The Pennsylvania State University. The Graduate School. Department of Biochemistry and Molecular Biology VIRULENCE AND INFECTION: INTERACTIONS BETWEEN
|
|
|
- Blake Hilary Griffith
- 6 years ago
- Views:
Transcription
1 The Pennsylvania State University The Graduate School Department of Biochemistry and Molecular Biology VIRULENCE AND INFECTION: INTERACTIONS BETWEEN BORDETELLA BRONCHISEPTICA AND THE IMMUNE SYSTEM A Dissertation in Biochemistry, Microbiology, and Molecular Biology by Sarah J. Muse 2014 Sarah J. Muse Submitted in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy August 2014
2 The dissertation of Sarah J. Muse was reviewed and approved* by the following: Eric T. Harvill Professor of Microbiology and Infectious Disease Dissertation Advisor Chair of Committee Andrea Mastro Professor of Microbiology and Cell Biology Robert F. Paulson Professor of Veterinary and Biomedical Sciences K. Sandeep Prabhu Professor of Immunology and Molecular Toxicology Scott B. Selleck Professor and Head, Department of Biochemistry and Molecular Biology *Signatures are on file in the Graduate School
3 iii ABSTRACT Infectious diseases are a source of serious problems for humanity. By understanding how pathogenic bacteria utilize virulence factors to interact with their host, researchers can increase their knowledge of the mechanisms underlying infections and develop improved preventative measures and treatments to combat disease. In these studies, the respiratory pathogen B. bronchiseptica was used to study different aspects of virulence. B. bronchiseptica arnt, which encodes a protein that modifies the lipid A by adding glucosamine, was shown to be required for resistance to cationic antimicrobial peptides. Although arnt does not alter TLR4 agonist activity, the presence of arnt was necessary for transmission of B. bronchiseptica between hosts in the mouse model. These data indicate a novel role for lipid A modification in influencing between host infection dynamics. In a second series of experiments, we investigated how a pathogen could overcome early innate immune responses by manipulating host cell signaling. B. bronchiseptica infection of cultured macrophages revealed that clpv, a component of the Type VI secretion system (T6SS), is involved in decreasing levels of caspase-1, an important proinflammatory signaling molecule and inflammasome component. The decrease in caspase-1 is mediated through induction of autophagy. This mechanism suggests a novel interplay between a recently characterized bacterial secretion system and inflammasome signaling and hints at a bacterial virulence strategy to manipulate potentially damaging immune responses. Finally, Hcp, a core component of the T6SS, was investigated and was found to be necessary for bacterial processes, such as cell division and growth.
4 iv TABLE OF CONTENTS List of Figures... vi List of Tables... viii Acknowledgements... ix Abbreviations... xi Chapter 1 Introduction Host-pathogen interactions... 2 Bordetella as a model... 6 Bordetella Virulence Factors... 7 Preface References Chapter 2 Enzymatic Modification of the Lipid A by ArnT protects B. bronchiseptica against Cationic Peptides and Is Required for Transmission Abstract Introduction Materials and Methods Results Discussion Author Contributions References Chapter 3 B. bronchiseptica and the Caspase-1 Inflammasome Abstract Introduction Materials and Methods Results Discussion Author Contributions References Chapter 4 Hcp is necessary for normal B. bronchiseptica growth and morphology Abstract Introduction Materials and Methods Results Discussion Author Contributions
5 v References Chapter 5 Summary and Significance Synopsis Enzymatic Modification of the Lipid A by an ArnT Protects B. bronchiseptica Against Cationic Peptides and Is Required for Transmission Summary and Implications Future Directions Bordetella bronchiseptica and the Inflammasome Summary and Implications Future Directions Hcp is necessary for normal B. bronchiseptica growth and morphology Summary and Implications Future Directions Conclusion References Appendix A Parameters of B. bronchiseptica Infection in Relation to Caspase
6 vi LIST OF FIGURES Figure 1.1. Bacterial-Host Interaction Model Figure 1.2. Inflammasome Signaling Pathway Figure 1.3. BvgAS System Figure 1.4. Pertussis Toxin and Adenylate Cyclase Toxin Figure 1.5. Bordetella Adhesins Figure 1.6. Schematic of the basic structure of lipopolysaccharide Figure 1.7. Type III Secretion System (T3SS) Figure 1.8. Type VI Secretion System (T6SS) Figure 2.1. Ions corresponding to glucosamine additions in RB50 are not seen in RB50ΔarnT Figure 2.2. ArnT was not required for induction of TNF TNFα in murine macrophages, complement resistance, or adherence to the lung epithelial cells Figure 2.3. ArnT was required for resistance to killing by cationic antimicrobial peptides Figure 2.4. ArnT did not contribute to growth or resistance in the respiratory tract in a high dose model of respiratory infection Figure 2.5. ArnT decreased the mean infectious dose of B. bronchiseptica Figure 2.6. ArnT was required for transmission of B. bronchiseptica between mice Figure 2.7. ArnT did not affect neutrophil recruitment or shedding from the host Figure 3.1. IL-1 and the pro-inflammatory immune response Figure 3.2. clpv is required for mortality in B. bronchiseptica infected IL-1R -/- mice Figure 3.3. IL-1R is required for control of B. bronchiseptica respiratory colonization Figure 3.4. The Type VI Secretion System (T6SS) Figure 3.5. Deletion of clpv does not affect induction of IL-1β in murine lungs during infection Figure 3.6. Caspase-1 Inflammasome Complex Figure 3.7. Pathogen Interaction with the Inflammasome
7 vii Figure 3.8. Autophagy and the Inflammasome Figure 3.9. Effect of clpv and Caspase-1 during B. bronchiseptica Colonization in the Mouse Model Figure Model of T6SS affecting the inflammasome in macrophages Figure clpv was required for reduction of capase-1 protein in cultured macrophages Figure clpv is required for IL-1β production Figure clpv did not affect Casp1 mrna levels Figure clpv is necessary for in decreased levels of Nlrp3 protein Figure clpv is required for induction of autophagy and degradation of caspase Figure 4.1. The Type VI Secretion System (T6SS) Figure 4.2. Hcp protein Figure 4.3. Construction and confirmation of RB50Δhcp Figure 4.4. hcp contributed to bacterial growth Figure 4.5. hcp is required for polymyxin B resistance Figure 4.6. RB50Δhcp optical density did not correspond to colony forming units Figure 4.7. hcp is required for normal morphology in B. bronchiseptica Figure 4.8. Model of hcp in B. bronchiseptica growth Figure A.1. Measuring T6SS dependent caspase-1 activity Figure A.2. The Type III Secretion System is required for decreased levels of caspase-1 protein
8 viii LIST OF TABLES Table 4.1. Primer Sequences
9 ix ACKNOWLEDGEMENTS Chapter 2 of this dissertation Enzymatic modification of lipid A by ArnT protects Bordetella bronchiseptica against cationic peptides and is required for transmission, has been published in Infection and Immunity; 2014 Feb;82(2): All permissions have been obtained regarding the reproduction of text and figures of this manuscript within the dissertation as well as for diagrams in Chapters 1, 3, and 4. First, I would like to thank my advisor Dr. Eric Harvill for his help and guidance throughout my research and my committee members Dr. Kenneth Keiler, Dr. Andrea Mastro, Dr. Robert Paulson, and Dr. K. Sandeep Prabhu for their input and advice. I would also like to thank current members of the Harvill lab, Dr. Laura Goodfield (for heroic efforts in template formatting), Dr. William Smallridge, Liron Bendor, Dr. Yury Ivanov, Tyler Malys, Kai Hu, and Dr. Bodo Lintz, as well as past lab members Dr. Olivier Rolin, David Place, Dr. Sara Hester, Dr. Xuqing Zhang, Dr. Laura Weyrich, Dr. Jihye Park, Dr. Alexia Karanikas, Heather Feaga, Nathan Jacobs, Chetan Safi, and Melissa Augustino for their invaluable support and patience over the years. In addition, I would like to acknowledge our collaborators Dr. Andrew Preston at the University of Bath, UK, Dr. Robert Ernst and Lauren Hittle at the University of Maryland School of Dentistry, and Dr. Shokrollah Elahi Dr. Volker Gerdts at the University of Saskatchewan, Canada, and Dr. Sarah Barchinger, formerly from the Ades lab. I would like to extend my thanks to Dr. Mary Kennett, Dr. Pamela Hankey, and the National Institutes of Health (NIH) for providing me training and funding during my
10 x research through the Animal Models of Inflammation Training Grant. I would also like to thank Dr. Girish Kirimanjeswara and his lab for assistance with the caspase-1 project. Special thanks are given to Rachel Markley for help with the bone marrow derived macrophages and Dr. Kalyan Dewan for technical suggestions. In addition, I would like to acknowledge the excellent staff members at the Penn State Core Facilities for help with my work. I would like to thank Missy Hazen, John Cantolina, Dr. Ruth Nissly, and Dr. Greg Ning from the Microscopy and Cytometry Facility for all their hours of training and help. I would also like to thank Dr. Tatiana Laremore and Dr. Philip Smith from the Proteomics and Mass Spectrometry Core Facility for help with 2D gel proteomics and bacterial protein identification. Finally, I would like to thank my parents and friends for their constant support and understanding. Thank you to Matthew Moreau for editorial input for the Hcp project. Special thanks to Nick, Anand, Greg, Sheila, and Jan for helping me with this process. In the words of Julian of Norwich, All shall be well, and all shall be well and all manner of things shall be well.
11 xi ABBREVIATIONS 3MA: 3-methyl adenine ACT: Adenylate cyclase toxin BG: Bordet-Gengou agar BMDM: Bone marrow derived macrophage CAMP: Cationic antimicrobial peptide CDC: Center for Disease Control CFU: Colony forming unit DMEM: Dulbecco s modified Eagle s medium DNT: Dermonecrotic toxin ELISA: Enzyme Linked Immunosorbent Assay FBS: Fetal bovine serum FHA: Filamentous hemagglutinin Fim: Fimbriae GlcN: Glucosamine Hcp: haemolysin coregulated protein HRP: Horseradish Peroxidase ID50: Dose to infect 50% of individuals IL: Interleukin LB: Luria Bertani broth LBP: lipopolysaccharide binding protein LPS: lipopolysaccharide
12 xii MALDI-TOF: matrix-assisted laser desorption ionization time of flight mbd3: mouse β-defensin 3 MOI: Multiplicity of infection NLR: Nod-like receptor protein PAMP: Pathogen associated molecular pattern pbd1: Porcine β-defensin 1 PBS: Phosphobuffered saline Prn: Pertactin PRR: Pattern recognition receptor PT: Pertussis toxin SDS: Sodium dodecyl sulfate T3SS: Type III secretion system T6SS: Type VI secretion system TLR: Toll-like receptor TNF: Tumor necrosis factor USDA: United States Department of Agriculture
13 Chapter 1 Introduction.
14 2 Infectious diseases, which occur when transmissible pathogens interact with their hosts, are a source of major problems for humanity. According to the World Health Organization, infectious diseases are responsible for 16.2% of all deaths worldwide, account for four of the top ten causes of death, and are the predominant cause of death for people from low-income countries (2). Besides the staggering contribution to mortality, infectious diseases also cause disruption on an emotional, social, and economic level for the population (41). As estimated by the CDC, the annual cost of infectious diseases, both direct and indirect, is approximately $120 billion (78). In addition to directly affecting global human health, infectious diseases that afflict animals are also a burden on agriculture. The USDA reports that infectious diseases are responsible for billions of dollars lost by livestock producers each year (99). The incidence of infectious disease is an eminent and pervasive threat to human health and well-being (through human restricted pathogens and the rise of zoonoses (60)) as well as the food supply and world economy (5). To combat this critical dilemma, scientists are working to improve current protective measures as well as develop novel strategies though increased understanding of how pathogens cause disease. Host-pathogen interactions Pathogens are organisms that have characteristics that enable them to cause damage to their hosts. Types of pathogens include viruses, fungi, other eukaryotic microorganisms, and bacteria. Pathogenic bacteria, the focus of this dissertation, are classified thusly due to their ability to cause tissue damage and/or cell death in the host.
15 3 However, successful infectious pathogens have multiple key traits that enable the optimal utilization of their host environment : the capacity to overcome initial barriers and colonize a specific microenvironment; the ability to compete with commensal microbiota within a niche; the capability to survive, evade, and possibly manipulate host immune defenses; and the ability to transmit to a new host (28). The first objective for pathogenic bacteria is to prevail against the challenges imposed by the host microenvironment and establish an infection (Figure 1.1). The bacteria must navigate physical impediments, such as the skin or the mucosa at preferred points of entry (35). Then, the bacteria must contend with the mechanical perturbation of the mucocilliary interface (88) as well as innate immune defenses such as lysozymes (26), antimicrobial peptides (104), and complement (59). The bacteria must produce appropriate adherence factors (17) and have the ability to provide sufficient protection to survive and colonize the target environmental niche (80). Another important aspect of pathogenicity, which was not widely considered until recently, is the competition of invading microbes with commensal microbiota (34). Because of the adaptations of the commensal microbe species, they create a barrier against invasive species (45). Pathogens that are competing for the same space or pool of resources need methods to interfere with homeostasis and gain an advantage. The competition between the pathogen and commensals could involve direct killing (49), the disruption of biofilms (70), or differences in ability to withstand bacteriophage (10), or manipulation of an immune response (13). The host immune response is a complex system that is structured to create multiple layers of defenses against invading pathogens. A cursory overview of the
16 4 immune processes illustrates potential problems for the pathogenic bacteria in question. In addition to the previously mentioned difficulties, pathogens must also contend with other aspects of innate immunity (34). The bacteria would also encounter resident immune cells or even epithelia that respond to bacterial pathogen associated molecular patterns (PAMPs) (54). The immune cells would take advantage of highly conserved pattern recognition receptors (PRRs) (71) such as the transmembrane TLR (Toll-like receptor) or the cytosolic NLR (Nod-like receptor protein) (Figure 1.2) (42). Downstream signaling events, such as TLR4 activation of MyD88 and NF-kB (63) or even inflammasome complex formation between a Nod-like receptor, caspase-1, and Asc (92), result in induction of pro-inflammatory cytokines (36) as well as cell recruitment to the site of infection (50). Another obstacle would be to avoid or survive engulfment by phagocytic cells (12) as well as prevent antigen presentation and recruitment of the adaptive immune response (8). That process would influence the recruitment and differentiation of T cells (91), B cells (93), and ultimately the humoral immune response (37). To avoid detection or favorably alter the type of immune response elicited, pathogens rely on the production and delivery of specific virulence factors that manipulate host defense cellular properties (15), ensuring bacterial survival. After successfully replicating in the host environment, the infectious pathogenic bacteria must then attempt to transmit. Virulence factors provide a selective advantage in enabling the bacteria to move to another host without causing the death of the current host (102). Experiments with the mouse infection model indicate that bacterial transmission correlates with the generation of localized inflammation (74). The presence
17 5 of virulence factors such as the Type III secretion system and adenylate cyclase toxin is required for increased recruitment of polymorphonuclear leukocytes (PMN) and increased levels of bacterial transmission from the murine nasal cavity (89). The presence of PMNs could contribute to transmission by increasing bacterial shedding levels by promoting a coughing or sneezing response (14) and augmenting the flow of mucus out of the site of infection (6). These findings indicate that virulence factor pathogenicity could aid in the spread of infection. To accomplish all of these objectives, successful bacterial pathogens rely on the production of virulence factors. These molecules or proteins have specific functions that enable the bacteria to subvert host defenses, acquire resources, as well as facilitate transmission. Virulence factor properties are numerous and varied, but all abilities work to promote bacterial survival in the various conditions encountered as the infection progresses. However, caveats exist concerning the appropriate use and regulation of virulence factors. The ability of many virulence factors to promote pathogenicity are highly dependent on the ecological niche of the bacteria (25). Virulence factors are not necessarily multipurpose, nor are they effective for all stages of the pathogen s life cycle. For example, the B. bronchiseptica Type III secretion system and Bvg regulated virulence factors cause disease and allow for persistence in swine but do not necessarily contribute to transmission (76, 77). The successful utilization of virulence factors is constrained by functional, temporal, and environmental considerations. Therefore, understanding the molecular mechanisms of virulence factors requires the use of a model system for both host and pathogen.
18 6 To investigate virulence strategies utilized by bacteria and the effects that those traits have on host organs and immune system, the respiratory pathogen Bordetella bronchiseptica was used. Bordetella as a model B. bronchiseptica is one of the most commonly studied species of the Bordetella genus and is classified as a member of the classical bordetellae (67). The other members are B. pertussis, the causative agent of whooping couch in humans (19), and B. parapertussis (61), strains of which have been isolated from patients with whooping cough symptoms but has also been shown to cause disease in sheep (21). B. bronchiseptica, B. pertussis, and B. parapertussis strains are considered subspecies of a single species adapted to various hosts (67) because of similarities in DNA sequence (100), insertion sequence (IS) element polymorphisms (95), multilocus enzyme electrophoresis (MLEE) typing (75), and metabolic characteristics (81). B. bronchiseptica is considered a progenitor-like subspecies in relation to the other members of the classical bordetellae based on measurements of genetic diversity (82) and has a broad host range (38). Notably, B. bronchiseptica has been shown to cause a form of atrophic rhinitis, a swine respiratory disease (1) that results in facial distortions that lead to difficulties in feeding and the development of respiratory distress. Atrophic rhinitis has been detected in over twenty percent of farms surveyed and results in costly herd losses every year (1, 16). Further contributing to agricultural difficulties, B. bronchiseptica infection increases swine susceptibility to other respiratory diseases,
19 7 such as Haemophilus parasuis (22) and Pasteurella multocida (24), as well as the severity of co-infection with porcine respiratory coronavirus (23). In addition to infecting swine, B. bronchiseptica has been shown to causes disease in a wide range of mammals, including dogs (57), rabbits (62), guinea pigs (43), koalas (69), sea otters (94), and even humans (82). It is especially noteworthy that B. bronchiseptica also infects mice (67). This enables the use of a murine infection model to study the natural interaction between host and pathogen while enabling investigators to take advantage of the extensive body of literature and tools available for the study of mice. Using the mouse model, as well as through experimentation with cell culture and molecular biology techniques, researchers are able to mechanistically delineate the immunomodulatory and damage-inducing phenomena that occur during host-pathogen interactions by identifying and characterizing virulence factors in B. bronchiseptica. Bordetella Virulence Factors The major virulence regulatory system in Bordetella is the BvgAS two component system (31). BvgS acts as a sensor kinase and, in response to environmental stimuli, acts through a phosphorelay system to phosphorylate the BvgA regulator, resulting in dimerization and activation and repression of transcription (Figure 1.3A) (32, 98). This system serves as a master regulator for Bordetella virulence and controls the upregulation and downregulation of virulence gene sets in the various Bvg phases (Figure 1.3B) (72). In the Bvg + phase, virulence associated genes such as toxins and adherence
20 8 factors are upregulated (73), but they are downregulated in the Bvg - phase in favor of expression of metabolic and motility genes (3, 4). The genes expressed in this phase are necessary for successful B. bronchiseptica colonization of the host. The Bvg- phase may be required for survival outside of the host and in nutrient limiting conditions. An intermediate Bvg i phase also exists in which adherence factors and metabolic genes are upregulated but the other gene sets from both phases are not (97, 103). Changes in Bvg phase directly influence the virulence factors produced in response to the environment. A major class of virulence factors is bacterial toxins. The most famous toxin associated with Bordetella is pertussis toxin (PT) (85). PT is Bvg regulated (27) and functions by causing ADP-ribosylation of host proteins and altering G protein signaling (Figure 1.4 A,B) (56). Although the genome of B. bronchiseptica contains the coding regions necessary to produce PT, expression of the locus has not been observed (7). Adenylate cyclase toxin (ACT) is a calmodulin-sensitive adenylate cyclase/hemolysin that is expressed in Bvg + phase (67). However, non-bvg regulated factors, such as CO2 (47), have also been shown to influence its expression. ACT induces the overproduction of camp (Figure 1.4C,D), resulting in impaired phagocytosis (30) and induction of apoptosis in macrophages (44). B. bronchiseptica produces adenylate cyclase toxin and dermonecrotic toxin which have been shown to interact with host cell pathways. Dermonecrotic toxin (DNT) is highly conserved among the bordetellae and was discovered when B. pertussis extracts caused necrosis on the skin of treated mice (33). Properties of DNT include activation of Rho GTPase to inhibit cell differentiation and proliferation (52), disruption of the host cell cycle (58), and inhibition of antibody
21 9 response in mice (96). DNT is also a key virulence factor in causing the aforementioned atrophic rhinitis in swine (51). Another important type of virulence factor in B. bronchiseptica is adhesins. Fimbriae (Fim2/3), filamentous hemagglutinin (FHA), and pertactin (Prn) (Figure 1.5 A, B, C) enable B. bronchiseptica to adhere to respiratory tract cells (39) and also serve as components of the acellular pertussis vaccine. Fim and FHA are both necessary for tracheal colonization and are important for induction of an antibody response (53, 68). The membrane component lipopolysaccharide (LPS) (Figure 1.6) can also influence the ability of bacteria to survive and evade the host immune response. LPS consists of 3 main regions: the lipid A, which serves to anchor the LPS into the outer membrane of Gram negative bacteria (65); the core oligosaccharide, which commonly linked to the lipid A through 2-keto-3-deoxyoctulosonic acid; and the O antigen, which attaches to the core and consists of repeating polysaccharide chains (86). LPS can protect B. bronchiseptica from antimicrobial peptides (40), but the lipid A portion also interacts with TLR4 (66), resulting in the generation of a proinflammatory response. The addition of modifications to the lipid A can, in turn, influence changes in the host immune response. Previous work has shown that PagP palmitoyl transferase is necessary for colonization (87) and resistance to complement (84). The O-antigen component of LPS can also influence the bacterial interaction with the host. The presence of O-antigen is necessary for B. bronchiseptica colonization of the respiratory tract (46) and acts to protect bacteria from antimicrobial peptides and complement deposition (9). The bordetellae have 3 horizontally acquired O-antigen
22 10 types (48), and these polysaccharides have different levels of immungenicity, affecting resistance to antibody mediated killing. Secretion systems function to export proteins required for virulence. In B. bronchiseptica, two secretion systems are associated with pathogenesis: the Type III secretion system (T3SS) and the recently characterized Type VI secretion system (T6SS). The T3SS consists of a needle-like injection apparatus powered by a system designated ATPase (bscn) that transports effector proteins externally or into host cells (Figure 1.7) (79). The T3SS is required for caspase-1 dependent necrosis (96) in culture as well as inhibition of NF-kB signaling (105). In the mouse model, the T3SS is necessary for colonization in the lungs and inhibits the proinflammatory interferon (IFN)-γ that is required for antibody mediated clearance (83). The B. bronchiseptica T6SS was recently identified (20) and an initial characterization was performed (101). The T6SS consists of 13 core components as well as 13 Bordetella specific genes (Figure 1.8A). These proteins form a T4 phage tail-like secretion apparatus which has been shown to be contact dependent in other systems. Important conserved system components include Hcp, which has been shown to form nanotubule structures through which effectors can pass (Figure 1.8B) (18); VgrG, the tip of the secretion apparatus which interacts with membrane components present in target cells (90); IcmF, a conserved inner membrane protein that has been shown to contribute to ATP hydrolysis and binding (64); and ClpV, a AAA+ ATPase which serves as the main power source for the system and enables contraction of the apparatus (55). The T6SS is thought to assemble by polymerization of the Hcp-based tubule in an ATP dependent manner after the base-plate complex of proteins that span the inner and
23 11 outer membranes are brought together (29). Then, the VipA/VipB sheath is hypothesizes to form in the uncontracted position around the Hcp tubule, which is capped with the VgrG protein. After the system is stimulated, the VipA/VipB sheath contracts, enabling effector proteins, Hcp, and VgrG to be delivered into the target cell or the extracellular milieu (Figure 1.8C) (11). clpv is required for colonization in the mouse model, contributes to lung pathology (101), inhibits antibody mediated clearance by suppression of proinflammatory cytokines (L. S. Weyrich, unpublished), and is required for B. bronchiseptica mediated displacement of the nasal cavity microbiota (L.S. Weyrich, unpublished data). Additonally, it has been shown to affect IL-6, IL-1β, IL-17, and IL-10 cytokine responses in vitro and IFN-γ in vivo (101). Besides Hcp, none of the effector molecules secreted by the B. bronchiseptica T6SS have been identified. However, potential candidates include the proteins encoded by loci BB0797 (tssd) and BB0798 (tsse), which form a putative toxin-antitoxin system. Additional work has also been performed analyzing the role of BB0804 (tssj), which encodes a gp25-like lysosome and is hypothesized to be part of the base-plate of the apparatus or possibly secreted. Preliminary results have shown that TssJ does not affect cytotoxicity in culture and is not required for colonization of the respiratory tract in the mouse (H. Feaga, unpublished). However, during infection, the presence of TssJ has been shown to be required for the expression of the pro-inflammatory cytokines IL-6 and IL-1β as well as the chemokines KC and MIP1-α. B. bronchiseptica is capable of producing a wide array of virulence factors to facilitate its survival, replication, and transmission to a new host. By performing
24 12 experiments to characterize the interface of virulence strategies with the immune system, we hope to create a more detailed understanding of how virulence factors function in order to counter their effects and prevent disease. Preface The focus of this dissertation is investigating how the respiratory pathogen B. bronchiseptica utilizes virulence strategies to interact with the host and survive. The work presented in Chapter 2 features arnt, which encodes an enzyme that adds a glucosamine modification on the B. bronchiseptica lipid A. This virulence factor provides resistance to antimicrobial peptides and influences the ability of the bacteria to initially colonize the nasal cavity as well as its ability to affect leukocyte recruitment and facilitate transmission. The material in Chapter 2 was previously published in Infection and Immunity. In Chapter 3, an examination of a novel mechanism of virulence factor mediated manipulation of host immune cells is presented. In cultured macrophage cells, the presence of clpv, which encodes the major power source of the T6SS, is required for a decrease in caspase-1, a central inflammatory signaling protein. This decrease is facilitated through the clpv dependent induction of autophagy, implicating the involvement of the T6SS in a host cell degradation pathway to decrease inflammatory signaling. The experiments described in Chapter 4 concerned the investigation of Hcp, a core component of the T6SS (a system previously implicated in virulence in culture and
25 13 the mouse model), which is required for essential bacterial properties such as membrane stability and normal cell division. The overall results are summarized in Chapter 5, and the significance of the findings and virulence strategies on host-pathogen interactions is discussed.
26 14 References Swine United States Department of Agriculture, vol. II. Reference of swine health and health management in the United States The top 10 causes of death. World Health Organization. 3. Akerley, B. J., and J. F. Miller Flagellin gene transcription in Bordetella bronchiseptica is regulated by the BvgAS virulence control system. J Bacteriol 175: Akerley, B. J., D. M. Monack, S. Falkow, and J. F. Miller The bvgas locus negatively controls motility and synthesis of flagella in Bordetella bronchiseptica. J Bacteriol 174: Anderson, P. P The Future World Food Situation and the Role of Plant Diseases. Plant Health Instructor. 6. Antunes, M. B., D. A. Gudis, and N. A. Cohen Epithelium, cilia, and mucus: their importance in chronic rhinosinusitis. Immunol Allergy Clin North Am 29: Arico, B., and R. Rappuoli Bordetella parapertussis and Bordetella bronchiseptica contain transcriptionally silent pertussis toxin genes. J Bacteriol 169: Atif, S. M., S. E. Winter, M. G. Winter, S. J. McSorley, and A. J. Baumler. Salmonella enterica serovar Typhi impairs CD4 T cell responses by reducing antigen availability. Infect Immun. 9. Banemann, A., H. Deppisch, and R. Gross The lipopolysaccharide of Bordetella bronchiseptica acts as a protective shield against antimicrobial peptides. Infect Immun 66: Barr, J. J., R. Auro, M. Furlan, K. L. Whiteson, M. L. Erb, J. Pogliano, A. Stotland, R. Wolkowicz, A. S. Cutting, K. S. Doran, P. Salamon, M. Youle, and F. Rohwer. Bacteriophage adhering to mucus provide a non-host-derived immunity. Proc Natl Acad Sci U S A 110: Basler, M., M. Pilhofer, G. P. Henderson, G. J. Jensen, and J. J. Mekalanos. Type VI secretion requires a dynamic contractile phage tail-like structure. Nature 483: Baxt, L. A., A. C. Garza-Mayers, and M. B. Goldberg. Bacterial subversion of host innate immune pathways. Science 340: Belkaid, Y., and T. W. Hand. Role of the Microbiota in Immunity and Inflammation. Cell 157: Bemis, D. A., W. R. Shek, and C. B. Clifford Bordetella bronchiseptica infection of rats and mice. Comp Med 53: Bliska, J. B., X. Wang, G. I. Viboud, and I. E. Brodsky. Modulation of innate immune responses by Yersinia type III secretion system translocators and effectors. Cell Microbiol 15: Boessen, C. R., J. B. Kliebenstein, R. P. Cowart, K. C. Moore, and C. R. Burbee Effective use of slaughter checks to determine economic losses from morbidity in swine. Acta Vet Scand Suppl 84: Boland, T., R. A. Latour, and F. J. Stutzenberger Molecular basis of bacterial adhesion, p Handbook of Bacterial Adhesion. Springer. 18. Bonemann, G., A. Pietrosiuk, and A. Mogk. Tubules and donuts: a type VI secretion story. Mol Microbiol 76: Bordet, J., and O. Gengou Note complã mentaire sur le microbe de la coqueluche, vol.
27 20. Boyer, F., G. Fichant, J. Berthod, Y. Vandenbrouck, and I. Attree Dissecting the bacterial type VI secretion system by a genome wide in silico analysis: what can be learned from available microbial genomic resources? BMC Genomics 10: Brinig, M. M., K. B. Register, M. R. Ackermann, and D. A. Relman Genomic features of Bordetella parapertussis clades with distinct host species specificity. Genome Biol 7:R Brockmeier, S. L Prior infection with Bordetella bronchiseptica increases nasal colonization by Haemophilus parasuis in swine. Vet Microbiol 99: Brockmeier, S. L., C. L. Loving, T. L. Nicholson, and M. V. Palmer Coinfection of pigs with porcine respiratory coronavirus and Bordetella bronchiseptica. Vet Microbiol 128: Brockmeier, S. L., M. V. Palmer, S. R. Bolin, and R. B. Rimler Effects of intranasal inoculation with Bordetella bronchiseptica, porcine reproductive and respiratory syndrome virus, or a combination of both organisms on subsequent infection with Pasteurella multocida in pigs. Am J Vet Res 62: Buboltz, A. M., T. L. Nicholson, M. R. Parette, S. E. Hester, J. Parkhill, and E. T. Harvill Replacement of adenylate cyclase toxin in a lineage of Bordetella bronchiseptica. J Bacteriol 190: Callewaert, L., and C. W. Michiels. Lysozymes in the animal kingdom. J Biosci 35: Carbonetti, N. H., N. Khelef, N. Guiso, and R. Gross A phase variant of Bordetella pertussis with a mutation in a new locus involved in the regulation of pertussis toxin and adenylate cyclase toxin expression. J Bacteriol 175: Casadevall, A., and L. A. Pirofski Host-pathogen interactions: redefining the basic concepts of virulence and pathogenicity. Infect Immun 67: Cascales, E., and C. Cambillau. Structural biology of type VI secretion systems. Philos Trans R Soc Lond B Biol Sci 367: Confer, D. L., and J. W. Eaton Phagocyte impotence caused by an invasive bacterial adenylate cyclase. Science 217: Cotter, P. A., and J. F. Miller BvgAS-mediated signal transduction: analysis of phase-locked regulatory mutants of Bordetella bronchiseptica in a rabbit model. Infect Immun 62: Cotter, P. A., and J. F. Miller A mutation in the Bordetella bronchiseptica bvgs gene results in reduced virulence and increased resistance to starvation, and identifies a new class of Bvg-regulated antigens. Mol Microbiol 24: Cowell, J. L., E. L. Hewlett, and C. R. Manclark Intracellular localization of the dermonecrotic toxin of Bordetella pertussis. Infect Immun 25: David, R. Microbiome: Pathogens and commensals fight it out. Nat Rev Microbiol 10: Davis, K. M., H. T. Akinbi, A. J. Standish, and J. N. Weiser Resistance to mucosal lysozyme compensates for the fitness deficit of peptidoglycan modifications by Streptococcus pneumoniae. PLoS Pathog 4:e de Jong, H. K., C. M. Parry, T. van der Poll, and W. J. Wiersinga. Host-pathogen interaction in invasive Salmonellosis. PLoS Pathog 8:e Deknuydt, F., T. Nordstrom, and K. Riesbeck. Diversion of the host humoral response: a novel virulence mechanism of Haemophilus influenzae mediated via outer membrane vesicles. J Leukoc Biol. 38. Diavatopoulos, D. A., C. A. Cummings, L. M. Schouls, M. M. Brinig, D. A. Relman, and F. R. Mooi Bordetella pertussis, the causative agent of whooping cough, 15
28 evolved from a distinct, human-associated lineage of B. bronchiseptica. PLoS Pathog 1:e Edwards, J. A., N. A. Groathouse, and S. Boitano Bordetella bronchiseptica adherence to cilia is mediated by multiple adhesin factors and blocked by surfactant protein A. Infect Immun 73: Erles, K., and J. Brownlie. Expression of beta-defensins in the canine respiratory tract and antimicrobial activity against Bordetella bronchiseptica. Vet Immunol Immunopathol 135: Fonkwo, P. N Pricing infectious disease. The economic and health implications of infectious diseases. EMBO Rep 9 Suppl 1:S Fukata, M., and M. Arditi. The role of pattern recognition receptors in intestinal inflammation. Mucosal Immunol 6: Griffith, J. W., K. M. Brasky, and C. M. Lang Experimental pneumonia virus of mice infection of guineapigs spontaneously infected with Bordetella bronchiseptica. Lab Anim 31: Gueirard, P., A. Druilhe, M. Pretolani, and N. Guiso Role of adenylate cyclasehemolysin in alveolar macrophage apoptosis during Bordetella pertussis infection in vivo. Infect Immun 66: Harvill, E. T. Cultivating our "frienemies": viewing immunity as microbiome management. MBio Harvill, E. T., A. Preston, P. A. Cotter, A. G. Allen, D. J. Maskell, and J. F. Miller Multiple roles for Bordetella lipopolysaccharide molecules during respiratory tract infection. Infect Immun 68: Hester, S. E., M. Lui, T. Nicholson, D. Nowacki, and E. T. Harvill. Identification of a CO2 responsive regulon in Bordetella. PLoS One 7:e Hester, S. E., J. Park, L. L. Goodfield, H. A. Feaga, A. Preston, and E. T. Harvill. Horizontally acquired divergent O-antigen contributes to escape from cross-immunity in the classical bordetellae. BMC Evol Biol 13: Hibbing, M. E., C. Fuqua, M. R. Parsek, and S. B. Peterson. Bacterial competition: surviving and thriving in the microbial jungle. Nat Rev Microbiol 8: Hickey, M. J., and C. L. Westhorpe. Imaging inflammatory leukocyte recruitment in kidney, lung and liver--challenges to the multi-step paradigm. Immunol Cell Biol 91: Horiguchi, Y. Swine atrophic rhinitis caused by pasteurella multocida toxin and bordetella dermonecrotic toxin. Curr Top Microbiol Immunol 361: Horiguchi, Y., T. Nakai, and K. Kume Effects of Bordetella bronchiseptica dermonecrotic toxin on the structure and function of osteoblastic clone MC3T3-e1 cells. Infect Immun 59: Inatsuka, C. S., S. M. Julio, and P. A. Cotter Bordetella filamentous hemagglutinin plays a critical role in immunomodulation, suggesting a mechanism for host specificity. Proc Natl Acad Sci U S A 102: Janeway, C. A., Jr Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harb Symp Quant Biol 54 Pt 1: Kapitein, N., G. Bonemann, A. Pietrosiuk, F. Seyffer, I. Hausser, J. K. Locker, and A. Mogk. ClpV recycles VipA/VipB tubules and prevents non-productive tubule formation to ensure efficient type VI protein secretion. Mol Microbiol 87: Katada, T. The inhibitory G protein G(i) identified as pertussis toxin-catalyzed ADPribosylation. Biol Pharm Bull 35:
29 57. Keil, D. J., and B. Fenwick Strain- and growth condition-dependent variability in outer membrane protein expression by Bordetella bronchiseptica isolates from dogs. Am J Vet Res 60: Lacerda, H. M., G. D. Pullinger, A. J. Lax, and E. Rozengurt Cytotoxic necrotizing factor 1 from Escherichia coli and dermonecrotic toxin from Bordetella bronchiseptica induce p21(rho)-dependent tyrosine phosphorylation of focal adhesion kinase and paxillin in Swiss 3T3 cells. J Biol Chem 272: Lambris, J. D., D. Ricklin, and B. V. Geisbrecht Complement evasion by human pathogens. Nat Rev Microbiol 6: Lefranà ois, T., and T. Pineau. Public health and livestock: Emerging diseases in food animals. Animal Frontiers 4: Long, G. H., A. T. Karanikas, E. T. Harvill, A. F. Read, and P. J. Hudson. Acellular pertussis vaccination facilitates Bordetella parapertussis infection in a rodent model of bordetellosis. Proc Biol Sci 277: Long, G. H., D. Sinha, A. F. Read, S. Pritt, B. Kline, E. T. Harvill, P. J. Hudson, and O. N. Bjornstad. Identifying the age cohort responsible for transmission in a natural outbreak of Bordetella bronchiseptica. PLoS Pathog 6:e Lu, Y. C., W. C. Yeh, and P. S. Ohashi LPS/TLR4 signal transduction pathway. Cytokine 42: Ma, L. S., F. Narberhaus, and E. M. Lai. IcmF family protein TssM exhibits ATPase activity and energizes type VI secretion. J Biol Chem 287: Maeshima, N., and R. C. Fernandez. Recognition of lipid A variants by the TLR4-MD- 2 receptor complex. Front Cell Infect Microbiol 3: Mann, P. B., D. Wolfe, E. Latz, D. Golenbock, A. Preston, and E. T. Harvill Comparative toll-like receptor 4-mediated innate host defense to Bordetella infection. Infect Immun 73: Mattoo, S., and J. D. Cherry Molecular pathogenesis, epidemiology, and clinical manifestations of respiratory infections due to Bordetella pertussis and other Bordetella subspecies. Clin Microbiol Rev 18: Mattoo, S., J. F. Miller, and P. A. Cotter Role of Bordetella bronchiseptica fimbriae in tracheal colonization and development of a humoral immune response. Infect Immun 68: McKenzie, R. A., A. D. Wood, and P. J. Blackall Pneumonia associated with Bordetella bronchiseptica in captive koalas. Aust Vet J 55: McMillan, A., M. Dell, M. P. Zellar, S. Cribby, S. Martz, E. Hong, J. Fu, A. Abbas, T. Dang, W. Miller, and G. Reid. Disruption of urogenital biofilms by lactobacilli. Colloids Surf B Biointerfaces 86: Medzhitov, R Recognition of microorganisms and activation of the immune response. Nature 449: Melvin, J. A., E. V. Scheller, J. F. Miller, and P. A. Cotter. Bordetella pertussis pathogenesis: current and future challenges. Nat Rev Microbiol 12: Merkel, T. J., S. Stibitz, J. M. Keith, M. Leef, and R. Shahin Contribution of regulation by the bvg locus to respiratory infection of mice by Bordetella pertussis. Infect Immun 66: Monack, D. M. Salmonella persistence and transmission strategies. Curr Opin Microbiol 15: Musser, J. M., E. L. Hewlett, M. S. Peppler, and R. K. Selander Genetic diversity and relationships in populations of Bordetella spp. J Bacteriol 166:
30 76. Nicholson, T. L., S. L. Brockmeier, C. L. Loving, K. B. Register, M. E. Kehrli, Jr., and S. M. Shore. The Bordetella bronchiseptica type III secretion system is required for persistence and disease severity but not transmission in swine. Infect Immun 82: Nicholson, T. L., S. L. Brockmeier, C. L. Loving, K. B. Register, M. E. Kehrli, Jr., S. E. Stibitz, and S. M. Shore. Phenotypic modulation of the virulent Bvg phase is not required for pathogenesis and transmission of Bordetella bronchiseptica in swine. Infect Immun 80: Noah, D., and G. Fidas The global infectious disease threat and its implications for the United States. DTIC Document. 79. Panina, E. M., S. Mattoo, N. Griffith, N. A. Kozak, M. H. Yuk, and J. F. Miller A genome-wide screen identifies a Bordetella type III secretion effector and candidate effectors in other species. Mol Microbiol 58: Papo, N., and Y. Shai A molecular mechanism for lipopolysaccharide protection of Gram-negative bacteria from antimicrobial peptides. J Biol Chem 280: Park, J., Y. Zhang, A. M. Buboltz, X. Zhang, S. C. Schuster, U. Ahuja, M. Liu, J. F. Miller, M. Sebaihia, S. D. Bentley, J. Parkhill, and E. T. Harvill. Comparative genomics of the classical Bordetella subspecies: the evolution and exchange of virulenceassociated diversity amongst closely related pathogens. BMC Genomics 13: Parkhill, J., M. Sebaihia, A. Preston, L. D. Murphy, N. Thomson, D. E. Harris, M. T. Holden, C. M. Churcher, S. D. Bentley, K. L. Mungall, A. M. Cerdeno-Tarraga, L. Temple, K. James, B. Harris, M. A. Quail, M. Achtman, R. Atkin, S. Baker, D. Basham, N. Bason, I. Cherevach, T. Chillingworth, M. Collins, A. Cronin, P. Davis, J. Doggett, T. Feltwell, A. Goble, N. Hamlin, H. Hauser, S. Holroyd, K. Jagels, S. Leather, S. Moule, H. Norberczak, S. O'Neil, D. Ormond, C. Price, E. Rabbinowitsch, S. Rutter, M. Sanders, D. Saunders, K. Seeger, S. Sharp, M. Simmonds, J. Skelton, R. Squares, S. Squares, K. Stevens, L. Unwin, S. Whitehead, B. G. Barrell, and D. J. Maskell Comparative analysis of the genome sequences of Bordetella pertussis, Bordetella parapertussis and Bordetella bronchiseptica. Nat Genet 35: Pilione, M. R., and E. T. Harvill The Bordetella bronchiseptica type III secretion system inhibits gamma interferon production that is required for efficient antibodymediated bacterial clearance. Infect Immun 74: Pilione, M. R., E. J. Pishko, A. Preston, D. J. Maskell, and E. T. Harvill pagp is required for resistance to antibody-mediated complement lysis during Bordetella bronchiseptica respiratory infection. Infect Immun 72: Pittman, M The concept of pertussis as a toxin-mediated disease. Pediatr Infect Dis 3: Preston, A., A. G. Allen, J. Cadisch, R. Thomas, K. Stevens, C. M. Churcher, K. L. Badcock, J. Parkhill, B. Barrell, and D. J. Maskell Genetic basis for lipopolysaccharide O-antigen biosynthesis in bordetellae. Infect Immun 67: Preston, A., E. Maxim, E. Toland, E. J. Pishko, E. T. Harvill, M. Caroff, and D. J. Maskell Bordetella bronchiseptica PagP is a Bvg-regulated lipid A palmitoyl transferase that is required for persistent colonization of the mouse respiratory tract. Mol Microbiol 48: Reynolds, H. Y Modulating airway defenses against microbes. Curr Opin Pulm Med 8: Rolin, O., W. Smallridge, M. Henry, L. Goodfield, D. Place, and E. T. Harvill. Tolllike receptor 4 limits transmission of Bordetella bronchiseptica. PLoS One 9:e
31 90. Sha, J., J. A. Rosenzweig, E. V. Kozlova, S. Wang, T. E. Erova, M. L. Kirtley, C. J. van Lier, and A. K. Chopra. Evaluation of the roles played by Hcp and VgrG type 6 secretion system effectors in Aeromonas hydrophila SSU pathogenesis. Microbiology 159: Shaler, C. R., C. Horvath, R. Lai, and Z. Xing. Understanding delayed T-cell priming, lung recruitment, and airway luminal T-cell responses in host defense against pulmonary tuberculosis. Clin Dev Immunol 2012: Sollberger, G., G. E. Strittmatter, M. Garstkiewicz, J. Sand, and H. D. Beer. Caspase-1: the inflammasome and beyond. Innate Immun 20: Spera, J. M., C. K. Herrmann, M. S. Roset, D. J. Comerci, and J. E. Ugalde. A Brucella virulence factor targets macrophages to trigger B-cell proliferation. J Biol Chem 288: Staveley, C. M., K. B. Register, M. A. Miller, S. L. Brockmeier, D. A. Jessup, and S. Jang Molecular and antigenic characterization of Bordetella bronchiseptica isolated from a wild southern sea otter (Enhydra lutris nereis) with severe suppurative bronchopneumonia. J Vet Diagn Invest 15: Stibitz, S., and M. S. Yang Genomic plasticity in natural populations of Bordetella pertussis. J Bacteriol 181: Stockbauer, K. E., A. K. Foreman-Wykert, and J. F. Miller Bordetella type III secretion induces caspase 1-independent necrosis. Cell Microbiol 5: Sukumar, N., M. Mishra, G. P. Sloan, T. Ogi, and R. Deora Differential Bvg phase-dependent regulation and combinatorial role in pathogenesis of two Bordetella paralogs, BipA and BcfA. J Bacteriol 189: Uhl, M. A., and J. F. Miller Autophosphorylation and phosphotransfer in the Bordetella pertussis BvgAS signal transduction cascade. Proc Natl Acad Sci U S A 91: United States Department of Agriculture, R. E. E. I. S INFECTIOUS DISEASE PATHOGENESIS STUDY AND PREVENTION DEVELOPMENT van der Zee, A., H. Groenendijk, M. Peeters, and F. R. Mooi The differentiation of Bordetella parapertussis and Bordetella bronchiseptica from humans and animals as determined by DNA polymorphism mediated by two different insertion sequence elements suggests their phylogenetic relationship. Int J Syst Bacteriol 46: Weyrich, L. S., O. Y. Rolin, S. J. Muse, J. Park, N. Spidale, M. J. Kennett, S. E. Hester, C. Chen, E. G. Dudley, and E. T. Harvill. A Type VI secretion system encoding locus is required for Bordetella bronchiseptica immunomodulation and persistence in vivo. PLoS One 7:e Wickham, M. E., N. F. Brown, E. C. Boyle, B. K. Coombes, and B. B. Finlay Virulence is positively selected by transmission success between mammalian hosts. Curr Biol 17: Williams, C. L., P. E. Boucher, S. Stibitz, and P. A. Cotter BvgA functions as both an activator and a repressor to control Bvg phase expression of bipa in Bordetella pertussis. Mol Microbiol 56: Yang, D., O. Chertov, and J. J. Oppenheim The role of mammalian antimicrobial peptides and proteins in awakening of innate host defenses and adaptive immunity. Cell Mol Life Sci 58: Yuk, M. H., E. T. Harvill, P. A. Cotter, and J. F. Miller Modulation of host immune responses, induction of apoptosis and inhibition of NF-kappaB activation by the Bordetella type III secretion system. Mol Microbiol 35:

32 20 Figure 1.1. Bacterial-Host Interaction Model. Bacterial pathogens must overcome multiple challenges over the course of infection. Initial obstacles include overcoming physical barriers to colonization such as mucus or the beating of cilia, withstanding antimicrobial peptides, complement, and lysozymes, outcompeting the commensal microflora, and prevention of engulfment by resident phagocytes. After the establishment of the pathogen, pro-inflammatory cytokine and chemokine responses initiated by the epithelium and resident immune cells and antigen presentation lead to the recruitment of leukocytes as well as T and B lymphocytes to the site of infection and the initiation of a humoral immune response. Modification of figure created by O. Rolin.

33 21 Figure 1.2. Inflammasome Signaling Pathway. Maturation and secretion of IL-1β requires 2 signals: the priming signal leads to synthesis of pro-il-1β, pro-il-18, and other components of the inflammasome, such as NLRP3, and the second signal results in assembly of the inflammasome, activation of caspase-1, and release of mature cytokines IL-1β and IL-18 into the extracellular milieu. Currently, the nature of the second signal is debated. The 3 proposed models of activation are shown: 1) extracellular ATP, which activates the purinergic P2X 7 receptor and causes subsequent recruitment of pannexin-1 hemichannel to the plasma membrane and K + efflux; 2) lysosomal rupture after engulfment of crystalline or particulate agonists; and 3) reactive oxygen species (ROS), which upregulate NLRP3 expression and activate the inflammasome. PAMP, pathogen-associated molecular pattern; DAMP, danger-associated molecular pattern; TLR, Toll-like receptor; dsdna, double-stranded DNA. Reprinted with permission from the American Physiological Society, Am J Physiol Lung Cell Mol Physiol Oct 15;303(8):L
 BvgS is a polydomain histidine sensor kinase that contains (from the amino to the carboxyl terminus) two periplasmically located venus flytrap domains (VFT1 and VFT2), a transmembrane domain, a")
34 22 Figure 1.3. BvgAS System. A) BvgS is a polydomain histidine sensor kinase that contains (from the amino to the carboxyl terminus) two periplasmically located venus flytrap domains (VFT1 and VFT2), a transmembrane domain, a PAS domain, a histidine kinase domain (HK), a receiver domain (Rec) and a histidine phosphoryl transfer domain (Hpt). BvgA is a response regulator protein that becomes autophosphorylated, and the phosphoryl group is then transferred to the Rec domain, followed by the Hpt and finally to the Rec domain of BvgA. Phosphorylated BvgA dimerizes and activates the expression of virulence-associated genes. B) Bvg Phase Gene Expression Profiles. The Bvg + phase occurs when BvgAS is fully active and is characterized by maximal expression of genes that encode adhesins and toxins and minimal expression of motility genes and bipa. The Bvg phase occurs when BvgAS is inactive and is characterized by maximal expression of bvgr and motility genes and minimal expression of toxins, adhesins and bipa. The Bvg i phase occurs when BvgAS is partially active and is characterized by the maximal expression of adhesins and bipa (expressed exclusively in this phase) and minimal expression of toxins and motility genes. Reprinted with permission from Nature Publishing Group, Nat Rev Microbiol Apr;12(4):
 Model of PT ADP-ribosylation.")

35 23 Figure 1.4. Pertussis Toxin and Adenylate Cyclase Toxin. A) Structure of Pertussis Toxin (PT) indicating the A subunit (catalytic subunit) and the B pentamer (transport subunits). B) Model of PT ADP-ribosylation. PT binds to a sialoglycoprotein host cell receptor, is endocytosed and trafficked to the endoplasmic reticulum (ER) where the B pentamer dissociates from the A subunit. The A subunit then traffics on exosomes to the cytoplasmic membrane, where it ADP-ribosylates the α-subunit of heterotrimeric G proteins, altering the ability of the G proteins to regulate multiple enzymes and pathways, including their ability to inhibit cyclic AMP (camp) formation. C) Structure of Adenylate Cyclase Toxin (ACT) indicating the adenylate cyclase domain connect via a hydrophobic segment to the RTX domain. D) Model of ACT function. The RTX domain of ACT interacts with complement receptor 3 (CR3). The hydrophobic segments of the linker region form pores in the membrane, enabling the passage of cations and the adenylate cyclase domain is translocated into the cytoplasm. Adenylate cyclase activity is stimulated by binding to calmodulin in the host cell, leading to an increase in camp production. Reprinted with permission from Nature Publishing Group, Nat Rev Microbiol Apr;12(4):

36 24 Figure 1.5. Bordetella Adhesins.. A) Filamentous haemagglutinin (FHA) is a TpsA exoprotein that is translocated across the outer membrane through its cognate TpsB pore protein, FhaC. This translocation occurs via the two-partner secretion pathway. Processing during translocation removes the carboxyterminal prodomain (yellow) from the full-length FhaB protein to produce the mature ~250 kda FHA protein. B) Bordetella fimbriae are type I pili. Fim2 and Fim3 are the major pilin subunits and FimD is likely to be the fimbrial tip protein. Both Fims and FHA are required for adherence to ciliated epithelial cells and modulating the host immune system. C) Pertactin is a classical autotransporter which is thought to play a role in cell adherence. The C-terminal ~30 kda domain (orange) forms a channel in the outer membrane, which is required for the translocation of the ~70 kda β-helical passenger domain (blue) to the cell surface. Reprinted with permission from Nature Publishing Group, Nat Rev Microbiol Apr;12(4):274-88
37 25 Figure 1.6. Schematic of the basic structure of lipopolysaccharide. LPS consists of three regions: from the bottom, lipid A (chair structure indicates di-glucosamine head group, red circles indicate phosphate groups, wavy lines indicate acyl chains),core sugars, and O-antigen, which consists of repeating units (denoted in brackets, with an n )of oligosaccharides. Reprinted via the Creative Commons Attribution (CC BY) license, Front Cell Infect Microbiol Feb 12;3:3

38 26 Figure 1.7. Type III Secretion System (T3SS). Schematic representation of components of T3S S, indicting conserved cytoplasmic, membrane spanning, and extracellular components of the apparatus. Reprinted with permission from the American Society of Microbiology, Microbiol Mol Biol Rev Jun;76(2):


39 27 Figure 1.8. Type VI Secretion System (T6SS). A) T6SS locus from B. bronchiseptica strain RB50. Each arrow corresponds to relative gene length. Homologues of T6SS core components are labeled. B) Model of assembled T6SS, indicating central tube and conserved components. C) Model of T6SS Apparatus Action. Reprinted with permission from Nature Publishing Group, Nature Feb 26;483(7388):182-6 and Creative Commons Attribution (CC BY) license, Front Microbiol Jul 18;2:155
40 28 Chapter 2 Enzymatic Modification of the Lipid A by ArnT protects B. bronchiseptica against Cationic Peptides and Is Required for Transmission.
41 29 Abstract Pathogen transmission cycles require many steps: initial colonization, growth and persistence, shedding, and transmission to new hosts. Alterations in the membrane components of the bacteria, including lipid A, the membrane anchor of lipopolysaccharide, could affect any of these steps via its structural role in protecting bacteria from host innate immune defenses, including antimicrobial peptides and signaling through TLR4. To date, lipid A has only been shown to affect the withinhost dynamics of infection rather than the between-host dynamics of transmission. We investigated the effects of lipid A modification in a mouse infection and transmission model. Disruption of the Bordetella bronchiseptica locus (BB4268) revealed that ArnT is required for addition of glucosamine (GlcN) to B. bronchiseptica lipid A. ArnT modification of lipid A did not change its TLR4 agonist activity in J774 cells, but deleting arnt decreased resistance to killing by cationic antimicrobial peptides, such as polymyxin B and β-defensins. In the standard infection model, mutation of arnt did not affect B. bronchiseptica colonization, growth, persistence throughout respiratory tract, recruitment of neutrophils to the nasal cavity, or shedding of the pathogen. However, the number of bacteria necessary to colonize a host (ID50) was five-fold higher for the arnt mutant. Furthermore, the arnt mutant was defective in transmission between hosts. These results revealed novel functions of the ArnT lipid A modification and highlight the sensitivity of low dose infections and transmission experiments for illuminating aspects of infectious diseases between hosts. Factors such as ArnT can have important effects on
42 30 the burden of disease and are potential targets for interventions that can interrupt transmission.
43 31 Introduction Lipopolysaccharide (LPS), the major component of the outer membrane of Gramnegative bacteria, is known to affect interactions with the host in a variety of ways that have been illuminated using host infection models. Upon initial contact with the host mucosa, LPS can protect pathogens from innate host defenses, such as complement and cationic antimicrobial peptides (CAMPs) (26). LPS is extracted from bacterial membranes by LPS binding protein (LBP), which then transfers the LPS to the CD14, a glycosyl-phosphatidylinositol-anchored protein (22). Then CD14 presents the LPS to the Toll-like receptor 4 (TLR4)-MD-2 complex, which recognizes the lipid A through interaction with hydrophobic regions in the binding pocket of MD-2 (16). The TLR4- MD-2 complex dimerizes and acts as a scaffold for MyD88 or TRIF (14), creating a signaling cascade that results in mobilization of the transcription factor NF-κB (16), induction of the expression of proinflammatory cytokines, such as TNF-α and IL-6 (33), as well as chemokines in cells of the innate immune system (13). TLR4 signaling also facilitates the recruitment of adaptive immune responses, particularly through the activation of dendritic cells (DC), which are induced by LPS to migrate to regional lymph nodes and present antigens to T cells (12). Lipid A-TLR4 interactions are therefore central to host-pathogen dynamics during infections by Gram-negative bacteria. Consequently, it is not surprising that pathogens regulate their lipid A structure through a number of covalent modifications which can affect interactions with host immunity (21, 26).
44 32 Bordetella bronchiseptica is a Gram-negative coccobacillus, closely related to B. pertussis and B. parapertussis and the causative agents of whooping cough in humans. B. bronchiseptica is highly infectious in mice, providing a model system in which the role of specific Bordetella virulence factors during infection can be probed in the context of a natural host infection (19). Adhesins, toxins, and other factors that enable B. bronchiseptica to thrive within the host are chiefly controlled by the two-component regulatory system, BvgAS (1, 20). These virulence-associated genes are expressed maximally in the Bvg + phase and transcriptionally repressed in the Bvg phase (4). Modifications of the lipid A of B. bronchiseptica are regulated by BvgAS (15, 17, 18). B. bronchiseptica lipid A consists of a glucosamine disaccharide backbone anchored to the bacterial outer membrane by a series of acyl groups (15) (Figure 2.1 C, D, E structure of the lipid A). Normally, B. bronchiseptica lipid A is penta-acylated with 3- OH C14 acyl groups at the 2 and 2 positions and a 3-OH C10 at the 3 position. The 3 position is empty due to the deacylase activity of the outer membrane enzyme PagL (15). Secondary or piggyback acylations at the 2 position are either a 2OH-C12 or C12 with the presence of 2OH-C12 dependent on the lipid A dioxygenase, LpxO (15). PagP is a Bvg-regulated lipid A palmitoyl transferase that adds palmitate as a secondary acylation at the 3 position, generating a hexa-acylated structure (24, 25). Finally, the major lipid A species contains a single phosphate at the C-4, although some molecules are also phosphorylated at the C-1 position (17). In monophosphorylated lipid A species, the C-4 phosphate is decorated with a GlcN molecule; however, in lipid A molecules that possess two phosphate groups, only one GlcN modification is observed at either the C-1 or the C-4 position. Orthologues of the lipid A modification enzyme, ArnT, which
45 33 decorates the lipid A phosphates of Salmonella typhimurium with aminoarabinose are conserved among Bordetella species (34). The modification of phosphate groups with aminoarabinose decreases the net negative charge on LPS and renders S. typhimurium resistant to the antimicrobial cationic peptide, polymyxin B (11). In B. pertussis, the ArnT activity of the homologue LgmB is induced in the Bvg + phase and mediates the addition of GlcN to both terminal phosphate groups of the lipid A, which is associated with increased stimulation of TLR4 activity in the HEK-Blue assay and upon infection of human macrophages (17, 18, 30). Deletion of arnt did not affect resistance to killing by polymyxin B (17). ArnT-mediated addition of GlcN has also been reported for B. parapertussis (9) and B. bronchiseptica strain 4650 (18). In this story, we characterized the function of the B. bronchiseptica arnt homologue, BB4268, by the construction and analysis of an arnt mutant in B. bronchiseptica strain RB50, henceforth referred to as RB50ΔarnT. Similar to its function in B. pertussis, we found that B. bronchiseptica arnt was required for Bvg + phasedependent addition of GlcN to the lipid A. No change in the TLR4 agonist activity of the mutant strain was observed; however, loss of resistance against polymyxin B and β- defensin mediated killing was detected. Loss of arnt had no effect on bacterial growth or persistence when bacteria were seeded throughout the respiratory tract by standard high dose inoculation; however, the arnt mutant was not transmitted between mice, even though the mutant was shed from index cases at the same level as wild type. Furthermore, RB50ΔarnT required approximately a 5-fold increase in mean inoculation dose compared to wild type to initiate infections. Together these results showed that deleting arnt had no observable effects in standard virulence and pathogenesis assays but
46 did affect LPS modification, which had a major impact on shedding and transmission of B. bronchiseptica. 34 Materials and Methods Bacterial strains and growth. Bordetella bronchiseptica strains RB50 (4), RB50Δwbm (2) and RB50ΔarnT were maintained on Bordet-Gengou agar (Difco) supplemented with 10% defibrinated sheep blood (Hema Resources) and 200 μg/ml streptomycin (Sigma-Aldrich) and cultured in Stainer-Scholte broth (3) at 37 C until grown to mid-log phase approximate OD600 of 0.5. For the mbd3 killing assay, E. coli K12 bacteria were grown in Luria Bertani broth at 37 C until mid-log phase. Mutation of BB4268. The genomic DNA template was made by resuspending several colonies of plategrown bacteria in 0.5 ml of water, boiling in a water bath for 5 min, spinning at top speed in a bench- top microcentrifuge for 2 minutes and taking 0.2 ml of supernatant. 1 μl of supernatant was used per PCR reaction. Each PCR reaction comprised genomic DNA template, buffer as directed by the manufacturer, dntps (25 mm each), 20 ng of each primer, 5% (v/v) DMSO, 5 mm MgCl2 and 2.5 units of TAQ DNA polymerase (Promega). Primers used to amplify an approximately 1 kb section of B. bronchiseptica 4268 were 5 ATGTAGCCGACCAGCTTG 3 and 5 ATCCATGCAACCCCATGC 3, corresponding to bases and respectively, of the B. bronchiseptica RB50 genome sequence, Genbank accession number BX (23).
47 35 PCR reactions were incubated at 94 C for 5 min followed by 30 cycles of 94 C for 75 s, 60 C for 75 s and 72 C for 90 s, followed by a final step of 72 C for 7 min. The PCR product was cloned into pgem-t Easy (Promega) according to the manufacturer s instructions. A non-polar kanamycin resistance cassette was ligated into a unique StuI site residing in the middle of the cloned BB4268 region. The resulting BB4268-kan region was subcloned into pex100t (29) and the resulting construct was moved into the conjugation donor strain SM10 (31) by transformation. Bacterial conjugations were performed as described previously (2). The expected chromosomal rearrangements in conjugants were confirmed by Southern hybridization analyses. B. bronchiseptica arnt was amplified by PCR using primers incorporating NdeI and HindIII restriction endonuclease recognition sites at the 5 and 3 ends of the amplicon respectively. Following digestion of the PCR product with NdeI and HindIII, this fragment was cloned behind the B. bronchiseptica pagp promoter into pbbrkanpagp (25), replacing the pagp CDS in this construct. The arnt containing construct was moved into wild type B. bronchiseptica and the B. bronchiseptica arnt mutant by conjugation as described (25). Lipid A purification. LPS was purified from 1 liter, overnight B. bronchiseptica cultures as described previously (35). Briefly, the bacteria were harvested, lyophilized, and treated with DNase I and proteinase. The bacteria were then boiled, and LPS was extracted by collecting the phenol fraction after using the hot phenol-water method. Further treatment of LPS with RNase A, DNase I, and proteinase K ensured removal of contaminating
48 36 nucleic acids and proteins (7). Hydrolysis of LPS to isolate lipid A was accomplished with 1% sodium dodecyl sulfate (SDS) at ph 4.5 as described (3). Confirmation of lipid A structures. The lipid A structures were confirmed by matrix-assisted laser desorption ionization time of flight (MALDI-TOF) mass spectrometry. LPS was isolated using a rapid small-scale isolation method (36). Cell culture pellets (1 10 ml of an overnight culture) were resuspended in a 1.0 ml aliquot of TriReagent (Molecular Research Center; Cincinnati, OH, USA) and incubated at room temperature for 15 min. Chloroform (200 ml) was added and the samples were vortexed and incubated at room temperature for 15 min. Samples were centrifuged for 10 min at 13,400xg and the aqueous layers were collected. Water (500 ml) was added to the lower layer and vortexed. After 30 min, the samples were centrifuged as above and the aqueous layers were pooled. Two more aliquots of water were added to each sample for a total of four extractions. The combined aqueous layers were frozen and lyophilized. The LPS was then hydrolyzed to lipid A. Lyophilized LPS was resuspended in 0y.5 ml 1% sodium dodecyl sulfate (SDS) in 10 mm sodium acetate buffer, ph 4.5 (3). Samples were incubated at 100C for 1 h, frozen, and lyophilized. The dried pellets were washed in 0.1 ml of water and 1 ml of acidified ethanol (100 ml 4 N HCl in 20 ml 95% EtOH). Samples were centrifuged at 2300xg for 5 min and the supernatant discarded. The lipid A pellet was further washed (twice for a total of three washes) in 1 ml of 95% EtOH. The entire series of washes was repeated twice. A final wash step was carried out in 100% ethanol. Lipid A was extracted in a mixture of chloroform, methanol, and water (3:1:0.25, vol/vol/vol). One microliter of this extract was then spotted onto a MALDI
49 37 target plate followed by 1 ul of Norharmane matrix and air dried. Samples were analyzed on a Bruker AutoFlex Speed (Bruker Daltonics, Billerica, MA) mass spectrometer, which was calibrated using Agilent Tuning Mix (Agilent Technologies, Foster City, CA). In vitro Macrophage Stimulation and TNFα Detection. J774 murine macrophages were cultured in Dulbecco s modified Eagle s medium (DMEM, Difco) supplemented with 10% fetal bovine serum, 1% penicillin-streptomycin, 1% nonessential amino acids, and 1% sodium pyruvate weight/volume. The cells were grown to approximately 85% confluency in a 96 well plate. In order to compare TNFα secretion, J774 cells were inoculated with RB50 or RB50ΔarnT at an MOI of 0.001, 0.01, 0.1 or 1. Medium was harvested 2 hours post-inoculation, and an ELISA to detect TNFα concentrations was performed according to the manufacturer s instructions (R&D Systems). These assays were performed two times in quadruplicate. Adherence Assay. Rat epithelial (L2) cells were grown to 80% confluence in 96 well plates using Dulbecco s modified Eagle medium (DMEM, Difco)-F-12 medium supplemented with 10% fetal bovine serum. These cells were inoculated with 10 4 CFU of RB50 or RB50ΔarnT (MOI 0.2). Plates were then centrifuged for 5 min at 250 g, followed by incubation at 37 C with 5% CO2 for 40 min. Wells were then washed four times with 1 ml of the growth medium to remove non-adherent bacteria. L2 cells were then treated with 0.5 ml of 0.125% trypsin (Sigma Aldrich), followed by incubation for 10 min at 37 C. The total volume of each well was brought up to 1 ml with growth medium and homogenized by pipetting. Dilutions were plated on BG plates containing 40 μg of
50 38 streptomycin/ml to determine CFU counts, which were then used to calculate the proportion of adherent bacteria, expressed as a percentage of the original inoculum. Serum Killing Assay. Approximately 10 3 CFU RB50, RB50Δwbm (O-antigen mutant), or RB50ΔarnT in 50 µl of PBS were incubated with serum from mice naïve to Bordetella, at concentrations ranging from 0 15% solution by volume. After 1 h incubation at 37 C followed by 5 min incubation on ice, the entire 50 μl sample was plated onto Bordet- Gengou blood agar containing streptomycin (20 μg/ml). Colonies were enumerated after 2-days of incubation at 37 C. This assay was performed two times in quadruplicate. β-defensin Killing Assay CFU RB50 and RB50ΔarnT were incubated with 0, 5, and 10 μg/ml of synthetic pbd-1 (6) and mbd3 (R&D Systems) in 100 µl of PBS at 37 C for 2 hours after which the reaction mixture was plated on BG agar and incubated at 37 C for 2 days to calculate CFU. For the mbd3 assay, E. coli K12 bacteria were used as a positive control. Polymyxin B Susceptibility Assay. Cultures of RB50 or RB50ΔarnT were diluted to 10 6 CFU/mL into a final 1mL volume of PBS, PBS containing 10mg/mL polymyxin B, or PBS containing 100 mg/ml of polymyxin B. Suspensions were incubated for 2 hours at 37 C, following which, the number of organisms remaining in each sample was determined by quantitative culture on BG agar plates.
51 39 Colonization studies. C57BL/6 (wild type), and C3H/HEJ (TLR4-deficient) mice were obtained from Jackson Laboratories and bred in our Bordetella-free, specific pathogen-free facilities at The Pennsylvania State University. Bacteria grown overnight to an optical density at 600 nm of approximately 0.3 in liquid culture were diluted in PBS to approximately 2 x 10 6 CFU/mL. For a high dose/ high volume inoculation, 50 µl of the inoculum (10 4 CFU) was pipetted onto the external nares of 4 6 week old mice that had been lightly sedated with 5% isoflurane in oxygen. For low dose/low volume inoculations, bacterial cultures were further diluted in PBS to concentrations of 10 3 CFU/mL and for high dose/low volume to 4 x 10 4 CFU/mL. Mice were inoculated with doses of 5 CFU and 200 CFU respectively in 5μL using the previously described procedure. Groups of three or four animals were sacrificed on days 3, 7, 14, and 28 post-inoculation or as indicated, and the nasal cavity, trachea, and lungs were excised. Bacterial numbers in the respiratory tract were quantified by homogenization of each tissue in PBS followed by plating onto Bordet-Gengou blood agar containing streptomycin (20 μg/ml). Colonies were enumerated after 2 days of growth at 37 C. All protocols were reviewed and approved by The Pennsylvania State University Institutional Animal Care and Use Committee (IACUC), and all animals were handled in accordance with institutional guidelines. Shedding Analysis. Shedding was assessed by lightly swabbing the external nares for 10 seconds using a Dacron-polyester tipped swab. Swab tips were cut off and placed into 1 ml of PBS. Samples were vortexed vigorously and cultured on Bordet Gengou agar (Himedia).
52 40 Analysis of Leukocyte Recruitment. Prior to dissection, ml of PBS was perfused through the left ventricle of mice while venous runoff was collected from the orbit. Nasal bones were dissected and placed in 1 ml of DMEM containing 5% FBS and 1 mg/ml collagenase D. Samples were incubated for 45 min at 37 o C and subsequently disaggregated into a single cell suspension by mechanical disruption over a 70 µm mesh screen. Subsequently, 2x10 6 cells per well were then added to 96 well plates. Samples were resuspended in FC blocking buffer (200:1 anti-cd16/32 BD Biosciences in PBS + 2% FBS) and incubated on ice for 20 min. Following wash, cell surface markers were labeled with the following antibodies in PBS + 2% FBS: anti-cd45 APC-cy7 400:1 (BD Biosciences), anti-cd11b Horizon V450 (BD Biosciences), anti-ly6g APC (E Bioscience). Statistical Analysis. Data analysis between groups was performed using a One-Way Analysis of Variance test to evaluate statistical significance with p values <0.05 considered significant. The Reed Muench calculation was used for determination of ID50 (27). Results ArnT Is Required for Modification of Lipid A with Glucosamine. To investigate the role of ArnT enzymatic activity in GlcN modification of lipid A, the B. bronchiseptica RB50 arnt homologue, previously designated BB4268, was mutated to generate strain RB50ΔarnT. Purified lipid A isolated from RB50 or RB50ΔarnT was analyzed by matrix-assisted laser desorption ionization time of flight
53 41 (MALDI-TOF) mass spectrometry in the negative ion mode. Bordetella bronchiseptica lipid A had previously been characterized by Preston et al. (25). Peaks indicating a glucosamine addition were present at m/z 1651 and 1667, representing the monophosphorylated penta-acylated lipid A with a two acyl-oxo-acyl C12 or 2OH-C12 respectively, and m/z 1731, corresponding to the 1651 species with diphosphate additions (Figure 2.1A). The presence of the palmitate in acyl-oxo-acyl linkage at the C3 position was previously characterized and shown to be facilitated by the B. bronchiseptica PagP palmitoyl transferase (25). The lipid A form containing the phosphate addition was previously confirmed to be heterogeneously present in the bacterial population (15). Both of these peaks were entirely absent in the RB50ΔarnT spectrum (Figure 2.1B). Complementation with the arnt gene restored the phenotype (data not shown), demonstrating that ArnT was required for addition of glucosamine to Bordetella bronchiseptica lipid A. All other characteristic peaks were observed for both wild type and RB50ΔarnT. TLR4 Stimulation in Murine Macrophages, Serum Resistance and Adhesion Were Not Affected by arnt Mutation. ArnT-mediated modification of lipid A has been shown to enhance stimulation of TLR4 by B. pertussis LPS (17). To determine if the increased TLR4 agonist activity of B. bronchiseptica LPS was dependent on GlnN addition, cultures of murine macrophages (J774) were inoculated with either wild type strain RB50 or the arnt mutant. Release of the pro-inflammatory cytokine TNFα into the medium was used as a surrogate measurement of TLR4 receptor activity. Cultures were inoculated with MOIs ranging from to 1 and the concentration of TNFα in the medium was determined 2 hours
54 42 after inoculation by quantitative ELISA (Figure 2.2A). The amount of TNFα detected in the supernatant of cells exposed to B. bronchiseptica increased in a dose dependent manner; however, at each MOI, the amount of TNFα released by macrophages was similar between RB50 and RB50ΔarnT inoculated cell cultures. These data suggest that agonist activity of lipid A for TLR4 was not affected by ArnT-mediated addition of glucosamine. B. bronchiseptica is protected from the antimicrobial activities of serum complement by its LPS; strains lacking either O-antigen, such as (2), or the outer core oligosaccharide and O-antigen portions of LPS are highly susceptible to killing by serum complement. The B. bronchiseptica RB50 mutant with a deletion of wbm, a locus necessary for the assembly of the O antigen, lacks this structure and has the aforementioned defect in resistance to complement-mediated killing (2). To determine whether mutation of arnt affected serum resistance, approximately 10 3 CFU of RB50, RB50Δwbm, or RB50ΔarnT were incubated with concentrations of serum ranging from 0-15% (by volume). Whereas RB50Δwbm was killed by 5% serum, serum concentrations of up to 15% had no effect on either wild type or arnt mutant bacteria, demonstrating that mutation of arnt did not affect serum resistance (Figure 2.2B). Loss of the GlcN substitution might alter the structure of the outer membrane and destabilize interactions that facilitate bacterial adherence to respiratory epithelial cells. To test this, L2 rat lung epithelial cells were inoculated with between 5 1,000 CFU of the wild type strain or RB50ΔarnT. No significant difference was observed in the number of wild type or mutant bacteria recovered from the trypsin treated epithelial cells
55 43 (Figure 2.2C). This result suggests that binding of RB50 to the respiratory epithelium was not affected by deletion of arnt. Glucosamine Additions to Lipid A Contribute to Resistance against Cationic Antimicrobial Peptides. Host epithelial cells and leukocytes produce small cationic peptides, defensins, and cathelicidins that exhibit antimicrobial activity against numerous bacterial species (5, 8, 32). Defensins are amphipathic molecules proposed to intercalate into the bacterial cell membrane and use like-charge repulsion to disrupt membrane integrity (8). Disruption of the arnt locus rendered S. typhimurium susceptible to killing by cationic peptides. To test whether arnt provided a similar adaptation for B. bronchiseptica, we compared the ability of RB50 and RB50ΔarnT to survive in the presence of varying concentrations of antimicrobial peptides. RB50 was resistant to killing by polymyxin B at concentrations up to 100 µg/ml. However, greater than 90% of RB50ΔarnT were killed by 10 µg/ml, and 99.99% of the mutant bacteria were killed by 100 μg/ml of polymyxin B (Figure 2.3A). In addition, wild type RB50 survived a concentration of 10 µg/ml of the porcine β-defensin 1 (pbd1), whereas more than 99% of RB50ΔarnT were killed by 5 µg/ml of pbd1. Surprisingly, RB50 and the arnt mutant showed no significant difference in susceptibility to killing by mouse β-defensin 3, which shares homology with pbd1 (Figure 2.3C). Our results suggested that modification of lipid A by GlcN rendered RB50 more resistant to killing by some, but not all, antimicrobial cationic peptides.
56 44 B. bronchiseptica arnt Is Not Necessary for Infection of the Mouse Respiratory Tract. To determine whether decreased resistance to CAMPs would result in reduced fitness within a host, we compared the ability of the RB50 and the arnt mutant to grow and persist during experimental infections of mice. Following inoculation of C57BL/6 mice with 10 4 CFU of either RB50 or RB50ΔarnT in 50 µl of PBS, mice were dissected after 3, 7, 14 and 28 days. At each time point the number of CFU recovered from the respiratory tract of mice inoculated with RB50ΔarnT were similar to that recovered from RB50 inoculated mice (Figure 2.4). This result suggested that addition of GlcN to the lipid A was not required for growth and persistence of B. bronchiseptica in the murine respiratory tract when introduced in a high dose inoculum. RB50ΔarnT Is Unable to Colonize Mice When Inoculated in Low Dose. Although RB50ΔarnT grew and persisted similarly to the wild type strain following high dose inoculation challenge, we hypothesized that high dose infections may not accurately reproduce the interactions taking place when B. bronchiseptica initially seeds the respiratory mucosa. For example, CAMPs may not be sufficiently potent to control the initial influx of infectious organisms, or the epithelial cells that produce CAMPs may be rapidly damaged by bacterial toxins. A functional defect resulting from decreased CAMP resistance could therefore be more effectively determined using low dose inoculations. To approximate a mean infectious dose, 5, 50 or 200 CFU of either RB50 or RB50ΔarnT were inoculated in a 5 µl droplet onto the external nares of C57BL/6 mice. Seven days post- inoculation, the presence of B. bronchiseptica in the nasal cavity was determined (Figure 2.5A). Five CFU of wild type
57 45 RB50 was sufficient to infect 5 of 10 mice whereas 50 CFU resulted in stable colonization of 11/12 mice. Additionally, 12/12 mice became infected following inoculation with 200 CFU of RB50. By comparison, 5 CFU RB50ΔarnT infected only 1/16 mice, 50 CFU infected 4/12 mice and 200 CFU infected 12/ 12 mice (Figure 2.5B). Using the Reed Muench calculation (27), we determined that the ID50 of wild type RB50 was approximately 6.2 CFU whereas the ID50 of RB50ΔarnT was 30 CFU. RB50ΔarnT Fails to Transmit between Mice. Although the presence of ArnT did not affect the infectious burden following high dose inoculation, subtle functional deficiency resulting from mutation of arnt would be more likely resolved by analysis of transmission efficiency. We have previously demonstrated that RB50 transmits efficiently between C3H/HeJ mice (28). To evaluate whether RB50ΔarnT can be transmitted between animals, mice were inoculated with 500 CFU of RB50 or RB50ΔarnT. Inoculated individuals (index mice) were placed in a cage with 2-3 naïve mice (secondary mice). After co-housing for 21 days, mice were dissected and bacterial numbers in the nasal cavity were determined (Figure 2.6). Five out of 6 secondary mice exposed to RB50-inoculated index mice became colonized, whereas none of the secondary mice exposed to RB50ΔarnT-infected index mice became colonized (Figure 2.6). RB50ΔarnT Is not Defective at Being Shed by the Host. Although the inability of RB50ΔarnT to be transmitted corresponded with an increased ID50, this did not exclude the possibility that RB50ΔarnT might also be shed at a reduced rate. We have demonstrated that the shedding intensity of index mice correlated with the probability of transmission to other hosts (28). To determine whether
58 46 RB50ΔarnT was shed at a lower rate as compared to wild type RB50, C57BL/6 mice were inoculated with 500 CFU of either RB50 or RB50ΔarnT. Shedding was monitored by swabbing the external nares at multiple points over the course of 14 days. Neutrophil recruitment to the nasal cavity, previously shown to correlate with shedding intensity (10) was also determined at 7 and 14 days after inoculation. Shedding intensity of RB50ΔarnT was not different from that of RB50 at any time over a 14 day course (Figure 2.7B). Furthermore, neutrophil numbers in the nasal cavity of mice inoculated with RB50ΔarnT were similar to those obtained from RB50 inoculated mice at day 7 and day 14 post-inoculation (Figure 2.7A). These results demonstrate that ArnT is not required to induce neutrophil recruitment or enhance bacterial shedding from the murine nasal cavity. Discussion Phenotypic assessment of an arnt-deficient derivative of B. bronchiseptica strain RB50 showed that this gene was required for the modification of lipid A with GlcN (15). Deletion of arnt was associated with decreased resistance to killing mediated by cationic antimicrobial peptides (CAMPs). ArnT did not affect the growth or persistence within the host but was required for transmission. The failure of mice to transmit RB50ΔarnT corresponded with a five-fold increase in the mean infectious dose. Together these results suggested that ArnT-mediated addition of glucosamine to lipid A confers resistance to certain CAMPs and increases the frequency with which pathogens that seed the nasal mucosa are able to successfully colonize and initiate infections.
59 47 The protein encoded by arnt shares homology with Salmonella ArnT, a periplasmic enzyme that catalyzes the transfer of aminoarabinose from an undecaprenyl donor to the phosphate groups of lipid A and results in resistance to CAMPs (34). This protein is 100% identical at the amino acid level to the B. pertussis ArnT homologue, encoded by the gene designated BP0398. While the B. pertussis ArnT homologue mediates substitution of lipid A by GlcN (18), this activity was not associated with decreased resistance to CAMP-mediated attack but rather with increased TLR4 agonist potency (11). Deletion of the B. bronchiseptica ortholog did not alter the TLR4 stimulatory activity of the bacteria, suggesting that ArnT activity is not required for maximal stimulation of TLR4 by B. bronchiseptica. The differences between the observed effects of ArnT in B. bronchiseptica and B. pertussis may relate to their relative TLR4 stimulatory activity, which differ by an order of magnitude. The other structural differences between the LPS (actually LOS for B. pertussis) of these two organisms, and the basis for these differences, remain to be elucidated. B. bronchiseptica is strongly resistant to the bactericidal activity of porcine β- defensin 1 (6). In the absence of arnt, B. bronchiseptica is more susceptible to the bactericidal effects of polymyxin B and pbd1. These results are analogous to findings reported for deletion mutants of arnt in S. typhimurium. As with 4-amino L-arabinose, substitution of the phosphate group by GlcN could decrease the anionic character of the outer membrane. The charge neutralizing effect of GlcN may therefore represent an adaptation that destabilizes CAMP binding within the membrane and thus mitigates its activity. It is unclear why RB50ΔarnT is more susceptible to killing by pbd1 but is unaffected in its sensitivity to mbd3. Although CAMPs are hypothesized to have a
60 48 common mechanism of action based on conserved structural motifs, the multitude of genes encoding CAMPs in eukaryotic genomes, and the divergence of sequences from one host species to another suggests that individual CAMPs may have some specificity regarding the microbes they target and/or their mechanisms of action (8). Alternatively, it is possible that mbd3 is not highly active on its own but acts in cooperation with other antimicrobial mechanisms in vivo. Since ArnT activity is increased in the Bvg + phase, which is thought to contribute to growth during infection, one would expect this gene to contribute to fitness within the host. When comparing RB50 and RB50ΔarnT in mouse infections, differences between the two strains were observed upon titration of the infectious dose toward the lower limits. When mice were inoculated with RB50ΔarnT in a volume and dose sufficient to seed the entire respiratory tract with high bacterial numbers, no difference between the wild type parental strain and the arnt mutant was detected. When comparing the fitness of the wild type and the mutant introduced in very low numbers (50 CFU or less), the mean infectious dose required to stably infect 50% of mice (ID50) with RB50ΔarnT was approximately five-fold greater than the ID50 of the wild type strain. These data suggest that the likelihood of B. bronchiseptica successfully colonizing the respiratory tract of a new host is greatly enhanced by ArnT. The lipid A modification mediated by ArnT is likely an adaptation to host antimicrobial defenses, which while effective against small numbers of pathogens, may be overwhelmed by increased doses of bacteria. When pathogens seed mucosal surfaces and begin the process of invasion and colonization, CAMP molecules, in particular β-defensins, are among the first elements of host resistance encountered. Resistance to CAMP killing may play a role in the survival of
61 49 bacteria during the initial colonization process. However, due to the number of genes encoding CAMPs, knockout systems may not be a practical way of demonstrating that ArnT activity enables B. bronchiseptica to colonize mice by conferring resistance to CAMP mediated attack. Our results indicate that ArnT mediated GlcN modification of lipid A and contributes significantly to the infectiousness and transmissibility of B. bronchiseptica. We also demonstrated that directly quantifying transmission and infectivity using low dose infection models can be a more sensitive approach for probing interactions that are important for the initial colonization of new hosts, aspects not clearly observed when higher dose inocula are delivered. Whether this inocula-based difference occurs because important host defense measures are overwhelmed or because of some other mechanism remains to be determined. However, an approach focusing on lower, more natural, doses and on transmission between animals, revealed phenotypes dependent on ArnT that are likely to have important implications to the spread of infection, and therefore the burden of this infectious disease at the population scale.
62 50 Author Contributions Olivier Rolin 1,2*, Sarah J. Muse 1,3*, Chetan Safi 1, Shokrollah Elahi 4, Volker Gerdts 5, Lauren E. Hittle 6, Robert K. Ernst 6, Eric T. Harvill 1, and Andrew Preston 7 1 Department of Veterinary and Biomedical Sciences, The Pennsylvania State University, University Park, PA USA. 2 Graduate Program in Immunology and Infectious Disease. 3 Graduate Program in Biochemistry, Microbiology and Molecular Biology. 4 Seattle Biomedical Research Institute, Seattle, WA USA. 5 Vaccine and Infectious Disease Organization, University of Saskatchewan, Saskatoon, SK Canada. 6 Department of Microbial Pathogenesis, University of Maryland School of Dentistry, Baltimore, MD, USA. 7 Department of Biology and Biochemistry, University of Bath, Bath, U.K. * Contributed equally to this manuscript Conceived and designed experiments: OR, SJM, CS, SE, VG, LEH, RE, ETH, AP Performed experiments: OR, SJM, CS, SE, LEH, AP Analyzed the data: OR, SJM, CS, SE, VG, LEH, RE, AP Wrote the paper: OR, SJM, ETH, AP
63 51 References 1. Arico, B., V. Scarlato, D. M. Monack, S. Falkow, and R. Rappuoli Structural and genetic analysis of the bvg locus in Bordetella species. Mol Microbiol 5: Burns, V. C., E. J. Pishko, A. Preston, D. J. Maskell, and E. T. Harvill Role of Bordetella O antigen in respiratory tract infection. Infect Immun 71: Caroff, M., A. Tacken, and L. Szabo Detergent-accelerated hydrolysis of bacterial endotoxins and determination of the anomeric configuration of the glycosyl phosphate present in the "isolated lipid A" fragment of the Bordetella pertussis endotoxin. Carbohydr Res 175: Cotter, P. A., and J. F. Miller BvgAS-mediated signal transduction: analysis of phase-locked regulatory mutants of Bordetella bronchiseptica in a rabbit model. Infect Immun 62: Doss, M., M. R. White, T. Tecle, and K. L. Hartshorn. Human defensins and LL-37 in mucosal immunity. J Leukoc Biol 87: Elahi, S., R. M. Buchanan, S. Attah-Poku, H. G. Townsend, L. A. Babiuk, and V. Gerdts The host defense peptide beta-defensin 1 confers protection against Bordetella pertussis in newborn piglets. Infect Immun 74: Fischer, W., H. U. Koch, and R. Haas Improved preparation of lipoteichoic acids. Eur J Biochem 133: Ganz, T Defensins: antimicrobial peptides of vertebrates. C R Biol 327: Geurtsen, J., M. Dzieciatkowska, L. Steeghs, H. J. Hamstra, J. Boleij, K. Broen, G. Akkerman, H. El Hassan, J. Li, J. C. Richards, J. Tommassen, and P. van der Ley Identification of a novel lipopolysaccharide core biosynthesis gene cluster in Bordetella pertussis, and influence of core structure and lipid A glucosamine substitution on endotoxic activity. Infect Immun 77: Gopinath, S., A. Hotson, J. Johns, G. Nolan, and D. Monack. The systemic immune state of super-shedder mice is characterized by a unique neutrophildependent blunting of TH1 responses. PLoS Pathog 9:e Herrera, C. M., J. V. Hankins, and M. S. Trent. Activation of PmrA inhibits LpxT-dependent phosphorylation of lipid A promoting resistance to antimicrobial peptides. Mol Microbiol 76: Jakubzick, C., F. Tacke, J. Llodra, N. van Rooijen, and G. J. Randolph Modulation of dendritic cell trafficking to and from the airways. J Immunol 176:
64 13. Kawai, T., and S. Akira. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol 11: Lu, Y. C., W. C. Yeh, and P. S. Ohashi LPS/TLR4 signal transduction pathway. Cytokine 42: MacArthur, I., J. W. Jones, D. R. Goodlett, R. K. Ernst, and A. Preston. Role of pagl and lpxo in Bordetella bronchiseptica lipid A biosynthesis. J Bacteriol 193: Maeshima, N., and R. C. Fernandez. Recognition of lipid A variants by the TLR4-MD-2 receptor complex. Front Cell Infect Microbiol 3: Marr, N., A. M. Hajjar, N. R. Shah, A. Novikov, C. S. Yam, M. Caroff, and R. C. Fernandez. Substitution of the Bordetella pertussis lipid A phosphate groups with glucosamine is required for robust NF-kappaB activation and release of proinflammatory cytokines in cells expressing human but not murine Toll-like receptor 4-MD-2-CD14. Infect Immun 78: Marr, N., A. Tirsoaga, D. Blanot, R. Fernandez, and M. Caroff Glucosamine found as a substituent of both phosphate groups in Bordetella lipid A backbones: role of a BvgAS-activated ArnT ortholog. J Bacteriol 190: Mattoo, S., and J. D. Cherry Molecular pathogenesis, epidemiology, and clinical manifestations of respiratory infections due to Bordetella pertussis and other Bordetella subspecies. Clin Microbiol Rev 18: Merkel, T. J., S. Stibitz, J. M. Keith, M. Leef, and R. Shahin Contribution of regulation by the bvg locus to respiratory infection of mice by Bordetella pertussis. Infect Immun 66: Miller, S. I., R. K. Ernst, and M. W. Bader LPS, TLR4 and infectious disease diversity. Nat Rev Microbiol 3: Miyake, K Innate immune sensing of pathogens and danger signals by cell surface Toll-like receptors. Semin Immunol 19: Parkhill, J., M. Sebaihia, A. Preston, L. D. Murphy, N. Thomson, D. E. Harris, M. T. Holden, C. M. Churcher, S. D. Bentley, K. L. Mungall, A. M. Cerdeno-Tarraga, L. Temple, K. James, B. Harris, M. A. Quail, M. Achtman, R. Atkin, S. Baker, D. Basham, N. Bason, I. Cherevach, T. Chillingworth, M. Collins, A. Cronin, P. Davis, J. Doggett, T. Feltwell, A. Goble, N. Hamlin, H. Hauser, S. Holroyd, K. Jagels, S. Leather, S. Moule, H. Norberczak, S. O'Neil, D. Ormond, C. Price, E. Rabbinowitsch, S. Rutter, M. Sanders, D. Saunders, K. Seeger, S. Sharp, M. Simmonds, J. Skelton, R. Squares, S. Squares, K. Stevens, L. Unwin, S. Whitehead, B. G. Barrell, and D. J. Maskell Comparative analysis of the genome sequences of Bordetella pertussis, Bordetella parapertussis and Bordetella bronchiseptica. Nat Genet 35: Pilione, M. R., E. J. Pishko, A. Preston, D. J. Maskell, and E. T. Harvill pagp is required for resistance to antibody-mediated complement lysis during Bordetella bronchiseptica respiratory infection. Infect Immun 72:
65 25. Preston, A., E. Maxim, E. Toland, E. J. Pishko, E. T. Harvill, M. Caroff, and D. J. Maskell Bordetella bronchiseptica PagP is a Bvg-regulated lipid A palmitoyl transferase that is required for persistent colonization of the mouse respiratory tract. Mol Microbiol 48: Raetz, C. R., C. M. Reynolds, M. S. Trent, and R. E. Bishop Lipid A modification systems in gram-negative bacteria. Annu Rev Biochem 76: Reed, L. J., Muench, H A Simple Method of Estimating Fifty Per Cent Endpoints. Am J Epidemiol 27: Rolin, O., W. Smallridge, M. Henry, L. Goodfield, D. Place, and E. T. Harvill. Toll-like receptor 4 limits transmission of Bordetella bronchiseptica. PLoS One 9:e Schweizer, H. P., and T. T. Hoang An improved system for gene replacement and xyle fusion analysis in Pseudomonas aeruginosa. Gene 158: Shah, N. R., S. Albitar-Nehme, E. Kim, N. Marr, A. Novikov, M. Caroff, and R. C. Fernandez. Minor modifications to the phosphate groups and the C3' acyl chain length of lipid A in two Bordetella pertussis strains, BP338 and , independently affect Toll-like receptor 4 protein activation. J Biol Chem 288: Simon, R., U. Priefer, and A. Puhler A Broad Host Range Mobilization System for In Vivo Genetic Engineering: Transposon Mutagenesis in Gram Negative Bacteria. Nat Biotech 1: Song, J., B. L. Bishop, G. Li, M. J. Duncan, and S. N. Abraham TLR4- initiated and camp-mediated abrogation of bacterial invasion of the bladder. Cell Host Microbe 1: Takeuchi, O., and S. Akira. Pattern recognition receptors and inflammation. Cell 140: Trent, M. S., A. A. Ribeiro, S. Lin, R. J. Cotter, and C. R. Raetz An inner membrane enzyme in Salmonella and Escherichia coli that transfers 4- amino-4-deoxy-l-arabinose to lipid A: induction on polymyxin-resistant mutants and role of a novel lipid-linked donor. J Biol Chem 276: Westphal, O., Jann, J.K Bacterial lipopolysaccharides: extraction with phenol-water and further applications of the procedure. Methods Carbohydr. Chem. 5: Yi, E. C., and M. Hackett Rapid isolation method for lipopolysaccharide and lipid A from gram-negative bacteria. Analyst 125:


 (B) RB50ΔarnT mutant strain no glucosamine peaks are present.")
66 54 Figure 2.1. Ions corresponding to glucosamine additions in RB50 are not seen in RB50ΔarnT.. Mass spectra of (A) B. bronchiseptica RB50, ions corresponding to glucosamine additions mass-to-charge ratio (m/z) of 1651, 1667, and 1731 are indicated (*) (B) RB50ΔarnT mutant strain no glucosamine peaks are present. Lipid A structures corresponding to m/z 1570 (C), 1808 (D) and 1651 (E).
67 55 Figure 2.2. ArnT was not required for induction of TNF TNFα in murine macrophages, complement resistance, or adherence to the lung epithelial cells. (A) J774 macrophages were incubated for 2 hours in the presence of RB50 (white bars) or RB50ΔarnT (black bars) at an MOI of 0.001, 0.01, 0.1 or 1.0. Grey bars represent cultures treated with media only. (B) 10 3 CFU of either B. bronchiseptica strain RB50 (white squares), RB50ΔarnT (black diamonds) or RB50Δwbm (black circles) were incubated for 1 hour in PBS only or PBS with 5, 10 or 15 percent naive mouse serum. Symbols represent the mean log10 CFU ± standard error of four individual samples recovered after incubation. The limit of detection is marked by a dashed line. (C) Increasing MOI of RB50 (white squares) or RB50ΔarnT (black diamonds) were inoculated onto cultured L2 rat lung epithelial cells. The quantity of CFU of RB50 (white squares) or RB50ΔarnT (black diamonds) adhering to epithelial cells following incubation was determined by culture. Symbols represent the mean CFU ± standard error of four individual samples.
68 56 Figure 2.3. ArnT was required for resistance to killing by cationic antimicrobial peptides CFU of bacteria were incubated for 2 hours in PBS (white bars) or PBS with increasing concentrations (striped bars, black bars) of (A) polymyxin B (B) pbd1 or (C) synthetic mbd3. B. bronchiseptica strains RB50 or RB50ΔarnT were incubated with mbd3, or E coli K12, whose susceptibility to mbd3 was previously shown, was used as a control to determine whether mbd3 was active (B). Bars represent the mean log10 CFU ± standard error of 4 individual samples in each group. * indicates a p value of <0.05, ** indicates a p value of <0.01. The limit of detection is marked by a dashed line.
69 57 Figure 2.4. ArnT did not contribute to growth or resistance in the respiratory tract in a high dose model of respiratory infection. Groups of 4 mice were inoculated with 5x10 4 CFU of RB50 (white squares) or RB50ΔarnT (black diamonds) in 50 µl of PBS. Symbols represent the mean log10 CFU ± standard error recovered from either the (A) nasal cavity, (B) trachea, or (C) lungs at 3, 7, 14 and 28 days following inoculation. The limit of detection is marked by a dashed line.
70 58 Figure 2.5. ArnT decreased the mean infectious dose of B. bronchiseptica. (A) C57BL/6 Mice were inoculated with 5 CFU in 5 µl of either RB50 (left) or RB50ΔarnT (right). Dots represent the CFU recovered from the nasal cavity of individual mice dissected seven days after inoculation. The whiskers span the 1 st and 4 th quartile. The limit of detection is marked by a dashed line. The proportion of infected mice in each group is listed above the box and whisker plot. (B) Percent of C57BL/6 mice infected after inoculation with either 5 CFU (10 mice/group), 50 CFU (12 mice/group) or 200 CFU (16 mice/group) of RB50 (white squares) or RB50ΔarnT (black diamonds).

71 59 Figure 2.6. ArnT was required for transmission of B. bronchiseptica between mice. Two index C3H/HeJ mice per group were inoculated with 500 CFU of either RB50 or RB50ΔarnT in 5μl PBS. Index mice were housed with 2-3 secondary mice for 3 weeks following inoculation. The quantity of CFU of RB50 (left) or RB50ΔarnT (right) recovered from the nasal cavity of individual secondary mice is represented by dots. Box spans the interquartile range, and the median value is represented by a bar. The whiskers span the 1 st and 4 th quartile. The proportion of infected mice in each group is listed above the box and whisker plot. The limit of detection is marked by a dashed line.
72 60 Figure 2.7. ArnT did not affect neutrophil recruitment or shedding from the host. C57BL/6 mice were infected with 500 CFU in 5 µl PBS of either RB50 or RB50ΔarnT. (A) Mice were sacrificed at either 7 or 14 days after inoculation and the mean ± standard error neutrophil counts from nasal cavities of mice infected with RB50 (white bars) or RB50ΔarnT (black bars) were obtained by flow cytometric detection of CD45 + / CD11b + / Ly6G + cells. (B) Shedding was detected by culture of bacteria from a 10 second swab of the external nares. Symbols represent the mean log10 CFU ± standard error shed from four mice/group infected with RB50 (white squares) or RB50ΔarnT (black diamonds). The limit of detection is marked by a dashed line.
73 Chapter 3 B. bronchiseptica and the Caspase-1 Inflammasome.
74 62 Abstract Successful bacterial pathogens have evolved numerous strategies to evade and manipulate host immune responses. These can include the ability to overcome early innate immune responses in order to prevent detection and favorably shift the type of immune responses elicited by the infection. Caspase-1 is a key inflammasome component that activates a pro-inflammatory signaling cascade and has been shown to be important for the control of invading pathogens. Using the animal pathogen B. bronchiseptica as a model, we investigated how bacterial infection can result in decreased caspase-1 levels; we also uncovered a novel requirement for the recently characterized Type VI Secretion System (T6SS). Although no differences in colonization or cytokine expression were detected between wild-type and Casp1 -/- mice during infection, exposing cultured macrophages to a functional B. bronchiseptica T6SS resulted in a reduction of Nlrp3 protein, an important NLR bacterial sensing protein, indicating the involvement of multiple components of the inflammasome. The presence of the T6SS did not affect Casp1or Nlrp3 mrna levels, indicating that the decrease did not take place at the level of transcription. Tissue culture inhibitor assays revealed that the B. bronchiseptica T6SS was required for the induction of autophagy and that this process caused the decrease in caspase-1. Understanding this mechanism illustrates the ability of B. bronchiseptica to manipulate host microbe sensing pathways to reduce inflammasome function and may allow for the future development of novel treatments for bacterial disease.
75 63 Introduction The presence of pathogenic bacteria in the body initiates a series of molecular signaling events that lead to activation of resident immune cells, the expression of proinflammatory cytokines, chemokine cascades, cell recruitment to the site of infection, and the generation of an adaptive immune response. Bacterial infections initiate an acute inflammatory response in the respiratory tract. Damage caused by the pathogen results in the expression of pro-inflammatory cytokines such as members of the IL-1 family, TNFα, and IL-6 (43) by resident phagocytic cells as well as epithelial tissue (49), resulting in chemokine signaling via CXCL10 (60), CCL20 (34), KC, LIX, or MIP-2 (58) and leukocyte extravasation through the microvasculature. The cytokines produced will also determine the type of immune response (ex. type 1 or 17 versus type 2) (44), T cell responses at the site of infection (2), and the generation of an adaptive immune response (26, 28) (Figure 3.1A). Understanding the mechanisms and interactions underlying the innate immune response will create a more complete picture of the dynamics of the overall immune system response to infection. The IL-1 signaling pathway (Figure 3.1B) consists of the generation and processing of precursor cytokines IL-1α and IL-1β, which in turn signal through the IL- 1R. This results in activation of NF-κB signaling as well as JNK, ERK1/2, p38 pathways, which induce the expression of IL-1 target inflammatory genes as well as Il1a
76 64 and Il1b (55). IL-1β precursor protein has been shown to be cleaved and prepared for secretion by protein complexes called inflammasomes, which assemble in response to pro-inflammatory stimuli (19). In this work, we used the respiratory pathogen B. bronchiseptica to study IL-1 and inflammasome signaling. B. bronchiseptica is closely related to the subspecies B. pertussis and B. parapertussis, the causative agents of whooping cough in patients (37). Therefore, researching the pathogenesis of B. bronchiseptica infection can reveal bacterial strategies for evading and manipulating the host immune response and lead to novel treatments for both human and veterinary respiratory diseases. Murine survival assays have shown that the IL-1R is necessary during B. bronchiseptica infection (Figure 3.2 A,B; L. Bendor and A. Karanikas, unpublished). IL- 1R -/- mice were shown to succumb to lethal bordetellosis, with entire cohorts of mice inoculated with B. bronchiseptica strain RB50 at doses of 5 x 10 5 and 1 x 10 5 CFU dying by days 3 and 4 post-inoculation respectively. The LD50 of RB50 in IL-1R -/- mice appears to be between 5 x 10 4 and 1 x 10 5 CFU, whereas death of wild-type mice was not observed with the infectious dose used in this experiment during the 12 day time period. Measurement of early colonization in the respiratory tract of inoculated mice revealed that RB50 colonized to significantly higher levels in the absence of IL-1R in the lungs and nasal cavity (Figure 3.3A,C), but no changes in colonization were observed in the trachea (Figure 3.3B). These results suggest that IL-1R is required for the control of bacterial numbers, which, if uncurbed, could be the cause of death in the mice. Interestingly, no mortality was observed in either wild-type or IL-1R deficient mice when they were inoculated with bacteria lacking a functional Type VI secretion
77 65 system (T6SS) (Figure 3.2 C,D). These data indicate that the T6SS is necessary for the lethal phenotype and could play an important role in interacting with the IL-1 signaling pathways. The genome of B. bronchiseptica laboratory strain RB50 contains a contiguous 26 gene locus that is predicted to encode T6SS associated genes (57). Type VI secretion systems are present in Gram negative bacteria and are structurally similar to T4 bacteriophage tail spikes (Figure 3.4) (23). The secretion apparatus consists of a membrane spanning protein complex, including core components Hcp, a multimerized protein that forms the tube-like structure for effector secretion; VgrG, which forms the tip of the complex that enables it to interact with the membranes of other cells; and ClpV, the AAA+ ATPase acting as the primary power source for the system (47). In the absence of clpv, the T4 phage-like sheath is unable to properly assemble and contract (3), rendering the secretion apparatus stuck and nonfunctional. The T6SS has been linked to virulence in eukaryotic systems. In Aeromonas hydrophila (51) and Legionella pneumophila (41), the T6SS is associated with increased macrophage cytotoxicity; and in Vibrio cholerae (35) and H. hepaticus (9) infections in the gut, the T6SS has been shown to lead to an increase in inflammation. In B. bronchiseptica, infections with the mouse model have shown that a functional T6SS is necessary for colonization of the respiratory tract as well as levels of lung pathology (57). These data indicate that the B. bronchiseptica T6SS could influence inflammatory processes during infection. In addition, the presence of the clpv component is required for production of key proinflammatory cytokines in cultured
78 66 macrophages, indicating that this system could influence the host innate immune response. Due to the superior natural host infection model and dramatic phenotypes observed, the knowledge acquired from studying how the T6SS in B. bronchiseptica interacts with the innate immune system can be used to better understand how human pathogens (Y. pestis, S. typhimurium, E. tarda, F. tularensis, P. aeruginosa, V. cholerae (40) ) that also have Type VI secretion systems, manipulate the immune system. One potential mechanism by which the T6SS could affect murine survival is by inducing increased levels of IL-1 cytokine, leading to localized inflammation and the generation of immune responses that could control the numbers and location of B. bronchiseptica. Therefore, in the absence of IL-1 signaling, the bacteria would be unchecked by this response and could spread systemically, leading to septic shock and death. However, no significant T6SS dependent changes in total IL-1β levels were detected in either the early cytokine response (Figure 3.5A) in the lungs or over the course of infection (Figure 3.5B). A possible explanation could be that the T6SS affects changes in active IL-1β levels, facilitated by caspase-1 inflammasome function (56). Work by Dunne et al. (15) has shown that adenylate cyclase toxin induces Nlrp3-caspase- 1 inflammasome mediated activation of IL-1β during B. pertussis infection, resulting in a robust, protective Th17 immune response. These data highlight the potential importance of inflammasome signaling during host-immune interactions with bordetellae species. One of the key mediators of inflammation is the cysteine protease caspase-1 (Figure 3.6A). Caspase-1 activation results in cleavage and activation of IL-1β and IL-18 (14) pro-inflammatory cytokines, as well as alterations in NF-kB signaling (31) and
79 67 changes in cell death pathways (1). These localized signaling events can affect many important immune processes which include promoting T cell differentiation (10) and upregulating cytokine production in immune cells (42). The caspase-1 inflammasome consists of a Nod-like receptor (prominent members include Nlrp1, Nlrp3, and Nlrc4 (21)) or Aim2 (24) cytosolic sensor protein (Figure 3.6B), which interacts though a Pyrin-domain with the scaffolding protein Asc (50), and caspase-1, which is recruited by Asc through interaction with the CARD domain and cleaved for activation (16). Activation of various members of the NLR family and assembly of the inflammasome complexes are stimulated by different ligands, such as lethal toxin produced by Bacillus anthracis and the muramyl dipeptide peptidoglycan component for Nlrp1 (17, 33); a wide variety of microbial toxins and ionophores or endogenous danger molecules for Nlrp3 (62); bacterial flagellin and T3SS structural components for Nlrc4 (38); and cytosolic DNA for Aim2 (18). One of the major inflammasome functions involves detection of microbial components (Figure 3.7A). This process enables the detection of pathogens, leading to an appropriate immune signaling response to facilitate their clearance. The Nlrp3 (nucleotide-binding domain and leucine-rich repeat protein 3) inflammasome is one of the best characterized complexes; is involved in detecting physiological stresses such as arthritis or gout as well as microbial threats; and is induced in a two-step process in which an initial signal through the TLR or other proinflammatory stimulatory pathways results in increased expression of the genes encoding Nlrp3 and Il1b. The second signal (one of the aforementioned bacterial or endogenous
80 68 ligands) results in the activation of the Nlrp3 complex formation (29). In Nlrp3 -/- macrophages, caspase-1 activation is decreased and susceptibility to killing is increased during V. parahaemolyticus and Yersinia infection (22, 61), indicating that Nlrp3 is necessary for control of complex assembly in response to microbes. However, pathogenic bacteria have developed ways to inhibit inflammasome function and favorably alter the mechanics of the subsequent host immune response to counteract detection and the subsequent immune response (Figure 3.7B). A M. tuberculosis zinc metalloprotease, Zmp1, is required to prevent caspase-1 activation (36). The presence of F. tularensis genes mvin and ripa decreases AIM2 inflammasome activation (54). In addition, Type III secretion system (T3SS) effectors play prominent roles in inflammasome inhibition. P. aeruginosa PA103 T3SS effector ExoU acts to inhibit the activation of caspase-1 (52) Yersinia T3SS effector protein, YopK, impedes recognition of bacterial components by the Nlrp3 and Nlrc4 inflammasome (4), and YopM binds directly to caspase-1 to prevent the formation of the complex (32). Another manner through which inflammasome activity can be limited is via activation of autophagy (46). Selective macroautophagy is a specialized degradative process which results in the targeted destruction of large cellular protein complexes, organelles, and even foreign bodies (30). Key steps in the process include autophagosome formation through the class III PI3K and Beclin-1 ;sequestration of the ubiquitinylated protein of interest; autophagosome-lysosomal fusion; and ultimately degradation and transport of components (Figure 3.8A) (53). Selective autophagy has also been shown to be involved in xenophagy to remove invading microorganisms (27).
81 69 Autophagy interfaces with the phagocytosis pathway by ubiquitinylating bacteria taken up or detected in the cell and targeting them for destruction during the autophagosomal fusion. Numerous intracellular bacteria have evolved effectors to avoid this destructive process, such as M. tuberculosis, that secretes ESAT-6 to block autophagosomal fusion, or cholera toxin from Vibrio cholera that presents the assembly of autophagy machinery (25). However, other bacteria utilize autophagy in a different survival strategy by using it to destroy inflammasome complexes and thus eliminate the pro-inflammatory downstream signals. Manipulation of autophagy by bacterial pathogens has been shown to influence bacterial interactions with the inflammasome and is a likely candidate to play a role in this system. Pseudomonas aeruginosa prevents caspase-1 from cleaving Trif, resulting in less inflammasome activation, lower autophagy, and higher bacterial numbers (26). Conversely, Vibrio parahaemolyticus secretes effector protein VopQ to induce autophagy and prevent Nlrc4 inflammasome activation (Figure 3.8B) (22). Our results indicate that B. bronchiseptica strain RB50 caused a clpv dependent decrease in caspase-1 protein in cultured macrophages as well as a decrease in Nlrp3 protein, an important inflammasome component. This clpv dependent process occurred due to selective autophagy. These novel findings implicate the Type VI secretion system in enabling B. bronchiseptica to manipulate inflammatory pathways and alter hostpathogen dynamics.
82 70 Materials and Methods Ethics Statement This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Institutional Animal Care and Use Committee at The Pennsylvania State University at University Park, PA (#31297 Bordetella-host Interaction). All animals were euthanized using carbon dioxide inhalation to minimize animal suffering. Bacterial Strains. Bordetella bronchiseptica strains RB50 (12) and RB50ΔclpV (57) were grown on Bordet- Gengou (BG) agar (HiMedia) supplemented with 15% defibrinated sheep blood (Hema Resources & Supply, Inc.) and 20 μg/ml streptomycin (Sigma-Aldrich) or grown to mid-log phase in Stainer-Scholte broth (48) supplemented with 500μg/mL heptakis (Sigma-Aldrich) shaking overnight at 37 C. Animal experiments. Wild type C57BL/6 mice were obtained from Jackson Laboratories, Bar Harbor, ME and and Casp1 -/- mice were obtained as a gift from the Flavell Lab at Yale University. Mice were bred and maintained at a specific pathogen-free facility at The Pennsylvania State University, University Park, PA, and all experiments were carried out in accordance with all institutional guidelines. All animal experiments were done as previously described (57). Mice were sedated with 5% isoflurane (IsoFlo, Abbott Laboratories) in oxygen and inoculated by pipetteing 50 µl PBS containing the indicated number of CFU onto the external nares of the mouse. For quantification of bacterial numbers, mice were euthanized with CO2 inhalation, and the respiratory organs were harvested. Tissues were homogenized in PBS, serially diluted and plated onto BG (Hi-Media) agar plates with 20 µg/ml streptomycin. Colonies were enumerated after 2
83 71 days of growth at 37 C. Survival curves were generated as previously described (6). Mice were observed over a 28 day period; any mouse exhibiting lethal bordetellosis, indicated by ruffled fur, labored breathing, and diminished responsiveness, was euthanized immediately to prevent unnecessary suffering. Bone Marrow Derived Macrophages. Bone marrow-derived macrophages were generated from bone marrow isolated from the femurs of C57BL/6 mice as previously described (60). Briefly, to extract the marrow, the bones were crushed with a sterile pestle through a 70 μm nylon cell strainer (BD Biosciences), and the cell suspension was pelleted and resuspended in Dulbecco s Modified Eagle s Medium/ F-12, 1:1 Mixture, with g/l Glucose, 15 mm HEPES, and L-Glutamine (BioWhittaker) containing 10% FBS (Gemini) and 20% L-929 cell supernatant media before seeding the plates. The L-929 supernate contains protein factors such as GM-CSF for macrophage differentiation. Cells were seeded into 100 x 15 mm non-tissue culture treated Petri dishes (VWR) and grown in 10mL of 20% L-929 supernate differentiation media in 5% CO2 incubator (Binder) at 37 C. After 6 days, the cells were checked for morphology and adherence. The cells were then removed from the plates using Cellstripper (Cellgro), resuspended in media and seeded into 6-well (Greiner Bio- One) or 24-well tissue culture-treated plates (Costar) overnight at densities of 1 x 10 6 and 2 x 10 5 cells per well, respectively. Cell culture infections. Bone marrow-derived macrophages were primed with 100ng/mL E. coli 011:B4 LPS (Sigma-Aldrich) for three hours. After three hours, all wells were aspirated and replenished with fresh media, or media containing RB50 or RB50ΔclpV at MOIs of 10. In assays to inhibit autophagy, cells were pre-treated for 30 minutes with media containing 10mM 3-methyladenine (3-MA) (Sigma-Aldrich). To ensure contact between the bacteria and the cultured macrophages, the plates were centrifuged at 500 x g for 5 minutes. After 2 or 4 hours, the medium was
84 72 removed, wells were washed with 1 x PBS, and cell lysates were harvested. All assays were performed in triplicate. RNA Extraction. Cell lysates were harvested in 1mL Trizol (Life Technologies) for RNA extraction. Trizol extraction was performed according to previously established methods (8). The extracted nucleic acid was treated with RNase free DNAse I (Ambion) for 20 minutes at 37 C to remove DNA contamination, and then the concentration of RNA was measured by Nanodrop (Thermo Scientific). RT-qPCR. 1μg of RNA from each sample was used to synthesize cdna using the Superscript III First Strand Synthesis System (Invitrogen) according to manufacturer s instructions. The resulting cdna was used as a template for qpcr. SYBR GreenER (Invitrogen) Master Mix was used with primers for Casp1 (5 -GCC CAC TGC TGA TAG GGT GA-3 (forward) and 5 -CCC GGG AAG AGG TAG AAA CG-3 (reverse)), Nlrp3 (5 -ATC AAC AGG CGA GAC CTC TG- 3 (forward) and 5 -GTC CTC CTG GCA TAC CAT AGA-3 (reverse) ), or Gapdh (5 -TGA CCT CAA CTA CAT GGT CTA CA-3 (forward) and 5 -CTT CCC ATT CTC GGC CTT G-3 (reverse)). ROX was used as a passive reference dye. 2 μl cdna and 23 μl Master Mix were added to each well of a 96-well optical reaction plate (Denville), and each plate was run using an Applied Biosystems StepOne Real-Time PCR System. All reactions were performed with 3 biological replicates in technical duplicate with no-template and no-reverse transcriptase controls. The data were analyzed using the 2 -ΔΔCt method (45), and expression of all genes was normalized to the Gapdh reference gene. The data are mean increases relative to samples harvested after 0 hours treatment.
85 73 Western Blots Cell culture lysates were harvested in Laemmli sample buffer (BioRad) with 2- Mercaptoethanol (Sigma-Aldrich), and protein concentrations were quantified using the Pierce 660 nm Protein assay in conjunction with the Nanodrop (Thermo Scientific). For detection of IL- 1β, cell culture supernatants were concentrated by a methanol/chloroform precipitation method described previously (15). Samples were run on a 12% polyacrylamide gel under reducing conditions and transferred to a polyvinylidene difluoride (PVDF) membrane (BioRad). Membranes were probed with primary antibodies against caspase-1 p10 (Santa Cruz, sc-514, 1:1000), Nlrp3 (Abcam, ab4207, 1:500), anti-il-1β (Abcam, ab9722) or β-actin (Sigma Aldrich, A2006, 1:12,000) and secondary goat anti-rabbit-hrp (Southern Biotech, , 1:10,000). The membrane signals were visualized using Immobilon Chemiluminescence detection reagent (Millipore) and the ChemiDoc XRS+ System (BioRad). Statistical Analyses. Significant differences between groups were determined using the 1-way ANOVA using MINITAB and Student s t-test with p values <0.05 considered significant. Results Caspase-1 is not required for B. bronchiseptica respiratory tract colonization. To determine if the previously observed IL-1R dependent increases in B. bronchiseptica colonization and lethality occurred due to the effects of caspase-1 signaling, colonization by wild-type RB50 (which has an intact T6SS) and RB50ΔclpV bacteria in C57BL/6 was measured (Figure 3.9 A, B, C), as well as RB50 in C57BL/6 and Casp1 -/- mice inoculated with CFU (Figure 3.9 D, E, F). The levels of
86 74 colonization of RB50 were significantly higher in comparison those of RB50ΔclpV, illustrating that clpv is required for colonization of the respiratory tract. However, no statistically significant differences in bacterial numbers were observed between the wildtype and Casp1 -/- mice, and none of the Casp1 -/- mice succumbed to bordetellosis during the 28 day time course. Therefore, it is unlikely that the T6SS dependent ability of bacteria to grow to large numbers in the absence of IL-1R is dependent on the caspase-1 mediated ability to activate IL-1β. It is also possible, because the IL-1R -/- mice were generated on the F2.129 background and were not C57BL/6 mice, that the previous phenotype was not observed due to genetic background specific effects (due to strain specific enhancers or alleles in inflammatory genes). However, it is possible that the T6SS could act on multiple points of the pathway. These results could also indicate that the T6SS is able to downregulate caspase-1 in the wild-type mice, thus enabling it to colonize at equivalent levels regardless of the presence of caspase-1. Caspase-1 could act on specialized immune cells such as macrophages and the result of the signaling involves a more specific, localized response. Therefore, the effect of the T6SS on caspase-1 in cultured macrophages was assayed (Figure 3.10). B. bronchiseptica reduces caspase-1 protein in a clpv-dependent manner. To determine the effect of B. bronchiseptica strain RB50 on the levels of caspase- 1 protein in macrophages, primed bone marrow-derived macrophages (BMDM) cells were inoculated at an MOI of 10 with RB50, RB50ΔclpV (lacks a functional T6SS), or medium as a control. Western blot analysis showed that there were lower levels of procaspase-1 and active caspase-1 protein in the macrophages treated with RB50 in comparison to medium alone (Figure 3.11 A,B ). However, the macrophages treated with
87 75 RB50ΔclpV did not show any changes in either form of caspase-1 in comparison to those treated with only medium, indicating that clpv was required for lower levels of caspase- 1. One of the canonical functions of caspase-1 is its ability to proteolytically cleave the precursor form of IL-1β, an important proinflammatory cytokine, into the active form, enabling the secretion of the mature IL-1β from the cell (59). To determine the effect of the B. bronchiseptica T6SS component clpv on IL-1β, BMDM cells were stimulated with LPS and treated with media, RB50, or RB50ΔclpV at an MOI of 10 for 2 or 4 hours. As a positive control, additional cells were treated with 5mM ATP to provide the second signal necessary for IL-1β secretion, and unprimed cells were included as a negative control (Figre 3.12A). Higher levels of total IL-1β were present in samples treated with RB50 in comparison to those treated with RB50ΔclpV, but the ratio of active IL-1β to total did not change in a clpv dependent manner (Figure 3.12B). Based on these findings, clpv is necessary for increased levels of total IL-1β protein. However, these data indicate that the clpv dependent decrease in caspase-1 does not result in a downstream decrease in active IL-1β. This is likely processed through one of the noncanonical activation pathways. clpv dependent reduction in caspase-1 occurred at the protein level rather than reduction in Casp1 mrna levels. One possible mechanism by which B. bronchiseptica affected caspase-1 protein levels was to alter the mrna levels of caspase-1. RT-qPCR analysis performed on BMDMs treated with RB50 or RB50ΔclpV (Figure 3.13). Analysis was performed using the 2 -ΔΔCt method (45), and expression of all genes was normalized to the Gapdh
88 76 reference gene. The data are mean increases relative to samples harvested after 0 hours treatment. The infected macrophages showed no significant clpv dependent changes in Casp1 mrna levels, signifying that the lower levels of the caspase-1 protein occurred in translation or are due to degradation. Reduction in caspase-1 protein correlates with lower levels of Nlrp3 protein. Because caspase-1 functions as a key member of canonical inflammasomes, it is likely that if caspase-1 protein levels were decreased in a clpv dependent manner, other protein components of the complex would be affected as well. Nlrp3 is a NOD-like receptor that serves as a sensor for the complex and has been shown to be stimulated by numerous pathogens (5, 11, 39). Western blot analysis to detect caspase-1, Nlrp3, and beta actin was performed on whole cell lysate collected from LPS primed BMDMs treated with media, RB50, and RB50ΔclpV (Figure 3.14 A,B). Densitometry analysis indicated that there were lower levels of Nlrp3 in cells treated with RB50 in comparison to those treated with RB50ΔclpV. Therefore, the decrease in caspase-1 protein levels may have been due to the decrease in components of assembled Nlrp3 inflammasomes. To determine if the presence of the T6SS affected the mrna levels of inflammasome components, RT-qPCR analysis was performed on samples extracted from BMDMs treated with RB50 or RB50ΔclpV. Fold induction of Nlrp3 (Figure 3.14C) relative to untreated macrophages showed no statistically significant clpv dependent changes in mrna levels. These data indicated that the decrease in Nlrp3 occurred at the level of the protein.
89 77 clpv is required for induction of autophagy and degradation of caspase-1 protein. A major protein degradation pathway that interacts with immune signaling components is selective autophagy (30). Recent findings have shown that inflammasomes can be selectively targeted for destruction in autophagosomes. To determine whether autophagy played a role in the clpv dependent decrease of caspase-1, BMDMs were incubated with medium or 10mM 3-methyadenine (3MA) and then treated with RB50 or RB50ΔclpV. Treatment with 3MA inhibited the process of autophagosomal formation, and prevented autophagy from occurring. Western blot analysis performed to detect caspase-1 and actin (Figure 3.15A,B) showed that the clpv dependent decrease in caspase-1 when cells were treated with RB50 did not occur when the 3MA inhibitor was added. The presence of 3MA also does not affect IL-1β activation levels. Taken as a whole, these results indicate that the presence of the B. bronchiseptica T6SS component clpv is required for the activation of the selective autophagy pathway and decreased the levels of caspase-1. Discussion Caspase-1, a prominent pro-inflammatory signaling protein, has been shown to influence the overall immune response through signaling at the site of infection and leading to larger shifts in the global immune response type. In culture, clpv, a gene encoding a core component of the B. bronchiseptica T6SS, was shown to be necessary for the induction of autophagy and decrease in caspase-1. These results suggest a novel requirement for T6SS in bacterial interaction with the inflammasome as well as
90 78 autophagosomal degradation. Changes in Nlrp3 indicated that the T6SS mediated effect could act on the Nlrp3 inflammasome complex as a whole and result in the possible ubiquitinylation and degradation of the entire complex via autophagy. The clpv dependent decrease in caspase-1 could be due to direct interaction between T6SS effectors acting on host cell or disruption of bacterial systems by dysregulating the T6SS and other B. bronchiseptica virulence factors. For example, in Burkholderia mallei, the T6SS shares regulatory pathways with the T3SS, which has previously been shown to influence inflammasome function (7). The T3SS might also be required for decreases in caspase-1 protein levels, and future work will be conducted to further explore these possibilities. In addition, this alteration in immune signaling could affect the intracellular phase for B. bronchiseptica (20). Bacterial pathogens have been shown to manipulate autophagy to survive intracellularly in addition to promoting inflammasome degradation (28). The T6SS induction of autophagy could come at the expense of the ability of B. bronchiseptica to occupy intracellular niche. However, the potential alteration of the inflammatory immune response could be more beneficial for the entire bacterial population. Identification of a T6SS effector that could induce the decrease of the inflammasome complex could be used as a potential recombinant protein treatment, which could reduce inflammation by decreasing Nlrp3 inflammasome signaling in chronic inflammatory conditions such as arthritis and gout. Chemical inhibitors utilizing a similar strategy of decreasing inflammasome activation are currently undergoing clinical trials (13); however, care must be taken to prevent subjecting the patient to a risk
91 79 of septic shock if the resident bacteria can no longer be properly controlled, such as was observed in the IL-1R -/- mouse model. Overall, bacterial manipulation of the autophagy pathway correlated to a decrease the levels of a major inflammasome component. This decrease could allow bacteria to change the response of resident macrophages and thus affect larger, more global immune responses in the respiratory tract. These data have uncovered a novel bacterial immune interaction involving a functional T6SS and its requirement for an autophagy dependent decrease in caspase-1, a key inflammatory signaling protein. These results can contribute to our understanding of immune-pathogen dynamics and may aid in the development of novel treatments for infectious disease.
92 80 Author Contributions Sarah J. Muse 1,2, David E. Place 1,3, Laura L. Goodfield 1,3, Sara E. Hester 1,2, Alexia T. Karanikas 1,3, Liron Bendor 1,4, Laura S. Weyrich 1,2, Eric T. Harvill 1 1 Department of Veterinary and Biomedical Sciences, The Pennsylvania State University, University Park, Pennsylvania, USA 2 Graduate Program in Biochemistry, Microbiology and Molecular Biology 3 Graduate Program in Immunology and Infectious Diseases 4 Graduate Program in Cell and Developmental Biology 5 Graduate Program in Genetics Conceived and designed experiments: SJM, DEP, LLG, SEH, ATK, LB, ETH Performed experiments: SJM, DEP, LLG, ATK, LB Analyzed the data: SJM, DEP, LLG, ATK, LB Wrote the paper: SJM, ETH
93 81 References 1. Aachoui, Y., V. Sagulenko, E. A. Miao, and K. J. Stacey. Inflammasome-mediated pyroptotic and apoptotic cell death, and defense against infection. Curr Opin Microbiol 16: Andreasen, C., D. A. Powell, and N. H. Carbonetti Pertussis toxin stimulates IL-17 production in response to Bordetella pertussis infection in mice. PLoS One 4:e Basler, M., M. Pilhofer, G. P. Henderson, G. J. Jensen, and J. J. Mekalanos. Type VI secretion requires a dynamic contractile phage tail-like structure. Nature 483: Brodsky, I. E., N. W. Palm, S. Sadanand, M. B. Ryndak, F. S. Sutterwala, R. A. Flavell, J. B. Bliska, and R. Medzhitov. A Yersinia effector protein promotes virulence by preventing inflammasome recognition of the type III secretion system. Cell Host Microbe 7: Broz, P., K. Newton, M. Lamkanfi, S. Mariathasan, V. M. Dixit, and D. M. Monack. Redundant roles for inflammasome receptors NLRP3 and NLRC4 in host defense against Salmonella. J Exp Med 207: Buboltz, A. M., T. L. Nicholson, L. S. Weyrich, and E. T. Harvill Role of the type III secretion system in a hypervirulent lineage of Bordetella bronchiseptica. Infect Immun 77: Burtnick, M. N., and P. J. Brett. Burkholderia mallei and Burkholderia pseudomallei cluster 1 type VI secretion system gene expression is negatively regulated by iron and zinc. PLoS One 8:e Chomczynski, P., and N. Sacchi The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: twenty-something years on. Nat Protoc 1: Chow, J., and S. K. Mazmanian. A pathobiont of the microbiota balances host colonization and intestinal inflammation. Cell Host Microbe 7: Chung, Y., S. H. Chang, G. J. Martinez, X. O. Yang, R. Nurieva, H. S. Kang, L. Ma, S. S. Watowich, A. M. Jetten, Q. Tian, and C. Dong Critical regulation of early Th17 cell differentiation by interleukin-1 signaling. Immunity 30: Costa, A., R. Gupta, G. Signorino, A. Malara, F. Cardile, C. Biondo, A. Midiri, R. Galbo, P. Trieu-Cuot, S. Papasergi, G. Teti, P. Henneke, G. Mancuso, D. T. Golenbock, and C. Beninati. Activation of the NLRP3 inflammasome by group B streptococci. J Immunol 188: Cotter, P. A., and J. F. Miller BvgAS-mediated signal transduction: analysis of phase-locked regulatory mutants of Bordetella bronchiseptica in a rabbit model. Infect Immun 62: Di Virgilio, F. The therapeutic potential of modifying inflammasomes and NOD-like receptors. Pharmacol Rev 65: Dinarello, C. A Interleukin-1 beta, interleukin-18, and the interleukin-1 beta converting enzyme. Ann N Y Acad Sci 856: Dunne, A., P. J. Ross, E. Pospisilova, J. Masin, A. Meaney, C. E. Sutton, Y. Iwakura, J. Tschopp, P. Sebo, and K. H. Mills. Inflammasome activation by adenylate
94 cyclase toxin directs Th17 responses and protection against Bordetella pertussis. J Immunol 185: Fang, R., K. Tsuchiya, I. Kawamura, Y. Shen, H. Hara, S. Sakai, T. Yamamoto, T. Fernandes-Alnemri, R. Yang, E. Hernandez-Cuellar, S. R. Dewamitta, Y. Xu, H. Qu, E. S. Alnemri, and M. Mitsuyama. Critical roles of ASC inflammasomes in caspase-1 activation and host innate resistance to Streptococcus pneumoniae infection. J Immunol 187: Faustin, B., L. Lartigue, J. M. Bruey, F. Luciano, E. Sergienko, B. Bailly-Maitre, N. Volkmann, D. Hanein, I. Rouiller, and J. C. Reed Reconstituted NALP1 inflammasome reveals two-step mechanism of caspase-1 activation. Mol Cell 25: Fernandes-Alnemri, T., J. W. Yu, P. Datta, J. Wu, and E. S. Alnemri AIM2 activates the inflammasome and cell death in response to cytoplasmic DNA. Nature 458: Fettelschoss, A., M. Kistowska, S. LeibundGut-Landmann, H. D. Beer, P. Johansen, G. Senti, E. Contassot, M. F. Bachmann, L. E. French, A. Oxenius, and T. M. Kundig. Inflammasome activation and IL-1beta target IL-1alpha for secretion as opposed to surface expression. Proc Natl Acad Sci U S A 108: Forde, C. B., R. Parton, and J. G. Coote Bioluminescence as a reporter of intracellular survival of Bordetella bronchiseptica in murine phagocytes. Infect Immun 66: Franchi, L., T. Eigenbrod, R. Munoz-Planillo, and G. Nunez The inflammasome: a caspase-1-activation platform that regulates immune responses and disease pathogenesis. Nat Immunol 10: Higa, N., C. Toma, Y. Koizumi, N. Nakasone, T. Nohara, J. Masumoto, T. Kodama, T. Iida, and T. Suzuki. Vibrio parahaemolyticus effector proteins suppress inflammasome activation by interfering with host autophagy signaling. PLoS Pathog 9:e Ho, B. T., T. G. Dong, and J. J. Mekalanos. A view to a kill: the bacterial type VI secretion system. Cell Host Microbe 15: Hornung, V., A. Ablasser, M. Charrel-Dennis, F. Bauernfeind, G. Horvath, D. R. Caffrey, E. Latz, and K. A. Fitzgerald AIM2 recognizes cytosolic dsdna and forms a caspase-1-activating inflammasome with ASC. Nature 458: Huang, J., and J. H. Brumell. Bacteria-autophagy interplay: a battle for survival. Nat Rev Microbiol 12: Jabir, M. S., N. D. Ritchie, D. Li, H. K. Bayes, P. Tourlomousis, D. Puleston, A. Lupton, L. Hopkins, A. K. Simon, C. Bryant, and T. J. Evans. Caspase-1 cleavage of the TLR adaptor TRIF inhibits autophagy and beta-interferon production during Pseudomonas aeruginosa infection. Cell Host Microbe 15: Jin, M., X. Liu, and D. J. Klionsky. SnapShot: Selective autophagy. Cell 152: e Jo, E. K., J. M. Yuk, D. M. Shin, and C. Sasakawa. Roles of autophagy in elimination of intracellular bacterial pathogens. Front Immunol 4: Kim, J. J., and E. K. Jo. NLRP3 inflammasome and host protection against bacterial infection. J Korean Med Sci 28: Kraft, C., M. Peter, and K. Hofmann. Selective autophagy: ubiquitin-mediated recognition and beyond. Nat Cell Biol 12: Lamkanfi, M., M. Kalai, X. Saelens, W. Declercq, and P. Vandenabeele Caspase-1 activates nuclear factor of the kappa-enhancer in B cells independently of its enzymatic activity. J Biol Chem 279:
95 32. LaRock, C. N., and B. T. Cookson. The Yersinia virulence effector YopM binds caspase-1 to arrest inflammasome assembly and processing. Cell Host Microbe 12: Levinsohn, J. L., Z. L. Newman, K. A. Hellmich, R. Fattah, M. A. Getz, S. Liu, I. Sastalla, S. H. Leppla, and M. Moayeri. Anthrax lethal factor cleavage of Nlrp1 is required for activation of the inflammasome. PLoS Pathog 8:e Lopez-Boado, Y. S., L. M. Cobb, and R. Deora Bordetella bronchiseptica flagellin is a proinflammatory determinant for airway epithelial cells. Infect Immun 73: Ma, A. T., S. McAuley, S. Pukatzki, and J. J. Mekalanos Translocation of a Vibrio cholerae type VI secretion effector requires bacterial endocytosis by host cells. Cell Host Microbe 5: Master, S. S., S. K. Rampini, A. S. Davis, C. Keller, S. Ehlers, B. Springer, G. S. Timmins, P. Sander, and V. Deretic Mycobacterium tuberculosis prevents inflammasome activation. Cell Host Microbe 3: Mattoo, S., and J. D. Cherry Molecular pathogenesis, epidemiology, and clinical manifestations of respiratory infections due to Bordetella pertussis and other Bordetella subspecies. Clin Microbiol Rev 18: Miao, E. A., C. M. Alpuche-Aranda, M. Dors, A. E. Clark, M. W. Bader, S. I. Miller, and A. Aderem Cytoplasmic flagellin activates caspase-1 and secretion of interleukin 1beta via Ipaf. Nat Immunol 7: Munoz-Planillo, R., L. Franchi, L. S. Miller, and G. Nunez A critical role for hemolysins and bacterial lipoproteins in Staphylococcus aureus-induced activation of the Nlrp3 inflammasome. J Immunol 183: Pukatzki, S., S. B. McAuley, and S. T. Miyata The type VI secretion system: translocation of effectors and effector-domains. Curr Opin Microbiol 12: Purcell, M., and H. A. Shuman The Legionella pneumophila icmgcdjbf genes are required for killing of human macrophages. Infect Immun 66: Puren, A. J., P. Razeghi, G. Fantuzzi, and C. A. Dinarello Interleukin-18 enhances lipopolysaccharide-induced interferon-gamma production in human whole blood cultures. J Infect Dis 178: Ross, P. J., E. C. Lavelle, K. H. Mills, and A. P. Boyd Adenylate cyclase toxin from Bordetella pertussis synergizes with lipopolysaccharide to promote innate interleukin-10 production and enhances the induction of Th2 and regulatory T cells. Infect Immun 72: Ross, P. J., C. E. Sutton, S. Higgins, A. C. Allen, K. Walsh, A. Misiak, E. C. Lavelle, R. M. McLoughlin, and K. H. Mills. Relative contribution of Th1 and Th17 cells in adaptive immunity to Bordetella pertussis: towards the rational design of an improved acellular pertussis vaccine. PLoS Pathog 9:e Schmittgen, T. D., and K. J. Livak Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc 3: Shi, C. S., K. Shenderov, N. N. Huang, J. Kabat, M. Abu-Asab, K. A. Fitzgerald, A. Sher, and J. H. Kehrl. Activation of autophagy by inflammatory signals limits IL-1beta production by targeting ubiquitinated inflammasomes for destruction. Nat Immunol 13: Silverman, J. M., Y. R. Brunet, E. Cascales, and J. D. Mougous. Structure and regulation of the type VI secretion system. Annu Rev Microbiol 66: Stainer, D. W., and M. J. Scholte A simple chemically defined medium for the production of phase I Bordetella pertussis. J Gen Microbiol 63:
96 49. Strieter, R. M., S. L. Kunkel, M. P. Keane, and T. J. Standiford Chemokines in lung injury: Thomas A. Neff Lecture. Chest 116:103S-110S. 50. Stutz, A., G. L. Horvath, B. G. Monks, and E. Latz. ASC speck formation as a readout for inflammasome activation. Methods Mol Biol 1040: Suarez, G., J. C. Sierra, M. L. Kirtley, and A. K. Chopra. Role of Hcp, a type 6 secretion system effector, of Aeromonas hydrophila in modulating activation of host immune cells. Microbiology 156: Sutterwala, F. S., L. A. Mijares, L. Li, Y. Ogura, B. I. Kazmierczak, and R. A. Flavell Immune recognition of Pseudomonas aeruginosa mediated by the IPAF/NLRC4 inflammasome. J Exp Med 204: Tanida, I., T. Ueno, and E. Kominami LC3 and Autophagy. Methods Mol Biol 445: Ulland, T. K., B. W. Buchan, M. R. Ketterer, T. Fernandes-Alnemri, D. K. Meyerholz, M. A. Apicella, E. S. Alnemri, B. D. Jones, W. M. Nauseef, and F. S. Sutterwala. Cutting edge: mutation of Francisella tularensis mvin leads to increased macrophage absent in melanoma 2 inflammasome activation and a loss of virulence. J Immunol 185: Weber, A., P. Wasiliew, and M. Kracht. Interleukin-1 (IL-1) pathway. Sci Signal 3:cm Wen, H., E. A. Miao, and J. P. Ting. Mechanisms of NOD-like receptor-associated inflammasome activation. Immunity 39: Weyrich, L. S., O. Y. Rolin, S. J. Muse, J. Park, N. Spidale, M. J. Kennett, S. E. Hester, C. Chen, E. G. Dudley, and E. T. Harvill. A Type VI secretion system encoding locus is required for Bordetella bronchiseptica immunomodulation and persistence in vivo. PLoS One 7:e Widney, D. P., Y. Hu, A. K. Foreman-Wykert, K. C. Bui, T. T. Nguyen, B. Lu, C. Gerard, J. F. Miller, and J. B. Smith CXCR3 and its ligands participate in the host response to Bordetella bronchiseptica infection of the mouse respiratory tract but are not required for clearance of bacteria from the lung. Infect Immun 73: Yu, H. B., and B. B. Finlay The caspase-1 inflammasome: a pilot of innate immune responses. Cell Host Microbe 4: Zhang, X., T. Goel, L. L. Goodfield, S. J. Muse, and E. T. Harvill. Decreased leukocyte accumulation and delayed Bordetella pertussis clearance in IL-6-/- mice. J Immunol 186: Zheng, Y., S. Lilo, I. E. Brodsky, Y. Zhang, R. Medzhitov, K. B. Marcu, and J. B. Bliska. A Yersinia effector with enhanced inhibitory activity on the NF-kappaB pathway activates the NLRP3/ASC/caspase-1 inflammasome in macrophages. PLoS Pathog 7:e Zhong, Y., A. Kinio, and M. Saleh. Functions of NOD-Like Receptors in Human Diseases. Front Immunol 4:
 IL-1α and IL-1β synthesis and signal transduction pathway. Reprinted with permission from Nature Publishing Group, (A) Nat Rev Immunol.")
97 85 Figure 3.1. IL-1 and the pro-inflammatory immune response. A) Pro-inflammatory cytokine and cell type specific response to infection. B) IL-1α and IL-1β synthesis and signal transduction pathway. Reprinted with permission from Nature Publishing Group, (A) Nat Rev Immunol Nov;8(11):829-35, (B) Nat Rev Rheumatol Jan;10(1):44-56.
 or RB50ΔclpV (C,D), and monitored for survival up to 12 days post-inoculation.")
98 86 Figure 3.2. clpv is required for mortality in B. bronchiseptica infected IL-1R -/- mice. Groups of wild-type F2.129 or IL-1R -/- mice were inoculated with 5 x 10 5 CFU RB50 (A,B) or RB50ΔclpV (C,D), and monitored for survival up to 12 days post-inoculation. *Courtesy of A. Karanikas and L. Bendor
 and IL-1R -/- mice (square) were inoculated with 5 x 10 5 CFU RB50, and bacteria were enumerated in the lungs A), trachea B), and nasal cavity C) after 0, 12, 24, 36,")
99 87 Figure 3.3. IL-1R is required for control of B. bronchiseptica respiratory colonization. Groups of wild-type (diamond) and IL-1R -/- mice (square) were inoculated with 5 x 10 5 CFU RB50, and bacteria were enumerated in the lungs A), trachea B), and nasal cavity C) after 0, 12, 24, 36, and 48 hours. Symbols represent the mean CFU ± standard error of four individual samples. * indicates p<0.05. *Courtesy of A. Karanikas
")
100 88 Figure 3.4. The Type VI Secretion System (T6SS). A model for type VI secrection system assembly and function. A putative model that integrates the current data is proposed. An inner membrane channel formed by the IcmF like and IcmH like proteins interacts at the cytoplasmic side of the IM with a complex composed of the probable cytosolic type VI secrection (T6S) subunits and the ClpV AAA + ATPase. Recruitment of the ClpV multimer is induced by the regulation of forkhead associated (FHA) phosphorylation through the activities of PpkA and PppA, and by the presence of the Hcp protein. A multimer of the putative lipoprotein in association with periplasmic subunits is shown at the outer membrane. Putative routes for substrate translocation are depicted through the cell envelope and the host cell membrane (blue arrow) including a one step mechanism through a unique channel, and a two step mechanism, in which both steps are catalysed by T6S subunits with transient accumulation in the periplasm (P). This hypothetical model shows a trimeric VgrG inserted into the OM through the amino terminal domain and puncturing the host cell through the needle like structure formed by the central domains, releasing the activity domain into the host cytosol (for eukaryotic like activities) or in the medium (for binding or adhesion activities). J, L, M and H (TssJ, TssL, TssM and TssH respectively) represent the T6S core components, following the nomenclature of Shalom et al, Reprinted with permission from Wiley Online Library, EMBO Rep Aug;9(8):
101 89 Figure 3.5. Deletion of clpv does not affect induction of IL-1β in murine lungs during infection. Groups of C57BL/6 mice were inoculated with 5 x 105 CFU RB50 (black bars) or RB50ΔclpV (white bars). The early induction of IL-1β protein levels in the lungs was measured 4, 24, or 48 hours post-inoculation A), and the IL-1β levels were also measured over the course of infection 0,3,7,14, 28, and 49 days post-inoculation B). Bar represent the mean CFU ± standard error of four individual samples
 from microorganisms or damage-associated molecular patterns (DAMPs) from endogenous danger")
102 90 Figure 3.6. Caspase-1 Inflammasome Complex. A) Mechanism of Inflammasome complex formation. The auto-repression of NLRP3 is removed in the presence of pathogen associated molecular patterns (PAMPs) from microorganisms or damage-associated molecular patterns (DAMPs) from endogenous danger signals. This results in exposure of the NACHT domain. In turn, NLRP3 oligomerizes and recruits apoptosis-associated speck-like protein containing a CARD (ASC; also known as PYCARD) and pro-caspase 1, triggering the activation of caspase 1 and the maturation and secretion of pro-inflammatory cytokines such as interleukin-1β (IL-1β) and IL-18. B) The three biochemically characterized inflammasomes. The NLRP1 inflammasome consists of NLRP1, ASC, and caspases-1 and -5. Little is known about the agonists that activate NLRP1. Anthrax lethal toxin, MDP, and decreased cytosolic ATP have been reported to stimulate this inflammasome. NAIP and NLRC4 form a caspase-1 inflammasome in response to bacterial flagellin and T3SS rod proteins. NLRP3, on the other hand, is activated by a wide range of agonists including a number of MAMPs and DAMPs. Reprinted with permission from Nature Publishing Group, Nat Rev Immunol Mar;10(3):210-5 and via the Creative Commons Attribution (CC BY) license, Front Immunol Oct 16;4:333.

103 Figure 3.7. Pathogen Interaction with the Inflammasome. 91
104 92 A) Pathogen components that activate inflammasomes. B) Microbial and endogenous inhibitors of inflammasome activation. Reprinted with permission from Nature Publishing Group, Nat Immunol Mar 19;13(4): and Nat Immunol Mar 19;13(4):333-42
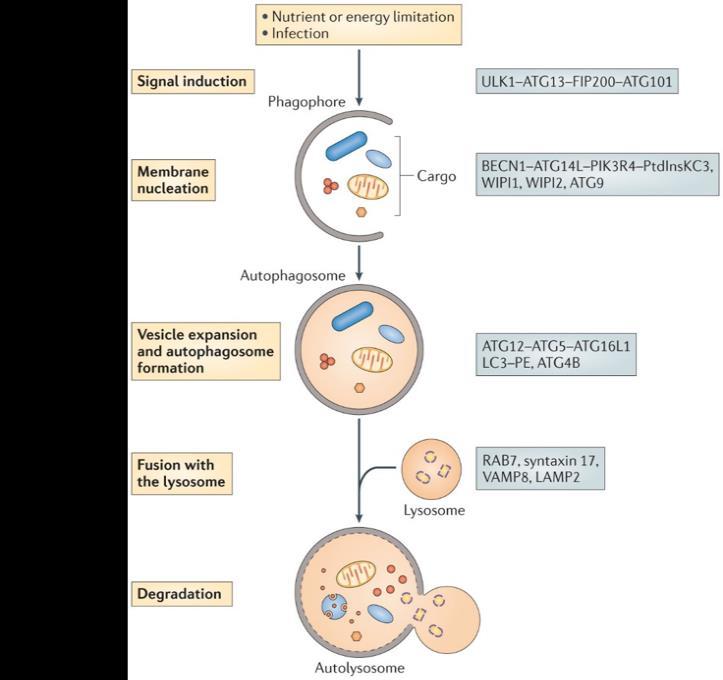
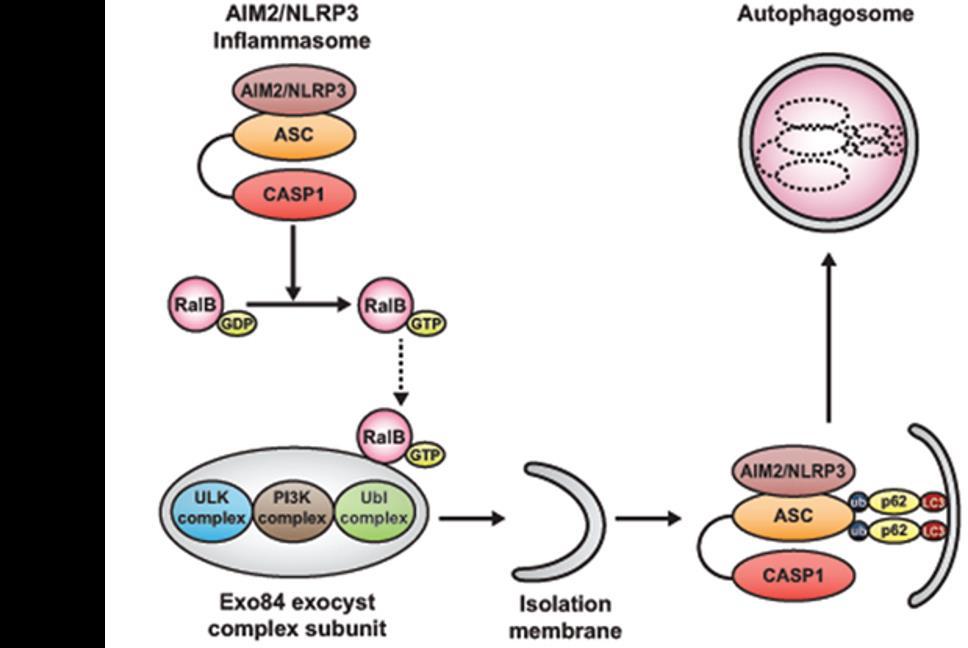
105 93
106 94 Figure 3.8. Autophagy and the Inflammasome. A) Model overview of autophagy pathway. B) Degradation of inflammasome through autophagy pathway. Reprinted with permission from Nature Publishing Group, Nat Rev Microbiol Feb;12(2): and via the Creative Commons Attribution (CC BY) license, Front Immunol Mar 12;3:44. Figure 3.9. Effect of clpv and Caspase-1 during B. bronchiseptica Colonization in the Mouse Model. Colonization of RB50 verses RB50ΔclpV in C57BL/6 mice at an inoculation dose of CFU in 50 µl in the nasal cavity A), trachea B), and lung C). Colonization of RB50 in C57BL/6 and Casp1 -/- mice at an inoculation dose of CFU in 50 µl in the nasal cavity D), trachea E), and lungs F). Symbols represent the mean CFU ± standard error of four individual samples *= p<0.05
107 95


108 96 Figure Model of T6SS affecting the inflammasome in macrophages. Model of formation of the T6SS Apparatus.
109 97 Figure clpv was required for reduction of capase-1 protein in cultured macrophages. A) Western blot analysis to detect caspase-1 and actin (reference) was performed on whole cell lysates from bone marrow derived macrophages that were treated for 2 hours with medium, RB50, or RB50ΔclpV (MOI 10). B) Quantification was performed to normalize signal to reference levels (caspase-1: actin).
 for 2 hours with or without the presence of the 3MA inhibitor.")
110 98 Figure clpv is required for IL-1β production. LPS primed bone marrow-derived macrophages were treated with media, RB50, RB50ΔclpV (MOI 10) for 2 hours with or without the presence of the 3MA inhibitor. Macrophages treated with 5mM ATP and cells that were not primed with LPS were included as controls. Western blot analysis was used to measure IL-1β in methanol-precipitated supernatant and whole cell lysate.
111 99 Figure clpv did not affect Casp1 mrna levels. RT-qPCR analysis was performed on samples extracted from bone marrow derived macrophages. Cultured macrophages were treated with RB50 or RB50ΔclpV (MOI 10) for 0.5, 1, or 2 hours, and fold induction of Casp1 was measured relative to untreated macrophages using the 2 -ΔΔCt method with Gapdh as a reference gene. Relative expression is shown as mean + standard error, * indicates p<0.05.
112 100 Figure clpv is necessary for in decreased levels of Nlrp3 protein. (A) Western blot analysis to detect caspase-1 and NLRP3 was performed on whole cell lysate collected from LPS primed bone marrow derived macrophages treated with media, RB50, or RB50ΔclpV. ). B) Quantification was performed to normalize signal to reference levels (Nlrp3: actin). RT-qPCR analysis was performed on samples extracted from cultured macrophages treated with RB50 or RB50ΔclpV (MOI 10) for 0.5, 1, or 2 hours, and fold induction of Nlrp3 C) in bone marrow- derived macrophages was measured relative to untreated macrophages using the 2 -ΔΔCt method with Gapdh as a reference gene. Relative expression is shown as mean + standard error, *p<0.05.

 for 2 hours.")
113 101 Figure clpv is required for induction of autophagy and degradation of caspase-1. Bone marrow derived macrophages were incubated with media or 10mM 3MA and then treated with RB50 or RB50ΔclpV (MOI 10) for 2 hours. Western blot analysis was performed to detect caspase-1 A) and actin B).
114 102 Chapter 4 Hcp is necessary for normal B. bronchiseptica growth and morphology.
115 103 Abstract The Type VI secretion system (T6SS) is a T4 phage-like apparatus that is present in numerous species of Gram negative bacteria and has been shown to affect bacterial interspecies competition as well as virulence against eukaryotic host cells. Previously, we described a T6SS locus in B. bronchiseptica, and have shown that clpv, which encodes an ATPase that provides power for the system, is required for manipulation of macrophages and colonization in a mouse model. To determine the role of a gene encoding another conserved core component of the T6SS apparatus, hcp, an in-frame deletion was generated in B.bronchiseptica strain RB50. Deletion of hcp increased lag phase when grown in Stainer-Scholte media but not when the strain was grown in LB broth or medium supplemented with yeast extract. In addition, RB50Δhcp was more susceptible than wild type to polymyxin B, suggesting membrane alterations. Finally, hcp was required for normal morphology in B. bronchiseptica; the absence of hcp resulted in enlarged, filamentous bacteria, indicating dysfunctional cell replication. These results showed the importance of the tube component of the T6SS for maintenance of homeostasis within the bacterium.
116 104 Introduction The Type VI secretion system (T6SS) is found in a wide range of Gram negative bacteria (25% of proteobacteria) (25) and has been shown to affect bacterial interspecies competition (1) as well as virulence against eukaryotic host cells (4). Type VI secretion systems are encoded by a locus of conserved genes and share a large degree of homology with a T4 phage tail (5, 16). The proteins form a membrane spanning complex, complete with an ATPase /chaperone and secretion tube, and topped by a potential cell puncturing multimeric protein (Figure 4.1A) (16). The conserved core components include ClpV (24), VgrG (20), VipA/B (7), and Hcp (38). Hcp (haemolysin coregulated protein) dimerizes to form a hexameric ring structure (15, 31) that has been shown in vitro to polymerize and form the tube portion of the T6SS (3, 8). Hcp shares homology with the T4 phage gp19 tail protein (25) and forms a tube structure analogous to the contractile phage sheath (6). VgrG interacts with the polymerized Hcp tube, and the VipA/B polymerized sheath assembles in the cytoplasm, requiring ClpV ATPase activity (2). The VipA/B tail sheath has been shown to act as a scaffold for head-to-tail Hcp subunit assembly and is thought to propel the assembled Hcp nanotube and VgrG spike out of the cell and through the target cell membrane (9, 19). Hcp has been shown to be required for secretion of multiple classes of effectors, and its secretion is considered the hallmark of a functioning T6SS (41). Hcp can act as a chaperone for effectors, such as the Tse2 toxin in P. aeruginosa, as well as serving to stabilizing effectors that interact with VgrG through the PAAR motif, such as Tse5 and
117 105 Tse6 (37, 41). Hcp is thought to enable the transport and release of these effectors and is not thought to interact with them in their final, active conformation. Hcp is also required for virulence against amoeba (44), yeast (18), host cells (45), and in some systems, plays a role in quorum sensing (21) and biofilm formation when the T6SS is necessary (22). In addition, Hcp has been shown to be necessary for bacterial growth (1) as well as under stressed environmental conditions (17, 42). We investigated the role of hcp in B. bronchiseptica strain RB50. B. bronchiseptica is closely related to the subspecies B. pertussis and B. parapertussis, the causative agents of whooping cough (28). Therefore, understanding the pathogenesis of B. bronchiseptica infection can reveal bacterial strategies for evading and manipulating the host immune response and can lead to novel treatments for both human and veterinary respiratory diseases. Previous research in the lab has been performed to characterize the T6SS locus in RB50 (Figure 4.1 B). The locus consists of 26 genes, including all the core components as well as 13 novel, Bordetella specific genes (40). SWISS-Model analysis indicates that the B. bronchiseptica Hcp shares strong structural similarity (RMSD=0.06) with the P. aeruginosa PAO1 Hcp crystal structure (Figure 4.2A) (36). Recombinant Hcp (Figure 4.2B, L. Bendor, unpublished) was imaged using negative stain TEM and shown to have the characteristic hexameric shape associated with the protein complex. Preliminary experiments with the protein showed that it was immunogenic, indicating that its presence could shape the immune response against the T6SS (36). Previous work has shown that clpv is required for in vitro macrophage cytotoxicity as well as induction of interleukin (IL)-1β, IL-6, IL-17, and IL-10 production
118 106 in J774 macrophages (40). clpv is necessary for alterations in cytokine production, lung pathology, delayed lower respiratory tract clearance, and long term nasal colonization in the mouse model (40). Therefore, we are interested in determining the role played by hcp, another key T6SS component and have examined how disruption of this gene can influence membrane stability and bacterial growth and morphology. Materials and Methods Bacterial Strains and Growth B. bronchiseptica strain RB50 (12), RB50ΔclpV (40), and RB50Δhcp were maintained on Bordet Gengou (BG) agar plates (Difco) containing 10% sheep blood (Hema Resources) with 20 µg/ml streptomycin (Sigma). E. coli K-12 were maintained on Luria Bertani (LB) plates and cultured in LB broth. B. bronchiseptica strains were cultured in Stainer-Scholte (39) or LB liquid media until mid-log phase, shaking overnight at 37ºC. Growth curves were performed as previously described (32). Construction of RB50Δhcp Strain The RB50Δhcp strain was constructed using an allelic exchange strategy as previously described (40). The first 3 codons of hcp (BB0802) and the 755 base pairs (bp) upstream were amplified via PCR using primers flanked with BamHI on the 5 end and EcoRI on the 3 end (Table 1). The last 6 codons of hcp and the 584 bp downstream were amplified via PCR using primers flanked with EcoRI on the 5 end and BamHI on the 3 end (Table 1). These fragments were PCR purified (Qiagen), BamHI digested
119 107 (New England Biolabs), gel purified (Qiagen), and ligated overnight at 4 C (New England Biolabs), and amplified with the 5 F and 3 R primers as described above. The 1,364 bp knock-out construct was then ligated into the TOPO-TA vector (Invitrogen), transformed into Mach1 DH5α cells (Invitrogen), and verified by sequencing. The 1,364 bp construct was digested from TOPO-TA, gel-purified, and ligated overnight into the BamHI-digested pss4245 allelic exchange vector (courtesy of S. Stibitz). Triparental mating was conducted with DH5α containing pss4545δhcp, DH5α containing pss1827, and B. bronchiseptica strain RB50 grown on BG plates with 50 mm MgSO4 (Bvg conditions). The mating was performed for 4 hrs on a BG-10 mm MgCl2-50 mm MgSO4 plate at 37 C. B. bronchiseptica containing pss4245δhcp was positively selected by growth on BG-streptomycin-kanamycin-50 mm MgSO4 plates and incubated for 2 days at 37 C, which was performed twice. The resulting colonies were streaked onto BG plates and incubated for 2 days at 37 C, which resulted in colonies lacking pss4245 and containing either the wild type hcp or knockout construct. Colonies were then screened for the presence of either the wild type or knockout construct using PCR. The absence of pss4245 was confirmed by growth on BG-streptomycin plates and lack of growth on BG-kanamycin plates. Polymyxin B Susceptibility Assay Cultures of RB50, RB50ΔclpV and RB50Δhcp were diluted to 10 4 CFU/mL into a final 100 µl volume of PBS or PBS containing 0.09, 0.19, 0.39, 0.78, 1.56, 3.12, 6.25, 12.5, or 25 µg/ml polymyxin B. Suspensions were incubated for 2 hours at 37 C, following which, the number of organisms remaining in each sample was determined by quantitative culture on BG agar plates.
120 108 Brightfield and Transmission Electron Microscopy RB50 and RB50Δhcp cells grown to mid-log phase in either LB or Stainer- Scholte broth were visualized using an Olympus BX61 microscope. One ml of RB50 and RB50Δhcp cells grown to mid-log phase in either LB or Stainer-Scholte broth were centrifuged at 10,000 x g for 1 minute. The cells were fixed overnight at 4ºC in 2% glutaraldehyde in 0.05 M sodium cacodylate, ph 7.2. The pellets were washed 3 times in 0.1 M sodium cacodylate and incubated for 1 hour in 2% osmium tetroxide in 0.1 M sodium cacodylate, ph 7.2. Then the pellets were washed once with 0.1 M sodium cacodylate, 2 times with dh20 water, and stained for 1 hour in 2% aqueous uranyl acetate. The samples were dehydrated in acetone and then infiltrated with Epon araldite resin for incubation overnight at 60ºC. Ultrathin sections of the embedded sample were cut onto grids, and samples were visualized using the JEM 1200 EXII electron microscope (JEOL). Statistical Analysis Data analysis was performed using a two-way Student s T-test, with significant values of p>0.05. Results Deletion of hcp from B.bronchiseptica strain RB50 To investigate the role of hcp in B. bronchiseptica, we constructed an in-frame deletion of hcp (BB0802) in B. bronchiseptica strain RB50 using the pss4245 allelic exchange vector (10) to generate RB50Δhcp (Figure 4.3A). PCR was performed to
121 109 confirm the deletion; amplification of the wild type hcp gene resulted in a 1,945 bp product, and amplification of the region after deletion of hcp resulted in a 1,364 bp product (Figure 4.3B). These results indicate that the RB50Δhcp strain was successfully generated and could be used for further experimentation. hcp contributes to bacterial growth in a Bvg independent, media specific manner When assessing the growth of RB50Δhcp in Stainer-Scholte broth (39), a defined medium used to culture bordetellae and optimize production of virulence factors, we observed that the mutant lacking hcp had an increased time in lag phase in comparison to wild type RB50 (Figure 4.4A). Because bacteria use the lag phase of growth to adapt to new environmental conditions and synthesize proteins and metabolites necessary for cell replication, it is possible that the presence of hcp is necessary for the function of these pathways. Therefore, RB50 and RB50Δhcp were cultured in the nutrient rich LB broth, and growth was assessed over time (11). When RB50Δhcp was cultured in LB broth, the lag phase was decreased (6 hours versus 12 hours), indicating that the hcp dependent alteration in growth dynamics was medium specific (Figure 4.4B). The BvgAS two-component system in Bordetella serves as a master regulator of gene sets, including virulence factors such as adenylate cyclase toxin in the Bvg + phase as well as motility genes such as flagellin in the Bvg - phase, and bipa in the Bvg i phase (12, 32). Because Stainer-Scholte media was developed to maximize virulence factor production, the bordetellae grow in the Bvg + phase when cultured in Stainer-Scholte broth. However, different sets of genes expressed in the various Bvg phases, including the Bvg - phase, which enables the bordetellae to grow under nutrient limiting conditions (13, 33). To determine whether the Bvg phase of the bacteria contributed to the effect of
122 110 hcp on length of lag phase, RB50 and RB50Δhcp were cultured in Stainer-Scholte broth supplemented with 40mM MgSO4 to shift the phase to Bvg- (Figure 4.4C. However, the change in Bvg phase did not decrease the lag phase of RB50Δhcp, indicating that the effect of hcp on bacterial growth kinetics was not due to Bvg-dependent production of virulence factors. RB50Δhcp showed decreased lag phase when grown in a 1:1 mixture Stainer-Scholte and LB broth (6 hours versus the original 12 hours) (Figure 4.4D), illustrating that the mutant adapted more rapidly to growth in LB broth. The addition of yeast extract to Stainer-Scholte also for a shorter lag phase for RB50Δhcp to 4 hours (Figure 4.4E). hcp is required for resistance to polymyxin B Changes in growth kinetics could be caused by disruption of the bacterial membrane. In the absence of hcp, the other T6SS structural components could form a dysfunctional complex, changing architecture, composition, or properties of the outer and inner membrane. Alternatively, without hcp acting as a chaperone, T6SS effectors could be released into the cytoplasm, causing the misfolding of proteins or oxidative stress responses which have been shown to alter membrane properties (34). To test whether hcp is required for membrane stability, RB50 and RB50Δhcp were treated with multiple concentrations of polymyxin B (Figure 4.5). Polymyxin B is an amphipathic molecule that works through indirect interaction with the lipid A and subsequently acts to perturb the bacterial membrane (14). Susceptibility to polymyxin B has been shown to be associated with changes in outer membrane protein expression (26, 27). RB50Δhcp was significantly more susceptible to polymyxin B than RB50 (decreased below the threshold of detection after exposure to 0.39µg/mL). These data show that hcp was necessary for
123 111 resistance to killing with this cationic protein and suggested that hcp plays a role in membrane stability, which could explain effects on growth. hcp is required for normal morphology of B. bronchiseptica To examine the growth defect more closely, RB50Δhcp was grown in LB and Stainer-Scholte media, and the culture turbidity and number of colony forming units (cfu) were measured over time (Figure 4.6 A,B). Although RB50Δhcp grown in Stainer- Scholte media has an extended lag phase in comparison to RB50Δhcp grown in LB broth, the bacteria grown in Stainer-Scholte media have equivalent numbers of cfu/ml to those grown in LB after 12 and 24 hours and significantly higher numbers of viable bacteria at the 6 and 18 hour timepoints. RB50Δhcp grown in Stainer-Scholte has an increased ratio of cfu/ml to optical density (Figure 4.6 C) in comparison to RB50Δhcp grown in LB broth. These data suggest that growing RB50Δhcp in LB broth could enable the bacteria to increase in biomass without the generation of more viable cells. These data could indicate that in the absence of hcp, when grown under nutrient rich conditions, B. bronchiseptica could be replicating normally and then dying, or the bacteria could be enlarged due to continued cell growth without division. Gram stain pictures and quantification of area indicated that RB50Δhcp were filamentous in shape and were 4 times larger in area than the wild type when grown on BG agar (Figure 4.7 A,B). Therefore, in the absence of hcp, filamentation occurs in nutrient rich conditions. These data added further evidence to the possibility of growth without cell division by illustrating that the presence of hcp affected cell morphology. Therefore, we investigated whether the RB50Δhcp bacteria grown in LB broth became elongated and filamentous (increased nutrients partially enabling the bacteria to
124 112 increase in size but still unable to divide), while RB50Δhcp grown in Stainer-Scholte medium did not (no increase in cell size or division). Transmission electron microscopy analysis was performed and showed that RB50Δhcp bacteria did grow as filaments in comparison to wild type when cultured in LB broth (Figure 4.8 A,B). The RB50Δhcp cells cultured in Stainer-Scholte broth comprised a more heterogeneous population of both fibrous bacteria and bacteria which were closer to wild type in size but with a more rounded shape (Figure 4.8 C,D). These findings could indicate that when grown in Stainer-Scholte, the bacteria lacking hcp do not form filaments as readily, or they become filamentous but can undergo some cell division as they reach the end of lag phase. Overall, these results suggest that hcp is required for normal B. bronchiseptica cell replication. Discussion This work has uncovered a role for Hcp, a core component of the T6SS, in bacterial function and viability of B. bronchiseptica. These data underscore the importance of understanding the roles of individual secretion system components in relation to the maintenance of global homeostasis of multiple bacterial systems rather than simply in the context of the system function and secretion of effectors. In other systems, bacterial filamentation has been shown to occur in response to the presence of bacterial stress (23). One possible explanation for the observed phenotype is that the disruption of a secretion system that spans the outer and inner membrane could create membrane stress. Also, because of the Hcp chaperone activity,
125 113 T6SS effector proteins could be released into the cytoplasm, resulting in damage or even triggering the misfolded protein response. This possibility could be tested by pulling down Hcp in the cytoplasm to determine which proteins interact, or putative effectors in the cytoplasm could be detected using antibodies generated against recombinant proteins encoded by the T6SS locus. Future directions also include determining how the absence of hcp creates problems with cell replication. To directly show that hcp is required for normal septal formation, visualization of the cell division process could be performed via microscopy using a membrane dye, such as FM 46-4 (43) or immunofluorescence with antibodies against FtsZ, a tubulin like protein that enables Z ring assembly (30). The absence of hcp could result in induction of SOS response, which has been shown to halt cell division in the presence of DNA damage. The initiation of this pathway could occur through potential damaging agents generated by cytoplasmic effectors, direct damage to the nucleoid (tssd) encodes a putative effector with nuclease activity), or via crosstalk with the aforementioned stress pathways. Overall, these findings suggested that Hcp, a single secretion system component, can alter other critical functions besides virulence and illustrates the drastic alterations which can be facilitated by subtle disruptions of protein apparati. These data open up new avenues of research in defining how components of the T6SS can affect integral processes of bacterial physiology.
126 114 Author Contributions Sarah J. Muse 1,2, Yury V. Ivanov 1, Kai Hu 1,3, Liron Bendor 1,3, Eric T. Harvill 1 1 Department of Veterinary and Biomedical Sciences, The Pennsylvania State University, University Park, PA Graduate Program in Biochemistry, Microbiology and Molecular Biology 3 Graduate Program in Genetics Conceived and designed experiments: SJM, YVI, ETH Performed the experiments: SJM, YVI, KH, LB Analyzed the data: SJM, YVI, KH, ETH Wrote the paper: SJM, ETH
127 115 References 1. Alteri, C. J., S. D. Himpsl, S. R. Pickens, J. R. Lindner, J. S. Zora, J. E. Miller, P. D. Arno, S. W. Straight, and H. L. Mobley. Multicellular bacteria deploy the type VI secretion system to preemptively strike neighboring cells. PLoS Pathog 9:e Aschtgen, M. S., M. Gavioli, A. Dessen, R. Lloubes, and E. Cascales. The SciZ protein anchors the enteroaggregative Escherichia coli Type VI secretion system to the cell wall. Mol Microbiol 75: Ballister, E. R., A. H. Lai, R. N. Zuckermann, Y. Cheng, and J. D. Mougous In vitro self-assembly of tailorable nanotubes from a simple protein building block. Proc Natl Acad Sci U S A 105: Bartonickova, L., T. Sterzenbach, S. Nell, F. Kops, J. Schulze, A. Venzke, B. Brenneke, S. Bader, A. D. Gruber, S. Suerbaum, and C. Josenhans. Hcp and VgrG1 are secreted components of the Helicobacter hepaticus type VI secretion system and VgrG1 increases the bacterial colitogenic potential. Cell Microbiol 15: Basler, M., B. T. Ho, and J. J. Mekalanos. Tit-for-tat: type VI secretion system counterattack during bacterial cell-cell interactions. Cell 152: Basler, M., M. Pilhofer, G. P. Henderson, G. J. Jensen, and J. J. Mekalanos. Type VI secretion requires a dynamic contractile phage tail-like structure. Nature 483: Bonemann, G., A. Pietrosiuk, A. Diemand, H. Zentgraf, and A. Mogk Remodelling of VipA/VipB tubules by ClpV-mediated threading is crucial for type VI protein secretion. Embo J 28: Bonemann, G., A. Pietrosiuk, and A. Mogk. Tubules and donuts: a type VI secretion story. Mol Microbiol 76: Brunet, Y. R., J. Henin, H. Celia, and E. Cascales. Type VI secretion and bacteriophage tail tubes share a common assembly pathway. EMBO Rep 15: Buboltz, A. M., T. L. Nicholson, L. S. Weyrich, and E. T. Harvill Role of the type III secretion system in a hypervirulent lineage of Bordetella bronchiseptica. Infect Immun 77: Conover, M. S., C. J. Redfern, T. Ganguly, N. Sukumar, G. Sloan, M. Mishra, and R. Deora. BpsR modulates Bordetella biofilm formation by negatively regulating the expression of the Bps polysaccharide. J Bacteriol 194: Cotter, P. A., and J. F. Miller BvgAS-mediated signal transduction: analysis of phase-locked regulatory mutants of Bordetella bronchiseptica in a rabbit model. Infect Immun 62: Cotter, P. A., and J. F. Miller A mutation in the Bordetella bronchiseptica bvgs gene results in reduced virulence and increased resistance to starvation, and identifies a new class of Bvg-regulated antigens. Mol Microbiol 24: Daugelavicius, R., E. Bakiene, and D. H. Bamford Stages of polymyxin B interaction with the Escherichia coli cell envelope. Antimicrob Agents Chemother 44: Douzi, B., S. Spinelli, S. Blangy, A. Roussel, E. Durand, Y. R. Brunet, E. Cascales, and C. Cambillau. Crystal structure and self-interaction of the type VI secretion tail-tube protein from enteroaggregative Escherichia coli. PLoS One 9:e Filloux, A. The rise of the Type VI secretion system. F1000Prime Rep 5: Gueguen, E., E. Durand, X. Y. Zhang, Q. d'amalric, L. Journet, and E. Cascales. Expression of a Type VI Secretion System Is Responsive to Envelope Stresses through the OmpR Transcriptional Activator. PLoS One 8:e66615.
128 18. Haapalainen, M., H. Mosorin, F. Dorati, R. F. Wu, E. Roine, S. Taira, R. Nissinen, L. Mattinen, R. Jackson, M. Pirhonen, and N. C. Lin. Hcp2, a secreted protein of the phytopathogen Pseudomonas syringae pv. tomato DC3000, is required for fitness for competition against bacteria and yeasts. J Bacteriol 194: Hachani, A., L. P. Allsopp, Y. Oduko, and A. Filloux. The VgrG Proteins Are "a la Carte" Delivery Systems for Bacterial Type VI Effectors. J Biol Chem 289: Hachani, A., N. S. Lossi, A. Hamilton, C. Jones, S. Bleves, D. Albesa-Jove, and A. Filloux. Type VI secretion system in Pseudomonas aeruginosa: secretion and multimerization of VgrG proteins. J Biol Chem 286: Ishikawa, T., P. K. Rompikuntal, B. Lindmark, D. L. Milton, and S. N. Wai Quorum sensing regulation of the two hcp alleles in Vibrio cholerae O1 strains. PLoS One 4:e Jones, C., L. Allsopp, J. Horlick, H. Kulasekara, and A. Filloux. Subinhibitory concentration of kanamycin induces the Pseudomonas aeruginosa type VI secretion system. PLoS One 8:e Jones, T. H., K. M. Vail, and L. M. McMullen. Filament formation by foodborne bacteria under sublethal stress. Int J Food Microbiol 165: Kapitein, N., G. Bonemann, A. Pietrosiuk, F. Seyffer, I. Hausser, J. K. Locker, and A. Mogk. ClpV recycles VipA/VipB tubules and prevents non-productive tubule formation to ensure efficient type VI protein secretion. Mol Microbiol 87: Leiman, P. G., M. Basler, U. A. Ramagopal, J. B. Bonanno, J. M. Sauder, S. Pukatzki, S. K. Burley, S. C. Almo, and J. J. Mekalanos Type VI secretion apparatus and phage tail-associated protein complexes share a common evolutionary origin. Proc Natl Acad Sci U S A 106: Llobet, E., C. March, P. Gimenez, and J. A. Bengoechea Klebsiella pneumoniae OmpA confers resistance to antimicrobial peptides. Antimicrob Agents Chemother 53: Macfarlane, E. L., A. Kwasnicka, M. M. Ochs, and R. E. Hancock PhoP-PhoQ homologues in Pseudomonas aeruginosa regulate expression of the outer-membrane protein OprH and polymyxin B resistance. Mol Microbiol 34: Mattoo, S., and J. D. Cherry Molecular pathogenesis, epidemiology, and clinical manifestations of respiratory infections due to Bordetella pertussis and other Bordetella subspecies. Clin Microbiol Rev 18: Mizuno, T., and S. Mizushima Signal transduction and gene regulation through the phosphorylation of two regulatory components: the molecular basis for the osmotic regulation of the porin genes. Mol Microbiol 4: Monahan, L. G., A. T. Liew, A. L. Bottomley, and E. J. Harry. Division site positioning in bacteria: one size does not fit all. Front Microbiol 5: Mougous, J. D., M. E. Cuff, S. Raunser, A. Shen, M. Zhou, C. A. Gifford, A. L. Goodman, G. Joachimiak, C. L. Ordonez, S. Lory, T. Walz, A. Joachimiak, and J. J. Mekalanos A virulence locus of Pseudomonas aeruginosa encodes a protein secretion apparatus. Science 312: Nicholson, T. L., A. M. Buboltz, E. T. Harvill, and S. L. Brockmeier Microarray and functional analysis of growth phase-dependent gene regulation in Bordetella bronchiseptica. Infect Immun 77: Porter, J. F., R. Parton, and A. C. Wardlaw Growth and survival of Bordetella bronchiseptica in natural waters and in buffered saline without added nutrients. Appl Environ Microbiol 57:
129 34. Raivio, T. L Envelope stress responses and Gram-negative bacterial pathogenesis. Mol Microbiol 56: Rowley, G., M. Spector, J. Kormanec, and M. Roberts Pushing the envelope: extracytoplasmic stress responses in bacterial pathogens. Nat Rev Microbiol 4: Saxena, M Bordetella Bronchiseptica Gene BB0802 Encodes a Hexameric Immunogenic Protein;A Hcp Homologue. The Pennsylvania State University, University Park, PA. 37. Silverman, J. M., D. M. Agnello, H. Zheng, B. T. Andrews, M. Li, C. E. Catalano, T. Gonen, and J. D. Mougous. Haemolysin coregulated protein is an exported receptor and chaperone of type VI secretion substrates. Mol Cell 51: Silverman, J. M., Y. R. Brunet, E. Cascales, and J. D. Mougous. Structure and regulation of the type VI secretion system. Annu Rev Microbiol 66: Stainer, D. W., and M. J. Scholte A simple chemically defined medium for the production of phase I Bordetella pertussis. J Gen Microbiol 63: Weyrich, L. S., O. Y. Rolin, S. J. Muse, J. Park, N. Spidale, M. J. Kennett, S. E. Hester, C. Chen, E. G. Dudley, and E. T. Harvill. A Type VI secretion system encoding locus is required for Bordetella bronchiseptica immunomodulation and persistence in vivo. PLoS One 7:e Whitney, J. C., C. M. Beck, Y. A. Goo, A. B. Russell, B. N. Harding, J. A. De Leon, D. A. Cunningham, B. Q. Tran, D. A. Low, D. R. Goodlett, C. S. Hayes, and J. D. Mougous. Genetically distinct pathways guide effector export through the type VI secretion system. Mol Microbiol. 42. Wu, C. F., J. S. Lin, G. C. Shaw, and E. M. Lai. Acid-induced type VI secretion system is regulated by ExoR-ChvG/ChvI signaling cascade in Agrobacterium tumefaciens. PLoS Pathog 8:e Yao, Z., D. Kahne, and R. Kishony. Distinct single-cell morphological dynamics under beta-lactam antibiotics. Mol Cell 48: Zheng, J., B. Ho, and J. J. Mekalanos. Genetic analysis of anti-amoebae and antibacterial activities of the type VI secretion system in Vibrio cholerae. PLoS One 6:e Zhou, Y., J. Tao, H. Yu, J. Ni, L. Zeng, Q. Teng, K. S. Kim, G. P. Zhao, X. Guo, and Y. Yao. Hcp family proteins secreted via the type VI secretion system coordinately regulate Escherichia coli K1 interaction with human brain microvascular endothelial cells. Infect Immun 80:
.")
 B.")
130 118 Figure 4.1. The Type VI Secretion System (T6SS). A) Model of formation of the T6SS Apparatus. B) B. bronchiseptica strain RB50 T6SS locus.
 Negative Stain TEM imaging of recombinant B. bronchiseptica Hcp protein.")

131 119 Figure 4.2. Hcp protein. A) Crystal Structure of Hcp1 protein from P. aeruginosa. B) Negative Stain TEM imaging of recombinant B. bronchiseptica Hcp protein. C) SWISS Model Alignment of Hcp1 from P. aeruginosa PAO1 (PDB entry 1Y12) and model of B. bronchiseptica BB802. Reprinted with permission from American Association for the Advancement of Science, Science Jun 9;312(5779):
132 120 Figure 4.3. Construction and confirmation of RB50Δhcp. A) Schematic for generating RB50Δhcp using the pss4245 allelic exchange vector. B) PCR confirmation of knock out generation. Size markers are designated on the left.
133 121 Figure 4.4. hcp contributed to bacterial growth. OD600 values were measured for RB50 and RB50Δhcp grown in Stainer-Scholte broth (A) and LB broth (B) as well as C) Stainer Scholte broth supplemented with 40mM MgS04. OD600 values were measured for RB50 and RB50Δhcp grown in D) a 1:1 mixture of Stainer Scholte broth and LB broth and E) Stainer Scholte supplemented with yeast extract. * indicates p<0.05
134 122 Figure 4.5. hcp is required for polymyxin B resistance. RB50 and RB50Δhcp were incubated for 2h at 37 C in PBS with multiple concentrations of polymyxin B (0-25 µg/ml, twofold dilutions) and plated on BG agar for bacterial enumeration.
135 123 Figure 4.6. RB50Δhcp optical density did not correspond to colony forming units. Growth curve of RB50Δhcp in LB broth and Stainer Scholte measuring A) OD600and B) CFU/mL. Ratio of CFU/mL vs. OD600 is represented by C). * indicates p<0.05
136 124
 Gram-stain of RB50 (left) and RB50Δhcp (right) grown on BG agar.")

137 125 Figure 4.7. hcp is required for normal morphology in B. bronchiseptica. A) Gram-stain of RB50 (left) and RB50Δhcp (right) grown on BG agar. B) Quantification of bacterial volume of RB50 and RB50Δhcp using ImageJ for length measurement and approximating cylindrical shape. Transmission electron microscopy images of RB50 and RB50Δhcp grown to mid-log phase in Stainer Scholte (C,D) and LB broth (E,F).
138 Figure 4.8. Model of hcp in B. bronchiseptica growth. 126
139 127 Table 4.1. Primer Sequences. Primer Name Sequence Hcp KO 5 F BamHI AGA GGA TCC GTT CGT TCA AGC TGT ACG Hcp KO 5 R - EcoRI Hcp KO 3 F - EcoRI Hcp KO 3 R Bam HI CAC AAG GAC TGG ACC GAT GAA TTC TAG AAA GAA TTC CTG GTC GCG TGG GAC ATC TTC TCG ACG GCA GCT ATG GGA TCC TAA
140 Chapter 5 Summary and Significance.
141 129 Synopsis The development of new treatments to fight infection depends on understanding the mechanisms by which bacteria use virulence factors to survive and propagate between host systems. The goal of this work was to investigate how B. bronchiseptica uses its virulence factors to survive and propagate. Multiple strategies pursued by the bacterium were investigated, such as novel virulence and transmission properties for arnt, a mechanism by which B. bronchiseptica can alter inflammasome cell signaling in host immune cells, and how hcp, a Type VI secretion system component, can have important functions for the bacterium in the absence of host interaction. These experiments shed light on strategies used by B. bronchiseptica to survive, manipulate the host in a beneficial manner, and facilitate transmission to another host. Enzymatic Modification of the Lipid A by an ArnT Protects B. bronchiseptica Against Cationic Peptides and Is Required for Transmission By investigating a strategy used by pathogens to adapt to their host environment, we uncovered a novel virulence property for arnt, which encodes a protein that modifies the lipid A by adding glucosamine. As previously observed in other systems, arnt is required for resistance to cationic antimicrobial peptides; however, novel findings from our studies show that arnt is also required for transmission between hosts, indicating how virulence factors can influence multiple aspects of a pathogen s life cycle.
142 130 Summary and Implications To investigate the role played by the glucosamine (GlcN) lipid A modification, we generated the RB50ΔarnT deletion mutant and verified that the arnt was required for the GlcN addition using mass spectrometry (Figure 2.1A-E). Interestingly, the presence of arnt did not affect TLR4 signaling (measured by TNF-α levels) in J774 cells (Figure 2.2A), which differed from the phenotype observed from the B. pertussis arnt deficient mutant which was used to infect human THP-1 cells (12). However, this distinction could be due to the fact that infection with B. bronchiseptica stimulates TLR4 more highly than infection with B. pertussis (12, 13), as well as the fact that there are other structural differences (i.e. B. pertussis has a lipooligosaccharide) (2). The lipid A modification provided resistance to antimicrobial peptides (Figure 2.3), which is a trait that may have evolved to counter the barrage of cationic peptides encountered as the bacteria initially try to colonize the mucosa (6, 8). However, the ability of arnt to affect colonization in the respiratory tract depends on the dose of bacteria administered. Using low dose model of infection illustrates that glucosamine modification could enable a smaller number of bacteria to establish an infection in the nasal cavity (Figure 2.5), whereas a difference is not detected using the more standard high dose model (Figure 2.4). This fact draws attention to subtle bacterial properties that may ensure survival during a natural infection but would not be detected using the high dose infection model. One of the most interesting findings indicates that arnt is required for transmission between hosts (Figure 2.6) without affecting cell recruitment (Figure 2.7A)
143 131 or bacterial shedding (Figure 2.7B). Pursuing how bacteria can influence transmission can lead to the development of better treatments that prevent the spread of disease rather than limiting pathology or symptoms. Examination of how virulence factor modifications could affect transmission could also be important for vaccine development (17), especially given the recent B. pertussis resurgence (14) and the consideration of using detoxified endotoxin (4) as a potential additional component for improving the current acellular vaccine. Future Directions The results of these experiments are the first to indicate that modifications of the bacterial lipid A could influence transmission. A major question in the study of infectious disease, which is difficult to model, concerns how pathogens spread between hosts. Previous work in the field has shown a correlation between neutrophil recruitment to the site of infection and levels of transmission and shedding (9, 20). However, the influence of lipid A modifications, if not altering TLR4 signaling, indicates that other factors could influence B. bronchiseptica transmission, such as the promoting the ability to reliably colonize a potential host rather than increasing the numbers of bacteria released from the index animal through immune system manipulation. One possible future direction includes determining whether the lipid A modification can affect transmission using defensins or other antimicrobial peptide deficient mice as well as assaying for changes in local cytokine profiles linked to transmission rather than broader cell recruitment.
144 132 Bordetella bronchiseptica and the Inflammasome Another aspect of bacterial utilization of virulence factors is how they can alter the immune inflammatory response to their benefit. IL-1 signaling has been shown to be required during bordetellae infection (7, 16). Previous research using the mouse model has shown that IL-1R is required for control of B. bronchiseptica colonization (Figure 3.3 A,B,C) as well as survival of the host during infection (Figure 3.2 A,B), preventing rapid expansion of bacterial numbers that presumably escape the respiratory tract and kill the host through septic shock. However, when the mice are inoculated with a B. bronchiseptica strain lacking a functional T6SS, the subsequent infection in the absence of IL-1R is no longer lethal (Figure 3.2 C,D). No T6SS specific differences in IL-1β levels were detected early in infection or throughout the time course (Figure 3.4 A,B). However, activation of IL-1β could still be involved in the T6SS specific response. To investigate the involvement of IL-l signaling pathway upstream of the IL-1R, we explored the relationship between the T6SS and the caspase-1 inflammasome (22) in mouse model as well as in cultured macrophages. Summary and Implications The findings of these experiments indicate that a complex relationship exists between caspase-1 and the B. bronchiseptica T6SS (Figure 3.10). In cultured macrophages, a decrease in caspase-1 protein was observed in the presence of a functional T6SS (Figure 3.11) which did not result in a corresponding decrease in activated IL-1β (Figure 3.12). However, caspase-1 has multiple other targets (10, 11),
145 133 and these data could indicate that the decrease in caspase-1 could affect the host immune response in other ways besides the hypothesized mechanism. The presence of the T6SS does not result in a decrease in Casp1 or Nlrp3 mrna levels (Figure 3.13, 3.14C) but does affect levels of Nlrp3 protein (Figure 3.14A,B), an important inflammasome component (5, 19, 22). These data raise the possibility that the entire inflammasome complex is being decreased. After treatment with the 3MA autophagy inhibitor(18), the decrease in caspase-1 is abrogated (Figure 3.15), suggesting that the autophagy pathway is required for the clpv-mediated decrease in caspase-1 levels. Results from the murine infectionshint that the clpv dependent decrease in caspase-1 observed in culture could also affect infection in the animal model..during B. bronchiseptica infection, clpv is required for colonization of the respiratory tract. However, in mice deficient in caspase-1, no reduction in colonization is observed. One explanation of this phenomenon is that wild-type B. bronchiseptica, which have a functional T6SS, are able to decrease caspase-1 levels in wild type mice, a future possibility to be explored. Future Directions These findings open up many new avenues in pursuing how a newly characterized secretion system can interact with an important immune signaling pathway. One of the main points of interest would be determining how the T6SS influences autophagy and thus levels of inflammasome proteins. If the observed effect occurs due to a direct interaction between the T6SS and the host immune pathways, creating a series of deletion
146 134 mutants lacking genes encoding putative T6SS effector proteins could determine whether a specific secreted protein is responsible for manipulating host responses. Identifying the T6SS effectors responsible for the decrease in inflammasome levels could also be accomplished by generating and screening a transposon library. It is also possible that disrupting the T6SS locus affects other bacterial virulence systems, so determining if other known virulence factors can mediate a similar effect is another strategy of interest. For example, mutants deficient in Type III secretion system function or Bvg phase locked mutants could be tested to determine if they affect inflammasome levels. Another important research aspect would be to determine the mechanism underlying the caspase-1 degradation, including measuring ubiquitinylation state and using pulldown assays to determine if bacterial effector proteins are interacting directly with the inflammasome. Pursuing these avenues of research would enable the scientific community to gain knowledge about important early immune responses as well as understanding novel bacterial secretion system mechanisms. Hcp is necessary for normal B. bronchiseptica growth and morphology The final aspect of this study involves determining how regulation of a virulence factor also affects bacterial systems and growth. In the simplest terms, a virulence factor is a component that enables the pathogen to inflict damage on the host (3). This perspective highlights the importance of bacterial function and viability in determining whether or not designated virulence systems are required for host-pathogen interactions.
147 In this study, we investigate a major component of the T6SS, Hcp, and how it affects the growth of B. bronchiseptica. 135 Summary and Implications The in-frame deletion strain of B. bronchiseptica that lacks hcp (Figure 4.3) has an increased lag phase in comparison to wild-type when grown in Stainer-Scholte broth which is compensated for by growth in the rich LB broth (Figure 4.4). However, RB50Δhcp has different optical density to CFU ratios in Stainer-Scholte and LB media (Figure 4.5) due to filamentous morphology (Figure 4.7). Altered resistance to Polymyxin B (Figure 4.5) could also indicate a membrane perturbation, possibly due to stress caused by a misassembled secretion system apparatus or due to a buildup of unchaperoned effector molecules in the cytoplasm. These results illustrate how disruption of one system can have large scale effects, such as filamentation, and can result in negative consequences for the pathogen. Future Directions Future directions include determining the mechanism underlying how the absence of hcp creates problems with bacterial growth and morphology. Directly testing whether hcp is required for normal septal formation could be performed using a membrane dye, such as FM 46-4, to directly visualize the division process (21) or immunofluorescence to visualize FtsZ polymerization during Z ring assembly (15).
148 136 Another important question to investigate would be whether the absence of hcp could result in induction of SOS response, which has been shown to halt cell division in the presence of DNA damage (1) or potentially cell envelope stress. Microarray analysis and comparisons of expression profiles would indicate an increase in genes having promoters with LexA boxes in RB50Δhcp versus wild type or possibly induction of the RpoS regulon in the case of an SOS response and an increase in expression of genes under the control of alternate sigma factors in the case of membrane or oxidative stress. The response pathway initiation could occur through potential damaging agents generated by cytoplasmic effectors, direct damage to the nucleoid (tssd encodes a putative effector with nuclease activity), or via crosstalk with the aforementioned stress pathways. This possibility could be tested by creating double mutant strains of bacteria lacking both hcp and the putative effector proteins and observing morphology and growth parameters. Conclusion In conclusion, this research has investigated multiple aspects of bacterial virulence and how the B. bronchiseptica pathogen can interact with the immune response. Pathogens use multiple strategies to ensure their survival, such as the ArnT mediated glucosamine lipid A modification, which alters a pathogen associated molecular pattern to improve resistance to an innate immune defense as well as enable them to transmit to a new host. The T6SS enables the bacterium to manipulate host processes to eliminate a key pro-inflammatory response. Absence of Hcp, a T6SS component generally considered in the context of secretion system specific functions, results in severe changes
149 137 in bacterial growth and morphology. These different aspects of pathogenesis illustrate the diversity and complexity of the ways in which bacterial pathogens interact with their host and the host immune response to survive. As always, further work is necessary to better understand bacterial virulence strategies and to develop disease prophylactics and treatments in the ever evolving fight against infection.
150 138 References 1. Baharoglu, Z., and D. Mazel. SOS, the formidable strategy of bacteria against aggressions. FEMS Microbiol Rev. 2. Burns, V. C., E. J. Pishko, A. Preston, D. J. Maskell, and E. T. Harvill Role of Bordetella O antigen in respiratory tract infection. Infect Immun 71: Casadevall, A., and L. A. Pirofski Host-pathogen interactions: redefining the basic concepts of virulence and pathogenicity. Infect Immun 67: Casella, C. R., and T. C. Mitchell Putting endotoxin to work for us: monophosphoryl lipid A as a safe and effective vaccine adjuvant. Cell Mol Life Sci 65: Costa, A., R. Gupta, G. Signorino, A. Malara, F. Cardile, C. Biondo, A. Midiri, R. Galbo, P. Trieu-Cuot, S. Papasergi, G. Teti, P. Henneke, G. Mancuso, D. T. Golenbock, and C. Beninati. Activation of the NLRP3 inflammasome by group B streptococci. J Immunol 188: Doss, M., M. R. White, T. Tecle, and K. L. Hartshorn. Human defensins and LL-37 in mucosal immunity. J Leukoc Biol 87: Dunne, A., P. J. Ross, E. Pospisilova, J. Masin, A. Meaney, C. E. Sutton, Y. Iwakura, J. Tschopp, P. Sebo, and K. H. Mills. Inflammasome activation by adenylate cyclase toxin directs Th17 responses and protection against Bordetella pertussis. J Immunol 185: Ganz, T Defensins: antimicrobial peptides of vertebrates. C R Biol 327: Gopinath, S., A. Hotson, J. Johns, G. Nolan, and D. Monack. The systemic immune state of super-shedder mice is characterized by a unique neutrophil-dependent blunting of TH1 responses. PLoS Pathog 9:e Jabir, M. S., N. D. Ritchie, D. Li, H. K. Bayes, P. Tourlomousis, D. Puleston, A. Lupton, L. Hopkins, A. K. Simon, C. Bryant, and T. J. Evans. Caspase-1 cleavage of the TLR adaptor TRIF inhibits autophagy and beta-interferon production during Pseudomonas aeruginosa infection. Cell Host Microbe 15: Lamkanfi, M., M. Kalai, X. Saelens, W. Declercq, and P. Vandenabeele Caspase-1 activates nuclear factor of the kappa-enhancer in B cells independently of its enzymatic activity. J Biol Chem 279: Marr, N., A. M. Hajjar, N. R. Shah, A. Novikov, C. S. Yam, M. Caroff, and R. C. Fernandez. Substitution of the Bordetella pertussis lipid A phosphate groups with glucosamine is required for robust NF-kappaB activation and release of proinflammatory cytokines in cells expressing human but not murine Toll-like receptor 4-MD-2-CD14. Infect Immun 78: Marr, N., A. Tirsoaga, D. Blanot, R. Fernandez, and M. Caroff Glucosamine found as a substituent of both phosphate groups in Bordetella lipid A backbones: role of a BvgAS-activated ArnT ortholog. J Bacteriol 190: Melvin, J. A., E. V. Scheller, J. F. Miller, and P. A. Cotter. Bordetella pertussis pathogenesis: current and future challenges. Nat Rev Microbiol 12: Monahan, L. G., A. T. Liew, A. L. Bottomley, and E. J. Harry. Division site positioning in bacteria: one size does not fit all. Front Microbiol 5: Park, J., Y. Zhang, A. M. Buboltz, X. Zhang, S. C. Schuster, U. Ahuja, M. Liu, J. F. Miller, M. Sebaihia, S. D. Bentley, J. Parkhill, and E. T. Harvill. Comparative genomics of the classical Bordetella subspecies: the evolution and exchange of virulenceassociated diversity amongst closely related pathogens. BMC Genomics 13:545.
151 17. Rolin, O., W. Smallridge, M. Henry, L. Goodfield, D. Place, and E. T. Harvill. Tolllike receptor 4 limits transmission of Bordetella bronchiseptica. PLoS One 9:e Tanida, I., T. Ueno, and E. Kominami LC3 and Autophagy. Methods Mol Biol 445: Tschopp, J., and K. Schroder. NLRP3 inflammasome activation: The convergence of multiple signalling pathways on ROS production? Nat Rev Immunol 10: Weyrich, L. S., O. Y. Rolin, S. J. Muse, J. Park, N. Spidale, M. J. Kennett, S. E. Hester, C. Chen, E. G. Dudley, and E. T. Harvill. A Type VI secretion system encoding locus is required for Bordetella bronchiseptica immunomodulation and persistence in vivo. PLoS One 7:e Yao, Z., D. Kahne, and R. Kishony. Distinct single-cell morphological dynamics under beta-lactam antibiotics. Mol Cell 48: Zhou, Y., J. Tao, H. Yu, J. Ni, L. Zeng, Q. Teng, K. S. Kim, G. P. Zhao, X. Guo, and Y. Yao. Hcp family proteins secreted via the type VI secretion system coordinately regulate Escherichia coli K1 interaction with human brain microvascular endothelial cells. Infect Immun 80:
152 Appendix A Parameters of B. bronchiseptica Infection in Relation to Caspase-1 Figure A.1. Measuring T6SS dependent caspase-1 activity. Bone marrow derived macrophages were incubated with media, RB50, or RB50ΔclpV (MOI 10) for 2 hours and treated with medium alone or the BafA1 autophagy inhibitor. Activity of caspase-1 was determined using fluormetric assay based on the detection of AFC (7-amino-4-trifluoromethyl coumarin). Bars represent the mean fluorescent units + standard error.
, and RB50ΔbscN (WD3) (inactive T3SS) at an MOI of 10 for 2 hours using β-actin loading")
153 141 Figure A.2. The Type III Secretion System is required for decreased levels of caspase-1 protein Immunoblot of Caspase-1 using RAW whole cell lysates from cells inoculated with RB50, RB50ΔclpV (inactive T6SS), and RB50ΔbscN (WD3) (inactive T3SS) at an MOI of 10 for 2 hours using β-actin loading control (S. Hester, unpublished).
Role of Antibodies in Immunity to Bordetella Infections
 INFECTION AND IMMUNITY, Apr. 2003, p. 1719 1724 Vol. 71, No. 4 0019-9567/03/$08.00 0 DOI: 10.1128/IAI.71.4.1719 1724.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Role of
INFECTION AND IMMUNITY, Apr. 2003, p. 1719 1724 Vol. 71, No. 4 0019-9567/03/$08.00 0 DOI: 10.1128/IAI.71.4.1719 1724.2003 Copyright 2003, American Society for Microbiology. All Rights Reserved. Role of
Bordetella evolution: lipid A and Toll-like receptor 4
 IEIIS Meeting minireview Bordetella evolution: lipid A and Toll-like receptor 4 Iain MacArthur 1, Paul B. Mann 2 *, Eric T. Harvill 2, Andrew Preston 1 1 Department of Molecular and Cellular Biology, University
IEIIS Meeting minireview Bordetella evolution: lipid A and Toll-like receptor 4 Iain MacArthur 1, Paul B. Mann 2 *, Eric T. Harvill 2, Andrew Preston 1 1 Department of Molecular and Cellular Biology, University
The Bvg Virulence Control System Regulates Biofilm Formation in Bordetella bronchiseptica
 JOURNAL OF BACTERIOLOGY, Sept. 2004, p. 5692 5698 Vol. 186, No. 17 0021-9193/04/$08.00 0 DOI: 10.1128/JB.186.17.5692 5698.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. The
JOURNAL OF BACTERIOLOGY, Sept. 2004, p. 5692 5698 Vol. 186, No. 17 0021-9193/04/$08.00 0 DOI: 10.1128/JB.186.17.5692 5698.2004 Copyright 2004, American Society for Microbiology. All Rights Reserved. The
The Pennsylvania State University. The Graduate School. College of Agricultural Science UNDERSTANDING HOW VACCINATION AND PARTICULAR VIRULENCE
 The Pennsylvania State University The Graduate School College of Agricultural Science UNDERSTANDING HOW VACCINATION AND PARTICULAR VIRULENCE FACTORS CONTRIBUTE TO BORDETELLA TRANSMISSION A Dissertation
The Pennsylvania State University The Graduate School College of Agricultural Science UNDERSTANDING HOW VACCINATION AND PARTICULAR VIRULENCE FACTORS CONTRIBUTE TO BORDETELLA TRANSMISSION A Dissertation
VACCINE-INDUCED-IMMUNITY-MEDIATED COMPETITION BETWEEN ENDEMIC BORDETELLAE AND HOST IMMUNITY AGAINST THEM
 The Pennsylvania State University The Graduate School College of Agricultural Sciences VACCINE-INDUCED-IMMUNITY-MEDIATED COMPETITION BETWEEN ENDEMIC BORDETELLAE AND HOST IMMUNITY AGAINST THEM A Dissertation
The Pennsylvania State University The Graduate School College of Agricultural Sciences VACCINE-INDUCED-IMMUNITY-MEDIATED COMPETITION BETWEEN ENDEMIC BORDETELLAE AND HOST IMMUNITY AGAINST THEM A Dissertation
Role of the Type III Secretion System in a Hypervirulent Lineage of Bordetella bronchiseptica
 INFECTION AND IMMUNITY, Sept. 2009, p. 3969 3977 Vol. 77, No. 9 0019-9567/09/$08.00 0 doi:10.1128/iai.01362-08 Copyright 2009, American Society for Microbiology. All Rights Reserved. Role of the Type III
INFECTION AND IMMUNITY, Sept. 2009, p. 3969 3977 Vol. 77, No. 9 0019-9567/09/$08.00 0 doi:10.1128/iai.01362-08 Copyright 2009, American Society for Microbiology. All Rights Reserved. Role of the Type III
Probing the Function of Bordetella bronchiseptica Adenylate Cyclase Toxin by Manipulating Host Immunity
 INFECTION AND IMMUNITY, Mar. 1999, p. 1493 1500 Vol. 67, No. 3 0019-9567/99/$04.00 0 Copyright 1999, American Society for Microbiology. All Rights Reserved. Probing the Function of Bordetella bronchiseptica
INFECTION AND IMMUNITY, Mar. 1999, p. 1493 1500 Vol. 67, No. 3 0019-9567/99/$04.00 0 Copyright 1999, American Society for Microbiology. All Rights Reserved. Probing the Function of Bordetella bronchiseptica
Impact of Antimicrobial Resistance on Human Health. Robert Cunney HSE HCAI/AMR Programme and Temple Street Children s University Hospital
 Impact of Antimicrobial Resistance on Human Health Robert Cunney HSE HCAI/AMR Programme and Temple Street Children s University Hospital AMR in Foodchain Conference, UCD, Dec 2014 Sir Patrick Dun s Hospital
Impact of Antimicrobial Resistance on Human Health Robert Cunney HSE HCAI/AMR Programme and Temple Street Children s University Hospital AMR in Foodchain Conference, UCD, Dec 2014 Sir Patrick Dun s Hospital
THE COST OF COMPANIONSHIP
 THE COST OF COMPANIONSHIP Jared Gillingham and Robert Burlage Concordia University School of Pharmacy Mequon, WI Synopsis: Infectious diseases are always a concern, but when you are a person in an at-risk
THE COST OF COMPANIONSHIP Jared Gillingham and Robert Burlage Concordia University School of Pharmacy Mequon, WI Synopsis: Infectious diseases are always a concern, but when you are a person in an at-risk
Phenotypic modulation of the Bvg+ phase is not required for pathogenesis and. transmission of Bordetella bronchiseptica in swine
 IAI Accepts, published online ahead of print on 12 December 2011 Infect. Immun. doi:10.1128/iai.06016-11 Copyright 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights
IAI Accepts, published online ahead of print on 12 December 2011 Infect. Immun. doi:10.1128/iai.06016-11 Copyright 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights
THE PENNSYLVANIA STATE UNIVERSITY SCHREYER HONORS COLLEGE DEPARTMENT OF BIOCHEMISTRY AND MOLECULAR BIOLOGY
 THE PENNSYLVANIA STATE UNIVERSITY SCHREYER HONORS COLLEGE DEPARTMENT OF BIOCHEMISTRY AND MOLECULAR BIOLOGY THE ROLE OF FIMBRIAE IN BORDETELLA COLONIZATION MARGARET CURRY DUNAGIN Spring 2010 A thesis submitted
THE PENNSYLVANIA STATE UNIVERSITY SCHREYER HONORS COLLEGE DEPARTMENT OF BIOCHEMISTRY AND MOLECULAR BIOLOGY THE ROLE OF FIMBRIAE IN BORDETELLA COLONIZATION MARGARET CURRY DUNAGIN Spring 2010 A thesis submitted
bvg Repression of Alcaligin Synthesis in Bordetella bronchiseptica Is Associated with Phylogenetic Lineage
 JOURNAL OF BACTERIOLOGY, Nov. 1995, p. 6058 6063 Vol. 177, No. 21 0021-9193/95/$04.00 0 Copyright 1995, American Society for Microbiology bvg Repression of Alcaligin Synthesis in Bordetella bronchiseptica
JOURNAL OF BACTERIOLOGY, Nov. 1995, p. 6058 6063 Vol. 177, No. 21 0021-9193/95/$04.00 0 Copyright 1995, American Society for Microbiology bvg Repression of Alcaligin Synthesis in Bordetella bronchiseptica
WHY IS THIS IMPORTANT?
 CHAPTER 20 ANTIBIOTIC RESISTANCE WHY IS THIS IMPORTANT? The most important problem associated with infectious disease today is the rapid development of resistance to antibiotics It will force us to change
CHAPTER 20 ANTIBIOTIC RESISTANCE WHY IS THIS IMPORTANT? The most important problem associated with infectious disease today is the rapid development of resistance to antibiotics It will force us to change
Consequences of Antimicrobial Resistant Bacteria. Antimicrobial Resistance. Molecular Genetics of Antimicrobial Resistance. Topics to be Covered
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
MID 23. Antimicrobial Resistance. Consequences of Antimicrobial Resistant Bacteria. Molecular Genetics of Antimicrobial Resistance
 Antimicrobial Resistance Molecular Genetics of Antimicrobial Resistance Micro evolutionary change - point mutations Beta-lactamase mutation extends spectrum of the enzyme rpob gene (RNA polymerase) mutation
Antimicrobial Resistance Molecular Genetics of Antimicrobial Resistance Micro evolutionary change - point mutations Beta-lactamase mutation extends spectrum of the enzyme rpob gene (RNA polymerase) mutation
Antimicrobial Resistance
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of empiric antimicrobial therapy Increased number of hospitalizations Increased length
Antimicrobial Resistance Acquisition of Foreign DNA
 Antimicrobial Resistance Acquisition of Foreign DNA Levy, Scientific American Horizontal gene transfer is common, even between Gram positive and negative bacteria Plasmid - transfer of single or multiple
Antimicrobial Resistance Acquisition of Foreign DNA Levy, Scientific American Horizontal gene transfer is common, even between Gram positive and negative bacteria Plasmid - transfer of single or multiple
Methicillin-Resistant Staphylococcus aureus
 Methicillin-Resistant Staphylococcus aureus By Karla Givens Means of Transmission and Usual Reservoirs Staphylococcus aureus is part of normal flora and can be found on the skin and in the noses of one
Methicillin-Resistant Staphylococcus aureus By Karla Givens Means of Transmission and Usual Reservoirs Staphylococcus aureus is part of normal flora and can be found on the skin and in the noses of one
Microarray and Functional Analysis of Growth Phase-Dependent Gene Regulation in Bordetella bronchiseptica
 INFECTION AND IMMUNITY, Oct. 2009, p. 4221 4231 Vol. 77, No. 10 0019-9567/09/$08.00 0 doi:10.1128/iai.00136-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Microarray and Functional
INFECTION AND IMMUNITY, Oct. 2009, p. 4221 4231 Vol. 77, No. 10 0019-9567/09/$08.00 0 doi:10.1128/iai.00136-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. Microarray and Functional
Bordetella bronchiseptica: A Candidate Mucosal Vaccine Vector
 University of Tennessee, Knoxville Trace: Tennessee Research and Creative Exchange Doctoral Dissertations Graduate School 5-2002 Bordetella bronchiseptica: A Candidate Mucosal Vaccine Vector Sreekumari
University of Tennessee, Knoxville Trace: Tennessee Research and Creative Exchange Doctoral Dissertations Graduate School 5-2002 Bordetella bronchiseptica: A Candidate Mucosal Vaccine Vector Sreekumari
Inefficient Toll-Like Receptor-4 Stimulation Enables Bordetella parapertussis to Avoid Host Immunity
 Inefficient Toll-Like Receptor-4 Stimulation Enables Bordetella parapertussis to Avoid Host Immunity Daniel N. Wolfe 1 a b, Anne M. Buboltz 1,2, Eric T. Harvill 1 * 1 Department of Veterinary and Biomedical
Inefficient Toll-Like Receptor-4 Stimulation Enables Bordetella parapertussis to Avoid Host Immunity Daniel N. Wolfe 1 a b, Anne M. Buboltz 1,2, Eric T. Harvill 1 * 1 Department of Veterinary and Biomedical
Bacterial Pneumonia in Sheep, The Domestic Bighorn Sheep Interface, and Research at ADRU
 Bacterial Pneumonia in Sheep, The Domestic Bighorn Sheep Interface, and Research at ADRU USAHA Committee on Sheep and Goats Providence, RI October 27, 2015 PLC M. A. Highland, DVM, DACVP, PhD candidate
Bacterial Pneumonia in Sheep, The Domestic Bighorn Sheep Interface, and Research at ADRU USAHA Committee on Sheep and Goats Providence, RI October 27, 2015 PLC M. A. Highland, DVM, DACVP, PhD candidate
Antimicrobial Resistance
 Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of Change in the approach to the administration of empiric antimicrobial therapy Increased
Antimicrobial Resistance Consequences of Antimicrobial Resistant Bacteria Change in the approach to the administration of Change in the approach to the administration of empiric antimicrobial therapy Increased
MATTHEW S. CONOVER. A Dissertation Submitted to the Graduate Faculty of WAKE FOREST UNIVERSITY GRADUATE SCHOOL OF ARTS AND SCIENCES
 AN EXAMINATION OF THE FUNCTION AND THE TRANSCRIPTIONAL REGULATION OF THE BPS POLYSACCHARIDE IN BORDETELLA PERTUSSIS PATHOGENESIS AND BIOFILM DEVELOPMENT BY MATTHEW S. CONOVER A Dissertation Submitted to
AN EXAMINATION OF THE FUNCTION AND THE TRANSCRIPTIONAL REGULATION OF THE BPS POLYSACCHARIDE IN BORDETELLA PERTUSSIS PATHOGENESIS AND BIOFILM DEVELOPMENT BY MATTHEW S. CONOVER A Dissertation Submitted to
Staphylococcus aureus
 Staphylococcus aureus Significant human pathogen. SSTI Biomaterial related infections Osteomyelitis Endocarditis Toxin mediated diseases TSST Staphylococcal enterotoxins Quintessential Pathogen? Nizet
Staphylococcus aureus Significant human pathogen. SSTI Biomaterial related infections Osteomyelitis Endocarditis Toxin mediated diseases TSST Staphylococcal enterotoxins Quintessential Pathogen? Nizet
Significant human pathogen. SSTI Biomaterial related infections Osteomyelitis Endocarditis Toxin mediated diseases TSST Staphylococcal enterotoxins
 Staphylococcus aureus Significant human pathogen. SSTI Biomaterial related infections Osteomyelitis Endocarditis Toxin mediated diseases TSST Staphylococcal enterotoxins Quintessential Pathogen? Nizet
Staphylococcus aureus Significant human pathogen. SSTI Biomaterial related infections Osteomyelitis Endocarditis Toxin mediated diseases TSST Staphylococcal enterotoxins Quintessential Pathogen? Nizet
Index. Note: Page numbers of article titles are in boldface type.
 Index Note: Page numbers of article titles are in boldface type. A Abdominal viscera, examination of, in investigation of emerging infectious diseases of food animals, 6 American Veterinary Medical Association,
Index Note: Page numbers of article titles are in boldface type. A Abdominal viscera, examination of, in investigation of emerging infectious diseases of food animals, 6 American Veterinary Medical Association,
Lecture 6: Fungi, antibiotics and bacterial infections. Outline Eukaryotes and Prokaryotes Viruses Bacteria Antibiotics Antibiotic resistance
 Lecture 6: Fungi, antibiotics and bacterial infections Outline Eukaryotes and Prokaryotes Viruses Bacteria Antibiotics Antibiotic resistance Lecture 1 2 3 Lecture Outline Section 4 Willow and aspirin Opium
Lecture 6: Fungi, antibiotics and bacterial infections Outline Eukaryotes and Prokaryotes Viruses Bacteria Antibiotics Antibiotic resistance Lecture 1 2 3 Lecture Outline Section 4 Willow and aspirin Opium
Vaccines for Cats. 2. Feline viral rhinotracheitis, FVR caused by FVR virus, also known as herpes virus type 1, FHV-1
 Vaccines for Cats Recent advances in veterinary medical science have resulted in an increase in the number and type of vaccines that are available for use in cats, and improvements are continuously being
Vaccines for Cats Recent advances in veterinary medical science have resulted in an increase in the number and type of vaccines that are available for use in cats, and improvements are continuously being
Why Don t These Drugs Work Anymore? Biosciences in the 21 st Century Dr. Amber Rice October 28, 2013
 Why Don t These Drugs Work Anymore? Biosciences in the 21 st Century Dr. Amber Rice October 28, 2013 Outline Drug resistance: a case study Evolution: the basics How does resistance evolve? Examples of
Why Don t These Drugs Work Anymore? Biosciences in the 21 st Century Dr. Amber Rice October 28, 2013 Outline Drug resistance: a case study Evolution: the basics How does resistance evolve? Examples of
O Antigen Protects Bordetella parapertussis from Complement
 INFECTION AND IMMUNITY, Apr. 2008, p. 1774 1780 Vol. 76, No. 4 0019-9567/08/$08.00 0 doi:10.1128/iai.01629-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. O Antigen Protects
INFECTION AND IMMUNITY, Apr. 2008, p. 1774 1780 Vol. 76, No. 4 0019-9567/08/$08.00 0 doi:10.1128/iai.01629-07 Copyright 2008, American Society for Microbiology. All Rights Reserved. O Antigen Protects
An#bio#cs and challenges in the wake of superbugs
 An#bio#cs and challenges in the wake of superbugs www.biochemj.org/bj/330/0581/bj3300581.htm ciss.blog.olemiss.edu Dr. Vassie Ware Bioscience in the 21 st Century November 14, 2014 Who said this and what
An#bio#cs and challenges in the wake of superbugs www.biochemj.org/bj/330/0581/bj3300581.htm ciss.blog.olemiss.edu Dr. Vassie Ware Bioscience in the 21 st Century November 14, 2014 Who said this and what
Mechanisms and Pathways of AMR in the environment
 FMM/RAS/298: Strengthening capacities, policies and national action plans on prudent and responsible use of antimicrobials in fisheries Final Workshop in cooperation with AVA Singapore and INFOFISH 12-14
FMM/RAS/298: Strengthening capacities, policies and national action plans on prudent and responsible use of antimicrobials in fisheries Final Workshop in cooperation with AVA Singapore and INFOFISH 12-14
Different mechanisms of vaccine-induced and infection-induced immunity to Bordetella bronchiseptica
 Microbes and Infection 9 (2007) 442e448 Original article Different mechanisms of vaccine-induced and infection-induced immunity to Bordetella bronchiseptica Lakshmi Gopinathan b, Girish S. Kirimanjeswara
Microbes and Infection 9 (2007) 442e448 Original article Different mechanisms of vaccine-induced and infection-induced immunity to Bordetella bronchiseptica Lakshmi Gopinathan b, Girish S. Kirimanjeswara
copyright Joette Calabrese, Inc.
 1 copyright Joette Calabrese, Inc. 2 copyright Joette Calabrese, Inc. 3 4 5 6 7 I want to read you a very striking quote from my friend Sarah Pope s blog: The Healthy Home Economist: The over reliance
1 copyright Joette Calabrese, Inc. 2 copyright Joette Calabrese, Inc. 3 4 5 6 7 I want to read you a very striking quote from my friend Sarah Pope s blog: The Healthy Home Economist: The over reliance
Antibiotics: Conflict and Communication in Microbial Communities
 Antibiotics: Conflict and Communication in Microbial Communities Antibiotics mediate species interactions in natural habitats, affecting the dynamics of microbial coevolution Daniel C. Schlatter and Linda
Antibiotics: Conflict and Communication in Microbial Communities Antibiotics mediate species interactions in natural habitats, affecting the dynamics of microbial coevolution Daniel C. Schlatter and Linda
Selective toxicity. Antimicrobial Drugs. Alexander Fleming 10/17/2016
 Selective toxicity Antimicrobial Drugs Chapter 20 BIO 220 Drugs must work inside the host and harm the infective pathogens, but not the host Antibiotics are compounds produced by fungi or bacteria that
Selective toxicity Antimicrobial Drugs Chapter 20 BIO 220 Drugs must work inside the host and harm the infective pathogens, but not the host Antibiotics are compounds produced by fungi or bacteria that
of Conferences of OIE Regional Commissions organised since 1 June 2013 endorsed by the Assembly of the OIE on 29 May 2014
 of Conferences of OIE Regional Commissions organised since 1 June 2013 endorsed by the Assembly of the OIE on 29 May 2014 2 12 th Conference of the OIE Regional Commission for the Middle East Amman (Jordan),
of Conferences of OIE Regional Commissions organised since 1 June 2013 endorsed by the Assembly of the OIE on 29 May 2014 2 12 th Conference of the OIE Regional Commission for the Middle East Amman (Jordan),
BvgAS Is Sufficient for Activation of the Bordetella pertussis ptx Locus in Escherichia coli
 JOURNAL OF BACTERIOLOGY, Nov. 1995, p. 6477 6485 Vol. 177, No. 22 0021-9193/95/$04.00 0 Copyright 1995, American Society for Microbiology BvgAS Is Sufficient for Activation of the Bordetella pertussis
JOURNAL OF BACTERIOLOGY, Nov. 1995, p. 6477 6485 Vol. 177, No. 22 0021-9193/95/$04.00 0 Copyright 1995, American Society for Microbiology BvgAS Is Sufficient for Activation of the Bordetella pertussis
Investigation of the molecular biology and contribution to virulence of Bordetella bronchiseptica urease
 University of Wollongong Research Online University of Wollongong Thesis Collection University of Wollongong Thesis Collections 1999 Investigation of the molecular biology and contribution to virulence
University of Wollongong Research Online University of Wollongong Thesis Collection University of Wollongong Thesis Collections 1999 Investigation of the molecular biology and contribution to virulence
Dier & Kruid Prof. Dr. J. Fink-Gremmels DVM, PhD, Dip ECVPT
 Dier & Kruid 03-06-2015 Prof. Dr. J. Fink-Gremmels DVM, PhD, Dip ECVPT J.Fink@uu.nl Antibiotics secondary metabolites produced under conditions of stress Fungi imperfecti (Penicillium Fusarium) Streptomyces
Dier & Kruid 03-06-2015 Prof. Dr. J. Fink-Gremmels DVM, PhD, Dip ECVPT J.Fink@uu.nl Antibiotics secondary metabolites produced under conditions of stress Fungi imperfecti (Penicillium Fusarium) Streptomyces
The O Antigen Is a Critical Antigen for the Development of a Protective Immune Response to Bordetella parapertussis
 INFECTION AND IMMUNITY, Nov. 2009, p. 5050 5058 Vol. 77, No. 11 0019-9567/09/$12.00 doi:10.1128/iai.00667-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. The O Antigen Is a Critical
INFECTION AND IMMUNITY, Nov. 2009, p. 5050 5058 Vol. 77, No. 11 0019-9567/09/$12.00 doi:10.1128/iai.00667-09 Copyright 2009, American Society for Microbiology. All Rights Reserved. The O Antigen Is a Critical
Overview. There are commonly found arrangements of bacteria based on their division. Spheres, Rods, Spirals
 Bacteria Overview Bacteria live almost everywhere. Most are microscopic ranging from 0.5 5 m in size, and unicellular. They have a variety of shapes when viewed under a microscope, most commonly: Spheres,
Bacteria Overview Bacteria live almost everywhere. Most are microscopic ranging from 0.5 5 m in size, and unicellular. They have a variety of shapes when viewed under a microscope, most commonly: Spheres,
National Research Center
 National Research Center Update of immunodiagnosis of cystic echinococcosis cysts Global distribution of zoonotic strains of Echinococcus granulosus (Adapted from Eckert and Deplazes, 2004) Echinococcus
National Research Center Update of immunodiagnosis of cystic echinococcosis cysts Global distribution of zoonotic strains of Echinococcus granulosus (Adapted from Eckert and Deplazes, 2004) Echinococcus
Antibiotics & Resistance
 What are antibiotics? Antibiotics & esistance Antibiotics are molecules that stop bacteria from growing or kill them Antibiotics, agents against life - either natural or synthetic chemicals - designed
What are antibiotics? Antibiotics & esistance Antibiotics are molecules that stop bacteria from growing or kill them Antibiotics, agents against life - either natural or synthetic chemicals - designed
Attorneys for Plaintiffs Hells Canyon Preservation Council and The Wilderness Society UNITED STATES DISTRICT COURT FOR THE DISTRICT OF IDAHO
 Lauren M. Rule (ISB # 6863 ADVOCATES FOR THE WEST PO Box 1612 Boise ID 83701 (208 342-7024 lrule@advocateswest.org Attorney for Plaintiff Western Watersheds Project Jennifer R. Schemm (OSB #97008 602 O
Lauren M. Rule (ISB # 6863 ADVOCATES FOR THE WEST PO Box 1612 Boise ID 83701 (208 342-7024 lrule@advocateswest.org Attorney for Plaintiff Western Watersheds Project Jennifer R. Schemm (OSB #97008 602 O
Restriction Endonuclease Analysis Discriminates Bordetella bronchiseptica Isolates
 JOURNAL OF CLINICAL MICROBIOLOGY, Dec. 2000, p. 4387 4393 Vol. 38, No. 12 0095-1137/00/$04.00 0 Restriction Endonuclease Analysis Discriminates Bordetella bronchiseptica Isolates RANDY E. SACCO,* KAREN
JOURNAL OF CLINICAL MICROBIOLOGY, Dec. 2000, p. 4387 4393 Vol. 38, No. 12 0095-1137/00/$04.00 0 Restriction Endonuclease Analysis Discriminates Bordetella bronchiseptica Isolates RANDY E. SACCO,* KAREN
ESBL Producers An Increasing Problem: An Overview Of An Underrated Threat
 ESBL Producers An Increasing Problem: An Overview Of An Underrated Threat Hicham Ezzat Professor of Microbiology and Immunology Cairo University Introduction 1 Since the 1980s there have been dramatic
ESBL Producers An Increasing Problem: An Overview Of An Underrated Threat Hicham Ezzat Professor of Microbiology and Immunology Cairo University Introduction 1 Since the 1980s there have been dramatic
Medical Bacteriology- Lecture 14. Gram negative coccobacilli. Zoonosis. Brucella. Yersinia. Francesiella
 Medical Bacteriology- Lecture 14 Gram negative coccobacilli Zoonosis Brucella Yersinia Francesiella 1 Zoonosis: A disease, primarily of animals, which is transmitted to humans as a result of direct or
Medical Bacteriology- Lecture 14 Gram negative coccobacilli Zoonosis Brucella Yersinia Francesiella 1 Zoonosis: A disease, primarily of animals, which is transmitted to humans as a result of direct or
Transition cows have decreased immune function. The transition period. Inflammation, Immune Function, and the Transition Cow.
 Overview Inflammation, Immune Function, and the Transition Cow Barry Bradford Kansas State University Herd Health & Nutrition Conferences April 2016 Immunity and inflammation in the transition cow Long
Overview Inflammation, Immune Function, and the Transition Cow Barry Bradford Kansas State University Herd Health & Nutrition Conferences April 2016 Immunity and inflammation in the transition cow Long
Electron Microscopic Observations on Ciliated Epithelium of Tracheal Organ Cultures Infected with Bordetella bronchiseptica
 Microbiol. Immunol. Vol. 33 (2), 111-121, 1989 Electron Microscopic Observations on Ciliated Epithelium of Tracheal Organ Cultures Infected with Bordetella bronchiseptica Kachiko SEKIYA,*,1 Yutaka FUTAESAKU,2
Microbiol. Immunol. Vol. 33 (2), 111-121, 1989 Electron Microscopic Observations on Ciliated Epithelium of Tracheal Organ Cultures Infected with Bordetella bronchiseptica Kachiko SEKIYA,*,1 Yutaka FUTAESAKU,2
Antibiotic Resistance. Antibiotic Resistance: A Growing Concern. Antibiotic resistance is not new 3/21/2011
 Antibiotic Resistance Antibiotic Resistance: A Growing Concern Judy Ptak RN MSN Infection Prevention Practitioner Dartmouth-Hitchcock Medical Center Lebanon, NH Occurs when a microorganism fails to respond
Antibiotic Resistance Antibiotic Resistance: A Growing Concern Judy Ptak RN MSN Infection Prevention Practitioner Dartmouth-Hitchcock Medical Center Lebanon, NH Occurs when a microorganism fails to respond
INTERACTIONS BETWEEN ENDEMIC BORDETELLA SPECIES AND HOST IMMUNITY
 The Pennsylvania State University The Graduate School College of Agricultural Sciences INTERACTIONS BETWEEN ENDEMIC BORDETELLA SPECIES AND HOST IMMUNITY A Thesis in Pathobiology by Daniel Nathan Wolfe
The Pennsylvania State University The Graduate School College of Agricultural Sciences INTERACTIONS BETWEEN ENDEMIC BORDETELLA SPECIES AND HOST IMMUNITY A Thesis in Pathobiology by Daniel Nathan Wolfe
Eric T. Harvill, Dept. of Veterinary and Biomedical Sciences, Penn State. Vivek Kapur, Dept. of Veterinary and Biomedical Sciences, Penn State
 Genomic Analysis of the Classical Bordetella Eric T. Harvill, Dept. of Veterinary and Biomedical Sciences, Penn State Vivek Kapur, Dept. of Veterinary and Biomedical Sciences, Penn State Ying Zhang, Dept.
Genomic Analysis of the Classical Bordetella Eric T. Harvill, Dept. of Veterinary and Biomedical Sciences, Penn State Vivek Kapur, Dept. of Veterinary and Biomedical Sciences, Penn State Ying Zhang, Dept.
Running title: Contribution of Bordetella Bps to Biofilm Formation and Respiratory Disease in
 IAI Accepted Manuscript Posted Online 30 May 2017 Infect. Immun. doi:10.1128/iai.00261-17 Copyright 2017 American Society for Microbiology. All Rights Reserved. 1 2 The Bordetella Bps Polysaccharide is
IAI Accepted Manuscript Posted Online 30 May 2017 Infect. Immun. doi:10.1128/iai.00261-17 Copyright 2017 American Society for Microbiology. All Rights Reserved. 1 2 The Bordetella Bps Polysaccharide is
THE HUMAN MICROBIOME: THE INFECTION PREVENTIONIST S BEST FRIEND
 THE HUMAN MICROBIOME: THE INFECTION PREVENTIONIST S BEST FRIEND Michigan Communicable Disease Conference May 4, 2017 Richard A. Van Enk, Ph.D., CIC Director, Infection Prevention and Epidemiology vanenkr@bronsonhg.org
THE HUMAN MICROBIOME: THE INFECTION PREVENTIONIST S BEST FRIEND Michigan Communicable Disease Conference May 4, 2017 Richard A. Van Enk, Ph.D., CIC Director, Infection Prevention and Epidemiology vanenkr@bronsonhg.org
Xochitl Morgan: The human microbiome; the role of commensals in health and disease.
 MICR332: Health Microbiology 18 points; Semester 2 Course prescription: Disease mechanisms of key microbial pathogens, including bacteria, protozoa, and fungi. Treatment and control of microbial diseases.
MICR332: Health Microbiology 18 points; Semester 2 Course prescription: Disease mechanisms of key microbial pathogens, including bacteria, protozoa, and fungi. Treatment and control of microbial diseases.
Burton's Microbiology for the Health Sciences. Chapter 9. Controlling Microbial Growth in Vivo Using Antimicrobial Agents
 Burton's Microbiology for the Health Sciences Chapter 9. Controlling Microbial Growth in Vivo Using Antimicrobial Agents Chapter 9 Outline Introduction Characteristics of an Ideal Antimicrobial Agent How
Burton's Microbiology for the Health Sciences Chapter 9. Controlling Microbial Growth in Vivo Using Antimicrobial Agents Chapter 9 Outline Introduction Characteristics of an Ideal Antimicrobial Agent How
READER S DIGEST OVERVIEW: BIGHORN SHEEP. Peregrine Wolff, DVM
 READER S DIGEST OVERVIEW: RESPIRATORY DISEASE IN BIGHORN SHEEP Peregrine Wolff, DVM Nevada Department of Wildlife During the Lewis & Clark expedition (1804 1806) There may have been 2 million bighorn sheep
READER S DIGEST OVERVIEW: RESPIRATORY DISEASE IN BIGHORN SHEEP Peregrine Wolff, DVM Nevada Department of Wildlife During the Lewis & Clark expedition (1804 1806) There may have been 2 million bighorn sheep
Redefining Infection Management. Proven Clinical Outcomes
 Proven Clinical Outcomes Proof of Bacteria-Binding1 In the first 30 seconds, 1 square centimeter of Cutimed Sorbact binds wound bacteria - after 2 hours, the amount of bacteria bound are more than would
Proven Clinical Outcomes Proof of Bacteria-Binding1 In the first 30 seconds, 1 square centimeter of Cutimed Sorbact binds wound bacteria - after 2 hours, the amount of bacteria bound are more than would
Federal Expert Select Agent Panel (FESAP) Deliberations
 Deliberations") Federal Expert Select Agent Panel (FESAP) Deliberations FESAP and Biennial Review Established in 2010 and tasked with policy issues relevant to the security of biological select agents and toxins Per recommendations
Federal Expert Select Agent Panel (FESAP) Deliberations FESAP and Biennial Review Established in 2010 and tasked with policy issues relevant to the security of biological select agents and toxins Per recommendations
Typhoid fever - priorities for research and development of new treatments
 Typhoid fever - priorities for research and development of new treatments Isabela Ribeiro, Manica Balasegaram, Christopher Parry October 2017 Enteric infections Enteric infections vary in symptoms and
Typhoid fever - priorities for research and development of new treatments Isabela Ribeiro, Manica Balasegaram, Christopher Parry October 2017 Enteric infections Enteric infections vary in symptoms and
BIOL 2900 D 4.00 Microbiology in Health/Disease
 SYLLABUS BIOL 2900 - D Spring, 2017 Course: Microbiology in Health and Disease Instructor: Prafull C. Shah Office Hours: Before or after classes, or by appointment by Email to pcshah@valdosta.edu. Semester
SYLLABUS BIOL 2900 - D Spring, 2017 Course: Microbiology in Health and Disease Instructor: Prafull C. Shah Office Hours: Before or after classes, or by appointment by Email to pcshah@valdosta.edu. Semester
Block Objectives: Basic Infectious Diseases Block
 Course: Intro to Infectious Diseases IID-BASID-01 Virtual Lab: Infectious Disease Laboratory Session 1 Identify etiologic bacterial organisms from clinical case studies based on the following: Discriminating
Course: Intro to Infectious Diseases IID-BASID-01 Virtual Lab: Infectious Disease Laboratory Session 1 Identify etiologic bacterial organisms from clinical case studies based on the following: Discriminating
Toll-Like Receptor 4 Limits Transmission of Bordetella bronchiseptica
 Toll-Like Receptor 4 Limits Transmission of Bordetella bronchiseptica Olivier Rolin 1,2, Will Smallridge 1,2, Michael Henry 1, Laura Goodfield 1,2, David Place 1,2, Eric T. Harvill 1 * 1 Department of
Toll-Like Receptor 4 Limits Transmission of Bordetella bronchiseptica Olivier Rolin 1,2, Will Smallridge 1,2, Michael Henry 1, Laura Goodfield 1,2, David Place 1,2, Eric T. Harvill 1 * 1 Department of
Course: Microbiology in Health and Disease
 SYLLABUS BIOL 2900 SECTION D SPRING 2012 Course: Microbiology in Health and Disease BIPIN PATEL Office Hours: Before or after Class or by appointment Semester Begins JANUARY 09 TO MAY 04 2012 2900 D 4.00
SYLLABUS BIOL 2900 SECTION D SPRING 2012 Course: Microbiology in Health and Disease BIPIN PATEL Office Hours: Before or after Class or by appointment Semester Begins JANUARY 09 TO MAY 04 2012 2900 D 4.00
Course: Microbiology in Health and Disease Office Hours: Before or after Class or by appointment
 SYLLABUS BIOL 2900 SECTIONS C AND D Spring, 2011 Course: Microbiology in Health and Disease Office Hours: Before or after Class or by appointment Semester Begins on January 10, 2011 and ends on May 2,
SYLLABUS BIOL 2900 SECTIONS C AND D Spring, 2011 Course: Microbiology in Health and Disease Office Hours: Before or after Class or by appointment Semester Begins on January 10, 2011 and ends on May 2,
Virulence of Bordetella bronchiseptica: Role of Adenylate Cyclase-Hemolysin
 INFEcrION AND IMMUNITY, OCt. 1993, p. 472-478 Vol. 61, No. 1 19-9567/93/1472-7$2./ Copyright 1993, American Society for Microbiology Virulence of Bordetella bronchiseptica: Role of Adenylate Cyclase-Hemolysin
INFEcrION AND IMMUNITY, OCt. 1993, p. 472-478 Vol. 61, No. 1 19-9567/93/1472-7$2./ Copyright 1993, American Society for Microbiology Virulence of Bordetella bronchiseptica: Role of Adenylate Cyclase-Hemolysin
Randall Singer, DVM, MPVM, PhD
 ANTIBIOTIC RESISTANCE Randall Singer, DVM, MPVM, PhD Associate Professor of Epidemiology Department of Veterinary and Biomedical Sciences University of Minnesota Overview How does resistance develop? What
ANTIBIOTIC RESISTANCE Randall Singer, DVM, MPVM, PhD Associate Professor of Epidemiology Department of Veterinary and Biomedical Sciences University of Minnesota Overview How does resistance develop? What
Antimicrobial use in poultry: Emerging public health problem
 Antimicrobial use in poultry: Emerging public health problem Eric S. Mitema, BVM, MS, PhD CPD- Diagnosis and Treatment of Poultry Diseases FVM, CAVS, 6 th. August, 2014 AMR cont Antibiotics - Natural or
Antimicrobial use in poultry: Emerging public health problem Eric S. Mitema, BVM, MS, PhD CPD- Diagnosis and Treatment of Poultry Diseases FVM, CAVS, 6 th. August, 2014 AMR cont Antibiotics - Natural or
Filamentous Hemagglutinin of Bordetella bronchiseptica Is Required for Efficient Establishment of Tracheal Colonization
 INFECTION AND IMMUNITY, Dec. 1998, p. 5921 5929 Vol. 66, No. 12 0019-9567/98/$04.00 0 Copyright 1998, American Society for Microbiology. All Rights Reserved. Filamentous Hemagglutinin of Bordetella bronchiseptica
INFECTION AND IMMUNITY, Dec. 1998, p. 5921 5929 Vol. 66, No. 12 0019-9567/98/$04.00 0 Copyright 1998, American Society for Microbiology. All Rights Reserved. Filamentous Hemagglutinin of Bordetella bronchiseptica
Bordetella pertussis Infection or Vaccination Substantially Protects Mice against B. bronchiseptica Infection
 Bordetella pertussis Infection or Vaccination Substantially Protects Mice against B. bronchiseptica Infection Elizabeth M. Goebel 1,2, Xuqing Zhang 1,3, Eric T. Harvill 1 * 1 Department of Veterinary and
Bordetella pertussis Infection or Vaccination Substantially Protects Mice against B. bronchiseptica Infection Elizabeth M. Goebel 1,2, Xuqing Zhang 1,3, Eric T. Harvill 1 * 1 Department of Veterinary and
Inhibiting Microbial Growth in vivo. CLS 212: Medical Microbiology Zeina Alkudmani
 Inhibiting Microbial Growth in vivo CLS 212: Medical Microbiology Zeina Alkudmani Chemotherapy Definitions The use of any chemical (drug) to treat any disease or condition. Chemotherapeutic Agent Any drug
Inhibiting Microbial Growth in vivo CLS 212: Medical Microbiology Zeina Alkudmani Chemotherapy Definitions The use of any chemical (drug) to treat any disease or condition. Chemotherapeutic Agent Any drug
Understanding and prevention of transmission of antibiotic resistance between bacterial populations and One Health reservoirs
 Priority Topic D - Transmission Understanding and prevention of transmission of antibiotic resistance between bacterial populations and One Health reservoirs The overarching goal of this priority topic
Priority Topic D - Transmission Understanding and prevention of transmission of antibiotic resistance between bacterial populations and One Health reservoirs The overarching goal of this priority topic
Boosting Bacterial Metabolism to Combat Antibiotic Resistance
 Boosting Bacterial Metabolism to Combat Antibiotic Resistance The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published
Boosting Bacterial Metabolism to Combat Antibiotic Resistance The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published
Eur. J. Immunol : Antibody-mediated bacterial 1
 Seite 2 Eur. J. Immunol. 2004. 34: Antibody-mediated bacterial 1 Antibody-mediated bacterial clearance from the lower respiratory tract of mice requires complement component C3 Elizabeth J. Pishko, Girish
Seite 2 Eur. J. Immunol. 2004. 34: Antibody-mediated bacterial 1 Antibody-mediated bacterial clearance from the lower respiratory tract of mice requires complement component C3 Elizabeth J. Pishko, Girish
22/10/2013. antibiotic resistance an ecological problem. this part of the session. an ecological perspective. selection pressure and evolution
 antibiotic resistance an ecological problem Jon Iredell critical.infection@sydney.edu.au NHMRC Centre for Research Excellence in Critical Infection Marie Bashir Institute Westmead Millennium Institute
antibiotic resistance an ecological problem Jon Iredell critical.infection@sydney.edu.au NHMRC Centre for Research Excellence in Critical Infection Marie Bashir Institute Westmead Millennium Institute
Molecular Analysis of β-lactamase Genes in Antibiotic Resistant Bacteria
 Bowling Green State University ScholarWorks@BGSU Honors Projects Honors College Spring 5-1-2017 Molecular Analysis of β-lactamase Genes in Antibiotic Resistant Bacteria Neisha Medina Candelaria neisham@bgsu.edu
Bowling Green State University ScholarWorks@BGSU Honors Projects Honors College Spring 5-1-2017 Molecular Analysis of β-lactamase Genes in Antibiotic Resistant Bacteria Neisha Medina Candelaria neisham@bgsu.edu
Antimicrobial Therapy
 Chapter 12 The Elements of Chemotherapy Topics - Antimicrobial Therapy - Selective Toxicity - Survey of Antimicrobial Drug - Microbial Drug Resistance - Drug and Host Interaction Antimicrobial Therapy
Chapter 12 The Elements of Chemotherapy Topics - Antimicrobial Therapy - Selective Toxicity - Survey of Antimicrobial Drug - Microbial Drug Resistance - Drug and Host Interaction Antimicrobial Therapy
In Vitro and In Vivo Characterization of a Bordetella bronchiseptica Mutant Strain with a Deep Rough Lipopolysaccharide Structure
 INFECTION AND IMMUNITY, Apr. 2002, p. 1791 1798 Vol. 70, No. 4 0019-9567/02/$04.00 0 DOI: 10.1128/IAI.70.4.1791 1798.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. In Vitro
INFECTION AND IMMUNITY, Apr. 2002, p. 1791 1798 Vol. 70, No. 4 0019-9567/02/$04.00 0 DOI: 10.1128/IAI.70.4.1791 1798.2002 Copyright 2002, American Society for Microbiology. All Rights Reserved. In Vitro
Evolution of the Bordetella autotransporter Pertactin: identifications of regions subject to positive selection
 Evolution of the Bordetella autotransporter Pertactin: identifications of regions subject to positive selection Marcel Hijnen 1,2, Dimitri Diavatopoulos 1,2 and Frits R. Mooi 1,2 Both authors contributed
Evolution of the Bordetella autotransporter Pertactin: identifications of regions subject to positive selection Marcel Hijnen 1,2, Dimitri Diavatopoulos 1,2 and Frits R. Mooi 1,2 Both authors contributed
PREPARED FOR: U.S. Army Medical Research and Materiel Command Fort Detrick, Maryland X Approved for public release; distribution unlimited
 Award Number: W8XWH--- TITLE: Defining the Role of Autophagy Kinase ULK Signaling in Therapeutic Response of Tuberous Sclerosis Complex to Inhibitors PRINCIPAL INVESTIGATOR: Reuben J. Shaw, Ph.D. CONTRACTING
Award Number: W8XWH--- TITLE: Defining the Role of Autophagy Kinase ULK Signaling in Therapeutic Response of Tuberous Sclerosis Complex to Inhibitors PRINCIPAL INVESTIGATOR: Reuben J. Shaw, Ph.D. CONTRACTING
Mechanism of antibiotic resistance
 Mechanism of antibiotic resistance Dr.Siriwoot Sookkhee Ph.D (Biopharmaceutics) Department of Microbiology Faculty of Medicine, Chiang Mai University Antibiotic resistance Cross-resistance : resistance
Mechanism of antibiotic resistance Dr.Siriwoot Sookkhee Ph.D (Biopharmaceutics) Department of Microbiology Faculty of Medicine, Chiang Mai University Antibiotic resistance Cross-resistance : resistance
Florida Health Care Association District 2 January 13, 2015 A.C. Burke, MA, CIC
 Florida Health Care Association District 2 January 13, 2015 A.C. Burke, MA, CIC 11/20/2014 1 To describe carbapenem-resistant Enterobacteriaceae. To identify laboratory detection standards for carbapenem-resistant
Florida Health Care Association District 2 January 13, 2015 A.C. Burke, MA, CIC 11/20/2014 1 To describe carbapenem-resistant Enterobacteriaceae. To identify laboratory detection standards for carbapenem-resistant
Dr. P. P. Doke. M.D., D.N.B., Ph.D., FIPHA. Professor, Department of Community Medicine, Bharati Vidyapeeth Medical College, Pune
 Dr. P. P. Doke M.D., D.N.B., Ph.D., FIPHA Professor, Department of Community Medicine, Bharati Vidyapeeth Medical College, Pune 1 Anti microbial resistance is now a global geometrically increasing threat
Dr. P. P. Doke M.D., D.N.B., Ph.D., FIPHA Professor, Department of Community Medicine, Bharati Vidyapeeth Medical College, Pune 1 Anti microbial resistance is now a global geometrically increasing threat
EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS WORK-PROGRAMME PROPOSAL Version 2 VISAVET. Universidad Complutense de Madrid
 FOR BOVINE TUBERCULOSIS WORK-PROGRAMME PROPOSAL Version 2 VISAVET. Universidad Complutense de Madrid") EUROPEAN COMMISSION HEALTH & CONSUMERS DIRECTORATE-GENERAL Directorate D Animal Health and Welfare Unit D1- Animal health and Standing Committees EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS
EUROPEAN COMMISSION HEALTH & CONSUMERS DIRECTORATE-GENERAL Directorate D Animal Health and Welfare Unit D1- Animal health and Standing Committees EUROPEAN REFERENCE LABORATORY (EU-RL) FOR BOVINE TUBERCULOSIS
Cell Wall Inhibitors. Assistant Professor Naza M. Ali. Lec 3 7 Nov 2017
 Cell Wall Inhibitors Assistant Professor Naza M. Ali Lec 3 7 Nov 2017 Cell wall The cell wall is a rigid outer layer, it completely surrounds the cytoplasmic membrane, maintaining the shape of the cell
Cell Wall Inhibitors Assistant Professor Naza M. Ali Lec 3 7 Nov 2017 Cell wall The cell wall is a rigid outer layer, it completely surrounds the cytoplasmic membrane, maintaining the shape of the cell
Antimicrobial Resistance Initiative
 Antimicrobial Resistance Initiative Antimicrobial Resistance Initiative Resistance to antimicrobial agents has become a threat to public health all over the world. Microorganisms become resistant to antimicrobial
Antimicrobial Resistance Initiative Antimicrobial Resistance Initiative Resistance to antimicrobial agents has become a threat to public health all over the world. Microorganisms become resistant to antimicrobial
Correlation of. Animal Science Biology & Technology, 3/E, by Dr. Robert Mikesell/ MeeCee Baker, 2011, ISBN 10: ; ISBN 13:
 Correlation of Animal Science Biology & Technology, 3/E, by Dr. Robert Mikesell/ MeeCee Baker, 2011, ISBN 10: 1435486374; ISBN 13: 9781435486379 to Indiana s Agricultural Education Curriculum Standards
Correlation of Animal Science Biology & Technology, 3/E, by Dr. Robert Mikesell/ MeeCee Baker, 2011, ISBN 10: 1435486374; ISBN 13: 9781435486379 to Indiana s Agricultural Education Curriculum Standards
Antibiotic Resistance: Implications on Genetic Improvement and Disease Resistance. D. Hurnik, University of Prince Edward Island
 Antibiotic Resistance: Implications on Genetic Improvement and Disease Resistance. What are Antibiotics? D. Hurnik, University of Prince Edward Island Antibiotics are compounds that kill bacteria and are
Antibiotic Resistance: Implications on Genetic Improvement and Disease Resistance. What are Antibiotics? D. Hurnik, University of Prince Edward Island Antibiotics are compounds that kill bacteria and are
CHAPTER 18 THE COCCI OF MEDICAL IMPORTANCE. Learning Objectives
 CHAPTER 18 THE COCCI OF MEDICAL IMPORTANCE Gram-positive and gram-negative cocci that cause infection are presented. The difference between commensal and pathogenic strains is explained, because many of
CHAPTER 18 THE COCCI OF MEDICAL IMPORTANCE Gram-positive and gram-negative cocci that cause infection are presented. The difference between commensal and pathogenic strains is explained, because many of
Antibiotic Resistance in Bacteria
 Antibiotic Resistance in Bacteria Electron Micrograph of E. Coli Diseases Caused by Bacteria 1928 1 2 Fleming 3 discovers penicillin the first antibiotic. Some Clinically Important Antibiotics Antibiotic
Antibiotic Resistance in Bacteria Electron Micrograph of E. Coli Diseases Caused by Bacteria 1928 1 2 Fleming 3 discovers penicillin the first antibiotic. Some Clinically Important Antibiotics Antibiotic
VETERINARY BIOMEDICAL SCIENCES (VBSC)
") Veterinary Biomedical Sciences (VBSC) 1 VETERINARY BIOMEDICAL SCIENCES (VBSC) VBSC 5000 Master s Research and Thesis Prerequisites: Graduate standing. Description: Research problem for meeting requirements
Veterinary Biomedical Sciences (VBSC) 1 VETERINARY BIOMEDICAL SCIENCES (VBSC) VBSC 5000 Master s Research and Thesis Prerequisites: Graduate standing. Description: Research problem for meeting requirements
Recommended for Implementation at Step 7 of the VICH Process on 15 December 2004 by the VICH Steering Committee
 VICH GL27 (ANTIMICROBIAL RESISTANCE: PRE-APPROVAL) December 2003 For implementation at Step 7 - Final GUIDANCE ON PRE-APPROVAL INFORMATION FOR REGISTRATION OF NEW VETERINARY MEDICINAL PRODUCTS FOR FOOD
VICH GL27 (ANTIMICROBIAL RESISTANCE: PRE-APPROVAL) December 2003 For implementation at Step 7 - Final GUIDANCE ON PRE-APPROVAL INFORMATION FOR REGISTRATION OF NEW VETERINARY MEDICINAL PRODUCTS FOR FOOD
Our vision. To be a game-changer in the development of sustainable, prophylactic and therapeutic veterinary products.
 product information Our vision To be a game-changer in the development of sustainable, prophylactic and therapeutic veterinary products. www.farmpharma.se Company Registration Number: 559157-4131 15 A
product information Our vision To be a game-changer in the development of sustainable, prophylactic and therapeutic veterinary products. www.farmpharma.se Company Registration Number: 559157-4131 15 A
Growth Phase- and Nutrient Limitation-Associated Transcript Abundance Regulation in Bordetella pertussis
 INFECTION AND IMMUNITY, Oct. 2006, p. 5537 5548 Vol. 74, No. 10 0019-9567/06/$08.00 0 doi:10.1128/iai.00781-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Growth Phase- and
INFECTION AND IMMUNITY, Oct. 2006, p. 5537 5548 Vol. 74, No. 10 0019-9567/06/$08.00 0 doi:10.1128/iai.00781-06 Copyright 2006, American Society for Microbiology. All Rights Reserved. Growth Phase- and
Name(s): Period: Date:
: Period: Date:") Evolution in Action: Antibiotic Resistance HASPI Medical Biology Lab 21 Background/Introduction Evolution and Natural Selection Evolution is one of the driving factors in biology. It is simply the concept
Evolution in Action: Antibiotic Resistance HASPI Medical Biology Lab 21 Background/Introduction Evolution and Natural Selection Evolution is one of the driving factors in biology. It is simply the concept
Cercetări bacteriologice, epidemiologice şi serologice în bruceloza ovină ABSTRACT
 ABSTRACT Thesis entitled BACTERIOLOGICAL, EPIDEMIOLOGICAL AND SEROLOGICAL RESEARCHES IN BRUCELLOSIS OVINE is scientific and practical reasons the following: - Infectious epididymitis in Romania, described
ABSTRACT Thesis entitled BACTERIOLOGICAL, EPIDEMIOLOGICAL AND SEROLOGICAL RESEARCHES IN BRUCELLOSIS OVINE is scientific and practical reasons the following: - Infectious epididymitis in Romania, described
Tackling the need for new antibacterial drugs
 Tackling the need for new antibacterial drugs Wendy Lawson Lead Pharmacist, Infectious Diseases Imperial College Healthcare NHS Trust, London & Antibiotic Action Champion Timeline of Antibiotic Discovery
Tackling the need for new antibacterial drugs Wendy Lawson Lead Pharmacist, Infectious Diseases Imperial College Healthcare NHS Trust, London & Antibiotic Action Champion Timeline of Antibiotic Discovery
E-BOOK # BACTERIAL DISEASES IN HUMANS EBOOK
 15 November, 2017 E-BOOK # BACTERIAL DISEASES IN HUMANS EBOOK Document Filetype: PDF 475.49 KB 0 E-BOOK # BACTERIAL DISEASES IN HUMANS EBOOK Communicable diseases, also known as infectious diseases or
15 November, 2017 E-BOOK # BACTERIAL DISEASES IN HUMANS EBOOK Document Filetype: PDF 475.49 KB 0 E-BOOK # BACTERIAL DISEASES IN HUMANS EBOOK Communicable diseases, also known as infectious diseases or