structures have proven to be useful for the elucidation of evolutionary relationships and for delineating specimens to the species level, including closely related protozoan parasites (42-56). We hypothesize that mitochondrial genomes will similarly aid in our understanding of Piroplasmida phylogeny. In this study, we describe the sequence and structure of seven previously uncharacterized Piroplasmida mitochondrial genomes, including annotation of proteinencoding genes cytochrome b (cytb) and cytochrome c oxidase subunits I and III (cox1 and cox3) as well as ribosomal subunit fragments. Additionally, we have identified conserved ribosomal subunit fragment sequences from 10 previously reported Piroplasmida mitochondrial genomes. Phylogenetic analysis of these mitochondrial genome sequences concatenated with 18S sequences identified five distinct Piroplasmida lineages with relatively strong statistical support: 1) Babesia sensu stricto, 2) Theileria and Cytauxzoon, 3) Theileria equi, 4) Babesia conradae ( Western Babesia group), and 5) the Babesia microti group. These five groups, which can also be identified by analysis of COX1 amino acid sequence, are further supported by unique mitochondrial genome structures as well as known biological features of the organisms. These findings emphasize the need for reclassification of Piroplasmida species. MATERIALS AND METHODS Parasite species Mitochondrial genomes were characterized for seven Piroplasmida species that commonly infect companion animals (dogs and cats; Table 1). Blood samples previously confirmed to be infected with these parasites (18S amplification and sequencing) were readily available from the North Carolina State University College of Veterinary Medicine 49
68 Vector-Borne Disease Diagnostic Laboratory. One infected blood sample was utilized for each mitochondrial genome characterized. All additional parasite sequences utilized in this study are summarized in Table 1. Table 1. Species and sequences utilized in phylogenetic analysis. Species Current Categorization a MT Genome GenBank 18S GenBank Host Effected Accession Number Accession Number Babesia caballi Babesia sensu stricto Equine AB Z15104 Babesia bigemina Babesia sensu stricto Bovine AB HQ Babesia bovis Babesia sensu stricto Bovine AB AY Babesia canis canis Babesia sensu stricto Canine KC b AY Babesia canis rossi Babesia sensu stricto Canine KC b L19079 Babesia canis vogeli Babesia sensu stricto Canine KC b AY Babesia conradae Babesia sensu latu (Western group?) Canine KC b AF Babesia gibsoni Babesia sensu stricto Canine AB EU Babesia microti Babesia sensu latu (Microti group?) Murine, Human FO082868, AB c U09844 Babesia microti-like sp. (T. annae) Babesia sensu latu (Microti group?) Canine KC b AF Babesia rodhaini Babesia sensu latu (Microti group?) Murine AB M87656 Babesia sp. Coco Babesia sensu stricto Canine KC b EU Cytauxzoon felis Unclear: Theileria? Unique group? Feline KC b AY Theileria annulata Theileria Bovine NW M64243 Theileria equi Unclear: Theileria? Unique group? Equine AB EU Theileria orientalis Theileria Bovine AB HM Theileria parva Theileria Bovine AB L02366 Plasmodium falciparum N/A Human AY Z23263 a Names of sub-groups as previously defined in references (7) and (11) b Denotes mitochondrial genomes that were first characterized in this study c Two mitochondrial genomes that vary in sequence and structure have been reported for B. microti; both are utilized in this study DNA isolation DNA was extracted from 200 µl of anti-coagulated infected whole blood samples using a commercial kit according to manufacturer s instructions (QIAamp DNA Blood Mini Kit, Qiagen Inc., Valencia, CA). PCR amplification of mitochondrial genomes Using conserved regions of previously reported Piroplasmida mitochondrial genomes as a guide (52), primers were designed to PCR-amplify near-full length mitochondrial genomes of C. felis, B. canis canis, B. canis rossi, B. canis vogeli, B. conradae, and Babesia sp. Coco in three overlapping fragments (Table 2, see Figure S1 in Supplemental Material). Additional PCR assays were designed as needed for each species to obtain additional 50
69 mitochondrial genome sequence (see Tables S1-S6 and Figure S1-S3 in Supplemental Material). Each 50 µl reaction contained 1 µl of DNA template, 50 pmol of each primer, 10 nmol dntps, 75 nmol of MgCl2, 3.75 U AmpliTaq Gold DNA polymerase and a 1X concentration of GeneAmp PCR Gold Buffer (Applied Biosystems, Carlsbad, CA). Thermal cycling conditions consisted of an initial denaturation at 94 ºC for 5 minutes, followed by amplification cycles (94 ºC for 20 seconds, ºC for 30 seconds, and 68 ºC for minutes) and a final extension step at 72 ºC for 7 minutes (Techne Inc., Burlington, NJ). Annealing temperatures were optimized as needed utilizing a temperature gradient, and extension times and cycle number varied with amplicon length. Table 2. Primers utilized in PCR amplification of Piroplasmida mitochondrial genomes. Amplicon Forward Primer Reverse Primer MT Genome Fragment 1 a GGAAGTGGWACWGGWTGGAC ACTTTGAACACACTGCTCG MT Genome Fragment 2 a AGGCATGCAATACCGAACAGG AAGGTACGCCRGGGATAACAGG MT Genome Fragment 3 a AAGGTATGGTGAGACGACATGG CTTAACCCAACTCACGTACC cox1 b, c GGAAGTGGWACWGGWTGGAC TTCGGTATTGCATGCCTTG cytb b TTAGTGAAGGAACTTGACAGGT CGGTTAATCTTTCCTATTCCTTACG cox3 b ACTGTCAGCTAAAACGTATC ACAGGATTAGATACCCTGG cox3 (Babesia microti group) b CTCGATATTAATCTTAAAGTACAGGAC ACTCATATCTATTACCACTATAGGC a Primers were designed based on sequences of previously reported related Piroplasmida mitochondrial genomes. Three primer sets were utilized for the amplification of a near-full length mitochondrial genome for the majority of species (5 out of 7) characterized in this study. For additional primers used see Tables S1-S7 in Supplemental Material. b After sequencing of the mitochondrial genomes was complete, primers were designed in highly conserved regions for amplification of partial cox1 and full length cytb and cox3 in all species, and are recommended for amplification of these genes in future studies. c Recommended primer set for amplification of cox1 for phylogenetic analysis Sequences of the 5 end of the cox1 gene and mitochondrial telomeric regions were determined by inverted PCR (52). Primer pairs directed at terminal inverted repeats (TIR) would presumably self-anneal, leading to amplification of the remainder of the mitochondrial genome (see Figure S1 and Tables S1-S3 in Supplemental Material). A proofreading DNApolymerase with exonuclease activity (LA Taq) was used to remove any unpaired bases that 51
70 would interfere with the inverted PCR self-annealing. Each 50 µl PCR reaction contained 1 µl of DNA template, 50 pmol of each primer, 10 nmol dntps, 2.5 U LA Taq DNA polymerase and a 10X concentration of LA PCR Buffer II plus Mg +2 (Takara Bio Inc., Shiga, Japan). Thermal cycling conditions consisted of an initial denaturation at 95 ºC for 5 minutes, followed by 40 amplification cycles (94 ºC for 20 seconds and 68 ºC for 2.25 minutes) and a final extension step at 72 ºC for 7 minutes. Amplicons produced from the inverted PCR reactions (C. felis, B. canis canis, and B. canis rossi) were directly cloned according to manufacturer s instructions (pgem-t Easy vector system, Promega, San Luis Obispo, CA) and transformed into TOP-10 competent E. coli (Invitrogen, Grand Island, NY). Plasmids containing inserts of the appropriate size were isolated according to manufacturer s instructions (QIAprep Spin Miniprep Kit, Qiagen, Inc., Valencia, CA). Due to the unique mitochondrial genome structure of B. microti-like sp., an alternative PCR approach was required to amplify a near-full length mitochondrial genome (see Figure S4 and Table S7 in Supplemental Material). Primers were designed based on previously reported mitochondrial genome sequences for B. microti (FO and AB624353) and B. rodhaini (AB624357; 57, 58). The region of the mitochondrial genome containing cox1 and cytb was amplified in four overlapping fragments, while the region of the mitochondrial genome containing cox3 was amplified in two overlapping fragments. Each 50 µl reaction contained 1 µl of DNA template, 50 pmol of each primer, 10 nmol dntps, 75 nmol of MgCl2, 2.5 U AmpliTaq Gold DNA polymerase and a 1X concentration of GeneAmp PCR Gold Buffer (Applied Biosystems, Carlsbad, CA). Thermal cycling conditions consisted of an initial denaturation at 95 ºC for 5 minutes, followed by 45 amplification cycles (95 ºC for 20 seconds, ºC for 30 seconds, and 68 ºC for
71 minutes) and a final extension step at 72 ºC for 7 minutes; amplification of Fragment 4 required modified extension temperature due to high AT content of the amplicon (60 ºC during cyclic extension and 68 ºC during final extension; 59). Annealing temperatures were optimized as needed utilizing a temperature gradient, and extension times and cycle number varied with amplicon length. Because assumed internal inverted repeats (57, 60) were not relevant for phylogenetic analyses, amplification and sequencing of these regions were not pursued. Positive controls consisted of confirmed DNA extracted from Babesia gibsoniinfected canine whole blood and negative controls consisted of DNA extracted from uninfected canine whole blood and water (no DNA). For B. microti-like sp., no appropriate positive control was available. All amplicons were confirmed via electrophoresis on ethidium bromide-stained 1% agarose (Genesee Scientific, San Diego, CA). Amplification of B. conradae cox3-like gene Because a putative cox3 gene was not found in the mitochondrial genome of B. conradae, primers were designed based on highly conserved sequence flanking the cox3 gene in all Babesia, Theileria, and Cytauxzoon species referenced and examined in this paper (see Table S6 in Supplemental Material). A 700-bp amplicon was produced using PCR. Each 50 µl reaction contained 1 µl of DNA template, 50 pmol of each primer, 10 nmol dntps, 75 nmol of MgCl2, 2.5 U AmpliTaq Gold DNA polymerase and a 1X concentration of GeneAmp PCR Gold Buffer (Applied Biosystems, Carlsbad, CA). Thermal cycling conditions consisted of an initial denaturation at 94 ºC for 5 minutes, followed by 45 amplification cycles (94 ºC for 20 seconds, 50 ºC for 30 seconds, and 68 ºC for 1.5 minutes) and a final extension step at 72 ºC for 7 minutes. Positive and negative controls and 53
72 assessment of amplicons were identical to other mitochondrial PCRs; no amplicons were produced from DNA extracted from uninfected canine blood. Sequencing Purified amplicons (QIAquick PCR purification kit, Qiagen Inc., Valencia, CA) and plasmids were sequenced bi-directionally (MCLAB, South San Francisco, CA and Genewiz, South Plainfield, NJ). Additional primers were used for sequencing when necessary to obtain complete bi-directional sequence (see Tables S1-S7 in Supplemental Material). Sequence chromatograms were carefully inspected for heterogeneity, and contigs were assembled using the BioEdit Sequence Alignment Editor (North Carolina State University, Raleigh, NC). For those amplicons with sequence that could not be resolved directly (see Tables S1-S7 in Supplemental Material), PCR products were cloned using the pgem-t Easy vector system as described above. Genome annotation Protein-encoding genes (cox1, cox3, cytb) were identified by screening mitochondrial genomes for open reading frames. Putative genes were then queried against mitochondrial sequences of related parasites, and identified orthologues were aligned for confirmation. To identify putative rrna gene fragments, mitochondrial sequences were queried against previously reported rrna sequences from T. parva (Z23263; 61) using blastn under default algorithm parameters (NCBI BLAST). For identification of rrna fragments of B. microtilike sp., B. rodhaini, and B. microti, mitochondrial sequences for these species were further queried against rrna sequences of B. microti (AB624353) kindly provided by Kenji Hikosaka (51). Additionally, alignment of identified rrna sequences with previously annotated mitochondrial genomes was utilized in determining termini of rrna fragments as 54
73 needed (ClustalW, BioEdit Sequence Alignment Editor, North Carolina State University, Raleigh, NC). Alignment, substitution model choice and phylogenetic inference Alignments for mitochondrial and 18S sequences were carried out under standard configurations using ClustalW (BioEdit Sequence Alignment Editor, North Carolina State University, Raleigh, NC), and all sequences were cropped to the length of the shortest sequence in the alignment so as to prevent bias for nucleotides that were only amplified from select samples. Alignments were further edited, verified, and manually adjusted as needed in MEGA 6.05 (62). Protein coding genes were translated to amino acids to guide nucleotide alignment and to construct amino acid data sets. Highly variable regions where positional homology was uncertain or ad hoc, especially in ribosomal RNAs, were identified by inspection and excluded from phylogenetic analyses. The resulting alignment for each gene was concatenated using SequenceMatrix v (63); only those RNA fragments that were identified for all species were included (see Table S8 in Supplemental Material). Alignments and phylogenetic data sets are archived in the DRYAD public data repository ( Nucleotide substitution models were chosen by PartitionFinder (64) using the Bayesian Information Criterion (BIC). PartitionFinder was also used to find the best partitioning scheme based on BIC using the greedy option. For phylogenetic tree reconstruction, two optimality criteria were used: Maximum Likelihood (ML) and Bayesian Analysis (MB). For the maximum likelihood analysis, we used RaxML v8 (65) on concatenated datasets. Plasmodium falciparum was set as the outgroup and was used as the random seed value. No secondary structure options 55
74 were used. Tree inference was carried out under the GTRGAMMA model, and multiparametric bootstrapping was performed for iterations. For Bayesian analysis, we used MrBayes (66) with the MCMCMC (Metropolis coupled Markov Chain Monte Carlo) algorithm. According to results of likelihood ratio tests in PartitionFinder (64), seven unique partitions were identified and each was independently estimated under a specific site model. We carried out two simultaneous runs using the standard configuration with eight chains for every 108 generations, saving a tree every 1000 generations after discarding a burnin of 25%. Complete sampling was analyzed using ML criterion without partitioning. Clade support was assessed by examining Bayesian posterior probabilities (PP) from a post-burnin sample of optimal trees. ML Bootstrap percentages were compared to Bayesian posterior probabilities for each node to inform clade support. Resulting tree topologies were visualized in Figtree v1.4 (Rambaut and Drummond 2014, RESULTS Mitochondrial genome structures We present a detailed characterization of complete mitochondrial genome sequences from B. canis rossi, B. canis canis, and C. felis, and near-complete mitochondrial genome sequences from B. canis vogeli, B. conradae and Babesia sp. Coco, and B. microti-like sp. (T. annae). B. canis canis, B. canis rossi, B. canis vogeli, Babesia sp. Coco, and C. felis shared mitochondrial genome organization with previously reported Babesia sensu stricto and Theileria species (Figure 2A; 52). Linear mitochondrial genomes ranged in size from 5.6 to 5.9 kb, and included the protein-encoding genes cox1, cox3, and cytb, as well as multiple 56
 are indicated in white.")

75 Figure 2. Mitochondrial genome structures of Piroplasmida species characterized in this study. Genes shown above the central line are coded on the sense strand, while those below are on the antisense strand. Protein-coding genes (cox1, cox3, and cytb) are indicated in white. Large subunit rrna fragments are in light gray, small subunit rrna fragments are in dark gray, and miscellaneous conserved RNA fragments are in black. A) Mitochondrial genome sequences of C. felis, B. canis rossi, B. canis vogeli, B. canis canis, and Babesia sp. Coco maintained the mitochondrial genome structure that is characteristic of traditional Babesia sensu stricto and Theileria species, while B) the inferred Babesia microti-like sp. mitochondrial genome structure appears to be similar to that of B. microti and B. rodhaini, suggesting it has a flip-flop mitochondrial genome structure. Assumed inverted repeats A and B (indicated as IR-A and IR-B) were not confirmed due to lack of relevant sequence for phylogenetic analysis. C) Babesia conradae had a unique mitochondrial genome, which lacked cox3 and had a duplicated inversion that included the 3 end of cox1 and RNA17 and RNA18. Additionally, a collection of rrna fragments (RNA6, RNA7, RNA15, LSUC, and SSUF) found in Babesia sensu stricto, C. felis, and Theileria mitochondrial genomes was conserved but inverted as a unit adjacent to the duplicated inversion. 57
76 rrna fragments (Figure 2A). Similar to related species, terminal inverted repeats (TIR) were identified and characterized for B. canis rossi, B. canis canis, and C. felis (52). However, multiple attempts at inverted PCR were unsuccessful for B. canis vogeli and Babesia sp. Coco. Therefore, for these species the 5 end of cox1 could not be characterized and the presence or absence of TIRs could not be confirmed. Two separate pieces of the B. microti-like sp. mitochondrial genome were amplified that collectively comprised 5.9 kb (Figure 2B). Organization of protein-coding genes and rrna fragments on these fragments suggest that the B. microti-like sp. has a flip-flop mitochondrial genome structure similar to that of its close relative, B. microti (57, 60). Amplification spanning inverted repeats was not attempted due to lack of relevant sequence for phylogenetic analysis. In contrast, B. conradae had a novel mitochondrial genome structure (Figure 2C). While it had similar size (5.6 kb) and organization of cox1, cytb, and some rrna fragments as Babesia sensu stricto species, a cox3 gene was not identified at the predicted location within the mitochondrial genome. Instead, between cox1 and LSU1, there was an inverted duplication that included the 3 end of cox1 as well as RNA17 and RNA18 (Figure 2C). We attempted to identify a cox3 gene using primers matching RNA8 and RNA11 sequences, which flank cox3 in other Babesia s.s. and Theileria species (Figure 2A, Supplementary Table S6). This PCR produced a 700 base pair amplicon that contained a cox3-like sequence. The sequence (KF410591) had a cytochrome c oxidase characteristic heme-copper oxidase domain and shared 21-25% identity with Theileria, Babesia, and Plasmodium COX3 proteins (NCBI BLAST, blastx; Simple Modular Architecture Research Tool; 67, 68). The 58
77 Figure 3. Phylogenetic analysis of concatenated mitochondrial genome and 18S nucleotide sequence identifies five distinct lineages within Piroplasmida. Statistical support for clades are indicated at each node: posterior probabilities from Bayesian analysis (10 million generations of Markov chain Monte Carlo) are listed above nodes, while bootstrap values from Maximum Likelihood analysis (200 bootstrap replicates for A, 100 bootstrap replicates for B-C) are listed below nodes. Trees are drawn to scale, with branch lengths measured in the number of substitutions per site. A) Analysis of concatenated mitochondrial and 18S nucleotide sequences (6006 total characters) produce a robust tree with high statistical support of topology; see Table S8 for specific sequences included in analysis. Analysis of concatenated mitochondrial and 18S nucleotide sequences with cox3 sequences excluded (B; 5292 total characters) and mitochondrial nucleotide sequence alone (C; 4395 total characters) produced the same topology as A, but had less statistical support for some nodes, particularly the placement of T. equi and to a lesser extent the placement of B. conradae and C. felis. Maximum Likelihood analysis of mitochondrial sequences alone did not recover Group 3 (T. equi) as a distinct clade, which is denoted with an asterisk (*). The five lineages identified by analysis of concatenated mitochondrial and 18S nucleotide sequences are depicted in D (branch lengths not to scale). 59
78 60

79 61
80 location of this cox3-like gene with respect to the mitochondrial genome of B. conradae remains unknown. Phylogenetic relationships Phylogenetic analyses of all available mitochondrial sequences and 18S yielded a tree topology with strong statistical support at all nodes (Figure 3A). Many of the clades recovered (e.g. Babesia sensu stricto, Babesia conradae, Babesia microti group) correspond well with groups described previously (Figure 1). In contrast, Cytauxzoon felis was grouped in a clade with other Theileria species, while Theileria equi was placed as a sister group to the clade containing Babesia sensu stricto, Cytauxzoon felis, and other Theileria spp. Because it is unclear whether cox3-like sequences of T. equi and B. conradae are true cox3 orthologues, cox3 sequences were excluded from analysis, resulting in an identical topology that had less statistical support for the placement of T. equi (Figure 3B). When mitochondrial sequences (excluding cox3) were analyzed alone, the true placement of T. equi became unclear, and statistical support for the placement of B. conradae also decreased (Figure 3C). Although bootstrap support for the placement of C. felis decreased as less sequences were analyzed, the more extensive and stringent Bayesian analysis consistently supported this placement (Figure 3B-3C). All analyses strongly support that organisms in the B. microti group diverged at an early time point from the rest of the Piroplasmida (Figure 3A-3C). Therefore, our analysis supports the presence of five distinct lineages, including the Babesia microti group, Babesia sensu stricto, Theileria, B. conradae, and T. equi, although the precise placement of the latter group remains unclear (Figure 3D). This phylogeny is further supported by the unique mitochondrial genome structures (Figure 4) and known biological traits (Figure 5) of each of the five proposed groups. 62

81 Individual mitochondrial genes were also assessed to see if phylogenetic analysis of shorter, more easily obtained sequences could recapitulate results shown in Figure 3. Analysis of COX1 putative amino acid sequence yielded a similar topology, although interestingly, this analysis had strong statistical support for the divergence of T. equi prior to B. conradae (Figure 6). Analyses of other genes yielded tree topologies that were dissimilar to analysis of concatenated mitochondrial and 18S sequences, and often had low statistical support at deep nodes (see Figures S5-S9 in Supplemental Material). Figure 4. Mitochondrial genome structures further support recognition of the five groups identified by phylogenetic analysis of concatenated mitochondrial and 18S sequences. Genes are indicated with names (cox1, cox3, cytb) and ribosomal sequences are indicated in gray with L for large subunit and S for small subunit. Genes placed above the black central line on the diagram are coded on the sense strand of DNA, while those below the line are coded on the anti-sense strand. The presence or absence of TIRs in B. conradae s mitochondrial genome has not been confirmed. Branches not drawn to scale. 63
82 DISCUSSION In this study, we have utilized mitochondrial genome sequences and structures to infer the phylogeny of organisms in the order Piroplasmida. Analysis of the concatenated sequences of all identified mitochondrial genes with 18S gene sequence supported the existence of five distinct lineages of Piroplasmida, although the exact taxonomic location of the group consisting of T. equi remains unclear. Recognition of these five groups is further supported by mitochondrial genome structure and organism features, and the groups can be molecularly distinguished from one another through phylogenetic analysis of COX1 amino acid sequence alone. These results have a number of implications for the current organization of Piroplasmida, and raise further questions regarding the evolutionary history of a number of species. Although Babesia organisms have been informally divided into species exhibiting classical Babesia traits (Babesia sensu stricto) and those that don t (Babesia sensu latu), mitochondrial genome sequences and structures indicate Babesia encompasses at least 3 distinct groups, which we refer to as the Babesia microti group, the Western Babesia group, and Babesia sensu stricto. Analysis of mitochondrial genome sequences and structures, despite including only four representatives from the clade, clearly indicates that the group we call the Babesia microti group represents a distinct lineage that diverged long ago from all other Piroplasmida (Figure 3-4, Figure 6). This group includes the Babesia microti-like sp. found in Spanish dogs, whose mitochondrial genome was first characterized in this study. Although this organism has been renamed a number of times (Theileria annae, Babesia cf. microti, and most recently Babesia vulpes), this analysis clearly indicates that it should be 64
83 Figure 5. Biology of Piroplasmida organisms is consistent with phylogeny inferred from analysis of concatenated mitochondrial and 18S sequences. A) Organisms in Babesia sensu stricto (red) do not infect leukocytes and can be transmitted transovarially in the tick host, two traits unique to the group. All other Piroplasmida (gray) are thought to be limited to transstadial transmission in the tick host and infect leukocytes prior to erythrocytes. Notably, details regarding tick hosts, transmission in the tick, and infection of nucleated cells for Group 4 remains unknown (noted by question mark), and infection of nucleated host cells has only been demonstrated for a single species in Group 5 (24). Characteristics of species in Groups 2 and 3 (outlined with dashed red line) are further summarized in B. B) While many organisms in Group 2 and 3 have been demonstrated to infect leukocytes, the specific leukocyte infected isn t clade-specific and hasn t even been confirmed for some species (e.g., Cytauxzoon felis). Additionally, the shared biological features of organisms in Group 2 support their distinction from the organism in Group 3, T. equi. T. equi exclusively infects equine hosts, and disease is caused by parasite infection of erythrocytes rather than the brief schizogonous phase in leukocytes. However, there is evidence indicating that organisms in Group 2 have evolved more complex methods of host leukocyte manipulation. Species within Group 2 that diverged earliest (Cytauxzoon felis) exclusively infect carnivores and have grossly enlarged schizont-infected cells, which suggests a blocking of host cell apoptosis. The remaining Theileria species in Group 2 exclusively infect ruminants. Organisms in the next clade to diverge in Group 2, including Theileria orientalis, also have grossly enlarged schizont-infected cells. This group is commonly known as the non-transforming Theileria species. This is in contrast to the transforming Theileria species, which includes T. annulata and parva. Species in this clade have perfected host cell manipulation, and reversible transform infected host leukocytes into a proliferative neoplastic state, which supports the replicating parasite. Branches not drawn to scale. 65
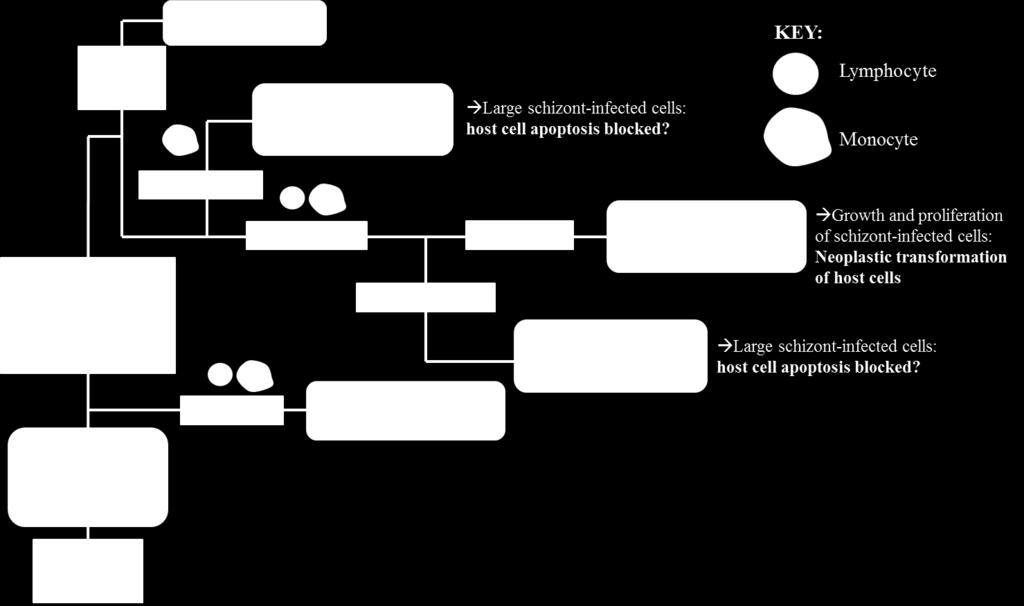

84 A B? 66
85 categorized in the Babesia microti group (69). Furthermore, this analysis underscores the fact that this group is not only distinct, but also highly divergent from other Piroplasmida (Figure 3, Figure 6). The marked diversity of organisms in the Babesia microti group also confirms its autonomy. More extensively than any other Piroplasmida clade, organisms in the Babesia microti group have adaptively radiated to infect a wide variety of hosts in a number of niches across the globe (20, 69, 70). This large group likely consists of multiple subgroups, and although our study lacks enough species to properly address topology within the Babesia microti group, our analysis concurs with studies that subdivide Babesia microti organisms from Babesia rodhaini (Figure 3, Figure 6; 20, 69). Despite the divergent nature of this group and implication that it represents an ancient lineage, infection of nucleated cells in the vertebrate host has been demonstrated for only a single species in the Babesia microti group (Figure 5; 24). Thus, it remains unclear whether invasion of nucleated cells is a primitive trait that has not been identified for other species in this group, or if it is a character trait that has been gained and then lost in the evolution of the Piroplasmida. Regardless, it is clear that the Babesia microti group represents a unique lineage, and a new name should be accordingly assigned to this group to distinguish it from Babesia. Similarly, analysis of concatenated mitochondrial genome and 18S sequence grouped Babesia conradae as a distinct lineage (Figure 3A-3B), which we have named the Western Babesia group. Statistical support for this clade decreased when mitochondrial nucleotide or COX1 amino acid sequence alone was analyzed (Figure 3C, Figure 6), but this can likely be attributed to the fact that only a single species from this group was represented in analysis. We hypothesize that as more mitochondrial genomes are characterized for species in this group (e.g. Babesia duncani/wa1, Babesia lengau, Babesia behnkei), statistical support will 67
86 increase and confirm this taxonomic location. Furthermore, Babesia conradae had a novel mitochondrial genome structure that lacked a cox3 gene, and instead had an inverted repeat of an adjacent region of the mitochondrial genome (Figure 2, Figure 4). The purpose of this sequence is unclear, but it may play a role in facilitating recombination during mitochondrial genome replication like the internal repeats (IRs) found in the Babesia microti group (57). While this unique feature supports the notion of the Western Babesia group being distinguished from Babesia, the scarcity of information available for this group make it impossible to corroborate molecular analyses with biological data. Although our analysis and others (Figure 1) suggest this is a relatively older lineage that diverged first after the Figure 6. Phylogenetic analysis of COX1 amino acid sequence recovers the same five Piroplasmida groups as concatenated mitochondrial and 18S nucleotide sequences. Statistical support for clades are indicated at each node: posterior probabilities from Bayesian analysis (10 million generations of Markov chain Monte Carlo) are listed above nodes, while bootstrap values from Maximum Likelihood analysis (100 bootstrap replicates) are listed below nodes. Tree is drawn to scale, with branch lengths measured in the number of substitutions per site. Analysis of only 429 characters produced a tree with strong support for most branches, although recovery of B. conradae was not as highly supported by Maximum Likelihood analysis. 68
87 Babesia microti group, organisms in this group have only been recognized within the last few decades (21, 71-75). Originally thought to be localized in the Western United States, the discovery of Babesia lengau and lengau-like species in Africa and Europe verified that organisms in this clade are distributed worldwide (72, 73, 75). Currently described vertebrate hosts include ungulates, humans, and carnivores, while tick vectors remain unknown (21, 71-75). Disease severity varies between species, but is always due to intraerythrocytic organisms, as no intraleukocytic schizonts have been identified (Figure 5, 21, 22). Clearly, more studies are needed to fully understand the evolution of this group and to verify its taxonomy, including characterization of more mitochondrial genomes and most importantly, more extensive investigation into the ability of the organisms to invade nucleated cells. Importantly Babesia conradae was easily distinguished from Babesia sensu stricto in this study (Figure 3). When first discovered, Babesia conradae was identified as Babesia gibsoni USA, a nomenclature that persisted for years and continues to cause confusion in the literature today (74). However, Babesia sensu stricto, which includes the real Babesia gibsoni, clearly represents a distinct group as evidenced by both molecular and biological data (Figure 3-5). Previous studies have further subdivided this group into species that infect ungulates and species that infect carnivores (10). While our limited analysis supports this observation, we refrain from recognizing these subgroups, as evidence is mounting to suggest that Babesia sensu stricto species can infect multiple vertebrate hosts (1, 70). Despite this, it is interesting to note that Babesia sp. Coco has only been identified in domestic dogs, yet is taxonomically grouped with species that traditionally infect ungulates. Because Babesia sp. Coco has only been identified in immunocompromised dogs (76, 77), 69
88 we speculate that carnivores are not the natural host and that wild ungulates should be considered when searching for a reservoir host. Analysis of mitochondrial genomes also indicates that reorganization of the genera currently referred to as Theileria and Cytauxzoon should be considered. In contrast with phylogenetic analyses of 18S alone (Figure 1; 7, 9-11, 29), our analysis of concatenated mitochondrial and 18S sequences supports the placement of Cytauxzoon felis within the Theileria clade while excluding Theileria equi (Figure 3). A close taxonomic relationship between Cytauxzoon and these Theileria species is further supported by shared mitochondrial genome structures (Figure 4) as well as the unique biological features shared between the organisms (Figure 5). While previous classification schemes of Piroplasmida proposed recognizing groups based on the specific leukocyte infected (i.e., Cytauxzoon vs. Theileria), the discovery of Piroplasmida species that infect multiple leukocyte lineages indicates that such an approach is invalid (Figure 5; 25-28, 40, 78). Alternatively, we propose that different Piroplasmida lineages may be better defined by the extent of their interaction with host cells and the strategies they employed to enhance their propagation. For instance, in contrast to all other Piroplasmida lineages recovered in this study, evidence of advanced host leukocyte manipulation has been observed in representative organisms from all three branches within the Theileria/Cytauxzoon clade (Figure 5). While the most recently diverged Theileria species have perfected this strategy by transforming host cells into a neoplastic-like state, the older non-transforming Theileria and Cytauxzoon lineages show evidence of rudimentary blocking of host cell apoptosis by the presence of grossly enlarged schizont-infected leukocytes (78-81). Notably, these features have not been described for all species in this clade, including the newly discovered Cytauxzoon species in Europe, Asia, 70
89 and Africa (34-37). However, the extent of interaction between host and parasite is likely host-specific, as evidenced by the more limited schizogony that occurs in C. felis-infected bobcats compared to in cats (82). Thus, it is likely that other Cytauxzoon species infect leukocytes but that the stage isn t recognized due to its transience in parasite-adapted hosts. Additionally, in our phylogenetic analysis of concatenated mitochondrial and 18S sequences, Theileria equi was recovered as a unique lineage that is a sister group to the clade containing Babesia sensu stricto and all other Theileria and Cytauxzoon species. However, of the five groups identified by our analysis, T. equi had the lowest statistical support (Figure 3), and hence the definitive taxonomic placement of this organism with respect to Theileria/Cytauxzoon remains unclear. Nevertheless, despite lacking robust statistical support from phylogenetic analysis, the radically divergent mitochondrial genome structure (Figure 4), lack of evidence for host leukocyte manipulation (Figure 5), and results of previous phylogenetic analyses of 150 proteins (39) supports the notion that T. equi is distinct from both Theileria and Babesia. Further mitochondrial genome characterization for other Piroplasmida proposed to be closely related to T. equi, including Babesia bicornis (7), will likely aid in definitively resolving the taxonomy of T. equi. Although we would recommend characterization of the full mitochondrial genome sequence and structure in addition to 18S for inferring phylogeny of Piroplasmida, this strategy may be cumbersome if rapid identification of novel Piroplasmida species is desired. Phylogenetic analysis of partial COX1 amino acid sequence was able to recover all five groups identified in this study with relatively strong statistical support, albeit with a slightly altered topology (Figure 6). Therefore, in instances where analysis of the complete 71
90 mitochondrial genome is impractical, amplification of cox1 using primers listed in Table 2 is recommended. In conclusion, in this study we have characterized seven new Piroplasmida mitochondrial genomes, one of which (B. conradae) possessed a novel arrangement of mitochondrial genes. When analyzing concatenated mitochondrial gene and 18S nucleotide sequences, we recovered five distinct groups of Piroplasmida species. While the phylogeny derived from this analysis is further supported by both mitochondrial genome structures and known biological features of the groups, more work needs to be done in characterizing the latter for many Piroplasmida species. Future phylogenetic studies of Piroplasmida should include mitochondrial genome sequences and structures, or at the very least COX1 sequence. Importantly, the results of this study question the validity of currently recognized Piroplasmida genera, and call for additional analyses and subsequent reclassification of Piroplasmida. ACKNOWLEDGEMENTS The authors thank Karen Gore for her technical assistance. This research was funded by a charitable organization which wishes to remain anonymous. There were no conflicts of interest in this study. REFERENCES 1. Solano-Gallego L, Baneth G Babesiosis in dogs and cats--expanding parasitological and clinical spectra. Veterinary Parasitology 181: Meinkoth JH, Kocan AA Feline cytauxzoonosis. Vet Clin North Am Small Anim Pract 35:89-101, vi. 72
91 3. Bishop R, Musoke A, Morzaria S, Gardner M, Nene V Theileria: intracellular protozoan parasites of wild and domestic ruminants transmitted by ixodid ticks. Parasitology 129 Suppl:S Gohil S, Herrmann S, Gunther S, Cooke BM Bovine babesiosis in the 21st century: advances in biology and functional genomics. Int J Parasitol 43: Wise LN, Kappmeyer LS, Mealey RH, Knowles DP Review of equine piroplasmosis. J Vet Intern Med 27: Vannier EG, Diuk-Wasser MA, Ben Mamoun C, Krause PJ Babesiosis. Infect Dis Clin North Am 29: Schnittger L, Rodriguez AE, Florin-Christensen M, Morrison DA Babesia: A world emerging. Infect Genet Evol 12: Allsopp MT, Cavalier-Smith T, De Waal DT, Allsopp BA Phylogeny and evolution of the piroplasms. Parasitology 108 ( Pt 2): Allsopp MT, Allsopp BA Molecular sequence evidence for the reclassification of some Babesia species. Annals of the New York Academy of Sciences 1081: Criado-Fornelio A, Martinez-Marcos A, Buling-Sarana A, Barba-Carretero JC Molecular studies on Babesia, Theileria and Hepatozoon in southern Europe. Part II. Phylogenetic analysis and evolutionary history. Veterinary Parasitology 114: Lack JB, Reichard MV, Van Den Bussche RA Phylogeny and evolution of the Piroplasmida as inferred from 18S rrna sequences. International Journal for Parasitology 42: Irwin PJ Canine babesiosis: from molecular taxonomy to control. Parasites & Vectors 2 Suppl 1:S Sivakumar T, Hayashida K, Sugimoto C, Yokoyama N Evolution and genetic diversity of Theileria. Infect Genet Evol 27: Neitz WO, Thomas AD Cytauxzoon sylvicaprae gen. nov., spec. nov., a protozoon responsible for a hitherto undescribed disease in the duiker, Sylvicapra grimmia (Linne). Onderstepoort J Vet Sci Anim Ind 23: Chauvin A, Moreau E, Bonnet S, Plantard O, Malandrin L Babesia and its hosts: adaptation to long-lasting interactions as a way to achieve efficient transmission. Vet Res 40:37. 73
92 16. Kjemtrup AM, Kocan AA, Whitworth L, Meinkoth J, Birkenheuer AJ, Cummings J, Boudreaux MK, Stockham SL, Irizarry-Rovira A, Conrad PA There are at least three genetically distinct small piroplasms from dogs. Int J Parasitol 30: Kjemtrup AM, Thomford J, Robinson T, Conrad PA Phylogenetic relationships of human and wildlife piroplasm isolates in the western United States inferred from the 18S nuclear small subunit RNA gene. Parasitology 120 ( Pt 5): Zahler M, Rinder H, Gothe R Genotypic status of Babesia microti within the piroplasms. Parasitol Res 86: Zahler M, Rinder H, Zweygarth E, Fukata T, Maede Y, Schein E, Gothe R 'Babesia gibsoni' of dogs from North America and Asia belong to different species. Parasitology 120 ( Pt 4): Goethert HK, Telford SR, 3rd What is Babesia microti? Parasitology 127: Conrad PA, Kjemtrup AM, Carreno RA, Thomford J, Wainwright K, Eberhard M, Quick R, Telford SR, 3rd, Herwaldt BL Description of Babesia duncani n.sp. (Apicomplexa: Babesiidae) from humans and its differentiation from other piroplasms. Int J Parasitol 36: Kjemtrup AM, Wainwright K, Miller M, Penzhorn BL, Carreno RA Babesia conradae, sp. Nov., a small canine Babesia identified in California. Vet Parasitol 138: Reichard MV, Van Den Bussche RA, Meinkoth JH, Hoover JP, Kocan AA A new species of Cytauxzoon from Pallas' cats caught in Mongolia and comments on the systematics and taxonomy of piroplasmids. J Parasitol 91: Mehlhorn H, Raether W, Schein E, Weber M, Uphoff M [Light and electron microscopy studies of the developmental cycle and effect of pentamidine on the morphology of intra-erythrocyte stages of Babesia microti]. Dtsch Tierarztl Wochenschr 93: Grootenhuis JG, Young AS, Dolan TT, Stagg DA Characteristics of Theileria species (eland) infections in eland and cattle. Res Vet Sci 27: Spooner RL, Innes EA, Glass EJ, Brown CG Theileria annulata and T. parva infect and transform different bovine mononuclear cells. Immunology 66:
93 27. Spooner RL, Innes EA, Glass EJ, Millar P, Brown CG Bovine mononuclear cell lines transformed by Theileria parva or Theileria annulata express different subpopulation markers. Parasite Immunol 10: Young AS, Grootenhuis JG, Leitch BL, Schein E The development of Theileria=Cytauxzoon taurotragi (Martin and Brocklesby, 1960) from eland in its tick vector Rhipicephalus appendiculatus. Parasitology 81: Nijhof AM, Pillay V, Steyl J, Prozesky L, Stoltsz WH, Lawrence JA, Penzhorn BL, Jongejan F Molecular characterization of Theileria species associated with mortality in four species of African antelopes. J Clin Microbiol 43: Thomas SE, Wilson DE, Mason TE Babesia, Theileria and Anaplasma spp. infecting sable antelope, Hippotragus niger (Harris, 1838), in southern Africa. Onderstepoort J Vet Res 49: Uilenberg G, Franssen FF, Perie NM Relationships between Cytauxzoon felis and African piroplasmids. Vet Parasitol 26: Levine ND The protozoan phylum Apicomplexa. CRC Press, Boca Raton, Fla. 33. Levine ND Taxonomy of the Piroplasms. Transactions of the American Microscopal Society 90: Ketz-Riley CJ, Reichard MV, Van den Bussche RA, Hoover JP, Meinkoth J, Kocan AA An intraerythrocytic small piroplasm in wild-caught Pallas's cats (Otocolobus manul) from Mongolia. J Wildl Dis 39: Millan J, Naranjo V, Rodriguez A, de la Lastra JM, Mangold AJ, de la Fuente J Prevalence of infection and 18S rrna gene sequences of Cytauxzoon species in Iberian lynx (Lynx pardinus) in Spain. Parasitology 134: Carli E, Trotta M, Chinelli R, Drigo M, Sinigoi L, Tosolini P, Furlanello T, Millotti A, Caldin M, Solano-Gallego L Cytauxzoon sp. infection in the first endemic focus described in domestic cats in Europe. Vet Parasitol 183: Leclaire S, Menard S, Berry A Molecular characterization of Babesia and Cytauxzoon species in wild South-African meerkats. Parasitology 142: Mehlhorn H, Schein E Redescription of Babesia equi Laveran, 1901 as Theileria equi Mehlhorn, Schein Parasitol Res 84: Kappmeyer LS, Thiagarajan M, Herndon DR, Ramsay JD, Caler E, Djikeng A, Gillespie JJ, Lau AO, Roalson EH, Silva JC, Silva MG, Suarez CE, Ueti MW, Nene VM, Mealey RH, Knowles DP, Brayton KA Comparative genomic analysis and phylogenetic position of Theileria equi. BMC Genomics 13:
94 40. Ramsay JD, Ueti MW, Johnson WC, Scoles GA, Knowles DP, Mealey RH Lymphocytes and macrophages are infected by Theileria equi, but T cells and B cells are not required to establish infection in vivo. PLoS One 8:e Morrison DA, Ellis JT Effects of nucleotide sequence alignment on phylogeny estimation: a case study of 18S rdnas of apicomplexa. Mol Biol Evol 14: Moritz C, Dowling, T. E., Brown, W. M Evolution of animal mitochondrial DNA: Relevance for population biology and systematics. Annual Review of Ecology and Systematics 18: Gou H, Guan G, Liu A, Ma M, Xu Z, Liu Z, Ren Q, Li Y, Yang J, Chen Z, Yin H, Luo J A DNA barcode for Piroplasmea. Acta Trop 124: He L, Zhang Y, Zhang QL, Zhang WJ, Feng HH, Khan MK, Hu M, Zhou YQ, Zhao JL Mitochondrial genome of Babesia orientalis, apicomplexan parasite of water buffalo (Bubalus babalis, Linnaeus, 1758) endemic in China. Parasit Vectors 7: Boore JL Animal mitochondrial genomes. Nucleic Acids Res 27: Sankoff D, Leduc G, Antoine N, Paquin B, Lang BF, Cedergren R Gene order comparisons for phylogenetic inference: evolution of the mitochondrial genome. Proc Natl Acad Sci U S A 89: Kher CP, Doerder FP, Cooper J, Ikonomi P, Achilles-Day U, Kupper FC, Lynn DH Barcoding Tetrahymena: discriminating species and identifying unknowns using the cytochrome c oxidase subunit I (cox-1) barcode. Protist 162: Chantangsi C, Lynn DH Phylogenetic relationships within the genus Tetrahymena inferred from the cytochrome c oxidase subunit 1 and the small subunit ribosomal RNA genes. Mol Phylogenet Evol 49: Le TH, Blair D, McManus DP Mitochondrial genomes of human helminths and their use as markers in population genetics and phylogeny. Acta Trop 77: Ogedengbe JD, Hanner RH, Barta JR DNA barcoding identifies Eimeria species and contributes to the phylogenetics of coccidian parasites (Eimeriorina, Apicomplexa, Alveolata). International Journal for Parasitology 41: Hikosaka K, Kita K, Tanabe K Diversity of mitochondrial genome structure in the phylum Apicomplexa. Mol Biochem Parasitol 188:
95 52. Hikosaka K, Watanabe Y, Tsuji N, Kita K, Kishine H, Arisue N, Palacpac NM, Kawazu S, Sawai H, Horii T, Igarashi I, Tanabe K Divergence of the mitochondrial genome structure in the apicomplexan parasites, Babesia and Theileria. Mol Biol Evol 27: Savolainen V, Cowan RS, Vogler AP, Roderick GK, Lane R Towards writing the encyclopedia of life: an introduction to DNA barcoding. Philosophical Transactions of the Royal Society of London Series B, Biological sciences 360: Tian Z, Luo J, Zheng J, Xie J, Shen H, Yin H, Luo J, Tian M, Yuan X, Wang F, Liu G Phylogenetic analysis of Babesia species in China based on cytochrome b (COB) gene. Infect Genet Evol doi: /j.meegid Vaidya AB, Mather MW Mitochondrial evolution and functions in malaria parasites. Annu Rev Microbiol 63: Wilson RJ, Williamson DH Extrachromosomal DNA in the Apicomplexa. Microbiol Mol Biol Rev 61: Hikosaka K, Tsuji N, Watanabe YI, Kishine H, Horii T, Igarashi I, Kita K, Tanabe K Novel type of linear mitochondrial genomes with dual flip-flop inversion system in apicomplexan parasites, Babesia microti and Babesia rodhaini. BMC Genomics 13: Cornillot E, Hadj-Kaddour K, Dassouli A, Noel B, Ranwez V, Vacherie B, Augagneur Y, Bres V, Duclos A, Randazzo S, Carcy B, Debierre-Grockiego F, Delbecq S, Moubri-Menage K, Shams-Eldin H, Usmani-Brown S, Bringaud F, Wincker P, Vivares CP, Schwarz RT, Schetters TP, Krause PJ, Gorenflot A, Berry V, Barbe V, Ben Mamoun C Sequencing of the smallest Apicomplexan genome from the human pathogen Babesia microti. Nucleic Acids Res 40: Su XZ, Wu Y, Sifri CD, Wellems TE Reduced extension temperatures required for PCR amplification of extremely A+T-rich DNA. Nucleic Acids Res 24: Cornillot E, Dassouli A, Garg A, Pachikara N, Randazzo S, Depoix D, Carcy B, Delbecq S, Frutos R, Silva JC, Sutton R, Krause PJ, Mamoun CB Whole genome mapping and re-organization of the nuclear and mitochondrial genomes of Babesia microti isolates. PLoS One 8:e Feagin JE, Harrell MI, Lee JC, Coe KJ, Sands BH, Cannone JJ, Tami G, Schnare MN, Gutell RR The fragmented mitochondrial ribosomal RNAs of Plasmodium falciparum. PLoS One 7:e
96 62. Tamura K, Stecher G, Peterson D, Filipski A, Kumar S MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol 30: Vaidya G, Lohman, D. J., Meier, R SequenceMatrix: concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 27: Lanfear R, Calcott B, Ho SY, Guindon S Partitionfinder: combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol Biol Evol 29: Stamatakis A RAxML version 8: a tool for phylogenetic analysis and postanalysis of large phylogenies. Bioinformatics 30: Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Hohna S, Larget B, Liu L, Suchard MA, Huelsenbeck JP MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol 61: Letunic I, Doerks T, Bork P SMART 7: recent updates to the protein domain annotation resource. Nucleic Acids Res 40:D Schultz J, Milpetz F, Bork P, Ponting CP SMART, a simple modular architecture research tool: identification of signaling domains. Proc Natl Acad Sci U S A 95: Baneth G, Florin-Christensen M, Cardoso L, Schnittger L Reclassification of Theileria annae as Babesia vulpes sp. nov. Parasit Vectors 8: Yabsley MJ, Shock BC Natural history of Zoonotic Babesia: Role of wildlife reservoirs. Int J Parasitol Parasites Wildl 2: Quick RE, Herwaldt BL, Thomford JW, Garnett ME, Eberhard ML, Wilson M, Spach DH, Dickerson JW, Telford SR, 3rd, Steingart KR, Pollock R, Persing DH, Kobayashi JM, Juranek DD, Conrad PA Babesiosis in Washington State: a new species of Babesia? Ann Intern Med 119: Bajer A, Alsarraf M, Bednarska M, Mohallal EM, Mierzejewska EJ, Behnke- Borowczyk J, Zalat S, Gilbert F, Welc-Faleciak R Babesia behnkei sp. nov., a novel Babesia species infecting isolated populations of Wagner's gerbil, Dipodillus dasyurus, from the Sinai Mountains, Egypt. Parasit Vectors 7: Bosman AM, Oosthuizen MC, Peirce MA, Venter EH, Penzhorn BL Babesia lengau sp. nov., a novel Babesia species in cheetah (Acinonyx jubatus, Schreber, 1775) populations in South Africa. J Clin Microbiol 48:
97 74. Kjemtrup AM, Conrad PA A review of the small canine piroplasms from California: Babesia conradae in the literature. Vet Parasitol 138: Giadinis ND, Chochlakis D, Kritsepi-Konstantinou M, Makridaki E, Tselentis Y, Kostopoulou D, Karatzias H, Psaroulaki A Haemolytic disease in sheep attributed to a Babesia lengau-like organism. Vet Rec 170: Sikorski LE, Birkenheuer AJ, Holowaychuk MK, McCleary-Wheeler AL, Davis JM, Littman MP Babesiosis caused by a large Babesia species in 7 immunocompromised dogs. J Vet Intern Med 24: Birkenheuer AJ, Neel J, Ruslander D, Levy MG, Breitschwerdt EB Detection and molecular characterization of a novel large Babesia species in a dog. Vet Parasitol 124: Dobbelaere D, Heussler V Transformation of leukocytes by Theileria parva and T. annulata. Annu Rev Microbiol 53: Susta L, Torres-Velez F, Zhang J, Brown C An in situ hybridization and immunohistochemical study of cytauxzoonosis in domestic cats. Vet Pathol 46: Hagiwara K, Takahashi K, Taniyama H, Kawamoto S, Kurosawa T, Ikuta K, Ishihara C Detection of Theileria sergenti schizonts in bovine lymph node. Int J Parasitol 27: Sato M, Kamio T, Kawazu S, Taniguchi T, Minami T, Fujisaki K Histological observations on the schizonts in cattle infected with Japanese Theileria sergenti. J Vet Med Sci 55: Blouin EF, Kocan AA, Kocan KM, Hair J Evidence of a limited schizogonous cycle for Cytauxzoon felis in bobcats following exposure to infected ticks. J Wildl Dis 23:
98 SUPPLEMENTAL MATERIAL Figure S1. Schematic of PCR amplification of B. canis canis, B. canis rossi, and C. felis mitochondrial genomes. Primers were designed to amplify near full length mitochondrial genomes in three overlapping fragments. Primers for fragments 1-3 and TIRs are indicated with arrows (forward primers: F1-F3, reverse primers: R1-R3, TIR primers: TIR F/R). Genes shown above the central line are coded on the sense strand, while those below are on the antisense strand. Protein-coding genes (cox1, cox3, and cytb) are indicated in white. Large subunit rrna fragments are in light gray, small subunit rrna fragments are in dark gray, and miscellaneous conserved RNA fragments are in black. 80
99 Figure S2. Schematic of PCR amplification of B. canis vogeli and Babesia sp. Coco mitochondrial genomes. Because attempts at TIR amplifications were unsuccessful, primers were designed to amplify near full length mitochondrial genomes of species in five overlapping fragments. Primers for fragments 0-4 are indicated with arrows (forward primers: F0-F4, reverse primers: R0-R4). Protein-coding genes (cox1, cox3, and cytb) are indicated in white. Large subunit rrna fragments are in light gray, small subunit rrna fragments are in dark gray, and miscellaneous conserved RNA fragments are in black. 81
100 Figure S3. Schematic of PCR amplification of B. conradae mitochondrial genome. Because initial attempts at amplifying near full length mitochondrial genome was unsuccessful, primers were designed to amplify near full length B. conradae mitochondrial genome in six overlapping fragments. Primers for fragments 0-4 are indicated with arrows (forward primers: F0-F4, reverse primers: R0-R4). Proteincoding genes (cox1, cox3, and cytb) are indicated in white. Large subunit rrna fragments are in light gray, small subunit rrna fragments are in dark gray, and miscellaneous conserved RNA fragments are in black. 82

101 Figure S4. Schematic of PCR amplification of B. microti-like sp. mitochondrial genome. Because of the mitochondrial genome structure of close relative B. microti is known to differ from that of Babesia sensu stricto species, an alternative approach was employed to amplify the B. microti-like sp. mitochondrial genome. Primers were designed to amplify near full length mitochondrial genomes of species in six fragments that formed two separate contigs. Amplification across assumed inverted repeats (indicated by IR-A and IR-B) was not pursued due to lack of informative phylogenetic sequence in this region; hence, a single contig of the mitochondrial genome was not obtained. Primers for fragments 1-6 are indicated with arrows (forward primers: F1-F6, reverse primers: R1-R6). Protein-coding genes (cox1, cox3, and cytb) are indicated in white. Large subunit rrna fragments are in light gray, small subunit rrna fragments are in dark gray, and miscellaneous conserved RNA fragments are in black. 83
102 Table S1. Primers utilized in additional C. felis PCR assays. Purpose Sequence Amplicon c Inverted PCR a ATACCTGTCAAGTTCCTTCACTA TIR F TTGACACTACTAAAAGCACTAGC TIR R Additional Sequencing b CAATGTGTAGAATCAGTTAGATCATTGG Fragment 1 (F) ACGTATCAATATTCTCTACTCTGTTACC Fragment 2 (F) ACTGAATATAATATCGCAGTAGAAGTAG Fragment 2 (F) CAAGAGTGTAAGGTTCATACCATAG CTGATAAATTGGATAATTCTGACTTAGTG ATCTAGTGACAAGATATGAATCAACAC CATTGAGCATGG AAATAACGGAATG TATTCCTATAATTGCACCTATAGATAACAC GGGATATTTACATTCTAGATGGTCAAAT AGGAATTGATATTAGACGTCAGAAAGTAC TATCATGTGTTGATTCATATCTTGTCAC Fragment 3 (F) Fragment 3 (F) Fragment 3 (F) Fragment 3 (F) Fragment 1 (R) Fragment 2 (R) Fragment 3 (R) Fragment 3 (R) a Primers were designed to amplify terminal inverted repeats (TIR); cloning was required for full sequence resolution b Additional primers were designed to obtain complete bi-directional sequencing of mitochondrial fragments. c (F)=Forward Primer, (R)=Reverse Primer Table S2. Primers utilized in additional B. canis canis PCR assays. Purpose Sequence Amplicon c Inverted PCR a TACCTGTCAAATTCCTTCACTAAC TIR F TTGCTCCACTCATTGCAC TIR R Internal Sequencing b ACA ACTGGAGTTATATTAGGAAATGC Fragment 1 (F) ATGGAATCAGTATATTCCAGGGTATC Fragment 2 (F) CTGATAAATTGGATAATTCTGACTTAGTG GAATCAAATTAA ACAACATGTTCCACTG GAATCAATTCCAGATAATGGATTAGTACT ATACACATTGTGCATGGAAGTAACG TCCTAAGAAATGCATTGGAATGAATG ACGTATCAATATTCTCTACTCTGTTACC TATGGGCTTTTTTGCATATGAGATG CAAATGAGTTATTGGGGAGC GGAATAGGAAAGATTAACCGCTATC Fragment 3 (F) Fragment 3 (F) Fragment 3 (F) Fragment 3 (F) Fragment 1 (R) Fragment 2 (R) Fragment 2 (R) Fragment 3 (R) Fragment 3 (R) a Primers were designed to amplify terminal inverted repeats (TIR); cloning was required for full sequence resolution b Additional primers were designed to obtain complete bi-directional sequencing of mitochondrial fragments. c (F)=Forward Primer, (R)=Reverse Primer 84
103 Table S3. Primers utilized in additional B. canis rossi PCR assays. Purpose Sequence Amplicon c Inverted PCR a ATACCTGTCAAGTTCCTTCACTA TIR F GTTTGACCTATTGATTTTAAAGCACC TIR R Additional Sequencing b CATTTTCACTTTGTTCTATCAATTGGAGC Fragment 1 (F) ATGGAATCAGTATATTCCAGGGTATC Fragment 2 (F) ACGTATCAATATTCTCTACTCTGTTACC Fragment 2 (F) CTGATA AATTGGATAATTCTGACTTAGTG GAATCAAATTAAACAACATGTTCCACTG GAATCAATTCCAGATAATGGATTAGTACT ATACACATTGTGCATGGAAGTAACG CCAAAAAGTGCATTGGTATGAAAG TATGGGCTTTTTTGCATATGAGATG AGGAATTGATATTAGACGTCAGAAAGTAC GGAATAGGAAAGATTAACCGCTATC CAAATGAGTTATTGGGGAGC Fragment 3 (F) Fragment 3 (F) Fragment 3 (F) Fragment 3 (F) Fragment 1 (R) Fragment 2 (R) Fragment 3 (R) Fragment 3 (R) Fragment 3 (R) a Primers were designed to amplify terminal inverted repeats (TIR); cloning was required for full sequence resolution b Additional primers were designed to obtain complete bi-directional sequencing of mitochondrial fragments. c (F)=Forward Primer, (R)=Reverse Primer Table S4. Primers utilized in additional B. canis vogeli PCR assays. Purpose Sequence Amplicon c Additional mitochondrial AGAACAGAATTGAGTATGAGTG Fragment 0 (F) d genome PCR amplification a CACTCATAGCACTAGCTATTCC Fragment 0 (R) d CTCATTGTGCATGAAAATAGCG ATACTAGATAGGGAACGAACTGC Fragment 4 (F) Fragment 4 (R) Additional Sequencing b ACAACTGGAGTTATATTAGGAAATGC Fragment 1 (F) GCGAATAACGTAGATGAGTTAGTTG Fragment 2 (F) ACGTATCAATATTCTCTACTCTGTTACC Fragment 2 (F) GAATCAAATTAAACAACATGTTCCACTG TGTAAGCAAATACCATTCTGGTAC CTCATTGTGCATGAAAATAGCG TCCTAAGAAATGCATTGGAATGAATG ACGTTACAGGTACTTTAAACGTAG GGAATAGGAAAGATTAACCGCTATC CAAATGAGTTATTGGGGAGC Fragment 3 (F) Fragment 3 (F) Fragment 3 (F) Fragment 1 (R) Fragment 2 (R) Fragment 3 (R) Fragment 3 (R) a B. canis vogeli required alternative PCR assays to obtain additional mitochondrial genome sequence as TIR PCR attempts were unsuccessful b Additional primers were designed to obtain complete bi-directional sequencing of mitochondrial fragments. c (F)=Forward Primer, (R)=Reverse Primer d PCR amplicons required cloning for full sequence resolution 85
104 Table S5. Primers utilized in additional Babesia sp. Coco PCR assays Purpose Sequence Amplicon c Additional mitochondrial AGAACTGAATTAAGCATGAGTG Fragment 0 (F) genome PCR amplification a TCCTAAGAAATGCATTGGAATGAATG Fragment 0 (R) GAATCAATTCCAGATAATGGATTAGTACT Fragment 4 (F) ATACTAGATAGGGAACGAACTGC Fragment 4 (R) Additional Sequencing b CAATTCCACCAAAAAGTCCAGTC Fragment 0 (R) CATTTTCACTTTGTTCTATCAATTGGAGC Fragment 1 (F) ATGGAATCAGTATATTCCAGGGTATC Fragment 2 (F) ACGTATCAATATTCTCTACTCTGTTACC Fragment 2 (F) GAATCAAATTAAACAACATGTTCCACTG Fragment 3 (F) CTGATAAATTGGATAATTCTGACTTAGTG Fragment 3 (F) CATTGAGCATGGAAATAACGGAATG Fragment 3 (F) GAATCAATTCCAGATAATGGATTAGTACT Fragment 3 (F) TCCTAAGAAATGCATTGGAATGAATG Fragment 1 (R) ACGTTACAGGTACTTTAAACGTAG Fragment 2 (R) CAAATGAGTTATTGGGGAGC Fragment 3 (R) GGAATAGGAAAGATTAACCGCTATC Fragment 3 (R) a Babesia sp. Coco required alternative PCR assays to obtain additional mitochondrial genome sequence as TIR PCR attempts were unsuccessful b Additional primers were designed to obtain complete bi-directional sequencing of mitochondrial fragments. c (F)=Forward Primer, (R)=Reverse Primer 86
105 Table S6. Primers utilized in additional Babesia conradae PCR assays Purpose Sequence Amplicon d Additional mitochondrial AGAACAGAATTGAGTATGAGTG Fragment 0 (F) e genome PCR amplification a TCCTAATATAACTCCAGTAGTACCTCC Fragment 0 (R) e ACAACTGGAGTTATATTAGGAAATGC Fragment 1.5 (F) GACAGTGAACTTGTAGCTGAAC Fragment 1.5 (R) ATACGTGCCGAGCAGTGTG Fragment 2 (F) CCTTGAAATCCAGGAATAATACC Fragment 4 (F) ATACTAGATAGGGAACGAACTGC Fragment 4 (R) Additional Sequencing b CTCTATCAACTGATAAACCAACAGATC Fragment 0 (R) ACAACTGGAGTTATATTAGGAAATGC Fragment 1 (F) ACATCATTGTAGCTCCTACTGTAG Fragment 2 (F) AATGAGGAGCGTCTGT Fragment 2 (F) CTTCGCATTAAACAGTAGGTTCC Fragment 3 (F) GATACACATTGAGCATGAAAATAACG Fragment 3 (F) CCTTGAAATCCAGGAATAATACC Fragment 3 (F) TCCTAATATAACTCCAGTAGTACCTCC Fragment 1 (R) CCTATGCATTTCTTAGGGTTTTCC Fragment 2 (R) AGGAATTGATATTAGACGTCAGAAAGTAC Fragment 3 (R) CAAATGAGTTATTGGGGAGC Fragment 3 (R) GGAATAGGAAAGATTAACCGCTATC Fragment 3 (R) cox3 PCR c ACAGGATTAGATACCCTGG cox3 (F) ACTGTCAGCTAAAACGTATC cox3 (R) a Due to its unique mitochondrial genome structure as well as the inability to successfully perform TIR PCR, B. conradae required alternative PCR assays to obtain additional mitochondrial genome sequence b Additional primers were designed to obtain complete bi-directional sequencing of mitochondrial fragments c Primers designed to amplify cox3 from B. conradae yielded a cox3-like amplicon d (F)=Forward Primer, (R)=Reverse Primer e PCR amplicons required cloning for full sequence resolution 87
106 Table S7. Primers utilized in additional Babesia microti-like sp. (T. annae) PCR assays. Purpose Sequence Amplicon c Mitochondrial genome TATTAGCTACCTTTGGCTATC Fragment 1 (F) PCR amplification a AGTATTAGCACATACACTTCTGG Fragment 1 (R) GGAAGTGGWACWGGWTGGAC Fragment 2/cox1 (F) TTCGGTATTGCATGCCTTG Fragment 2/cox1 (R) ACAAAGACTAATGCTTGAAGTGG Fragment 3 (F) GTAACTCAGTCAGACCTTTG Fragment 3 (R) TTAGTGAAGGAACTTGACAGGT Fragment 4/cytb (F) CGGTTAATCTTTCCTATTCCTTACG Fragment4.cytb (R) CTCGATATTAATCTTAAAGTACAGGAC Fragment 5 (F) ATCTAGTGCCAGCAGTAG AGCTTACTAATGAATTGTCTCG CCTTCTATAATTATGCTAATTTCAGTAG Fragment 5 (R) Fragment 6 (F) Fragment 6 (R) Additional sequencing b AGTATTAGCACATACACTTCTGG Fragment 2/cox1 (R) CATCTGATATCTACATGATGGC Fragment 4/cytb (F) CATTTCGTAGAAGCTGTATGG Fragment 5 (F) TTCAGCTACAAGTTCACTGTC Fragment 5 (F) TTGCTCACACAGTCAATACG Fragment 5 (R) ACTCATATCTATTACCACTATAGGC Fragment 5 (R) a Due to its mitochondrial genome structure, B. microti-like sp. required alternative PCR assays to obtain additional mitochondrial genome sequence b Additional primers were designed to obtain complete bi-directional sequencing of mitochondrial fragments c (F)=Forward Primer, (R)=Reverse Primer 88
107 Table S8. Locations of protein-coding genes and rrna fragments within Piroplasmida mitochondrial genomes. Gene/fragment coordinates with a white background are coded on the sense strand, while those highlighted in yellow are on the antisense strand. Some genes/fragments were not identified in some mitochondrial genomes (indicated in green and with No ID ) while others were duplicated for some mitochondrial genomes (indicated in blue). Coordinates initially reported in separate studies are not reported in this table (noted in gray, previously published ). Genes/fragments that were identified for all species were included in phylogenetic analysis (highlighted in red). Species and GenBank Accession Number Cytauxzoon Babesia canis Babesia canis Babesia canis Babesia sp. felis canis rossi vogeli Coco Babesia Babesia Babesia Babesia Babesia Babesia bovis Theileria conradae microti-like gibsoni bigemina caballi parva (T. annae) Theileria Theileria Theileria annulata orientalis equi Babesia Babesia Babesia rodhaini microti microti (linear)* (circular) Gene KC KC KC KC KC KC KC AB AB AB AB AB NW AB AB AB AB FO cox cox No ID^ cytb LSU LSU Previously published LSU LSU LSU LSU LSUA * LSUB * LSUC No ID No ID No ID No ID No ID No ID No ID No ID No ID No ID SSUA * SSUB * SSUD * SSUE * SSUF * RNA * RNA No ID * RNA * RNA RNA No ID No ID No ID No ID No ID RNA No ID No ID No ID No ID No ID RNA No ID No ID No ID No ID No ID No ID RNA No ID No ID No ID No ID No ID RNA , , RNA * , RNA * ^Although a cox3-like gene was amplified, its location relative to the mitochondrial genome is unknown *Provided by Kenji Hikosaka 89
108 Figure S5. Phylogenetic analysis of cox1 nucleotide sequence does not recover the five distinct lineages identified by analysis of concatenated mitochondrial and 18S sequences. Statistical support for clades are indicated at each node: posterior probabilities from Bayesian analysis (10 million generations of Markov chain Monte Carlo) are listed above nodes, while bootstrap values from Maximum Likelihood analysis (100 bootstrap replicates) are listed below nodes. Tree is drawn to scale, with branch lengths measured in the number of substitutions per site. Analysis of 1287 total characters produced a tree with decent statistical support, but the Babesia conradae clade was grouped within the Theileria clade, which does not reflect the results of analysis of concatenated mitochondrial and 18S sequences. 90
109 Figure S6. Phylogenetic analysis of cytb nucleotide sequence does not recover the five distinct lineages identified by analysis of concatenated mitochondrial and 18S sequences. Statistical support for clades are indicated at each node: posterior probabilities from Bayesian analysis (10 million generations of Markov chain Monte Carlo) are listed above nodes, while bootstrap values from Maximum Likelihood analysis (100 bootstrap replicates) are listed below nodes. Tree is drawn to scale, with branch lengths measured in the number of substitutions per site. Analysis of 1095 total characters produced a tree with poor statistical support for many nodes and a topology that does not reflect the results of analysis of concatenated mitochondrial and 18S sequences. 91
110 Figure S7. Phylogenetic analysis of CYTB amino acid sequence does not recover the five distinct lineages identified by analysis of concatenated mitochondrial and 18S sequences. Statistical support for clades (bootstrap values from Maximum Likelihood analysis, 100 bootstrap replicates) are indicated below each node. Tree is drawn to scale, with branch lengths measured in the number of substitutions per site. Analysis of 365 total characters produced a tree with poor statistical support for many nodes and a topology that does not reflect the results of analysis of concatenated mitochondrial and 18S sequences. 92
111 Figure S8. Phylogenetic analysis of concatenated cox1 and cytb nucleotide sequence does not recover the five distinct lineages identified by analysis of concatenated mitochondrial and 18S sequences. Statistical support for clades are indicated at each node: posterior probabilities from Bayesian analysis (10 million generations of Markov chain Monte Carlo) are listed above nodes, while bootstrap values from Maximum Likelihood analysis (100 bootstrap replicates) are listed below nodes. Tree is drawn to scale, with branch lengths measured in the number of substitutions per site. Analysis of 2382 total characters produced a tree with poor statistical support for multiple nodes and a topology that does not reflect the results of analysis of concatenated mitochondrial and 18S sequences. 93
112 Figure S9. Phylogenetic analysis of concatenated COX1 and CYTB amino acid sequence recovers the five distinct lineages identified by analysis of concatenated mitochondrial and 18S sequences but with poorer statistical support than analysis COX1 amino acid alone. Statistical support for clades (bootstrap values from Maximum Likelihood analysis, 100 bootstrap replicates) are indicated below each node. Tree is drawn to scale, with branch lengths measured in the number of substitutions per site. Analysis of 794 total characters produced a tree with topology identical to that of analysis of COX1 amino acid sequence, but had poorer statistical support. 94
113 CHAPTER 3: Pharmacogenomics of Cytauxzoon felis cytochrome b: implications for atovaquone and azithromycin therapy in domestic cats with cytauxzoonosis Megan E. Schreeg* 1, Henry S. Marr 1, Jaime Tarigo 1, Leah A. Cohn 2, Michael G. Levy 1, Adam J. Birkenheuer 1 1. North Carolina State University College of Veterinary Medicine, Raleigh, NC. 2. University of Missouri, College of Veterinary Medicine, Columbia, MO. *Responsible for all laboratory work, statistical analyses, and majority of writing As published in: Journal of Clinical Microbiology, 2013 Sep; 51(9): doi: /JCM ABSTRACT Cytauxzoon felis, an emerging virulent protozoan parasite that infects domestic cats, is treated with atovaquone and azithromycin (A&A). Atovaquone targets parasite cytochrome b. We characterized the C. felis cytochrome b gene (cytb) in cats with cytauxzoonosis, and found a cytb genotype that was associated with survival in A&A-treated cats. TEXT Cytauxzoonosis is an emerging disease in domestic and wild felines in North and South America caused by the tick-transmitted apicomplexan protozoan parasite, Cytauxzoon felis (1, 2). Without treatment, cytauxzoonosis is fatal in up to 97% of domestic cats (3). Recent advances in treatment combining atovaquone and azithromycin (A&A) have reduced the mortality rate to 40% (3). Azithromycin targets the mitochondrial ribosomes of the parasite, while atovaquone targets protozoal cytochrome b (cytb), disrupting electron transport in the parasite mitochondria (4, 5). 95
114 In related parasites, including Babesia and Plasmodium species, resistance to atovaquone treatment has been attributed to mutations in the cytb gene (6-11). However, to this point, no similar pharmacogenomic studies have been performed characterizing the C. felis cytb gene. The purpose of this study is to determine whether or not response to A&A treatment is associated with C. felis cytb genotype. Therefore, we characterized and compared C. felis cytb genotypes from cats with cytauxzoonosis that were treated with A&A or another type of antiprotozoal drug, imidocarb dipropionate, which does not interact with cytb (12). Sixty-nine pre-treatment DNA samples from cats with cytauxzoonosis were available from a previous study (3). Total DNA was isolated from 200 µl of infected feline whole blood using a commercial kit according to kit instructions (QIAamp DNA Blood Mini Kit, Qiagen Inc., Valencia, CA). All samples were confirmed to be infected by using a Cytauxzoon felis-specific PCR assay (13). Cats were treated with A&A (n=45) or imidocarb dipropionate (n=24) as described and clinical outcome was recorded (3). Full length C. felis cytb was amplified in three overlapping fragments (Figure 1). Primers for fragment 1 (forward: 5 -CTTAACCCA ACTCACGTACC -3, reverse: 5 - ATCTAGTGACAAGATATGAATCAACAC -3 ), fragment 2 (forward: 5 - ACCTTGGTCATGGTATTCAG-3, reverse: 5 -GATCTAGCTTCAACCAATGC-3 ), and fragment 3 (forward: 5 -GCATAGATGTTCAAGTACTAATCC-3, reverse: 5 - GGTTAATCTTTCCTATTCCTTACG-3 ) were designed based on previously reported C. felis cytb sequence (GenBank Accession KC207821; Figure 1). Each 50 µl reaction contained 1 µl of DNA template, 50 pmol of each primer, 10 nmol dntps, 1.75 U Expand High Fidelity Enzyme mix and a 1X concentration of Expand High Fidelity Buffer with 96
115 Figure 1. PCR amplification of C. felis cytb gene in three overlapping fragments. Full-length cytb gene (1092 bp) was PCR amplified from 69 samples in three overlapping fragments. Primers are indicated with labeled arrows. Annealing temperatures for PCR assays were optimized for each fragment: Fragment 1=53.8 degrees C, Fragment 2=56 degrees C, and Fragment 3=53 degrees C. Fragment 2 includes the putative atovaquone binding sites (*) (6). MgCl2 (Roche, Mannheim, Germany). Thermal cycling conditions consisted of an initial denaturation at 95 ºC for 5 minutes, followed by 45 amplification cycles (95 ºC for 20 seconds, ºC for 30 seconds, and 60 ºC for 45 seconds) and a final extension step at 68 ºC for 7 minutes (Techne Inc., Burlington, NJ). Annealing temperatures for fragments 1-3 were 53.8, 56, 53 respectively (Figure 1); a 60 ºC extension temperature was found to be superior to 72 ºC, presumably due to high AT nucleotide content (14). Positive controls consisted of C. felis-infected feline blood samples and negative controls consisted of water (no DNA). Amplicons were visualized on an agarose gel, purified, and sequenced bidirectionally (MCLAB, South San Francisco, CA); chromatograms were carefully inspected for heterogeneity. Any secondary peaks present at 30% or more of the primary nucleotide peaks in both forward and reverse sequence were edited accordingly using IUPAC ambiguity codes (Vector NTI, Invitrogen, Grand Island, NY; Figure 2). Contigs were assembled using a commercially available software package (BioEdit Sequence Alignment Editor, North 97
116 Figure 2. Presence of secondary peaks in C. felis cytb sequence as determined by Vector NTI. All sequences were analyzed bidirectionally to detect the presence of secondary peaks (for example, the thymine present as a peak secondary to cytosine at position 114 in the sequence above). Nucleotide sequence for the sample was edited accordingly using IUPAC ambiguity code (for example, Y in the sequence above). Carolina State University, Raleigh, NC). Sequence data for ribosomal internal transcribed spacer (ITS) regions were characterized for 61 out of 69 samples (3) and assessed for any potential associations with cytb genotype. Associations between categorical variables were analyzed using two-tailed Fisher exact probability tests with p < 0.05 considered significant (VassarStats, Poughkeepsie, NY). A total of 30 C. felis cytb genotypes were characterized (Figure 3). The majority of samples (46/69) showed evidence of infection with C. felis possessing a single genotype of cytb, while the remaining samples (23/69) had two or more cytb genotypes present. In the vertebrate host C. felis exists as haploid forms, which replicate asexually. While the majority 98
.")
117 of genetic material is encoded on chromosomes, some apicomplexan genes, including cytb, are encoded on extra-chromosomal fragments of DNA within the apicoplast or mitochondria (15). In apicomplexan parasites, up to 20 copies of the mitochondrial genome can exist within one non-dividing haploid organism (15). Therefore samples containing two or more genotypes could represent a single haploid C. felis organism with multiple different copies of the cytb gene or multiple haploid organisms with different cytb genes. One single cytb genotype was found in 13 samples and was designated as cytochrome b genotype 1 (cytb1). Compared to cytb1, 35 different locations throughout the cytb gene were found to have single nucleotide substitutions. Of the 30 different cytb genotypes, only Figure 3. Characterization of 30 novel C. felis cytb genotypes. Thirty different genotypes were characterized from 69 total C. felis samples collected from cats with cytauxzoonosis. Thirty-five different point mutations were discovered throughout the gene, 11 of which altered nucleotide sequence in or near the putative atovaquone binding sites (denoted by gray shading and brackets at the bottom). Genotype 1 (cytb1) was present alone in 13 samples. Asterisks indicate mutations conferring amino acid changes. Nucleotides differing from the cytb1 sequence are in black boxes. AA, amino acid; NUC, nucleotide; GT, genotype; Tx, treatment (A&A, atovaquone and azithromycin; IMID, imidocarb dipropionate; BOTH, different cats possessing this genotype were treated with A&A or imidocarb); %SUR, percent survival of cats infected with the indicated cytb genotype. Gray shading in the headings indicates the 3 different cytb PCR amplicons (fragments 1 to 3). 99

118 nine genotypes had non-synonymous substitutions conferring amino acid changes in the cytb gene, with three of these in or near the atovaquone-binding site (Figure 3, Figure 4). Due to a small sample size, only cytb1 was assessed for association with survival in each treatment group. Cytb1 was found to be positively associated with survival (p=0.017) as compared to all other genotypes in cats treated with A&A (Table 1). However, there was no association between cytb1 and survival (p=0.608) in cats treated with imidocarb dipropionate (Table 1). While no statistical comparisons could be performed, all three cats infected with genotypes conferring amino acid changes in or near the putative atovaquonebinding site died; only one of these cats was treated with A&A (Figure 3). Some genotypes, including cytb1, were found only in select states (see Table S1 in Supplemental Material). Cytb1 was only found in Arkansas and Missouri. Association between cytb1 and survival in the subpopulation of cats treated with A&A from Arkansas and Missouri remained significant (p=0.001) when only these samples were considered, while there was again no association (p=0.545) between cytb1 and survival in cats from AR Figure 4. Evidence of missense mutations in or near putative atovaquone-binding sites of C. felis CYTB. Putative amino acid sequences of C. felis CYTB genotypes with missense mutations in the atovaquone-binding sites were aligned with the P. falciparum CYTB sequence. Blue-bracketed regions indicate putative atovaquone-binding sites, predicted by alignment with P. falciparum CYTB atovaquone-binding sites (6). Red arrows indicate sites of previously characterized mutations in related Plasmodium and Babesia species linked to atovaquone resistance. Green arrows indicate sites of missense mutations (compared to the amino acid sequence of cytb1) discovered in C. felis CYTB in this study. 100
119 Table 1. Correlation between survival rate of cats treated with A&A and C. felis cytb genotype a a Survival rate of cats infected with C. felis cytb1 was compared to survival rate of cats infected with strains of all other genotypes combined ( non-cytb1 ). Results were analyzed using a two-tailed Fisher exact probability test. and MO treated with imidocarb dipropionate. (see Table S2 in Supplemental Material). Furthermore, when comparing survival rates of cats from AR and MO to cats from OK, NC, and TN where cytb1 was not present, there was no significant difference in survival rates (see Table S3 in Supplemental Material), indicating that the increased survival benefit of cytb1does not merely reflect a decreased virulence of C. felis in AR and MO. We found no association between the most common ITS genotype (ITSc, GenBank accession EU450802/EU450804) and survival or ITS genotype and cytochrome b genotype (see Tables S4 to S6 in Supplemental Material; 16, 17). In this study, we found that the C. felis cytb gene sequence is highly variable. Despite the high variability of the cytb gene and a relatively small sample size, we were able to detect a cytb genotype (cytb1) that was associated with survival in cats treated with A&A (Table 1). We anticipate that with a larger sample size the cytb 1 genotype would be detected in other regions, albeit at lower proportions. Additionally, we believe it is likely that other genotypes, such as cytb genotype 3, may confer a survival benefit in cats treated with A&A (Figure 3). 101
120 Studies involving a larger sample size across a larger geographic range should be pursued to further assess these hypotheses. Despite sharing identical amino acid sequence with nearly all other cytb genotypes (Table 1), the cytb1 genotype was associated with survival in cats treated with A&A. While a synonymous substitution does not cause an amino acid change, silent mutations can result in changes in protein amount, structure, or function (18, 19). For instance, differences in cytb1 nucleotide sequence could have an effect on mrna structure, stability, and translation kinetics (codon preference; 20). Evidence with human multidrug resistance 1 gene (MDR1) has shown that synonymous substitutions can alter the kinetics of translation, leading to alterations in protein folding and intracellular function (18, 21). Likewise, synonymous substitutions in cytb could possibly impact protein folding or structure and alter atovaquone binding. Another possibility is that the cytb1 genotype is a genetic marker for alterations in promoter regions or neighboring mitochondrial genes. These genes include cytochrome c oxidase subunits I and III (cox1 and cox3), which are involved downstream of cytb in the electron transport chain, and large subunit (LSU) ribosomal RNA fragments believed to be involved in translation of mitochondrial genes (5). Alterations in any of these genes could impact the metabolic efficiency of the mitochondria and fitness of the organism, rendering the parasite more susceptible to A&A treatment. Further studies are needed to discern the complete pharmacogenomic role of C. felis cytb genotypes. In conclusion, cytb genotype appears promising for predicting survival in cats with cytauxzoonosis treated with A&A. We are evaluating assays such as high resolution melting curve analysis to rapidly characterize cytb genotypes from clinical samples to provide prognostic information for cats with cytauxzoonosis. 102
121 ACKNOWLEDGEMENTS We would like to acknowledge the technical assistance of Karen Gore. This work was supported by The ALSAM Foundation. We do not have any conflicts of interest to declare. REFERENCES 1. Andre MR, Adania CH, Machado RZ, Allegretti SM, Felippe PA, Silva KF, Nakaghi AC, Dagnone AS Molecular detection of Cytauxzoon spp. in asymptomatic Brazilian wild captive felids. J Wildl Dis 45: Brown HM, Lockhart JM, Latimer KS, Peterson DS Identification and genetic characterization of Cytauxzoon felis in asymptomatic domestic cats and bobcats. Vet Parasitol 172: Cohn LA, Birkenheuer AJ, Brunker JD, Ratcliff ER, Craig AW Efficacy of atovaquone and azithromycin or imidocarb dipropionate in cats with acute cytauxzoonosis. J Vet Intern Med 25: Mather MW, Henry KW, Vaidya AB Mitochondrial drug targets in apicomplexan parasites. Curr Drug Targets 8: Vaidya AB, Mather MW Mitochondrial evolution and functions in malaria parasites. Annu Rev Microbiol 63: Korsinczky M, Chen N, Kotecka B, Saul A, Rieckmann K, Cheng Q Mutations in Plasmodium falciparum cytochrome b that are associated with atovaquone resistance are located at a putative drug-binding site. Antimicrob Agents Chemother 44: Sakuma M, Setoguchi A, Endo Y Possible emergence of drug-resistant variants of Babesia gibsoni in clinical cases treated with atovaquone and azithromycin. J Vet Intern Med 23: Srivastava IK, Morrisey JM, Darrouzet E, Daldal F, Vaidya AB Resistance mutations reveal the atovaquone-binding domain of cytochrome b in malaria parasites. Mol Microbiol 33: Syafruddin D, Siregar JE, Marzuki S Mutations in the cytochrome b gene of Plasmodium berghei conferring resistance to atovaquone. Mol Biochem Parasitol 104:
122 10. Vaidya AB, Mather MW Atovaquone resistance in malaria parasites. Drug Resist Updat 3: Wormser GP, Prasad A, Neuhaus E, Joshi S, Nowakowski J, Nelson J, Mittleman A, Aguero-Rosenfeld M, Topal J, Krause PJ Emergence of resistance to azithromycin-atovaquone in immunocompromised patients with Babesia microti infection. Clin Infect Dis 50: Wickramasekara Rajapakshage BK, Yamasaki M, Hwang SJ, Sasaki N, Murakami M, Tamura Y, Lim SY, Nakamura K, Ohta H, Takiguchi M Involvement of Mitochondrial Genes of Babesia gibsoni in Resistance to Diminazene Aceturate. J Vet Med Sci 74: Birkenheuer AJ, Le JA, Valenzisi AM, Tucker MD, Levy MG, Breitschwerdt EB Cytauxzoon felis infection in cats in the mid-atlantic states: 34 cases ( ). J Am Vet Med Assoc 228: Su XZ, Wu Y, Sifri CD, Wellems TE Reduced extension temperatures required for PCR amplification of extremely A+T-rich DNA. Nucleic Acids Res 24: Wilson RJ, Williamson DH Extrachromosomal DNA in the Apicomplexa. Microbiol Mol Biol Rev 61: Brown HM, Berghaus RD, Latimer KS, Britt JO, Rakich PM, Peterson DS Genetic variability of Cytauxzoon felis from 88 infected domestic cats in Arkansas and Georgia. J Vet Diagn Invest 21: Brown HM, Modaresi SM, Cook JL, Latimer KS, Peterson DS Genetic variability of archived Cytauxzoon felis histologic specimens from domestic cats in Georgia, J Vet Diagn Invest 21: Sauna ZE, Kimchi-Sarfaty C, Ambudkar SV, Gottesman MM Silent polymorphisms speak: how they affect pharmacogenomics and the treatment of cancer. Cancer Res 67: Sauna ZE, Kimchi-Sarfaty C Understanding the contribution of synonymous mutations to human disease. Nat Rev Genet 12: Sauna ZE, Kimchi-Sarfaty C, Ambudkar SV, Gottesman MM The sounds of silence: synonymous mutations affect function. Pharmacogenomics 8: Shabalina SA, Spiridonov NA, Kashina A Sounds of silence: synonymous nucleotides as a key to biological regulation and complexity. Nucleic Acids Res 41:
123 SUPPLEMENTAL MATERIAL Table S1. Frequency of C. felis cytb genotypes in five different states. 105


124 Table S2. Survival rate of cats treated with A&A is associated with C. felis cytb genotypes localized to geographic range of cytb1. In this study, cytb1 was found only in samples from Arkansas and Missouri. We assessed the sub-population of samples from these states to determine if the association between cytb1 and survival and A&A treatment remained; results from assessing this sub-population show the same trend as present in the entire study population. Table S3. Regions with cytb1 do not have higher survival rate regardless of treatment when compared to regions lacking cytb1. C. felis cytb1 was discovered only in Arkansas and Missouri; to determine if all genotypes in these two states conferred a higher survival rate, survival rates in Arkansas and Missouri were compared to survival rates of samples from Tennessee, North Carolina, and Oklahoma. In either treatment group, there was not a significant difference in survival rate between regions with cytb1 and those without cytb1. 106

125 Table S4. Frequency of C. felis cytb genotypes in relation to ITS genotypes. ITS genotypes were characterized for 61/69 samples used in this study. 107


126 Table S5. C. felis cytb1 is not associated with ITS genotypes. ITS genotypes were available for 61/69 samples used in this study. There was no association between cytb1, the most prevalent cytb genotype, and ITS genotypes. Data were analyzed using a two-tailed Fisher exact probability test. Table S6. ITSc genotype is not associated with increased survival. In this study, there was no apparent association between survival rate and the most common ITS genotype (ITSc). 108
127 CHAPTER 4: Rapid high resolution melt analysis of Cytauxzoon felis cytochrome b to aid in the prognosis of cytauxzoonosis Megan E. Schreeg* 1, Henry S. Marr 1, Jaime L. Tarigo 1, Leah A. Cohn 2, Michael G. Levy 1, Adam J. Birkenheuer 1 1. North Carolina State University, College of Veterinary Medicine, Raleigh, NC, USA 2. University of Missouri, College of Veterinary Medicine, Columbia, MO, USA *Responsible for all laboratory work, statistical analyses, and majority of writing As published in: Journal of Clinical Microbiology, In press. Accepted manuscript posted online May 27 th, doi: /JCM ABSTRACT Cytauxzoon felis is a virulent tick-transmitted protozoan parasite that infects felines. Cytauxzoonosis was previously believed to be uniformly fatal in domestic cats. Treatment combining atovaquone and azithromycin (A&A) has been associated with survival rates over 60%. Atovaquone, a ubiquinone analogue, targets C. felis cytochrome b (cytb), of which 30 unique genotypes have been identified. The C. felis cytb genotype cytb1 is associated with increased survival in cats treated with A&A. The purpose of this study was to design a PCR panel that could distinguish C. felis cytb1 from other cytochrome b genotypes. Primer pairs were designed to span five different nucleotide positions where single nucleotide polymorphisms (SNPs) had been identified in the C. felis cytb gene. Through the use of high-resolution melt (HRM) analysis, this panel was predicted to distinguish cytb1 from other cytb genotypes. Assays were validated using samples from 69 cats with cytauxzoonosis that had their C. felis cytb genotypes previously characterized. The 109
128 PCR panel identified C. felis cytb1 with 100% sensitivity and 98.2% specificity. High resolution melt analysis can rapidly provide prognostic information for clients considering A&A treatment in cats with cytauxzoonosis. INTRODUCTION Cytauxzoonosis is an emerging disease in domestic and wild felids in North and South America caused by the tick-transmitted apicomplexan parasite Cytauxzoon felis (1-8). Cytauxzoonosis was originally thought to be uniformly fatal in domestic cats (4, 5), but our understanding of the epidemiology of C. felis is evolving. Recent evidence indicates that some cats survive C. felis infection without any evidence of clinical disease and/or history of antiprotozoal therapy (9-13). Whether this change is due to increased recognition of subclinical infection, differences in infectious dose, alternative mechanisms of transmission, or differences in virulence between strains is unclear. However, for cats presented to veterinary hospitals with acute cytauxzoonosis, mortality rates remain high. Even with advances in treatment, mortality rates ranged from 40-74% in a prospective randomized clinical trial (14). In the same study, atovaquone and azithromycin (A&A) was associated with improved survival rates compared to imidocarb dipropionate (14), which was previously considered the treatment of choice (15, 16). The majority of cats with acute cytauxzoonosis that die do so within 2-5 days of presentation (4, 14). Given this rapid clinical course, it is paramount to initiate A&A therapy as soon as possible. Azithromycin is believed to target the mitochondrial ribosomes of the parasite, while atovaquone is presumed to target protozoal cytochrome b (cytb), disrupting electron transport in the parasite mitochondria (17, 18). In related parasites, mutations in the putative atovaquone-binding site of cytb have been associated with response to A&A treatment (19-110
129 21). A recent study has identified a C. felis cytb genotype (cytb1) that was associated with increased survival rates in cats treated with A&A (22). Cats infected with this genotype (cytb1) that were treated with imidocarb dipropionate, which does not target cytb, did not have a significantly improved survival rate compared to imidocarb dipropionate-treated cats infected with non-cytb1 genotypes (22). Therefore, the improved survival rate of cats infected with C. felis cytb1 appears to be specifically associated with A&A treatment. It is unknown if this association is a direct result of the cytb1 nucleotide or amino acid sequence, or if cytb1 is simply a genetic marker for other mechanisms of susceptibility to A&A treatment (22). Identification of C. felis cytb1 in a clinical sample from a cat with cytauxzoonosis could provide useful prognostic information for a client deciding whether or not to pursue A&A therapy, which can cost thousands of dollars. Accurate cytb genotyping may be challenging due to its location in the mitochondrial genome. Eukaryotic cells possess multiple copies of mitochondrial genomes and different mitochondrial sequences can co-exist within the same cell or tissue, a condition known as heteroplasmy (23-26). While it is currently unknown how many copies of the mitochondrial genome exist in C. felis organisms, in related apicomplexan parasites up to 150 copies of the mitochondrial genome can exist within one non-dividing haploid organism (27). Characterization of C. felis cytb genotypes revealed heteroplasmic single nucleotide polymorphisms (SNPs) in 16 out of 30 C. felis cytb genotypes, representing 33% (23/69) of total samples (22). Due to the rapid clinical course, high mortality rate, cost of treatment, and prevalence of heteroplasmy, an assay to detect C. felis cytb1 must be rapid, costeffective, and sensitive enough to discriminate heteroplasmic SNPs. Quantitative real-time PCR coupled with high resolution melt (HRM) analysis fulfills these criteria (23, 28-34). 111
130 The objective of this study is to develop an assay that can rapidly and accurately identify C. felis cytb1 in clinical samples using a quantitative real-time PCR panel coupled with HRM analysis. Herein we describe an assay that can detect C. felis cytb1 in feline blood samples with 100% sensitivity and 98.2% specificity. This assay can provide useful prognostic information for owners of cats with cytauxzoonosis. MATERIALS AND METHODS Sixty-nine pre-treatment DNA samples from cats with cytauxzoonosis were available from a previous study (14). Total DNA was isolated from 200 µl of infected feline whole blood using a commercial kit according to kit instructions (QIAamp DNA Blood Mini Kit, Qiagen Inc., Valencia, CA). All samples were confirmed to be infected by PCR amplification of a portion of the C. felis 18S rrna gene as previously described (35). Cytauxzooon felis cytb genotypes had been previously characterized using bi-directional sequencing. Any secondary nucleotide peaks present at a height 30% of the primary nucleotide peak height on DNA sequence chromatograms were detected by a computer program and denoted using IUPAC ambiguity codes (Vector NTI, Invitrogen, Grand Island, NY; 22). A total of 30 unique C. felis cytb genotypes have been identified; thirty-five nucleotide positions were determined to have single-nucleotide polymorphisms (SNPs; Table 1; 22). It was predicted that cytb1 could be distinguished from all other cytb genotypes by assessment of as few as five nucleotide positions (Table 2). Primer pairs were designed to span each of the five nucleotide positions in the C. felis cytb gene (Table 3). Each 25 µl PCR reaction contained 12.5 µl of 2X SsoAdvanced SYBR Green Supermix (Bio-Rad Laboratories, Inc., Hercules, CA), 12.5 pmol of each primer, and 1 µl of DNA 112
 were previously identified at 35 different nucleotide positions in the 1092 base pair C. felis cytb gene.")
131 Table 1. Nucleotide positions evaluated for the identification of cytb1. Single nucleotide polymorphisms (highlighted in dark gray) were previously identified at 35 different nucleotide positions in the 1092 base pair C. felis cytb gene. Nucleotide positions selected for assessment by HRM analysis are indicated by the shaded columns. template. Thermal cycling conditions (CFX96, Bio-Rad Laboratories, Inc., Hercules, CA) consisted of an initial denaturation step at 98 C for 3.5 minutes followed by 40 amplification cycles (98 C for 20 seconds and 60 C for 30 seconds) and a melt curve step (62 C to 80 C, increasing at increments of 0.1 C every 10 seconds). Annealing/extension temperature was optimized to eliminate the production of non-specific amplicons and initial PCR products were confirmed via gel electrophoresis. Amplicons that had abnormal amplification Table 2. Predicted and actual specificity of the PCR panel. cytb Nucleotide(s) Analyzed Predicted cytb1 Specificity Actual cytb1 Specificity 750 A 80.4% (45/56) 75% (42/56) 750 A T 92.9% (52/56) 85.7% (48/56) 750 A T A 96.4% (54/56) 91.1% (51/56) 750 A T A +274 C 98.2% (55/56) 96.4% (54/56) 750 A T A +274 C G 100% (56/56) 98.2% (55/56) *Specificity defined as (Number of True Negatives/(Number of True Negatives + False Positives)) 113
132 curves (e.g., non-sigmoidal) were excluded and PCRs were repeated prior to high resolution melt (HRM) analysis (9 repeated PCRs/1035 total PCRs). Following PCR, amplicons were analyzed via high resolution melt (HRM) analysis with auto-cluster detection analysis set at 50% melt curve shape sensitivity and 0.10 ºC Tm difference threshold (Precision Melt Analysis, Bio-Rad Laboratories, Inc., Hercules, CA). Assays were performed in triplicate for each sample; only samples clustering identically with corresponding positive controls in all three replicates were considered true matches to sequence data. Any sample that had insufficient DNA volume for triplicate assays was diluted 1:10 in water (n=4 DNA samples). Positive controls consisted of samples previously characterized as having mitochondrial homoplasmy at the five evaluated C. felis cytb nucleotide positions and negative controls consisted of C. felis-negative cat DNA and water (no template). Initial amplicon sequences were confirmed via TA cloning (PGEM T Easy, Promega, Madison, WI) and bi-directional sequencing (MCLAB, South San Francisco, CA). Table 3. Primers sequences for amplification of five SNP regions analyzed by HRM and full length C. felis cytb. Target Forward Primer Reverse Primer Amplicon Size (bp) Nucleotide 750 GGAGATGTTGATAATTCAATATTGGC CATTCAGGAACAATATGCAATGG 65 Nucleotide 947 GTCCATTATCATAGAGATTGGACAG TGACATTCTTCCAATGCATCC 84 Nucleotide 399 CAAATGGTCAAATGAGCTATTGG CCCAATAAAACAAATTTGTAATGACTG 57 Nucleotide 274 CTTTATGTTTTTACATGTATTGAAAGGG CATGACCAAGGTAGATATCTACTAG 63 Nucleotide 858 TTAGCAGGACTGATTGCTATG GCTTCAACCAATGCTACAAG 59 Full-length cytb AGGATACAGGGCTATAACCAAC GTACTCTGGCTATGTCAATTTCTAC 1203 To investigate discrepancies between chromatogram and HRM results for nucleotide 750 that impacted the specificity of the PCR panel, HRM analysis was performed using mixtures of cytb clones that simulated mitochondrial heteroplasmy at nucleotide 750. Full length C. felis cytb1 (adenine at nucleotide position 750) and cytb2 (guanine at nucleotide position 750) were amplified (Table 3) and cloned (PGEM T Easy, Promega, Madison, WI). 114
133 Plasmids were diluted to clinically relevant concentrations (e.g., cycle threshold results between 15-25), mixed in varying ratios, and subjected to PCR and HRM analysis of C. felis cytb nucleotide 750 to assess the sensitivity of HRM to differentiate heteroplasmic SNPs at nucleotide position 750 (Figure 3). Predicted and actual specificity of detection of C. felis cytb1 by HRM was calculated for the cumulative assessment of one, two, three, four, or five nucleotide positions (Table 2). Positive predictive values were estimated for the cumulative assessment of each nucleotide position based on the 18.8% prevalence (13/69 samples) of C. felis cytb1 in the sample population and the limits of the 95% confidence interval (95% CI: %, Table 4; 36). Table 4. Positive predictive value (PPV) for identification of C. felis cytb1 is highest when all five nucleotide positions in the PCR panel are analyzed. cytb Nucleotide(s) Analyzed Specificity of cytb1 detection PPV with cytb1 population prevalence of 8% PPV with cytb1 population prevalence of 18.8% PPV with cytb1 population prevalence of 26.7% 750 A A T A T A A T A C 750 A T A C G RESULTS Overall performance of PCR Panel The PCR panel had a sensitivity of 100% (13/13 cytb1 samples correctly identified) and specificity of 98.2% (55/56 non-cytb1 samples correctly identified; Table 2). Positive predictive values for identification of C. felis cytb1 by cumulative analysis of each nucleotide positions ranged from 25.8% to 95.3% and are summarized in Table 4. Representative high 115
134 resolution melt curves and differential fluorescence curves for each nucleotide analyzed are shown in Figure 1A-1E. Predicted vs. actual performance by nucleotide position Of the five nucleotide positions assessed in this panel, nucleotide 750 was predicted to differentiate the largest cohort of non-cytb1 samples from cytb1. Based on chromatogram data, it was predicted in silico that analysis of nucleotide 750 alone would identify 45/56 non-cytb1 samples (Specificity=80.4%). However, actual HRM analysis of nucleotide 750 alone only identified 42/56 non-cytb1 samples (Specificity=75%). Analysis of nucleotide 750 failed to identify four non-cytb1 samples, yet additionally identified a single non-cytb1 sample that was predicted to only have a SNP at nucleotide 947 (Table 2). Based on chromatogram data, in silico analysis of nucleotides 750 and 947 together was predicted to identify 52/56 non-cytb1 samples (Specificity=92.9%). When taking into account the actual results of HRM analysis of nucleotide 750, it was now predicted that analysis of nucleotides 750 and 947 together would identify 48/56 non-cytb1 samples (Specificity=85.7%). Actual HRM analysis of nucleotides 750 and 947 was consistent with this prediction (Table 2). Based on chromatogram data, in silico analysis of nucleotides 750, 947, and 399 together was predicted to identify 54/56 non-cytb1 samples (Specificity=96.4%). A total of three samples had SNPs at nucleotide 399 as identified by chromatogram data. One of these was predicted to be identified as non-cytb1 by HRM analysis of nucleotide 750; however, HRM analysis of nucleotide 750 failed to identify this sample. Thus, HRM analysis of nucleotide 399 was required to identify this sample as non-cytb1 (Table 2). When taking into account this information as well as the actual results of HRM analysis of the previous 116
135 Figure 1: Identification of C. felis cytb1 by HRM analysis of five cytb nucleotides. High resolution melt curves and differential fluorescence curves of 69 samples are shown for one of three replicate PCRs for each of the five nucleotide positions analyzed. Green and red boxes on melt curves indicate the pre- and post-melt regions (respectively) used for data normalization. RFU, relative fluorescence units. Red clusters of melt curves represent samples with the nucleotide sequence for cytb1 at each respective position, while clusters of other colors represent samples with SNPs at that location. Two samples (circled on difference curves and denoted with an asterisk) had anomalous clustering patterns that occurred at all nucleotides analyzed in at least one replicate PCR. Clustering patterns of these two samples could not be explained by chromatogram data but did not impact the sensitivity/specificity of the overall assay. A) Nucleotide position 750: The red cluster represents samples with adenine at nucleotide 750, while the green cluster represents samples with guanine at nucleotide 750. Blue and orange clusters represent samples with apparent heteroplasmy. Based on chromatogram data, the circled samples had a guanine at nucleotide 750, yet clustered outside of the expected spectrum of results (denoted by purple cluster). B) Nucleotide position 947: The red cluster represents samples with thymine at nucleotide 947, while the green cluster represents samples with cytosine at nucleotide 947. Based on chromatogram data, the circled samples had a thymine at nucleotide 947, yet clustered between clusters for thymine and cytosine (denoted by blue cluster). C) Nucleotide position 399: The red cluster represents samples with adenine at nucleotide 399, while the green cluster represents samples with guanine at nucleotide 399. The blue cluster represents samples with apparent heteroplasmy. Based on chromatogram data, the circled samples had an adenine at nucleotide 399, yet clustered with a sample confirmed to have heteroplasmy at nucleotide 399 (for replicate PCR data shown, only one sample demonstrates this clustering). D) Nucleotide position 274: The red cluster represent samples with cytosine at nucleotide 274, while green cluster represent samples with thymine at nucleotide 274. Blue and orange clusters represent samples with apparent heteroplasmy. Interestingly, two samples (circled on difference curves and denoted with an asterisk) had anomalous clustering patterns that occurred at all nucleotides analyzed in at least one replicate PCR. Based on chromatogram data, the circled samples had a cytosine at nucleotide 274, yet clustered outside of the expected spectrum of results (denoted by purple cluster). E) Nucleotide position 858: The red cluster represents samples with guanine at nucleotide 858, while the green cluster represents samples with an adenine at nucleotide 858. Notably, two samples (circled on difference curves and denoted with an asterisk) had anomalous clustering patterns that occurred at all nucleotides analyzed in at least one replicate PCR. Based on chromatogram data, the circled samples had a guanine at nucleotide 858, yet in 1/3 PCR replicates, the samples clustered outside of the expected spectrum of results (data from PCR replicate shown does not have this clustering pattern). 117

136 118
137 two nucleotides, it was now predicted that analysis of these three nucleotides together would identify 51/56 non-cytb1 samples (Specificity=91.1%). Actual HRM analysis was consistent with this prediction (Table 2). Based on chromatogram data, in silico analysis of nucleotides 750, 947, 399, and 274 together was predicted to identify 55/56 non-cytb1 samples (Specificity=98.2%). A total of eight samples had SNPs at nucleotide 274 as identified by chromatogram. Seven of these were predicted to be identified as non-cytb1 by HRM analysis of nucleotide 750; however, HRM analysis of nucleotide 750 failed to identify two of these seven samples. Thus, HRM analysis of nucleotide 274 was required to identify both of these samples as non-cytb1 (Table 2). When taking into account this information as well as the actual results of HRM analysis of the previous three nucleotides, it was now predicted that analysis of these four nucleotides together would identify 54/56 non-cytb1 samples (Specificity=96.4%). Actual HRM analysis was consistent with this prediction (Table 2). Finally, based on chromatogram data, in silico analysis of nucleotides 750, 947, 399, 274, and 858 together was predicted to identify 56/56 non-cytb1 samples (Specificity=100%). When taking into account the actual results of HRM analysis of the previous four nucleotides, it was now predicted that analysis of all five nucleotides together would identify 55/56 non-cytb1 samples (Specificity=98.2%). Actual HRM analysis was consistent with this prediction (Table 2). Agreement between HRM analysis and chromatogram data A total of 345 nucleotide positions were analyzed (5 nucleotide positions for 69 samples; see Table S1 in Supplemental Material). Agreement between HRM analysis and automated detection of secondary peaks on chromatograms was fair (Kappa=0.3564). HRM 119
, and failed to detect heteroplasmy at 6/345 nucleotide positions (1.7%). Discordant results at nucleotides 947, 399, 274, and 858 did not impair our ability to identify C. felis cytb1.")
.")
. These findings were confirmed by HRM analysis of plasmid mixtures simulating mitochondrial heteroplasmy at nucleotide 750 (Figure 3).")
138 analysis matched automated secondary peak detection at 307/345 nucleotide positions (89%), suggested the presence of additional heteroplasmy at 32/345 nucleotide positions (9.3%), and failed to detect heteroplasmy at 6/345 nucleotide positions (1.7%). Discordant results at nucleotides 947, 399, 274, and 858 did not impair our ability to identify C. felis cytb1. However, failure to detect heteroplasmy at nucleotide position 750 did impact our ability to identify C. felis cytb1 and therefore we further analyzed these discrepancies. HRM analysis of nucleotide 750 failed to detect relatively high A750G heteroplasmy that were identified by automated secondary peak detection (Figure 2). However, HRM analysis of nucleotide 750 detected low G750A heteroplasmy that were below the limit of automated secondary peak detection but could be identified by careful manual inspection of chromatograms (Figure 2). These findings were confirmed by HRM analysis of plasmid mixtures simulating mitochondrial heteroplasmy at nucleotide 750 (Figure 3). Representative sample where HRM analysis detected low G750A heteroplasmy at nucleotide 750 that is below the threshold of automated secondary peak detection: Figure 2. HRM analysis failed to detect relatively high A750G heteroplasmy yet was able to detect low G750A heteroplasmy at C. felis cytb nucleotide 750 in clinical samples. Red cluster is anticipated to have only adenine at nucleotide 750, green cluster is anticipated to have only guanine at nucleotide 750, and orange and blue clusters are anticipated to have heteroplasmy at nucleotide 750. Representative sample where HRM analysis failed to detect relatively high A750G heteroplasmy that is detected by automated secondary peak detection: 120
 Difference curve for triplicate analysis of clone mixtures and clinical samples B) Number of replicates per cluster for each sample analyzed When analyzed by HRM, two non-cytb1 samples repeatedly")
.")
139 A B Figure 3. Analysis of C. felis cytb clone mixtures confirms inability of HRM analysis to consistently discriminate heterogeneity in complex mixtures of genotypes. A) Difference curve for triplicate analysis of clone mixtures and clinical samples B) Number of replicates per cluster for each sample analyzed When analyzed by HRM, two non-cytb1 samples repeatedly had unexplained clustering patterns. For all nucleotide positions analyzed, these samples were either identified as a unique cluster or were placed on the fringe of their expected cluster (Figure 1A-1E). However, upon careful manual inspection of chromatograms, there was no evidence of SNPs at the targeted nucleotide or any other nucleotide positions within the amplicon. These results did not impact our ability to detect cytb1. DISCUSSION When treated with atovaquone and azithromycin (A&A), cats infected with Cytauxzoon felis cytb1 have had improved survival rates compared to cats infected with other 121
140 C. felis cytb genotypes (22). In this study we developed an assay that can rapidly identify C. felis cytb1 in clinical samples with 100% sensitivity and 98.2% specificity using a quantitative real-time PCR panel and HRM analysis. This test is intended to aid pet owners and clinicians in deciding whether or not to treat a cat for cytauxzoonosis with A&A, which can cost thousands of dollars. The cost of this assay for clients is likely to be comparable to currently offered molecular diagnostic panels ($ ). From the time the sample arrives in the laboratory, the assay can be completed in less than three hours, which is important for clinical decision making given the rapid disease progression and high mortality rate associated with acute cytauxzoonosis. While 100% of cats (8/8) infected with C. felis cytb1 survived when treated with A&A, it is important to note that over 50% of cats (20/37) infected with non-cytb1 genotypes also survived when treated with A&A (22). Therefore, while this assay will provide useful prognostic information for treating cats with cytauxzoonosis, other factors also need to be considered when implementing A&A treatment. First, a variety of factors besides C. felis cytb genotype may contribute to whether or not a cat will survive infection when treated with A&A. In a previous study that assessed the efficacy of A&A and imidocarb dipropionate for treatment of cats with acute cytauxzoonosis, cats that died tended to have higher parasitemia, higher serum bilirubin, and more severe leukopenia compared to cats that survived infection (14). However, treatment groups (A&A vs. imidocarb dipropionate) were not considered when these associations were assessed (14). Additionally, the majority of cats that died did so within 24 hours of admission to a veterinary clinic (14), suggesting that in some cases the disease may have already progressed beyond the point for any available treatment to be effective. 122
141 Second, given the heterogeneity of the C. felis cytb gene sequence (22), it is likely that uncharacterized C. felis cytb genotypes exist that have SNPs at nucleotide positions that are not assessed by this assay. Some of these uncharacterized genotypes may be A&Aresistant, yet falsely identified as C. felis cytb1 (type I error). Alternatively, some characterized and uncharacterized non-cytb1 genotypes may also be susceptible to A&A treatment. If future studies demonstrate that other C. felis cytb genotypes are also associated with improved survival rates or uncharacterized genotypes are falsely identified as cytb1, an extended PCR panel analyzing additional nucleotide positions can be developed. Additionally, the limitations of HRM analysis in detecting heteroplasmy impacted this assay. HRM analysis was unable to detect heteroplasmy in one non-cytb1 sample, resulting in one false positive result and a specificity of 98.2% (Table 2). Although the impact was minimal, this problem could also be solved by development of an extended PCR panel. Lastly, this study revealed the inability of HRM analysis to consistently discriminate heterogeneity in complex mixtures of genotypes. While these inconsistencies had minimal impact on the current study, the shortcomings of HRM analysis should be considered in the design and interpretation of any HRM assay. We encountered multiple scenarios in which there was disagreement between chromatogram evaluation and HRM analysis. First, in some cases HRM analysis was able to detect heteroplasmy that was present on chromatograms but below the limit of automated secondary peak detection (Figures 2-3, see Table S1 in Supplemental Material). This highlights the importance of manual chromatogram inspection for detection of heteroplasmy. Second, in other cases HRM analysis failed to detect heteroplasmy despite having clear evidence of heteroplasmy on chromatograms (Figures 2-123
142 3, see Table S1 in Supplemental Material). The reasons behind these discordant results are unclear; however, we hypothesize that the surrounding nucleotide sequences and overall GC content of the amplicon may impact HRM analysis. Third, in some cases HRM analysis detected heteroplasmy for which there was no chromatogram evidence even with manual inspection (Figure 1, see Table S1 in Supplemental Material). It is unclear whether HRM analysis was detecting actual heteroplasmy beyond the resolution of both automated and manual chromatogram inspection or if factors unrelated to amplicon sequence (e.g., individual sample ph, salt concentration, etc.) were skewing HRM results. Last, for some samples, HRM results differed between replicates (see Table S1 in Supplemental Material). Thus, if HRM analysis is to be performed on complex mixtures of genotypes, multiple analyses of the same sample may be required for accurate genotyping. In conclusion, this study describes an assay that can rapidly provide prognostic information to clients considering treatment options for cats with cytauxzoonosis. Through utilization of quantitative PCR and HRM analysis, we were able to develop an assay that identified C. felis cytb1 with 100% sensitivity and 98.2% specificity. Due to the variability of disease progression in individual cases of cytauxzoonosis, the heterogeneity of C. felis cytb sequence, and the inherent limitations of HRM analysis, multiple factors should be considered in the clinical decision-making process. ACKNOWLEDGEMENTS: Funding for this study was provided by the ALSAM Foundation. No conflicts of interest were present in this study. 124
143 REFERENCES: 1. Glenn BL, Stair EL Cytauxzoonosis in domestic cats: report of two cases in Oklahoma, with a review and discussion of the disease. J Am Vet Med Assoc 184: Hoover JP, Walker DB, Hedges JD Cytauxzoonosis in cats: eight cases ( ). J Am Vet Med Assoc 205: Wagner JE A fatal cytauxzoonosis-like disease in cats. J Am Vet Med Assoc 168: Ferris DH A progress report on the status of a new disease of American cats: cytauxzoonosis. Comp Immunol Microbiol Infect Dis 1: Birkenheuer AJ, Le JA, Valenzisi AM, Tucker MD, Levy MG, Breitschwerdt EB Cytauxzoon felis infection in cats in the mid-atlantic states: 34 cases ( ). J Am Vet Med Assoc 228: Jackson CB, Fisher T Fatal cytauxzoonosis in a Kentucky cat (Felis domesticus). Vet Parasitol 139: Andre MR, Adania CH, Machado RZ, Allegretti SM, Felippe PA, Silva KF, Nakaghi AC, Dagnone AS Molecular detection of Cytauxzoon spp. in asymptomatic Brazilian wild captive felids. J Wildl Dis 45: MacNeill AL, Barger AM, Skowronski MC, Lanka S, Maddox CW Identification of Cytauxzoon felis infection in domestic cats from southern Illinois. J Feline Med Surg doi: / x Haber MD, Tucker MD, Marr HS, Levy JK, Burgess J, Lappin MR, Birkenheuer AJ The detection of Cytauxzoon felis in apparently healthy freeroaming cats in the USA. Vet Parasitol 146: Rizzi TE, Reichard MV, Cohn LA, Birkenheuer AJ, Taylor JD, Meinkoth JH Prevalence of Cytauxzoon felis infection in healthy cats from enzootic areas in Arkansas, Missouri, and Oklahoma. Parasit Vectors 8: Walker DB, Cowell RL Survival of a domestic cat with naturally acquired cytauxzoonosis. J Am Vet Med Assoc 206: Meinkoth J, Kocan AA, Whitworth L, Murphy G, Fox JC, Woods JP Cats surviving natural infection with Cytauxzoon felis: 18 cases ( ). J Vet Intern Med 14:
144 13. Brown HM, Lockhart JM, Latimer KS, Peterson DS Identification and genetic characterization of Cytauxzoon felis in asymptomatic domestic cats and bobcats. Vet Parasitol 172: Cohn LA, Birkenheuer AJ, Brunker JD, Ratcliff ER, Craig AW Efficacy of atovaquone and azithromycin or imidocarb dipropionate in cats with acute cytauxzoonosis. J Vet Intern Med 25: Greene CE, Latimer K, Hopper E, Shoeffler G, Lower K, Cullens F Administration of diminazene aceturate or imidocarb dipropionate for treatment of cytauxzoonosis in cats. J Am Vet Med Assoc 215: , Greene CE Infectious diseases of the dog and cat, 2nd ed. W.B. Saunders, Philadelphia. 17. Mather MW, Henry KW, Vaidya AB Mitochondrial drug targets in apicomplexan parasites. Curr Drug Targets 8: Vaidya AB, Mather MW Mitochondrial evolution and functions in malaria parasites. Annu Rev Microbiol 63: Sakuma M, Setoguchi A, Endo Y Possible emergence of drug-resistant variants of Babesia gibsoni in clinical cases treated with atovaquone and azithromycin. J Vet Intern Med 23: Korsinczky M, Chen N, Kotecka B, Saul A, Rieckmann K, Cheng Q Mutations in Plasmodium falciparum cytochrome b that are associated with atovaquone resistance are located at a putative drug-binding site. Antimicrob Agents Chemother 44: Siregar JE, Syafruddin D, Matsuoka H, Kita K, Marzuki S Mutation underlying resistance of Plasmodium berghei to atovaquone in the quinone binding domain 2 (Qo(2)) of the cytochrome b gene. Parasitol Int 57: Schreeg ME, Marr HS, Tarigo J, Cohn LA, Levy MG, Birkenheuer AJ Pharmacogenomics of Cytauxzoon felis cytochrome b: Implications for atovaquone and azithromycin therapy in domestic cats with cytauxzoonosis. J Clin Microbiol 51: Kurelac I, Lang M, Zuntini R, Calabrese C, Simone D, Vicario S, Santamaria M, Attimonelli M, Romeo G, Gasparre G Searching for a needle in the haystack: comparing six methods to evaluate heteroplasmy in difficult sequence context. Biotechnol Adv 30: Sobenin IA, Mitrofanov KY, Zhelankin AV, Sazonova MA, Postnov AY, Revin VV, Bobryshev YV, Orekhov AN Quantitative assessment of heteroplasmy 126
145 of mitochondrial genome: perspectives in diagnostics and methodological pitfalls. Biomed Res Int 2014: White DJ, Wolff JN, Pierson M, Gemmell NJ Revealing the hidden complexities of mtdna inheritance. Mol Ecol 17: Wong LJ Diagnostic challenges of mitochondrial DNA disorders. Mitochondrion 7: Wilson RJ, Williamson DH Extrachromosomal DNA in the Apicomplexa. Microbiol Mol Biol Rev 61: Tong SY, Giffard PM Microbiological applications of high-resolution melting analysis. J Clin Microbiol 50: Vossen RH, Aten E, Roos A, den Dunnen JT High-resolution melting analysis (HRMA): more than just sequence variant screening. Hum Mutat 30: Montgomery JL, Sanford LN, Wittwer CT High-resolution DNA melting analysis in clinical research and diagnostics. Expert Rev Mol Diagn 10: Dobrowolski SF, Hendrickx AT, van den Bosch BJ, Smeets HJ, Gray J, Miller T, Sears M Identifying sequence variants in the human mitochondrial genome using high-resolution melt (HRM) profiling. Hum Mutat 30: Polakova KM, Lopotova T, Klamova H, Moravcova J High-resolution melt curve analysis: initial screening for mutations in BCR-ABL kinase domain. Leuk Res 32: van der Stoep N, van Paridon CD, Janssens T, Krenkova P, Stambergova A, Macek M, Matthijs G, Bakker E Diagnostic guidelines for high-resolution melting curve (HRM) analysis: an interlaboratory validation of BRCA1 mutation scanning using the 96-well LightScanner. Hum Mutat 30: Vondrackova A, Vesela K, Hansikova H, Docekalova DZ, Rozsypalova E, Zeman J, Tesarova M High-resolution melting analysis of 15 genes in 60 patients with cytochrome-c oxidase deficiency. J Hum Genet 57: Birkenheuer AJ, Marr H, Alleman AR, Levy MG, Breitschwerdt EB Development and evaluation of a PCR assay for the detection of Cytauxzoon felis DNA in feline blood samples. Vet Parasitol 137: Rogan WJ, Gladen B Estimating prevalence from the results of a screening test. Am J Epidemiol 107:
 by chromatogram and HRM analysis; HRM analysis was performed")
, suggested the presence of additional heteroplasmy at 32/345 nucleotide")

146 SUPPLEMENTAL MATERIAL Table S1. Summary of treatment, clinical outcome, cytb genotype, and discrepancies between HRM analysis and automated secondary peak detection. Five different nucleotide positions were analyzed for 69 samples (n=345 total different nucleotides analyzed) by chromatogram and HRM analysis; HRM analysis was performed in triplicate for all samples. HRM analysis matched automated secondary peak analysis at 307/345 nucleotide positions (89%), suggested the presence of additional heteroplasmy at 32/345 nucleotide positions (9.3%), and failed to detect heteroplasmy at 6/345 nucleotide positions (1.7%). Discrepancies between chromatogram and HRM analysis are denoted according to the key shown to the right. A&A=Atovaquone and Azithromycin; Imidocarb=Imidocarb Dipropionate; Percents under "manual chromatogram inspection" indicate percent of denoted nucleotide at nucleotide position as determined by manual inspection of chromatograms; Unique cluster=hrm cluster outside of expected spectrum of results 128
147 CHAPTER 5: PCR amplification of a multi-copy mitochondrial gene (cox3) for early detection of Cytauxzoon felis infection Megan E. Schreeg 1*, Henry S. Marr 1, Emily H. Griffith 2, Jaime L. Tarigo 1,3, David M. Bird 4, Mason V. Reichard 5, Leah A. Cohn 6, Michael G. Levy 1, Adam J. Birkenheuer 1 1. North Carolina State University, College of Veterinary Medicine, Raleigh, NC 2. North Carolina State University, College of Sciences, Raleigh, NC 3. University of Georgia, College of Veterinary Medicine, Athens, GA 4. North Carolina State University, College of Agriculture and Life Sciences, Raleigh, NC 5. Oklahoma State University, College of Veterinary Medicine, Stillwater, OK 6. University of Missouri, College of Veterinary Medicine, Columbia, MO *Responsible for all laboratory work, interpretation of statistical analyses, and majority of writing ABSTRACT Cytauxzoon felis is a tick-transmitted protozoan parasite that infects felids. Clinical disease caused by acute C. felis infection rapidly progresses in domestic cats, leading to high morbidity and mortality. Accurately diagnosing cytauxzoonosis as soon as possible during acute infection would allow for earlier initiation of antiprotozoal therapy which could lead to higher survival rates. Molecular detection of parasite rrna genes (18S) by PCR has previously been shown to be a sensitive method of diagnosing C. felis infections. Based on evidence from related apicomplexan species, we hypothesized that C. felis mitochondrial genes would exist at higher copy numbers than 18S and would be a more sensitive diagnostic target. In this study we have designed a PCR assay targeting the C. felis mitochondrial gene 129
148 cytochrome c oxidase subunit III (cox3). Herein we demonstrate that 1) the cox3 PCR can detect as low as 1 copy of DNA target and can detect C. felis in samples with known mitochondrial sequence heterogeneity, 2) cox3 copy number is increased relative to 18S in blood and tissue samples from acutely infected cats, and 3) the cox3 PCR is more sensitive than 18S PCR for detection of C. felis in samples from early acute infection. INTRODUCTION Cytauxzoonosis is an emerging and highly virulent disease in domestic and wild felids in North and South America caused by the tick-transmitted apicomplexan parasite Cytauxzoon felis (1-8). Although recent studies have documented cats surviving C. felis infection without evidence of clinical disease and/or receiving antiprotozoal treatment (9-13), morbidity and mortality remains high for cats with acute cytauxzoonosis presenting to veterinary hospitals. These cats initially show vague clinical signs of lethargy, anorexia, and pyrexia; if not treated, the disease rapidly progresses and within 2-5 days culminates in systemic inflammatory response syndrome, disseminated intravascular coagulation, multiorgan failure and death in most cats (14, 15). Survival of cats with acute cytauxzoonosis that do not receive antiprotozoal treatment remains low, with reported survival rates ranging from 0.2-3% (4, 5). However, recent advances in treatment combining atovaquone and azithromycin (A&A) have increased survival rates to over 60% (14). Given the rapid clinical course of the disease, it is vital to diagnose Cytauxzoon felis as soon as possible so treatment can be initiated. Unfortunately, accurate early diagnosis of cytauxzoonosis can be difficult given the initial nondescript clinical presentation. As such, a diagnosis of cytauxzoonosis often isn t reached until the disease is advanced. Current methods of C. felis identification include 130
149 blood smear analysis, fine needle aspirate of tissues, or molecular diagnostic techniques namely, polymerase chain reaction (PCR; 5, 16-19). Identification of ring-shaped merozoites or schizont-engorged leukocytes on peripheral blood smears or fine-needle aspirates of tissues are diagnostic for the disease. However, these methods are not as sensitive as PCR, particularly in the early stages of infection when parasitemia remains low, and greater than 50% of cats may progress to death without microscopic evidence of parasitemia (4, 5, 16). Currently, parasite genes encoding ribosomal RNA (18S and internal transcribed spacer, or ITS) in the chromosomal genome are the primary PCR targets utilized for C. felis identification (16, 18, 19). However, it has been demonstrated for related apicomplexan parasites (Babesia, Theileria, and Plasmodium) that genes encoded on the mitochondrial genome, including cytochrome c oxidase subunits (cox1, cox3) and cytochrome b (cytb), are more sensitive PCR targets than rrna genes (20-23). This is presumably due to the increased number of mitochondria (24-30) and mitochondrial genomes, the latter of which can be times more numerous than the chromosomal genome in a single parasite (31-35). Recent studies conducted by our lab revealed that clinical samples from cats with cytauxzoonosis often contain a complex mixture of C. felis cytb genotypes (36, 37), a condition known as mitochondrial heteroplasmy (38, 39). This suggests that a single C. felis organism has multiple mitochondrial genome copies. However, the C. felis mitochondrial genome copy number is unknown, and it remains undetermined if PCR assays targeting mitochondrial genes will be more sensitive for detection of C. felis in clinical samples than assays amplifying rrna genes. In this study, we have designed a PCR assay targeting C. felis cox3. We first demonstrate that this assay can detect as low as one copy of DNA target and is able to detect 131
150 C. felis in samples with known mitochondrial genome sequence variability, suggesting that cox3 is a suitable diagnostic PCR target for detecting a variety of C. felis strains. Second, we demonstrate that cox3 exists at a higher copy number than 18S in blood and tissues from acutely infected cats, but that cox3 and 18S copy numbers are similar in blood from cats that have been infected for over a year. Third, we demonstrate that a PCR targeting cox3 is more sensitive (p<0.05) than 18S at detecting C. felis in the early stage of infection. This new PCR assay targeting mitochondrial cox3 will aid in early detection of Cytauxzoon felis infection in cats with acute cytauxzoonosis. MATERIALS AND METHODS Samples For determining the ability of cox3 PCR to detect C. felis in samples with known mitochondrial sequence heterogeneity, blood samples (n=30) from acutely infected cats were available from a previous study (14). All samples had tested positive for the C. felis 18S PCR and had their C. felis cytb genotype characterized (14, 16, 37). For determining relative mitochondrial genome copy number, blood and/or fresh frozen tissue samples (spleen, liver, lymph node, or lung) were available from cats with acute cytauxzoonosis, cats recently recovered from acute cytauxzoonosis, or cats with chronic C. felis infection. All samples had been collected prior to anti-protozoal treatment. Cats with acute cytauxzoonosis were defined by the presence of clinical signs and identification of schizonts on blood smears and/or imprints of tissues harvested at necropsy; cats recently recovered from acute cytauxzoonosis had blood samples collected one month after the disappearance of clinical signs of acute infection (schizonts no longer detected); and cats with chronic C. felis infection had survived acute infection (schizonts no longer detected) and 132
151 were further subdivided based on whether cats had been chronically infected for less than 6 months or more than 1 year. For determination of PCR sensitivity, serially-collected blood samples were available from cats experimentally infected with C. felis. Seven purpose bred cats were obtained from a commercial supplier (Liberty Research, Inc., USA) and were between 11 and 18 months old at the time of the study. Cats were cared for according to the principles outlined in the National Institutes of Health Guide for the Care Use of Laboratory Animals and were housed in AAALAC accredited facilities within sealed, climate controlled isolation rooms with 12 hour light/dark cycles. All animal use was approved by the University of Missouri Animal Use and Care Committee (protocol number 7909). Cats were experimentally infected with C. felis using tick transmission as previously described (40-42). Briefly, fifty adult Amblyomma americanum ticks (25 females and 25 males), which had been fed to repletion as nymphs on a cat chronically infected with C. felis, were placed between the shoulder blades of anesthetized cats (dexdomitor (15-20 μg/kg) and acepromazine (0.1 mg/kg) IV). Stockinettes were secured around the thorax to prevent tick escape/removal, and cats were housed in individual cages placed inside a plastic basin lined with double sided tape to trap any ticks that left the cat. Blood was collected every other day from each cat starting at 7 days post infection (DPI). Blood was drawn from the jugular vein into EDTA tubes and was refrigerated until further processing. Prior to sampling, topical anesthetic cream (lidocaine/prilocaine emulsion, APP Pharmaceuticals, LLC, Schaumberg, IL) was applied to the skin over the venous access site. 133
152 DNA Isolation Total DNA was isolated from infected feline whole blood using an automated work station (Qiagen BioRobot M48 Robotic Workstation) and commercially available kit according to manufacturer s instructions (QIAGEN MagAttract DNA Mini M48 Kit, Qiagen Inc., Valencia, CA). DNA was eluted into a blood equivalent volume (e.g., DNA from 200 µl of blood was eluted into 200 µl of H2O). For tissue samples obtained from cats with naturally acquired cytauxzoonosis, DNA was isolated from approximately 0.1 grams of macerated tissue using a commercial kit according to kit instructions (QIAamp DNEasy Blood and Tissue Kit, Qiagen Inc., Valencia, CA). Assessment of PCR inhibitors in DNA samples All DNA samples were tested for the presence of PCR inhibitors by quantifying the cycle threshold (Ct) value of a spiked internal positive control (10,000 copies of puc19 cloning plasmid, Invitrogen Corporation, Carlsbad, CA) that should not be endogenously present in any feline DNA samples. Ct values of the internal positive control (IPC) were compared to the Ct value of an equivalent concentration of puc19 diluted in water. PCR inhibition was defined as the IPC crossing the cycle threshold two or more cycles after puc19 alone. DNA samples positive for PCR inhibition were diluted accordingly until the IPC Ct value was within two cycles of puc19 alone (Table 1). Primers (Forward: 5 - TGCTGCAAGGCGATTAAGTTG-3, Reverse: 5 - AATGTGAGTTAGCTCACTCATTAGGC-3 ) were designed to span the multiple cloning site of puc19 and create a 252 bp amplicon. Each 25 µl PCR reaction contained 12.5 µl of 2X SsoAdvanced SYBR Green Supermix (Bio-Rad Laboratories, Inc., Hercules, CA), 12.5 pmol of each primer, 10,000 copies of puc19, and 5 µl of DNA sample. Thermal cycling 134
153 conditions (CFX96, Bio-Rad Laboratories, Inc., Hercules, CA) consisted of an initial denaturation step at 98 C for 3.5 minutes followed by 45 amplification cycles (98 C for 20 seconds and 60 C for 30 seconds) and a melt curve step (60 C to 95 C, increasing at increments of 0.5 C every 5 seconds). Annealing/extension temperature was optimized to eliminate the production of non-specific amplicons as confirmed by melt curve analysis and gel electrophoresis of PCR products. Amplification efficiency of puc19 as determined by puc19 plasmid serial dilutions was 98.8%. Negative control consisted of water (no plasmid or feline DNA). cox3 PCR Primers were designed to produce a 190 base pair amplicon of C. felis cox3 (Forward: 5'-GCATATCTTCAAATTACAGATACAC-3', Reverse: 5'- CCAGTAACTGTTTAGTGTAGTTAAC-3', GenBank Accession KC207821). Each 25 µl PCR reaction contained 12.5 µl of 2X SsoAdvanced SYBR Green Supermix (Bio-Rad Laboratories, Inc., Hercules, CA), 12.5 pmol of each primer, and 5 µl of DNA sample. Thermal cycling conditions (CFX96, Bio-Rad Laboratories, Inc., Hercules, CA) consisted of an initial denaturation step at 98 C for 3.5 minutes followed by 45 amplification cycles (98 C for 20 seconds and 60 C for 30 seconds) and a melt curve step (60 C to 95 C, increasing at increments of 0.5 C every 5 seconds). Annealing/extension temperature was optimized to eliminate the production of non-specific amplicons and was confirmed by melt curve analysis and gel electrophoresis of PCR products. Positive controls consisted of DNA extracted from C. felis-infected feline whole blood; during initial PCR development, positive controls were sequenced bi-directionally to confirm cox3 amplification (Genewiz, South Plainfield, NJ). Negative controls consisted of uninfected feline DNA samples and water. 135
154 No amplicons were produced when negative controls were utilized as template during initial PCR development. To determine the efficiency and limit of detection of the assay, serial dilutions ( copies per reaction) of a cox3 plasmid were prepared and subjected to PCR amplification. Plasmids were initially prepared by directly cloning cox3 amplicon according to manufacturer s instructions (pgem-t Easy vector system, Promega, San Luis Obispo, CA) and transformation into TOP-10 competent E. coli (Invitrogen, Grand Island, NY). Plasmids containing inserts of the appropriate size were isolated (QIAprep Spin Miniprep Kit, Qiagen, Inc., Valencia, CA) and sequenced bi-directionally (MCLAB, South San Francisco, CA). 18S PCR A 284 base pair fragment of C. felis 18S rrna was amplified using previously described primers (Forward: 5 - GCGAATCGCATTGCTTTATGCT-3, Reverse: 5 - CCAATTGATACTCCGGAAAGAG-3 ; 16). PCR conditions and methods for determining PCR efficiency and limit of detection were as described for cox3. Positive and negative controls consisted of DNA extracted from C. felis-infected feline whole blood and no DNA (water), respectively. Determination of relative mitochondrial genome copy number 18S and cox3 PCRs were performed as described above in triplicate for all samples (n=35), and Ct values of C. felis mitochondrial genome (cox3) and chromosomal genome (18S) were assessed by quantitative real time PCR. Relative copy number difference between cox3 and 18S rrna was quantified using the 2 ΔCt method (43, 44). 136
155 Determination of sensitivity Individual C. felis cox3 and 18S molecules within the extracted DNA solution should have a Poisson distribution, and as such, testing a single sample multiple times will increase the probability of a detecting low concentration target molecules placed into a reaction (16, 43, 44). Therefore, starting with the earliest time point of sample collection (7 DPI), ten replicates of both 18S and cox3 PCRs were performed for each DNA sample. The number of positive 18S and cox3 PCR results were recorded for each sample. Serially collected samples were tested accordingly until a time point was reached where all ten replicates for both 18S and cox3 were positive for all cats. An equivalent number of negative controls with water alone as template were assessed concurrently with each DNA sample to test for amplicon contamination during PCR set up. All results were confirmed by both melt curve analysis and agarose gel electrophoresis. Any false positives (i.e., negative control water samples that were PCR positive) were further confirmed by bi-directional sequencing (Genewiz, South Plainfield, NJ). Stringent measures (i.e., practice of clean to dirty flow, preparation of single-use reagent aliquots, etc.) were taken to prevent amplicon contamination during PCR set up. Statistical Analysis To test for differences in relative mitochondrial genome copy number between blood samples acquired from cats during different stages of infection (acute, recently recovered, chronic < 6 months, chronic > 1 year), natural logs of the data points were fit to a linear mixed model where individual cat was set as a random effect and time point was set as a fixed effect. Natural logs of the data were used due to heteroskedasticity in residuals. 137
156 To test differences in relative mitochondrial genome copy number between the different tissues of acutely infected cats, natural logs of the data points were fit to a linear mixed model where individual cat was set as a random effect and tissue type was set as a fixed effect. Natural logs of the data were used to improve the fit of the model. In order to compare the performance of the cox3 and 18S PCRs, the number of positives from each of the two tests over DPI were assessed using logistic regression. DPI and test type (cox3 or 18s) were independent variables, and the response was the number of positives divided by the number of tests. False positive rate (number of water samples testing positive/total water samples tested) of each PCR was compared to the proportion of positive-testing samples for each date using a Chi-squared test for equivalent proportions. RESULTS cox3 and 18S have identical limits of detection and can detect different C. felis genotypes Efficiencies of cox3 (97.6%) and 18S (99.1%) were determined by serial dilutions of respective plasmids, and both assays were able to detect as few as one copy of their respective target. Additionally, the known sequence variability of C. felis cytochrome b (37), which is adjacent to cox3 in the mitochondrial genome, created concern that cox3 primers would be less sensitive at detecting C. felis than 18S due to the potential for sequence mismatches between cox3 primers and DNA template. To test if C. felis mitochondrial sequence variability would impact the sensitivity of the cox3 PCR, DNA samples (n=30) that represented the 30 characterized C. felis cytochrome b (cytb) genotypes were subjected to cox3 and 18S PCR. Both targets were successfully amplified and confirmed by melt curve analysis from all samples (Data not shown). 138
157 C. felis mitochondrial genome copy number (cox3) is increased relative to a chromosomal gene (18S) during acute cytauxzoonosis Mitochondrial genome copy number was increased nearly ten-fold compared to 18S in blood samples from acutely infected cats (Figure 1). Furthermore, there was a significant difference (p<0.05) in relative mitochondrial genome copy number in blood from acutely infected cats compared to blood from cats chronically infected for over a year (Figure 1). There was no significant difference (p>0.05) in relative mitochondrial genome copy in blood from acutely infected cats compared to blood from cats recently recovered from acute cytauxzoonosis or cats infected for less than six months (Figure 1). Because mitochondrial copy number decreased with chronic infection, we predicted that schizont life stage of C. Figure 1. Cytauxzoon felis mitochondrial genome copy number is increased in the blood of cats infected for less than 6 months but not cats infected for over a year. Mitochondrial genome copy number was calculated relative to 18S using the 2 ΔCt method. Bars represent mean mitochondrial copy number of 3-8 samples that were subjected to cox3 and 18S PCRs three times each. Standard error is denoted by error bars; different letters represent statistically significant differences (p<0.05) between means. 139

158 felis, which is only present during acute infection, may be contributing to the increased mitochondrial genome copy number observed in blood samples from acute infection. To test this, we evaluated mitochondrial genome copy number in tissue samples from acutely infected cats, which were confirmed by microscopy to contain schizonts and should be enriched in schizonts compared to peripheral blood obtained from the same cats at the same time. However, we found no significant difference (p>0.05) in mitochondrial genome copy number in blood, lymph node, spleen, or lung acquired from acutely infected cats (Figure 2). In fact, there was actually a significantly higher mitochondrial genome copy number in the blood compared to one schizont-laden tissue (liver; Figure 2). Figure 2. In acutely infected cats, Cytauxzoon felis mitochondrial genome copy number in blood samples is equal to or higher than that in tissue samples. Mitochondrial genome copy number was calculated relative to 18S using the 2 ΔCt method. Bars represent means of 4-10 samples that were subjected to PCRs three times each. Standard error is denoted by error bars; different letters represent statistically significant differences (p<0.05) between means. 140

159 cox3 PCR is more sensitive than 18S PCR at detecting early C. felis infection Because of the increased copy number of the mitochondrial genome observed in samples from acutely infected cats, we hypothesized that cox3 would be a more sensitive diagnostic target in detecting C. felis during early infection. The cox3 PCR was more sensitive (p<0.05) than the 18S PCR at detecting C. felis infection at both 7 DPI and 9 DPI (Figure 3). At 7 DPI, the cox3 PCR had a sensitivity of 24.59% (15/61 true positives detected) while the 18S PCR had a significantly lower (p<0.05) sensitivity of 4.92% (3/61 true positives detected). At 9 DPI, the cox3 PCR had a sensitivity of 92.86% (65/70 true positives detected) while the 18S PCR had a significantly lower (p<0.05) sensitivity of 70% (49/70 true positives detected). For each individual sample tested at 7 DPI and 9 DPI, cox3 PCR was either more sensitive or as equally sensitive as 18S PCR at detecting C. felis (Table Figure 3. Cytauxzoon felis cox3 is more sensitive than 18S at detecting infection at 7 and 9 DPI. Samples from 7 cats collected at different days post infection (DPI) were subjected to replicate cox3 and 18S PCR testing. Bars represent percentage of replicates testing positive for each test. *p<
 than the respective proportion of water samples testing positive.")
160 1). Both tests had a sensitivity of 100% by 11 DPI. There was a 0.05% false positive rate (1 positive test/200 true negatives) for both the cox3 and 18S PCRs. However, for each date the proportion of feline samples testing positive for both cox3 and 18S PCRs were significantly higher (p<0.05) than the respective proportion of water samples testing positive. Table 1. cox3 PCR detects C. felis with increased or equal sensitivity as 18S PCR for all samples. Samples were tested at 7, 9, and 11 DPI. Samples were diluted as needed as determined by PCR inhibitor testing. a Two samples were unable to be tested ten times for each PCR due to low volume of DNA sample b cox3 was more sensitive (p<0.05) than 18S at detecting C. felis infection at 7 and 9 DPI DISCUSSION This study describes a diagnostic PCR assay that targets a mitochondrial gene (cox3) for the detection of Cytauxzoon felis DNA in clinical samples. This PCR assay is able to detect a wide range of documented C. felis genotypes and is more sensitive than a 142
161 chromosomal gene (18S PCR assay) at detecting early C. felis infection (Figure 3, Table 1). While we believe that this assay will be clinically useful for identifying C. felis infection, there are a number of factors to consider when implementing this test as a diagnostic tool. First, while the cox3 assay was able to detect C. felis infection with a higher sensitivity than 18S at 7 and 9 DPI, no cats in this study became clinically ill until 11 DPI or later, a time point where the cox3 and 18S PCRs were equally sensitive in detecting C. felis. It is unclear whether the improved sensitivity of C. felis detection by cox3 over 18S will be clinically relevant, However, it appears possible that a naturally infected cat with cytauxzoonosis could test positive for cox3 and negative for 18S at the early onset of clinical disease. Second, a potential pitfall to using a mitochondrial gene as a diagnostic target is the prevalent heteroplasmy of the C. felis mitochondrial genome. While we were able to demonstrate that samples representing all of the previously characterized C. felis cytb genotypes tested positive for the cox3 assay, it is possible that other C. felis strains exist that may have mutations that prevent annealing of the cox3 primers. For this reason, it may be beneficial to target multiple loci (e.g. cox3, 18S, cytb (36), etc.) for the definitive identification of C. felis in a diagnostic setting. In this study we found that the C. felis cox3 copy number is increased approximately tenfold relative to 18S during acute infection but is present at a 1:1 ratio with 18S during chronic infections established for over a year (Figure 1). Therefore, the relative concentrations of cox3 and 18S could be beneficial for differentiating between acute and chronic Cytauxzoon felis infection, which is becoming more important given the increasing prevalence of C. felis infected cats with no history of clinical disease (13). 143
162 In addition to being diagnostically useful, the dynamic change of the mitochondrial genome copy number over time may shed light on the biology of C. felis. We initially considered the increased mitochondrial genome copy number seen during acute infection to be associated with schizogony, as is the case in related Theileria species (28, 29). However, relative mitochondrial genome copy numbers remained increased in samples where schizonts were not detectable, including the blood of cats recently recovered from cytauxzoonosis and cats chronically infected for less than six months (Figure 1). Additionally, schizont-laden tissues had equivalent or even fewer mitochondrial genome copies than blood samples where few or no schizonts were visualized (Figure 2). Therefore, the mere presence of schizonts does not appear to explain the increased mitochondrial genome copy number in acutely infected cats. The increased mitochondrial genome number observed during acute infection could be attributed to the parasite s higher metabolic demand. In related organisms, mitochondria and mitochondrial genome copy number are highest in metabolically active parasite stages, including intraerythrocytic trophozoites and gametocytes (24-27, 30, 45-47). Although these stages have not been specifically defined in the C. felis life cycle, features characteristic of trophozoites (presence of a food vacuole and multiple mitochondria (24, 26, 27)) and gametocytes (presence of multiple mitochondria (30)) have been previously described in acutely infected cats (48, 49), while intraerythrocytic replication (multiple organisms per cell, diversity in organism morphology) has been observed in both this study (Figure 4) and others (48, 49). However, these features have not been observed during chronic infection (18, 50) when mitochondrial genome copy number is decreased (Figure 1). These findings 144
 that detects early Cytauxzoon felis infection with higher sensitivity than 18S.")
163 suggest that different intraerythrocytic life stages of C. felis exist and should be further characterized in future studies. In conclusion, we have developed a PCR assay targeting a mitochondrial gene (cox3) that detects early Cytauxzoon felis infection with higher sensitivity than 18S. This test should aid in the detection of early cytauxzoonosis and could assist in differentiating acute a nd chronic infections. Figure 4. Cytauxzoon felis piroplasms are highly pleiomorphic during acute infection. Panels A-D are representative images from the blood smear of an acutely infected cat in this study. Samples from this cat were included in relative quantification of C. felis mitochondrial genome copy number. Organisms traditionally have been described as signet rings (Panel D, arrowhead), but can be amoeboid or elongated (*), may be replicating (#), and can vary in size. Occasionally multiple parasites (arrow) can be seen within a single erythrocyte. 145
164 ACKNOWLEDGEMENTS We would like to acknowledge the clinical and technical service of all individuals who aided in care of cats, including Meredith Sherrill, Hilton Outi, Jennifer Thomas, and Talli Hogen among others. This research was funded by a charitable organization which wishes to remain anonymous. There were no conflicts of interest in this study. REFERENCES 1. Wagner JE A fatal cytauxzoonosis-like disease in cats. J Am Vet Med Assoc 168: Glenn BL, Stair EL Cytauxzoonosis in domestic cats: report of two cases in Oklahoma, with a review and discussion of the disease. J Am Vet Med Assoc 184: Hoover JP, Walker DB, Hedges JD Cytauxzoonosis in cats: eight cases ( ). J Am Vet Med Assoc 205: Ferris DH A progress report on the status of a new disease of American cats: cytauxzoonosis. Comp Immunol Microbiol Infect Dis 1: Birkenheuer AJ, Le JA, Valenzisi AM, Tucker MD, Levy MG, Breitschwerdt EB Cytauxzoon felis infection in cats in the mid-atlantic states: 34 cases ( ). J Am Vet Med Assoc 228: Jackson CB, Fisher T Fatal cytauxzoonosis in a Kentucky cat (Felis domesticus). Vet Parasitol 139: Andre MR, Adania CH, Machado RZ, Allegretti SM, Felippe PA, Silva KF, Nakaghi AC, Dagnone AS Molecular detection of Cytauxzoon spp. in asymptomatic Brazilian wild captive felids. J Wildl Dis 45: MacNeill AL, Barger AM, Skowronski MC, Lanka S, Maddox CW Identification of Cytauxzoon felis infection in domestic cats from southern Illinois. J Feline Med Surg doi: / x Brown HM, Lockhart JM, Latimer KS, Peterson DS Identification and genetic characterization of Cytauxzoon felis in asymptomatic domestic cats and bobcats. Vet Parasitol 172:
165 10. Haber MD, Tucker MD, Marr HS, Levy JK, Burgess J, Lappin MR, Birkenheuer AJ The detection of Cytauxzoon felis in apparently healthy freeroaming cats in the USA. Vet Parasitol 146: Meinkoth J, Kocan AA, Whitworth L, Murphy G, Fox JC, Woods JP Cats surviving natural infection with Cytauxzoon felis: 18 cases ( ). J Vet Intern Med 14: Walker DB, Cowell RL Survival of a domestic cat with naturally acquired cytauxzoonosis. J Am Vet Med Assoc 206: Rizzi TE, Reichard MV, Cohn LA, Birkenheuer AJ, Taylor JD, Meinkoth JH Prevalence of Cytauxzoon felis infection in healthy cats from enzootic areas in Arkansas, Missouri, and Oklahoma. Parasit Vectors 8: Cohn LA, Birkenheuer AJ, Brunker JD, Ratcliff ER, Craig AW Efficacy of atovaquone and azithromycin or imidocarb dipropionate in cats with acute cytauxzoonosis. J Vet Intern Med 25: Conner BJ, Hanel, R.M., Brooks, M., Cohn, L.A., Birkenheuer, A.J Coagulation abnormalities in 5 cats with naturally occurring cytauxzoonosis. Journal of Veterinary Emergency and Critical Care doi:doi: /vec Birkenheuer AJ, Marr H, Alleman AR, Levy MG, Breitschwerdt EB Development and evaluation of a PCR assay for the detection of Cytauxzoon felis DNA in feline blood samples. Vet Parasitol 137: Holman PJ, Snowden KF Canine hepatozoonosis and babesiosis, and feline cytauxzoonosis. Vet Clin North Am Small Anim Pract 39: , v. 18. Brown HM, Latimer KS, Erikson LE, Cashwell ME, Britt JO, Peterson DS Detection of persistent Cytauxzoon felis infection by polymerase chain reaction in three asymptomatic domestic cats. J Vet Diagn Invest 20: Bondy PJ, Jr., Cohn LA, Tyler JW, Marsh AE Polymerase chain reaction detection of Cytauxzoon felis from field-collected ticks and sequence analysis of the small subunit and internal transcribed spacer 1 region of the ribosomal RNA gene. J Parasitol 91: Salem GH, Liu X, Johnsrude JD, Dame JB, Roman Reddy G Development and evaluation of an extra chromosomal DNA-based PCR test for diagnosing bovine babesiosis. Mol Cell Probes 13: Bilgic HB, Karagenc T, Shiels B, Tait A, Eren H, Weir W Evaluation of cytochrome b as a sensitive target for PCR based detection of T. annulata carrier animals. Vet Parasitol 174:
166 22. Haanshuus CG, Mohn SC, Morch K, Langeland N, Blomberg B, Hanevik K A novel, single-amplification PCR targeting mitochondrial genome highly sensitive and specific in diagnosing malaria among returned travellers in Bergen, Norway. Malar J 12: Buling A, Criado-Fornelio A, Asenzo G, Benitez D, Barba-Carretero JC, Florin- Christensen M A quantitative PCR assay for the detection and quantification of Babesia bovis and B. bigemina. Vet Parasitol 147: Conrad PA, Kelly BG, Brown CG Intraerythrocytic schizogony of Theileria annulata. Parasitology 91 ( Pt 1): Uchida TA, Yanagawa H, Mori T, Shiraishi S Ultrastructural observations of the intra-erythrocytic merozoite in Theileria sergenti. Trop Med Parasitol 36: Conrad PA, Denham D, Brown CG Intraerythrocytic multiplication of Theileria parva in vitro: an ultrastructural study. Int J Parasitol 16: Fawcett DW, Conrad PA, Grootenhuis JG, Morzaria SP Ultrastructure of the intra-erythrocytic stage of Theileria species from cattle and waterbuck. Tissue Cell 19: Jura WG, Brown CG, Rowland AC Ultrastructural characteristics of in vitro parasite-lymphocyte behaviour in invasions with Theileria annulata and Theileria parva. Vet Parasitol 12: Shaw MK, Tilney LG How individual cells develop from a syncytium: merogony in Theileria parva (Apicomplexa). J Cell Sci 101 ( Pt 1): Mehlhorn H, Schein, E, Ahmed, S. J Theileria, p In Kreier JP (ed), Parasitic Protozoa, 2 ed, vol 7. Academic Press, Inc., San Diego, CA. 31. Wilson RJ, Williamson DH Extrachromosomal DNA in the Apicomplexa. Microbiol Mol Biol Rev 61: Preiser PR, Wilson RJ, Moore PW, McCready S, Hajibagheri MA, Blight KJ, Strath M, Williamson DH Recombination associated with replication of malarial mitochondrial DNA. EMBO J 15: Joseph JT, Aldritt SM, Unnasch T, Puijalon O, Wirth DF Characterization of a conserved extrachromosomal element isolated from the avian malarial parasite Plasmodium gallinaceum. Mol Cell Biol 9: Vaidya AB, Arasu P Tandemly arranged gene clusters of malarial parasites that are highly conserved and transcribed. Mol Biochem Parasitol 22:
167 35. Hikosaka K, Tsuji N, Watanabe YI, Kishine H, Horii T, Igarashi I, Kita K, Tanabe K Novel type of linear mitochondrial genomes with dual flip-flop inversion system in apicomplexan parasites, Babesia microti and Babesia rodhaini. BMC Genomics 13: Schreeg ME, Marr HS, Tarigo JL, Cohn LA, Levy MG, Birkenheuer AJ Rapid high resolution melt analysis of Cytauxzoon felis cytochrome b to aid in the prognosis of cytauxzoonosis. J Clin Microbiol doi: /jcm Schreeg ME, Marr HS, Tarigo J, Cohn LA, Levy MG, Birkenheuer AJ Pharmacogenomics of Cytauxzoon felis Cytochrome b: Implications for Atovaquone and Azithromycin Therapy in Domestic Cats with Cytauxzoonosis. J Clin Microbiol 51: White DJ, Wolff JN, Pierson M, Gemmell NJ Revealing the hidden complexities of mtdna inheritance. Mol Ecol 17: Sobenin IA, Mitrofanov KY, Zhelankin AV, Sazonova MA, Postnov AY, Revin VV, Bobryshev YV, Orekhov AN Quantitative assessment of heteroplasmy of mitochondrial genome: perspectives in diagnostics and methodological pitfalls. Biomed Res Int 2014: Reichard MV, Thomas JE, Arther RG, Hostetler JA, Raetzel KL, Meinkoth JH, Little SE Efficacy of an imidacloprid 10 % / flumethrin 4.5 % collar (Seresto(R), Bayer) for preventing the transmission of Cytauxzoon felis to domestic cats by Amblyomma americanum. Parasitol Res 112 Suppl 1: Reichard MV, Edwards AC, Meinkoth JH, Snider TA, Meinkoth KR, Heinz RE, Little SE Confirmation of Amblyomma americanum (Acari: Ixodidae) as a vector for Cytauxzoon felis (Piroplasmorida: Theileriidae) to domestic cats. J Med Entomol 47: Reichard MV, Meinkoth JH, Edwards AC, Snider TA, Kocan KM, Blouin EF, Little SE Transmission of Cytauxzoon felis to a domestic cat by Amblyomma americanum. Vet Parasitol 161: Pfaffl MW Quantification strategies in real-time PCR, p In Bustin SA (ed), A-Z of quantitative PCR. International University Line, La Jolla, CA. 44. Pfaffl MW Relative quantification, p In Dorak MT (ed), Real-time PCR. Taylor and Francis Group, Oxford. 45. Krungkrai J The multiple roles of the mitochondrion of the malarial parasite. Parasitology 129:
168 46. Learngaramkul P, Petmitr S, Krungkrai SR, Prapunwattana P, Krungkrai J Molecular characterization of mitochondria in asexual and sexual blood stages of Plasmodium falciparum. Mol Cell Biol Res Commun 2: Petmitr S, Krungkrai J Mitochondrial cytochrome b gene in two developmental stages of human malarial parasite Plasmodium falciparum. Southeast Asian J Trop Med Public Health 26: Simpson CF, Harvey JW, Carlisle JW Ultrastructure of the intraerythrocytic stage of Cytauxzoon felis. Am J Vet Res 46: Kier AB, Wagner JE, Kinden DA The pathology of experimental cytauxzoonosis. J Comp Pathol 97: Lewis KM, Cohn LA, Birkenheuer AJ Lack of evidence for perinatal transmission of Cytauxzoon felis in domestic cats. Vet Parasitol 188:
169 CHAPTER 6: DNA vaccination of domestic cats against Cytauxzoon felis: Approach, outcome, and future directions Megan E. Schreeg 1 *, Henry S. Marr 1, Jaime L. Tarigo 1,2, Meredith K. Sherrill 3, Hilton K. Outi 3, Elizabeth H. Scholl 4, David M. Bird 4, Adam Vigil 5, Chris Hung 5, Rie Nakajima 5, Li Liang 5, Jennifer E. Thomas 6, Michael G. Levy 1, Mason V. Reichard 6, Philip L. Felgner 5, Leah A. Cohn 3, Adam J. Birkenheuer 1 1. North Carolina State University, College of Veterinary Medicine, Raleigh, NC 2. University of Georgia, College of Veterinary Medicine, Athens, GA 3. University of Missouri, College of Veterinary Medicine, Columbia, MO 4. North Carolina State University, College of Agriculture and Life Sciences, Raleigh, NC 5. University of California Irvine, School of Medicine, Irvine, CA 6. Oklahoma State University, College of Veterinary Medicine, Stillwater, OK *Assisted with vaccine production, animal care, and microarray preparation; responsible for data analysis and majority of writing ABSTRACT Cytauxzoonosis is an emerging disease of felids caused by the tick-transmitted apicomplexan parasite Cytauxzoon felis, an organism rapidly spreading across the United States. Cytauxzoonosis is particularly virulent for domestic cats, and most untreated cats that present to veterinary hospitals succumb to disease. The best treatment available (atovaquone and azithromycin) only confers a 60% survival rate and can cost thousands of dollars, limiting its widespread use. Disease prevention is currently limited to indoor confinement and prophylactic acaricides, but immunization against C. felis would be a more cost-effective 151
170 and pragmatic approach to preventing cytauxzoonosis. Because the parasite is not yet cultivable in vitro, we have searched the C. felis genome and identified 33 genes encoding candidate immunogens from which we have designed two DNA vaccines: an expression library vaccine including all candidates (CF-Library) and a vaccine containing the most well described candidate, cf76 (CF-1). We hypothesized that these vaccines could protect against infection and/or disease, and tested this hypothesis in a non-randomized, partially blinded pilot study. Regrettably, all vaccinated cats became infected and developed disease. However, cats vaccinated with CF-Library had the highest survival rate (100%), and one cat in this group demonstrated a comprehensive serological response to vaccination and required no therapeutic intervention. Serological response to vaccination varied highly between cats, suggesting that antigens were not uniformly delivered and/or expressed. Consequently, although neither vaccine as currently designed is recommended for further development, further investigation into the utility of these antigens in a different vaccine platform is warranted. INTRODUCTION Cytauxzoon felis is a tick-transmitted apicomplexan parasite that is the causative agent of cytauxzoonosis in domestic and wild felids in North and South America (1-8). Since its initial discovery in Missouri in 1976, Cytauxzoon felis has been increasingly recognized in feline species throughout the United States. To date, C. felis infection has been detected in domestic cats in 17 states (6, 8) and in bobcats in two additional states where the disease has yet to be observed in domestic cats (9, 10). Although the bobcat was originally considered the primary disease reservoir (11), recent evidence suggests that domestic cats can also be reservoirs, as more cats are surviving C. felis infection without any evidence of 152
171 clinical disease and/or history of antiprotozoal therapy (12-16). The reason behind this apparent change in disease epidemiology is unclear. However, the increased number of subclinically infected cats combined with widespread wild felid reservoirs and competent tick vectors (Amblyomma americanum and Dermacentor variabilis) is likely to enhance the rapid spread of C. felis infection in domestic cats throughout the Western Hemisphere. Despite the evolving epidemiology of C. felis infection in domestic cats, morbidity and mortality remain high for cats presenting to veterinary hospitals with acute cytauxzoonosis. Within 2-5 days of clinical presentation the disease culminates in systemic inflammatory response syndrome, disseminated intravascular coagulation, multi-organ failure and death in most cats (17, 18). Up to 99.8% of domestic cats infected with C. felis presented to veterinary hospitals die without treatment (4), and even with the best treatment currently available (atovaquone and azithromycin), mortality remains at 40% (17). Additionally, atovaquone and azithryomycin therapy in conjunction with supportive care can cost thousands of dollars, a factor which can be prohibitive for clients seeking treatment. When considering the high morbidity and mortality, lack of efficacious and affordable treatment options, and the rapid geographic spread of cytauxzoonosis, prevention of disease appears to be the optimal control strategy. Currently, prevention of cytauxzoonosis is completely dependent on application of prophylactic acaricides and keeping cats indoors (19). However, these prevention strategies may not be viable options for some cat owners, as an estimated million pet cats are allowed to roam outdoors in the United States (20, 21). Furthermore, effective acaricide prophylaxis may be limited by lack of owner compliance or cost of acaricides, as a recent study indicated as few as 38% of cats presenting to a veterinary teaching hospital received any form of tick prevention (22). 153
172 Therefore, we propose that immunization against C. felis would serve as a more practical method of prevention. Two previous studies have demonstrated that cats that mount an immune response against C. felis and survive are protected from clinical disease following subsequent infections, suggesting that an appropriately designed vaccine could also prevent illness (4, 23). Due to the inability to currently cultivate Cytauxzoon felis in vitro, our lab has obtained and mined the complete Cytauxzoon felis genome in an effort to search for vaccine candidates (24), a concept known as reverse vaccinology (25). To date, we have identified 33 different vaccine candidates that are differentially reactive against serum from C. felisinfected cats by protein microarray (24; Schreeg et al., unpublished data). However, it is unclear which (if any) of these candidates will induce a protective immune response against cytauxzoonosis, and solving this by synthesis and assessment of all 33 proteins may be laborintensive and cost-prohibitive. Alternatively, DNA vaccines can be manufactured at a low cost and with relative ease, and can incorporate a number of candidates if desired (26). For instance, expression library immunization (ELI) is a DNA vaccination strategy in which individuals are collectively immunized with a library of expression plasmids containing either a portion of or the entire genome of the infectious agent (27, 28). This approach can be utilized to induce a broad immune response against multiple antigens, or can be used as a high-throughput strategy to screen the genome of the infectious agent for vaccine candidates. Expression library vaccines have been shown to induce protection against a variety of protozoal infections in mice, including Plasmodium, Leishmania, and Trypanosoma species (26, 29-33). By extension, we hypothesize that a small-scale expression library vaccine containing 154
173 the 33 C. felis vaccine candidates will induce protection against cytauxzoonosis in cats experimentally infected with C. felis. The USDA recognizes five different levels of protection vaccines can provide against a pathogen: 1) prevention of infection, 2) prevention of disease, 3) aid in prevention of disease, 4) aid in control of disease, and 5) other claims. Because cytauxzoonosis is highly pathogenic, an effective vaccine against this disease would probably need to achieve one of the first two USDA label claims. The purpose of this pilot study was to assess if either of these claims could be met by vaccination with one of two DNA vaccines. The first vaccine is a small-scale expression library vaccine (CF-Library), while the second vaccine (CF-1) contains cf76, the most well-characterized of the candidates in the expression library vaccine (24). Herein we describe a pilot study evaluating efficacy of CF-Library and CF-1 vaccines against Cytauxzoon felis. While neither vaccine prevented infection or disease, CF-Library showed promise for aiding in disease control in some cats, and those candidates included in CF-Library warrant further investigation. MATERIALS AND METHODS DNA Isolation, RNA Isolation and cdna Synthesis Cytauxzoon felis genomic DNA was extracted from leukoreduced blood (Purecell NEO Neonatal High Efficiency Leukocyte Reduction Filter for Red Cell Aliquots, PALL Corp., Port Washington, NY) using the QIAamp DNA Blood Mini Kit according to manufacturer s instructions (Qiagen, Valencia, CA). RNA was extracted from liver tissue of a cat infected with C. felis using the ZR Tissue and Insect RNA Kit according to manufacturer s instructions (Zymo Research, Irvine, CA). Using this RNA as template, 155
174 cdna was synthesized with Taqman Reverse Transcription Reagents following the two step RT-PCR protocol according to manufacturer s instructions (Roche, Mannheim, Germany). Amplification of Vaccine Candidates A total of 33 different C. felis vaccine candidates were identified by protein microarray in previous studies (24; Schreeg et al., unpublished data). Candidates were amplified by polymerase chain reaction (PCR) using primers listed in Table S1 in Supplemental Material. Primers included 3 cassettes homologous to cloning vector to allow subsequent cloning via homologous recombination (Forward cassette: 5 - ACCGAGCTCGGCCACCATGG-3, Reverse cassette: 5 - CACACTGGACTAGTGGATCTA-3 ) Each 25 μl reaction contained 1 μl of genomic C. felis DNA template or cdna (see Table S1, Supplemental Material), 25 pmol of each primer, 5 nmol of deoxynucleoside triphosphates (dntps), 1.75 U of Expand high-fidelity enzyme mix, and a 1X concentration of Expand high-fidelity buffer with MgCl2 (Roche, Mannheim, Germany). Thermal cycling conditions consisted of an initial denaturation step at 94 C for 5 min, followed by 40 amplification cycles (94 C for 30 seconds, 55 C for 15 seconds, 50 C for 15 seconds, and 68 C for 4 minutes) and a final extension step at 68 C for 7 min (Techne Inc., Burlington, NJ). Annealing temperatures and primer concentrations were altered as needed to optimize amplification of individual candidates. Despite multiple attempts at optimizing PCR conditions, one candidate (Candidate 7) was recalcitrant to amplification and therefore was not included in the CF-Library vaccine. Linear Amplification of pvax1 pvax1 (Invitrogen, Carlsbad, CA), an expression vector designed for use in DNA vaccines, was utilized as the delivery vehicle for the DNA vaccine. The circular plasmid was 156
175 linearized by overnight digestion with BamHI at 37 ºC, and digest was purified using the QIAquick PCR Purification Kit and verified by agarose gel electrophoresis (Qiagen, Valencia, CA). This linearized pvax1 was used as template to generate the linear acceptor vector. Primers were designed (Forward: 5 - TAGATCCACTAGTCCAGTGTG-3, Reverse: 5'- CCATGGTGGCCGAGCTCGGTACCAAGC -3') to amplify the linear acceptor vector and incorporate a Kozak sequence upstream of the cloning site and a stop codon downstream of the coding site (see Table S1 in Supplemental Material). The 25 μl reaction consisted of 25 pmols of each primer, 5 pmol of dntps, U of U of Expand highfidelity enzyme mix, and a 1X concentration of Expand high-fidelity buffer with MgCl2 (Roche, Mannheim, Germany). Thermal cycling conditions consisted of an initial denaturation step at 95 C for 5 min, followed by 35 amplification cycles (94 C for 20 seconds, 50 C for 30 seconds, and 68 C for 3.5 minutes) and a final extension step at 72 C for 5 min (Techne Inc., Burlington, NJ). Homologous Recombination and Plasmid Isolation Amplified C. felis open reading frames (ORFs) were cloned into linearized pvax1 expression vector using in vivo homologous recombination adapted from previously described methods (34). Briefly, each transformation consisted of 10 μl of chemically competent TOP 10 E. coli cells (Invitrogen, Carlsbad, CA) and the linearized vector and ORF mixed in a 1:1 molar ratio (30 ng of pvax1 and the appropriate amount of ORF based on amplicon size). Mixture was incubated on ice for 30 minutes, heat shocked at 42 C for 1 minute, then incubated in 250 μl of super optimal broth with catabolite repression (SOC) at 37 ºC for 1 hour. After plating subsequent bacterial cultures on lysogeny broth (LB)/kanamycin (50 µg/ml) agar for overnight growth at 37 ºC, individual colonies were 157
176 screened by PCR using universal T7 and BGH reverse primers. Each 25 μl reaction consisted of 25 pmols of each primer, 5 nmol dntps, 37.5 nmol of MgCl2, 1.25 U AmpliTaq Gold DNA polymerase, and a 1X concentration of GeneAmp PCR Gold Buffer (Applied Biosystems, Carlsbad, CA). Thermal cycling conditions consisted of an initial denaturation step at 95 C for 5 min, followed by 30 amplification cycles (95 C for 20 seconds, 53 C for 20 seconds, and 72 C for 4 minutes) and a final extension step at 72 C for 5 min (Techne Inc., Burlington, NJ). PCR positive colonies were selected and grown in 2 ml liquid LB/kanamycin (50 µg/ml) overnight, and individual plasmids were purified from 1.5 ml of the resultant culture (QIAprep Spin Miniprep Kit, Qiagen, Inc., Valencia, CA). Plasmids were sequenced bi-directionally using T7 and BGH reverse primers to confirm successful homologous recombination; internal sequencing primers were designed and utilized as needed to obtain complete bi-directional sequencing (MCLAB, South San Francisco, CA). Production of Vaccines Individual colonies confirmed to contain plasmids of interest were selected and grown in 150 ml LB/kanamycin (50 µg/ml) as per Zyppy Plasmid Maxiprep Kit recommendations (Zymo Research, Irvine, CA). Plasmids were isolated according to manufacturer s instructions (Zyppy Plasmid Maxiprep Kit, Zymo Research, Irvine, CA) or were commercially prepared (Genewiz, South Plainfield, NJ). Multiple maxipreparations were performed as needed to attain the desired final amount of plasmid ( μg). Plasmids were further concentrated by ethanol precipitation as necessary and were resuspended in endotoxin-free tissue culture grade distilled water. Two different vaccines were developed in this study, referred to as CF-Library and CF-1. CF-Library consisted of 32 successfully cloned candidates, while CF-1 consisted of 158
, four cats vaccinated with CF-Library prior to infection (blue")
177 Figure 1. Cytauxzoon felis DNA vaccination pilot study design. A total of 18 cats were divided into four groups assessed in this study: three cats vaccinated with CF-1 prior to infection (red box), four cats vaccinated with CF-Library prior to infection (blue box), eight cats that were not vaccinated prior to infection (green box), and three cats that were neither vaccinated nor infected (gray box). The eight unvaccinated, infected cats were involved in a separate study testing the efficacy of novel chemotherapeutics against cytauxzoonosis. If it became evident that vaccines or chemotherapeutics were not halting the progression of disease, cats were additionally given atovaquone and azithromycin to attempt to prevent death; these cats are noted accordingly. A&A=atovaquone and azithromycin, 4X ABX=pradofloxacin, doxycycline, clindamycin, and metronidazole. both the C-terminal region and full-length C. felis cf76 (see Table S1 in Supplemental Material), the most well-characterized of the candidates (24). Each vaccine dosage contained a total of 1 mg of DNA. CF-1 contained 500 μg each of C-terminal region cf76 and fulllength cf76, while CF-Library contained μg of each of the 32 plasmids. Animals A total of 18 purpose bred cats were obtained from a commercial supplier (Liberty Research, Inc., USA) and were between 11 and 18 months old at the time of the study. Test groups are summarized in Figure 1. Briefly, seven cats received vaccines: three cats (331, 623, and 638) were immunized with CF-1 and four cats (77, 308, 339, and 835) were immunized with CF-Library. An additional eleven cats did not receive vaccines: eight of 159


178 these were infected with C. felis (positive controls for infection) and three were infested with C. felis-naïve ticks (negative controls for infection). Unvaccinated cats were also simultaneously enrolled in separate studies testing the efficacy of novel chemotherapeutics against cytauxzoonosis and the immune response to tick infestation and/or C. felis infection. Investigators were blinded to whether vaccinated cats received CF-Library or CF-1. Cats were cared for according to the principles outlined in the National Institutes of Health Guide for the Care Use of Laboratory Animals and were housed in AAALAC accredited facilities within sealed, climate controlled isolation rooms with 12 hour light/dark cycles. All animal use was approved by the University of Missouri Animal Use and Care Committee (protocol number 7909). Immunization Schedule The schedule of immunization and infection is summarized in Figure 2. Cats within both vaccine test groups received three immunizations intramuscularly (IM). Prior to each immunization the injection site was shaved. Cats were initially immunized with 1 mg of DNA (0.7 ml volume) delivered IM via needle and syringe in the left cranial thigh Figure 2. Timeline of vaccination, infection, and sample collection for vaccinated cats. 160
179 (quadriceps femoris muscle). The second immunization (1 mg of DNA in 0.7 ml) was administered 24 days after the initial immunization and was delivered IM via needle and syringe in the right cranial thigh (quadriceps femoris muscle). The third and final immunization (1 mg of DNA in 0.25 ml) was administered 44 days after the initial immunization and was delivered transdermally in the caudal medial right thigh via VET JET transdermal vaccination system (Merial, Inc., Athens, Georgia). When using the VET JET device, cats were sedated with dexdomitor (15 to 20 ug/kg) and acepromazine (0.1 mg/kg) delivered intravascularly (IV). Infection Model Vaccinated cats were infected with C. felis 59 days after initial immunization (Figure 2). Cats were experimentally infected with C. felis using tick transmission as previously described (19, 35, 36). Briefly, fifty adult Amblyomma americanum ticks (25 females and 25 males), which had been fed to repletion as nymphs on a cat chronically infected with C. felis, were placed between the shoulder blades of anesthetized cats (dexdomitor (15-20 μg/kg) and acepromazine (0.1 mg/kg) IV). Stocking nets were secured around the thorax to prevent tick escape/removal, and cats were housed in individual cages placed inside a plastic basin lined with double sided tape to trap any ticks that left the cat. Uninfected cats were identically infested with C. felis-naïve ticks. Ticks were removed from cats days post infestation, and the number of free and attached ticks as well as relative engorgement of ticks were recorded for each cat (see Table S2 in Supplemental Material). Monitoring and Clinical Evaluation Once infected, cats were inspected at least four times a day to watch for the appearance of clinical signs of cytauxzoonosis. Physical exams were performed daily, and 161
180 heart rate, respiratory rate, body temperature (measured rectally or by subcutaneous temperature chip (IPTT-3000, Bio Medic Data Systems, Seaford, DE)), and general appearance and attitude of cats were recorded 2-4 times daily. Starting at 12 days post-infection and finishing at the resolution of clinical symptoms or death, blood was collected for complete blood count (CBC) and serum biochemical profile. Frequency and extent of testing performed for each individual cat was dictated by severity of illness and discretion of attending veterinarian. For this reason and due to varying disease progression of individual cats, peaks and/or nadirs of clinical parameters were compared. Blood was collected into anticoagulant (EDTA) for complete blood count (CBC, Sysmex XT2000i V Automated Hematology Analyzer) and into red top tubes for serum biochemical profile (Olympus 400AUe Chemistry Analyzer). Blood was drawn from the jugular vein or from a subcutaneous jugular venous access port (VAP; CompanionPort CP4, Access Technologies, Skokie, IL). At least 15 minutes prior to sampling, topical anesthetic cream (lidocaine/prilocaine emulsion, APP Pharmaceuticals, LLC, Schaumberg, IL) was applied to the skin over the venous access site. To draw blood from cats with VAPs, an initial aliquot of blood (0.5 ml) containing heparin solution from the vascular access port was discarded then 3 ml of blood was aspirated for use in analysis. Supportive Care Supportive care was initiated on a case by case basis at the discretion of attending veterinarian. Specific therapies administered varied between cats and were tailored to the clinical needs of the individual (summarized in Table S3 of Supplemental Material). The following treatments were administered for at least one cat: subcutaneous (SQ) or intravenous (IV) fluid therapy (crystalloids (PlasmaLyte) with or without KCl, rates varied 162
181 by individual), whole blood or hyperimmune plasma transfusions, anticoagulant (200 U/kg SQ heparin TID; additional heparin added to fluids as needed), antiemetics (0.5-1 mg/kg SQ or IV maropitant q24h, mg/kg oral metoclopramide TID, and/or 1 mg/kg IV dolasetron q24h), appetite stimulants (3.75 mg (1/4 tablet) oral mirtazapine q72h), pain management (0.02 mg/kg SQ buprenorphine TID), nutritional support (commercial liquid diet (Clinicare) with or without potassium supplementation (Tumil K), volumes varied by individual), diuretics (2-2.5 mg/kg IV furosemide as needed), antacids (0.5 mg/kg IV famotidine q12-24h), anticonvulsants (1-3 mg/kg oral phenobarbital q12h), topical ointments (zinc oxide cream q12h), antimuscarinic eye drops (tropicamide, 1 drop q24h), ocular ointment (triple antibiotic ointment, q8h), and antibacterials (7.5 mg/kg oral pradofloxacin q24h). Antiprotozoal Therapy Antiprotozoal therapy was initiated on a case by case basis at the discretion of attending veterinarian. Three different treatments regimens were utilized in this study: two were experimental therapies that were administered to unvaccinated infected cats as a part of a different study (Figure 1). The experimental therapies were Coartem (20 mg artemether mg lumefantrine/tablet (Novartis), 2 tablets BID for 3 days) and a combination of 4 oral antibiotics, referred to through the study as 4X Abx (pradofloxacin 7.5 mg/kg q24h, doxycycline 10 mg/kg q12h, metronidazole 25 mg/kg q12h, clindamycin 10 mg/kg q12h until clinical improvement). For any cat that developed advanced cytauxzoonosis despite vaccination or experimental treatments, atovaquone and azithromycin (A&A, atovaquone (Mepron, GlaxoSmithKline) PO 15 mg/kg q8h for 10 days, azithromycin PO q24h for 10 days) was administered in an attempt to prevent death. 163
182 Assessment of Infection, Disease, and Survival Infection was confirmed by PCR amplification of parasite DNA (ITS-1 and/or 18S) as previously described (35, 37, 38) and if possible by identification of parasites via microscopic evaluation of blood smears. Cytauxzoonosis was defined as cats being febrile (body temperature > F) and having at least one of the following three biochemical/hematological abnormalities: hyperbilirubinemia (>0.3 mg/dl), neutropenia (<2.5 x10 3 segmented neutrophils/µl), or thrombocytopenia (<300 x10 3 segmented neutrophils/µl). Survival rates of groups were compared using the Freeman Halton extension (39) of the Fisher exact probability test with significance set at p < 0.05 (VassarStats, Poughkeepsie, NY). Protein Microarray A protein microarray containing 673 putative C. felis proteins, including all vaccine candidates, was available from a previous study (Schreeg et al., unpublished data). Briefly, all putative proteins were expressed (PXT7 vector) via in vitro transcription-translation (IVTT; RTS E. coli HY 100 kit, Roche Mannheim, Germany), and printed on nitrocellulosecoated glass FAST slides (Whatman, GE Healthcare Life Sciences, Pittsburgh, PA) using an Omni Grid 100 microarray printer (Genomic Solutions, Ann Arbor, MI) as previously described (40; Schreeg et al., unpublished data). Controls spotted onto chips included IVTT reactions without plasmid and purified IgG, and protein expression was confirmed by probing for polyhistidine and hemagglutinin tags included within the PXT7 vector. After the completion of the clinical trial portion of this study, the protein microarray was probed as previously described (40) with sera obtained from cats from each test group. Sera was collected from vaccinated cats at 5-7 different time points throughout the study (Figure 2). 164
183 Sera was collected from all unvaccinated infected cats and uninfected cats three months prior to infection and two months post infection. Additional sera was also available from specific pathogen free (SPF) cats. All sera had been frozen at -80 ºC until protein microarray analysis. Briefly, sera was first preadsorbed with E. coli lysate to reduce background reactivity against E. coli proteins from IVTT reactions. Arrays were rehydrated in Protein Array Blocking Buffer (Whatman, GE Healthcare Life Sciences, Pittsburgh, PA) then probed with the sera (diluted to 1:200 in blocking buffer) overnight at 4 C with constant agitation. Slides were then rinsed 10 mm Tris buffer (TBS, ph 8.0) containing 0.05% Tween-20 (TTBS), then incubated in biotin-conjugated, goat anti-cat immunoglobulin (anti-iggfcc, Jackson Immuno Research, West Grove, PA) diluted 1/200 in blocking buffer. After rinsing the slides, bound antibodies were detected by incubation with streptavidin conjugated SureLightH P-3 (Columbia Biosciences, Frederick, MD). The slides were then washed three times in TTBS and three times in TBS followed by a final water wash. The slides were air dried after brief centrifugation and analyzed using a Perkin Elmer ScanArray Express HT microarray scanner (Waltham, MA). RESULTS Infection and Disease Criteria All cats infested with C. felis positive ticks met the criteria for both infection and cytauxzoonosis (Table 1), with the majority (n=13/15 infected cats) exceeding the study criteria for cytauxzoonosis by exhibiting at least two of the three biochemical/hematological abnormalities that were considered disease-defining. No parasites were detected for cats infested with C. felis naïve ticks and cats did not develop cytauxzoonosis. Because these 165
184 negative control cats never became febrile or clinically ill, only limited laboratory data was collected over the course of the study (Table 1). Table 1. Summary of notable vital signs, laboratory findings, and clinical outcomes for individual cats. Biochemical/hematological values outside of reference range are underlined for each individual. For additional clinical observations and laboratory findings, see Supplemental Material and Appendix. Body Temperature Total Bilirubin Segmented PMN Platelet Nadir Infection Test Group Cat Cytauxzoonosis Supportive Antiprotozoal Peak ( F) Peak (mg/dl)* Nadir (x10 3 /µl)* (x10 3 Survival /µl)* Confirmed Criteria Met Care Therapy CF-Library Yes Yes No None Alive Yes Yes Yes A&A Alive Yes Yes Yes A&A Alive Yes Yes Yes A&A Alive CF Yes Yes Yes A&A Dead Yes Yes Yes A&A Dead Yes Yes Yes A&A Alive Unvaccinated, Yes Yes Yes 4X Abx + A&A Alive Infected Yes Yes Yes 4X Abx + A&A Alive Yes Yes Yes Coartem Alive Yes Yes Yes Coartem Alive Yes Yes Yes 4X Abx + A&A Dead Yes Yes Yes Coartem Alive Yes Yes Yes Coartem + A&A Dead Yes Yes Yes Coartem + A&A Alive Unvaccinated, 264 N/A N/A No N/A No None Alive Uninfected 276 N/A N/A No N/A No None Alive 880 N/A N/A No N/A No None Alive Segmented PMN=segmented neutrophil count, Band PMN=Band neutrophil count, A&A=atovaquone and azithromycin, 4X Abx=pradofloxacin, doxycycline, clindamycin, and metronidazole. *References ranges: Bilirubin: mg/dl, Segmented neutrophils: x 10 3 /µl, Platelets: x 10 3 /µl Supportive Care Supportive care for individual cats is summarized in Table 1 (see Table S3 in Supplemental Material for additional details on extent of supportive care administered to each individual). Only three out of four infected cats receiving CF-Library vaccine required supportive care. All infected cats receiving the CF-1 vaccine (3/3) and all unvaccinated infected cats (8/8) required some level of supportive care. None of the uninfected cats (n=3) showed signs of clinical illness and hence did not receive supportive care. Antiprotozoal Treatment Antiprotozoal treatment for individual cats is summarized in Table 1. Only three out of four infected cats receiving CF-Library vaccine required antiprotozoal treatment. All infected cats receiving the CF-1 vaccine (3/3) and all unvaccinated infected cats (8/8) 166
.")
185 required antiprotozoal treatment. None of uninfected cats (n=3) showed signs of clinical illness and hence did not receive antiprotozoal treatment. Survival Survival of individual cats is summarized in Table 1 and Figure 3. Survival rate of the infected cats vaccinated against CF-Library was 100% (4 out of 4 cats survived). Survival rate of the infected cats vaccinated against CF-1 was 33% (1 out of 3 cats survived). Survival rate of the unvaccinated infected cats was 75% (6 out of 8 cats survived). Survival rate of uninfected cats was 100% (3 out of 3 cats survived). However, there was no significant difference between survival rates of groups (p > 0.05). Figure 3. Cats vaccinated with CF-Library have a higher survival rate (100%) than other infected cats. Cats vaccinated with CF-1 had a 33% survival rate and unvaccinated, infected cats had a survival rate of 75%. Unvaccinated, uninfected cats also had a 100% survival rate (data not shown). DPI, days post infection. Serological Profiles of Individual Cats in Response to Vaccination Serological responses to vaccination varied highly between individuals (Figure 4). All three cats vaccinated with CF-1 had a serological response against partial and/or full- 167
.")

186 length cf76 after vaccination but prior to infection (Figure 4); however, only one of these cats survived (Figure 3, Table 1). Additionally, all three of these cats had serological responses to candidates that they were not vaccinated against prior to infection (Figure 4). Of the four cats vaccinated with CF-Library, one cat showed minimal serological response to Figure 4. Serological response of individual vaccinated cats to candidates throughout study. Arrays containing the 32 vaccine candidates included in CF-Library and in CF-1 were probed with sera samples from individual cats. Heat map shows normalized signal intensity with red strongest, white weakest, and gray intermediate; raw intensity values are listed within each spot. Rows denote 32 different candidates included in vaccines listed in descending order of reactivity; candidates were included in CF-1, while all listed candidates were included in CF-Library. Results are organized by individual cats (identified by number at top), and survival status of each cat is indicated as A (alive) or D (dead). Individual columns within each cat s array represent serum samples collected at different time points through study (labeled by the day in the study the sample was collected; refer to Figure 2 for timeline). There was no correlation between immunization protocol, individual reactivity, and survival for most cats, with the exception of Cat 77, who had widespread reactivity against all candidates in the CF-Library vaccine prior to infection and subsequently had milder disease and did not require supportive care or antiprotozoal therapy. 168
187 vaccination (308), two had serological reactivity against more than half of the candidates (339 and 835), and one had serological reactivity to nearly all candidates (77, Figure 4). This cat (77) was the only infected cat in the study that didn t require supportive care or antiprotozoal therapy (Table 1). Post-infection serum from all unvaccinated infected cats was highly reactive against putative C. felis antigens, while serum from all uninfected and SPF cats showed minimal reactivity (see Figure S1 in Supplemental Material). DISCUSSION The purpose of this pilot study was to assess the feasibility of preventing C. felis infection or clinical cytauxzoonosis by vaccination with CF-Library or CF-1. Neither vaccine in this study was able to meet these criteria, making both CF-Library and CF-1 unsuitable vaccines against cytauxzoonosis in their current forms using the described immunization routes and schedule. Despite this, it is interesting to note that cats vaccinated with CF-Library all survived (Figure 3), generally required less therapeutic intervention (Table 1 and Table S3 in Supplemental Material), and tended to have less extreme physical and laboratory abnormalities (lower body temperature peak, higher body temperature nadir, and fewer cats with bilirubinemia and neutropenia; Table 1 and Appendix). Collectively, this suggests that CF-Library may have aided in control of disease, meeting the fourth USDA vaccine label claim. However, the study was not intended to assess control of disease and stringent evaluation of the differences in clinical parameters between groups was not pursued. A major limitation of this study was the inability to screen individual serological responses prior to challenging cats with C. felis. Serological responses varied greatly from cat to cat, clearly suggesting differences in efficiency of vaccine delivery and/or antigen 169
188 expression (Figure 4). A number of steps are required for a DNA vaccine to be effective, including 1) delivery into a cell, 2) expression of the DNA as protein, 3) exposure of protein to appropriate immune cells, 4) development an innate immune response, and 5) development of a vaccine-specific adaptive immune response (see Figure 8 in Chapter 1; 26, 41). Deficiencies at any of these levels will likely lead to vaccine failure. Accordingly, the absent (308) or partial (331, 623, 638, 339, and 835) serological response of the majority of cats in this study to their respective vaccine suggests at least some level of vaccine failure. Furthermore, all cats vaccinated with CF-1 (331, 623, 638) had serological responses to antigens that were not included in CF-1 (e.g. Candidates 21, 22, 14). We hypothesize that this represents production of non-specific cross-reactive antibodies stimulated by the innate immune response to vaccination (42). Immunological cross-reactivity between hemoprotozoan parasites and a variety of other pathogens has been previously demonstrated (43-45), and many of these candidates (e.g. Candidates 21 and 22) are homologous to proteins expressed by other organisms that cats may have been previously exposed to (e.g. Coccidia). Such highly conserved antigens may not be ideal for inducing a C. felis-specific immune response and therefore further examination of the suitability of these candidates in a vaccine is needed. Collectively these serological results indicate that future vaccine trials should confirm uniform seroconversion of vaccinated cats prior to challenge. Other limitations present in this study include low number of cats, non-randomization of therapeutic intervention between groups, and inability to definitively standardize infectious dose of C. felis administered to cats. However, these limitations were of little consequence for analyzing study outcomes, as CF-Library and CF-1 clearly did not meet the intended vaccination standard of preventing infection or disease. 170
189 Interestingly, one cat (77) developed a comprehensive, albeit not robust, serological response to vaccination (Figure 4). Furthermore, on day 72 of the study (7 days post infection) serological response against all candidates decreased, suggesting that antibodies against these candidates had been depleted during early infection (Figure 4). This cat was the only individual in the study that required no supportive care or antiprotozoal therapy (Table 1). Collectively, this indicates that the 32 candidates in CF-Library may have afforded some level of disease protection for Cat 77. These results warrant further investigation into the incorporation of the 32 candidates in a vaccine, although a different delivery platform that could consistently induce an immune response for all vaccinated cat would be required. Potential alternative delivery platforms include a protein subunit vaccine or a recombinant virus expressing candidates. Unlike an expression library vaccine, however, these strategies would likely be limited to fewer proteins. Hence, further studies need to be done to narrow down to fewer candidates. This could include repeating expression library vaccination with smaller pools of candidates (28, 29) or further characterizing both cellular and humoral immune responses to candidates that multiple cats responded to in this study, such as cf76 (Candidates 32-33, Figure 4). In conclusion, this pilot study demonstrated that as designed and administered, CF- Library and CF-1 are unacceptable vaccines for prevention of cytauxzoonosis. However, CF-Library may have aided in disease control, suggesting that the 32 candidates may be promising for incorporation into a different vaccine platform that can efficiently stimulate an immune response in all cats. 171
190 ACKNOWLEDGEMENTS We would like to acknowledge the clinical expertise of Talli Hogen in the care of cats, the clinical pathology aid from Erin Burton in retrieving laboratory data, and the technical service of all veterinary technicians and students at the University of Missouri who aided in care of cats. This research was funded by a charitable organization which wishes to remain anonymous. There were no conflicts of interest in this study. REFERENCES 1. Glenn BL, Stair EL Cytauxzoonosis in domestic cats: report of two cases in Oklahoma, with a review and discussion of the disease. J Am Vet Med Assoc 184: Hoover JP, Walker DB, Hedges JD Cytauxzoonosis in cats: eight cases ( ). J Am Vet Med Assoc 205: Wagner JE A fatal cytauxzoonosis-like disease in cats. J Am Vet Med Assoc 168: Ferris DH A progress report on the status of a new disease of American cats: cytauxzoonosis. Comp Immunol Microbiol Infect Dis 1: Birkenheuer AJ, Le JA, Valenzisi AM, Tucker MD, Levy MG, Breitschwerdt EB Cytauxzoon felis infection in cats in the mid-atlantic states: 34 cases ( ). J Am Vet Med Assoc 228: Jackson CB, Fisher T Fatal cytauxzoonosis in a Kentucky cat (Felis domesticus). Vet Parasitol 139: Andre MR, Adania CH, Machado RZ, Allegretti SM, Felippe PA, Silva KF, Nakaghi AC, Dagnone AS Molecular detection of Cytauxzoon spp. in asymptomatic Brazilian wild captive felids. J Wildl Dis 45: MacNeill AL, Barger AM, Skowronski MC, Lanka S, Maddox CW Identification of Cytauxzoon felis infection in domestic cats from southern Illinois. J Feline Med Surg doi: / x Birkenheuer AJ, Marr HS, Warren C, Acton AE, Mucker EM, Humphreys JG, Tucker MD Cytauxzoon felis infections are present in bobcats (Lynx rufus) in 172
191 a region where cytauxzoonosis is not recognized in domestic cats. Vet Parasitol 153: Shock BC, Birkenheuer AJ, Patton LL, Olfenbuttel C, Beringer J, Grove DM, Peek M, Butfiloski JW, Hughes DW, Lockhart JM, Cunningham MW, Brown HM, Peterson DS, Yabsley MJ Variation in the ITS-1 and ITS-2 rrna genomic regions of Cytauxzoon felis from bobcats and pumas in the eastern United States and comparison with sequences from domestic cats. Vet Parasitol 190: Glenn BL, Kocan AA, Blouin EF Cytauxzoonosis in bobcats. J Am Vet Med Assoc 183: Haber MD, Tucker MD, Marr HS, Levy JK, Burgess J, Lappin MR, Birkenheuer AJ The detection of Cytauxzoon felis in apparently healthy freeroaming cats in the USA. Vet Parasitol 146: Walker DB, Cowell RL Survival of a domestic cat with naturally acquired cytauxzoonosis. J Am Vet Med Assoc 206: Rizzi TE, Reichard MV, Cohn LA, Birkenheuer AJ, Taylor JD, Meinkoth JH Prevalence of Cytauxzoon felis infection in healthy cats from enzootic areas in Arkansas, Missouri, and Oklahoma. Parasit Vectors 8: Meinkoth J, Kocan AA, Whitworth L, Murphy G, Fox JC, Woods JP Cats surviving natural infection with Cytauxzoon felis: 18 cases ( ). J Vet Intern Med 14: Brown HM, Lockhart JM, Latimer KS, Peterson DS Identification and genetic characterization of Cytauxzoon felis in asymptomatic domestic cats and bobcats. Vet Parasitol 172: Cohn LA, Birkenheuer AJ, Brunker JD, Ratcliff ER, Craig AW Efficacy of atovaquone and azithromycin or imidocarb dipropionate in cats with acute cytauxzoonosis. J Vet Intern Med 25: Conner BJ, Hanel, R.M., Brooks, M., Cohn, L.A., Birkenheuer, A.J Coagulation abnormalities in 5 cats with naturally occurring cytauxzoonosis. Journal of Veterinary Emergency and Critical Care doi:doi: /vec Reichard MV, Thomas JE, Arther RG, Hostetler JA, Raetzel KL, Meinkoth JH, Little SE Efficacy of an imidacloprid 10 % / flumethrin 4.5 % collar (Seresto(R), Bayer) for preventing the transmission of Cytauxzoon felis to domestic cats by Amblyomma americanum. Parasitol Res 112 Suppl 1: Salo AL, Stone E A survey of the views of US veterinary teaching faculty to owned cat housing practices. J Feline Med Surg doi: / x
192 21. Loyd KA, Hernandez SM, Abernathy KJ, Shock BC, Marshall GJ Risk behaviours exhibited by free-roaming cats in a suburban US town. Vet Rec 173: Gates MC, Nolan TJ Factors influencing heartworm, flea, and tick preventative use in patients presenting to a veterinary teaching hospital. Prev Vet Med 93: Motzel SL, Wagner JE Treatment of experimentally induced cytauxzoonosis in cats with parvaquone and buparvaquone. Vet Parasitol 35: Tarigo JL, Scholl EH, Bird DM, Brown CC, Cohn LA, Dean GA, Levy MG, Doolan DL, Trieu A, Nordone SK, Felgner PL, Vigil A, Birkenheuer AJ A novel candidate vaccine for cytauxzoonosis inferred from comparative apicomplexan genomics. PLoS One 8:e Rappuoli R Reverse vaccinology, a genome-based approach to vaccine development. Vaccine 19: Smooker PM, Rainczuk A, Kennedy N, Spithill TW DNA vaccines and their application against parasites--promise, limitations and potential solutions. Biotechnol Annu Rev 10: Talaat AM, Stemke-Hale K Expression library immunization: a road map for discovery of vaccines against infectious diseases. Infect Immun 73: Barry MA, Howell DP, Andersson HA, Chen JL, Singh RA Expression library immunization to discover and improve vaccine antigens. Immunol Rev 199: Shibui A, Nakae S, Watanabe J, Sato Y, Tolba ME, Doi J, Shiibashi T, Nogami S, Sugano S, Hozumi N Screening of novel malaria DNA vaccine candidates using full-length cdna library. Exp Parasitol 135: Alberti E, Acosta A, Sarmiento ME, Hidalgo C, Vidal T, Fachado A, Fonte L, Izquierdo L, Infante JF, Finlay CM, Sierra G Specific cellular and humoral immune response in Balb/c mice immunised with an expression genomic library of Trypanosoma cruzi. Vaccine 16: Piedrafita D, Xu D, Hunter D, Harrison RA, Liew FY Protective immune responses induced by vaccination with an expression genomic library of Leishmania major. J Immunol 163: Melby PC, Ogden GB, Flores HA, Zhao W, Geldmacher C, Biediger NM, Ahuja SK, Uranga J, Melendez M Identification of vaccine candidates for experimental visceral leishmaniasis by immunization with sequential fractions of a cdna expression library. Infect Immun 68:
193 33. Smooker PM, Setiady YY, Rainczuk A, Spithill TW Expression library immunization protects mice against a challenge with virulent rodent malaria. Vaccine 18: Doolan DL, Mu Y, Unal B, Sundaresh S, Hirst S, Valdez C, Randall A, Molina D, Liang X, Freilich DA, Oloo JA, Blair PL, Aguiar JC, Baldi P, Davies DH, Felgner PL Profiling humoral immune responses to P. falciparum infection with protein microarrays. Proteomics 8: Reichard MV, Edwards AC, Meinkoth JH, Snider TA, Meinkoth KR, Heinz RE, Little SE Confirmation of Amblyomma americanum (Acari: Ixodidae) as a vector for Cytauxzoon felis (Piroplasmorida: Theileriidae) to domestic cats. J Med Entomol 47: Reichard MV, Meinkoth JH, Edwards AC, Snider TA, Kocan KM, Blouin EF, Little SE Transmission of Cytauxzoon felis to a domestic cat by Amblyomma americanum. Vet Parasitol 161: Bondy PJ, Jr., Cohn LA, Tyler JW, Marsh AE Polymerase chain reaction detection of Cytauxzoon felis from field-collected ticks and sequence analysis of the small subunit and internal transcribed spacer 1 region of the ribosomal RNA gene. J Parasitol 91: Birkenheuer AJ, Marr H, Alleman AR, Levy MG, Breitschwerdt EB Development and evaluation of a PCR assay for the detection of Cytauxzoon felis DNA in feline blood samples. Vet Parasitol 137: Freeman GH, Halton JH Note on an exact treatment of contingency, goodness of fit and other problems of significance. Biometrika 38: Vigil A, Ortega R, Jain A, Nakajima-Sasaki R, Tan X, Chomel BB, Kasten RW, Koehler JE, Felgner PL Identification of the feline humoral immune response to Bartonella henselae infection by protein microarray. PLoS One 5:e Williams JA, Carnes AE, Hodgson CP Plasmid DNA vaccine vector design: impact on efficacy, safety and upstream production. Biotechnol Adv 27: Crampton SP, Voynova E, Bolland S Innate pathways to B-cell activation and tolerance. Ann N Y Acad Sci 1183: Gabrielsen AA, Jr., Jensen JB, Boland MT Identification and partial characterization of exoantigens derived from medium used to culture Plasmodium falciparum. Am J Trop Med Hyg 32:
194 44. Parra M, Liu X, Derrick SC, Yang A, Tian J, Kolibab K, Kumar S, Morris SL Molecular analysis of non-specific protection against murine malaria induced by BCG vaccination. PLoS One 8:e Herzenberg AM, Barta JR, Desser SS Monoclonal antibodies raised against coccidia and malarial parasites recognize antigenic epitopes found in lankesterellid and adeleorin parasites. J Parasitol 81:
195 SUPPLEMENTAL MATERIAL Table S1. PCR and cloning information for vaccine candidates. Vaccine Candidate Forward Primer Reverse Primer Size (BP) PCR Template 1 ACCGAGCTCGGCCACCATGGTGATGAGACTTGTAGGTAAAGT CACACTGGACTAGTGGATCTATGATCCCGTACCTTGTGAAA 1152 DNA 2 ACCGAGCTCGGCCACCATGGGTCGTAGACGTATTGT CACACTGGACTAGTGGATCTATTGTTGTGTGGTACGTAGAG 2004 DNA 3 ACCGAGCTCGGCCACCATGGTTGTTGGTATTTTAATCGG CACACTGGACTAGTGGATCTATTGTCTTGGGAGTGGGATTT 249 DNA 4 ACCGAGCTCGGCCACCATGGATCTAGGGTACATAAACGC CACACTGGACTAGTGGATCTAAATCGATTGAATTAATGTTA 2106 DNA 5 ACCGAGCTCGGCCACCATGGTGATGAAATCGTTCACATTTCT CACACTGGACTAGTGGATCTAGATCTTAAGTGTGAAAAGGA 762 DNA 6 ACCGAGCTCGGCCACCATGGTGATGAATTTTAAGGTAGTTGG CACACTGGACTAGTGGATCTATGTAGTTTGTCTCTGTTGTT 1128 DNA 7^ ACCGAGCTCGGCCACCATGGTGATGCAAGAGAATAAGTTATT CACACTGGACTAGTGGATCTATTGAGCAGGGGTAGAAGCGC 1114 DNA 8 ACCGAGCTCGGCCACCATGGTGATGCTAGTGTCTAATGACTA CACACTGGACTAGTGGATCTATACACCCCCAGGATATAAAC 1332 DNA 9 ACCGAGCTCGGCCACCATGGTGATGTTACAGTATATTGTACATCAATT CACACTGGACTAGTGGATCTAAATGAATTCCAAGTCATCATTAGATGG 1732 DNA 10 ACCGAGCTCGGCCACCATGGATTTTTCAATATTTGT CACACTGGACTAGTGGATCTAACTTAAAATAGATACTAGGA 804 DNA 11 ACCGAGCTCGGCCACCATGGTGATGACCATGAGTTACAGTAA CACACTGGACTAGTGGATCTATGATGTACCTGTAGGTTTCG 846 DNA 12 ACCGAGCTCGGCCACCATGGTGATGAAGAATGAGACATATAA CACACTGGACTAGTGGATCTAATTATATTCTGCTTTGCCCT 1772 DNA 13 ACCGAGCTCGGCCACCATGGAATCCTATTTTAGGAA CACACTGGACTAGTGGATCTAACCATCTTCCACAAATAGTC 2946 DNA 14 ACCGAGCTCGGCCACCATGGCATTTGATCTATTACA CACACTGGACTAGTGGATCTATTCATTTTTGTTGTGATCGT 180 DNA 15 ACCGAGCTCGGCCACCATGGTGAAGCAACAATTATTAAGAGT CACACTGGACTAGTGGATCTAAACATTGCAAGCTGAATATT 1686 DNA 16 ACCGAGCTCGGCCACCATGGTGATGAGACTATTACTCTTCAT CACACTGGACTAGTGGATCTATTGTATTTTACAGAAATCAT 1224 DNA 17 ACCGAGCTCGGCCACCATGGTGAGGCCTACCAGAAGTGATAA CACACTGGACTAGTGGATCTAATCACTTAGTAAATGAGCTT 270 DNA 18 ACCGAGCTCGGCCACCATGGTGATGACCAATGAAGATCATGG CACACTGGACTAGTGGATCTAGTTTAGGTGTTTTTTCATTC 3804 DNA 19 ACCGAGCTCGGCCACCATGGTGATTGTTACCAAGGCACCAGA CACACTGGACTAGTGGATCTATGGTATAGATGGTTTAGTAA 1499 DNA 20 ACCGAGCTCGGCCACCATGGTGAGGAAATTGAGGGACTCTGG CACACTGGACTAGTGGATCTAATTAGATTCATTTTCTTTGT 126 DNA 21 ACCGAGCTCGGCCACCATGGTGAAATTCAATCCCAATGCAA CACACTGGACTAGTGGATCTATAATGTTTTAGCTAGTAATGCATGCTC 759 DNA 22 ACCGAGCTCGGCCACCATGGTGATGTTGATTGAGGAGCCACT CACACTGGACTAGTGGATCTAATCTGAGAATGAGTTGAATG 1365 cdna 23 ACCGAGCTCGGCCACCATGGATGGTTTGTTGGGTAT CACACTGGACTAGTGGATCTATATGTTTTGTGGTGGATCTA 3186 DNA 24 ACCGAGCTCGGCCACCATGGTGATATTGGATGTTGAGCCTAA CACACTGGACTAGTGGATCTATACTATTTCATATGCACCTT 951 DNA 25 ACCGAGCTCGGCCACCATGGTGATGCTATGGATAGAATTTAA CACACTGGACTAGTGGATCTACTTTTCCATCTTCTTTGCGA 156 DNA 26 ACCGAGCTCGGCCACCATGGTAGAGAATAAGAAGGT CACACTGGACTAGTGGATCTATTCATCGAAATCTTTGTTTA 1422 DNA 27 ACCGAGCTCGGCCACCATGGTGATGACGGAAGCATTAAATAA CACACTGGACTAGTGGATCTATGCTGCTAATGTTAAAAATG 1416 DNA 28 ACCGAGCTCGGCCACCATGGTGATGAAAAATATAGTTGTTCT CACACTGGACTAGTGGATCTAATCACCAACTAGATGATGAG 1050 DNA 29 ACCGAGCTCGGCCACCATGGGTGAACAGCATGTAGAATCTAAAG CACACTGGACTAGTGGATCTAACAACATTCCATATCTACAGGATTTG 690 cdna 30 ACCGAGCTCGGCCACCATGGAATGCTTAGAATCCATAGG CACACTGGACTAGTGGATCTAATTCAATATGGATATGTTCA 192 DNA 31 ACCGAGCTCGGCCACCATGGTGAAATATAGGGAGCAACGTAA CACACTGGACTAGTGGATCTAGAAATAATTGAAATGGGGCT 795 DNA 32* ACCGAGCTCGGCCACCATGGAATTTTTATTAATGTTTGTGGTGC CACACTGGACTAGTGGATCTAAACTAGTGTTAATGATAAC 2277 DNA 33* ACCGAGCTCGGCCACCATGGAAGAACATTCAATAGCT CACACTGGACTAGTGGATCTAAACTAGTGTTAATGATAAC 832 DNA Red=pVAX1 cassette sequence, blue=kozak sequence, green=nucleotides for engineered stop codon (entire stop codon underlined) *Candidate 32=Full length cf76, Candidate 33=C-terminal region of cf76. In addition to being in CF-Library, these two candidate comprise CF-1 ^Not included in CF-Library (unable to amplify) 177
196 Table S2. Additional clinical observations and information for individual cats. Initial A&A Therapy Test Group Cat ID Attached Ticks Unattached Ticks Total Ticks Tick Comments DPI febrile DPI afebrile DPI most severe illness DPI improved Therapy Added CF-Library engorged; 18 unengorged N/A No /2 engorged; 1/2 unengorged N/A Yes /3 engorged; 2/3 unengorged N/A Yes /4 engorged; 2/4 unengorged N/A Yes CF /3 engorged; 2/3 unengorged N/A Yes 11 EUTHANIZED 16 EUTHANIZED /4 engorged; 2/4 unengorged N/A Yes 13 EUTHANIZED 16 EUTHANIZED engorged; 14 unengorged N/A Yes Unvaccinated, Many big ticks, few engorged 4X Abx Yes Infected Many engorged ticks 4X Abx Yes Unclear Lots of small ticks, few engorged Coartem No All small Coartem No N/A 4X Abx Yes EUTHANIZED Large ticks, unengorged Coartem No Lots of big ticks, some engorged Coartem Yes FOUND DEAD Mostly small ticks, couple engorged Coartem Yes Unvaccinated, N/A N/A N/A N/A N/A N/A N/A Uninfected N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A A&A=atovaquone and azithromycin, 4X ABX=pradofloxacin, doxycycline, clindamycin, and metronidazole. 178
197 Table S3. Summary of supportive care administered to individual cats. Cats vaccinated with CF-Library received less overall supportive care. Categories Test Group Cat Fluids Anticoagulant Analgesic Antiemetic Nutritional Support Appetite stimulant Transfusion Other Survival CF-Library 77 No No No No No No No No Alive 308 Yes (SQ) No No Yes No Yes Yes (hyperimmune plasma) Anticonvulsant (phenobarbital) Alive 339 Yes (SQ) No No No No Yes No No Alive 835 Yes (IV + Bolus) Yes Yes Yes Yes Yes Yes (whole blood) Antacid (famotidine) Alive CF Yes (IV) Yes Yes Yes Yes No No Ocular TAO + tropicamide* Dead 623 Yes (IV) Yes Yes Yes Yes No No No Dead 638 Yes (SQ/IV) Yes Yes Yes Yes No No No Alive Unvaccinated, 13 Yes (IV) Yes Yes Yes Yes Yes No Zinc oxide on perineum^ Alive Infected 47 Yes (IV) Yes Yes Yes Yes Yes Yes (whole blood) Antibacterial (pradofloxacin) #, zinc oxide on perineum^ Alive 59 Yes (IV) Yes Yes Yes Yes Yes Yes (hyperimmune plasma) Antibacterial (pradofloxacin) # Alive 84 Yes (IV) Yes Yes Yes No No No No Alive 577 Yes (IV) Yes Yes Yes Yes Yes No Diuretic (furosemide) Dead 775 Yes (IV) Yes Yes Yes No No No No Alive 797 Yes (IV + Bolus) Yes Yes Yes Yes Yes Yes (whole blood) No Dead 816 Yes (IV) Yes Yes Yes Yes (+ Tumil K) Yes Yes (hyperimmune plasma) Diuretic (furosemide) Alive Unvaccinated, 264 No No No No No No No No Alive Uninfected 276 No No No No No No No No Alive 880 No No No No No No No No Alive SQ=subcutaneous, IV=intravenous, Bolus=IV bolus of fluids administered in addition to baseline fluids, Anticoagulant=heparin (SQ or IV), analgesic=buprenorphine, antiemetic=maropitant, metoclopramide, and/or dolasetron, nutritional support=clinicare, Tumil K=potassium supplement, appetite stimulant=mirtazapine, TAO=triple antibiotic ointment *Used to treat blepharospasm in Cat 331 ^Used to treat rectal bleeding and swelling in Cats 13 and 47 # Pradofloxacin administered independent of 4X Abx treatment regimen for Cat

198 Figure S1. Cats infected with Cytauxzoon felis have a serological response to C. felis antigens but uninfected cats do not. Arrays containing a total of 673 putative C. felis antigens, including the 32 vaccine candidates, were probed with sera samples from unvaccinated cats; a selection of seroreactive antigens are shown. Heat map shows normalized signal intensity with red strongest, white weakest, and gray intermediate. Rows denote antigens, while columns denote individual serum samples. Unvaccinated cats infected with C. felis are reactive to antigens post-infection but not preinfection. In contrast, unvaccinated cats that were infested with naïve ticks show minimal response to C. felis antigens after ticks are removed, indicating that response is specific to C. felis infection. SPF cats also are minimally responsive to C. felis antigens. 180
199 CHAPTER 7: Conclusions In this study, we utilized mitochondrial and chromosomal genomes to acquire critical information about Cytauxzoon felis, a highly virulent parasite of domestic cats that is uncultivable and for which little information is known. First, using mitochondrial genome sequences and structures we demonstrated that Cytauxzoon felis and Theileria species are in fact a single taxonomic group, and proposed that the common bond within this group was a highly developed ability to manipulate infected host cells. Second, we investigated if there was a link between treatment efficacy of atovaquone and azithromycin (A&A) and the gene sequence of the drug target, mitochondrial C. felis cytochrome b (cytb). We discovered that the C. felis cytb gene was highly heteroplasmic, yet identified a C. felis cytochrome b genotype (cytb1) that was associated with an increased survival rate in cats A&A-treated cats. Third, we developed a prognostic assay that was able to detect C. felis cytb1 with 100% sensitivity and 98.2% specificity. This assay is cost-effective, can be rapidly performed, and will aid clinicians and pet owners in making treatment decisions. Fourth, we demonstrated that C. felis has multiple mitochondrial genome copies relative to 18S, and used this fact to develop a diagnostic assay targeting mitochondrial cox3 that is more sensitive than 18S for early detection of infection. Last, we developed two DNA vaccines comprised of candidates identified by mining the C. felis chromosomal genome and tested the efficacy of these vaccines in a pilot study. Although the vaccines were unable to prevent cytauxzoonosis, this study set the groundwork for future research into developing a subunit vaccine against C. felis. In conclusion, we have resourcefully increased our knowledge base of the biology and clinical approach to Cytauxzoon felis by use of mitochondrial and chromosomal genomics. 181
200 APPENDIX 182
201 CLINICAL DATA FOR CATS IN CYTAUXZOON FELIS DNA VACCINATION PILOT STUDY Table 1. Daily body temperature ( F) peaks (A) and nadirs (B) of individual cats. Daily averages are listed at the bottom of each group and highlighted in gray. Dates highlighted in yellow denote when cats were clinically ill. Peak and nadir body temperatures for each individual during illness are noted in the last column ( Illness Peak/Nadir ) and group averages of these values are indicated in bold within the average row. All temperatures were measured rectally or with a subcutaneous temperature chip (IPTT-3000, Bio Medic Data Systems, Seaford, DE) with the exception of the dates outlined in black for Cat 47, which were measured in the axilla. These axillary measurements were not included in assessment of peak and nadir temperatures. DPI, days post infection. Dates where data was not collected are noted ( N/A ). A) DPI ILLNESS GROUP CAT PEAK CF-Library N/A N/A N/A N/A N/A N/A N/A N/A AVERAGE N/A N/A CF N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A AVERAGE N/A N/A Unvaccinated, Infected N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A AVERAGE
202 B) DPI ILLNESS GROUP CAT NADIR CF-Library N/A N/A N/A N/A N/A N/A N/A N/A 99.6 AVERAGE N/A N/A CF N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 100 AVERAGE N/A N/A Unvaccinated, Infected N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 98.3 AVERAGE
203 A Figure 1. Daily body temperature peaks (A) and nadirs (B) of individual cats over time. Each cat has a unique marker, while groups are denoted accordingly by different colored lines (CF- Library=Blue, CF-1=Red, Unvaccinated, infected=green). DPI, days post infection. Notably, cats from both vaccine groups tend to have a slightly lower maximum body temperature. B 185

204 A B Figure 2. Average daily body temperature peaks (A) and nadirs (B) of different test groups over time. DPI, days post infection. Figure 3. Overall average body temperature peaks and nadirs of different test groups. Notably, unvaccinated infected cats tended to have higher body temperature peaks and lower body temperature nadirs than vaccinated cats. 186
205 Table 2. Daily heart rate (beats per minute) peaks (A) and nadirs (B) of individual cats. Daily averages are listed at the bottom of each group and highlighted in gray. Dates highlighted in yellow denote when cats were clinically ill. Peak and nadir heart rates for each individual during illness are noted in the last column ( Illness Peak/Nadir ) and group averages of these values are indicated in bold within the average row. DPI, days post infection. Dates where data was not collected are noted ( N/A ). A) DPI ILLNESS GROUP CAT PEAK CF-Library 77 N/A N/A N/A N/A N/A N/A N/A N/A 220 N/A N/A N/A N/A N/A N/A N/A N/A N/A 240 AVERAGE N/A N/A CF N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 260 AVERAGE N/A N/A Unvaccinated, Infected N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 240 AVERAGE
206 B) DPI ILLNESS GROUP CAT NADIR CF-Library 77 N/A N/A N/A N/A N/A N/A N/A N/A 220 N/A N/A N/A N/A N/A N/A N/A N/A N/A 180 AVERAGE N/A N/A CF N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 180 AVERAGE N/A N/A Unvaccinated, Infected N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 168 AVERAGE

207 A Figure 4. Daily heart rate peaks (A) and nadirs (B) of individual cats over time. Each cat has a unique marker, while groups are denoted accordingly by different colored lines (CF-Library=Blue, CF-1=Red, Unvaccinated, infected=green). DPI, days post infection. B 189

208 A B Figure 5. Average daily heart rate peaks (A) and nadirs (B) of different test groups over time. DPI, days post infection. Figure 6. Overall average heart rate peaks and nadirs of different test groups. 190
209 Table 3. Daily respiratory rate (breaths per minute) peaks (A) and nadirs (B) of individual cats. Daily averages are listed at the bottom of each group and highlighted in gray. Dates highlighted in yellow denote when cats were clinically ill. Peak and nadir respiratory rates for each individual during illness are noted in the last column ( Illness Peak/Nadir ) and group averages of these values are indicated in bold within the average row. DPI, days post infection. Dates where data was not collected are noted ( N/A ). A) DPI ILLNESS GROUP CAT PEAK CF-Library N/A N/A N/A N/A N/A N/A N/A N/A 70 AVERAGE N/A N/A CF N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 60 AVERAGE N/A N/A Unvaccinated, Infected N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 60 AVERAGE
210 B) DPI ILLNESS GROUP CAT NADIR CF-Library N/A N/A N/A N/A N/A N/A N/A N/A 32 AVERAGE N/A N/A CF N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 32 AVERAGE N/A N/A Unvaccinated, Infected N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 20 AVERAGE
. DPI, days post infection.")
211 A Figure 7. Daily respiratory rate peaks (A) and nadirs (B) of individual cats over time. Each cat has a unique marker, while groups are denoted accordingly by different colored lines (CF-Library=Blue, CF-1=Red, Unvaccinated, infected=green). DPI, days post infection. Notably, unvaccinated cats and cats vaccinated with CF-1 tend to have higher respiratory rate peaks around the initiation of illness (Days 12-13). B 193
212 A B Figure 8. Average daily respiratory rate peaks (A) and nadirs (B) of different test groups over time. DPI, days post infection. Figure 9. Overall average respiratory rate peaks and nadirs of different test groups. Cats vaccinated with CF- Library tended to have lower peak respiratory rates, perhaps indicating reduced stress or reduced dyspnea. 194
213 Table 4. Albumin values (g/dl) for individual cats over the course of illness. Number of albumin measurements taken for each individual over the course of illness was dictated by severity of disease; dates where no data was collected are denoted ( N/A ). Peak and nadir albumin measurements were tabulated for each individual, and averages of each were calculated for each group. DPI, days post infection. DPI AVG PEAK AVG NADIR GROUP CAT PEAK NADIR OF GROUP OF GROUP CF-Library 77 N/A 3 N/A 2.9 N/A N/A N/A 2.8 N/A N/A 2.9 N/A 2.7 N/A N/A N/A N/A N/A N/A 3.1 N/A 2.6 N/A N/A N/A N/A N/A N/A 2.9 N/A 2.1 N/A N/A N/A 2.7 N/A CF N/A 2.7 N/A 2.2 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 2.6 N/A 2.1 N/A N/A N/A N/A N/A Unvaccinated, N/A 3 N/A 2.4 N/A N/A 2.7 N/A N/A Infected N/A 2.7 N/A 2.5 N/A N/A N/A N/A N/A 2 N/A N/A N/A N/A 2.7 N/A 2.5 N/A 2.8 N/A N/A N/A N/A N/A N/A N/A 1.7 N/A N/A N/A N/A N/A N/A N/A 2.7 N/A 2.5 N/A N/A 2.8 N/A N/A N/A 2.8 N/A 2.6 N/A N/A N/A 1.8 N/A N/A N/A N/A N/A N/A N/A N/A
214 Figure 10. Albumin levels of individual cats over time. Each cat has a unique marker, while groups are denoted accordingly by different colored lines (CF- Library=Blue, CF-1=Red, Unvaccinated, infected=green). DPI, days post infection. Figure 11. Overall average albumin peaks and nadirs of different test groups. Cats vaccinated with CF-Library tended to have higher albumin levels. 196
215 Table 5. Plasma protein values (g/dl) for individual cats over the course of illness. Number of plasma protein measurements taken for each individual over the course of illness was dictated by severity of disease; dates where no data was collected are denoted ( N/A ). Peak and nadir plasma protein measurements were tabulated for each individual, and averages of each were calculated for each group. Note that measurements were available for unvaccinated, uninfected cats at two time points. DPI, days post infection. DPI AVG PEAK AVG NADIR GROUP CAT PEAK NADIR OF GROUP OF GROUP CF-Library N/A N/A 7.6 N/A N/A N/A 8.2 N/A N/A N/A 7.0 N/A N/A N/A N/A N/A N/A N/A 6.8 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 7.2 N/A CF N/A N/A 5.9 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 6.2 N/A N/A N/A N/A N/A Unvaccinated, N/A 6.4 N/A 6.8 N/A N/A 8 N/A N/A Infected N/A 7.4 N/A 7.4 N/A N/A N/A N/A 6.1 N/A N/A N/A N/A 7.0 N/A 7.8 N/A N/A N/A N/A N/A N/A 5 N/A N/A N/A N/A N/A N/A N/A 6.6 N/A 6.1 N/A N/A 7.2 N/A N/A 6.9 N/A 7.2 N/A N/A N/A 5.5 N/A N/A N/A N/A N/A N/A N/A Unvaccinated, N/A N/A 7.4 N/A N/A N/A N/A N/A N/A N/A Uninfected N/A N/A 6.6 N/A N/A N/A N/A N/A N/A N/A N/A 7.0 N/A N/A N/A N/A N/A N/A N/A N/A
.")
216 Figure 12. Plasma protein levels of individual cats over time. Each cat has a unique marker, while groups are denoted accordingly by different colored lines (CF- Library=Blue, CF-1=Red, Unvaccinated, infected=green, Unvaccinated, uninfected=black). DPI, days post infection. Figure 13. Overall average plasma protein peaks and nadirs of different test groups. Cats vaccinated with CF- Library tended to have higher plasma protein nadirs compared to other infected cats. 198
217 Table 7. Total bilirubin values (mg/dl) for individual cats over the course of illness. Number of bilirubin measurements taken for each individual over the course of illness was dictated by severity of disease; dates where no data was collected are denoted ( N/A ). Peak and nadir total bilirubin measurements were tabulated for each individual, and averages of each were calculated for each group. DPI, days post infection. DPI AVG PEAK AVG NADIR GROUP CAT PEAK NADIR OF GROUP OF GROUP CF-Library 77 N/A 0.1 N/A 0.1 N/A N/A N/A 1.9 N/A N/A 0.1 N/A 0.1 N/A N/A N/A N/A N/A N/A 0.1 N/A 0.2 N/A N/A N/A N/A N/A N/A 0.1 N/A 3.7 N/A N/A N/A 1.8 N/A CF N/A 0.1 N/A 1.5 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 0.1 N/A 2.7 N/A N/A N/A N/A N/A Unvaccinated, N/A 1.1 N/A 2.3 N/A N/A 0.9 N/A N/A Infected N/A 0.1 N/A 2 N/A N/A N/A N/A N/A 3.4 N/A N/A N/A N/A 0.1 N/A 0.2 N/A 0.1 N/A N/A N/A N/A N/A N/A N/A 4.8 N/A N/A N/A N/A N/A N/A N/A 2.1 N/A 0.4 N/A N/A 0.3 N/A N/A N/A 0.1 N/A 1.3 N/A N/A N/A 3.7 N/A N/A N/A N/A N/A N/A N/A N/A
218 Figure 14. Total bilirubin levels of individual cats over time. Each cat has a unique marker, while groups are denoted accordingly by different colored lines (CF- Library=Blue, CF-1=Red, Unvaccinated, infected=green). DPI, days post infection. Notably, most vaccinated cats (7/8) had total bilirubin that peaked at or above 2 mg/dl. However, only ¼ cats vaccinated with CF-Library had total bilirubin peak above 2 mg/dl. Figure 15. Overall average total bilirubin peaks and nadirs of different test groups. Unvaccinated cats tended to have higher total bilirubin peak levels compared to vaccinated cats. 200
219 Table 8. Hematocrit values (%) for individual cats over the course of illness. Number of hematocrit measurements taken for each individual over the course of illness was dictated by severity of disease; dates where no data was collected are denoted ( N/A ). Peak and nadir hematocrit measurements were tabulated for each individual, and averages of each were calculated for each group. Note that measurements were available for unvaccinated, uninfected cats at two time points. DPI, days post infection. DPI AVG PEAK AVG NADIR GROUP CAT PEAK NADIR OF GROUP OF GROUP CF-Library N/A N/A 33 N/A N/A N/A 27 N/A N/A N/A 24 N/A N/A N/A N/A N/A N/A N/A 24 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 19 N/A CF N/A N/A 20 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 21 N/A N/A N/A N/A N/A Unvaccinated, N/A 26 N/A 29 N/A N/A 23 N/A N/A Infected N/A 26 N/A 23 N/A N/A N/A N/A 24 N/A N/A N/A N/A 22 N/A 25 N/A N/A N/A N/A N/A N/A 22 N/A N/A N/A N/A N/A N/A N/A 30 N/A 20 N/A N/A 22 N/A N/A 30 N/A 27 N/A N/A N/A 17 N/A N/A N/A N/A N/A N/A N/A Unvaccinated, N/A N/A N/A N/A N/A N/A N/A N/A N/A Uninfected N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A
220 Figure 16. Hematocrit of individual cats over time. Each cat has a unique marker, while groups are denoted accordingly by different colored lines (CF-Library=Blue, CF-1=Red, Unvaccinated, infected=green, Unvaccinated, uninfected=black). DPI, days post infection. Hematocrit decreased over time for all infected cats. Figure 17. Overall average hematocrit peaks and nadirs of different test groups. There is little difference between the hematocrit nadirs of different groups of infected cats. 202
221 Table 9. Red blood cell count (RBC, millions/µl) for individual cats over the course of illness. Number of RBC measurements taken for each individual over the course of illness was dictated by severity of disease; dates where no data was collected are denoted ( N/A ). Peak and nadir RBC measurements were tabulated for each individual, and averages of each were calculated for each group. DPI, days post infection. DPI AVG PEAK AVG NADIR GROUP CAT PEAK NADIR OF GROUP OF GROUP CF-Library N/A N/A 9.08 N/A N/A N/A 7.04 N/A N/A N/A 5.77 N/A N/A N/A N/A N/A N/A N/A 7.14 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 4.15 N/A CF N/A N/A 5.11 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 5.15 N/A N/A N/A N/A N/A Unvaccinated, N/A 6.22 N/A 6.8 N/A N/A 5.53 N/A N/A Infected N/A 6.16 N/A 5.29 N/A N/A N/A N/A N/A 5.75 N/A N/A N/A N/A 7.41 N/A 5.82 N/A 6.44 N/A N/A N/A N/A N/A N/A N/A 4.9 N/A N/A N/A N/A N/A N/A N/A 7.5 N/A 4.85 N/A N/A 5.2 N/A N/A N/A 6.74 N/A 6.02 N/A N/A N/A 3.82 N/A N/A N/A N/A N/A N/A N/A N/A
222 Figure 18. RBC counts of individual cats over time. Each cat has a unique marker, while groups are denoted accordingly by different colored lines (CF- Library=Blue, CF-1=Red, Unvaccinated, infected=green). DPI, days post infection. RBCs decreased over time for all infected cats. Figure 19. Overall average RBC peaks and nadirs of different test groups. There is little difference between RBC nadirs of different groups of infected cats. 204
223 Table 10. Hemoglobin values (g/dl) for individual cats over the course of illness. Number of hemoglobin measurements taken for each individual over the course of illness was dictated by severity of disease; dates where no data was collected are denoted ( N/A ). Peak and nadir total hemoglobin measurements were tabulated for each individual, and averages of each were calculated for each group. DPI, days post infection. DPI AVG PEAK AVG NADIR GROUP CAT PEAK NADIR OF GROUP OF GROUP CF-Library N/A N/A 11 N/A N/A N/A 8.5 N/A N/A N/A 8.1 N/A N/A N/A N/A N/A N/A N/A 8.4 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 6.1 N/A CF N/A N/A 7 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 6.9 N/A N/A N/A N/A N/A Unvaccinated, N/A 8.7 N/A 9.7 N/A N/A 7.9 N/A N/A Infected N/A 8.9 N/A 7.7 N/A N/A N/A N/A N/A 8.2 N/A N/A N/A N/A 10 N/A 7.7 N/A 8.6 N/A N/A N/A N/A N/A N/A N/A 7.1 N/A N/A N/A N/A N/A N/A N/A 10 N/A 6.4 N/A N/A 7 N/A N/A N/A 9.9 N/A 9 N/A N/A N/A 5.6 N/A N/A N/A N/A N/A N/A N/A N/A
224 Figure 20. Total hemoglobin levels of individual cats over time. Each cat has a unique marker, while groups are denoted accordingly by different colored lines (CF- Library=Blue, CF-1=Red, Unvaccinated, infected=green). Hemoglobin decreased over time for all infected cats. Figure 21. Overall average hemoglobin peaks and nadirs of different test groups. 206
225 Table 11. Mean cell volume (MCV, fl) for individual cats over the course of illness. Number of MCV measurements taken for each individual over the course of illness was dictated by severity of disease; dates where no data was collected are denoted ( N/A ). Peak and nadir MCV measurements were tabulated for each individual, and averages of each were calculated for each group. DPI, days post infection. DPI AVG PEAK AVG NADIR GROUP CAT PEAK NADIR OF GROUP OF GROUP CF-Library N/A N/A 36 N/A N/A N/A 38 N/A N/A N/A 42 N/A N/A N/A N/A N/A N/A N/A 33 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 47 N/A CF N/A N/A 40 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 41 N/A N/A N/A N/A N/A Unvaccinated, N/A 42 N/A 43 N/A N/A 42 N/A N/A Infected N/A 41 N/A 44 N/A N/A N/A N/A N/A 42 N/A N/A N/A N/A 39 N/A 38 N/A 39 N/A N/A N/A N/A N/A N/A N/A 45 N/A N/A N/A N/A N/A N/A N/A 40 N/A 41 N/A N/A 41 N/A N/A N/A 44 N/A 45 N/A N/A N/A 45 N/A N/A N/A N/A N/A N/A N/A N/A
226 Figure 22. MCV of individual cats over time. Each cat has a unique marker, while groups are denoted accordingly by different colored lines (CF- Library=Blue, CF-1=Red, Unvaccinated, infected=green). DPI, days post infection. Figure 23. Overall average MCV peaks and nadirs of different test groups. 208
227 Table 12. Mean cell hemoglobin (MCH, pg) for individual cats over the course of illness. Number of MCH measurements taken for each individual over the course of illness was dictated by severity of disease; dates where no data was collected are denoted ( N/A ). Peak and nadir MCH measurements were tabulated for each individual, and averages of each were calculated for each group. DPI, days post infection. DPI AVG PEAK AVG NADIR GROUP CAT PEAK NADIR OF GROUP OF GROUP CF-Library N/A N/A 12.1 N/A N/A N/A 12.1 N/A N/A N/A 14 N/A N/A N/A N/A N/A N/A N/A 11.8 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 14.7 N/A CF N/A N/A 13.7 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 13.4 N/A N/A N/A N/A N/A Unvaccinated, N/A 14 N/A 14.3 N/A N/A 14.3 N/A N/A Infected N/A 14.4 N/A 14.6 N/A N/A N/A N/A N/A 14.3 N/A N/A N/A N/A 13.5 N/A 13.2 N/A 13.4 N/A N/A N/A N/A N/A N/A N/A 14.5 N/A N/A N/A N/A N/A N/A N/A 13.3 N/A 13.2 N/A N/A 13.5 N/A N/A N/A 14.7 N/A 15 N/A N/A N/A 14.7 N/A N/A N/A N/A N/A N/A N/A N/A
228 Figure 24. MCH of individual cats over time. Each cat has a unique marker, while groups are denoted accordingly by different colored lines (CF- Library=Blue, CF-1=Red, Unvaccinated, infected=green). DPI, days post infection. Figure 25. Overall average MCH peaks and nadirs of different test groups. 210
229 Table 13. Mean corpuscular hemoglobin concentration (MCHC, g/dl) for individual cats over the course of illness. Number of MCHC measurements taken for each individual over the course of illness was dictated by severity of disease; dates where no data was collected are denoted ( N/A ). Peak and nadir MCHC measurements were tabulated for each individual, and averages of each were calculated for each group. DPI, days post infection. DPI AVG PEAK AVG NADIR GROUP CAT PEAK NADIR OF GROUP OF GROUP CF-Library N/A N/A 33.8 N/A N/A N/A 32.1 N/A N/A N/A 33.3 N/A N/A N/A N/A N/A N/A N/A 35.6 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 32.8 N/A CF N/A N/A 34.7 N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A N/A 33 N/A N/A N/A N/A N/A Unvaccinated, N/A 33.7 N/A 33.3 N/A N/A 33.8 N/A N/A Infected N/A 34.9 N/A 33.3 N/A N/A N/A N/A N/A 33.6 N/A N/A N/A N/A 34.8 N/A 35 N/A 34.3 N/A N/A N/A N/A N/A N/A N/A 32.6 N/A N/A N/A N/A N/A N/A N/A 33.4 N/A 32 N/A N/A 32.6 N/A N/A N/A 33.1 N/A 33.3 N/A N/A N/A 32.4 N/A N/A N/A N/A N/A N/A N/A N/A
.")
230 Figure 26. MCHC levels of individual cats over time. Each cat has a unique marker, while groups are denoted accordingly by different colored lines (CF- Library=Blue, CF-1=Red, Unvaccinated, infected=green). DPI, days post infection. Figure 27. Overall average MCHC peaks and nadirs of different test groups. 212
Outline 4/25/2009. Cytauxzoonosis: A tick-transmitted parasite of domestic and wild cats in the southeastern U.S. What is Cytauxzoonosis?
 Cytauxzoonosis: A tick-transmitted parasite of domestic and wild cats in the southeastern U.S. Michelle Rosen Center for Wildlife Health Department of Forestry, Wildlife, & Fisheries What is Cytauxzoonosis?
Cytauxzoonosis: A tick-transmitted parasite of domestic and wild cats in the southeastern U.S. Michelle Rosen Center for Wildlife Health Department of Forestry, Wildlife, & Fisheries What is Cytauxzoonosis?
Lecture 11 Wednesday, September 19, 2012
 Lecture 11 Wednesday, September 19, 2012 Phylogenetic tree (phylogeny) Darwin and classification: In the Origin, Darwin said that descent from a common ancestral species could explain why the Linnaean
Lecture 11 Wednesday, September 19, 2012 Phylogenetic tree (phylogeny) Darwin and classification: In the Origin, Darwin said that descent from a common ancestral species could explain why the Linnaean
MURDOCH RESEARCH REPOSITORY
 MURDOCH RESEARCH REPOSITORY http://researchrepository.murdoch.edu.au/20636/ Irwin, P.J. (2007) Blood, bull terriers and babesiosis: a review of canine babesiosis. In: 32nd Annual World Small Animal Veterinary
MURDOCH RESEARCH REPOSITORY http://researchrepository.murdoch.edu.au/20636/ Irwin, P.J. (2007) Blood, bull terriers and babesiosis: a review of canine babesiosis. In: 32nd Annual World Small Animal Veterinary
Blood protozoan: Plasmodium
 Blood protozoan: Plasmodium Dr. Hala Al Daghistani The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans: four species are associated The Plasmodium spp.
Blood protozoan: Plasmodium Dr. Hala Al Daghistani The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans: four species are associated The Plasmodium spp.
Blood protozoan: Plasmodium
 Blood protozoan: Plasmodium The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans:four species are associated The Plasmodium spp. life cycle can be divided
Blood protozoan: Plasmodium The causative agent of including Plasmodium vivax P. falciparum P. malariae P. ovale. malaria in humans:four species are associated The Plasmodium spp. life cycle can be divided
FACULTY OF VETERINARY MEDICINE
 FACULTY OF VETERINARY MEDICINE DEPARTMENT OF VETERINARY PARASITOLOGY AND ENTOMOLOGY M.Sc. AND Ph.D. DEGREE PROGRAMMES The postgraduate programmes of the Department of Veterinary Parasitology and Entomology
FACULTY OF VETERINARY MEDICINE DEPARTMENT OF VETERINARY PARASITOLOGY AND ENTOMOLOGY M.Sc. AND Ph.D. DEGREE PROGRAMMES The postgraduate programmes of the Department of Veterinary Parasitology and Entomology
Canine Anaplasmosis Anaplasma phagocytophilum Anaplasma platys
 Canine Anaplasmosis Anaplasma phagocytophilum Anaplasma platys It takes just hours for an infected tick to transmit Anaplasma organisms to a dog. What is canine anaplasmosis? Canine anaplasmosis is a disease
Canine Anaplasmosis Anaplasma phagocytophilum Anaplasma platys It takes just hours for an infected tick to transmit Anaplasma organisms to a dog. What is canine anaplasmosis? Canine anaplasmosis is a disease
Consequences of Antimicrobial Resistant Bacteria. Antimicrobial Resistance. Molecular Genetics of Antimicrobial Resistance. Topics to be Covered
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
MID 23. Antimicrobial Resistance. Consequences of Antimicrobial Resistant Bacteria. Molecular Genetics of Antimicrobial Resistance
 Antimicrobial Resistance Molecular Genetics of Antimicrobial Resistance Micro evolutionary change - point mutations Beta-lactamase mutation extends spectrum of the enzyme rpob gene (RNA polymerase) mutation
Antimicrobial Resistance Molecular Genetics of Antimicrobial Resistance Micro evolutionary change - point mutations Beta-lactamase mutation extends spectrum of the enzyme rpob gene (RNA polymerase) mutation
Antimicrobial Resistance
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Acquisition of Foreign DNA
 Antimicrobial Resistance Acquisition of Foreign DNA Levy, Scientific American Horizontal gene transfer is common, even between Gram positive and negative bacteria Plasmid - transfer of single or multiple
Antimicrobial Resistance Acquisition of Foreign DNA Levy, Scientific American Horizontal gene transfer is common, even between Gram positive and negative bacteria Plasmid - transfer of single or multiple
TOPIC CLADISTICS
 TOPIC 5.4 - CLADISTICS 5.4 A Clades & Cladograms https://upload.wikimedia.org/wikipedia/commons/thumb/4/46/clade-grade_ii.svg IB BIO 5.4 3 U1: A clade is a group of organisms that have evolved from a common
TOPIC 5.4 - CLADISTICS 5.4 A Clades & Cladograms https://upload.wikimedia.org/wikipedia/commons/thumb/4/46/clade-grade_ii.svg IB BIO 5.4 3 U1: A clade is a group of organisms that have evolved from a common
The Friends of Nachusa Grasslands 2016 Scientific Research Project Grant Report Due June 30, 2017
 The Friends of Nachusa Grasslands 2016 Scientific Research Project Grant Report Due June 30, 2017 Name: Laura Adamovicz Address: 2001 S Lincoln Ave, Urbana, IL 61802 Phone: 217-333-8056 2016 grant amount:
The Friends of Nachusa Grasslands 2016 Scientific Research Project Grant Report Due June 30, 2017 Name: Laura Adamovicz Address: 2001 S Lincoln Ave, Urbana, IL 61802 Phone: 217-333-8056 2016 grant amount:
In the first half of the 20th century, Dr. Guido Fanconi published detailed clinical descriptions of several heritable human diseases.
 In the first half of the 20th century, Dr. Guido Fanconi published detailed clinical descriptions of several heritable human diseases. Two disease syndromes were named after him: Fanconi Anemia and Fanconi
In the first half of the 20th century, Dr. Guido Fanconi published detailed clinical descriptions of several heritable human diseases. Two disease syndromes were named after him: Fanconi Anemia and Fanconi
CLADISTICS Student Packet SUMMARY Phylogeny Phylogenetic trees/cladograms
 CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
both are fatal diseases. In babesiosis blood comes out with the urine and hence it is also known as Red water disease. Theileria vaccines are not
 1.1 INTRODUCTION Animal husbandry plays an important role in Indian agriculture. Indians by large are vegetarian and as such the only source of animal protein is milk and milk products. With the increasing
1.1 INTRODUCTION Animal husbandry plays an important role in Indian agriculture. Indians by large are vegetarian and as such the only source of animal protein is milk and milk products. With the increasing
EFSA Scientific Opinion on canine leishmaniosis
 EFSA Scientific Opinion on canine leishmaniosis Andrea Gervelmeyer Animal Health and Welfare Team Animal and Plant Health Unit AHAC meeting 19 June 2015 PRESENTATION OUTLINE Outline Background ToR Approach
EFSA Scientific Opinion on canine leishmaniosis Andrea Gervelmeyer Animal Health and Welfare Team Animal and Plant Health Unit AHAC meeting 19 June 2015 PRESENTATION OUTLINE Outline Background ToR Approach
Ehrlichiosis, Anaplasmosis and other Vector Borne Diseases You May Not Be Thinking About Richard E Goldstein Cornell University Ithaca NY
 Ehrlichiosis, Anaplasmosis and other Vector Borne Diseases You May Not Be Thinking About Richard E Goldstein Cornell University Ithaca NY Canine Monocytic Ehrlichiosis Ehrlichia canis The common etiologic
Ehrlichiosis, Anaplasmosis and other Vector Borne Diseases You May Not Be Thinking About Richard E Goldstein Cornell University Ithaca NY Canine Monocytic Ehrlichiosis Ehrlichia canis The common etiologic
Interpreting Evolutionary Trees Honors Integrated Science 4 Name Per.
 Interpreting Evolutionary Trees Honors Integrated Science 4 Name Per. Introduction Imagine a single diagram representing the evolutionary relationships between everything that has ever lived. If life evolved
Interpreting Evolutionary Trees Honors Integrated Science 4 Name Per. Introduction Imagine a single diagram representing the evolutionary relationships between everything that has ever lived. If life evolved
An Overview of Canine Babesiosis
 Page 1 of 6 C. Wyatt Cleveland, DVM; David S. Peterson, DVM, PhD; and Kenneth S. Latimer, DVM, PhD Class of 2002 (Cleveland), Department of Medical Microbiology and Parasitology (Peterson), and Department
Page 1 of 6 C. Wyatt Cleveland, DVM; David S. Peterson, DVM, PhD; and Kenneth S. Latimer, DVM, PhD Class of 2002 (Cleveland), Department of Medical Microbiology and Parasitology (Peterson), and Department
Drd. OBADĂ MIHAI DORU. PhD THESIS ABSTRACT
 UNIVERSITY OF AGRICULTURAL SCIENCES AND VETERINARY MEDICINE ION IONESCU DE LA BRAD IAŞI FACULTY OF VETERINARY MEDICINE SPECIALIZATION MICROBIOLOGY- IMUNOLOGY Drd. OBADĂ MIHAI DORU PhD THESIS ABSTRACT RESEARCHES
UNIVERSITY OF AGRICULTURAL SCIENCES AND VETERINARY MEDICINE ION IONESCU DE LA BRAD IAŞI FACULTY OF VETERINARY MEDICINE SPECIALIZATION MICROBIOLOGY- IMUNOLOGY Drd. OBADĂ MIHAI DORU PhD THESIS ABSTRACT RESEARCHES
Why Don t These Drugs Work Anymore? Biosciences in the 21 st Century Dr. Amber Rice October 28, 2013
 Why Don t These Drugs Work Anymore? Biosciences in the 21 st Century Dr. Amber Rice October 28, 2013 Outline Drug resistance: a case study Evolution: the basics How does resistance evolve? Examples of
Why Don t These Drugs Work Anymore? Biosciences in the 21 st Century Dr. Amber Rice October 28, 2013 Outline Drug resistance: a case study Evolution: the basics How does resistance evolve? Examples of
The detection of Cytauxzoon felis in apparently healthy free-roaming cats in the USA
 Veterinary Parasitology 146 (2007) 316 320 www.elsevier.com/locate/vetpar The detection of Cytauxzoon felis in apparently healthy free-roaming cats in the USA Marion D. Haber a, Melissa D. Tucker a, Henry
Veterinary Parasitology 146 (2007) 316 320 www.elsevier.com/locate/vetpar The detection of Cytauxzoon felis in apparently healthy free-roaming cats in the USA Marion D. Haber a, Melissa D. Tucker a, Henry
AKC Canine Health Foundation Grant Updates: Research Currently Being Sponsored By The Vizsla Club of America Welfare Foundation
 AKC Canine Health Foundation Grant Updates: Research Currently Being Sponsored By The Vizsla Club of America Welfare Foundation GRANT PROGRESS REPORT REVIEW Grant: 00748: SNP Association Mapping for Canine
AKC Canine Health Foundation Grant Updates: Research Currently Being Sponsored By The Vizsla Club of America Welfare Foundation GRANT PROGRESS REPORT REVIEW Grant: 00748: SNP Association Mapping for Canine
BioSci 110, Fall 08 Exam 2
 1. is the cell division process that results in the production of a. mitosis; 2 gametes b. meiosis; 2 gametes c. meiosis; 2 somatic (body) cells d. mitosis; 4 somatic (body) cells e. *meiosis; 4 gametes
1. is the cell division process that results in the production of a. mitosis; 2 gametes b. meiosis; 2 gametes c. meiosis; 2 somatic (body) cells d. mitosis; 4 somatic (body) cells e. *meiosis; 4 gametes
InternationalJournalofAgricultural
 www.ijasvm.com IJASVM InternationalJournalofAgricultural SciencesandVeterinaryMedicine ISSN:2320-3730 Vol.5,No.1,February2017 E-Mail:editorijasvm@gmail.com oreditor@ijasvm.comm@gmail.com Int. J. Agric.Sc
www.ijasvm.com IJASVM InternationalJournalofAgricultural SciencesandVeterinaryMedicine ISSN:2320-3730 Vol.5,No.1,February2017 E-Mail:editorijasvm@gmail.com oreditor@ijasvm.comm@gmail.com Int. J. Agric.Sc
Cystic echinococcosis in a domestic cat: an Italian case report
 13th NRL Workshop, Rome, 24-25 May, 2018 Cystic echinococcosis in a domestic cat: an Italian case report Istituto Zooprofilattico Sperimentale (IZS) of Sardinia National Reference Laboratory for Cistic
13th NRL Workshop, Rome, 24-25 May, 2018 Cystic echinococcosis in a domestic cat: an Italian case report Istituto Zooprofilattico Sperimentale (IZS) of Sardinia National Reference Laboratory for Cistic
Vaccines for Cats. 2. Feline viral rhinotracheitis, FVR caused by FVR virus, also known as herpes virus type 1, FHV-1
 Vaccines for Cats Recent advances in veterinary medical science have resulted in an increase in the number and type of vaccines that are available for use in cats, and improvements are continuously being
Vaccines for Cats Recent advances in veterinary medical science have resulted in an increase in the number and type of vaccines that are available for use in cats, and improvements are continuously being
Ehrlichia and Anaplasma: What Do We Need to Know in NY State Richard E Goldstein DVM DACVIM DECVIM-CA The Animal Medical Center New York, NY
 Ehrlichia and Anaplasma: What Do We Need to Know in NY State Richard E Goldstein DVM DACVIM DECVIM-CA The Animal Medical Center New York, NY Learning Objectives The attendees will be familiar with the
Ehrlichia and Anaplasma: What Do We Need to Know in NY State Richard E Goldstein DVM DACVIM DECVIM-CA The Animal Medical Center New York, NY Learning Objectives The attendees will be familiar with the
Phenotype Observed Expected (O-E) 2 (O-E) 2 /E dotted yellow solid yellow dotted blue solid blue
 2 (O-E) 2 /E dotted yellow solid yellow dotted blue solid blue") 1. (30 pts) A tropical fish breeder for the local pet store is interested in creating a new type of fancy tropical fish. She observes consistent patterns of inheritance for the following traits: P 1 :
1. (30 pts) A tropical fish breeder for the local pet store is interested in creating a new type of fancy tropical fish. She observes consistent patterns of inheritance for the following traits: P 1 :
PLASMODIUM MODULE 39.1 INTRODUCTION OBJECTIVES 39.2 MALARIAL PARASITE. Notes
 Plasmodium MODULE 39 PLASMODIUM 39.1 INTRODUCTION Malaria is characterized by intermittent fever associated with chills and rigors in the patient. There may be enlargement of the liver and spleen in the
Plasmodium MODULE 39 PLASMODIUM 39.1 INTRODUCTION Malaria is characterized by intermittent fever associated with chills and rigors in the patient. There may be enlargement of the liver and spleen in the
Modern Evolutionary Classification. Lesson Overview. Lesson Overview Modern Evolutionary Classification
 Lesson Overview 18.2 Modern Evolutionary Classification THINK ABOUT IT Darwin s ideas about a tree of life suggested a new way to classify organisms not just based on similarities and differences, but
Lesson Overview 18.2 Modern Evolutionary Classification THINK ABOUT IT Darwin s ideas about a tree of life suggested a new way to classify organisms not just based on similarities and differences, but
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and
 Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
Antimicrobial Resistance
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of Change in the approach to the administration of empiric antimicrobial therapy Increased
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of Change in the approach to the administration of empiric antimicrobial therapy Increased
The Essentials of Ticks and Tick-borne Diseases
 The Essentials of Ticks and Tick-borne Diseases Presenter: Bobbi S. Pritt, M.D., M.Sc. Director, Clinical Parasitology Laboratory Co-Director, Vector-borne Diseases Laboratory Services Vice Chair of Education
The Essentials of Ticks and Tick-borne Diseases Presenter: Bobbi S. Pritt, M.D., M.Sc. Director, Clinical Parasitology Laboratory Co-Director, Vector-borne Diseases Laboratory Services Vice Chair of Education
ABSTRACT. Cytauxzoonosis is an emerging tick transmitted disease of domestic cats (Felis
 ABSTRACT TARIGO, JAIME. The Cytauxzoon felis Genome: A Guide To Vaccine Candidate Antigen Discovery For Cytauxzoonosis. (Under the direction of Dr. Adam Birkenheuer, Chair, and Dr. Gregg Dean, Vice Chair).
ABSTRACT TARIGO, JAIME. The Cytauxzoon felis Genome: A Guide To Vaccine Candidate Antigen Discovery For Cytauxzoonosis. (Under the direction of Dr. Adam Birkenheuer, Chair, and Dr. Gregg Dean, Vice Chair).
Species: Panthera pardus Genus: Panthera Family: Felidae Order: Carnivora Class: Mammalia Phylum: Chordata
 CHAPTER 6: PHYLOGENY AND THE TREE OF LIFE AP Biology 3 PHYLOGENY AND SYSTEMATICS Phylogeny - evolutionary history of a species or group of related species Systematics - analytical approach to understanding
CHAPTER 6: PHYLOGENY AND THE TREE OF LIFE AP Biology 3 PHYLOGENY AND SYSTEMATICS Phylogeny - evolutionary history of a species or group of related species Systematics - analytical approach to understanding
Ch 1.2 Determining How Species Are Related.notebook February 06, 2018
 Name 3 "Big Ideas" from our last notebook lecture: * * * 1 WDYR? Of the following organisms, which is the closest relative of the "Snowy Owl" (Bubo scandiacus)? a) barn owl (Tyto alba) b) saw whet owl
Name 3 "Big Ideas" from our last notebook lecture: * * * 1 WDYR? Of the following organisms, which is the closest relative of the "Snowy Owl" (Bubo scandiacus)? a) barn owl (Tyto alba) b) saw whet owl
UNIT III A. Descent with Modification(Ch19) B. Phylogeny (Ch20) C. Evolution of Populations (Ch21) D. Origin of Species or Speciation (Ch22)
 B. Phylogeny (Ch20) C. Evolution of Populations (Ch21) D. Origin of Species or Speciation (Ch22)") UNIT III A. Descent with Modification(Ch9) B. Phylogeny (Ch2) C. Evolution of Populations (Ch2) D. Origin of Species or Speciation (Ch22) Classification in broad term simply means putting things in classes
UNIT III A. Descent with Modification(Ch9) B. Phylogeny (Ch2) C. Evolution of Populations (Ch2) D. Origin of Species or Speciation (Ch22) Classification in broad term simply means putting things in classes
Visit ABLE on the Web at:
 This article reprinted from: Lessem, P. B. 2008. The antibiotic resistance phenomenon: Use of minimal inhibitory concentration (MIC) determination for inquiry based experimentation. Pages 357-362, in Tested
This article reprinted from: Lessem, P. B. 2008. The antibiotic resistance phenomenon: Use of minimal inhibitory concentration (MIC) determination for inquiry based experimentation. Pages 357-362, in Tested
Phylogeographic assessment of Acanthodactylus boskianus (Reptilia: Lacertidae) based on phylogenetic analysis of mitochondrial DNA.
 based on phylogenetic analysis of mitochondrial DNA.") Zoology Department Phylogeographic assessment of Acanthodactylus boskianus (Reptilia: Lacertidae) based on phylogenetic analysis of mitochondrial DNA By HAGAR IBRAHIM HOSNI BAYOUMI A thesis submitted in
Zoology Department Phylogeographic assessment of Acanthodactylus boskianus (Reptilia: Lacertidae) based on phylogenetic analysis of mitochondrial DNA By HAGAR IBRAHIM HOSNI BAYOUMI A thesis submitted in
American Association of Zoo Veterinarians Infectious Disease Committee Manual 2013 CYTAUXZOONOSIS
 Animal Group(s) Affected Felids, wild and domestic Transmission Tick-borne (Amblyomma americanum and Dermacentor variabilis) Clinical Signs Domestic cats and some exotic felids: some cats develop no clinical
Animal Group(s) Affected Felids, wild and domestic Transmission Tick-borne (Amblyomma americanum and Dermacentor variabilis) Clinical Signs Domestic cats and some exotic felids: some cats develop no clinical
Development and improvement of diagnostics to improve use of antibiotics and alternatives to antibiotics
 Priority Topic B Diagnostics Development and improvement of diagnostics to improve use of antibiotics and alternatives to antibiotics The overarching goal of this priority topic is to stimulate the design,
Priority Topic B Diagnostics Development and improvement of diagnostics to improve use of antibiotics and alternatives to antibiotics The overarching goal of this priority topic is to stimulate the design,
Course Offerings: Associate of Applied Science Veterinary Technology. Course Number Name Credits
 Course Offerings: Associate of Applied Science Veterinary Technology Course Number Name Credits Required Courses in Major: Fall Semester, First Year *VETT-101 Animal Health Careers 1-0-1 *VETT-102 Veterinary
Course Offerings: Associate of Applied Science Veterinary Technology Course Number Name Credits Required Courses in Major: Fall Semester, First Year *VETT-101 Animal Health Careers 1-0-1 *VETT-102 Veterinary
Overview. There are commonly found arrangements of bacteria based on their division. Spheres, Rods, Spirals
 Bacteria Overview Bacteria live almost everywhere. Most are microscopic ranging from 0.5 5 m in size, and unicellular. They have a variety of shapes when viewed under a microscope, most commonly: Spheres,
Bacteria Overview Bacteria live almost everywhere. Most are microscopic ranging from 0.5 5 m in size, and unicellular. They have a variety of shapes when viewed under a microscope, most commonly: Spheres,
Tick-borne Disease Testing in Shelters What Does that Blue Dot Really Mean?
 Tick-borne Disease Testing in Shelters What Does that Blue Dot Really Mean? 2017 ASPCA. All Rights Reserved. Your Presenter Stephanie Janeczko, DVM, MS, DABVP, CAWA Senior Director of Shelter Medical Programs
Tick-borne Disease Testing in Shelters What Does that Blue Dot Really Mean? 2017 ASPCA. All Rights Reserved. Your Presenter Stephanie Janeczko, DVM, MS, DABVP, CAWA Senior Director of Shelter Medical Programs
Protozoan Parasites Transmitted by Arthropod vectors
 Protozoan Parasites Transmitted by Arthropod vectors Spencer Greenwood Dept.of Biomedical Sciences Office: 3313 N AVC North Annex 566-6002 892-4686 Sgreenwood@upei.ca Protozoa Transmitted by Arthropod
Protozoan Parasites Transmitted by Arthropod vectors Spencer Greenwood Dept.of Biomedical Sciences Office: 3313 N AVC North Annex 566-6002 892-4686 Sgreenwood@upei.ca Protozoa Transmitted by Arthropod
Evolution of Birds. Summary:
 Oregon State Standards OR Science 7.1, 7.2, 7.3, 7.3S.1, 7.3S.2 8.1, 8.2, 8.2L.1, 8.3, 8.3S.1, 8.3S.2 H.1, H.2, H.2L.4, H.2L.5, H.3, H.3S.1, H.3S.2, H.3S.3 Summary: Students create phylogenetic trees to
Oregon State Standards OR Science 7.1, 7.2, 7.3, 7.3S.1, 7.3S.2 8.1, 8.2, 8.2L.1, 8.3, 8.3S.1, 8.3S.2 H.1, H.2, H.2L.4, H.2L.5, H.3, H.3S.1, H.3S.2, H.3S.3 Summary: Students create phylogenetic trees to
Course Curriculum for Master Degree in Internal Medicine/ Faculty of Veterinary Medicine
 Course Curriculum for Master Degree in Internal Medicine/ Faculty of Veterinary Medicine The Master Degree in Internal Medicine/Faculty of Veterinary Medicine is awarded by the Faculty of Graduate Studies
Course Curriculum for Master Degree in Internal Medicine/ Faculty of Veterinary Medicine The Master Degree in Internal Medicine/Faculty of Veterinary Medicine is awarded by the Faculty of Graduate Studies
Antimicrobial Resistance Initiative
 Antimicrobial Resistance Initiative Antimicrobial Resistance Initiative Resistance to antimicrobial agents has become a threat to public health all over the world. Microorganisms become resistant to antimicrobial
Antimicrobial Resistance Initiative Antimicrobial Resistance Initiative Resistance to antimicrobial agents has become a threat to public health all over the world. Microorganisms become resistant to antimicrobial
WHY IS THIS IMPORTANT?
 CHAPTER 20 ANTIBIOTIC RESISTANCE WHY IS THIS IMPORTANT? The most important problem associated with infectious disease today is the rapid development of resistance to antibiotics It will force us to change
CHAPTER 20 ANTIBIOTIC RESISTANCE WHY IS THIS IMPORTANT? The most important problem associated with infectious disease today is the rapid development of resistance to antibiotics It will force us to change
Author - Dr. Josie Traub-Dargatz
 Author - Dr. Josie Traub-Dargatz Dr. Josie Traub-Dargatz is a professor of equine medicine at Colorado State University (CSU) College of Veterinary Medicine and Biomedical Sciences. She began her veterinary
Author - Dr. Josie Traub-Dargatz Dr. Josie Traub-Dargatz is a professor of equine medicine at Colorado State University (CSU) College of Veterinary Medicine and Biomedical Sciences. She began her veterinary
Changing Trends and Issues in Canine and Feline Heartworm Infections
 Changing Trends and Issues in Canine and Feline Heartworm Infections Byron L. Blagburn College of Veterinary Medicine Auburn University Canine and feline heartworm diagnostic, treatment and prevention
Changing Trends and Issues in Canine and Feline Heartworm Infections Byron L. Blagburn College of Veterinary Medicine Auburn University Canine and feline heartworm diagnostic, treatment and prevention
How to talk to clients about heartworm disease
 Client Communication How to talk to clients about heartworm disease Detecting heartworm infection early generally allows for a faster and more effective response to treatment. Answers to pet owners most
Client Communication How to talk to clients about heartworm disease Detecting heartworm infection early generally allows for a faster and more effective response to treatment. Answers to pet owners most
University of Canberra. This thesis is available in print format from the University of Canberra Library.
 University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
Egyptian Marital status. Single Lecturer of infectious Diseases in Department of Animal Occupation:
 Contact Present address: Telephone : E-mail : Department of Animal Medicine (Infectious diseases), Faculty of Veterinary Medicine, Assiut University, Assiut 71526, Egypt 002-01004477501 (Egypt) amiraelhosary@yahoo.com
Contact Present address: Telephone : E-mail : Department of Animal Medicine (Infectious diseases), Faculty of Veterinary Medicine, Assiut University, Assiut 71526, Egypt 002-01004477501 (Egypt) amiraelhosary@yahoo.com
Suggested vector-borne disease screening guidelines
 Suggested vector-borne disease screening guidelines SNAP Dx Test Screen your dog every year with the SNAP Dx Test to detect exposure to pathogens that cause heartworm disease, ehrlichiosis, Lyme disease
Suggested vector-borne disease screening guidelines SNAP Dx Test Screen your dog every year with the SNAP Dx Test to detect exposure to pathogens that cause heartworm disease, ehrlichiosis, Lyme disease
Burton's Microbiology for the Health Sciences. Chapter 9. Controlling Microbial Growth in Vivo Using Antimicrobial Agents
 Burton's Microbiology for the Health Sciences Chapter 9. Controlling Microbial Growth in Vivo Using Antimicrobial Agents Chapter 9 Outline Introduction Characteristics of an Ideal Antimicrobial Agent How
Burton's Microbiology for the Health Sciences Chapter 9. Controlling Microbial Growth in Vivo Using Antimicrobial Agents Chapter 9 Outline Introduction Characteristics of an Ideal Antimicrobial Agent How
Inheritance of Livershunt in Irish Wolfhounds By Maura Lyons PhD
 Inheritance of Livershunt in Irish Wolfhounds By Maura Lyons PhD Glossary Gene = A piece of DNA that provides the 'recipe' for an enzyme or a protein. Gene locus = The position of a gene on a chromosome.
Inheritance of Livershunt in Irish Wolfhounds By Maura Lyons PhD Glossary Gene = A piece of DNA that provides the 'recipe' for an enzyme or a protein. Gene locus = The position of a gene on a chromosome.
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and
 Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
National Academic Reference Standards (NARS) Veterinary Medicine. February st Edition
 Veterinary Medicine. February st Edition") National Academic Reference Standards (NARS) Veterinary Medicine February 2009 1 st Edition Table of Contents Introduction to Veterinary Medical Education 1 National Academic Reference Standards 3 Curriculum
National Academic Reference Standards (NARS) Veterinary Medicine February 2009 1 st Edition Table of Contents Introduction to Veterinary Medical Education 1 National Academic Reference Standards 3 Curriculum
New Insights into the Treatment of Leishmaniasis
 New Insights into the Treatment of Leishmaniasis Eric Zini Snow meeting, 14 March 2009 Few drugs available for dogs Initially developed to treat human leishmaniasis, later adopted in dogs None eradicates
New Insights into the Treatment of Leishmaniasis Eric Zini Snow meeting, 14 March 2009 Few drugs available for dogs Initially developed to treat human leishmaniasis, later adopted in dogs None eradicates
Hepatic Copper Storage Disorder in the Dalmatian. Copper Is Essential For Life 7/7/18. Hepatic Copper Transport. Normal Copper Metabolism
 7/7/18 Hepatic Storage Disorder in the Dalmatian Is Essential For Life David C. Twedt DVM, DACVIM Energy production Nerve transmission Antioxidant function Iron metabolism http://s3.amazonaws.com/cdn-origin-etr.akc.org/wp-content/u
7/7/18 Hepatic Storage Disorder in the Dalmatian Is Essential For Life David C. Twedt DVM, DACVIM Energy production Nerve transmission Antioxidant function Iron metabolism http://s3.amazonaws.com/cdn-origin-etr.akc.org/wp-content/u
Dominance/Suppression Competitive Relationships in Loblolly Pine (Pinus taeda L.) Plantations
 Plantations") Dominance/Suppression Competitive Relationships in Loblolly Pine (Pinus taeda L.) Plantations by Michael E. Dyer Dissertation submitted to the Faculty of the Virginia Polytechnic Institute and Stand University
Dominance/Suppression Competitive Relationships in Loblolly Pine (Pinus taeda L.) Plantations by Michael E. Dyer Dissertation submitted to the Faculty of the Virginia Polytechnic Institute and Stand University
THE ROYAL COLLEGE OF VETERINARY SURGEONS DIPLOMA EXAMINATION IN VETERINARY DERMATOLOGY. Tuesday 22 August PAPER 1 (3 hours)
") DIPLOMA EXAMINATION IN VETERINARY DERMATOLOGY Tuesday 22 August 2000 PAPER 1 Candidates are required to answer FOUR questions only. 1. What is meant by the term staphylococcal virulence factors. Indicate
DIPLOMA EXAMINATION IN VETERINARY DERMATOLOGY Tuesday 22 August 2000 PAPER 1 Candidates are required to answer FOUR questions only. 1. What is meant by the term staphylococcal virulence factors. Indicate
Inhibiting Microbial Growth in vivo. CLS 212: Medical Microbiology Zeina Alkudmani
 Inhibiting Microbial Growth in vivo CLS 212: Medical Microbiology Zeina Alkudmani Chemotherapy Definitions The use of any chemical (drug) to treat any disease or condition. Chemotherapeutic Agent Any drug
Inhibiting Microbial Growth in vivo CLS 212: Medical Microbiology Zeina Alkudmani Chemotherapy Definitions The use of any chemical (drug) to treat any disease or condition. Chemotherapeutic Agent Any drug
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and
 Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere
INFECTIOUS DISEASE Symposium Proceedings
 INFECTIOUS DISEASE 2015 Symposium Proceedings The opinions expressed in the articles in this publication are those of the authors and do not necessarily reflect the official label recommendations and points
INFECTIOUS DISEASE 2015 Symposium Proceedings The opinions expressed in the articles in this publication are those of the authors and do not necessarily reflect the official label recommendations and points
Mechanisms and Pathways of AMR in the environment
 FMM/RAS/298: Strengthening capacities, policies and national action plans on prudent and responsible use of antimicrobials in fisheries Final Workshop in cooperation with AVA Singapore and INFOFISH 12-14
FMM/RAS/298: Strengthening capacities, policies and national action plans on prudent and responsible use of antimicrobials in fisheries Final Workshop in cooperation with AVA Singapore and INFOFISH 12-14
INTERNATIONAL CANINE HEALTH AWARDS Awards Presentation 24th May 2017
 INTERNATIONAL CANINE HEALTH AWARDS Awards Presentation 24th May 2017 Supported by the Vernon & Shirley Hill Foundation Administrated by the Kennel Club Charitable Trust PAGE 2 We are proud to support these
INTERNATIONAL CANINE HEALTH AWARDS Awards Presentation 24th May 2017 Supported by the Vernon & Shirley Hill Foundation Administrated by the Kennel Club Charitable Trust PAGE 2 We are proud to support these
Name: Date: Hour: Fill out the following character matrix. Mark an X if an organism has the trait.
 Name: Date: Hour: CLADOGRAM ANALYSIS What is a cladogram? It is a diagram that depicts evolutionary relationships among groups. It is based on PHYLOGENY, which is the study of evolutionary relationships.
Name: Date: Hour: CLADOGRAM ANALYSIS What is a cladogram? It is a diagram that depicts evolutionary relationships among groups. It is based on PHYLOGENY, which is the study of evolutionary relationships.
Warm Up What recommendations do you have for him? Choose a partner and list some suggestions in your lab notebook.
 Antibiotics 1. Warmup: Medical Scenario 2. Lecture: PPT Slides & Notes 3. Math Connection: Graphing Activity 4. Assessment: Final Recommendation for Medical Scenario Citing Evidence 5. Enrichment: Article
Antibiotics 1. Warmup: Medical Scenario 2. Lecture: PPT Slides & Notes 3. Math Connection: Graphing Activity 4. Assessment: Final Recommendation for Medical Scenario Citing Evidence 5. Enrichment: Article
Pain management: making the most of the latest options
 Vet Times The website for the veterinary profession https://www.vettimes.co.uk Pain management: making the most of the latest options Author : James Westgate Categories : Business, Business planning Date
Vet Times The website for the veterinary profession https://www.vettimes.co.uk Pain management: making the most of the latest options Author : James Westgate Categories : Business, Business planning Date
COMPARING DNA SEQUENCES TO UNDERSTAND EVOLUTIONARY RELATIONSHIPS WITH BLAST
 Big Idea 1 Evolution INVESTIGATION 3 COMPARING DNA SEQUENCES TO UNDERSTAND EVOLUTIONARY RELATIONSHIPS WITH BLAST How can bioinformatics be used as a tool to determine evolutionary relationships and to
Big Idea 1 Evolution INVESTIGATION 3 COMPARING DNA SEQUENCES TO UNDERSTAND EVOLUTIONARY RELATIONSHIPS WITH BLAST How can bioinformatics be used as a tool to determine evolutionary relationships and to
Pathogenesis of E. canis
 Tick-born disease Rhipicephalus sanguineus brown dog tick Rickettsia Ehrlichia canis Ehrlichia platys Anaplasma platys Pathogenesis of E. canis Incubation period: 8 20 days Mononuclear cells Liver, spleen,
Tick-born disease Rhipicephalus sanguineus brown dog tick Rickettsia Ehrlichia canis Ehrlichia platys Anaplasma platys Pathogenesis of E. canis Incubation period: 8 20 days Mononuclear cells Liver, spleen,
Bi156 Lecture 1/13/12. Dog Genetics
 Bi156 Lecture 1/13/12 Dog Genetics The radiation of the family Canidae occurred about 100 million years ago. Dogs are most closely related to wolves, from which they diverged through domestication about
Bi156 Lecture 1/13/12 Dog Genetics The radiation of the family Canidae occurred about 100 million years ago. Dogs are most closely related to wolves, from which they diverged through domestication about
Tick-Borne Disease Research Program
 Tick-Borne Disease Research Program Strategic Plan INTRODUCTION The Congressionally Directed Medical Research Programs (CDMRP) represents a unique partnership among the U.S. Congress, the military, and
Tick-Borne Disease Research Program Strategic Plan INTRODUCTION The Congressionally Directed Medical Research Programs (CDMRP) represents a unique partnership among the U.S. Congress, the military, and
VETERINARY BIOMEDICAL SCIENCES (VBSC)
") Veterinary Biomedical Sciences (VBSC) 1 VETERINARY BIOMEDICAL SCIENCES (VBSC) VBSC 5000 Master s Research and Thesis Prerequisites: Graduate standing. Description: Research problem for meeting requirements
Veterinary Biomedical Sciences (VBSC) 1 VETERINARY BIOMEDICAL SCIENCES (VBSC) VBSC 5000 Master s Research and Thesis Prerequisites: Graduate standing. Description: Research problem for meeting requirements
Phylum:Apicomplexa Class:Sporozoa
 Phylum:Apicomplexa Class:Sporozoa The most characteristic features of sporozoa are 1-unique appearance of most protozoa makes it possible for knowledge able person to identifiy them to level of genus and
Phylum:Apicomplexa Class:Sporozoa The most characteristic features of sporozoa are 1-unique appearance of most protozoa makes it possible for knowledge able person to identifiy them to level of genus and
 Tutorial 9 notes Super Bug: Antibiotics & Evolution Kristy J. Wilson Department of Pathology Emory University History of Antibiotics http://videos.howstuffworks.com/science-channel/29783-100-greatest-discoveries-penicillinvideo.htm
Tutorial 9 notes Super Bug: Antibiotics & Evolution Kristy J. Wilson Department of Pathology Emory University History of Antibiotics http://videos.howstuffworks.com/science-channel/29783-100-greatest-discoveries-penicillinvideo.htm
READER S DIGEST OVERVIEW: BIGHORN SHEEP. Peregrine Wolff, DVM
 READER S DIGEST OVERVIEW: RESPIRATORY DISEASE IN BIGHORN SHEEP Peregrine Wolff, DVM Nevada Department of Wildlife During the Lewis & Clark expedition (1804 1806) There may have been 2 million bighorn sheep
READER S DIGEST OVERVIEW: RESPIRATORY DISEASE IN BIGHORN SHEEP Peregrine Wolff, DVM Nevada Department of Wildlife During the Lewis & Clark expedition (1804 1806) There may have been 2 million bighorn sheep
17.2 Classification Based on Evolutionary Relationships Organization of all that speciation!
 Organization of all that speciation! Patterns of evolution.. Taxonomy gets an over haul! Using more than morphology! 3 domains, 6 kingdoms KEY CONCEPT Modern classification is based on evolutionary relationships.
Organization of all that speciation! Patterns of evolution.. Taxonomy gets an over haul! Using more than morphology! 3 domains, 6 kingdoms KEY CONCEPT Modern classification is based on evolutionary relationships.
TE 408: Three-day Lesson Plan
 TE 408: Three-day Lesson Plan Partner: Anthony Machniak School: Okemos High School Date: 3/17/2014 Name: Theodore Baker Mentor Teacher: Danielle Tandoc Class and grade level: 9-10th grade Biology Part
TE 408: Three-day Lesson Plan Partner: Anthony Machniak School: Okemos High School Date: 3/17/2014 Name: Theodore Baker Mentor Teacher: Danielle Tandoc Class and grade level: 9-10th grade Biology Part
SAVING LIVES in an antibiotic-resistant world by Julie O Connor
 SAVING LIVES in an antibiotic-resistant world by Julie O Connor 16 Imagine this scenario. At a metro Detroit hospital emergency room, a four-year old girl with a severe case of vomiting, diarrhea, fever
SAVING LIVES in an antibiotic-resistant world by Julie O Connor 16 Imagine this scenario. At a metro Detroit hospital emergency room, a four-year old girl with a severe case of vomiting, diarrhea, fever
ERG on multidrug-resistant P. falciparum in the GMS
 ERG on multidrug-resistant P. falciparum in the GMS Minutes of ERG meeting Presented by D. Wirth, Chair of the ERG Geneva, 22-24 March 2017 MPAC meeting Background At the Malaria Policy Advisory Committee
ERG on multidrug-resistant P. falciparum in the GMS Minutes of ERG meeting Presented by D. Wirth, Chair of the ERG Geneva, 22-24 March 2017 MPAC meeting Background At the Malaria Policy Advisory Committee
OIE international standards on Rabies:
 Regional cooperation towards eradicating the oldest known zoonotic disease in Europe Antalya, Turkey 4-5 December 2008 OIE international standards on Rabies: Dr. Lea Knopf Scientific and Technical Department
Regional cooperation towards eradicating the oldest known zoonotic disease in Europe Antalya, Turkey 4-5 December 2008 OIE international standards on Rabies: Dr. Lea Knopf Scientific and Technical Department
TEACHERS TOPICS A Lecture About Pharmaceuticals Used in Animal Patients
 TEACHERS TOPICS A Lecture About Pharmaceuticals Used in Animal Patients Elaine Blythe Lust, PharmD School of Pharmacy and Health Professions, Creighton University Submitted October 30, 2008; accepted January
TEACHERS TOPICS A Lecture About Pharmaceuticals Used in Animal Patients Elaine Blythe Lust, PharmD School of Pharmacy and Health Professions, Creighton University Submitted October 30, 2008; accepted January
UW College of Agriculture and Natural Resources Global Perspectives Grant Program Project Report
 UW College of Agriculture and Natural Resources Global Perspectives Grant Program Project Report COVER PAGE Award Period: Fall 2017 Fall 2018 Principle Investigator: Brant Schumaker Department: Veterinary
UW College of Agriculture and Natural Resources Global Perspectives Grant Program Project Report COVER PAGE Award Period: Fall 2017 Fall 2018 Principle Investigator: Brant Schumaker Department: Veterinary
BEHAVIOUR OF THE DOMESTIC DOG (Canis familiaris)
") THE INFLUENCE OF CEREBRAL LATERALISATION ON THE BEHAVIOUR OF THE DOMESTIC DOG (Canis familiaris) A thesis submitted for the Degree of DOCTOR OF PHILOSOPHY by Luke Aaron Schneider B. A. (Hons) School of
THE INFLUENCE OF CEREBRAL LATERALISATION ON THE BEHAVIOUR OF THE DOMESTIC DOG (Canis familiaris) A thesis submitted for the Degree of DOCTOR OF PHILOSOPHY by Luke Aaron Schneider B. A. (Hons) School of
Antimicrobial Therapy
 Chapter 12 The Elements of Chemotherapy Topics - Antimicrobial Therapy - Selective Toxicity - Survey of Antimicrobial Drug - Microbial Drug Resistance - Drug and Host Interaction Antimicrobial Therapy
Chapter 12 The Elements of Chemotherapy Topics - Antimicrobial Therapy - Selective Toxicity - Survey of Antimicrobial Drug - Microbial Drug Resistance - Drug and Host Interaction Antimicrobial Therapy
Fungal Disease. What is a fungus?
 Fungal Disease What is a fungus? A fungus is a living organism. It goes through a complicated life cycle and is able to spread in the environment by producing large numbers of spores that are easily dispersed
Fungal Disease What is a fungus? A fungus is a living organism. It goes through a complicated life cycle and is able to spread in the environment by producing large numbers of spores that are easily dispersed
Antimicrobial agents. are chemicals active against microorganisms
 Antimicrobial agents are chemicals active against microorganisms Antibacterial Agents Are chemicals active against bacteria Antimicrobials Antibacterial Antifungal Antiviral Antiparasitic: -anti protozoan
Antimicrobial agents are chemicals active against microorganisms Antibacterial Agents Are chemicals active against bacteria Antimicrobials Antibacterial Antifungal Antiviral Antiparasitic: -anti protozoan
Comparing DNA Sequences Cladogram Practice
 Name Period Assignment # See lecture questions 75, 122-123, 127, 137 Comparing DNA Sequences Cladogram Practice BACKGROUND Between 1990 2003, scientists working on an international research project known
Name Period Assignment # See lecture questions 75, 122-123, 127, 137 Comparing DNA Sequences Cladogram Practice BACKGROUND Between 1990 2003, scientists working on an international research project known
5/3/2018 3:09 AM Approved (Changed Course) ANHLT 151 Course Outline as of Fall 2017
 ANHLT 151 Course Outline as of Fall 2017") 5/3/2018 3:09 AM Approved (Changed Course) ANHLT 151 Course Outline as of Fall 2017 CATALOG INFORMATION Dept and Nbr: ANHLT 151 Title: VET LAB IMAGING PROC Full Title: Veterinary Laboratory and Imaging
5/3/2018 3:09 AM Approved (Changed Course) ANHLT 151 Course Outline as of Fall 2017 CATALOG INFORMATION Dept and Nbr: ANHLT 151 Title: VET LAB IMAGING PROC Full Title: Veterinary Laboratory and Imaging
Veterinary Parasitology
 Veterinary Parasitology 186 (2012) 159 164 Contents lists available at SciVerse ScienceDirect Veterinary Parasitology jo u rn al hom epa ge : www.elsevier.com/locate/vetpar The therapeutic efficacy of
Veterinary Parasitology 186 (2012) 159 164 Contents lists available at SciVerse ScienceDirect Veterinary Parasitology jo u rn al hom epa ge : www.elsevier.com/locate/vetpar The therapeutic efficacy of
Essential Principles of Horseshoeing
 A Primer for the Doug Butler Enterprises 21st Century approach to Farrier Education! Essential Principles of Horseshoeing A first-of-it s kind resource providing step-by-step instruction and corresponding
A Primer for the Doug Butler Enterprises 21st Century approach to Farrier Education! Essential Principles of Horseshoeing A first-of-it s kind resource providing step-by-step instruction and corresponding
Exotic Hematology Lab Leigh-Ann Horne, LVT, CWR Wildlife Center of Virginia
 Exotic Hematology Lab Leigh-Ann Horne, LVT, CWR Wildlife Center of Virginia lhorne@wildlifecenter.org Anne Lynch, LVT Cedarcrest Animal Clinic amllvt9@gmail.com Introduction While the general set-up for
Exotic Hematology Lab Leigh-Ann Horne, LVT, CWR Wildlife Center of Virginia lhorne@wildlifecenter.org Anne Lynch, LVT Cedarcrest Animal Clinic amllvt9@gmail.com Introduction While the general set-up for
What causes heartworm disease?
 Heartworm Disease: What causes heartworm disease? Heartworm disease (dirofilariasis) is a serious and potentially fatal disease in dogs and cats. It is caused by a blood-borne parasite called Dirofilaria
Heartworm Disease: What causes heartworm disease? Heartworm disease (dirofilariasis) is a serious and potentially fatal disease in dogs and cats. It is caused by a blood-borne parasite called Dirofilaria
Tick-Borne Disease Diagnosis: Moving from 3Dx to 4Dx AND it s MUCH more than Blue Dots! indications implications
 Tick-Borne Disease Diagnosis: Moving from 3Dx to 4Dx Richard B. Ford, DVM, MS Professor of Medicine Diplomate ACVIM and (Hon) ACVPM North Carolina State University Raleigh, NC In just the past 3 to 5 years,
Tick-Borne Disease Diagnosis: Moving from 3Dx to 4Dx Richard B. Ford, DVM, MS Professor of Medicine Diplomate ACVIM and (Hon) ACVPM North Carolina State University Raleigh, NC In just the past 3 to 5 years,
مادة االدوية المرحلة الثالثة م. غدير حاتم محمد
 م. مادة االدوية المرحلة الثالثة م. غدير حاتم محمد 2017-2016 ANTIMICROBIAL DRUGS Antimicrobial drugs Lecture 1 Antimicrobial Drugs Chemotherapy: The use of drugs to treat a disease. Antimicrobial drugs:
م. مادة االدوية المرحلة الثالثة م. غدير حاتم محمد 2017-2016 ANTIMICROBIAL DRUGS Antimicrobial drugs Lecture 1 Antimicrobial Drugs Chemotherapy: The use of drugs to treat a disease. Antimicrobial drugs: