Mastozoología Neotropical ISSN: Sociedad Argentina para el Estudio de los Mamíferos. Argentina
|
|
|
- Emory Cannon
- 6 years ago
- Views:
Transcription
1 Mastozoología Neotropical ISSN: Sociedad Argentina para el Estudio de los Mamíferos Argentina Amelia Chemisquy, M.; Prevosti, Francisco J. IT TAKES MORE THAN LARGE CANINES TO BE A SABRETOOTH PREDATOR Mastozoología Neotropical, vol. 21, núm. 1, 2014, pp Sociedad Argentina para el Estudio de los Mamíferos Tucumán, Argentina Available in: How to cite Complete issue More information about this article Journal's homepage in redalyc.org Scientific Information System Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Non-profit academic project, developed under the open access initiative
2 Mastozoología Neotropical, 21(1):27-36, Mendoza, 2014 Versión impresa ISSN Versión on-line ISSN Copyright SAREM, Artículo IT TAKES MORE THAN LARGE CANINES TO BE A SABRETOOTH PREDATOR M. Amelia Chemisquy 1 and Francisco J. Prevosti 1, 2 1 División Mastozoología, Museo Argentino de Ciencias Naturales Bernardino Rivadavia (MACN CONICET), Av. Angel Gallardo 470, 1405 Ciudad Autónoma de Buenos Aires, Argentina [correspondence: M. Amelia Chemisquy <amelych80@gmail.com>]. 2 Departamento de Ciencias Básicas, Universidad Nacional de Luján, Cruce de Rutas 5 y 7, 6700 Luján, Buenos Aires, Argentina. ABSTRACT. Monodelphis dimidiata is a small marsupial from southern South America. It is a true semelparous species that develops an extreme sexual dimorphism associated to the attaining of sexual maturity, both on the size, weight and skull morphology, including the development of sabre-like canines in males. A recent paper considered M. dimidiata males to be pigmy sabretooth predators, based mainly on morphometric analyses. Here we study the skull morphometry (including canine size) of M. dimidiata in comparison with other marsupials, living felids and extinct sabretooth predators, looking for convergences with the latter. We also put the hypothesis of M. dimidiata as a sabretooth predator in the context of its life history, reinterpreting the origin of its sabre-like canine and its suitability as a living analogue of primitive sabretooth predators. We found that the skull pattern of M. dimidiata is not different from other didelphid marsupials, and even Didelphis albiventris has canines of the same relative length. We consider that the large canines of M. dimidiata are a byproduct of the exacerbated growth of males, caused by their delayed eruption, as well as by their late apexification. Large canines are related to the particular reproductive cycle of this species instead of being an adaptation to hunt large preys, as was proposed for sabretooth cats. RESUMEN. Se precisa más que caninos grandes para ser un predador dientes de sable. Monodelphis dimidiata es un pequeño marsupial del sur de América del Sur. Es una especie con semelparía verdadera, que desarrolla un dimorfismo sexual extremo asociado a la madurez sexual, tanto en el tamaño, el peso y la morfología craneana, incluyendo el desarrollo de caninos tipo sable en los machos. Un trabajo reciente considera a los machos de M. dimidiata como depredadores dientes de sable pigmeos, sobre la base de análisis morfométricos. Aquí se estudia la morfometría del cráneo (incluyendo el tamaño de los caninos) de M. dimidiata en comparación con otros marsupiales, félidos vivientes y especies extintas de depredadores dientes de sable, en busca de convergencias con el último grupo. También se pone a prueba la hipótesis de M. dimidiata como depredador dientes de sable en el contexto de su historia de vida, reinterpretando el origen de su canino tipo sable y si es apropiado como un análogo viviente de los depredadores dientes de sable primitivos. Se encontró que la morfología general del cráneo de M. dimidiata no es diferente de otros marsupiales didélfidos, e incluso Didelphis albiventris tiene caninos del mismo largo relativo. Consideramos que los grandes caninos de M. dimidiata son un subproducto del crecimiento exacerbado de los machos, causado por su erupción retrasada, así como por su apexificación tardía. Los caninos grandes parecen estar relacionados con el ciclo reproductivo particular de esta especie en lugar de ser una adaptación a cazar presas grandes, como se propuso para los tigres dientes de sable. Key words: Canine growth. Didelphis albiventris. Marsupials. Monodelphis dimidiata. Palabras clave: Crecimiento de los caninos. Didelphis albiventris. Marsupiales. Monodelphis dimidiata. Recibido 19 marzo Aceptado 19 mayo Editor asociado: D Flores
3 28 Mastozoología Neotropical, 21(1):27-36, Mendoza, MA Chemisquy and FJ Prevosti INTRODUCTION Monodelphis dimidiata (Wagner, 1847) is a small didelphid marsupial from the Pampas and Atlantic Forest domains of Argentina, Brazil, Uruguay, and Paraguay (as M. sorex; Pine and Handley, 2007; Smith, 2008; Vilela et al., 2010). This small mammal has the peculiarity of being semelparous, meaning that its life cycle is reduced to one reproductive event (1 year in this case), where males die after reproduction and females after the weaning of the offspring (Pine et al., 1985; Baladrón et al., 2012). Associated with the attaining of sexual maturity, both sexes increase the growth rate, being this rate more accentuated in males, which are significantly larger than females (Pine et al., 1985). This extreme sexual dimorphism is not only evident in the size and weight of males, but also in the skull morphology, where males have larger cranial crests than females, enlarged postorbital processes on the frontals, wider zygomatic arches and sabre-like canines (Pine et al., 1985:217). In a recent paper, Blanco et al. (2013) rediscovered the sabre-like canines of M. dimidiata and considered the species (particularly males) as a pigmy sabretooth predator, based on a morphometric analysis and the comparison with measurements of sabretooth predators published by Emerson and Radinsky (1980), and the observations of animals held in captivity published by González and Claramunt (2000). Blanco et al. (2013) stated, using indices previously defined by Emerson and Radinsky (1980), that M. dimidiata have canines relatively larger than the other marsupials analyzed, and even the indices calculated by Blanco and coworkers were similar to those of sabretooth predators. Also, the authors found, in a principal component analysis of several species of marsupials, that males of M. dimidiata could be separated in the fifth principal component mainly by the loadings of several variables that could be related to a sabretooth condition: large upper canine height and anteroposterior length, short masseteric fossa length and lever arm of temporalis, long distance from the mandibular condyle to the M3, and short jaw (Blanco et al., 2013). The authors also found that the humerus of males of M. dimidiata is very robust, analogue to what happens in some sabretooth cats (e.g., Meachen-Samuels and Van Valkenburgh, 2010). This combination of characters led the authors to conclude that M. dimidiata might be a living analogue of the primitive sabretooth condition, with hyperthrophied canines, a wider gape, lower bite force, and a strong humerus. The conclusion of Blanco et al. (2013) is intriguing, because the morphological skull pattern, diet (mostly insectivorous), and size of M. dimidiata differ greatly from sabretooth cats (Pine et al., 1985; Busch and Kravetz, 1991; Pine and Handley, 2007). These differences make it difficult to see how further evolutionary, behavioral and ecological studies of M. dimidiata would provide a better understanding of the origin and behavior of sabretooths in the past (Blanco et al., 2013:9), and leave the possibility that the similarities found by the authors are only convergences related to other causal factors. In this paper we study the morphometry of the skull (including canine size) of M. dimidiata in comparison to other marsupials, living felids and extinct sabretooth predators (Felidae, Nimravidae, Sparassodonta), looking for convergences with the last group. We also put the findings of Blanco et al. (2013) in the context of the life history of M. dimidiata, reinterpreting the origin of its sabre-like canine and reassessing its suitability as a living analogue of primitive sabretooth predators. MATERIALS AND METHODS We reanalyzed the data of Blanco et al. (2013) adding two large males and one large female of M. dimidiata (ages 6/7 following Gardner, 1973), five Didelphis albiventris of ages 6 and 7, and two adults of each of the following living felids: Neofelis nebulosa, Panthera leo, Puma concolor and Leopardus tigrinus (Appendix 1, supplementary material). We also included in the analyses the sample of sabretooth predators (SP, hereafter) of the clades Felidae, Nimravidae, and Sparassodonta from Emerson and Radinsky (1980). Measurements C1W (upper canine mediolateral width) and OCPH (occipital height) were excluded since we did not have data for the SP and Neofelis, respectively. It is important to mention that both measurements
4 Monodelphis dimidiata IS NOT A SABRETOOTH 29 were also excluded by Blanco et al. (2013) to obtain a level of variation above the Jolliffe cut-off point and consequently are not relevant to the discussion. Since the taxonomic sample we are analyzing is a mixture of felids and marsupials, we consider that the length from the condyle to the third lower molar (COM3) in marsupials is analogue to the length from the condyle to the first lower molar (COM1) in Feliformia (see Jones, 2003). We performed a principal component analysis (PCA) using indices obtained from the 12 measurements (dividing the measurements by the skull length or the jaw length in order to standardize them), and one using the final 6 indices selected by Blanco et al. (2013). Since these indices are ratios, we log-transformed the data prior to performing the PCA (James and McCulloch, 1990). The variancecovariance matrix was used to obtain the eigenvalues and eigenvectors. Blanco et al. (2013) also used allometric equations based on their sample to estimate an expected value for each measurement. Then, they obtained an index as the ratio between the measured value and the expected value obtained from the equations, and compared those indices with the data from Emerson and Radinsky (1980). The problem with that approach is that Blanco et al. (2013) compared indices obtained using different allometric equations (since they did not include the data from Emerson and Radinsky in their estimations), which is incorrect and can lead to wrong conclusions. Other problem with this approach is that it does not take into account the phylogenetic signal of the data, biasing the allometric pattern obtained (e.g., Harvey and Pagel, 1991). To avoid these problems we prefer to estimate the relative sizes of the canines and other measurements, dividing each measurement by the geometric mean of all the measurements used in this study (Jungers et al., 1995). Finally, as another way of analyzing overall similarity, we performed a cluster analysis using the measurements above mentioned divided by their geometric mean (log-transformed). For the cluster analysis we used the unweighted pair-group average algorithm (UPGMA), computing the distance matrix using the Euclidean matrix. All the statistical analyses were performed using the software PAST vers (Hammer et al., 2001). RESULTS In the PCA where we used the 11 indices, the first principal component (PC1) explained 71.6% of total variance, while the second principal component (PC2) explained 9.8% of total variance. PC1 separated the SP from the remaining taxa (Fig. 1). Except for Dinictis, SP were placed on the negative end of PC2, while living felids were placed on its positive end (Fig. 1). M. dimidiata was mostly placed on the negative side of PC2, in the same morphospace of Didelphis albiventris and other species of Didelphis (Fig. 1). Regarding its position in PC1, M. dimidiata and D. albiventris were at the same coordinates than living felids (except Neofelis) and Sarcophilus harrisii (Fig. 1). The remaining marsupials were placed on the negative side of PC1. Canine height and length (C1H and C1L) and jaw height (JH) were the most influential variables for PC1, the three increasing towards its positive end (Table 1). For PC2 the variables that contributed the most were: zygomatic arch width (ZAW), occipital width (OCPW), temporal fossa length (TFL), jaw height (JH), masseteric fossa length (MFL) and the moment arm of the temporalis (MAT) towards the positive end, while tooth row length (TRL) and C1L increased towards the negative end of PC2 (Table 1). None of the remaining 9 PCs placed M. dimidiata close to the sabretooth predators. Using the 6 indices selected by Blanco et al. (C1L, C1H, JL, COM3, MFL, and MAT) the pattern was similar, except that some specimens of M. dimidiata, D. albiventris and the two specimens of S. harrisii were placed in the same morphospace that some living felids (not shown). Again, none of the PCs placed M. dimidiata close to the sabretooth predators. The relative height of canines of M. dimidiata (as shown by dividing C1H by the geometric mean) did not escape the morphospace of other marsupials, widely overlapping with the relative height of canines of D. albiventris (Fig. 2a). Among marsupials, the highest value belonged to a M. dimidiata female (MACN [Museo Argentino de Ciencias Naturales] 24458). There was no overlap between SP and any of the other taxa, being N. nebulosa the closest species to the sabretooth predators (Fig. 2a). The same pattern was observed for the relative length of canines, where M. dimidiata did not differ from other marsupials and SP did not overlap with any other taxa (Fig. 2b). Finally, the relative distance from the mandibular condyle to the
5 30 Mastozoología Neotropical, 21(1):27-36, Mendoza, MA Chemisquy and FJ Prevosti Fig. 1. Plots of PC1 versus PC2 from principal component analysis of the 11 indices, for marsupials, felids and sabretooth predators from the sample of Emerson and Radisnky (1980). Table 1 PC loadings (eigenvectors) obtained from the principal component analysis (PCA) of the 11 indices. Acronyms: C1H = upper canine height; C1L = upper canine anteroposterior length; COM3 = condyle to M3; JL = jaw length; OCPW = occipital width; SL = skull length; TFL = temporal fossa length; TRL = tooth row length; ZAW = zygomatic arch width; JH = jaw height; MFL = masseteric fossa length; MAT = moment arm of temporalis; PC, principal component. Indices PC1 PC2 PC % 9.76% 7.12% C1H/SL C1L/SL JL/SL OCPW/SL TFL/SL TRL/SL ZAW/SL COM3/JL JH/JL MFL/JL MAT/JL M3 was highly overlapped among marsupials, being D. albiventris the species with the highest distance (Fig. 2c). The complete set of measurements and their indices are available on Appendix 2 (supplementary material). The cluster analysis (cophenetic correlation coefficient = ) was consistent with the other analyses, and showed no relation between M. dimidiata and the SP. Most sabretooth predators (except Dinictis) were placed in a cluster distant from the remaining taxa. M. dimidiata grouped with D. albiventris, D. marsupialis, Caluromys lanatus and one specimen of Lutreolina crassicaudata. S. harrisii grouped with some of the living felids, while N. nebulosa was clustered with Dinictis (Fig. 3 supplementary material). DISCUSSION Our results clearly show that in a morphological context, M. dimidiata does not escape from the morphometric space of other didelphids (Figs. 1 and 3 [supplementary material]), and is far from the sabretooth predator morphotype (Fig. 1). The PCA including all the variables separated the SP along PC1 mainly by the size of the canines, and placed them on the positive end of PC1 and towards the negative side of PC2. In this analysis, M. dimidiata was the extant marsupial with higher PC1 scores, but was widely overlapped with D. albiventris and only one female (MACN 24458) had a slightly higher score. N. nebulosa, a felid considered by some authors to be a living analogue of sabretooth cats (Christiansen, 2006, 2008a, 2008b) was placed on the positive end of PC1, being the closest living taxa to SP. However, like other living felids, N. nebulosa was also on the positive end of PC2, mainly due to a wider zygomatic arch, larger temporal and masseteric
6 Monodelphis dimidiata IS NOT A SABRETOOTH 31 Fig. 2. Boxplots summarizing the information from the analyses of three indices (a) upper canine height/geometric mean, (b) upper canine anteroposterior length/geometric mean, and (c) condyle to M3 or M1/ geometric mean. muscle advantage, longer temporal fossa and wider braincase. SP were placed on the positive end of PC1, with more specialized taxa (e.g., Thylacosmilus, Barbourofelis) having higher PC1 scores, and negative PC2 scores, while less specialized species (e.g., Dinictis) showed positive PC2 scores and were placed forming a continuum between living felids and the highly specialized SP (Fig. 1). This pattern of continuum variation between felids and SP (though with derived SP occupying a different portion of the morphospace), and even the superposition between less specialized SP and felids, was found previously, using different taxonomic samples and geometric morphometric analyses (e.g., Slater and Van Valkenburgh, 2008; Christiansen, 2008; Prevosti et al., 2010). With the variables used here we can state that specialized SP have longer and larger upper canines, narrower skulls, longer dental series and lower mandible, and smaller fossae for temporal and masseteric muscles and shorter coronoid process, while less modified taxa are more similar to felids (specially to Neofelis), presenting the opposite pattern (Fig. 1). In this analysis is also clear that living marsupials do not invade the morphospace of SP, and that Monodelphis and Didelphis are in the periphery of the marsupial sample. Though Monodelphis and Didelphis showed PC1 scores similar to living felids, because of their relatively longer and larger canines, their skull anatomy is not different from other marsupials, and is strikingly different from living felids and SP (Figs. 1 and 4). The living marsupial that showed a more divergent anatomy in the PCA analysis was S. harrisii, which also has relatively larger canines, and a skull proportion similar to living felids (at least with the variables used here; Fig. 1). This agrees with the more predatory and scavenger habits reported for the Tasmanian devil (e.g., Strahan, 1998; Nowak, 2005). The PCA arrangement described in this paper contrasts with the analyses published by Blanco et al. (2013), which could be related to the inclusion of more didelphid specimens, but mainly to the inclusion of felids and sabretooth predators. However, the principal component analyses performed by Blanco et al. (2013) deserve two important criticisms. First, we think that the procedure followed by the authors to reduce the number of variables in the PCA is highly questionable, since they manipulated the data until they obtained the results they expected (i.e., M. dimidiata in a different morphospace than the remaining marsupials). Moreover, the reduced PCA has a limited representation of variables (i.e., different morphometric aspects of the skull), and reduces the entire skull shape
7 32 Mastozoología Neotropical, 21(1):27-36, Mendoza, MA Chemisquy and FJ Prevosti to six measurements, none of which refers to the cranium. Second, the authors used ratios as variables, but they did not log-transformed the data. Log-transforming the data is important not only because it helps normalizing the data and achieving homoscedasticity (Legendre and Legendre, 1998), but in the case of ratios, when one has a product of variances, the transformation helps making the relationship between variables linear, which is a requirement for PCA (James and McCulloch, 1990). We performed the same PCAs conducted by Blanco et al. (2013) (i.e., without adding specimens) but log-transforming the variables, and found that the pattern they described disappears, both for the complete and the reduced analysis (Fig. 5 supplementary material). Consequently, the authors should reconsider their findings, not only based on the discussion presented here, but also based on a more accurate statistical analysis of their data. The same tendency found in the PCA was observed when analyzing the relative size of M. dimidiata canines, which were not different from the relative size of D. albiventris canines, even overlapping with the canine size of the other marsupials analyzed. Also, the distance between the mandibular condyle and the third lower molar was no different in M. dimidiata versus other marsupials, and even the largest values belonged to a specimen of D. albiventris. Consequently, there is nothing on the morphological features analyzed here that indicates that M. dimidiata is a pigmy sabretooth predator (see Fig. 4 for a comparison among species). Even the size of the canines is discarded as a distinctive feature, since there is a wide overlap with D. albiventris. A lateral view of the skull of the specimen MACN (Fig. 4c) clearly resembles the sabre-like canines of M. dimidiata males (Fig. 4b). We believe that Blanco et al. (2013) failed to see that overlap not only due to the indices they used (see above), but also because the three specimens of D. albiventris that they selected were small, while we included the largest specimens of D. albiventris available (as well as we did with M. dimidiata). A striking result was the position of the M. dimidiata female MACN (Fig. 4a), which had the highest canines among marsupials and was the only specimen that escaped from the morphometric space of the marsupials in the reduced PCA. This suggests that some females also have the potential of developing a large body size and large canines, probably in cases where there are available good food sources, or in cases of reduced fecundity, as suggested by Gardner (1973) for Didelphis virginiana. Consequently, the issue of M. dimidiata as a pigmy sabretooth predator is not only associated with males, as stated by Blanco et al. (2013), but also with some females. If the sabretooth predator morph in M. dimidiata is mainly restricted to adult males, we should expect to find differences between sexes in diet, behavior and ecology, which is not the case. The diet analysis of M. dimidiata performed by Goin et al. (1992) supports the idea that females are not that different from males, since they found no differences in the stomach contents of both sexes, not even in adults of over 60 g (i.e., after attaining sexual maturity). In fact, up to date, there is no published evidence of differences in the diet between males and females. There are speculations that males would be more carnivorous than females (e.g. Pine et al., 1985), but in captivity females killed mice as avidly as males did (González and Claramunt, 2000). The fact that females kill and consume mice in the same way males do, goes against the behavioral explanation proposed by Blanco et al. (2013) for the convergence between M. dimidiata and sabretooth predators, since the average adult female has much shorter canines than males, as well as a shorter distance from the mandibular condyle to the m3. As previously suggested, it is clear that the large canines of males are not related (or at least not strongly related) to the feeding habits of M. dimidiata (Goin et al., 1992; González and Claramunt, 2000). In addition, contrary to other carnivorous didelphid marsupials such as L. crassicaudata (Mares et al., 1989), M. dimidiata is not considered to be an effective mammalian predator, and there are even some reports that, when put on the same cage, they coexisted with mice (Pine et al., 1985, and references therein). The majority of possible rodent preys described for M. dimidiata are smaller than this marsupial (Busch and Kravetz,
 Smilodon fatalis YPM 11532, (e) Hophlophoneus oreodontis YPMPU 13628, (f) Neofelis nebulosa FMNH 75831. Scale = 10 mm. 1991), only Oxymycterus rufus (= O.")
8 Monodelphis dimidiata IS NOT A SABRETOOTH 33 Fig. 4. Lateral view of the skull of (a) Monodelphis dimidiata MACN 24458, (b) M. dimidiata MACN 24440, (c) Didelphis albiventris MACN , (d) Smilodon fatalis YPM 11532, (e) Hophlophoneus oreodontis YPMPU 13628, (f) Neofelis nebulosa FMNH Scale = 10 mm. 1991), only Oxymycterus rufus (= O. rutilans) and Necromys obscurus (= Bolomys obscurus) could be considered to be similar in size to an adult M. dimidiata (52 g and 86 g respectively; Fornes and Massoia, 1965). Additionally, laboratory mice (Mus musculus) weight less than half the weight of a grown adult male (González and Martínez Lanfranco, 2010), so M. dimidiata rarely kills rodents larger than themselves. We believe that the morphological convergences between M. dimidiata and SP described by Blanco et al. (2013) are a byproduct of the ontogenetic development of M. dimidiata males, and have little to do with ecological or trophic factors. Once males attain sexual maturity, they start to grow faster than females, ending up being larger (Pine et al., 1985). Since males have this exacerbated growth once they have the complete dentition, it is expected that they will develop a large distance between the coronoid process and the last molar, since molars had already erupted in their definite size when males were much smaller and had smaller mandibles to accommodate the dentition. This increases the COM3 measurement, but merely as a consequence of the enlargement of the mandible, and not as a biomechanical requirement for a wider gape. Moreover, large females that did not develop large canines have a relative COM3 value similar to males with larger canines (Fig. 6 supplementary material), meaning that the long distance from the mandibular condyle to the third lower molar is not a modification to maintain mandible functionality with hypertrophied canines, as was suggested by Blanco et al. (2013), and is merely a consequence of the enlargement of the mandible after the eruption of the molars. Something similar happens with canines; Jones (2003) mentioned that canine teeth of marsupial carnivores never cease to erupt. A radiographic analysis of the canines of M. dimidiata showed that individuals that completed their adult dentition have a wide pulp cavity, with an open apex with thin walls on the root canal (Fig. 7a), meaning that the canines still have the potentiality of growing. Even in large sexually mature males and females the apex remains open, and apexification (i.e., tapering of the root apex) is not complete (Figs. 7b and 7c). This means that the long canines are also a byproduct of the exacerbated growth of males, and evidently the same factors that cause the development of the cranial crests, the enlargement of the postorbital processes on the frontals and the widening of the zygomatic
 MACN 22474, (b) MACN 22458, and (c) MACN 24440. arches also affect canine growth (and humerus robustness).")
9 34 Mastozoología Neotropical, 21(1):27-36, Mendoza, MA Chemisquy and FJ Prevosti Fig. 7. Radiographs showing the canine pulp cavity and the open apex in Monodelphis Dimidiata: (a) MACN 22474, (b) MACN 22458, and (c) MACN arches also affect canine growth (and humerus robustness). Consequently, this delay in canine eruption and the late apexification causes that species such as D. albiventris that continue to grow throughout their life (Gardner, 1973) develop large canines, as well as M. dimidiata males, which have an accelerated growth in the reproductive season (Pine et al., 1985). In the case of the large females of M. dimidiata, the situation is similar to the one in D. albiventris, where the canine enlargement is caused by the body growth. The acquisition of these long canines in M. dimidiata is clearly not related to predatory habits as it is in sabretooth predators (Akersten, 1985; Anton and Galobart, 1999). Finally, finding a living analogue for the extinct sabretooth cats is an appealing idea, and surely M. dimidiata could be somehow considered one. But similarly, it is possible to find analogues to different aspects of the sabertooth cat mechanics in different vertebrates. For example, snakes were used as a model for the sabretooth stabbing (Simpson, 1935; Scott, 1937), while varanid lizards were considered to be a good analogue based not only on tooth morphology, but also on ecological features (Akersten, 1985). There are many other examples of mammals which have long canines, such as many species of primates, the water deer (Hydropotes inermis), and the walrus (Odobenus rosmarus); all of them might be sharing some similarities with SP of the past. Even the hippos (Hippopotamus amphibius), with their wide gape and reduced coronoid process could be considered a living analogue of sabretooth predators. However, we feel that long canines and a wide gape are not enough to be considered a sabretooth predator, particularly in the case of M. dimidiata, which can hardly be considered a predator of large preys. We believe that evolutionary, behavioral and ecological studies of M. dimidiata will be as useful for understanding the origin and behavior of SP of the past as the same studies in D. albiventris, especially when taking into account the broad differences between the morphology, behavior and life history of M. dimidiata and any member of Felidae, Nimravidae or Sparassodonta. N. nebulosa, due to morphological similarities (see above) and phylogenetic relationships is clearly the best living analogue that we have for these extinct predators. ACKNOWLEDGEMENTS We are grateful to Juan José Kraus for helping with the X-rays and for the discussion on tooth formation and development; Gabriel Martin for the discussion on feeding habits of mouse opossums; Guillermo Cassini for the statistical help; Miriam Morales for the data on N. nebulosa; Ulyses Pardiñas for the data on rodents weights; David Flores (MACN), Damián Romero (Museo Municipal de Cs. Naturales de Mar del Plata), Bill Simpson (Field Museum of Natura History) and Walter Joyce (Yale Peabody Muesum) for allowing the analysis of material under their care; F. Abdala and an anonymous reviewer for their comments on the manuscript. CONICET provided financial support. This is
10 Monodelphis dimidiata IS NOT A SABRETOOTH 35 a contribution to PICT 2012 Nº 0256 to MAC, PICT and PIP to FP. LITERATURE CITED AKERSTEN WA Canine function in Smilodon (Mammalia, Felidae, Machairodontinae). Contributions in Science, Natural History Museum of Los Angeles County 356:1-22. ANTÓN M and A GALOBART Neck function and predatory behavior in the scimitar toothed cat Homotherium latidens (Owen). Journal of Vertebrate Paleontology 19: BALADRÓN AV, AI MALIZIA, MS BÓ, MS LIÉBANA, and MJ BECHARD Population dynamics of the southern short-tailed opossum (Monodelphis dimidiata) in the Pampas of Argentina. Australian Journal of Zoology 60: BLANCO RE, WW JONES, and N MILNE Is the extant southern short-tailed opossum a pigmy sabretooth predator? Journal of Zoology 291: BUSCH M and FO KRAVETZ Diet composition of Monodelphis dimidiata (Marsupialia, Didelphidae). Mammalia 55: CHRISTIANSEN P Sabertooth characters in the clouded leopard (Neofelis nebulosa Griffiths 1821). Journal of Morphology 267: CHRISTIANSEN P. 2008a. Evolution of skull and mandible shape in cats (Carnivora: Felidae). PLOS ONE 3:e2807. CHRISTIANSEN P. 2008b. Evolutionary convergence of primitive sabertooth craniomandibular morphology: The clouded leopard (Neofelis nebulosa) and Paramachairodus ogygia compared. Journal of Mammalian Evolution 15: EMERSON SB and LB RADINSKY Functional analysis of sabertooth cranial morphology. Paleobiology 6: FORNES A and E MASSOIA Micromamíferos (Marsupialia y Rodentia) recolectados en la localidad bonaerense de Miramar. Physis 25: GARDNER AL The systematics of the genus Didelphis (Marsupialia: Didelphidae) in North and Middle America. Special Publications, the Museum Texas Tech University 4:1-81. GOIN FJ, C VELÁZQUEZ, and O SCAGLIA Orientación de las crestas cortantes en el molar tribosfénico. Sus implicancias funcionales en didelfoideos (Marsupialia) fósiles y vivientes. Revista del Museo de La Plata (nueva serie) 9: GONZÁLEZ EM and S CLARAMUNT Behaviors of short-tailed opossums, Monodelphis dimidiata. Mammalia 64: GONZÁLEZ EM and JA MARTÍNEZ LANFRANCO Mamíferos de Uruguay. Ediciones de la Banda Oriental, Montevideo. HAMMER Ø, DAT HARPER, and PD RYAN PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica 4:1-9. HARVEY PH and MD PAGEL The comparative method in evolutionary biology. Oxford University, New York. JACKSON JE A User s Guide to Principal Components. Wiley, New York. JAMES FC and CE MCCULLOCH Multivariate analysis in ecology and systematics: Panacea or Pandora s Box? Annual Review of Ecology and Systematics 21: JONES ME Convergence in ecomorphology and guild structure among marsupial and placental carnivores. Pp , in: Predators with pouches: The biology of carnivorous marsupials (M Jones, C Dickman, and M Archer, eds.). CSIRO Publishing, Melbourne. JUNGERS WL, AB FALSETTI, and CE WALL Shape, relative size, and size-adjustments in morphometrics. Yearbook of Physical Anthropology 38: LEGENDRE P and L LEGENDRE Numerical ecology. 2nd edn. Elsevier, Amsterdam. MARES MA, RA OJEDA, and RM BARQUEZ Guide to the mammals of Salta Province, Argentina. University of Oklahoma Press, Norman and London. MEACHEN-SAMUELS J and B VAN VALKENBURGH Forelimb indicators of prey-size preference in the Felidae. Journal of Morphology 270: MEACHEN-SAMUELS J and B VAN VALKENBURGH Radiographs reveal exceptional forelimb strength in the sabertooth cat, Smilodon fatalis. PLOS ONE 5:e NOWAK RM Walker s marsupials of the world. The Johns Hopkins University Press, Baltimore. PINE RH and CO HANDLEY Jr Genus Monodelphis. Pp , in: Mammals of South America, Vol. 1 (AL Gardner, ed.). The University of Chicago Press, Chicago. PINE RH, PL DALBY, and JO MATSON Ecology, postnatal development, morphometrics, and taxanomic status of the short-tailed opossum, Monodelphis dimidiata, an apparently semelparous annual marsupial. Annals of the Carnegie Museum 54: SCOTT WB A history of land mammals in the western hemisphere. The Macmillan Company, New York. SIMPSON GG The function of saber-like canines in carnivorous mammals. American Museum Novitates 1130:1-12. SLATER GJ and B VAN VALKENBURGH Long in the tooth: Evolution of sabertooth cat cranial shape. Paleobiology 34: SMITH P FAUNA Paraguay Handbook of the Mammals of Paraguay Number 26 Monodelphis sorex. STRAHAN R The Mammals of Australia. Reed New Holland, London. VILELA JF, CA DE MORAES RUSSO, and JA DE OLIVEIRA An assessment of morphometric and molecular variation in Monodelphis dimidiata (Wagner, 1847) (Didelphimorphia: Didelphidae). Zootaxa 2646:26-42.
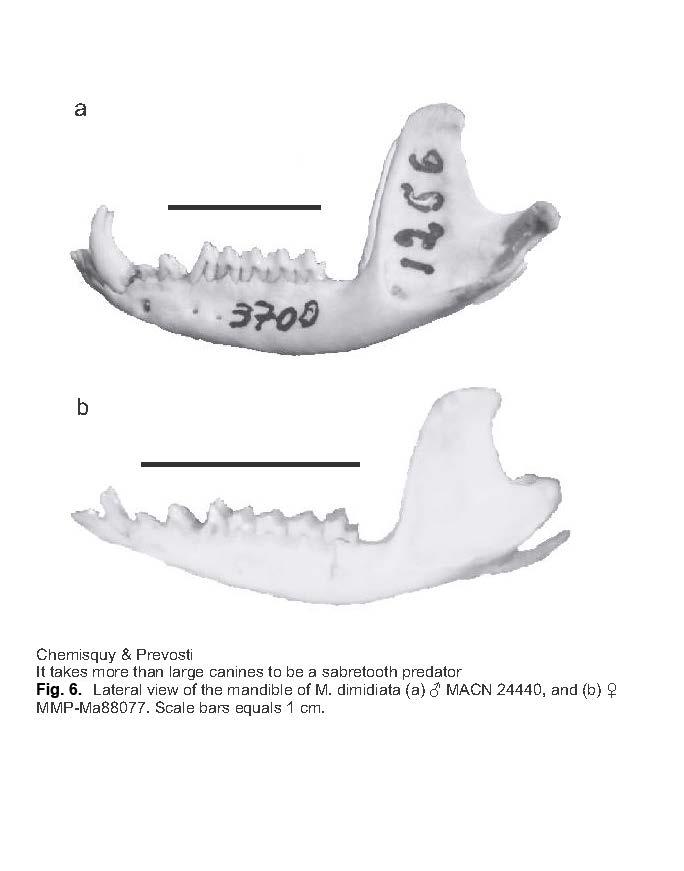
11 36 Mastozoología Neotropical, 21(1):27-36, Mendoza, MA Chemisquy and FJ Prevosti ONLINE SUPPLEMENTARY MATERIAL Fig. 3. Dendrogram obtained from the UPGMA analysis of the 11 measurements (divided by their geometric mean): upper canine height, upper canine anteroposterior length, condyle to M3, jaw length, occipital width, temporal fossa length, tooth row length, zygomatic arch width, masseteric fossa length, and moment arm of temporalis. Fig. 5. Repetition of the principal component analyses presented by Blanco et al. (2013) as evidence for the sabretooth condition of M. dimidiata, but log-transforming the variables; (a) PC1 vs. PC5 from the analysis using 14 indices; (b) PC1 vs. PC3 from the analysis using 6 indices. Fig. 6. Lateral view of the mandible of Monodelphis dimidiata: (a) MACN 24440, and (b) MMP-Ma Scale = 10 mm. Appendix 1. Measurements (in mm) of the specimens included in the analysis. Appendix 2. Cranial and mandibular indices of the species included in the analyses.
12
13
O'Regan HJ Defining cheetahs, a multivariante analysis of skull shape in big cats. Mammal Review 32(1):58-62.
:58-62.") O'Regan HJ. 2002. Defining cheetahs, a multivariante analysis of skull shape in big cats. Mammal Review 32(1):58-62. Keywords: Acinonyx jubatus/cheetah/evolution/felidae/morphology/morphometrics/multivariate
O'Regan HJ. 2002. Defining cheetahs, a multivariante analysis of skull shape in big cats. Mammal Review 32(1):58-62. Keywords: Acinonyx jubatus/cheetah/evolution/felidae/morphology/morphometrics/multivariate
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the
 Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the") Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
Fig. 5. (A) Scaling of brain vault size (width measured at the level of anterior squamosal/parietal suture) relative to skull size (measured at the distance between the left versus right temporomandibular
Chapter 2 Mammalian Origins. Fig. 2-2 Temporal Openings in the Amniotes
 Chapter 2 Mammalian Origins Fig. 2-2 Temporal Openings in the Amniotes 1 Synapsida 1. monophyletic group 2. Single temporal opening below postorbital and squamosal 3. Dominant terrestrial vertebrate group
Chapter 2 Mammalian Origins Fig. 2-2 Temporal Openings in the Amniotes 1 Synapsida 1. monophyletic group 2. Single temporal opening below postorbital and squamosal 3. Dominant terrestrial vertebrate group
CRANIAL EVIDENCE FOR SEXUAL DIMORPHISM AND GROUP LIVING IN THE EXTINCT AMERICAN LION (PANTHERA LEO ATROX)
") CRANIAL EVIDENCE FOR SEXUAL DIMORPHISM AND GROUP LIVING IN THE EXTINCT AMERICAN LION (PANTHERA LEO ATROX) Gold, David Department of Ecological and Evolutionary Biology Professor Matthew McHenry This study
CRANIAL EVIDENCE FOR SEXUAL DIMORPHISM AND GROUP LIVING IN THE EXTINCT AMERICAN LION (PANTHERA LEO ATROX) Gold, David Department of Ecological and Evolutionary Biology Professor Matthew McHenry This study
Mammalogy Lab 1: Skull, Teeth, and Terms
 Mammalogy Lab 1: Skull, Teeth, and Terms Be able to: Goals of today s lab Locate all structures listed on handout Define all terms on handout what they are or what they look like Give examples of mammals
Mammalogy Lab 1: Skull, Teeth, and Terms Be able to: Goals of today s lab Locate all structures listed on handout Define all terms on handout what they are or what they look like Give examples of mammals
Covariation in the skull modules of cats: the challenge of growing saber-like canines
 This article was downloaded by: [Smithsonian Institution Libraries] On: 28 January 2013, At: 08:24 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered
This article was downloaded by: [Smithsonian Institution Libraries] On: 28 January 2013, At: 08:24 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered
Title: Phylogenetic Methods and Vertebrate Phylogeny
 Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
Title: Phylogenetic Methods and Vertebrate Phylogeny Central Question: How can evolutionary relationships be determined objectively? Sub-questions: 1. What affect does the selection of the outgroup have
The melanocortin 1 receptor (mc1r) is a gene that has been implicated in the wide
 is a gene that has been implicated in the wide") Introduction The melanocortin 1 receptor (mc1r) is a gene that has been implicated in the wide variety of colors that exist in nature. It is responsible for hair and skin color in humans and the various
Introduction The melanocortin 1 receptor (mc1r) is a gene that has been implicated in the wide variety of colors that exist in nature. It is responsible for hair and skin color in humans and the various
Big Cat Rescue Presents. Tigrina or Oncilla
 Big Cat Rescue Presents Tigrina or Oncilla 1 Tigrina or Oncilla Big Cat Rescue 12802 Easy Street Tampa, Florida 33625 www.bigcatrescue.org Common Name: Oncilla Kingdom: Animalia Phylum: Chordata (Vertebrata)
Big Cat Rescue Presents Tigrina or Oncilla 1 Tigrina or Oncilla Big Cat Rescue 12802 Easy Street Tampa, Florida 33625 www.bigcatrescue.org Common Name: Oncilla Kingdom: Animalia Phylum: Chordata (Vertebrata)
CLADISTICS Student Packet SUMMARY Phylogeny Phylogenetic trees/cladograms
 CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
CLADISTICS Student Packet SUMMARY PHYLOGENETIC TREES AND CLADOGRAMS ARE MODELS OF EVOLUTIONARY HISTORY THAT CAN BE TESTED Phylogeny is the history of descent of organisms from their common ancestor. Phylogenetic
Evolution of Skull and Mandible Shape in Cats (Carnivora: Felidae)
") Evolution of Skull and Mandible Shape in Cats (Carnivora: Felidae) Per Christiansen* Department of Vertibrates, Zoological Museum, Copenhagen, Denmark Abstract The felid family consists of two major subgroups,
Evolution of Skull and Mandible Shape in Cats (Carnivora: Felidae) Per Christiansen* Department of Vertibrates, Zoological Museum, Copenhagen, Denmark Abstract The felid family consists of two major subgroups,
Introduction to Biological Anthropology: Notes 23 A world full of Plio-pleistocene hominins Copyright Bruce Owen 2011 Let s look at the next chunk of
 Introduction to Biological Anthropology: Notes 23 A world full of Plio-pleistocene hominins Copyright Bruce Owen 2011 Let s look at the next chunk of time: 3.0 1.0 mya often called the Plio-pleistocene
Introduction to Biological Anthropology: Notes 23 A world full of Plio-pleistocene hominins Copyright Bruce Owen 2011 Let s look at the next chunk of time: 3.0 1.0 mya often called the Plio-pleistocene
Main Points. 2) The Great American Interchange -- dispersal versus vicariance -- example: recent range expansion of nine-banded armadillos
 The Great American Interchange -- dispersal versus vicariance -- example: recent range expansion of nine-banded armadillos") Main Points 1) Mammalian Characteristics: Diversity, Phylogeny, and Systematics: -- Infraclass Eutheria -- Orders Scandentia through Cetacea 2) The Great American Interchange -- dispersal versus vicariance
Main Points 1) Mammalian Characteristics: Diversity, Phylogeny, and Systematics: -- Infraclass Eutheria -- Orders Scandentia through Cetacea 2) The Great American Interchange -- dispersal versus vicariance
Minnesota_mammals_Info_9.doc 11/04/09 -- DRAFT Page 1 of 64. Minnesota mammals
 Minnesota_mammals_Info_9.doc 11/04/09 -- DRAFT Page 1 of 64 Minnesota mammals This is a short guide to Minnesota mammals, with information drawn from Hazard s Mammals of, Walker s Mammals of the World,
Minnesota_mammals_Info_9.doc 11/04/09 -- DRAFT Page 1 of 64 Minnesota mammals This is a short guide to Minnesota mammals, with information drawn from Hazard s Mammals of, Walker s Mammals of the World,
Skulls & Evolution. 14,000 ya cro-magnon. 300,000 ya Homo sapiens. 2 Ma Homo habilis A. boisei A. robustus A. africanus
 Skulls & Evolution Purpose To illustrate trends in the evolution of humans. To demonstrate what you can learn from bones & fossils. To show the adaptations of various mammals to different habitats and
Skulls & Evolution Purpose To illustrate trends in the evolution of humans. To demonstrate what you can learn from bones & fossils. To show the adaptations of various mammals to different habitats and
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission.
 Bite Club: Comparative Bite Force in Big Biting Mammals and the Prediction of Predatory Behaviour in Fossil Taxa Author(s): Stephen Wroe, Colin McHenry, Jeffrey Thomason Source: Proceedings: Biological
Bite Club: Comparative Bite Force in Big Biting Mammals and the Prediction of Predatory Behaviour in Fossil Taxa Author(s): Stephen Wroe, Colin McHenry, Jeffrey Thomason Source: Proceedings: Biological
ANTHR 1L Biological Anthropology Lab
 ANTHR 1L Biological Anthropology Lab Name: DEFINING THE ORDER PRIMATES Humans belong to the zoological Order Primates, which is one of the 18 Orders of the Class Mammalia. Today we will review some of
ANTHR 1L Biological Anthropology Lab Name: DEFINING THE ORDER PRIMATES Humans belong to the zoological Order Primates, which is one of the 18 Orders of the Class Mammalia. Today we will review some of
LATE WINTER DIETARY OVERLAP AMONG GREATER RHEAS AND DOMESTIC HERBIVORES ON THE ARGENTINEAN FLOODING PAMPA
 LATE WINTER DIETARY OVERLAP AMONG GREATER RHEAS AND ID # 22-18 DOMESTIC HERBIVORES ON THE ARGENTINEAN FLOODING PAMPA G. Vacarezza 1, M.S. Cid 2,3, and F. Milano 1 1 Fac. Cs. Vet. (FCV), Univ. Nac. del
LATE WINTER DIETARY OVERLAP AMONG GREATER RHEAS AND ID # 22-18 DOMESTIC HERBIVORES ON THE ARGENTINEAN FLOODING PAMPA G. Vacarezza 1, M.S. Cid 2,3, and F. Milano 1 1 Fac. Cs. Vet. (FCV), Univ. Nac. del
Modern Evolutionary Classification. Lesson Overview. Lesson Overview Modern Evolutionary Classification
 Lesson Overview 18.2 Modern Evolutionary Classification THINK ABOUT IT Darwin s ideas about a tree of life suggested a new way to classify organisms not just based on similarities and differences, but
Lesson Overview 18.2 Modern Evolutionary Classification THINK ABOUT IT Darwin s ideas about a tree of life suggested a new way to classify organisms not just based on similarities and differences, but
Lab 8 Order Carnivora: Families Canidae, Felidae, and Ursidae Need to know Terms: carnassials, digitigrade, reproductive suppression, Jacobson s organ
 Lab 8 Order Carnivora: Families Canidae, Felidae, and Ursidae Need to know Terms: carnassials, digitigrade, reproductive suppression, Jacobson s organ Family Canidae Canis latrans ID based on skull, photos,
Lab 8 Order Carnivora: Families Canidae, Felidae, and Ursidae Need to know Terms: carnassials, digitigrade, reproductive suppression, Jacobson s organ Family Canidae Canis latrans ID based on skull, photos,
Main Points. 2) The Great American Interchange -- dispersal versus vicariance -- example: recent range expansion of nine-banded armadillos
 The Great American Interchange -- dispersal versus vicariance -- example: recent range expansion of nine-banded armadillos") Main Points 1) Diversity, Phylogeny, and Systematics -- Infraclass Eutheria -- Orders Scandentia through Cetacea 2) The Great American Interchange -- dispersal versus vicariance -- example: recent range
Main Points 1) Diversity, Phylogeny, and Systematics -- Infraclass Eutheria -- Orders Scandentia through Cetacea 2) The Great American Interchange -- dispersal versus vicariance -- example: recent range
Mastozoología Neotropical ISSN: Sociedad Argentina para el Estudio de los Mamíferos. Argentina
 Mastozoología Neotropical ISSN: 0327-9383 ulyses@cenpat.edu.ar Sociedad Argentina para el Estudio de los Mamíferos Argentina Schleich, Cristian E. EFFECT OF AGE AND TEMPERATURE ON THE VOCAL BEHAVIOR OF
Mastozoología Neotropical ISSN: 0327-9383 ulyses@cenpat.edu.ar Sociedad Argentina para el Estudio de los Mamíferos Argentina Schleich, Cristian E. EFFECT OF AGE AND TEMPERATURE ON THE VOCAL BEHAVIOR OF
Anatomy. Name Section. The Vertebrate Skeleton
 Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Name Section Anatomy The Vertebrate Skeleton Vertebrate paleontologists get most of their knowledge about past organisms from skeletal remains. Skeletons are useful for gleaning information about an organism
Sample Questions: EXAMINATION I Form A Mammalogy -EEOB 625. Name Composite of previous Examinations
 Sample Questions: EXAMINATION I Form A Mammalogy -EEOB 625 Name Composite of previous Examinations Part I. Define or describe only 5 of the following 6 words - 15 points (3 each). If you define all 6,
Sample Questions: EXAMINATION I Form A Mammalogy -EEOB 625 Name Composite of previous Examinations Part I. Define or describe only 5 of the following 6 words - 15 points (3 each). If you define all 6,
VERTEBRATE READING. Fishes
 VERTEBRATE READING Fishes The first vertebrates to become a widespread, predominant life form on earth were fishes. Prior to this, only invertebrates, such as mollusks, worms and squid-like animals, would
VERTEBRATE READING Fishes The first vertebrates to become a widespread, predominant life form on earth were fishes. Prior to this, only invertebrates, such as mollusks, worms and squid-like animals, would
What are taxonomy, classification, and systematics?
 Topic 2: Comparative Method o Taxonomy, classification, systematics o Importance of phylogenies o A closer look at systematics o Some key concepts o Parts of a cladogram o Groups and characters o Homology
Topic 2: Comparative Method o Taxonomy, classification, systematics o Importance of phylogenies o A closer look at systematics o Some key concepts o Parts of a cladogram o Groups and characters o Homology
Species: Panthera pardus Genus: Panthera Family: Felidae Order: Carnivora Class: Mammalia Phylum: Chordata
 CHAPTER 6: PHYLOGENY AND THE TREE OF LIFE AP Biology 3 PHYLOGENY AND SYSTEMATICS Phylogeny - evolutionary history of a species or group of related species Systematics - analytical approach to understanding
CHAPTER 6: PHYLOGENY AND THE TREE OF LIFE AP Biology 3 PHYLOGENY AND SYSTEMATICS Phylogeny - evolutionary history of a species or group of related species Systematics - analytical approach to understanding
Marsupial Mole. Notoryctes species. Amy Mutton Zoologist Species and Communities Branch Science and Conservation Division
 Marsupial Mole Notoryctes species Amy Mutton Zoologist Species and Communities Branch Science and Conservation Division Scientific classification Kingdom: Phylum: Class: Infraclass: Order: Family: Animalia
Marsupial Mole Notoryctes species Amy Mutton Zoologist Species and Communities Branch Science and Conservation Division Scientific classification Kingdom: Phylum: Class: Infraclass: Order: Family: Animalia
Mammalogy Lecture 8 - Evolution of Ear Ossicles
 Mammalogy Lecture 8 - Evolution of Ear Ossicles I. To begin, let s examine briefly the end point, that is, modern mammalian ears. Inner Ear The cochlea contains sensory cells for hearing and balance. -
Mammalogy Lecture 8 - Evolution of Ear Ossicles I. To begin, let s examine briefly the end point, that is, modern mammalian ears. Inner Ear The cochlea contains sensory cells for hearing and balance. -
Are the dinosauromorph femora from the Upper Triassic of Hayden Quarry (New Mexico) three stages in a growth series of a single taxon?
 three stages in a growth series of a single taxon?") Anais da Academia Brasileira de Ciências (2017) 89(2): 835-839 (Annals of the Brazilian Academy of Sciences) Printed version ISSN 0001-3765 / Online version ISSN 1678-2690 http://dx.doi.org/10.1590/0001-3765201720160583
Anais da Academia Brasileira de Ciências (2017) 89(2): 835-839 (Annals of the Brazilian Academy of Sciences) Printed version ISSN 0001-3765 / Online version ISSN 1678-2690 http://dx.doi.org/10.1590/0001-3765201720160583
PEABODY MUSEUM OF NATURAL HISTORY, YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 85 September 21, 1964 A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA STANLEY J. RIEL
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 85 September 21, 1964 A NEW OREODONT FROM THE CABBAGE PATCH LOCAL FAUNA, WESTERN MONTANA STANLEY J. RIEL
Main Points. 2) The Great American Interchange -- dispersal versus vicariance -- example: recent range expansion of nine-banded armadillos
 The Great American Interchange -- dispersal versus vicariance -- example: recent range expansion of nine-banded armadillos") Main Points 1) Diversity, Phylogeny, and Systematics -- Infraclass Metatheria continued -- Orders Diprotodontia and Peramelina -- Infraclass Eutheria -- Orders Lagomorpha through Cetacea 2) The Great American
Main Points 1) Diversity, Phylogeny, and Systematics -- Infraclass Metatheria continued -- Orders Diprotodontia and Peramelina -- Infraclass Eutheria -- Orders Lagomorpha through Cetacea 2) The Great American
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A.
 Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
Postilla PEABODY MUSEUM OF NATURAL HISTORY YALE UNIVERSITY NEW HAVEN, CONNECTICUT, U.S.A. Number 117 18 March 1968 A 7DIAPSID (REPTILIA) PARIETAL FROM THE LOWER PERMIAN OF OKLAHOMA ROBERT L. CARROLL REDPATH
1 Describe the anatomy and function of the turtle shell. 2 Describe respiration in turtles. How does the shell affect respiration?
 GVZ 2017 Practice Questions Set 1 Test 3 1 Describe the anatomy and function of the turtle shell. 2 Describe respiration in turtles. How does the shell affect respiration? 3 According to the most recent
GVZ 2017 Practice Questions Set 1 Test 3 1 Describe the anatomy and function of the turtle shell. 2 Describe respiration in turtles. How does the shell affect respiration? 3 According to the most recent
complex in cusp pattern. (3) The bones of the coyote skull are thinner, crests sharper and the
 The bones of the coyote skull are thinner, crests sharper and the") DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
DISTINCTIONS BETWEEN THE SKULLS OF S AND DOGS Grover S. Krantz Archaeological sites in the United States frequently yield the bones of coyotes and domestic dogs. These two canines are very similar both
Geoffroy s Cat: Biodiversity Research Project
 Geoffroy s Cat: Biodiversity Research Project Viet Nguyen Conservation Biology BES 485 Geoffroy s Cat Geoffroy s Cat (Leopardus geoffroyi) are small, little known spotted wild cat found native to the central
Geoffroy s Cat: Biodiversity Research Project Viet Nguyen Conservation Biology BES 485 Geoffroy s Cat Geoffroy s Cat (Leopardus geoffroyi) are small, little known spotted wild cat found native to the central
Phylogeny Reconstruction
 Phylogeny Reconstruction Trees, Methods and Characters Reading: Gregory, 2008. Understanding Evolutionary Trees (Polly, 2006) Lab tomorrow Meet in Geology GY522 Bring computers if you have them (they will
Phylogeny Reconstruction Trees, Methods and Characters Reading: Gregory, 2008. Understanding Evolutionary Trees (Polly, 2006) Lab tomorrow Meet in Geology GY522 Bring computers if you have them (they will
New York State Mammals. Morphology Ecology Identification Classification Distribution
 New York State Mammals Morphology Ecology Identification Classification Distribution ORDER: Didelphimorphia FAMILY: Didelphidae Common Name: Virginia opossum Scientific Name: (Didelphis virginiana) Marsupial
New York State Mammals Morphology Ecology Identification Classification Distribution ORDER: Didelphimorphia FAMILY: Didelphidae Common Name: Virginia opossum Scientific Name: (Didelphis virginiana) Marsupial
Title: Fossil Focus: Reimagining fossil cats IMPORTANT COPYRIGHT CITATION OF ARTICLE
 Title: Fossil Focus: Reimagining fossil cats Author(s): Andrew Cuff Volume: 8 Article: 4 Page(s): 1-10 Published Date: 01/04/2018 PermaLink: https://www.palaeontologyonline.com/articles/2018/patterns-palaeontology-earliestskeletons/
Title: Fossil Focus: Reimagining fossil cats Author(s): Andrew Cuff Volume: 8 Article: 4 Page(s): 1-10 Published Date: 01/04/2018 PermaLink: https://www.palaeontologyonline.com/articles/2018/patterns-palaeontology-earliestskeletons/
Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes
 Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Supplementary Information Exceptional fossil preservation demonstrates a new mode of axial skeleton elongation in early ray-finned fishes Erin E. Maxwell, Heinz Furrer, Marcelo R. Sánchez-Villagra Supplementary
Comparative Zoology Portfolio Project Assignment
 Comparative Zoology Portfolio Project Assignment Using your knowledge from the in class activities, your notes, you Integrated Science text, or the internet, you will look at the major trends in the evolution
Comparative Zoology Portfolio Project Assignment Using your knowledge from the in class activities, your notes, you Integrated Science text, or the internet, you will look at the major trends in the evolution
SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE
 PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
PROCEEDINGS OF THE UNITED STATES NATIONAL MUSEUM issued SWsK \ {^^m ^V ^^ SMITHSONIAN INSTITUTION U. S. NATIONAL MUSEUM Vol. 91 Washington : 1941 No. 3124 SOME LITTLE-KNOWN FOSSIL LIZARDS FROM THE OLIGOCENE
Do the traits of organisms provide evidence for evolution?
 PhyloStrat Tutorial Do the traits of organisms provide evidence for evolution? Consider two hypotheses about where Earth s organisms came from. The first hypothesis is from John Ray, an influential British
PhyloStrat Tutorial Do the traits of organisms provide evidence for evolution? Consider two hypotheses about where Earth s organisms came from. The first hypothesis is from John Ray, an influential British
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1
 Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
Geo 302D: Age of Dinosaurs LAB 4: Systematics Part 1 Systematics is the comparative study of biological diversity with the intent of determining the relationships between organisms. Humankind has always
UNIT III A. Descent with Modification(Ch19) B. Phylogeny (Ch20) C. Evolution of Populations (Ch21) D. Origin of Species or Speciation (Ch22)
 B. Phylogeny (Ch20) C. Evolution of Populations (Ch21) D. Origin of Species or Speciation (Ch22)") UNIT III A. Descent with Modification(Ch9) B. Phylogeny (Ch2) C. Evolution of Populations (Ch2) D. Origin of Species or Speciation (Ch22) Classification in broad term simply means putting things in classes
UNIT III A. Descent with Modification(Ch9) B. Phylogeny (Ch2) C. Evolution of Populations (Ch2) D. Origin of Species or Speciation (Ch22) Classification in broad term simply means putting things in classes
Wild boar (Sus scrofa ferus): productivity index in an experimental outdoor farm
: productivity index in an experimental outdoor farm") COMUNICACIÓN CORTA Wild boar (Sus scrofa ferus): productivity index in an experimental outdoor farm VIEITES, C.M. 1 ; BASSO, C.P. 1 and BARTOLONI, N. 2 ABSTRACT The European wild boar (Sus scrofa ferus)
COMUNICACIÓN CORTA Wild boar (Sus scrofa ferus): productivity index in an experimental outdoor farm VIEITES, C.M. 1 ; BASSO, C.P. 1 and BARTOLONI, N. 2 ABSTRACT The European wild boar (Sus scrofa ferus)
SENSITIZATION FOR THE AUTOCHTHONOUS BREEDS CONSERVATION VIA THE PUBLIC SHOWS OF ANIMALS
 SENSITIZATION FOR THE AUTOCHTHONOUS BREEDS CONSERVATION VIA THE PUBLIC SHOWS OF ANIMALS SENSIBILIZACION DE LA OPINION PUBLICA POR LA CONSERVACION DE RAZAS AUTOCTONAS A TRAVES DE LAS EXPOSICIONES DE ANIMALES
SENSITIZATION FOR THE AUTOCHTHONOUS BREEDS CONSERVATION VIA THE PUBLIC SHOWS OF ANIMALS SENSIBILIZACION DE LA OPINION PUBLICA POR LA CONSERVACION DE RAZAS AUTOCTONAS A TRAVES DE LAS EXPOSICIONES DE ANIMALES
Phylogenetics. Phylogenetic Trees. 1. Represent presumed patterns. 2. Analogous to family trees.
 Phylogenetics. Phylogenetic Trees. 1. Represent presumed patterns of descent. 2. Analogous to family trees. 3. Resolve taxa, e.g., species, into clades each of which includes an ancestral taxon and all
Phylogenetics. Phylogenetic Trees. 1. Represent presumed patterns of descent. 2. Analogous to family trees. 3. Resolve taxa, e.g., species, into clades each of which includes an ancestral taxon and all
6. The lifetime Darwinian fitness of one organism is greater than that of another organism if: A. it lives longer than the other B. it is able to outc
 1. The money in the kingdom of Florin consists of bills with the value written on the front, and pictures of members of the royal family on the back. To test the hypothesis that all of the Florinese $5
1. The money in the kingdom of Florin consists of bills with the value written on the front, and pictures of members of the royal family on the back. To test the hypothesis that all of the Florinese $5
Optimizing Phylogenetic Supertrees Using Answer Set Programming
 1 Online appendix for the paper Optimizing Phylogenetic Supertrees Using Answer Set Programming published in Theory and Practice of Logic Programming LAURA KOPONEN and EMILIA OIKARINEN and TOMI JANHUNEN
1 Online appendix for the paper Optimizing Phylogenetic Supertrees Using Answer Set Programming published in Theory and Practice of Logic Programming LAURA KOPONEN and EMILIA OIKARINEN and TOMI JANHUNEN
Fig Phylogeny & Systematics
 Fig. 26- Phylogeny & Systematics Tree of Life phylogenetic relationship for 3 clades (http://evolution.berkeley.edu Fig. 26-2 Phylogenetic tree Figure 26.3 Taxonomy Taxon Carolus Linnaeus Species: Panthera
Fig. 26- Phylogeny & Systematics Tree of Life phylogenetic relationship for 3 clades (http://evolution.berkeley.edu Fig. 26-2 Phylogenetic tree Figure 26.3 Taxonomy Taxon Carolus Linnaeus Species: Panthera
Dipsas trinitatis (Trinidad Snail-eating Snake)
") Dipsas trinitatis (Trinidad Snail-eating Snake) Family: Dipsadidae (Rear-fanged Snakes) Order: Squamata (Lizards and Snakes) Class: Reptilia (Reptiles) Fig. 1. Trinidad snail-eating snake, Dipsas trinitatis.
Dipsas trinitatis (Trinidad Snail-eating Snake) Family: Dipsadidae (Rear-fanged Snakes) Order: Squamata (Lizards and Snakes) Class: Reptilia (Reptiles) Fig. 1. Trinidad snail-eating snake, Dipsas trinitatis.
THE MONSTER OF TROY VASE IS NOT BASED ON A FOSSIL GIRAFFE. (Short title: MONSTER OF TROY VASE IS NOT A GIRAFFE)
") THE MONSTER OF TROY VASE IS NOT BASED ON A FOSSIL GIRAFFE (Short title: MONSTER OF TROY VASE IS NOT A GIRAFFE) Summary. It has been proposed that the Monster of Troy, depicted in a 6th Century BC Corinthian
THE MONSTER OF TROY VASE IS NOT BASED ON A FOSSIL GIRAFFE (Short title: MONSTER OF TROY VASE IS NOT A GIRAFFE) Summary. It has been proposed that the Monster of Troy, depicted in a 6th Century BC Corinthian
Mystery of Life Travelling Exhibition Vertebrate Kingdom
 Mystery of Life Travelling Exhibition Vertebrate Kingdom When science meets art, what will happen? Vertebrate exhibition, it s a perfect convergence of the technique and art, where you can learn not only
Mystery of Life Travelling Exhibition Vertebrate Kingdom When science meets art, what will happen? Vertebrate exhibition, it s a perfect convergence of the technique and art, where you can learn not only
Supporting Information
 Supporting Information Table S1. Sources of the historic range maps used in our analysis. Elevation limits (lower and upper) are in meters. Modifications to the source maps are listed in the footnotes.
Supporting Information Table S1. Sources of the historic range maps used in our analysis. Elevation limits (lower and upper) are in meters. Modifications to the source maps are listed in the footnotes.
d. Wrist bones. Pacific salmon life cycle. Atlantic salmon (different genus) can spawn more than once.
 can spawn more than once.") Lecture III.5b Answers to HW 1. (2 pts). Tiktaalik bridges the gap between fish and tetrapods by virtue of possessing which of the following? a. Humerus. b. Radius. c. Ulna. d. Wrist bones. 2. (2 pts)
Lecture III.5b Answers to HW 1. (2 pts). Tiktaalik bridges the gap between fish and tetrapods by virtue of possessing which of the following? a. Humerus. b. Radius. c. Ulna. d. Wrist bones. 2. (2 pts)
Panther Habitat. Welcome to the. Who Are Florida Panthers? Panther Classification
 Welcome to the Panther Habitat Panther Classification Class: Mammalia Order: Carnivora Family: Felidae Genus: Puma Species: Concolor Subspecies (Southern U.S): P.c. coryi Who Are Florida Panthers? The
Welcome to the Panther Habitat Panther Classification Class: Mammalia Order: Carnivora Family: Felidae Genus: Puma Species: Concolor Subspecies (Southern U.S): P.c. coryi Who Are Florida Panthers? The
Animal Diversity wrap-up Lecture 9 Winter 2014
 Animal Diversity wrap-up Lecture 9 Winter 2014 1 Animal phylogeny based on morphology & development Fig. 32.10 2 Animal phylogeny based on molecular data Fig. 32.11 New Clades 3 Lophotrochozoa Lophophore:
Animal Diversity wrap-up Lecture 9 Winter 2014 1 Animal phylogeny based on morphology & development Fig. 32.10 2 Animal phylogeny based on molecular data Fig. 32.11 New Clades 3 Lophotrochozoa Lophophore:
Spot the Difference: Using the domestic cat as a model for the nutritional management of captive cheetahs. Katherine M. Bell
 Spot the Difference: Using the domestic cat as a model for the nutritional management of captive cheetahs Katherine M. Bell Edited by Lucy A. Tucker and David G. Thomas Illustrated by Justine Woosnam and
Spot the Difference: Using the domestic cat as a model for the nutritional management of captive cheetahs Katherine M. Bell Edited by Lucy A. Tucker and David G. Thomas Illustrated by Justine Woosnam and
Morphological Variation in Anolis oculatus Between Dominican. Habitats
 Morphological Variation in Anolis oculatus Between Dominican Habitats Lori Valentine Texas A&M University Dr. Lacher Dr. Woolley Study Abroad Dominica 2002 Morphological Variation in Anolis oculatus Between
Morphological Variation in Anolis oculatus Between Dominican Habitats Lori Valentine Texas A&M University Dr. Lacher Dr. Woolley Study Abroad Dominica 2002 Morphological Variation in Anolis oculatus Between
Mammalogy Lecture 4A Metatherian Diversity
 Mammalogy Lecture 4A Metatherian Diversity I. Therians. Remember that metatherians and eutherians (i.e., marsupial and placental mammals) form a clade. II. Metatherians Marsupials are a monophyletic group.
Mammalogy Lecture 4A Metatherian Diversity I. Therians. Remember that metatherians and eutherians (i.e., marsupial and placental mammals) form a clade. II. Metatherians Marsupials are a monophyletic group.
Growth of feral pigs in New Zealand
 Acta Theriologica 35 (1-2): 77-88, 1990. PL ISSN 0001-7051 Growth of feral pigs in New Zealand Ryszard M. DZI^CIOLOWSKI, Colin M. H. CLARKE and Brian J. FREDRIC Dzi?ciolowski R. M., Clarke C. M. H. and
Acta Theriologica 35 (1-2): 77-88, 1990. PL ISSN 0001-7051 Growth of feral pigs in New Zealand Ryszard M. DZI^CIOLOWSKI, Colin M. H. CLARKE and Brian J. FREDRIC Dzi?ciolowski R. M., Clarke C. M. H. and
NOTES ON THE ECOLOGY AND NATURAL HISTORY OF TWO SPECIES OF EGERNIA (SCINCIDAE) IN WESTERN AUSTRALIA
 IN WESTERN AUSTRALIA") NOTES ON THE ECOLOGY AND NATURAL HISTORY OF TWO SPECIES OF EGERNIA (SCINCIDAE) IN WESTERN AUSTRALIA By ERIC R. PIANKA Integrative Biology University of Texas at Austin Austin, Texas 78712 USA Email: erp@austin.utexas.edu
NOTES ON THE ECOLOGY AND NATURAL HISTORY OF TWO SPECIES OF EGERNIA (SCINCIDAE) IN WESTERN AUSTRALIA By ERIC R. PIANKA Integrative Biology University of Texas at Austin Austin, Texas 78712 USA Email: erp@austin.utexas.edu
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * Dr. L.D. Boonstra. Paleontologist, South African Museum, Cape Town
 THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
THE GORGONOPSIAN GENUS, HIPPOSAURUS, AND THE FAMILY ICTIDORHINIDAE * by Dr. L.D. Boonstra Paleontologist, South African Museum, Cape Town In 1928 I dug up the complete skeleton of a smallish gorgonopsian
Adjustment Factors in NSIP 1
 Adjustment Factors in NSIP 1 David Notter and Daniel Brown Summary Multiplicative adjustment factors for effects of type of birth and rearing on weaning and postweaning lamb weights were systematically
Adjustment Factors in NSIP 1 David Notter and Daniel Brown Summary Multiplicative adjustment factors for effects of type of birth and rearing on weaning and postweaning lamb weights were systematically
Chapter 16: Evolution Lizard Evolution Virtual Lab Honors Biology. Name: Block: Introduction
 Chapter 16: Evolution Lizard Evolution Virtual Lab Honors Biology Name: Block: Introduction Charles Darwin proposed that over many generations some members of a population could adapt to a changing environment
Chapter 16: Evolution Lizard Evolution Virtual Lab Honors Biology Name: Block: Introduction Charles Darwin proposed that over many generations some members of a population could adapt to a changing environment
Cladistics (reading and making of cladograms)
") Cladistics (reading and making of cladograms) Definitions Systematics The branch of biological sciences concerned with classifying organisms Taxon (pl: taxa) Any unit of biological diversity (eg. Animalia,
Cladistics (reading and making of cladograms) Definitions Systematics The branch of biological sciences concerned with classifying organisms Taxon (pl: taxa) Any unit of biological diversity (eg. Animalia,
Owl Pellet Dissection A Study of Food Chains & Food Webs
 NAME Owl Pellet Dissection A Study of Food Chains & Food Webs INTRODUCTION: Owl pellets are masses of bone, teeth, hair, feathers and exoskeletons of various animals preyed upon by raptors, or birds of
NAME Owl Pellet Dissection A Study of Food Chains & Food Webs INTRODUCTION: Owl pellets are masses of bone, teeth, hair, feathers and exoskeletons of various animals preyed upon by raptors, or birds of
Mammalogy Laboratory 1 - Mammalian Anatomy
 Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
Mammalogy Laboratory 1 - Mammalian Anatomy I. The Goal. The goal of the lab is to teach you skeletal anatomy of mammals. We will emphasize the skull because many of the taxonomically important characters
Slide 1. Birds & Mammals. Chapter 15
 Slide 1 Birds & Mammals Chapter 15 Slide 2 What is a Bird? Vertebrate Endothermic Feathered 4 chambered heart Egg laying Fore-limbs adapted for flight Bones nearly hollow (allow for lighter weight) Slide
Slide 1 Birds & Mammals Chapter 15 Slide 2 What is a Bird? Vertebrate Endothermic Feathered 4 chambered heart Egg laying Fore-limbs adapted for flight Bones nearly hollow (allow for lighter weight) Slide
Ch. 17: Classification
 Ch. 17: Classification Who is Carolus Linnaeus? Linnaeus developed the scientific naming system still used today. Taxonomy What is? the science of naming and classifying organisms. A taxon group of organisms
Ch. 17: Classification Who is Carolus Linnaeus? Linnaeus developed the scientific naming system still used today. Taxonomy What is? the science of naming and classifying organisms. A taxon group of organisms
MAURICIO ANTÓN 1 *, MANUEL J. SALESA 1, JUAN FRANCISCO PASTOR 2, ISRAEL M. SÁNCHEZ 1, SUSANA FRAILE 1 and JORGE MORALES 1 INTRODUCTION
 Lin- Blackwell Science, LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082The nean Society of London, 2004? 2004 140? 207221 Original Article M. ANTÓN ET AL. MASTOID ANATOMY IN FELIDS Zoological
Lin- Blackwell Science, LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082The nean Society of London, 2004? 2004 140? 207221 Original Article M. ANTÓN ET AL. MASTOID ANATOMY IN FELIDS Zoological
Food Item Use by Coyote Pups at Crab Orchard National Wildlife Refuge, Illinois
 Transactions of the Illinois State Academy of Science (1993), Volume 86, 3 and 4, pp. 133-137 Food Item Use by Coyote Pups at Crab Orchard National Wildlife Refuge, Illinois Brian L. Cypher 1 Cooperative
Transactions of the Illinois State Academy of Science (1993), Volume 86, 3 and 4, pp. 133-137 Food Item Use by Coyote Pups at Crab Orchard National Wildlife Refuge, Illinois Brian L. Cypher 1 Cooperative
Question Set 1: Animal EVOLUTIONARY BIODIVERSITY
 Biology 162 LAB EXAM 2, AM Version Thursday 24 April 2003 page 1 Question Set 1: Animal EVOLUTIONARY BIODIVERSITY (a). We have mentioned several times in class that the concepts of Developed and Evolved
Biology 162 LAB EXAM 2, AM Version Thursday 24 April 2003 page 1 Question Set 1: Animal EVOLUTIONARY BIODIVERSITY (a). We have mentioned several times in class that the concepts of Developed and Evolved
Overall structure is similar to humans, but again there are differences. Some features that are unique to mammals: Found in eutherian mammals.
 Mammalian anatomy and physiology (part II): Nervous system: Brain: Sensory input: Overall structure is similar to humans, but again there are differences. Some features that are unique to mammals: Smell:
Mammalian anatomy and physiology (part II): Nervous system: Brain: Sensory input: Overall structure is similar to humans, but again there are differences. Some features that are unique to mammals: Smell:
Statistical support for the hypothesis of developmental constraint in marsupial skull evolution
 Bennett and Goswami BMC Biology 2013, 11:52 RESEARCH ARTICLE Statistical support for the hypothesis of developmental constraint in marsupial skull evolution C Verity Bennett 1* and Anjali Goswami 1,2 Open
Bennett and Goswami BMC Biology 2013, 11:52 RESEARCH ARTICLE Statistical support for the hypothesis of developmental constraint in marsupial skull evolution C Verity Bennett 1* and Anjali Goswami 1,2 Open
New York State Mammals. Order Lagomorpha Order Rodentia
 New York State Mammals Order Lagomorpha Order Rodentia FAMILY: LEPORIDAE Rabbits and hares Conspicuous tail Fenestra appears as bony latticework Some species molt seasonally Presence of a second incisor
New York State Mammals Order Lagomorpha Order Rodentia FAMILY: LEPORIDAE Rabbits and hares Conspicuous tail Fenestra appears as bony latticework Some species molt seasonally Presence of a second incisor
SOAR Research Proposal Summer How do sand boas capture prey they can t see?
 SOAR Research Proposal Summer 2016 How do sand boas capture prey they can t see? Faculty Mentor: Dr. Frances Irish, Assistant Professor of Biological Sciences Project start date and duration: May 31, 2016
SOAR Research Proposal Summer 2016 How do sand boas capture prey they can t see? Faculty Mentor: Dr. Frances Irish, Assistant Professor of Biological Sciences Project start date and duration: May 31, 2016
Museu de História Natural do Funchal. Madeira 31.XII.2012 No. 235
 1 ISSN 0523-7904 B O C A G I A N A Museu de História Natural do Funchal Madeira 31.XII.2012 No. 235 FIRST DATA ON BREEDING OF MANDARIN DUCK AIX GALERICULATA IN THE MADEIRAN ARCHIPELAGO BY DOMINGO TRUJILLO
1 ISSN 0523-7904 B O C A G I A N A Museu de História Natural do Funchal Madeira 31.XII.2012 No. 235 FIRST DATA ON BREEDING OF MANDARIN DUCK AIX GALERICULATA IN THE MADEIRAN ARCHIPELAGO BY DOMINGO TRUJILLO
Animal Evolution The Chordates. Chapter 26 Part 2
 Animal Evolution The Chordates Chapter 26 Part 2 26.10 Birds The Feathered Ones Birds are the only animals with feathers Descendants of flying dinosaurs in which scales became modified as feathers Long
Animal Evolution The Chordates Chapter 26 Part 2 26.10 Birds The Feathered Ones Birds are the only animals with feathers Descendants of flying dinosaurs in which scales became modified as feathers Long
SHORT NOTE Preferential use of one paw during feeding in the subterranean rodent Ctenomys talarum Cristian E. Schleich
 Belg. J. Zool., 146 (2) : 134 139 July 2016 SHORT NOTE Preferential use of one paw during feeding in the subterranean rodent Ctenomys talarum Cristian E. Schleich Laboratorio de Ecofisiología, Instituto
Belg. J. Zool., 146 (2) : 134 139 July 2016 SHORT NOTE Preferential use of one paw during feeding in the subterranean rodent Ctenomys talarum Cristian E. Schleich Laboratorio de Ecofisiología, Instituto
8/19/2013. Topic 5: The Origin of Amniotes. What are some stem Amniotes? What are some stem Amniotes? The Amniotic Egg. What is an Amniote?
 Topic 5: The Origin of Amniotes Where do amniotes fall out on the vertebrate phylogeny? What are some stem Amniotes? What is an Amniote? What changes were involved with the transition to dry habitats?
Topic 5: The Origin of Amniotes Where do amniotes fall out on the vertebrate phylogeny? What are some stem Amniotes? What is an Amniote? What changes were involved with the transition to dry habitats?
Giant croc with T. rex teeth roamed Madagascar
 Giant croc with T. rex teeth roamed Madagascar www.scimex.org/newsfeed/giant-croc-with-t.-rex-teeth-used-to-roam-in-madagascar Embargoed until: Publicly released: PeerJ A fossil of the largest and oldest
Giant croc with T. rex teeth roamed Madagascar www.scimex.org/newsfeed/giant-croc-with-t.-rex-teeth-used-to-roam-in-madagascar Embargoed until: Publicly released: PeerJ A fossil of the largest and oldest
muscles (enhancing biting strength). Possible states: none, one, or two.
. Possible states: none, one, or two.") Reconstructing Evolutionary Relationships S-1 Practice Exercise: Phylogeny of Terrestrial Vertebrates In this example we will construct a phylogenetic hypothesis of the relationships between seven taxa
Reconstructing Evolutionary Relationships S-1 Practice Exercise: Phylogeny of Terrestrial Vertebrates In this example we will construct a phylogenetic hypothesis of the relationships between seven taxa
May 17, SWBAT explain why scientists classify organisms SWBAT list major levels of hierarchy
 May 17, 2017 Aims: SWBAT explain why scientists classify organisms SWBAT list major levels of hierarchy Agenda 1. Do Now 2. Class Notes 3. Guided Practice 4. Independent Practice 5. Practicing our AIMS:
May 17, 2017 Aims: SWBAT explain why scientists classify organisms SWBAT list major levels of hierarchy Agenda 1. Do Now 2. Class Notes 3. Guided Practice 4. Independent Practice 5. Practicing our AIMS:
Williston, and as there are many fairly good specimens in the American
 56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
56.81.7D :14.71.5 Article VII.- SOME POINTS IN THE STRUCTURE OF THE DIADECTID SKULL. BY R. BROOM. The skull of Diadectes has been described by Cope, Case, v. Huene, and Williston, and as there are many
Josefina de Combellas, N Martinez and E Gonzalez. Instituto de Producción Animal, Facultad de Agronomia, Universidad Central de Venezuela, Maracay
 Trop Anim Prod 1980 5:3 261 A STUDY OF FACTORS WHICH INFLUENCE BIRTH AND WEANING WEIGHT IN LAMBS Josefina de Combellas, N Martinez and E Gonzalez Instituto de Producción Animal, Facultad de Agronomia,
Trop Anim Prod 1980 5:3 261 A STUDY OF FACTORS WHICH INFLUENCE BIRTH AND WEANING WEIGHT IN LAMBS Josefina de Combellas, N Martinez and E Gonzalez Instituto de Producción Animal, Facultad de Agronomia,
Age estimation in Iberian wildcats Felis by canine tooth sections
 Acta Theriologica 44 (3): 321-327, 1999. PL ISSN 0001-7051 Age estimation in Iberian wildcats Felis by canine tooth sections silvestris, Rosa GARCÍA-PEREA and Rocío A. BAQUERO Garcia-Perea R. and Baquero
Acta Theriologica 44 (3): 321-327, 1999. PL ISSN 0001-7051 Age estimation in Iberian wildcats Felis by canine tooth sections silvestris, Rosa GARCÍA-PEREA and Rocío A. BAQUERO Garcia-Perea R. and Baquero
INQUIRY & INVESTIGATION
 INQUIRY & INVESTIGTION Phylogenies & Tree-Thinking D VID. UM SUSN OFFNER character a trait or feature that varies among a set of taxa (e.g., hair color) character-state a variant of a character that occurs
INQUIRY & INVESTIGTION Phylogenies & Tree-Thinking D VID. UM SUSN OFFNER character a trait or feature that varies among a set of taxa (e.g., hair color) character-state a variant of a character that occurs
May 10, SWBAT analyze and evaluate the scientific evidence provided by the fossil record.
 May 10, 2017 Aims: SWBAT analyze and evaluate the scientific evidence provided by the fossil record. Agenda 1. Do Now 2. Class Notes 3. Guided Practice 4. Independent Practice 5. Practicing our AIMS: E.3-Examining
May 10, 2017 Aims: SWBAT analyze and evaluate the scientific evidence provided by the fossil record. Agenda 1. Do Now 2. Class Notes 3. Guided Practice 4. Independent Practice 5. Practicing our AIMS: E.3-Examining
Modern taxonomy. Building family trees 10/10/2011. Knowing a lot about lots of creatures. Tom Hartman. Systematics includes: 1.
 Modern taxonomy Building family trees Tom Hartman www.tuatara9.co.uk Classification has moved away from the simple grouping of organisms according to their similarities (phenetics) and has become the study
Modern taxonomy Building family trees Tom Hartman www.tuatara9.co.uk Classification has moved away from the simple grouping of organisms according to their similarities (phenetics) and has become the study
Bio 1B Lecture Outline (please print and bring along) Fall, 2006
 Fall, 2006") Bio 1B Lecture Outline (please print and bring along) Fall, 2006 B.D. Mishler, Dept. of Integrative Biology 2-6810, bmishler@berkeley.edu Evolution lecture #4 -- Phylogenetic Analysis (Cladistics) -- Oct.
Bio 1B Lecture Outline (please print and bring along) Fall, 2006 B.D. Mishler, Dept. of Integrative Biology 2-6810, bmishler@berkeley.edu Evolution lecture #4 -- Phylogenetic Analysis (Cladistics) -- Oct.
Mastozoología Neotropical ISSN: Sociedad Argentina para el Estudio de los Mamíferos. Argentina
 Mastozoología Neotropical ISSN: 0327-9383 ulyses@cenpat.edu.ar Sociedad Argentina para el Estudio de los Mamíferos Argentina Lucherini, Mauro; Soler, Lucía; Luengos Vidal, Estela A preliminary revision
Mastozoología Neotropical ISSN: 0327-9383 ulyses@cenpat.edu.ar Sociedad Argentina para el Estudio de los Mamíferos Argentina Lucherini, Mauro; Soler, Lucía; Luengos Vidal, Estela A preliminary revision
SKELETONS: Museum of Osteology Tooth and Eye Dentification Teacher Resource
 SKELETONS: Museum of Osteology Tooth and Eye Dentification Teacher Resource Grade Levels: 3 rd 5 th Grade 3 rd Grade: SC.3.N.1.1 - Raise questions about the natural world, investigate them individually
SKELETONS: Museum of Osteology Tooth and Eye Dentification Teacher Resource Grade Levels: 3 rd 5 th Grade 3 rd Grade: SC.3.N.1.1 - Raise questions about the natural world, investigate them individually
University of Canberra. This thesis is available in print format from the University of Canberra Library.
 University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
University of Canberra This thesis is available in print format from the University of Canberra Library. If you are the author of this thesis and wish to have the whole thesis loaded here, please contact
Gulf and Caribbean Research
 Gulf and Caribbean Research Volume 16 Issue 1 January 4 Morphological Characteristics of the Carapace of the Hawksbill Turtle, Eretmochelys imbricata, from n Waters Mari Kobayashi Hokkaido University DOI:
Gulf and Caribbean Research Volume 16 Issue 1 January 4 Morphological Characteristics of the Carapace of the Hawksbill Turtle, Eretmochelys imbricata, from n Waters Mari Kobayashi Hokkaido University DOI:
8/19/2013. What is convergence? Topic 11: Convergence. What is convergence? What is convergence? What is convergence? What is convergence?
 Topic 11: Convergence What are the classic herp examples? Have they been formally studied? Emerald Tree Boas and Green Tree Pythons show a remarkable level of convergence Photos KP Bergmann, Philadelphia
Topic 11: Convergence What are the classic herp examples? Have they been formally studied? Emerald Tree Boas and Green Tree Pythons show a remarkable level of convergence Photos KP Bergmann, Philadelphia
v:ii-ixi, 'i':;iisimvi'\>!i-:: "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi L I E) R.ARY OF THE VERSITY U N I or ILLINOIS REMO
 R.ARY OF THE VERSITY U N I or ILLINOIS REMO") "^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
"^ A%'''''-'^-''S.''v.--..V^'E^'-'-^"-t''gi v:ii-ixi, 'i':;iisimvi'\>!i-:: L I E) R.ARY OF THE U N I VERSITY or ILLINOIS REMO Natural History Survey Librarv GEOLOGICAL SERIES OF FIELD MUSEUM OF NATURAL
Your Eye, My Eye, and the Eye of the Aye Aye: Evolution of Human Vision from 65 Million Years Ago to the Present
 # 75 Your Eye, My Eye, and the Eye of the Aye Aye: Evolution of Human Vision from 65 Million Years Ago to the Present Dr. Christopher Kirk December 2, 2011 Produced by and for Hot Science - Cool Talks
# 75 Your Eye, My Eye, and the Eye of the Aye Aye: Evolution of Human Vision from 65 Million Years Ago to the Present Dr. Christopher Kirk December 2, 2011 Produced by and for Hot Science - Cool Talks
Interpreting Evolutionary Trees Honors Integrated Science 4 Name Per.
 Interpreting Evolutionary Trees Honors Integrated Science 4 Name Per. Introduction Imagine a single diagram representing the evolutionary relationships between everything that has ever lived. If life evolved
Interpreting Evolutionary Trees Honors Integrated Science 4 Name Per. Introduction Imagine a single diagram representing the evolutionary relationships between everything that has ever lived. If life evolved