CONTENTS. Effects of Various Litter Treatments on Growing Turkeys, by K. E. Nestor and P. A. Renner... 1
|
|
|
- Brendan Sutton
- 5 years ago
- Views:
Transcription

1
2 CONTENTS Effects of Various Litter Treatments on Growing Turkeys, by K. E. Nestor and P. A. Renner A Study of Crooked Toes in Turkeys, by K. E. Nestor The Influence of Egg Color on Hatchability of Turkey Eggs, by K. E* Nestor and P. A. Renner ? Production of Defective Eggs by Egg-and Meat-Type Turkey Hens Housed in Cages, by K. E. Nestor and W. L. Bacon The Influence of Ambient Temperature on Reproduction of Turkey Hens, by K. C. Nestor, S. P. Touchburn, and P. A. Renner A Possible Influence of Presence of Males on Egg Production of Turkey Hens, by K. E. Nestor and P. A. Renner The Relation Between Broodiness, Intensity of Lay, and Total Egg Production of Turkeys, by K, E. Nestor Reproductive Response of Japanese Quail to Intermittent Lighting Programs, by W. L. Bacon Intermittent Lighting Program for Turkey Hens, by K. I. Brown, W. L. Bacon, and K. E. Nestor Further Research on Manipulation of the Lighting Program for Control of Broodiness in Turkeys, by K. E. Nestor and W. L. Bacon The Effects of Accidental Pre-Lighting of Turkey Hens on Subsequent Egg Production, by K. E. Nestor Economic Significance of Mycoplasma meleagridis Infection in Turkeys, by Y. M. Saif and K. E, Nestor... 4 A New Variant of the Temperature Differential Procedure for Egg Dipping, by Y. M, Saif and K. E. Nestor Treatment of Turkey Hatching Eggs for the Control of Salmonella Infections, by Y, M. Saif Control of Mycoplasma meleagridis Infection in Turkey Hatching Eggs, by Y. M. Saif and K. E. Nestor Treatment of Turkey Semen to Eliminate Mycoplasma meleagridis, by Y. M. Saif and K. I. Brown The Inheritance and Relationship of Egg Quality and Reproduction Traits in Turkeys, by K. E. Nestor and C. R. Weaver
3 Exploring the Possible Value of Color Pattern Genes in Modern Turkeys, by K. E. Nestor * Low Intensity Light and Confinement Rearing of Turkey Males, by W. L. Bacon and S. P. Touchburn 58 Energy in Turkey Growing Diets, by S. P. Touchburn. 64 Growth Restriction of Large Type Turkey Breeders, by S. P. Touchburn and K. I. Brown 71 The Effects of Level and Form of Calcium in the Turkey Breeder Hen Diet on Egg Shell Quality and Reproductive Performance, by S. P. Touchburn and K. E. Nestor 76 A Study of Plasma Calcium Levels in Laying Turkey Hens, by Margery A. Musser, W. L. Bacon, and S. P. Touchburn 8 Concentration of a Yolk Precursor in the Plasma of Three Lines of Turkeys, by W. L. Bacon, K. E. Nestor, and Margery A. Musser 83 Development of Ovarian Follicles in Egg- and Meat-Type Turkeys, by W. L. Bacon and K. E. Nestor 85 Ovarian Follicular Development in Egg and Growth Lines of Quail, by W. L. Bacon, K. E. Nestor, and P. A. Renner 89 The Half Life of the Very Low Density Lipoprotein in Male and Laying Female Turkeys, by W. L. Bacon 92 Plasma Free Fatty Acid Levels in Laying Turkeys, by W. L. Bacon and Margery A. Musser 95 Progress in Turkey Semen Research, by K. I. Brown 97 ON THE COVER: Two flocks, one infected with Mycoplasma meleagridis (Mm) and the other free of Mm, were raised at the Ohio Agricultural Research and Development Center. The photos of both flocks were taken at 16 weeks of age (top: Mm-free, bottom: Mm-infected). The economic significance of Mycoplasma meleagridis is discussed in the article on page M

4 EFFECTS OF VARIOUS LITTER TREATMENTS ON GROWING TURKEYS Karl E. Nestor and Philip A. Renner Department of Poultry Science Leg problems have become increasingly important in the modern large white turkey. Although the specific cause or causes of leg weakness have not been discovered, various diseases, nutritional deficiencies, management factors, and breeding have been implicated. The influence of litter condition on leg weakness of turkeys has not been adequately studied. Therefore, an experiment was run in which the influences of various litter treatments were studied. Hardwood shavings litter was used in all trials. Pen size (1 ft. x 11 ft. 4 in.) was the same for all trials. In Trial 1, the influence of superphosphate (-2-) was studied. Superphosphate was added at the rate of 2.6 Ib. per pen each week, starting the fifth week after hatching and continuing until the birds were 8 weeks of age. The effect of addition of hydrated lime (1.4 Ib. per pen per week) was studied in Trial 2. In both trials, the litter was stirred approximately once each week in all pens, with the same amount of new litter (88 Ib. per pen) added each week to replace litter removed from around the feeders and waterers. In the last two trials, the influence of stirring vs. not stirring was studied. The primary difference between these two trials was that litter was removed from around feeders and waterers with new litter added in Trial 3, but not in Trial 4. All experimental treatments were started during the fifth week of age and continued until the birds were transferred to range at 8 weeks of age. All birds within a trial were reared intermingled on range. Moisture and ph measurements of the litter were made when the birds were 8 weeks of age. Body weights were obtained at 8 and 16 weeks of age. Mortality was summarized for the first 8 weeks, while the incidence of crooked toes and bad legs was recorded at 16 weeks of age. The various treatments had no significant effect on percent moisture in the litter when the birds were 8 weeks of age (Table 1). Addition of superphosphate lowered ph, while addition of hydrated lime increased ph. Stirring the litter in Trials 3 and 4 increased the ph slightly in comparison to pens in which the litter was not stirred. The only statistically significant difference in growth to 8 weeks of age was observed in the comparison of lime vs. no lime in Trial 2. Birds grown in pens in which no lime was added were heavier than those grown in pens in which lime was added. The difference between the two groups disappeared after the birds were placed on range. The birds grew slightly (but not significantly) faster in pens in which the litter was stirred in Trials 3 and 4. The reasons for the differences in growth resulting from litter treatment were not readily apparent. There were no significant differences between any of the treatments in frequency of crooked toes and bad legs. Mortality was not significantly different between any of the treatments. To estimate if ph and percent moisture of the litter had any effect on leg problems and mortality, data were summarized across treatments and are presented in Table 2. The frequency of crooked toes tended to decrease with increase in ph and -1.
5
6 decrease in litter moisture, although the correlations were not statistically significant. The mortality also tended to decrease with increase in moisture, but the effect was not statistically significant. The frequency of bad legs was not influenced by either ph or percent moisture within the ranges observed in this experiment. In studies of this kind, several factors could be important in the results obtained. One is bird density. The bird density in this experiment varied from tria to trial, but in no case was the bird density great. None of the pens in this experiment were extremely wet. The amount of change in ph also could be important. Perhaps the results would have been different if amounts of lime and superphosphate other than those arbitrarily used had been tested. The results suggest that modera changes in ph and moisture may influence the frequency of crooked toes. Table 2. The Effect of ph and Percent Moisture of Litter on Frequency of Crooked Toes, Frequency of Bad Legs, and Percent Mortality of Heavy Weight Turkeys. ph No. of Pens % Crooked Toes % Mortality % Bad Legs 7.4 to to to Correlation Moisture (%) to to to to to Correlation

7 A STUDY OF CROOKED TOES IN TURKEYS Karl E. Nestor Department of Poultry Science The condition in turkeys commonly called crooked toes is usually characterized by a lateral or medial deviation of either the second, third, or fourth digit, and quite frequently more than one digit is involved. One or both feet may be affected. A recent survey of the frequency of crooked toes in commercial flocks of turke> indicated that this condition is widespread, occurring in all 63 flocks surveyed (Supeene, 1971, Poultry Digest, 3: 349). Based on the results of this survey, the frequency of crooked toes was believed to be influenced by management, with little influence of genetics and nutrition. For two consecutive generations, a survey was made of the frequency of crooked toes in several medium and heavy weight strains maintained in genetic studies. The frequency of crooked toes among offspring from parents of which at least one had crooked toes was more than double the frequency among offspring of parents with normal toes (Table 1). This indicates that the tendency for crooked toes is inherited. Further evidence that crooked toes is influenced by inheritance is provided by the comparison of the frequency of crooked toes in medium weight vs. large weight strains in 197. Although no significant difference occurred between strains within each weight classification during this year, there was a highly significant difference between the incidence in the large and medium weight strains. Although nonsignificant, a similar difference between the two weight classifications existed in A highly significant strain difference in frequency of crooked toes was evident in 1971 for the medium weight strains. The medium weight strains used in this study were similar to the large weight white turkeys in 1957, while the large-bodied strains were established from commercial lines in The lower incidence of crooked toes in heavy strains indicates that commercial turkey breeders reduced the incidence from 1957 to The 24-week body weight of males with crooked toes was less than males with normal feet (21.6 vs Ib. for the medium weight strains and 29.2 vs. 3.1 Ib. for heavy weight strains) in 197. The differences approached significance at the 5 percent level. In 1971, a comparison was made only of families which had both normal toed and crooked toed individuals of the same sex in the same hatch. Such a paired comparison is much more efficient, even though based on smaller numbers. The results are presented in Table 2. Although the crooked toed brothers and sisters tended to be heavier than their normal toed sibs at 8 weeks of age, they did not grow as fast after this time, resulting in the normal toed brothers and sisters being significantly heavier at 16 and 24 weeks of age. The commercial turkey breeder may have reduced the incidence of crooked toes simply by selecting for increased body weight. Environmental factors also influence the incidence of crooked toes. Highly significant differences were observed between the frequency in four weekly hatches.4-
8 Table 1. The Occurrence of Crooked Toes (%) According to Phenotype of Parent, Line and Year. Medium Wt Strains Randombred Control Egg Semen Egg Selected Low Line High Line Average Heavy Wt Strains 197 Phenotype of Parent Crooked Toes Normal Toes »_ «.-._._ 16.4 Average of Both Phenotype of Parent Crooked Toes Normal Toes Average of Both A Randombred Control 16 Week Index Clutch Broody Average Total for Years No parents with crooked toes were present. Frequency of crooked toes differed highly significantly between types of ^parent. Frequency of crooked toes differed highly between weight classification in 197 but not in Difference between strains was highly significant only for the medium weight strains in Table 2. Body Weights of Full Sibs with Normal and Crooked Toes. No. Observations 8 Normal Toes Wks 16 Vvks 24 Wks Crooked Toes 8 Wks 16 Wks 24 Wks 8 Wks % of Normal Toed Sibs 16 Wks 24 Wks Medium Wt, Male Medium Wt, Female , Heavy Wt, Hale Heavy Wt, Female Average (61) 4, * 95.9** *Difference between normal and crooked toed individuals was significant (P<,5). **Difference between normal and crooked toed individuals was highly significant (P<.1). ( ) Total number of observations. -5-
9 (Table 3), even though they \\ere brooded in the same house under approximately similar management conditions. The results of this study indicate that the crooked toe condition is influenced by both inheritance and environment. Individuals with crooked toes tend to be smaller at older ages. Table 3. Frequency of Crooked Toes (%) in Different Hatches in Hatch Medium Wt Strains Randombred Control Egg Semen Egg-Growth Average Heavy Wt Strains Randombred Control 16 Week Index Average Overall Average ] Difference between hatches was highly significant.
10 THE INFLUENCE OF EGG COLOR ON HATCHABILITY OF TURKEY EGGS Karl E. Nestor and Philip A. Renner Department of Poultry Science Several strains of turkeys are used In OARDC genetic experiments. When setting eggs from these strains, it appeared that there was considerable variation in the color of eggs between strains. Individual hens within a given strain also laid eggs which were greatly different in color. The color of eggs from the various strains was evaluated periodically over a 12-week hatching period beginning when the hens first attained a 5 percent level of production. Shell color was scored from 1 to 5, with 1 being very dark speckled eggs and 5 being white eggs. At each determination, the eggs from each hen were independently rated by two people and the average of these two scores was used as the value for the hen. Four medium weight and three large-bodied strains were surveyed. The medium weight strains consisted of a randombred control population developed in 1957 and three selected sublines. One of these (egg) had been selected since 196 for increased egg production. Another (semen) had been selected since 1966 for increased semen yield. The other medium weight line (egg-growth) was a subline of the egg line selected for increased 16-week body weight. The large-bodied strains were a randombred control population established in 1967 from two elite large-bodied commercial strains, a subline (16 week) of the control selected for increased 16-week body weight, and a subline (index) of the control selected for both increased body weight and increased egg production by means of an index. The egg color, hatchability of fertile eggs, and egg weight of the different strains are shown in Table 1. Egg color did not differ significantly between strains within the medium weight or large-bodied groups. However, eggs laid by the egg strain were a little lighter and those laid by the semen line a little darker than the eggs laid by the corresponding randombred control. The large-bodied strains had darker eggs and poorer hatchability than the medium weight strains. Selection for increased egg production apparently reduced the variation in egg color, hatchability, and egg weight. Mean egg weight was also reduced by selection for high egg production. The average score for egg color declined as the hatching period advanced. This possibly indicates a decline in the amount of shell placed on the eggs later in the hatching period. Correlation coefficients were calculated between shell color, hatchability, and egg weight after the differences between strains were removed by statistical analysis. The correlation between shell color and hatchability was -,15, which was statistically significant (greater than zero). This indicates that darker colored eggs tend to hatch better. However, only 2 percent of the variation in hatchability can be accounted for by differences in shell color. Smaller eggs hatch better than larger eggs. A negative correlation of -.18, which was highly significant, was observed between egg weight and hatchability. Thus, variation in egg weight would account for about 3 percent of the variation in hatchabilty. The results of this study indicate strain variation In egg color. However, this variation contributes only slightly to differences in hatchability between hens. -7.
11 Table 1. Shell Color, Hatchability of Fertile Eggs and Egg Weight of Several Strains of Turkeys Over a 12-Week Hatching Period (1969). Mean + Standard Error Egg Color Hatchability (%) Egg Wt.(gm) Medium Weight Randombred Control *8+.99 Egg Semen Egg-Growth Large Weight Randombred Control Week Index
12 PRODUCTION OF DEFECTIVE EGGS BY EGG- AND MEAT-TYPE TURKEY HENS HOUSED IN CAGES Karl E. Nestor and Wayne L. Bacon Department of Poultry Science Recent research by Dr. R. George Jaap and coworkers in this department indicates that meat-type chicken females lay more abnormal eggs than egg-type chickens. Dr. Jaap theorized that inheritance which favors rapid protein anabolism (rapid body growth) may also favor rapid formation of lipoprotein in the liver and increased yolk production in ovarian follicles. However, a lack of synchronization exists between ovarian and oviduct functions of broiler pullets, and this was believed to be the major reason for the defective egg syndrome of these pullets. Results of OARDC studies (page 85) indicate that meat-type turkeys also have more ovarian ova (future yolks) in rapid development than egg-type turkeys, particularly early in the laying period. However, apparently some of these ova are lost because the egg-type turkeys lay more eggs. The question arises as to what happens to the extra ova in the meat-type turkey. Several possibilities exist. Among these are: 1) production of double yolked eggs; 2) loss of yolks through production of shell-less and thin shelled eggs, which are broken and not noticed under conditions of floor rearing; 3) attrition of follicles after rapid development has started; and 4) loss of yolks in the body cavity. The purpose of this experiment was to compare the production of abnormal eggs by egg- and meat-type hens housed in cages and to obtain some information of the performance of turkey hens in cages vs. hens in floor pens. The production of abnormal eggs by females of an egg strain and two growth strains was compared. The growth strains were a large-bodied randombred control strain and a subline of the randombred control population which was selected for increased 16-week body weight. The body weight at 5 percent production of these strains was 16,5, 23.2, and 26.8 lb., respectively, for the egg, randombred control, and 16-week strains. A sample of 48 females from each line was randomly selected, reared in confinement to 8 weeks of age, and then grown under range conditions until 24 weeks of age. At this time they were housed in cages measuring 15.4 inches wide, 24. inches deep, and 24. inches high, with a rigid plastic-covered wire floor. The females were restricted to a 6-hour light day for 6 weeks prior to lighting for egg production. Stimulatory lighting of 14 hours per day was given at 34 ± 1 weeks of age beginning Dec. 9, 197. The lights were turned on at 3 a.m. and were turned off at 5 p.m. Eggs were gathered periodically from 8 a.m. to 4 p.m. The time of lay and condition of the eggs were recorded. Those eggs gathered at 8 a.m. were considered laid from 4 p.m. to 8 a.m. A 24-hour period was based on eggs laid from 8 a.m. one day to 8 a.m. the following day. Eggs were recorded as apparently normal, membrane, soft-shelled, broken but with apparently normal shell, and slab-sided. Occasionally, the membrane and soft-shelled eggs would break and fall through the cage floor to the manure below. Such eggs were recorded and removed at each gathering to prevent errors in recording. Double-yolked eggs were identified by holding all eggs for 1 week and then comparing the eggs of each hen. Eggs classified as double-yolked were broken and checked for accuracy of classification. -9-
 and Two Large-Bodied Growth Lines (Randombred Control (RBC) and 16 Week).")
13 Table 1. Production of Normal and Abnormal Eggs by Caged Females from a Medium Weight Line Selected for Increased Egg Production (E) and Two Large-Bodied Growth Lines (Randombred Control (RBC) and 16 Week). No. Total Normal Broken Membrane Soft Double "Slab Two Eggs/24 Line Hens Eggs 2 Eggs 2 Eggs 2 Eggs 2 Shelled Yolked Sided 11 Hour Period 2 (No) (%) (%) (%) (%) (%) (%) (%) E RBC Week ^135-day production period beginning from the laying of the first egg. Highly significant line difference. The number of eggs laid in a 135-day production period was higher for egg line females than for meat-type females (Table 1), Egg production for the randombred control (RBC) was higher than that for the 16-week line. The meat-type hens had a larger percentage of broken eggs* membrane eggs, and soft-shelled eggs than the egg- ;ype hens. However, the difference between the egg and randombred control lines was not statistically significant for membrane and soft-shelled eggs. The frequency of double-yolked eggs did not differ significantly between eggand meat-type turkeys. OARDC research (page 85) suggests that a higher frequency of ova develop in pairs in the randombred control females than in the egg line females. Since there was no significant difference between these two strains in frequency of louble-yolked eggs, either one of the ova developing in pairs must be lost after wulation or the members of the pairs must be ovulated on different days. The frequency of two eggs laid per 24-hour period was greater (P<.1) in the 16-week line than in the egg or randombred control strains (Table 1). The frequency in the egg strain was greater than in the randombred control strain. The 16-week strain females laid a larger percentage of two membrane eggs per Jay than the other two strains (Table 2). Although the differences were not always statistically significant, the meat-type females laid a larger percentage of eggs in which both eggs were abnormal. This was partially the result of the higher frequency of abnormal eggs in these strains because a highly significant correlation of.25 was obtained between the frequency of abnormal eggs and frequency of two abnormal eggs laid during a 24-hour period. Three eggs were laid in a 24-hour period by six hens. difference between lines in this regard. There was no apparent The question arose as to whether the cage environment placed a stress on the turkey hen. To test this, a sample of hens obtained from the egg line during the same generation as the ones housed in cages were reared similarly to the egg line bens housed in cages, However, the hens in floor pens were brought into production during February at approximately 42 weeks of age. The average egg production of -1-
14
15 the egg line females housed in floor pens was similar (81,2 eggs) to that obtained in cages (8.8). The higher frequency of broken eggs from the growth type hens may have resulted from the hens being slightly taller, thus dropping their eggs a greater distance. The heavier hens also may have broken more eggs after they were laid as a result of their heavier weight. -12-
16 THE INFLUENCE OF AMBIENT TEMPERATURE ON REPRODUCTION OF TURKEY HENS Karl E. Nestor, Sherman P. Touchburn, and Philip A. Renner Department of Poultry Science It has been well established that seasonal declines in egg production and in fertility and hatchability of fertile eggs occur in spring-hatched turkey hens. Some experiments have indicated that the seasonal decline in fertility is mainly due to the female. Previous research indicates that environments with protection only against extreme temperatures result in better egg production than constantly high (65 F.) or constantly low (5 F.) temperature environments. The purpose of this experiment was to study the influence of ambient temperature with diurnal variation on reproduction of turkey hens. Four medium weight and three large-bodied lines of turkey hens were utilized in each of two studies. The hens were subjected to a restricted lighting program of 6 hours per day for an 8- week period prior to stimulatory lighting. On Feb. 1 when the hens were 39-2 weeks of age, stimulatory lighting of 14 hours per day was provided. Each hen was artificially inseminated with semen from a different male using a paired mating system. Males from all lines were housed in a pole shed structure, with the medium weight and large-bodied strains maintained in separate pens. The hens were inseminated when the first egg was laid. The second insemination was made 1 week later and bi-weekly inseminations were used thereafter. Egg production was recorded for 18 days, beginning when the first egg was laid. Fertility, hatchability of fertile eggs, and number of poults produced per hen were measured for a 12-week hatching season beginning when the hens first obtained a level of approximately 5 percent production. Body weights were recorded at the beginning of the hatching season. Egg weights for each hen were estimated by group weighing of the eggs laid in alternate weeks throughout the hatching season. Feed consumption was measured for four periods in Study 1: 1) Feb. 1 to March 27, 2) March 28 to May 22, 3) May 23 to July 9, and 4) July 1 to August 18; and three periods in Study 2: 1) Feb. 1 to March 17, 2) March 18 to May 19, and 3) May 2 to August 17. The hens were housed in the same insulated windowless breeder house in both studies. This house was equipped with fan ventilation and heating units which allowed the temperature to be partially controlled during cooler parts of the laying period but not during the warm summer months. Two experiments designated as A and B were used in Study 1, The average temperatures of these environments are presented in Table 1. The average temperature for each day was based on measurements made at 6: a.m., 12: m., 6: p.m., and 12: p.m. In both environments, an attempt was made to produce diurnal variation in the ambient temperature. The average difference between day and night temperatures was 2.9 F. during period 1 and 2.2 F. during period 2. No diurnal variation could be obtained in the two later periods as the result of high ambient temperature. Temperatures in the two environments did not begin overlapping until April 8. From April 8 until May 31 in the B environment, temperatures averaged less than those in -13-
17 Table 1. Average Temperatures ( F) of Environments Used in Studies 1 and 2. Study1 Study 2 Date A B Date C 21 3 to 3/ to 3/ /28 to 5/ /18 to 5/ /23 to /2 to 8/ /1 to 8/ the A environment., but there was some overlapping in the daily range. time, the temperatures were similar in the two environments. After this Two environments (C and D) were used in Study 2 (Table 1). An attempt was made to have environment C similar to environment B in Study 1 and to have environment D cooler than C earlier in the laying period. This was not possible because the outside temperature was higher than normal during this study. The average temperature of environment C was between that of environments A and B. Also, the temperature differential was maintained between environments C and D for only a short period of time. There was no overlapping of temperature until March 2. The average temperature was slightly different between the two environments until April 7, after which temperatures in both environments were similar. A diurnal variation in temperature similar to that obtained in Study 1 was maintained in this study. In both studies, hens were exposed to the experimentally imposed environment 7 days prior to stimulatory lighting. Prior to that time, all hens were given a similar environment. The cooler environment (B) in Study 1 resulted in greater egg production, egg weight, fertility, and number of poults produced per hen than the warmer environment (Table 2). In Study 2, there were no significant differences between environments Z and D for any of these traits. The results of Study 1 suggest that a moderately cool environment with diurnal variation in temperature may be beneficial to reproduction of turkey hens. The lack of a difference in Study 2 was probably the result of not maintaining the temperature differential between the two environments for a long enough period of time. In Study 1 there were significant interactions between environments and lines for number of poults produced per hen (P<.5) and for egg weight (P<.1). Means for these traits are presented in Table 3 according to line and environment. The cooler environment (B) resulted in a large increase in number of poults produced per hen in three of four medium weight lines and one of three large-bodied lines. There Aras very little difference between environments in the remaining three lines. The cooler environment resulted in larger average egg weights for the egg selected and 16-week lines than the warmer environments. Of the remaining five lines, four had slightly larger egg weights in the warmer environment. It appears that environmental temperature within the ranges used in this study did not ;onsistently influence egg weight.
18 Table 2. Reproduction of Turkey Hens Exposed to Four Temperature Environments. Environment No. 84 Hens Days Egg Production (No. /Bird) 12 Days 18 Days Days to First % Egg 1 Fertility- % Hatch. Fertile Eggs No. Poults /Hen Egg Weigh: (gna.) Study 1 A ,3 B ,8** 72.1** 96.5** 21.** 82.4* * 87,7* Study 2 C D * ^Days from stimulatory lighting to production of first egg. *Difference between environments within study was significant. Difference between environments within study was highly significant. Table 3. Number of Poults Produced Per Hen and Egg Weight According to Environment (A and B) and Line in Study 1. Line No. Poults/ No. Hens Hen A B A B Egg Weight (gin.) A B Medium Weight Randombred Control Egg Semen Egg Selected Average Large Bodied Randombred Control Week Index Average
19
20 The time required from stimulatory lighting to production of the first egg was less in the cooler environment (B) than in the warmer environment (A) in Study 1. The reverse was evident in Study 2. The reason for the different results in the two studies is unknown. Feed consumption per hen according to periods is presented in Table 4. In Study 1, feed consumption was significantly greater for the cooler environment only in period 2 (March 28 to May 22). Although there were no significant differences in period 1 (Feb. 1 to March 27), consumption was slightly higher in the warmer environment. Differences in egg production were significant for periods 2 and 3. These data suggest that the cooler environment may have stimulated an increase in egg production which in turn resulted in greater feed consumption. Feed consumption per egg produced was not significantly different between environments in either study. Interactions were observed between environmental temperature and line for egg weight, number of poults produced per hen, and days from stimulatory lighting to production of first egg. Several interactions were also observed in these same lines in a lighting experiment. These results indicate that strains selected for different purposes or which are greatly different in body weight may not react the same to environmental conditions and thus no general recommendation would strictly apply. Ideally, the optimum environment should be determined for each strain of bird. -17-
21 A POSSIBLE INFLUENCE OF PRESENCE OF MALES ON EGG PRODUCTION OF TURKEY HENS Karl E. Nestor and Philip A. Renner Department of Poultry Science The underlying physiological mechanisms influencing reproduction of turkey hens are not completely known. It is known that neurological mechanisms are involved* Some data indicating that the presence of turkey males might be beneficial to egg production of turkey hens was obtained during one generation in a genetic study. Six pens of turkey females were housed in adjacent pens to males in a 12-pen section of a window less breeder house. The remaining hens on the genetic study were housed in two other sections of the breeder house which were out of hearing distance of the males. The number of males in the pens was the same as the number of females. All strains were maintained intermingled in each pen. The temperature control in each section was set the same and periodically checked, but no recording of temperature was made during the course of the laying period. Egg production of the two groups of females was similar early (84 and 12 days of production) in the laying period (Table 1). However, the females housed adjacent to the males laid at a greater rate from 12 to 18 days of production than females housed away from the males. There was a difference in 18-day production of 8. and 5,1 eggs per hen for the medium weight and large weight lines. Only one of eight strains (index) exhibited greater egg production in the group housed separately from the males. A study was made in a later generation to further test the influence of presence of males on egg production. In this study, eight males were housed in two colony cages (four males in each cage) in a 12-pen section of the breeder house. The male cages were high enough so that all females in the section could see and hear the males. Control females were housed out of sight and hearing distance of the males. The environmental temperature was similar in all sections. The stimulus of males, if any, would not be expected to be as great in this study as in the previous one because many of the females were not as close to the males and fewer males were involved in the present study. The results are presented in Table 2. When all medium weight strains were considered, the presence of toms increased egg production, particularly late in the laying period during May through August. However, upon inspection of the data, it was observed that this effect was primarily on two relatively low producing strains. When only the data for these two strains were considered, a large difference of 19.2 eggs per hen was observed between hens housed in the presence of toms and those housed separately. The presence of toms had no influence on the high-producing strains of medium weight turkeys, even though they were housed in the same pens as the low-producing strains. The presence of toms stimulated the hens of all strains into production more rapidly following stimulatory lighting. This effect was not observed in the previous comparison (Table 1). The presence of toms did not appear to have a large effect on egg production of large-weight strains (Table 2). However, when the strains were grouped into highproducing and low-producing groups, a difference was apparent. The presence of toms increased the egg production of the low-producing strains but appeared to decrease the 18-day egg production of the high-producing strains. It is very doubtful if the presence of toms would reduce egg production under the conditions of this study. It is more likely that some adverse environmental condition influenced the egg pro- -18-
22
23

24 THE RELATION BETWEEN BROODINESS, INTENSITY OF LAY, AND TOTAL EGG PRODUCTION OF TURKEYS Karl E. Nestor Department of Poultry Science Total egg production of turkeys is a complex trait which is influenced by several component parts. Among these are time required from stimulatory lighting to production of first egg (i.e., a measure of responsiveness to light), broodiness traits (number of broody periods, average length of broody periods, and total days lost from broodiness), and intensity of lay traits (average clutch length, number of clutches, maximum clutch length, and rate of lay). The relation between the component parts and total egg production can be the result of genetic covariation (genetic correlation), environmental covariation (environmental correlation), or, a combination of the two (phenotypic correlation). Generally, correlation coefficients range from -1. to +1. and the size of the correlation coefficient measures the closeness of the relationship. Those approaching either -1. or +1, indicate a close association between the two traits in question. A negative sign on the correlation coefficient indicates that as one trait increases in magnitude, the other trait tends to diminish. A positive correlation coefficient indicates that both traits tend to increase or decrease together. The total variation observed in any trait is the result of genetic differences and environmental variation. The heritability (h2) of a trait measures the genetic variation present relative to total variation. It is possible that breeders might be able to make more progress in total egg production by selecting for one of the component parts. To test this, egg production records were analyzed for three strains of large-bodied turkeys over a period of 3 years. The records also were analyzed for four strains of medium weight turkeys over a period of 9 years. All hens were given stimulatory lighting (14 hours per day) when they were approximately 39 weeks of age on Feb. 1. Various changes in housing, nutrition, and management were made over the years. Phenotypic correlation coefficients were calculated after the data had been adjusted for the effects of lines, years, and lines x years interaction. These are presented in Table 1. All measures of broodiness were negatively correlated with total egg production. This indicates that as these broody traits increase, total egg production decreases. For the purpose of analysis, a broody period was considered as a period of non-production for 5 or more consecutive days. Thus some non-broody pauses may have been included. The magnitude of the correlations (Table 1) involving number of broody periods and average length of the broody periods were similar, while those for total days lost from broodiness were higher. This indicates that total days lost from broodiness is a superior measure of amount of broodiness. Clutches are the periods (measured in days) where one or more eggs are produced before a day when no eggs are produced. Maximum clutch length, number of clutches, and rate of lay were closely associated positively with total egg number (Table 1), Rate of lay was obtained by dividing the total eggs produced for a given period by the total days in the period minus those days lost from broodiness. -21-
25 Table 1. Phenotypic Correlation Coefficients Between 1 tal Number of Eggs Laid and Measurement of Broodiness and Intensity of Lay, Total No. of Eggs Laid (18 Days) Heavy Weight Medium Weight Crrelated with: Strains Strains Number Broody Periods^ -84 Days -,48-18 Days -.47 Average Length ofbroody Periods -34 Days Days Tota 1 Days Los t^ f rom Brood i.ne&s -84 Days Days Days Maximum Clutch Length.38 Number of Clutches -84 Days Days Days Days Days Average Clutch Length -84 Days Days Days Days Days Days from Lighting to First Egg -.12 a Rate of Lay -84 Days Days Days.85.7 Total Egg Production -84 Days Days Days Days.89 a Not significantly different from zero.all other correlation coefficients were significantly larger than zero. -22-
26 It is interesting that the number and length of clutches between 12 and 18 days had a closer association with total egg production for the 18-day period than similar measurements made over the total period. Likewise, the number of eggs produced during the period was more closely related to the total than the number of eggs produced during the periods of to 84 and 84 to 12 days. The rate of response to light at the beginning of lay did not have a significant effect on total egg production for the 18-day period. Table 2, Phenotypic Correlation Coefficients Between Measures of Broodiness and Intensity of Lay for a 18-Day Production Period In Large Bodied Turkeys. Correlated with: No. Clutch Rate of Maximum Clutch Days to Clutches Length Lay Length First Egg No. Broody Periods Ave, Length Broody Periods a -.1 a Total Days Lost From Broodiness, a Days to First Egg, Correlation coefficient was not significantly different from zero. All others were significant (P<.5). Table 3. Phenotypic Correlation Coefficients^ Between Different Measures of Broodiness. No. Broody Periods -18 Days Ave. Length of Broody Periods -84 Days Days Days Total Days Lost From Broodiness Days Days No. Broody Periods -84 Days -12 Days -18 Days Ave. Length of Broody Periods -84 Days -12 Days -18 Days All correlation coefficients were highly significantly different from zero. -23-

27 Generally, all measures of broodiness were negatively associated with measures of intensity of lay except for maximum clutch length (Table 2). This indicates that hens which go broody frequently do not lay at as high a rate when they are not broody* The number of broody periods was positively associated with the average length of broody periods (Table 3). In other words, hens which have more broody periods also have longer broody periods. The number and average length of the broody periods had a similar influence on total days lost from broodiness. The number of clutches was negatively correlated with average clutch length within a given period of time (Table 4). Both number of clutches and average length of clutches were positively correlated with rate of lay. Maximum clutch length was closely associated with average clutch length. The correlation coefficient of.7 indicates that 49% of the variance in average clutch length can be explained by variation in the size of the longest clutch. The heritability estimate of.29 for the 18-day egg production (Table 5) was similar to an estimate of.33 obtained for 84-day egg production based on nine generations of selection in an egg line. Total days lost from broodiness was the most highly inherited broodiness trait, while maximum clutch length and average clutch length were more highly heritable than number of clutches or rate of lay. All measures of broodiness and rate of response to light were negatively correlated genetically with the number of eggs produced (Table 6). The measures of intensity of lay were positively correlated with total number of eggs produced. Number of broody periods and total days lost from broodiness had little genetic association with maximum clutch length and average clutch length. This indicates that whenever a hen initiates a clutch, the clutch size is not restricted by broodiness tendencies. All measures of broodiness were negatively associated with the number of clutches. This was expected since any increase in broodiness would result in fewer days for clutch number to increase. From the tesults, based on heritabilities and phenotypic and genetic correlations, total days lost from broodiness and maximum or average clutch length are the best measures of broodiness and intensity of lay, respectively. The question arose as to whether selection for any of these traits would result in greater genetic gains than selecting directly for total egg number. When the same intensity of selection can be achieved when selecting for character Y as when selecting for character X, then the correlated response in X will be greater than the direct response if the genetic correlation (r a ) times the square root of the heritability of trait Y (hy) is greater than the square root of the heritability of trait X. The square root of heritability of total number of eggs was.53, which was larger than r a hy for maximum clutch length (.5), average clutch length (.45), or total days lost from broodiness (.39). Therefore, it appears that gains in total number of eggs would be greater by selecting for increased egg production itself than by selecting for a decrease in total days lost from broodiness, an increase in maximum clutch size, or an increase in average clutch size. -24-
28 Table 4. Pher nt/p \? ^ovi'i*.a'~ Intensity of Lay, ">'*: \," i f L, 1 ;*<(r >^'>/eei Measures of l*! 8 Clutches AVP LC?TJJJ, tvt C"! uteri Maximum i)~ iso Cratch Rate -84 Days of La 1-11 Day: No. of Clutches Days, ,85 -, a ) ,9 a.52.4:.31 Ave. Clutch -84 Days -18 Days, : Correlation coeff LcLorus wo.rn nor. sn gnili^antl^ difi>reof from zero. All others significant (?<.5) Table r 5. '*<or «tab! LI tv r*s tnr.it r«? from DauRHter-On-Dam Regressiooc Ineir Standard ^rrnra (s»e.) for a 18-Day ProductioD Period. Trait Total of * 29 ±' 32 Number of Length of Broody.364-* 12 Total Days,4fi-K!2 Days from to Egg -8ijh.il Maximum Clutch Number of Clutches.47--K3I.27-f-.ll Average Length of Clutches.9J4-11 Rate of Lay.28f.ll -25-
29 Table 6, Genetic Correlation Estimates (r) and Their Standard Errors (s*e.) Among Broodiness, Intensity of Lay and Total Number of for a 18-Day Production Period. Traits Correlated r+s.e..number _of...eggs...produced and: Number Broody Periods Length Broody Periods Total Days Lost From Broodiness Maximum Clutch Length Number of Clutches Average Clutch Length Days to First Egg Rate of Lay Length Broody Periods " Total Days Lost From Broodiness Maximum Clutch Length Number of Clutches -, Average Clutch Length Rate of Lay * 32 ±- 25 Days to First Egg L eng t h of Broody jp.griods^andj_ Total Days Lost From Broodiness Maximum Clutch Length Number of Clutches Average Clutch Length Rate of Lay Days to First Egg.42+, 12 To tal Days Lost From Broodiness and: Maximum Clutch Number of Clutches Average Clutch Length Rate of Lay Days to First Egg ' 13 Maximum Clu t cli L_ength...and, : Number of Clutches -»34+*19 Average Clutch Length,72+, 1 Rate of Lay Days to First Egg ~,S7+»G9 Average Clutch Length Rate of Lay Days to First Egg Average Clutch...Length...andj Rate of Lay Days to First Egg Days, to First.Egg Rate of Lay
30 REPRODUCTIVE OF TO Wayne L. Bacon Department of Poultry Science Controlled lighting programs have evolved in the commercial production of turkey hatching eggs to the point where they are a very important aspect of practical management in achieving optimum reproductive performance (Brown? 1966, Turkey Research Summary No. 17, pp. 2-5). In Coturnix quail, certain responses of the reproductive system to various lighting programs are very similar to those observed in turkeys, In quail, the relationship between sexual maturity and lighting programs has been studied. Wilson et al. (1962, Poultry Sci., 41: 17-22) found that sexual maturity was initiated at an earlier age and subsequently maintained if quail were exposed to a single light period of 12 or more hours per day. A lighting program consisting of 1 hour of light followed by 5 hours of darkness (1L:3D), repeated 6 times (6X) per day, also was found to be as efficient as constant illumination In initiating sexual maturity. Shifting quail reared under 6L:18D to 14L:1D initiated egg laying in 28 days. It is recommended (Brown, 1966, Turkey Research Summary No. 17, pp. 2-5) that female large white turkeys be restricted to 6 hours of light per 24-hour day at 21 weeks of age and be given stimulatory light of 14 hours per day at 31 weeks of age. Nestor and Brown (1972, Poultry Sci., 51: ) found that egg production was greater in large white turkeys exposed to 4.6 foot candles (FC) of light intensity than those given 3. and 1.6 FC. The lighting program is thus important in timing the induction of sexual maturity in turkeys and quail and in maintaining this state after it is attained. In large white turkeys, light intensity is also an important factor in maintaining egg production. The objectives of the presently reported experiments were: 1) to determine the number of 1-hour light periods, interspaced between dark periods of 1, 2, or 3 hours, necessary to induce sexual maturity in male and female quail and to maintain egg production in female quail equal to that of the stimulatory regimen of 14L:1D; and 2) to apply the results of these experiments to turkeys. A randomly mated breeding colony of Coturnix quail is maintained at the OARDC Poultry Research Unit. Day-old chicks derived from this population were battery reared to 7 weeks of age under 6 hours of light per day (6L:18D). At this time they were placed in individual bird pens (4 x 1 inches) and exposed to one of the lighting programs given in Table 1. Beginning the following day (day 1 of experiment), the following observations were made every second day (Experiments 1 and 2) or daily (Experiment 3): Females check for eggs. Males check for sexual development of the caudal gland. Give a visual score of for no development, 1 for gland with some foamy material and the appearance of a red color to the gland, or 2 an enlarged gland with foam and a definite pocketing or cavity in the gland. -27-

31 When a 2 was given for two observations in Experiments 1 and 2 or three observations in Experiment 3, the bird was no longer examined. Date of sexual maturity of males was the first day assigned a 1 with no intermittent scores of until the bird was no longer examined. The results of these experiments are given in Table 1 for the females and Table 2 for the males. Equivalent lighting periods of S, 6, 7, 9, 11, 13, 14, 16, 17, and 21 hours per day were given in the experiments. The equivalent lighting period is defined as the interval elapsing between the initiation of the first 1- hour light period to the end of the last light period, or for those exposed to 6L:18D and 14L:1D, the duration of the light period. The responses (sexual maturity in both males and females and reproductive performance in females) can be easily broken into three groups: 1) those with an equivalent lighting interval of less than 9 hours which were not stimulated, 2) those with an equivalent lighting interval of more than 13 hours which were stimulated, and 3) the one group with an equivalent lighting interval of 11 hours which was between the response of groups 1 and 2, In group 1 (those not stimulated), the 6L:18D group can be used as a control group. When this experiment was designed, it was postulated, based on the data of Follett and Earner (1966, Gen. Comp, Endocrinol., 7: ), that none of the birds in the 6L:18D treatment would become sexually mature. However, this is not what was observed (Table 1). Fifty-eight to 78 percent of all the hens in Group 1 treatments laid one or more eggs (average 43 to 58 eggs per hen), with a hen day percent from 65 to 77. The mean days to first egg, for those hens laying one or more eggs, was from 41 to 64 days after being placed on the treatments at 7 weeks of age. In the Group 1 males (Table 2), the average percent responding ranged from 47 to 83 and the average days to initial response ranged from 34 to 57 days. There is no apparent reason for the difference in these results and those of Follett and Farner (1966). In Group 2 treatments (stimulated), almost all birds were in production during the greater part of the 126-day experimental period (98-1% responded in all treatments), with the mean days to first egg ranging from 19 to 23 days. The Group 2 treatments also laid more eggs during the 126-day experimental period (range of mean number of eggs was 81 to 95) and had a slightly higher hen day percent production (range 77 to 88) than Group 1 treatments. The males from Group 2 treatments responded almost completely (98-1%) and very quickly to the stimulatory lighting, with days to first response ranging from 4,8 to 8.8. The Group 3 lighting treatment with an equivalent lighting period of 11 hours was intermediate in response of both males and females to those in treatment Groups 1 and 2. These experiments indicate that in quail, 1 hour of light followed by 2 or 3 hours of darkness, repeated often enough to give an equivalent lighting period of at least 13 hours, is as effective in inducing sexual maturity in both males and females, and maintaining egg production in females, as the stimulatory treatment of 14 hours of continuous light. In female turkeys given 1L:2D, 5X, no differences in production and broodiness traits were noted when compared to birds given continuous 14L:1D (page 31). -28-
32 Table 1. The Effect of Different Intermittent Lighting Treatments on Days to First Egg, Number of Eggs, Hen Day Percent Production, and Number of Hens Responding During the 126-Day Experimental Periods. Treatment Equivalent Lighting Period 1 Experiment Days to First Egg Mean s.d. Females Number Hen Day % of Eggs Production Mean s.d. Mean s.d. Responding in 126 Days No. % 1L: ID, 1L: ID, 3X 3X - 5 hr 5 hr /47 34/ L; ID, 4X 7 hr 2 55, / L; ID, 1L: ID, 5X 5X 9 hr 9 hr /47 35/ L: ID, 6X 11 hr / L: 2D, 1L: 2D, 3X 3X 7 hr 7 hr /47 29/ L: 2D, 4X 1 hr / L: 2D, 1L: 2D, 5X 5X 13 hr 13 hr /48 47/ L: 2D, 6X 16 hr /47 1 1L: 3D, 1L: 3D, 3X 3X 9 hr 9 hr /45 37/ L: 3D, 4X 13 hr 2 2, /48 1 1L: 3D, 1L: 3D, 5X 5X 17 hr 17 hr /48 47/ L: 3D, 6X 21 hr /47 1 6L:18D 6L:18D 6L:18D 6 hr 6 hr 6 hr , /47 28/48 36/ L:1D 14L:1D 14L:1D 14 hr 14 hr 14 hr , /48 45/46 48/ " Interval from beginning of first light period to end of last light period. -29-
33 Table 2. The Effect of Different Intermittent Lighting Periods on Days to Initial Response and Number of Males Responding in the 126-Day Experimental Periods. Treatment Equivalent Lighting Period 1 Experiment Days to Initial Response Mean s. d. Responding in 126 Days No. % 1L: ID, 1L: ID, 3X 3X 5 5 hr hr /47 31/ L: ID, 4X 7 hr /46 5 1L: 1L: ID, ID, 5X 5X 9 hr 9 hr /47 28/ L: ID, 6X 11 hr / L: 1L: 2D, 2D, 3X 3X 7 hr 7 hr /45 33/ L: 2D, 4X 1 hr / L: 1L: 2D, 2D, 5X 5X hr hr /48 48/ L: 2D, 6X 16 hr /48 1 1L: 1L: 3D, 3D, 3X 3X 9 hr 9 hr /47 34/ L: 3D, 4X 13 hr /46 1 1L: 1L: 3D, 3D, 5X 5X hr hr /46 46/ L: 3D, 6X 21 hr /48 1 6L: 18D 6L: 18D 6L: 18D 6 hr 6 hr 6 hr /46 26/45 21/ L: 1D 14L: 1D 14L: 1D hr hr hr /47 47/47 47/ Interval from beginning of first light period to end of last light period. -3-
34 INTERMITTENT LIGHTING PROGRAM FOR TURKEY HENS K. I. Brown, W. L. Bacon, and K. E. Nestor Department of Poultry Science Current recommendations for lighting turkey breeder hens are to restrict to a 6-hour day at 21 weeks of age, and to light to a 14-hour day at 29 to 32 weeks of age. The age of lighting to a 14-hour day may vary with the strain of turkey. In a recent study by Nestor and Brown (1972, Poultry Sci., 51: ), it was concluded that although no general recommendation for light intensity could be made, large-bodied strains of turkeys did lay more eggs when intensity is increased from 2 foot candles (FC) to 5 FC. Obtaining 5 FC intensity requires a minimum of 1 watt bulbs with reflectors on 1-foot centers 7 feet above the floor. This amount of light requires large amounts of electrical energy and may result in hyperactivity and lower feed efficiency. For that reason, it was decided to study the effects of intermittent light on turkey egg production. Bacon (page 27) has reported the effects of a series of lighting regimens on Coturnix quail. He clearly shows that the length of light day required to obtain maximum egg production is 13 hours or longer. Based on results with quail, it was decided to study the effects of 1 hour light, 2 hours dark, 5 times (1L:2D, 5X) on egg production of large weight turkey hens. Seventy-two large white turkey hens were distributed into six pens with 12 hens per pen. The light treatment for three pens was 14 hours light, 1 hours dark, and for the other three pens was 1L:2D, 5X. For the former group, the light intensity was 5 FC (4.3 to 5.7). For the latter group,.5 FC was provided continuously for 15 hours except for the 1-hour light periods of 5 FC. Egg production data were collected for 112 days beginning with production of the first egg. The days from lighting to production of the first egg were used as a measure of responsiveness to the light treatment. Broodiness was measured by number of broody periods and total days lost as a result of broodiness. A broody period was considered as 5 or more consecutive days of non-production regardless of nesting behavior. Intensity of lay was measured by number of clutches per hen, average number of clutches, and rate of lay. Rate of lay was calculated by dividing total egg production by total days in the period minus total days lost through broodiness. Although the reproductive performance of the intermittently lighted group trended lower than the continuously lighted group, there were no statistical differences in days to first egg, number of eggs per hen, number of clutches, eggs per clutch, or rate of lay (Table 1). There were significantly more broody periods per hen, longer broody periods, and total days broody in the 1L:2D, 5X group, which explains the slightly lower egg production. It is concluded that intermittent light will stimulate egg production in turkeys as well as continuous light* Further studies will be conducted to develop data on the optimum light regimen required to reduce the incidence of broodiness and for optimum egg production, and at the same time to use the least electrical energy. -31-
35 Table 1. Egg Production Traits of Large White Turkeys Exposed to Different Light Treatments. Light Treatment No. Hens Days to First Egg 112 Day Egg Prod, No./ Hen Broody Periods Ave. Length Total Days Broody No./ Hen Clutches Ave. Length Rate of Lay (no.) (no.) (no.) (no.) (days) (no.) (no.) (days) (%) lalrlod L:2D, 5X L at 5 foot candles. 1L, 5X at 5 foot candles with remaining 9L at.5-1. foot candles. -32-
, the authors presented a possible new approach to the control of broodiness of turkey hens.")
36 FURTHER RESEARCH ON MANIPULATION OF THE LIGHTING PROGRAM FOR CONTROL OF BROODINESS IN TURKEYS Karl E. Nestor and Wayne L. Bacon Department of Poultry Science In Turkey Research 197 (Research Summary 47, Dec. 197), the authors presented a possible new approach to the control of broodiness of turkey hens. This consisted of manipulating the normal lighting program of 14 hours per day by providing the laying bird with 1 or more days of continuous light periodically during the laying period. This method was compared with the very effective but timeconsuming method of broodiness control routinely used at the Research Center (Renner and Nestor, OARDC Turkey Research Summary 47, 197). The hens receiving the light treatment did not lay as many eggs as the control group of hens receiving the routine method of broodiness control. There was an indication that fewer hens went broody when exposed to the light treatment. A similar program was repeated on another group of hens. This group of hens was exposed to 48 hours of continuous light every 2 weeks throughout the laying period, beginning 3 weeks after the start of lay. In this experiment, both the control and the light-treated group (with the exception of the egg line) received the routine broody management treatment. Five medium weight and five large-bodied strains were used. The light treatment improved total egg production for a 18-day period for the relatively low-producing medium weight lines (randombred control, low line, and high line). This was accomplished by reducing the total days lost from broodiness and increasing intensity of lay (Table 1). The light treatment seriously reduced egg production in the egg and semen lines. In the egg line, the light treatment reduced the days lost from broodiness but decreased rate of lay. Broodiness was greater and intensity of lay was less in the group of semen line hens given the light treatment. The light treatment had no apparent effect on total egg production of the largebodied randombred control, a subline (16 weeks) of the control selected for increased 16-week body weight, or a subline (index) selected for both increased 16-week body weight and increased egg production. However, the light treatment reduced the total days broody in the randombred control but also reduced intensity of lay, so the net effect on egg production was nil. The light treatment reduced broodiness in a subline (broody) selected for decreased broodiness, resulting in slightly higher egg production. On the other hand, broodiness appeared to be increased by the light treatment in a subline (clutch) selected for increased clutch length. This resulted in lower egg production. Based on averages on all lines, the light treatment had no effect on total egg production, total days lost from broodiness, or intensity of lay. The light treatment characteristically changes the pattern of lay during light exposure and shortly thereafter. When the light treatment was given for 2 consecutive days, an increase in number of eggs laid was apparent on the second day of 24-hour light and on the following day when 14 hours of light were given. This was followed by a day or two of reduced egg production. The effect of the light treatment apparently varied from strain to strain. In some cases, the additional light depressed intensity of lay. Since the light treatment was beneficial in reducing broodiness in some lines, it is possible that if -33-
37 only broody hens were exposed to the additional light, the light treatment might reduce broodiness in all lines. An experiment is planned in which the additional light will be given only to those hens which are on the nests after the lights go out. These hens will receive 24 hours of light while the hens not on the nests will be lighted for only 14 hours per day. -34-
38
39 THE EFFECTS OF ACCIDENTAL PRE-LIGHTING OF TURKEY HENS ON SUBSEQUENT EGG PRODUCTION Karl E. Nestor Department of Poultry Science Flocks of turkey hens are occasionally given stimulatory lighting prior to the desired time as the result of malfunctioning of time clocks or other factors. The question arises as to whether such flocks can be used for economical production of turkey hatching eggs. One group of hens on genetic studies was accidentally lighted from a 9-hour light day to a 24-hour light day because of a time clock malfunction. The accidental lighting occurred when the hens were 3 weeks of age during the week of Nov. 21. It lasted for an undetermined length of time but with a known maximum of 11 days. When it was apparent that the stimulatory lighting had occurred, the light day was dropped to 4 hours. Feed was removed for 3 days and water was removed for 2 days beginning Dec. 6. Many hens came into egg production after the feed, water, and light restriction. Maximum egg production reached 2 percent. However, since the hens were not trapnested, it was not known which hens were laying. When it was apparent that the first restriction of feed and water was not sufficient to cause the hens to go out of production, a second restriction of feed and water was made beginning Dec. 21. This resulted in cessation of production by all hens by Jan. 4, when the hens were approximately 35 weeks of age. The hens were given stimulatory lighting of 14 hours per day on Feb. 1 when they were approximately 39 weeks of age. A similar group of hens which were neither pre-lighted nor restricted on feed and water served as controls. These control hens were restricted to a 4-hour light day prior to lighting for egg production. Hens from the pre-lighted and control groups were intermingled in pens at random. The hens which were accidentally pre-lighted came into production at a faster rate than the controls (Tables 1 and 2) when given 14 hours of light on Feb. 1. This indicates that even though egg production of the pre-lighted hens had ceased approximately 1 month prior to Feb. 1, these hens 1 reproductive systems apparently had not completely regressed, resulting in a quicker response to the stimulatory lighting. This resulted in superior egg production by most of the pre-lighted hens during February. Some of the medium weight pre-lighted hens maintained their superior egg production through April, 'Ihe pre-lighting had a greater effect on low-producing medium weight strains (Randombred Control and High line) than on higher producing strains (Egg strain and Low line) (Table 1). The major effect of pre-lighting occurred primarily after 12 weeks of lay. On the average, medium weight hens which were pre-lighted laid 1.9 fewer eggs per hen for an approximate 26-week (18-day) laying period. After approximately 17 weeks of lay, the difference was 4.7 eggs per hen. The effects of pre-lighting were not as severe on heavy weight hens as on medium weight hens (Table 2). Only hens of the Index and Fertility lines had greatly reduced egg production in the pre-lighted group. Chance sampling may have contributed to the large difference in the Index line, for only eight hens from this line were pre-lighted. The 18-day egg production for the Randombred Control and Clutch lines was greater in the pre-lighted group than in the control group. If the data for the -36-
40
41
42 Index lines are omitted* the control hens laid.7, *9, and 1.4 more eggs per hen than the pre-lighted group during the 84-day, 12-day, and 18-day production periods, respectively. The results obtained in this analysis indicate that pre-1ighting of turkey hens is somewhat detrimental to subsequent egg production. Medium weight hens are more greatly affected than heavy weight hens. The effect is more severe later in the laying period* Thus, economical egg production would be possible even after prelighting if the pre-1ighting was similar to that occurring in this study and if the time the hens were held into production was shortened from the 2-week production period which is commonly used. The pre-lighting may not have been as harmful if a period of more than 1 month elapsed between the pre-lighting and stimulatory lighting for egg production. However, the effect may have been more severe if the hens had been allowed to lay for a longer period of time after pre-lighting. -39-
43 ECONOMIC SIGNIFICANCE OF MYCOPLASMA MELEAGRIDIS INFECTION IN TURKEYS Y. M. Saif and K. E. Nestor Departments of Veterinary Science and Poultry Science Studies on different aspects of Mycoplasma meleagridis (Mm) infection in turkeys have been conducted at this institution and other institutions for the last several years. However, the economic significance of the infection was impossible to assess due to the lack of large flocks free of the Infection. Lately, a Mycoplasma-Free (MF) flock became available at the Nicholas Turkey Breeding Farm, Sonoma, Calif., and a study was initiated In cooperation between the OARDC and the Nicholas organization to assess the economic significance of the infection. Eggs from MF and Mycoplasma-Infected (MI) hens were shipped from California and set in Ohio Agricultural Research and Development Center Incubators. The number of eggs set and the fertility and hatchability data are shown in Table 1. The reasons for the low fertility of the MI eggs are not understood at this time. It is very doubtful that M. meleagridis Infection was entirely responsible for the extremely low fertility, since most commercial flocks are infected but do not experience such low fertility. The hatchability of the MF eggs was significantly better than the MI eggs. Eggs from the MF and MI flocks were incubated in separate incubators and when the poults hatched they were handled in separate rooms. At hatching time, the poults to be raised were sexed, banded, and inoculated with a penicillin-streptomycin preparation. At that time 54 MF and 45 MI poults were posted and the air sacs were examined for lesions and Mycoplasma. No airsac lesions or Mycoplasma were detected in the MF poults. However, nine of the MI poults had airsac lesions, and Mycoplasma was isolated from 12 of these birds. The poults were raised to 8 weeks of age in 12 floor pens on both sides of an aisle. The MF poults were housed on one side of the aisle and the MI poults were on the other side. The sexes from both groups were maintained separately. The air was exhausted towards the MI birds and attendants served the MF poults first. Feed consumed and mortality per pen were recorded. At 8 weeks of age, the birds were bled and weighed, and the trachea of each bird was swabbed to examine for Mycoplasma. Table 1. Fertility and Hatchability of Mycoplasma-Free and Mycoplasma-"Infected Eggs. No. Eggs No. Dead No. % No. % Set Embryos Fertile Fertile Hatched Hatched Mycoplasma Free Mycoplasma Infected -4-
44 The results of serologic testing and Mycoplasma Isolation indicated that none of 345 MF birds had Mycoplasma in the trachea and all the results of the serologic testing were negative. Among the 535 MI poults, 85% had Mycoplasma in the trachea and 7% were positive on serologic testing. These results present further evidence of the rarity of lateral transmission of M. meleagridis when the MF and MI birds are separated. Data of growth, mortality, and feed conversion for the first 8 weeks of age are presented in Table 2, Body weight and feed conversion were highly significantly greater in the MF group than in the MI group. The mortality to 8 weeks of age in the MI group was more than double that in the MF group, but this difference was not statistically significant. At 8 weeks of age, the MF and MI birds were transferred to two separate ranges. Significant changes in the MI birds have been noticed since that time and at the time of writing this report, when the birds were 13 weeks old. Cases of crooked necks, leg weakness, and runts had been on the increase since the birds were placed on range, but no such changes were noticed in the MF birds. The MF birds look homogenous in size, whereas the MI birds look different in size. There was no mortality in the MF birds, but ten MI birds died. The birds will be kept to 2 weeks of age, when they will be weighed and marketed. Data will be collected on condemnations at the packing plant. The preliminary data indicate that M. meleagridis infection is an economically important disease of turkeys. Table 2. Body Weight, Feed Conversion and Mortality to Eight Weeks of Age. No. of Birds at 8 Wks Body Wt. (Ibs) Feed Conversion Ibs Feed/ Ibs Gain % Mortalit; Mycoplasma Free Males Females Average Mycoplasma Infected Males Females Average ** ** Both Groups Males Females ' 73 ** * ^Difference between sexes was significant (P<.5). Difference between treatment or sex was highly significant (P<.1) -41-

45 A NEW VARIANT OF THE TEMPERATURE DIFFERENTIAL PROCEDURE FOR EGG DIPPING Y. M. Saif and K. E. Nestor Departments of Veterinary Science and Poultry Science Two procedures for egg dipping are commonly used at the present time the pressure differential (PD) and temperature differential (TD) procedures. The TO procedure in current use involves heating the eggs in incubators at a temperature of 1 F. for 2 or 3 hours, followed by dipping in a cold antibiotic solution. This procedure is referred to in this report as the air temperature differential procedure (AID). This report describes a variant of the ATI) which involves heating the eggs in water, followed by dipping in a cold antibiotic solution. This new procedure is a modification of a method used by an Ohio commercial hatchery for dipping turkey eggs. The term water temperature differential (WTD) procedure is used to refer to this procedure. An attempt was made to measure the temperature of eggs before heating by both the ATD and WTD procedures. For this purpose, groups of eggs (six each) maintained at different temperatures were dipped in water at different temperatures for different periods of time. Then a small opening was made in each egg, the temperature of the albumin was measured, and then the temperature of the albumin and yolk together was measured. The results are presented in Tables 1 and 2. Since the temperature of the interior of the egg after heating is important in any temperature differential procedure, the WTD procedure which resulted in interior temperature similar to that obtained by the normally used ATD procedure was used for the rest of this study. This procedure involved heating the eggs for 1 minutes at 14 F,, followed by dipping in an antibiotic solution maintained at 45 F. for 15 minutes. Comparisons between this procedure and the ATD and PD procedures also were made. The ATD procedure consisted of heating the eggs in an egg incubator for 3 hours at 99 F., followed by dipping in an antibiotic solution maintained at 45 F. for 15 minutes. For the PD procedure, a pump was used to create vacuum in a closed tank which contained an antibiotic solution in which the eggs were dipped. A negative pressure of 38 cm. Hg. was established and maintained for 5 minutes, and the eggs then remained in the dipping solution for 1 minutes under atmospheric pressure. The antibiotic used for the three procedures was gentamicin sulfate. The amount of antibiotic used for preparing dipping solutions in all trials was 1 p.p.m. After dipping, the amount of antibiotic in the yolk and albumin of a single egg or in groups of eggs was estimated by a biologic procedure. The results of testing for the amount of antibiotic absorbed by eggs dipped by the three dipping procedures are presented in Table 3. These indicate that under the conditions described, the WTD is as efficient a method as the ATD procedure. It was noticed that the amount of antibiotic absorbed by eggs subjected to the three different procedures was always highest with the WTD. When it became obvious that the WTD procedure is of value for egg dipping, an experiment was initiated to study its effect on fertility and hatchability. Four egg treatments were employed as follows: -42-
46 Table 1. Temperature Changes Following Water Heating^. Initial Temp. Incubation of Eggs *F Temp. F One Min/ Post Dipping Temperature ( F) Two Min. 2 Three Min. 2 Five Min. 2 Ten Min. 2 Alb. Min Alb. Mix Alb. Mix Alb. Mix Alb. Mix NT 3 NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT NT Average of temperature of 2 Period of incubation. 3 Not tested * six eggs. Table 2. Temperature Changes Following Air Heating 1. Initial Temp, of Eggs F Incubation and Temp. Time Post-Dipping Temp. ( F) F Mixture of Albumin and of a Yolk 76 2 hrs. at hrs. at hrs. at hrs. at hrs. at Average temperature of six eggs. -43-
47 1. Eggs not dipped. 2. Eggs heated in water for 1 minutes at 14 F. but not dipped. 3. Eggs heated in water for 1 minutes at 14 F., then dipped in water maintained at 45 F. for 15 minutes. 4. hggs heated in water for 1 minutes at 14 F., then dipped in a solution containing 1 p.p.m. gentamicin sulfate maintained at 45 F. for 15 minutes. These four treatments were repeated over a period of 5 weeks, using eggs produced on the day before the dipping. Approximately 3 eggs were used for each treatment. The results presented in Table 4 indicate that the WTD procedure did not influence the hatchability, and that dipping in gentamicin had no effect on fertility or hatchability of fertile eggs. Information presented in this report indicates that the WTD procedure can be a valuable procedure for egg dipping. Table 3, Antibiotic Absorption of Eggs Subjected to Different Treatments. Trial No. Method of Testing No. Eggs Ave. Amount of Antibiotic in Each Absorbed ( Treatment ATD^ FD J 1 Individual Eggs Individual Eggs ml from Each Egg Mixed ^Air temperature differential, ^Liquid temperature differential. Pressure differential. Table 4. Effect of Dipping by the Water Temperature Procedure on Fertility and Hatchability of Turkey Eggs. Differential Treatment % Fertility % Hatchability Eggs not heated or dipped Eggs heated» not dipped Eggs heated, dipped in water Eggs heated, dipped in gentamicin?eggs heated at 14 F for 1 minutes. *~Egg«s dipped at 45 F for 15 minutes S
48 TREATMENT OF TURKEY HATCHING EGGS FOR THE CONTROL OF SALMONELLA INFECTIONS Y. M. Saif Departments of Veterinary Science and Poultry Science In earlier reports, information was presented on the elimination of Arizona and control of Mycoplasma meleagridis infections in turkey eggs by dipping in solutions of the antibiotic gentamicin sulfate. In this report, information is presented on the use of gentamicin for the treatment of experimental Salmonella infections in turkey hatching eggs. Eggs laid over a period of 1 week before each experiment were used in this study. A pressure differential procedure was used to introduce Salmonella organisms into the eggs. Groups of eggs were infected with each of 2 serotypes of Salmonella. After infection, the eggs were maintained at approximately 42 F. for 48 hours. Then the eggs to be treated were incubated for 3 hours at 1 F., followed immediately by dipping in an antibiotic solution maintained at 45 F. for 15 minutes. The anibiotic solution contained 1 p.p.m. gentamicin sulfate dissolved in distilled water. Five trials were conducted employing 2 Salmonella serotypes for egg infection. Approximately 2 eggs were infected with each serotype. Half of each group of eggs were treated and the other half were left as infected untreated controls. The infected treated and infected nontreated eggs were incubated for 1 week at approximately 99 F. Then all eggs were opened (infected treated eggs were opened first) and three areas of each egg were swabbed. The swabs were transferred to appropriate culture media for growing Salmonella. The results of examination of eggs for Salmonella are presented in Table 1. No Salmonella were isolated from 177 eggs infected with 18 different serotypes, and 146 isolations were made from 179 infected nontreated eggs. One isolation was made from each of two eggs infected with S. amsterdam and 5* oranienburg* and 19 isolations were made from 2 eggs infected with both serotypes but not treated. The results indicate that it is possible to control Salmonella infections in turkey hatching eggs. Improvements in techniques might result in elimination of the infection. -45-
49 Table 1. Isolations of Salmonella from Infected-Treated and Infected-Nontreated Turkey Eggs. Trial No. Serotype Salmonella Infected Nontreated Eggs Isolations Infected Treated Eggs 1 S. S. 2 S. S. S. S. S. S. 3 S. S. S. S. 4 S. S. S. S. 5 S. S. S. S. drypool eimsbuettel saint-paul montevideo thompson newport s chwar z engrund binza derby heidelberg anatum typhimurium lutea senftenberg cerro tennessee amsterdam infantis pullorum oranienburg 8/1 1 6/1 7/1 1/1 6/9 9/1 1/1 8/1 5/1 8/1 5/1 8/1 1/1 1/1 1/1 1/1 1/1 9/1 7/1 9/1 /1 /8 /1 /9 /1 /1 /1 /1 /1 /1 /1 /1 /1 /1 /1 /1 1/1 /1 /1 1/1 Numerator «number of eggs examined, denominator = number of eggs with Salmonella. -46-
50 CONTROL OF IN TURKEY Y. M. Saif and K. E. Nestor Departments of Veterinary Science and Poultry Science In previous reports from the Ohio Agricultural Research and Development Center, it was indicated that pre-incubation dipping of turkey hatching eggs in the antibiotic tylosin was helpful in controlling Mycoplasma meleagridis infection. Other reports* (Turkey Research Summaries 197 and 1972) indicate the value of using the antibiotic gentamicin sulfate in eliminating Arizona and Salmonella infections from turkey hatching eggs. Because of the success in eliminating Arizona and Salmonella infections from the eggs, it was decided to study the effects of this drug on M. meleagridis infection. A temperature differential procedure was used for treating the eggs in this study. It consisted of incubating the eggs for 3 hours at 99 F., followed by dipping in a solution containing 1 p. p.m. of the antibiotic gentamicin sulfate for 15 minutes at 45 F. The solution also contained.35 percent of the iodine coinpound losan. A new dipping solution was prepared weekly. Eggs from two flocks of naturally infected hens were used. Eggs produced during a 1-week period were incubated at the same time. Some unhatched eggs, dead-inshell poults, and living poults from each hatch were cultured for M. meleagridis. and examined for air sac lesions. Samples were examined from ten hatches of Flock 1 and four hatches of Flock 2. Eggs used in seven hatches from Flock 1 were dipped, and those from three hatches served as undipped controls. All eggs from Flock 2 were dipped. The results of examination for air sac lesions and M. meleagridis isolations are presented in Tables 1 and 2. As indicated in Table 1, 294 of the poults which hatched from dipped eggs of Flock 1 were examined and only one (.34 percent) had air sac lesions and Mycoplasma. Of 259 dipped eggs, seven (2.7 percent) had Mycoplasma. Of the poults hatched from nondipped eggs, 11 were examined and 26 (23.6 percent) had air sac lesions and 43 (39.1 percent) had Mycoplasma. Ninetynine nondipped eggs were examined and 47 (47.5 percent) were found to have As reported in Table 2, no Mycoplasma was detected in hatched poults or nonhatched eggs and no air sac lesions were detected in the poults from Flock 2, It is interesting to note that Flock 2 had a higher percentage of infected hens than Flock 1. The results indicate that gentamicin was effective in reducing the incidence of the infection to a level where it would be possible to select serologically negative birds for breeding purposes,.47-
51 Table 1. "Mycoplasma Isolations and Air Sac Lesions from Eggs and Poults from Flock 1. Wk Treatment No. Examined No. with Air Sac Lesions Mycoplasma Isolations Not Assoc. with Lesions Assoc, tvlth Lesions Total Percent No. Examined Eggs Mycoplasma Isolation Total Percent 1 Dipped Dipped Dipped Dipped Nondipped Dipped Nondipped Dipped Nondipped Dipped Total Dipped Total Nondipped Table 2.-- Mycoplasma Isolations and Air Sac Lesions from Eggs and Poults from Flock 2. Wk Treatment No. Examined No. with Air Sac Lesions Mycoplasma Isolations Not Assoc. with Lesions Assoc, with Lesions Total Percent No. Examined Eggs Mycoplasma Isolation Total Percent 1 Dipped Dipped Dipped Dipped Total
52 OF TO ELIMINATE Y* M«Saif and K. I. Brown Departments of Veterinary Science and Poultry Science Mycoplasma meleagridis is transmitted from infected turkey males to noninfected hens via the semen. In this report, information is presented on attempts to control semen transmission. This work was done In vitro and In vivo. The In vitro work involved collecting semen from Mycoplasma-infected toms and treating it at room temperature, with the antibiotic gentamicin sulfate for different time periods. Data were collected on the effects of the semen treatment on fertility and hatchability. The semen was cultured before and after treatment for Mycoplasma. The antibiotic was dissolved in a semen diluent, and the semen was mixed with the diluent in a 1:2 ratio. The following drug concentrations were tested: 5, 1, 2, 4, 8, 1, 15, and 2 ug per ml. The only drug concentration which resulted in consistently complete elimination of the organisms was 2 ug per ml. It was discovered that the ph of the diluent is an important factor. A diluent ph of 7,5 is essential for the elimination of the organisms at the 2 ug per ml. level. At ph 7, it was possible to recover the organisms on some occasions, In the second stage of this study, the information gained from the in vitro studies was applied for the following In vivo experiment. A flock of 72 hens housed in six floor pens (12 each) was used. Before the first insemination, all birds were tested serologically for M. meleagridis antibodies and the vagina of each hen was swabbed for Mycoplasma isolation. The results indicated that all birds were free of M. meleagridis infection. Three treatment groups were used in each pen as follows: Treatment 1 Treatment 2 Treatment 3 four hens were inseminated with undiluted semen. four hens were inseminated with semen diluted with untreated diluent in the ratio of 1:2. four hens were inseminated with semen diluted with a diluent containing gentamicin in the ratio of 1:2. The final concentration of the semen-diluent mixture was 2 ug per ml. The diluent used had a ph of 7.5. Semen and semen-diluent mixtures for all three treatments were incubated for 3 minutes at 15 C. before use. The semen came from Mycoplasma-infected males. Inseminations were made weekly with the semen treatments shown above. The dose was,1 ml. for undiluted semen and.3 ml. for diluted semen. Therefore, approximately 7 billion total sperm and 56 billion motile sperm were inseminated each week. Vaginal swabs and blood were collected from each hen on the second, fourth, and seventh weeks of production. The results are presented in Table 1. Although complete elimination of Mycoplasma from the birds in Group 3 was not accomplished, the incidence is low when compared to Groups 1 and 2. The results indicate that semen treatment is of some benefit in controlling M. meleagridis in the semen. Further studies will be conducted to investigate the possibility of complete elimination of the infection from the semen, -49-
53 Eggs were set weekly and candled at 7 days. The eggs removed as infertile ere broken and examined macroscopically for dead germs. The data for the first ive sets are presented in Table 2, It is clear that for the first 5 weeks of production, semen treatments had no ignificant effect on fertility and hatchability. Table 1. Effect of Semen Treatment on Mycoplasma in the Reproductive Tract of Turkey Hens. Mycoplasma Isolation at 2nd Wk Mycoplasma Isolation at 4th Wk Mycoplasma Isolation at 7th Wk Nondiluted Semen 5/ /23 13/23 Diluted Semen 8/24 17/24 18/24 Diluted Antibiotic Treated Semen /24 /24 2/24 Numerator = number of infected hens, denominator = number of tested hens. Table 2. Effect of Semen Treatment on Fertility and Hatchability (5 weeks). No. % % Hatch. of Fertile % Hatch. of Total Treatment Eggs Fertile Eggs Eggs Nondiluted Semen Diluted Semen Diluted Antibiotic Treated Semen

54 THE INHERITANCE AND RELATIONSHIP OF EGG QUALITY AND REPRODUCTION TRAITS IN TURKEYS* Karl E. Nestor and C. R. Weaver Department of Poultry Science and Statistics Laboratory There is limited evidence on the inheritance of egg quality and some reproduction traits and on the relationship among egg quality and reproduction traits of turkeys. If egg quality traits are highly heritable and have a close genetic relationship to hatchability, it would be possible to improve hatchability by selecting for improvement in the egg traits. Estimates of the heritability of, and genetic and environmental covariation among, various reproduction traits were made using data from the first five generations of a line selected for increased egg production. Heritability (h^) is a measure of the total variation of a trait resulting from genetic variation. Genetic and environmental correlations measure the relationship between traits due to genetic and environmental causes, respectively. Generally, correlation coefficients range from -1. to +1,, with the magnitude of the correlation measuring the closeness of the relationship. A negative correlation indicates that as the magnitude of one trait increases, the magnitude of the second trait declines. On the other hand, a positive correlation means that the two traits tend to move up or down in magnitude together. The reproduction traits included in this study were percent fertility, percent hatchability of fertile eggs and of all eggs, number of poults produced per hen, egg production, egg weight, days from stimulatory lighting to production of first egg, number of broody periods per hen, and female body weight at 5 percent production* The hatching data were collected for a 12-week period beginning when the hens first attained 5 percent production. Egg production, egg weight, and number of broody periods per hen were measured for an 84-day period starting with production of the first egg, usually near the end of February each year. The heritability and correlation estimates were based on variation between full sib families and individuals within full sib families within years. The heritability estimates are presented in Table 1. In general, body weight at 5 percent production, egg weight, and hatchability of all eggs have high heritability (greater than.4), whereas all other reproduction traits except hatchability of fertile eggs have moderate heritability (.2 to.4). These results indicate that gains could be made by selective breeding in all reproduction traits measured. Those with higher heritability would be expected to be improved at a faster rate. Egg production was closely positively correlated genetically with fertility, hatchability, and number of poults produced per hen, and negatively correlated with egg weight, days from lighting to first egg, and number of broody periods per hen (Table 2). This indicates that selection for increased egg production would also result in increases in fertility, hatchability, and number of poults per hen. A positive genetic correlation was estimated between percent fertility and percent hatchability of fertile eggs. Fertility, hatchability of fertile eggs, and number of poults per hen were also positively correlated environmentally, but the *Supported in part by grants from the National Turkey Federation, Mount Morris, 111.
55 Table l.~heritablhty Estimates (h 2 ) and Their Standard Errors Obtained from Full Sib Analysis for Reproductive Traits in the Egg Line, Trait h 2 +Standard Error 84 Day Egg Production Days from Lighting to First Egg Female Body Wt at 5% Production Egg Weight Number of Broody Periods % Fertility % Hatchability Fertile Eggs % Hatchability All Eggs Number Poults per Hen for 8 Wks Table 2. Genetic Correlation Estimates (r fl ) and Their Standard Errors Obtained From Full Sib Analysis Among Reproductive Traits. Egg Hatch. No. Poults Production Fertility F.E. 1 Per Hen *;Q.L 91. Jy^* LJL qa+ in..7 *JT^. J.W 11*.% HhnVt I? IT 1 narcn» r. Ct * -^i* -^ QA4- SS. " H-T * J,J Hatch. A.E. 2 T> No. Poults Per Hen AS4-1 &. O.JT"* JLO QO r, -1 A * y OT^ -LU Egg Weight Days to First Egg^ No. Broody Periods Body Wt. at 5% Prod "^Hatchability of fertile eggs, ^Hatchability of all eggs. %ays required from stimulatory lighting (14 hours per day) to production of first egg. ^Standard error could not be calculated because estimate was greater than
56 A hierarchial mating design was used in a randombred control population of large weight white turkeys and in a medium weight strain selected for increased egg production to obtain heritability of, and genetic correlation estimates between, egg quality and reproduction traits. Genetic estimates were based on the sire + dam components of variance and covariance. Only egg quality measurements which had previously been shown to exhibit changes during the laying periods were included in the analysis. The egg quality traits included egg weight, yolk weight, percent yolk, percent dry matter in yolk, shell weight, percent shell, weight loss of eggs after 7 days of incubation, and percent incubation weight loss. The heritability estimate of egg weight was higher in March and April than in June (Table 5). The estimates in March and June were similar to the one based on data collected over an 8 to 12-week laying season beginning in March (Table Ij. The heritability of yolk weight also apparently declined with age of bird or length of Table 3. Heritability Estimates Obtained from Sire Plus Dam Components of Variance for Egg Quality and Reproductive Traits, + Standard Error Trait March April June Egg Weight Yolk Weight % Yolk % Dry Matter in Yolk Shell Weight % Shell Incubation Wt Loss % Incubation Wt Loss Wks -22 Wks % Fertility % Hatch. Fertile Eggs _.36 No. Poults Per Hen Egg Production, 22 Wks magnitude of the correlations was not as great as the corresponding genetic correlations. This means that environments which increase one reproduction trait tend to improve the other traits, but the effect is not as great as genetic factors. Egg production was strongly correlated on an environmental basis with only number of poults produced per hen. -53-
57 the laying season. However,, when adjusted for egg weight (percent yolk), the decline was not apparent. Percent dry matter in the yolk was highly heritable in March and April, but an abnormally low estimate was obtained in June. Shell weight and incubation weight loss expressed as percent of total weight were highly heritable in June, which indicates that genetic variation is evident late in the laying season. Ileritability estimates of fertility and hatchability of fertile eggs for an 8-week period (Table 3) were not greatly different than corresponding estimates made earlier (Table 1). Ileritability of fertility noticeably increased when a longer hatching period was observed* Herltability of egg production for a 22-week period of lay was higher than the estimate for a 12-week period of lay (.45 vs*,32). Egg weight in March, April, and June exhibited a consistent negative genetic association with percent fertility, number of poults produced per hen, and egg Table 4.--Genetic Correlation Estimates and Their Standard Errors^ Between Egg Measurements In March, April and June and Reproductive Traits for a 22-Week Period. Fertility % Hatch. Fertile Eggs No. Poults Egg Prod..Egg Weight March -.654;.29,31+, April June Unweighted Av % Shell March April June Unweighted Av. -.3, % Incubation Wt Loss March April June ~.37+> Unweighted Av % Yolk March April June * Unweighted Av..8.21,28.2 % Dry Matter Yolk March April June Unweighted Av Obtained from sire + dam components of covariance. -54-
58 production (Table 4). The genetic correlation estimates between percent shell and percent hatchability were positive and increased from the March to June measurements. The estimate between the percent shell of eggs laid in June and number of poults was positive. A negative genetic relationship was observed between egg production and percent shell later in the laying period (April and June), Percent incubation weight loss and hatchability were negatively correlated. April and June incubation weight losses were negatively correlated with number of poults produced. Percent yolk in March was closely related to hatchability and number of poults over the entire 22- week reproduction season. Percent dry matter in the yolk in March showed a large negative relationship with hatchability and number of poults. The heritability and genetic correlation estimates observed, although variable, indicate that genetic gains could be made in shell traits late in the laying season, and these gains should result in improved hatchability. Selection for increased percent yolk early in the laying season should also result in increased poult production throughout the entire season, but reduced egg weight might be expected. However, selection for increased amounts of shell (percent shell) would probably reduce egg production. It is very unlikely that selection for improvement in egg traits would be more efficient than selecting directly for egg production, fertility, hatchability, or number of poults, or a combination of these, because the heritability of these traits is moderate. -55-
59 EXPLORING THE POSSIBLE VALUE OF COLOR PATTERN GENES IN MODERN TURKEYS Karl E. Nestor Department of Poultry Science The turkey of today was developed from the North American wild turkey» The wild turkey still exists in the forests of several states,, including Ohio, Pennsylvania, and West Virginia. After domestication, turkeys were raised for a period of time primarily for their beauty. Then meat production became important and the bronze turkey emerged as a superior variety. Consumers began complaining about the colored pinfeathers and so the white turkey became the most popular. Many of the most beautiful turkey varieties of the past are almost extinct at the present time The question arose, do the genes controlling color patterns in turkeys have any influence on egg production, growth rate, semen yield, livability, or other traits of economic interest? Some preliminary evidence indicated that one of these genes (black-winged bronze) increased livability when present in the genetic background of a population of white turkeys. The black-winged bronze genes can be identified at hatching in white poults. Poults homozygous for this gene have a much lighter head and body color than poults carrying the bronze gene. In 1968, several of these white poults were observed at hatching. The number of white poults was accurately recorded in 1969, Two randombred control populations from the same source had similar gene frequencies (1,12 and 1.67%) with respect to this gene. A line established from the randombred control by selection for increased semen yield had a higher gene frequency (2,83%), This indicated that perhaps the black-winged bronze gene was increasing semen yield, thus being favored in the semen line. A preliminary comparison was made within families between individuals with normal and white down color for some economic traits. The data presented in Table 1 indicate that the black-winged bronze gene (white down color) actually reduced semen yield slightly, with little effect on sperm concentration. The frequency of abnormal sperm may have been increased slightly b> tho black-winged bronze gene. If the black-winged bronze gene reduced semen yield, why did this gene spread in a population selected entirely for increased semen yield" It appears that the major effect of the black-winged gene may have been in reducing mortality (Table 1). The above evidence, along with some data obtained by Dr. t. Buss at Pennsylvania State University which indicates that some of the color pattern genes are beneficial for growth, led Ohio researchers to take a giant step backward. Samples of all known color pattern genes in turkeys were obtained. These included bourbon red, narragansett, black, gray, royal palm, dark brown, light brown, black-winged bronze, slate, spotted, and bronze. These colored turkeys were crossed with large white turkeys to transfer the plumage color genes into a growth background similar to the modern commercial white turkeys. Special matings will be made to see if these genes have any beneficial effects on various traits of commercial interest. If the results indicate that these are favorable effects, this research may then lead to a giant step forward. Pure lines of these colored turkeys are being maintained separately to conserve the gene pools for future use. Based upon the number of breeders listed in Turkey World, the number of primary turkey breeders has dropped from 17 in 1967 to -56-
60 Table 1. Effect of Gene of Economic Traits, Down Color Normal White % Mortality Semen Yield (cc) Sperm Cone, (bill./cc) Bent Sperm (%) Male Body Wt. (Ib.) 8 Wks. 36 Wks. 24 Wks. Female Body Wt. (Ib.) 8 Wks. 16 Wks. 24 Wks C in Such a decline in number of primary breeders probably results in loss of useful genes which will never be recovered. Maintenance of the pure lines of colored turkeys will permit a source ol perhaps some of these genes for future utilization. The presence of these colored varieties, as well as the wild turkeys, allows the Poultry Science Department to show the historical development of the turkey. Such exhibits have received considerable interest. High school students are an especially important audience* This very interesting manner of showing the dynamic nature of the turkey industry may be instrumental in attracting more of these students to select Poultry Science as their major field in college. -57-
 may be more efficient in feed utilization and have improved growth rates than males grown under normaj light intensities of about 1 to 5 FC, However, late in the growing period")
61 LOW INTFMiir,' HfsHT OF AND ^ALH: L. Bacon and P. Touchburn Department of Poultry Science It has previously been reported that turkey males grown under a low light intensity of aliout.1 foot candles (FC) may be more efficient in feed utilization and have improved growth rates than males grown under normaj light intensities of about 1 to 5 FC, However, late in the growing period the reverse was offrn noted (Tourhhurn et al., 197. OARDC Research Summary 47). These data were not conclusive and therefore further studies were conducted. In Experiments 1 and 2, light intensities of 3, 1,.1, and,1 FC were used from 3 weeks of age to market age of 22 or 24 weeks. The diets fed were composed of practical ingredients in the relative quantities described by Naber and Touchburn (197, Ohio Cooperative Extension Service, Bull. 343), Fn these diets, protein and energy levels are adjusted according to the age of the birds. The birds were of two different Large strains and were started under' 24 hours of light and 1 FC intensity, M 1 week they were given 12 hours of light* per day, and at 3 weeks they were exposed to the experimental light intensities of S, 1,.1, or,1 HJ until the end of the experiment. The effects of age and light intensity on body weight for these experiments are given in Table 1. It is clear from these data that the birds reared under.1 FC were heavier up to 16 weeks of age (Experiments 1 and 2) and that from this time on they were not as heavy as those in the 3, 1, or,1 FC treatment groups. Interval weight gains (Table 2) more clearly demonstrate when the change in growth rate associated with light intensity occurs. The critical time was the 12 to 14 week interval in Experiment 1 and 1 to 12 week interval in Experiment 2. From this time on, the birds under the 3 and 1 FC treatments grew the most rapidly. Feed utilization efficiency (Table 3) was better in the.1 FC group than in the 3 and 1 FC groups in Experiment 1 up to 16 weeks of age, with no difference in the older birds. In Experiment 2, no difference was noted in the entire experiment* However, interval feed utilization efficiencies reflected interval growth rates (Table 4). In Experiment 1, those birds under.1 FC had the best feed utilization efficiencies up to 1 weeks of age,, but the poorest from 18 to 22 weeks of age. In Experiment 2, those under.1 FC did not show better feed utilization efficiencies at any stage of the experiment, but had the poorest feed utilization efficiencies from 12 to 22 weeks of age. From the data in Tables 1-4, it appears that superior growth rates were obtained under low intensity light up to at least 12 weeks of age, arid that there is a parallel effect of feed utilization efficiency. However, in the case of feed utilization efficiency, the effect is not as clear in Experiment 2 as in Experiment I. It is also apparent that after 14 to 16 weeks of age both interval growth rate and feed utilization efficiencies favor the higher light intensities. Experiment 3, utilizing two slightly different strains of large type male turkeys in each pen, was conducted to study the effects of changing from 1 FC to.1 FC and from.1 FC to 1 FC at 12 weeks of age. Otherwise this experiment was similar to Experiments 1 and 2, All birds were fed diets as outlined by Naber and Touchburn (197). Light treatment A was 1 FC from 3 to 22 weeks of age. Light -58-
62 Table 1, The Effect of Light Intensity on Body Weight (Ibs) in Experiments 1 and 2. Table 2. The Effect of Light Intensity on Interval Weight Gain (Ibs) in Experiments 1 and 2. Age 3 FC 1 FC Light Intensity.1 FC.1 FC Mean Wt. P ISD.1 or.5 1 Period 3 FC 1 FC Light Intensity.1 FC.1 FC Mean Wt. P LSD.1 or.5 1 Experiment I Experiment 1 4 wks * > < > < < < < < < < > <, < ^ wks O , >.5.99 < > < < < < < > < < < < Experiment 2 Experiment It IB , <.l 3.1 < < < < <.l < > > >, > > *> lb ^ < < < < < <.C < <, < < < > If P.<5, LSD at P *.5; if P<.1, LSD at P *.1. Ditrcrance from mean at e c ch age. If P<.5, LSD at P =.5; if P<.1, LSD "Tifference from mean at each age. at P *.1.
63 Table 3,--The Effect of Light Intensity on Gain/ Feed Accumulated from 3 Weeks of Age to Various Ages in Experiments 1 and 2. Table 4. The Effect of Light Intensity on Feed for Each Period in Experiments 1 and 2, Age 3 FC 1 FC Light *.1 FC Intensity.1 FC P LSD.1, or.5 Light Intensity Period 3 FC 1 FC.1 FC.1 FC P LSD.1 or,5 J O I 4 wks , , Experiment JL.584,562 * , , ,292 Experiment, , ,33^,329 2,69.66, , , , , ,562, ,324 >.5 >.5 >,5 < e 5,59 <.1.57 <.1.34 <,1.19 <.1,16 <,5,15 >,5 >,5 >.5 >,5 <<5,38 >.5 >,5 >,5 >,5 >,5 >,5 >.5 >,5 >.5 >.5 >*5 If P<*5» LSD at P «.5j if P<.1, LSD at P = wks , , ,23, , , Experiment ,312, , Experiment JL.627, , ,65, ,32.33,23,24, , , , >,5 <,5.67 >,5 <«1.38 >.5 <,5,42 >,5 >,5 >.5 >,5 <.1 <.1 <.1 *43 >,5 >.5 >,5 >.5 >,5 <.5.36 <.1.24 <.1,329 >.5 <«5.4S >,5 <,5,C47 L If P<,G5 f LSD at P =,5; if P<,1, LSD at? =.1.
64 treatment B was 1 FC from 3 weeks to 12 weeks and then.1 FC until 22 weeks of age. Light treatment C was.1 FC from 3 weeks to 12 weeks and then 1 FC from 12 weeks to 22 weeks. Light treatment D was.1 FC from 3 to 22 weeks. It was believed, based on the data in Tables 1-4, that the superior body weights and feed utilization efficiencies would be in treatment group C. Tables 5-8 give the results of this experiment. Body weights (Table 5) and interval weight gains (Table 6) are clearly superior in the birds reared to 12 weeks of age under.1 FC in comparison to those reared to this age under 1 FC. In feed utilization efficiency, there was an early effect of low intensity light on improving feed utilization efficiency (Tables 7 and 8), but this was later lost and was then not clearly in favor of any light treatment throughout the remainder of the experiment. Body weight and interval weight gains were clearly not benefited by switching birds from one light intensity to another at 12 weeks of age. It was believed that those in group C would be heaviest at 22 weeks of age and those in group B the lightest. The data show that while group B was indeed the lightest and significantly lighter than group A, group C was next to lightest and lighter than those grown entirely under.1 FC of light. Significant differences in the growth patterns of the two strains are shown by the mean values for body weight and interval gain (Tables 5 and 6, respectively). Table 5. The Effect of Light Intensity, Age and Strain on Body weight (Ibs) in Experiment 3. Age A Light Treatment" 1 " B C 1 P P LSD.1 or.5 2 Mean Body Weight Strain wks <.5 <.5 <.5 <.5 <.5 <.1 <.5 >.5 >.5 >.5 < ** 3.92** 7.4** 1.59** * 25.49* 26.72* 28.16* X A» 1 FC from 3 to 22 wks; B «1 FC from 3 to 12 wks,.1 FC from 12 to 22 wks; C».1 FC from 3 to 12 wks, 1 FC from 12 to 22 wks; and D».1 FC from 3 to 22 wks. 2 If P<.1, LSD».1; if P<.5, LSD -.5. **Strain 1 significantly different from Strain 2 P<.1. *Strain 1 significantly different from Strain 2 P<.5. ^Difference from over-all mean at each age.
65 Table 6. The Effect of Light Intensity, Age and Strain on Interval Weight Gain in Experiment 3. Period A Light Treatment" 1 " B C 1 D P LSD.1 or.5 2 Mean Body Weight Strain wks <.1 <.5 <.5 <.5 >.5 >.5 >.5 >.5 <.5 <.5 < ** 1.77** 3.11* ** ** L A = 1 FC from 3 to 22 wks; B = i FC from 3 to 12 wks,.1 FC from 12 to 22 wks; C =.1 FC from 3 to 12 wks, 1 FC from 12 to 22 wks; and D =.1 FC 9from 3 to 22 wks. "If P<.1, LSD =.1; if P<.5, LSD =.5. *Strain 1 significantly different from Strain 2 P<.1. Strain 1 significantly different from Strain 2 P<.5. Difference from over-all mean at each age. Table 7. The Effect of Light Treatment and Age on Gain/Feed Accumulated from 3 Weeks of Age to Various Ages in Experiment 3. Age A Light Treatment 1 B C D P LSD.1 or wks <.1 <.5 >.5 >.5 >.5 <.5 <.5 >.5 >.5 >.5 > A «1 FC from 3 to 22 wks; B * 1 FC from 3 to 12 wks,.1 FC from 12 to 22 wks; C».1 FC from 3 to 12 wks, 1 FC from 12 to 22 wks; and D =.1 FC from 3 to 22 wks. If P<.1» LSD -.1; if P<.5, LSD «
66 Table 8. The Effect of Light Treatment and Age on Interval Gain/Feed in Experiment 3. Period A Light Treatment 1 B C D P LSD.1 or wks <.1 >.5 >.5 >.5 <.1 <.1 >.5 >.5 >.5 >.5 > X A «1 FC from 3 to 22 wks; B» 1 FC from 3 to 12 wks*.1 FC from 12 to 22 wks; C «.1 FC from 3 to 12 wks, 1 FC from 12 to 22 wks; and D».1 FC from 3 to 22 wks. 2 If P<.1, LSD =.1; if P<.5, LSD».5. Strain 1 grew faster than Strain 2 during early growth but Strain 2 became the fastei growing strain later in the experiment. There was no interaction at any age. Strair effects were not available for feed utilization efficiencies, as all pens contained both strains. In general, based on these data, it is possible to recommend growing male turkeys for the turkey broiler market under.1 FC if they are marketed before 14 weeks of age. However, growing male turkeys beyond this age on.1 FC of light appears to be of questionable commercial value. Also, switching from.1 FC of light to 1. FC of light at 12 weeks of age is of no benefit, while switching from 1 FC to.1 FC at 12 weeks of age is clearly detrimental to grewth rate and feed utilization efficiency. -63-
67 ENERGY IN TURKEY GROWING DIETS S. P. Touchburn Department of Poultry Science Toward the later stages of growth, turkeys eat tremendous amounts of feed and the energy in these rations contributes 6 percent or more to the total cost of the feed. Calorie intake, through its influence on appetite, is the major factor controlling the intake of all nutrients in a complete mixed ration. Thus the contents of other critical nutrients (protein, vitamins, minerals) must be considered in relation to energy content. These relationships influence not only growth rate but also body composition. Seasonal temperature differences are also recognized as having effects on growth and feed utilization efficiency. Another variable, nutrient density or nutrient concentration, may further affect performance and definitely affects the total volume of feed ingredients and feces that must be handled. The trend is to reduce the fiber and increase the fat contents of feeds. Research at the OARDC has shown that turkeys perform well over a wide range of nutrient density provided a reasonable calorie-protein ratio is maintained. Studies of nutrient density inevitably lead to the use of added dietary fats, since they represent the most concentrated sources of calories. In the early experiments, levels of 2 percent of animal fat (yellow grease) or calorically equivalent levels of soybean oil or a hydrolyzed animal-vegetable fat product were well utilized by growing turkeys. Lxtra-Caloric Benefit of Fat An experiment was designed to study the influence of calorie-protein ratio on the effect of fat at 4-week intervals to 24 weeks of age in growing turkeys. Five replicates of 13 Large White male turkeys each were fed the experimental rations shown in Table 1. Low, intermediate, and high energy rations were formulated with and without added animal fat. The fat provided 25 percent of the dietary calories and was added at levels of 8 to 1.4 percent, depending on the age of the birds. The average weight gain and feed conversion data are given in Table 2. The highest energy ration without supplemental fat (Ration C) could not be formulated after the -4 week age interval using practical ingredients. I he intermediate energy ration without added fat (Ration B) tended to depress weight gains for the entire experiment, indicating that the calorie-protein ratio was too wide. In contrast, these same higher energy rations but containing added fat (Rations H and F) tended to increase weight gains and feed efficiencies. This effect became significant after 16 weeks of age. In Table 3, the efficiencies of feed utilization are shown expressed as a percent in relation to Ration A. Dietary fats exerted a beneficial effect on efficiency, especially at the intermediate and high energy levels (Rations E and F, respectively). Thus, a calorie-protein ratio which was excessive and tended to retard growth in a ration without added fat was rendered adequate by incorporation of fat. The high fat rations probably permitted a slight over-consumption of calories on the wide calorie-protein ratio diets. This was evidenced by an improvement in carcass fat as scored by visual evaluation of the carcasses at 24 weeks of age (Table 4). -64-
68 Table 1.--Design of Rations, Experiment 1 Age, Wks Protein % Low ME/P Calorie Level Intermed. ME/P High ME/P No Added Fat A B C Added Fat (8-1%) D E F Table 2.--Average Weight Gains and Feed Conversions, Experiment 1. (16-24 weeks age only.) Age, Wk: Ration 16-2 Gain F/G 2-24 Gain F/G 4-24 Gain F/G A B ,,* * * D E ** 5.1** 5.8** ** F ** 5.2** 5.7** ** Significant at 5% level of probability. **Significant at 1% level of probability. -65-
69 In another study, Wrolstad Small White turkeys were fed experimental diets from 5 to 14-1/2 weeks of age. Treatments involved narrow and wide calorie-protein ratios with and without supplemental fat to provide 25 percent of the diet calories. Each of the four diets was fed to three lots of males and three lots of females containing 15 birds each. The average weight gains are shown in Table 5. While growth rates were unaffected by the calorie-protein ratio, the addition of fat to the ration significantly increased the growth of the males. The efficiency of feed utilization (Table 6) was significantly better for the wide calorie-protein ratio diets. Supplemental fats, however, caused a further increase in efficiency. From the results of these two experiments, it appears that the beneficial effect of fat is independent of calorieprotein ratio. Table 3.--Efficiency of Feed Utilization Expressed as Percent of Predicted Efficiency Compared to the Unsupplemented Basal, Ration A. Experiment 1. Table 4.--Carcass Finish Score at 24 Weeks of Age. Experiment 1. Ration Ave.~ Age, Wk: Ration A 2.45 A B D E B D E F F Ave. of 4 independent observations on 1 samples each. Scale 1 to 4 with 4 representing maximum fat deposition. Table 5.--Effect of Supplementary Fat and Calorie- Protein Ratio on Weight Gains. Experiment 2. Calorie-Protein Ratio Added Fat Ave. Wt Gain Males, Ib /2 Wks Females, Ib Narrow Wide Narrow + Wide & a 9.97b 9.83b 6.9c 6.82c 6.81c 6.76c ^Figures sharing the same letter do not differ significantly. -66-
70 In the latter experiment, this effect of fat on efficiency became less apparent in the final interval, 12 to 14-1/2 weeks of age, but the explanation is provided by observing in Table 7 the significant improvement in carcass score which resulted from consumption of the ration containing supplemental fat* In other words, after 12 weeks of age the increased metabolic efficiency due to dietary fat still remained but was expressed as an increased body fat deposition rather than an improvement in feed efficiency. The explanation proposed for the greater metabolic efficiency of fat is that there is a saving in the energy needed to synthesize tissue fat from carbohydrate. Tissue deposition of dietary fat generates less heat than would the breaking down of carbohydrate and manufacturing of body fat. This lowered heat production, called Table 6.--Effect of Supplementary Fat and Calorie-Protein Ratio on Efficiency of Feed Utilization Expressed as Gain per Unit Feed. Experiment 2. Ration C/P Added Fat 5-8 Wks Males 8-12 Wks 12-14% Wks 5-8 Wks Females 8-12 Wks 12-14% Wks 1 Narrow Wide.496**.388**.285**.472**.35** Narrow Wide +.527**.413**.279*.479**.365**.242*' *Significant at 5% level of probability. ^^Significant at 1% level of probability. Table 7.--Effect of Supplementary Fat and Calorie-Protein Ratio on Carcass Finish Score at 14% Weeks of Age. Experiment 2. Added Ration Fat Males Females M & F Ave. 1 Narrow Wide Narrow ** 2.68* 2.68** 4 Wide * 2.75** 2.67** Significant at 5% level of probability. **Signi icant at 1% level of probability. -67-
 have confirmed the observation of an extra-caloric effect of fat for growing turkeys.")
71 Specific Dynamic Action, associated with the handling of fat by the bird could be important in reducing heat stress during hot weather. Jensen et al. (197, Poultry Sci., 49: 1967) have confirmed the observation of an extra-caloric effect of fat for growing turkeys. They estimate the fat to provide 32 percent more calories than the assigned metabolizable energy values. It is probable, however, that the degree to which metabolic efficiency is enhanced by dietary fat will vary depending on the age and the extent to which an animal is depositing fat. Obviously the extra-caloric effect will be greater during the later stages of growth. T emp erature Programme d Feeding Several feeding trials have indicated the value of temperature-programmed feeding for growing turkeys. In one such study, Large White torn poults which hatched November 17 were brooded conventionally except that after 5 weeks of age the temperature in the brooder house was gradually reduced to 4 F. by the time the birds were 8 weeks of age on January 6. Half of them were then distributed into six pens in half of an insulated building which was heated by radiant electric brooders laised to a height of 4 feet. The others were distributed into six pens in the other half of the house and subjected to cold conditions near ambient temperatures by thermostatic control of the exhaust fans. Temperatures in the pens were recorded continuously. The weekly mean differential between the warm and cold halves of the house ranged from 6 to 11 F. until \pril 12 when the birds were 21 weeks old. ihen warmer outside temperatures brought mean inside temperatures to about 6 K in both halves of the house, with daytime highs in the 7 f s. The regular six-ration schedule (Ohio Poultry Rations, 197. Ohio Cooperative Extension Service, Bull, 343) was fed to triplicate pens of 3 birds each in each half of the house. In the cold half, triplicate groups also received the same feeds except that they were advanced by 2 weeks from 8 weeks of age on. They thus followed a higher energy feeding program. In the warm half of the house, the opposite was true. To accommodate the reduced need for calories, the schedule was delayed by 2 weeks after 8 weeks of age. These birds thus followed a lower energy, higher protein feeding schedule, Table 8 shows the average weight gains, feed conversions, and feed costs per pound of gain. The feed costs were those charged by the Research Center feed mill and include charges for ingredients, mixing, pelleting, and overhead. These costs provide an important means of comparing the treatments. The effects of cooler environmental temperatures on growth are shown by the greater gains in the cold half of the house. The poorest feed conversion was obtained in the cold house on the regular feeding schedule. By advancing this schedule to provide higher energy-low protein feeds at earlier ages, it was possible to obtain equally good, if not better, rates of gain. Furthermore, feed conversion was also greatly improved on this advanced schedule and, since it involved more of the cheaper feeds, the feed costs per pound of gain were likewise improved. These two items were in fact almost identical to those of the birds on the regular schedule in the warm half of the house. However, no charges have been assessed for heating the warm half of the house. The fact that the birds fed the higher energy feeds in the cold house grew at least as well as those on the regular feeds shows that the latter were greatly over-consuming protein and other critical nutrients as they ate to satisfy the need for calories to maintain body temperature. -68-
72 Table 8.--Effect of Temperature Programmed Feeds on Large White Toms. Experiment 3. Warm House Cold House Feed Schedule Regular Delayed Regular Advanced -24 Weeks Gain , Feed/Gain , Cost/lb Gain, c Weeks Gain Feed/Gain Cost/lb Gain, ^Based on feed costs quoted by O.A.R.D.C. feed mill; include charges for ingredients, mixing, pelleting and overhead. The lack of response to the delayed (higher protein) feeding schedule in the warm environment was surprising. It was assumed that the higher protein diets would compensate for a lowered feed consumption in the warmer environment. However, Waldroup (1972, Proceedings, Arkansas Formula Feed Conf.) has observed a rise in the blood serum levels of those amino acids present in the diet in excess of the needs of growing chicks, \ppetite appears to be depressed by such a rise in serum amino acid content. Since practical poultry diets naturally contain excesses of certain amino acids, it is possible that the higher protein (delayed) feeding schedule in the warm environ ment actually contributed to a reduced intake. The lack of improvement in either growth or feed conversion, along with the increased cost of this feeding schedule, resulted in an increased cost of feed per pound of gain. This explanation suggests the need for research on the amino acid requirements of growing turkeys. With the above exception, the values for formulating feeds to meet the different energy needs according to season or type of housing are apparent. Summary Energy contributes the greatest cost to a poultry feed formula. The energy level of a complete poultry feed controls total intake because birds eat mainly to satisfy their energy needs. Manipulation of the calorie-protein ratio provides a means of altering the carcass composition. The trend toward increased nutrient density in poultry diets has led to greater use of fats because fats represent a concentrated source of energy. In the research reported here, turkeys were observed to grow well and utilize their feed efficiently over a wide range of protein and energy levels, provided the calorie-protein ratios were maintained within reasonable limits appropriate for a given age of bird. -69-
73 Growing turkeys were reported to be capable of effectively utilizing animal fat, vegetable oils, and mixed fats as supplements to their diet, even at levels of supplemental fat exceeding practical limits. Supplemental dietary fats were seen to improve growth rates. This phenomenon remains unexplained. The greater than expected caloric values of the diets containing fat which were observed were labeled the extra-caloric benefit of fat. This benefit was explained on the basis of the reduced heat increment or Specific Dynamic Action associated with the absorption and handling of fats in the animal body. Dietary fats increased carcass fat deposition, particularly during the later stages of growth. Environmental temperature was shown to have a direct bearing on feed intake. Temperature programmed feeding tended to improve feed utilization but, most importantly, reduced the feed cost of turkey production. The importance of amino acid balance in the diet and its effect on appetite may also be important considerations, particularly in a warm environment. This study emphasizes the important relationship among environmental temperature, energy content of the feed, age of bird, and carcass composition. -7-

74 GROWTH RESTRICTION OF LARGE TYPE TURKEY BREEDERS S. P. Touchburn and K. I. Brown Department of Poultry Science The restriction of feed intake during the growing period of chickens has been widely investigated. Although the results in the literature vary widely, there appears to be a tendency toward beneficial effects on the subsequent reproductive performance, particularly with the heavy broiler breeders. These advantages generally include reduction in body weight with a concomitant improvement in feed utilization efficiency, delay in sexual maturity resulting in increased initial egg size, increased rate of egg production, and improved livability. Very little research of this type has been reported for turkeys. Scott and Payne (1941, Poultry Sci., 2: ) limited the feed consumption of Broad Breasted Bronze turkeys in two experiments to 61 or 78 percent of ad libitum during a holding period from 26 to 42 or 45 weeks of age. The only effect on subsequent reproduction was a significant depression of hatchability by the more severe restriction. Anderson et al* (1963, Poultry Sci., 42: 8-15) restricted large type turkey pullets to 84 percent of full feed from 12 to 24 weeks of age and observed improvements in fertility and hatchability. Continuing the restriction to 4 weeks of age reduced the response. Those birds which were not returned to full feeding until just prior to egg production grossly over-ate during the first month of the breeding period. This was accompanied by a marked increase in weight and reduced hatchability. Touchburn et al. (1968, Poultry Sci., 47: ) conducted three trials with a medium sized strain of turkeys, restricting feed intake from 12 or 18 to 28 weeks of age and from 12 to 24 weeks of age. Restriction of intake and of growth rate were accomplished by manually limiting consumption to 7 or 8 percent of full feed 3 feeding a high fiber diet, or feeding a diet containing a compound known to be distasteful to the turkey. Rates of production were significantly lowered by restriction regardless of the method by which it was accomplished. More recently, Voitle and Harms (1972, Poultry Sci., 51: ) at the University of Florida reported initial results of restricted feeding the Marston straii of turkeys from 1 to 3 weeks of age. The control birds were fed a 21 percent protein diet from 1 to 24 weeks of age, followed by a 16.6 percent protein breeder diet. Treatment 2 birds were fed a 1.2 percent protein diet from 1 to 3 weeks. Treatment 3 were fed the 21 percent diet from 1 to 3 weeks of age on alternate days only, being given 1.5 times the amount consumed by the full-fed group. This treatment should have approximated restriction to 75 percent of the consumption of the full-fed group. By 26 weeks of age, these restrictions had limited the weights to 82 and 67 percent, respectively, of the body weight of control birds. Under conditions of natural lighting in springtime, the growth-restricting Treatments 2 and 3 significantly retarded the onset of egg production by 41 and 62 days, respectively. Further data on reproductive performance are not yet reported. Summers et al. (1969, Poultry Sci., 48: ), based on studies with broiler females, suggested that restriction during the early phases of growth might be more effective than restriction later in the growing period. -71-

75 Among turkey industry people, the comment is frequently heard that breeder turkeys should be restricted to prevent excessive fatness and to enhance reproductive performance. The current study was designed to ascertain the effect of restriction in today's Large White breeder turkey, Experimental Procedure Poults were hatched from eggs of the foundation breeding stock provided by a large foundation breeder. The poults used were Large White male line toms and Large White female line hens. The rations fed were the six-ration schedule from day-old to time of lighting for reproduction (26 weeks in males and 3 weeks in females), then the regular torn or hen breeder rations as outlined in Ohio Poultry Rations (197, Ohio Cooperative Extension Service, Bull, 343). After 3 weeks of age, one pen of each sex was fed ad libitum, the second and third treatments being attempts to limit growth to 8 and 9 percent of normal by daily weighing in the appropriate quantity of feed based on the weekly consumption of the respective full-fed birds. At 2 weeks of age, the birds were split into two pens per treatment. The daily lighting schedule was as follows: 24 hours at 5. foot candles intensity from day-old to 4 weeks of age,.5 foot candles from 4 weeks of age until time of lighting for reproduction, with daylength reduced to 12 hours at 6 weeks of age and then further reduced to 6 hours at 2 weeks of age. When the toms reached 26 weeks of age and the hens 3 weeks of age, they were exposed to a 14-hour day with a light intensity of 5. foot candles. One of the duplicate pens on each restriction level was switched to full feed while the other continued on the restrictive regimen. For the toms, this restriction continued throughout the reproductive period of the test. For the hens, the restrictive regimen was continued only 6 weeks from the time of stimulatory lighting or approximately 3 weeks after the start of egg production. The latter was an attempt to further retard onset of production. Individual records of egg production were obtained by trapnesting* Semen samples were collected from all toms at 34, 38, and 43 weeks of age. Measurements included total volume per ejaculate, percent sperm motility after 6 hours 1 storage at 4 C., number of motile sperm per ml. and per ejaculate, as well as the number of toms yielding semen at each collection. Results The effect of these feed restriction treatments on average body weights of the hens is shown in Table 1. By 3 weeks of age, the desired degree of restriction in body weight had been approximated, averaging 87 percent and 76 percent for the two treatments of which the goals had been 9 and 8 percent, respectively. Further restriction of one of the duplicate pens in each treatment had only a slight effect as noted in the 38-week weights. The 42-week weights suggest a slight trend toward minimizing the weight differences, but it is not likely that they would be eliminated by the end of the reproductive period. The number of days by which feed restriction delayed the achievement of 5 percent egg production after exposure to the stimulatory effect of a 14-hour day are given in Table 2. The restriction to 87 percent of body weight retarded onset of lay by 3 or 4 days. The more severe degree of restriction to 76 percent of body weight delayed onset of lay 6 or 8 days. In each case, the continuation of restriction after initiation of the 14-hour day had no further effect. -72-
76 Table 1, Effect of Feed Restriction on Body Weight. HENS Treatment Description Pen No. No. of Hens (3 wk) 3 Wk Lb (%) i Age in Weeks 38 Wk Lb (%) 42 Wk Lb (%) Full Fed Full Fed 7A 8A Mean Restr. to 3 Restr. to 36 to 9% Wt Wk. to 9% Wt Wk. 9A 1A Mean (87) (86) (87) (93) (92) (92) (94) (95) (94) Restr. to 3 Restr. to 36 to 8% Wt Wk. to 8% Wt Wk. 11A 12A Mean (75) (79) (76) (89) (85) (87) (9) (89) (89) percent of full-fed mean weight. Table 2. Effect of Feed Restriction on 77 Day Egg Production (July, August, and September). Treatment Description Pen No. No. of Hens Delay to 5% Prod. (days) Settable Eggs Per Hen (no.) Mean Full Fed Full Fed 7A 8A Restricted to 9% of normal wt to 3 wk, then full fed Restricted to 9% of wt to 36 wk, then full fed 9A 1A Restricted to 8% of wt to 3 wk, then full fed Restricted to 8% of wt to 36 wk, then full fed 11A 12A
77 The average number of settable eggs produced per hen during 77 days of lay (Table 2) show a striking increase as a result of restriction during the growing period. The less severe restriction to 87 percent of normal body weight at 3 weeks of age appears to be the best treatment. Continuation of the restriction after initiation of stimulatory lighting tended to reduce egg production. Although the 77-day egg production was fairly poor, averaging between 4 and 5 percent, it must be considered that this was during the hottest time of the year July, August, and early September. r ihe effect of feed restriction on average body weights of torus is shown in Table 3. At 26 weeks of age, the restricted birds weighed 83 and 73 percent of their full-fed counterparts. While this represents a greater restriction than occurred in the hens, semen collection was not started until the birds were 33 weeks of age and by 3 weeks of age the differences in body weight had moderated to the point where treated groups averaged 92 and 81 percent of the ad libitum fed controls. Continuation of feed restriction beyond 26 weeks of age was ineffective at the 9 percent level of intake but had a marked effect of inhibiting the tendency to catch up with the body weights of the full-fed birds. Restriction of feed intake caused no improvement in several traits relating to reproductive capacity of the toms (Table 4). The moderate restriction to 9 percent of control weight throughout the growing and breeding period was the only treatment which even equalled the performance of the full-fed treatment. All other restrictive treatments resulted in decreases in semen volume, sperm numbers, and percent motility. Table 3. Effect of Feed Restriction on Body Weight. TOMS Treatment Description Pen No. No. of Toms (26 wk) 26 Wk Lb OS) 1 Age in 3 Wk Lb (%) Weeks 38 m Lb (%) 42 Wk Lb (%) Full Fed Full Fed 5A 6A Mean Restr. to 9% wt to 26 wk Restr. to 9% wt continuously 3A 4A Mean (82) (84) 28.3 (83) 35,7 (92) 35,5 (91) 35.6 (92) (93) (89) (91) (93) ( (93) Restr. to 8% wt to 26 wk, Restr. to 8% wt continuously 1A 2A Mean (74) (81) 35.7 (86) (89) (72) 24,6 (73) 31.9 (8) (81) (79) (83) (78) 35,55 (83) percent of full-fed mean weight. -74-
78 Summary Restriction of feed intake of Large White turkeys from 3 weeks of age resulted in body weight reductions by 3 weeks of age to 9 and 8 percent of full-fed control bird weights. In hens, this restriction resulted in an increased production of settable eggs. Continuation of feed restriction for 6 weeks beyond the initiation of the 14-hour day reduced or eliminated this advantage. In toms, restriction to 9 percent of normal weight through both the growing and reproductive period resulted in semen production equal to that of the full-fed toms. The other restrictive treatments reduced the volume of semen, the number of motile sperm, and the percent motility of sperm. Table 4. Effect of Feed Restriction on Semen Production. Semen Trait Age (wks) 9% of Control to 26 Weeks then ad lib 9% of Control Wt 8% t>f Control to 26 Weeks then ad lib 8% of Control Wt Control Vol., ml Mean, , , % Producing Semen Mean , % Motility After 6-Hr Storage Mean , ,8 67, Motile Sperm/ml Mean Motile Sperm/Ejac Mean , ,
 eggs will contribute toward reducing the high cost of poults.")
79 THE EFFECTS OF LEVEL AND FORM OF CALCIUM IN THE TURKEY BREEDER HEN DIET ON EGG SHELL QUALITY AND REPRODUCTIVE PERFORMANCE S. P. Touchburn and K. E. Nestor Department of Poultry Science In a normal 2-week reproduction season, a large type turkey hen produces 7 eggs, usually yielding only about 4 poults. Any increase in the number of settable (sound-shelled) eggs will contribute toward reducing the high cost of poults. Shell quality is positively related to hatchability and therefore improvements in shell quality should further improve the economics of poult production. The work reported here is part of a major goal to determine the dietary physiological factors influencing blood calcium levels and their relationships to shell formation and hatchability in the turkey. Since such information relating to turkeys is extremely sparse, the initial phases of the research are aimed at establishing the norms for this species, Earlier work here (Nestor et al., 1972, Poultry Sci., 51: 14-11) provided data concerning the changes in various egg quality traits over a 7-month laying period. Two strains of turkeys, one selected for meat production and one selected for egg production, were used. Incubation weight loss of the eggs generally increased as the age of the hens increased. Shell weight and percent shell decreased with age of the hens, especially during the last few months of the laying period. The meat strain hens had significantly greater egg weight, shell weight, and incubation weight losses than those of the egg strain, Hatchability of fertile eggs and number of poults produced per hen consistently declined as the laying period advanced. Nestor et al. (1972, Poultry Sci., 51: ) showed that shell weight and percent shell of turkey eggs were positively associated with hatchability, while incubation weight loss was negatively associated with hatchability. Scott et al. (1971, Poultry Sci., 5: ) reported that replacement of two-thirds of the pulverized limestone of a normal laying hen diet with hen-size oyster shell significantly improved egg shell strength. They attributed this improvement to a constant metering of calcium from the gizzard into the bloodstream during the night when the gastro-intestinal tract was otherwise empty of food. In opposition to this explanation, Roland et al. (1972, Poultry Sci., 51: ) found that 87 percent of the calcium carbonate meter ed from the gizzard during a 24-hour period was metered out between 8 a.m. and 8 p.m., with very little metered out at night. E xp er iment a 1 P r p_ce dure The experiment reported here was designed to measure the effects of level and form of dietary calcium on egg shell traits and reproduction. Turkey hens of a strain selected for egg production were placed in individual laying cages and restricted to a 6-hour day from 24 weeks of age. They were fed the Ohio turkey hen breeder ration containing 2.25 percent calcium until 1 week prior to exposure to a 14-hour day, at which time each of the four experimental diets was fed to 36 hens. Diet 1 was the control (Ohio turkey hen breeder diet, Ohio Poultry Rations, 197, Ohio Cooperative Extension Service, Bull. 343) containing 2.25 percent calcium. Diet 2 was calculated to contain 1.75 percent calcium, which was believed to be a sub-optimal level. Diet 3 had the same calculated level of calcium as Diet 2 but -76-
80 with two-thirds of the ground limestone replaced by hen-size oyster shell flakes. Diet 4 contained 2.48 percent calcium and was considered to be approaching the maximum level which might be tolerated without adversely affecting reproduction. All diets contained 16 to 17 percent protein by analysis and were calculated to provide.83 percent total phosphorus. The hens were artificially inseminated weekly or bi-weekly throughout the 18- week reproductive period and the sound-shelled eggs were incubated. Individual records were obtained for feed consumption, egg production, egg weight, fertility, hatchability, and embryos dead at 7 and 28 days of incubation. Measures of shell quality included weight loss of eggs during the first week of incubation, shell (percent of egg weight), and the numbers of broken, cracked, thin-shelled, or shellless eggs produced. Results The mean values for egg weight and incubation measurements are shown in Table 1. Dietary levels or form of calcium had no significant effects on egg weight, weight loss of eggs during the first week of incubation, fertility, hatchability, or embryonic mortality at 7 or 28 days. The average numbers of eggs produced per hen during the 18-week reproductive period were significantly lower on the 1.75 percent calcium Diet 2 than on the 2.25 percent calcium control Diet 1 (Table 2). Replacing two-thirds of the dietary limestone of Diet 2 with oyster shell flakes (Diet 3) completely eliminated this depression. Egg production on Diet 4 which contained 2,48 percent calcium was equal to that of the control ration. The dietary treatments had no significant effects on any measure of shell quality (Table 2). These data included shell (percent of egg weight), percent normal eggs, and percentages of broken, shell-less, slab-sided, or weak-shelled eggs. From these results, it is apparent that turkey hens are capable of producing normal shelled eggs from a wide range of dietary calcium levels. The only clearcut difference was reduced egg production on the low calcium diet, which suggests Table 1. Effect of Dietary Treatment on Egg Weight, Incubation Weight Loss and Reproduction. ** Diet Egg Wt. Gm. Incubation Wt. Loss % Fertility % Hatch of Total % Hatchability % Embryos 7 Days Dead at: 28 Days 1. Control, 2.25% Ca A, % Ca A % Ca (1/3 limestone: 2/3 oyster shell) % Ca '-None of the differences was statistically significant. -77-
81
82 that the first limitation of calcium deficiency affected the amount of bound calcium available for yolk formation. Reduced rate of yolk formation would directly limit the rate of egg production. The positive effect on egg production observed when part of the calcium was supplied as oyster shell may suggest an effect on calcium availability which probably relates to time. This would agree with the hypothesis of Scott et al. (1971) of a constant metering of calcium from the gizzard into the bloodstream during the night when the gastro-intestinal tract was otherwise empty. The mean values of Table 2 provide a reasonable set of figures for turkey breeder hens maintained in wire cages. An average of 7 eggs per hen in 2 weeks from the time of stimulatory lighting is quite normal production. The 81.8 percent value for normal or settable eggs, however, is probably lower than it would be for a floor-housed flock. This is accounted for largely by the 1.9 percent of broken eggs. These represent apparently normal or sound-shelled eggs which were broken by impact with the wire or hard plastic bottom of the cage when they were laid. A 3 percent drop in feed consumption accompanies the achievement of peak egg production, a situation which seems far from ideal. This phenomenon will be the subject of further investigation. In Figure 1, the average feed consumption and average production of total eggs per hen are shown graphically. Egg production rises rapidly 2 weeks after initial exposure to the stimulatory effect of a 14-hour day. The first major drop in egg production at 11 weeks post lighting comes as a result of the first wave of broodiness. This is followed by a second, less dramatic broody period at 14 to 15 weeks which is followed by a rapid decline in production. -79-

83 A STUDY OF PLASMA CALCIUM LEVELS IN LAYING TURKEY HENS Margery A* Musser 9 W. L. Bacon, and S. P. Touchburn Department of Poultry Science Many studies have been made of calcium metabolism and shell quality in chickens but very little work has been done with turkeys. Poor egg shells reduce the profits of a turkey breeding flock in several ways. Many eggs are broken before they reach the incubator. Others are discarded because of structural weaknesses or cracks. Still others which appear normal are too porous and allow excessive moisture loss during incubation, which in turn reduces hatchability. To attack this problem of poor shell quality, more must be known about the mechanism of shell formation and the dynamics of calcium utilization. Determination of the changes in plasma calcium during the laying cycle is necessary before the dynamics of calcium metabolism can be understood. Calcium is required at a rate of 2 mg. per hour during the 15 hours of shell calcification. Each shell contains about 3 g. of calcium which is deposited around the egg in the form of calcium carbonate by the shell gland. The shell gland docs not store calcium and so all of the shell calcium must come from the blood plasma. The total amount of calcium in the plasma of a turkey hen is about 5 mg. Since this is only 1/6 of the quantity of calcium needed for each egg shell, this 5 mg. must be replaced on an average of every 15 minutes during shell calcification. Two things change in the hen before the start of egg production which help it meet this tremendous demand for calcium. The total blood calcium rises twofold or more and the bones undergo structural changes. Calcium is found in three forms in the blood plasma: ionic, in a simple organic compound, and protein bound. The first two make up the diffusible fraction. The protein bound fraction is part of a large lipoprotein complex associated with egg yolk formation which is produced from this time until egg production ceases. A rise in this fraction causes the rise in total blood calcium. Diffusible calcium stays fairly constant. The structural change in the bone is the formation of medullary bone inside the marrow cavities of bones which have a good blood supply. This newly formed bone provides a readily available source of calcium. Its presence is initiated and maintained by the action of estrogens circulating in the blood. When diffusible plasma calcium level drops, parathyroid hormone is released, causing calcium mobilization from this bone. Conversely, a rise in diffusible calcium level stimulates the release of calcitonin which causes deposition of calcium in this bone. Thus, this bone serves as a reservoir for maintaining the blood calcium levels in spite of the wide fluctuations in demand, while sparing the structural skeleton from depletion. The ultimate source of calcium is the diet. Absorption is dependent on an adequate supply of Vitamin Dj. The presence of parathyroid hormone causes the conversion of Vitamin Dj to a more active form which brings about increased absorption of calcium by the gut. In the present study, actual levels of bound and diffusible calcium were determined during the egg laying cycle of turkeys. The experimental work was carried out on egg line hens in the llth generation of selection for high egg production. These hens were on a 14-hour day (2 a.m. to 4 p.m.) and had been in production between 16 and 19 weeks when samples were collected. They were fed one of four diets containing from 1.75% to 2.48% calcium. All were individually caged in the same room.
84 Table 1. Plasma Cale^un Oorcent rut ions in Le\ ing Fen > Turing Egg tormatioi. Tine Number Collections 9 AM 1PM 5 PM 9 PM 1 AM AM 9 AM LSD^ 6 7 P<.1 P<.5 Diffusible Calcium S I ^1±* Bound Calcium 12.4^ ^ , Total Calcium J Mean in mg% + standard deviation. oa Least significant difference calculated by nethod c f Tukev On the day of sampling, hens which were selected had laid an egg before 9 a.m. that day. Blood samples were taken from the wing vein of each bird at 4-hour intervals over a 24-hour period. Only samples from birds with an egg in the uterus were analyzed. All samples after 5 p.m. were taken during shell calcification. Blood samples of 4-5 ml. were placed in heparinized tubes and immediately covered with mineral oil to prevent loss of C2- Escape of CO^ would change the blood ph and thus alter the ratio of bound to diffusible calcium. The blood was centrifuged to obtain the plasma* An aliquot of plasma still protected under oil was placed in a membrane cone which retained components of greater than 5, molecular weight and spun for 1 hour at 1 x gravity to obtain an ultrafiltrate. Aliquots of the ultrafiltrate were taken for determination of diffusible calcium. Aliquots of whole plasma were wet ashed with nitric and perchloric acids for determination of total calcium. All samples were then taken to appropriate dilutions and analyzed by atomic absorption. Individual values for bound calcium were obtained from the difference between the directly determined total and diffusible calcium values. Dietary treatment seemed to have no effect on diffusible or bound calcium levels and so the data from all birds were pooled. Means and standard deviations were calculated from the data of 18 birds for each of the seven sampling times (Table 1). Total calcium levels were lower than they would have been earlier in the production season. Table 2. Analysis of Variance for Diffusible, Bound and Total Calcium. Source Diffusible Calcium Bound Calcium Total Calcium df ms ms ms Time ** ** Error
85 Of the diffusible calcium means, only the one for collection 4 was significantly different (P<.5) from the others. This was the first collection taken after shell calcification had started. For both bound and total calcium, there was a significant drop between the values for period 1 and those for period 7. This drop may have been due partly to the stress of frequent handling of the birds during the 24 hours when samples were taken. For the 8 days prior to the experiment, egg production averaged 72%. During the 8 days following the experiment, three birds stopped laying and production averaged 42%. Calculations were made to determine the average amount of circulating plasma calcium present in each fraction at the time of collection 5. The results were 17, 35, and 52 mg. calcium for diffusible, bound, and total calcium, respectively. The average calcium content of each turkey egg shell was 3.25 g. A constant rate of deposition during the 15 hours of shell formation and a lack of equilibrium between bound and diffusible calcium was assumed in the following cases. If only the bound calcium were available for shell deposition, turnover time for all of the bound calcium would be 9.8 minutes. If only the diffusible calcium were used, its turnover time would be 4.6 minutes. If the two forms were in equilibrium and the total circulating calcium was available, the turnover time would be 14.3 minutes. These calculations illustrate how dynamic the system must be regarding calcium during egg shell formation in the turkey. -82-
 are formed in the liver under the influence of female sex steroid hormones (estrogens).")
86 CONCENTRATION OF A YOLK PRECURSOR IN THE PLASMA OF THREE LINES OF TURKEYS Wayne L. Bacon, Karl E. Nestor, and Margery A. Musser Department of Poultry Science It is well established that yolk low density lipoprotein precursors (LDF) are formed in the liver under the influence of female sex steroid hormones (estrogens). These are then released into the blood plasma and transported to the ovary where they are removed by follicles undergoing rapid development. Rapid development is the last phase of follicle development during which 98 6 of follicle growth occurs with the deposition of yellow yolk. It occurs in turkeys during the last 11 to 13 days prior to ovulation. Rapid development and ovulation are under control of the pituitary gonadotropic hormones, follicle stimulating hormone (FSH), and ovulation inducing hormone (OIH), respectively. It was postulated (Jaap, 1969, World 1 s Poultry Sci. J., 25:14-143) that inheritance which favors rapid body growth may also favor increased formation of LDF by the liver. Recently, a relatively simple method was developed to determine the blood plasma concentration of LDF. In the previous Turkey Research Summary (No. 47, 197), preliminary data based on a column chromatograph method was reported. This preliminary study indicated a higher concentration of plasma LDF in a meat line than an egg line of turkeys. This report gives more extensive results in comparisons of meat and egg lines of turkey hens, Ihree genetically different lines were compared. One line (E) was developed l>v selection for high egg production. When used, it was in its llth generation of selection. A second line was a modern randombred control (RBC) population, derived from two e*lite commercial growth lines in 1966, The third line (F) was a sub-line of the RBC selected for four generations for increased 16-week body weight. The latter two lines differ greatly in egg production potential from the egg line, but not from each other. In 1969, the egg line produced 114 eggs in 18 days while the growth strains averaged 81.5 eggs in 18 days. Hens used in this study were in full egg production and had laid eggs on the 3 days prior to sampling. The method for determining relative plasma LDF concentration is based on density gradient centrifugation of blood plasma and determining the optical density (O.D.) at 28 nm of the fraction containing the LDF. Total relative amounts of plasma LDF were calculated by multiplying body weight by.46 (McCartney, 1952, Poultry Sci.,31: ) to estimate plasma volume and then multiplying the relative plasma LDF concentration (O.D. at 28 nm) by plasma volume. The results of these comparisons are given in Table 1. No significant differences were observed between the three lines for relative concentration of plasma LDF. However, differences were noted between the three lines for body weights, and thus plasma volumes. The relative total amounts of plasma LDF showed that the RBC line had more total plasma LDF than the E line, while the F line was between the E and RBC lines. Thus, the heavier birds were shown to have more total plasma LDF, as was postulated. Considerable differences in plasma LDF concentration were noted within lines, as indicated by the relatively large standard errors in Table 1. This may indicate -83-
87 Table 1. Comparison of Body Weights, Optical Densities of the Plasma LDF and Estimated Total Plasma LDF in Three Lines of Turkeys. Line n Concentration (Optical Density) Factor Body Wt. (kg) Total Plasma LDF (Concentration X Plasma Volume) E RBC F 5.158* Isd Least significant difference (P<.1). Standard error. physiological differences in plasma LDF concentration associated with time in the egg production cycle, time of feed consumption, or nearness to a broody period. Recently, Bacon et al. (1972, Poultry Sci., 51: ) found no difference in the number of ovarian follicles in rapid development between the E and RBC lines when measured after 8 to 14 weeks of egg production. This may indicate a slower rate of removal of plasma LDF in the RBC than E lines by follicles in rapid development. -84-
88 DEVELOPMENT OF OVARIAN FOLLICLES AND MEAT-TYPE TURKEYS IN EGG- Wayne L. Bacon and Karl E. Uestor Department of Poultry Science Early in the laying period (13 to 41 days), meat-type turkeys have more ova in rapid development at autopsy and a greater ovarian ova weight than egg-type turkeys (Nestor et al., 197, Poultry Science, 49: ), while the duration of rapid development does not differ. The meat-type turkey hen thus produces more yolks than eggs. It was decided to collect additional data to see if the same differences between types of turkeys were present in the middle (56 to 97 days) as in the early portion of the laying period. The two lines compared in the first experiment were the egg line (E) and the randombred control (RBC), established in Samples of 4 females from each line were randomly selected when the birds were 24 weeks of age. Eight weeks prior to lighting for egg production (14 hours light per day at 39 weeks of age), they were given 6 hours of light per day. After 74 days of stimulatory lighting, or a mean of 56 days of egg production, the birds were caught daily and fed either a red or blue fat soluble dye. This allowed the determination of the duration of the rapid development period for each egg yolk for all subsequently laid eggs. After 41 days of dye feeding, all birds were killed and all ova in rapid development were removed and weighed. During the dye feeding portion of the experiment, egg weights, yolk weights, and the duration of rapid development and rest periods were determined for all eggs (Table 1). Significant differences were noted for all four measurements, with the RBC line being greater in each instance. This was also noted early in the laying season as reported by Nestor et al. (197). Data collected at autopsy are given in Table 2. No difference in the number of ovarian ova per hen was noted in this experiment. This is in contrast to early in the laying season when such a difference was noted. Ovarian ova weight was not significantly different between lines. Such a difference was noted early in the laying period. However, late in the laying period, ovarian number and weight were greater in the RBC than E line. Early in the laying period, no difference was noted between the lines in the occurrence of atretic follicles (Table 2). In Table 1. Egg Weight, Yolk Weight, Period of Rapid Development and Rest Period for All Eggs Laid During the Dye Feeding Period in Meat and Egg Strain Turkey Hens. Strain Egg Weight Yolk Weight (g) Rapid Development (days) Rest (days) RBC a 27. l a a 1.14 a E b b b 1.4 b Means with different superscript letters are significantly different -85-
89
90 contrast, in the middle of the production period during the present experiment, the RBC line had significantly more atretic follicles per hen than the E line. The incidence of hens with egg yolk in the body cavity was greater in the RBC than E line, indicating a possible loss of ova through not entering the reproductive tract. Differences in ovarian synchronization between the two lines can be estimated by examining the number of ova developing in pairs. When this is done, based on ovum weight, dye banding pattern, or both weight and dye banding pattern, the RBC had more pairs, or were less synchronized, than the E line hens. A second group of hens were autopsied after 79 days of production. This group is divided into two sub-groups of heavy and light weight strains regardless of egg production (Table 3). In both the heavy and light groups of hens, egg production was relatively poor (42 and 37%, respectively). The birds in the heavy group had a greater number of follicles in rapid development, more total ovarian ova weight, more atretic follicles per hen, and more yolks as pairs than the light weight group, although the differences were not significant (Table 3). It appears from these data that the excess yolk syndrome is more closely associated with body size than production level. Table 4. Phenotypic Correlation Coefficients Between Egg Weight, Yolk Weight, -\ Period of Rapid Development and Rest Period Based on Birds x and Individual Eggs z Egg Yolk Rapid Wt. Wt. Development Based on Birds Yolk Wt..577** JL.JL JLJU Rapid Development Rest.98** Based on Eggs Yolk Wt..445** JL.JL. JUJL Rapid Development Rest -.119**.19** -.57 Based on 123 degrees of freedom, strain effect removed. ^Based on 957 degrees of freedom, strain and individual bird effects removed. * P<.1. In the first experiment, phenotypic correlations were calculated between all combination pairs of egg and yolk weights, and the duration of the rest and rapid development periods for all eggs laid during the dye feeding period (Table 4). Egg weight was positively correlated with yolk weight and rapid development, indicating that in individual birds or individual eggs, when a large egg is laid it has a larger yolk and that this yolk takes longer to develop than in birds laying smaller eggs or in individual smaller eggs. These correlations were not significant earlier in the laying period (Nestor et al», 197). The significant correlation of yolk weight and rapid development also indicates that birds laying larger eggs or indi- -87-
91 vidual larger eggs have a longer period of rapid development. This correlation was also significant early in the laying period (Nestor et al,, 197). Egg weight and the duration of the rest period were positively correlated based on birds, indicating that a bird tending to lay large eggs tended to also have a longer rest period. This was not found early in the laying season. Curiously, however, when the individual bird effect was removed, the smaller eggs tended to have the larger rest periods. It is known that in the turkey the first egg of clutches tends to have a longer rest period than subsequent eggs. If these eggs tend to be smaller, this would explain this negative correlation in eggs after individual bird effects are removed. Rapid development and rest were not related. Heavy weight strains of turkeys normally are not as efficient producers of eggs as most medium and light weight strains (page 21). The reasons for this apparent effect of body size on egg production are not clearly understood, but can be ascribed to differences in genetic potential for egg production and immediate environment in which this genetic potential has to function, i.e., the bird's body exclusive of the ovary and reproductive tract. All data collected to date indicate a probable negative effect of the size of the immediate environment (body weight) on the genetic potential of the ovary and reproductive tract. Therefore, reductions in body weight may be beneficial to lessen the above mentioned negative effect of body weight on reproductive potential. This, in fact, is found to be true (page 71). -88-
92 OVARIAN FOLLICULAR DEVELOPMENT IN EGG AND GROWTH LINES OF QUAIL Wayne L. Bacon, Karl E. Nestor, and Philip A. Renner Department of Poultry Science In Japanese quail, the sequence of events occuring during follicular growth and maturation is of shorter duration, but otherwise quite similar to that found in chickens and turkeys. In turkeys, large heavy lines normally are not as efficient producers of eggs as are most medium and small weight strains. Early in the reproduction season, a heavy line of turkeys had more ovarian ova in rapid development than an egg line, while later in the reproductive season no difference was noted in the two lines. In both instances, the egg line laid the greater number of eggs. Recently, a large bodied (P) line of Japanese quail, based on increased 4-week body weight, has been developed (Marks, 1971, Poultry Science, 5: ). Ovarian follicular growth and maturation in the P line was compared to a corresponding control line (C), The results are discussed in relation to the earlier work with turkeys. In the first experiment, measurements were started after 9 weeks of egg production. In the second experiment, measurements were started after 3 weeks of production. In both experiments, the C and P lines produced about the same number of eggs per bird both before and during the experiments (Table 1). At autopsy, in both experiments, both lines contained about the same number of ovarian follicles (Table 1) The P line contained more ovarian weight as follicles in the younger birds in Experiment 2 (Table 1). This is in agreement with earlier work in turkeys where in younger hens the larger birds contained a greater weight of ovarian follicles than smaller birds. Egg and yolk weights were greater (P<.1) in the P line in both experiments, and were highly correlated (Table 3). This indicates that larger eggs have larger yolks. The durations of rapid development and rest were not different in Experiment 1 with the older birds, but were in Experiment 2 or in the younger birds. A correlation was found between the duration of rapid development and yolk weight in both experiments, while no correlations were found with the rest period. In general, these results in quail are in agreement with results obtained with turkeys (page 85). -89-
93
94 Table 2. Yolk Weights and Duration of Rapid Development and Rest Periods for Control (C) and Large Body Weight (P) Lines of Quail. Experiment 1 was conducted with birds in the middle of the laying period and Experiment 2 was early in the production period. Egg Wt (g) Yolk Wt (g) Rapid Development (days) Rest (days) Experiment 1 Line C a 3.3 a 5.96 a.1 a Line P b 3.57 b 6.22 a.22 a Experiment 2 Line C 9.16 al 2.78 a 6.4 a.7 a Line P 1.52 b 3.38 b 6.42 b.21 b "Means within experiment are significantly different (P<.1) if they have different superscripts. Table 3. All Possible Correlations Between Egg Weight, Yolk Weight, Rapid Development and Rest Based on Birds Within Strain. Rapid Yolk Wt Development Rest Experiment 1 n = 16 Egg Wt.87**.3.4 Yolk Wt Rapid Dev..39 Experiment 2 n = 36 Egg Wt.88** Yolk Wt Rapid Dev..39* ** *P<.5 P<.1-91-
95 THE HALF LIFE OF THE VERY LOW DENSITY LIPOPROTEIN IN MALE AND LAYING FEMALE TURKEYS Wayne L. Bacon Department of Poultry Science The onset of egg production causes many physiological changes in lipoprotein metabolism in chickens and turkeys. A serum lipoprotein fraction, the very low density lipoprotein fraction (VLDL) is different in males and non~laying females than a very low density lipoprotein fraction (LDF) found in laying females (Tanabc et al., 1961, Proc. Soc. Exper. Biol. and Med,, 16: 56-51). Triglyceride content of VLDL increases from a trace in non-laying females to about 1-2 mg. per ml. in LDF at the onset of egg production. The basic underlying physiological cause of this change is not completely understood. Possible factors which may cause an alteration in plasma LDF levels are summarized as follows: 1. A change in the rate of removal of VLDL between male and nonlaying females vs. that for LDF in laying females. 2. Increased rate of synthesis of triglyceride (TG) for LDF during egg production. There are probably more possible factors than those listed above, but the present discussion will be limited to these two. No evidence exists in the literature concerning a change in the rate of removal of VLDL in males and non-laying females vs. LDF in laying females. This was done in this laboratory as described below. Males or laying females were injected with a tracer dose of 1- ^C~ palmitate. Blood samples were collected at various times post injection and the VLDL or LDF was isolated by ultracentrifugation, Triglyceride fatty acid (TGFA) content of VLDL or LDF was determined, along with the amount of radioactivity in the 1GFA. This allowed the calculation of the VLDL or LDF half lives (t^) and clearance rates (Table 1). The mean VLDL TGFA t^ in males was 38 ± 13 minutes, while in laying females the mean LDF TGFA t^ was 434 ± 6 minutes, or about 14 times as long as for VLDL in males. The concentration of TGFA, expressed as u equiv. TGFA per ml, plasma, were.88 ±.57 in VLDL from males and 21.5 ± 2.1 for LDF in laying females. Total plasma TGFA from either VLDL or LDF was calculated by multiplying concentration by plasma volume. Plasma volume was calculated by the relationship of body weight x.46 (McCartney, 1952, Poultry Sci., 31: ). Total TGFA in male VLDL was 77 ± 598 u equiv. per bird and for laying female LDF, 765 i 156 u equiv. per bird. The turnover time was calculated by multiplying the t^'s by the constant The metabolic clearance rates were calculated by multiplying turnover time by total TGFA in VLDL or LDF per bird. This gave a value of 14. t 1.8 u equiv. TGFA per bird per min. for VLDL in males and u equiv. TGFA per bird per min. for LDF in females. These values are not statistically different (P>.5). To correct the metabolic clearance rate to g. per day as stearate, the u equiv. per min. was multiplied by 144 min. per day and the value multiplied by the molecular weight of stearic acid, 284, This gave a value of 5.71 ± 4.43 g, per day as -92-
96
97 stearic acid for male VLDL and 4.98 ± 1.3 g. per day as female LDF. These values were not statistically different (P>.5). It is apparent, however, that the clearance rate in males is much more variable than in laying females. From these data, the following conclusions concerning TGFA metabolism in laying females vs. male turkeys can be made: 1. t x * in males is much shorter than in laying females. 2. Plasma concentration and total amount of TGFA is much greater in laying females than in males. 3. Assuming constant conditions, metabolic clearance rate per bird per day of TGFA is about the same in both laying females and males, The question can now be asked: What is happening to the VLDL in males and LDF in laying females? In males, VLDL TGFA has essentially two possible routes of disposal. One is to be deposited in the adipose tissue stores as fat. The other is to be broken down to free fatty acids (FFA) and be recycled to be used as energy or resynthesis of more TGFA. The present data give no indication of the ultimate fate of the VLDL TGFA. In the laying female, it can be assumed that all of the LDF eventually is removed by follicles in rapid development and exits the bird via egg yolk. If it is assumed an egg yolk weighs 3 g., has 5% solids, and is 67% LDF (all values experimentally determined), it then contains 1 g. of LDF. LDF is 65% TG, of which 9% is TGFA. This gives 5.4 g. of TGFA per yolk. If the hen has a production rate of 1%, 5.4 g. compares favorable with the value of 4.98 ± 1.3 experimentally determined. If the hen lays at a rate of 6%, she would have an average production of 3.24 g. of LDF TGFA per day, a value still not too far from that calculated using the LDF t^ and LDF TGFA concentration. -94-
 levels in chickens may increase from about.3 u equiv. per ml. plasma in pullets to about 1. to 2, u equiv.")
98 PLASMA FREE FATTY ACID LEVELS IN LAYING TURKEYS Wayne L. Bacon and Margery A. Musser Department of Poultry Science It has been reported that plasma free fatty acid (FFA) levels in chickens may increase from about.3 u equiv. per ml. plasma in pullets to about 1. to 2, u equiv. per ml. in laying hens (Heald and Badman, 1963, Biophys. Biochim. Acta., 7: ). These determinations were made by a titrimetric method (Dole and Meinertz, 196, J. Biol. Chem., 235: ). A similar study has recently been reported for turkeys (Bajpayee and Brown, 1972, Poultry Sci., 51: ). Prior to long day lighting of 14L:1D for egg production, young turkey hens had an FFA concentration of (standard error) u equiv. per ml. plasma. After 21 days of long day lighting, the concentration increased to u equiv. per ml. The plasma FFA levels in this experiment were determined by a colorimetric procedure (Kvam et al., 1964, J. Pharmaceutical Sci., 4: ), which is based on forming the fatty acid (FA) copper salts and then determining the amount of copper in the FA! s after they are extracted. Unfortunately, phospholipids (PL f s) may also complex with copper, and when present in large amounts may give large errors in plasma FFA concentration when the copper content of the FA extract is assumed to be complexed only with FA f s. During the course of FFA metabolism studies, the concentration of plasma FFA in laying turkeys was found to be much lower than previously reported, ranging from.6 to.2 u equiv. per ml. In these samples, PL f s were removed and FA's extracted from the PL-free extracts before being determined by the method of Novak (1965, J. Lipid Res., 6: ). This method of FA determination is chemically similar to that of Kvam et al. except that the cobalt instead of copper salts of the FA are determined. A test was designed to determine whether removal of PL's from laying turkey plasma lipid extracts had an effect on plasma FFA levels. In the first trial, plasma was pooled from several laying turkeys and extracted, either with or without added palmitic acid, by the method of Trout et al. (196, J. Lipid Res., 1: ). Two samples with and two without added FA were set aside for colorimetric determination of FA by the Novak procedure. PL was removed from the remaining samples by silica gel (Bio Sil HA, BioRad, Richmond, Calif.) chromatography as follows. One and one-tenth g. of Bio Sil HA was weighed and covered with hexane: diethylether (H:D) 1:1. A column (.6 cm. id.), plugged with glass wool, was then filled with the Bio Sil slurry and allowed to run dry. The plasma extract was then added to the colbin in 1. ml. H:D. The test tube containing the lipid extract was then washed two times with additional 1. ml. aliquots of H:D and added to the column. After this, 7. ml. additional H:D were added to the column and it was allowed to run dry. This procedure removed the PL's, but allowed recovery of FA! s as indicated in Table 1. Two samples with added FA and two without added FA were set aside for FFA determination after Bio Sil HA chromatography. FA f s can be extracted from lipid extracts by partitioning with iso-octane and alkaline ethanol (Borgstrom, 1952, Acta. Physiologica. Scand., 25: ). The ethanol is then acidified and re-extracted with iso-octane to recover the FA f s. Two samples with and two without added FA were extracted and chromatographed as above and then partitioned in this manner. The results of this experiment are given in Table
99 Table 1. Fatty Acid Assayed From Trout Extract, Extract After Bio Sil HA Chromatography, and Extract After Both Bio Sil HA Chromatography and Alkaline EtOH Solvent Partition. All values are u equiv/ml plasma + standard error. Trout Extract After Bio Sil HA After EtOH Partition With FA Added , 3 Without FA Added Difference Added.4 u equiv palmitic acid per sample. Only 1 sample determined. It Is clearly seen from the data in Table 1 that removal of PL's by the relatively simple Bio Sil HA Chromatography step reduces detectable FFA levels by about 75%, while not reducing the recovery of added FA, After isolation of the FA f s by the alkaline EtOH partition, the values for samples with and without added FA were essentially the same as before EtOH solvent partition. These data raise the question of whether Heald and Badman and Bajpayee and Brown were detecting increases in circulating plasma FFA or whether they were in fact detecting increases in circulating plasma PL's, which are bound mainly to yolk lipoprotein precursors* It is well known that yolk lipoprotein precursors increase from a trace to over 2 mg. per ml. during the onset of egg production, and that the yolk lipoprotein precursors contain a considerable amount of PL's. Without removal of PL's from the llpid extracts before FFA determination, the above authors were probably detecting increased levels of circulating PL's rather than FFA's, which remain at a concentration comparable to that found in males or females prior to the onset of egg production. -96-

100 PROGRESS IN TURKEY SEMEN RESEARCH Keith I. Brown Department of Poultry Science The turkey hatching egg industry needs a semen extender which will allow storage for long periods of time without loss in fertilizing capacity. At QARDC, attempts are being made to solve this problem in two ways. First, research on freezing turkey semen is being conducted, If this could be done without loss of fertility and hatchability, semen could be stored for years. Secondly, attempts are being made to develop an extender which will allow several hours 1 storage in the liquid state. If semen can be stored 4 to 8 hours in the liquid state, it will be possible to control Mycoplasma meleagridis and other disease-bearing agents in the semen. In addition, it would be possible to make one semen collection and transport the semen long distances, thus making it unnecessary to house a flock of toms with each flock of hens. Research to date indicates that extender requirements are different for storage in the liquid state and for freezing (Table 1). High quality turkey semen (initial motility = 8%) diluted 1 part of semen to 7 parts of freezing extender (Extender 1) will be 2% motile after 24 hours' storage at 4 C. The same semen diluted 1:7 in the extender developed for storage in the liquid state (Extender II) will be 5 to 6% motile after 24 hours of storage at 4 C. If 7% ethylene glycol is added to each of the extenders, a 1:5 dilution is made, and the samples are frozen, the motility after thawing will be about 55% for Extender I and 25% for Extender II. Metabolic studies (oxygen consumption) indicate that a combination of glucose and citrate buffer are the major ingredients of Extender II which contribute to the livability of sperm diluted in this extender* A preliminary fertility trial (page 49) where semen diluted threefold with Extender II and stored for 3 minutes before inseminations are made indicates that there is no loss in fertility and hatchability. Further studies are planned to extend the storage time to 2, 4, 6, and 24 hours. Extensive research on freezing turkey semen has been conducted at the QARDC, One major requirement is a low ionic strength extender such as Extender I. Extensive tests have shown that the critical temperature for successful freezing is between -15 C. and -25 C. If the rate of temperature reduction is greater than 1 C, per second between -15 C. and -25 C., great cellular damage occurs. The rate of drop between 5 C. and -15 C. and below -25 C. can be extremely fast. In fact,, the faster one can freeze semen without the formation of intracellular ice crystals in the critical range between -15 C. and -25 C., the better the recovery after freeze. Based on these studies on freezing rate, the following procedure for freezing turkey semen is now used. The semen is collected into a tube held at 15 C. It is diluted (1 part of semen to 5 parts of extender) and equilibrated at 15 C. for 3 minutes. The cryophylactic agent used is 7% ethylene glycol. To freeze the semen,.5 ml. of diluted semen is placed in a 1 ml. ampule and the ampule is heat sealed. The ampule is passed through a series of baths as follows: -97-
101 Table 1. Turkey Semen Extenders. Chemical KC1 CaCl 2 MgCl 2 Extender I for Freezing (gm/liter) Extender II for Storage in the Liquid State (gm/liter) Glucose Inositol Citric Acid Sodium Salt * 2H 2 <3 Citric Acid Glutamate Sodium Salt Cysteine Tea Raff inose KOH NaOH MgSO 7H 2 Gentamicin Osmotic pressure «325 mosm. ph adjusted to 7,
102 Type of Bath C. bath 2. 4 C. bath C. ethyl alcohol C. ethyl alcohol C. ethyl alcohol 6. directly into liquid nitrogen at C. Time in Bath 3 minutes 1.5 minutes 2.5 minutes 1.5 minutes 1,5 minutes Using this freezing procedure and Extender I at a sixfold dilution and insemination dose of.5 ml. at weekly intervals, between 2 to 3% fertility with frozen semen can be routinely obtained. Summary Although considerable progress has been made in freezing turkey semen, the best fertilities obtained to date are between 2 and 3 r>. It is the author's opinion that no immediate improvement beyond 2 to 3% is in sight. On the other hand, extenders which allow several hours of storage in the liquid state appear possible in the near future. When this is accomplished, it will be possible to make one semen collection and transport the semen long distances, thus making it unnecessary to house a flock of toms with each flock of hens. However, none of the extenders currently on the market can be used to store semen even for short periods of time without loss in fertility and hatchability. Research to date indicates that extender requirements are different for storage in the liquid state and for freezing (Table 1). -99-
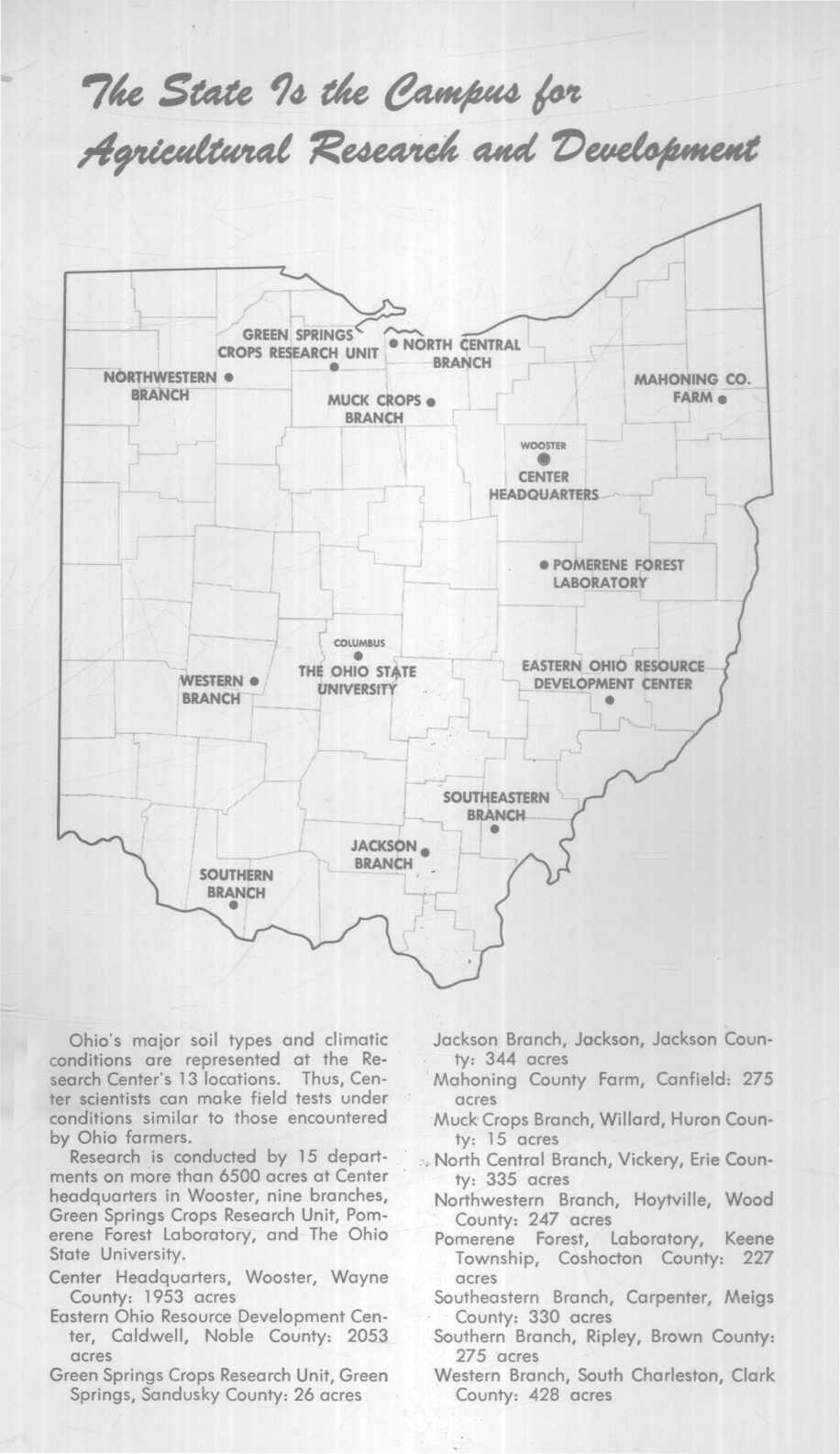
103
Selection for Egg Mass in the Domestic Fowl. 1. Response to Selection
 Selection for Egg Mass in the Domestic Fowl. 1. Response to Selection H. L. MARKS US Department of Agriculture, Science & Education Administration, Agricultural Research, uthern Regional Poultry Breeding
Selection for Egg Mass in the Domestic Fowl. 1. Response to Selection H. L. MARKS US Department of Agriculture, Science & Education Administration, Agricultural Research, uthern Regional Poultry Breeding
Performance of Broiler Breeders as Affected by Body Weight During the Breeding Season 1
 Performance of Broiler Breeders as Affected by Body Weight During the Breeding Season 1 H. R. WILSON and R. H. HARMS Department of Poultry Science, University of Florida, Gainesville, Florida 32611 (Received
Performance of Broiler Breeders as Affected by Body Weight During the Breeding Season 1 H. R. WILSON and R. H. HARMS Department of Poultry Science, University of Florida, Gainesville, Florida 32611 (Received
Recommended Resources: The following resources may be useful in teaching
 Unit D: Egg Production Lesson 1: Producing Layers Student Learning Objectives: Instruction in this lesson should result in students achieving the following objectives: 1. Discuss the materials and equipment
Unit D: Egg Production Lesson 1: Producing Layers Student Learning Objectives: Instruction in this lesson should result in students achieving the following objectives: 1. Discuss the materials and equipment
How Does Photostimulation Age Alter the Interaction Between Body Size and a Bonus Feeding Program During Sexual Maturation?
 16 How Does Photostimulation Age Alter the Interaction Between Body Size and a Bonus Feeding Program During Sexual Maturation? R A Renema*, F E Robinson*, and J A Proudman** *Alberta Poultry Research Centre,
16 How Does Photostimulation Age Alter the Interaction Between Body Size and a Bonus Feeding Program During Sexual Maturation? R A Renema*, F E Robinson*, and J A Proudman** *Alberta Poultry Research Centre,
FFA Poultry Career Development Event 2000 Poultry Judging Contest Arkansas State FFA Judging Contest
 FFA Poultry Career Development Event 2000 Poultry Judging Contest Arkansas State FFA Judging Contest Contestant Name: Contestant ID: 1. The per capita egg consumption is: A. 100 eggs per year B. 234 eggs
FFA Poultry Career Development Event 2000 Poultry Judging Contest Arkansas State FFA Judging Contest Contestant Name: Contestant ID: 1. The per capita egg consumption is: A. 100 eggs per year B. 234 eggs
FFA Poultry Career Development Event 2004 NEO Aggie Day. 1. With regard to egg storage, which of the following statements is FALSE?
 FFA Poultry Career Development Event 2004 NEO Aggie Day 1. With regard to egg storage, which of the following statements is FALSE? A. The longer the egg storage time, the higher the egg storage temperature
FFA Poultry Career Development Event 2004 NEO Aggie Day 1. With regard to egg storage, which of the following statements is FALSE? A. The longer the egg storage time, the higher the egg storage temperature
Effect of Calcium Level of the Developing and Laying Ration on Hatchability of Eggs and on Viability and Growth Rate of Progeny of Young Pullets 1
 1328 E. J. DAY AND B. C. DILWOETH for calcium:phosphorus ratios shows that toe ash was lowest for the birds receiving the rations containing the most narrow calcium:phosphorus ratio. Again, this observation
1328 E. J. DAY AND B. C. DILWOETH for calcium:phosphorus ratios shows that toe ash was lowest for the birds receiving the rations containing the most narrow calcium:phosphorus ratio. Again, this observation
CONTENTS ***** ***** *****
 CONTENTS ***** ***** ***** Implications of Selecting Poultry for High and Low Adrenal Response to Stress, by Keith I. Brown and Karl E. Nestor.... Effects of Stress on Rate of Gain and Feed Conversion
CONTENTS ***** ***** ***** Implications of Selecting Poultry for High and Low Adrenal Response to Stress, by Keith I. Brown and Karl E. Nestor.... Effects of Stress on Rate of Gain and Feed Conversion
EGG production of turkeys is not important
 A Study of Egg Production in Bronze Turkeys S. J. MAESDEN National Agricultural Research Center, Beltsville, Maryland EGG production of turkeys is not important commercially but good egg production during
A Study of Egg Production in Bronze Turkeys S. J. MAESDEN National Agricultural Research Center, Beltsville, Maryland EGG production of turkeys is not important commercially but good egg production during
Female Persistency Post-Peak - Managing Fertility and Production
 May 2013 Female Persistency Post-Peak - Managing Fertility and Production Michael Longley, Global Technical Transfer Manager Summary Introduction Chick numbers are most often reduced during the period
May 2013 Female Persistency Post-Peak - Managing Fertility and Production Michael Longley, Global Technical Transfer Manager Summary Introduction Chick numbers are most often reduced during the period
Female Persistency Post-Peak - Managing Fertility and Production
 Female Persistency Post-Peak - Managing Fertility and Production Michael Longley, Global Technical Transfer Manager May 2013 SUMMARY Introduction Chick numbers are most often reduced during the period
Female Persistency Post-Peak - Managing Fertility and Production Michael Longley, Global Technical Transfer Manager May 2013 SUMMARY Introduction Chick numbers are most often reduced during the period
FFA Poultry Career Development Event 2004 Poultry Judging District Contests
 FFA Poultry Career Development Event 2004 Poultry Judging District Contests 1. In a market broiler house, heaters should be turned on to preheat the house hours before the chicks arrival. A. 5-10 hours
FFA Poultry Career Development Event 2004 Poultry Judging District Contests 1. In a market broiler house, heaters should be turned on to preheat the house hours before the chicks arrival. A. 5-10 hours
Long-Term Selection for Body Weight in Japanese Quail Under Different Environments
 Long-Term Selection for Body Weight in Japanese Quail Under Different Environments H. L. MARKS USDA, Agricultural Research Service, Southeastern Poultry Research Laboratory, c/o The University of Georgia,
Long-Term Selection for Body Weight in Japanese Quail Under Different Environments H. L. MARKS USDA, Agricultural Research Service, Southeastern Poultry Research Laboratory, c/o The University of Georgia,
HAND BOOK OF POULTRY FARMING AND FEED FORMULATIONS
 HAND BOOK OF POULTRY FARMING AND FEED FORMULATIONS WHY POULTY FARMING? GENERAL ANATOMY OF POULTRY Feathers of fowl The Skin Skeletal System of Fowl Muscular System The respiratory system of fowl The digestive
HAND BOOK OF POULTRY FARMING AND FEED FORMULATIONS WHY POULTY FARMING? GENERAL ANATOMY OF POULTRY Feathers of fowl The Skin Skeletal System of Fowl Muscular System The respiratory system of fowl The digestive
Effect of EM on Growth, Egg Production and Waste Characteristics of Japanese Quail Abstract Introduction Experimental Procedures
 Effect of EM on Growth, Egg Production and Waste Characteristics of Japanese Quail S. Chantsavang, P. Piafupoa and O. Triwutanon Department of Animal Science, Kasetsart University, Bangkok, Thailand Abstract
Effect of EM on Growth, Egg Production and Waste Characteristics of Japanese Quail S. Chantsavang, P. Piafupoa and O. Triwutanon Department of Animal Science, Kasetsart University, Bangkok, Thailand Abstract
Unit D: Egg Production. Lesson 4: Producing Layers
 Unit D: Egg Production Lesson 4: Producing Layers 1 1 Terms broodiness caged layer production floor production layers 2 2 3 I. Layers are chickens that are used to produce large quantities of eggs. A.
Unit D: Egg Production Lesson 4: Producing Layers 1 1 Terms broodiness caged layer production floor production layers 2 2 3 I. Layers are chickens that are used to produce large quantities of eggs. A.
GENETICS INTRODUCTION. G. B. Havenstein,* 2 P. R. Ferket,* J. L. Grimes,* M. A. Qureshi, and K. E. Nestor
 GENETICS Comparison of the Performance of 1966- Versus 2003-Type Turkeys When Fed Representative 1966 and 2003 Turkey Diets: Growth Rate, Livability, and Feed Conversion 1 G. B. Havenstein,* 2 P. R. Ferket,*
GENETICS Comparison of the Performance of 1966- Versus 2003-Type Turkeys When Fed Representative 1966 and 2003 Turkey Diets: Growth Rate, Livability, and Feed Conversion 1 G. B. Havenstein,* 2 P. R. Ferket,*
Avian Reproductive System Female
 extension Avian Reproductive System Female articles.extension.org/pages/65372/avian-reproductive-systemfemale Written by: Dr. Jacquie Jacob, University of Kentucky For anyone interested in raising chickens
extension Avian Reproductive System Female articles.extension.org/pages/65372/avian-reproductive-systemfemale Written by: Dr. Jacquie Jacob, University of Kentucky For anyone interested in raising chickens
2015 Iowa State Poultry Judging CDE Written Exam Version A 1. What is the name of the portion of the digestive system that secretes hydrochloric acid
 1. What is the name of the portion of the digestive system that secretes hydrochloric acid and the enzyme pepsin? a. Rumen b. Gizzard c. Proventriculus d. Crop 2. In egg laying operations, production goals
1. What is the name of the portion of the digestive system that secretes hydrochloric acid and the enzyme pepsin? a. Rumen b. Gizzard c. Proventriculus d. Crop 2. In egg laying operations, production goals
The effects of shank length on incubation results of Japanese quails (Coturnix coturnix japonica) eggs and hatched chick shank length
 eggs and hatched chick shank length") The effects of shank length on incubation results of Japanese quails (Coturnix coturnix japonica) eggs and hatched chick shank length B. YILMAZ DIKMEN* and A. IPEK Faculty of Agriculture, Animal Science
The effects of shank length on incubation results of Japanese quails (Coturnix coturnix japonica) eggs and hatched chick shank length B. YILMAZ DIKMEN* and A. IPEK Faculty of Agriculture, Animal Science
RURAL INDUSTRIES RESEARCH AND DEVELOPMENT CORPORATION FINAL REPORT. Improvement in egg shell quality at high temperatures
 RURAL INDUSTRIES RESEARCH AND DEVELOPMENT CORPORATION FINAL REPORT Project Title: Improvement in egg shell quality at high temperatures RIRDC Project No.: US-43A Research Organisation: University of Sydney
RURAL INDUSTRIES RESEARCH AND DEVELOPMENT CORPORATION FINAL REPORT Project Title: Improvement in egg shell quality at high temperatures RIRDC Project No.: US-43A Research Organisation: University of Sydney
Key facts for maximum broiler performance. Changing broiler requires a change of approach
 Key facts for maximum broiler performance Changing broiler requires a change of approach Good chick quality = UNIFORMITY everywhere in the supply chain Performance 1. Professional breeder house / management
Key facts for maximum broiler performance Changing broiler requires a change of approach Good chick quality = UNIFORMITY everywhere in the supply chain Performance 1. Professional breeder house / management
Chapter 6 Breeder flock management
 Chapter 6 Breeder flock management The most important aspects of goose production are the management and feeding of the breeder flock as these can have a major impact on the reproductive rate, including
Chapter 6 Breeder flock management The most important aspects of goose production are the management and feeding of the breeder flock as these can have a major impact on the reproductive rate, including
THE OHIO TURKEY INDUSTRY. Robert E. Cook Chairman, Department of Poultry Science
 CONTENTS The Ohio Turkey Industry, by Robert E. Cook Recent Progress in G-enetic Improvement of Turkeys, by Karl E. Nestor and Philip A. Renner *..... 1 Efficient Use of Turkey Males, by Keith I* Brown...
CONTENTS The Ohio Turkey Industry, by Robert E. Cook Recent Progress in G-enetic Improvement of Turkeys, by Karl E. Nestor and Philip A. Renner *..... 1 Efficient Use of Turkey Males, by Keith I* Brown...
Allocating Feed to Female Broiler Breeders: Technical Bulletin #2
 Allocating Feed to Female Broiler Breeders: Technical Bulletin #2 Brenda Schneider 1, Martin Zuidhof 1, Frank Robinson 2 & Rob Renema 2 1 Alberta Agriculture, Food and Rural Development, 2 University of
Allocating Feed to Female Broiler Breeders: Technical Bulletin #2 Brenda Schneider 1, Martin Zuidhof 1, Frank Robinson 2 & Rob Renema 2 1 Alberta Agriculture, Food and Rural Development, 2 University of
FEEDING CHINESE RINGNECK PHEASANTS FOR EFFICIENT REPRODUCTION. Summary *
 FEEDING CHINESE RINGNECK PHEASANTS FOR EFFICIENT REPRODUCTION Robert E. Moreng, William K. Pfaff and Eldon W. Kienholz Summary * Two trials were conducted each using 240 Chinese Ringneck pheasant breeder
FEEDING CHINESE RINGNECK PHEASANTS FOR EFFICIENT REPRODUCTION Robert E. Moreng, William K. Pfaff and Eldon W. Kienholz Summary * Two trials were conducted each using 240 Chinese Ringneck pheasant breeder
Genetic improvement For Alternative Hen-Housing
 Genetic improvement For Alternative Hen-Housing Dr. Neil O Sullivan Hy-Line International 2015 Egg Industry Issues Forum Hy-Line International Genetic Excellence ! The Decision Process used in Breeding
Genetic improvement For Alternative Hen-Housing Dr. Neil O Sullivan Hy-Line International 2015 Egg Industry Issues Forum Hy-Line International Genetic Excellence ! The Decision Process used in Breeding
INCUBATION AND VITAL MORPHOLOGICAL TRAITS IN EGGS FROM AGE-RELATED TURKEYS
 Trakia Journal of Sciences, Vol. 7, No. 1, pp 63-67, 2009 Copyright 2009 Trakia University Available online at: http://www.uni-sz.bg ISSN 1313-7050 (print) ISSN 1313-3551 (online) Original Contribution
Trakia Journal of Sciences, Vol. 7, No. 1, pp 63-67, 2009 Copyright 2009 Trakia University Available online at: http://www.uni-sz.bg ISSN 1313-7050 (print) ISSN 1313-3551 (online) Original Contribution
Saskatchewan Breeder Meeting. July 7, 2015 Mark Belanger
 Saskatchewan Breeder Meeting July 7, 2015 Mark Belanger Where do Breeders fit Understand that the industry has changed and we are not doing business the same Goal of feeding the world is counting on chickens
Saskatchewan Breeder Meeting July 7, 2015 Mark Belanger Where do Breeders fit Understand that the industry has changed and we are not doing business the same Goal of feeding the world is counting on chickens
Improving Growth and Yield of Commercial Pheasants Through Diet Alteration and Feeding Program
 Improving Growth and Yield of Commercial Pheasants Through Diet Alteration and Feeding Program Sandra G. Velleman 1 and Nicholas B. Anthony 2 1 Department of Animal Sciences, The Ohio State University
Improving Growth and Yield of Commercial Pheasants Through Diet Alteration and Feeding Program Sandra G. Velleman 1 and Nicholas B. Anthony 2 1 Department of Animal Sciences, The Ohio State University
P O U LTOS CIE N G E
 P O U LTOS CIE N G E January, 1943? Vol. XXII, No. 1 The Relative Efficiency of Gains in Weight Made by Male and Female Bronze Turkeys* CONSIDERABLE data have been collected on feed used by turkeys at
P O U LTOS CIE N G E January, 1943? Vol. XXII, No. 1 The Relative Efficiency of Gains in Weight Made by Male and Female Bronze Turkeys* CONSIDERABLE data have been collected on feed used by turkeys at
FEEDER and FLOOR SPACE upon groy11ng TURKEYS. The effect of. in confinement. Wooster, Ohio OHIO AGRICULTURAL EXPERIMENT STATION J. W.
 RESEARCH CIRCULAR 87 JULY 1960 The effect of FEEDER and FLOOR SPACE upon groy11ng TURKEYS in confinement J. W. WYNE M. G. McCARTNEY R. D. CARTER V. D. CHAMBERLIN OHIO AGRICULTURAL EXPERIMENT STATION Wooster,
RESEARCH CIRCULAR 87 JULY 1960 The effect of FEEDER and FLOOR SPACE upon groy11ng TURKEYS in confinement J. W. WYNE M. G. McCARTNEY R. D. CARTER V. D. CHAMBERLIN OHIO AGRICULTURAL EXPERIMENT STATION Wooster,
ECONOMIC studies have shown definite
 The Inheritance of Egg Shell Color W. L. BLOW, C. H. BOSTIAN AND E.^W. GLAZENER North Carolina State College, Raleigh, N. C. ECONOMIC studies have shown definite consumer preference based on egg shell
The Inheritance of Egg Shell Color W. L. BLOW, C. H. BOSTIAN AND E.^W. GLAZENER North Carolina State College, Raleigh, N. C. ECONOMIC studies have shown definite consumer preference based on egg shell
EDUCATION AND PRODUCTION. Layer Performance of Four Strains of Leghorn Pullets Subjected to Various Rearing Programs
 EDUCATION AND PRODUCTION Layer Performance of Four Strains of Leghorn Pullets Subjected to Various Rearing Programs S. LEESON, L. CASTON, and J. D. SUMMERS Department of Animal and Poultry Science, University
EDUCATION AND PRODUCTION Layer Performance of Four Strains of Leghorn Pullets Subjected to Various Rearing Programs S. LEESON, L. CASTON, and J. D. SUMMERS Department of Animal and Poultry Science, University
POULTRY Allen County 4-H
 POULTRY Allen County 4-H Level 1 Grades 3-4-5 2017 $1.00 What you will do in this project: Enroll in the 4-H program by January 15. Complete the project by answering at least two of the activities in this
POULTRY Allen County 4-H Level 1 Grades 3-4-5 2017 $1.00 What you will do in this project: Enroll in the 4-H program by January 15. Complete the project by answering at least two of the activities in this
Body weight, feed coefficient and carcass characteristics of two strain quails and their reciprocal crosses
 1 Body weight, feed coefficient and carcass characteristics of two strain quails and their reciprocal crosses N.VALI 1, EDRISS, M.A. 2 and RAHMANI, H.R. 2 1 Department of Animal Sciences, faculty of Agriculture
1 Body weight, feed coefficient and carcass characteristics of two strain quails and their reciprocal crosses N.VALI 1, EDRISS, M.A. 2 and RAHMANI, H.R. 2 1 Department of Animal Sciences, faculty of Agriculture
Department of Animal and Poultry Sciences August 2, 1999
 Department of Animal and Poultry Sciences August 2, 1999 Erratic Oviposition and Defective Egg Syndrome (EODES) Effects of Genetic Selection for Body Weight Héctor L. Santiago ABSTRACT Reproductive performance
Department of Animal and Poultry Sciences August 2, 1999 Erratic Oviposition and Defective Egg Syndrome (EODES) Effects of Genetic Selection for Body Weight Héctor L. Santiago ABSTRACT Reproductive performance
Feeding the Commercial Egg-Type Replacement Pullet 1
 PS48 Feeding the Commercial Egg-Type Replacement Pullet 1 Richard D. Miles and Jacqueline P. Jacob 2 TODAY'S PULLET Advances in genetic selection make today's pullets quite different from those of only
PS48 Feeding the Commercial Egg-Type Replacement Pullet 1 Richard D. Miles and Jacqueline P. Jacob 2 TODAY'S PULLET Advances in genetic selection make today's pullets quite different from those of only
Arkansas State FFA Poultry Exam 2016
 Arkansas State FFA Poultry Exam 2016 Write answers on scantron. 1. For a typical egg laying operation, the production goals for a hen housed during a 52-80 week laying period is. a) 120 140 eggs b) 160-180
Arkansas State FFA Poultry Exam 2016 Write answers on scantron. 1. For a typical egg laying operation, the production goals for a hen housed during a 52-80 week laying period is. a) 120 140 eggs b) 160-180
Poultry Skillathon 2016
 Age Divisions: Junior (8-11) Intermediate (12-14) Senior (15-18) Exhibitors will participate in age-based Skillathons. This study guide includes all topics an exhibitor might be tested on. Youth will only
Age Divisions: Junior (8-11) Intermediate (12-14) Senior (15-18) Exhibitors will participate in age-based Skillathons. This study guide includes all topics an exhibitor might be tested on. Youth will only
Estelar CHAPTER-6 RAISING AND PRODUCTION OF POULTRY BIRDS
 CHAPTER-6 RAISING AND PRODUCTION OF POULTRY BIRDS Raising and production of poultry birds 6.1 INTRODUCTION Poultry- fish farming is the integration of poultry animals like chicken, duck and geese with
CHAPTER-6 RAISING AND PRODUCTION OF POULTRY BIRDS Raising and production of poultry birds 6.1 INTRODUCTION Poultry- fish farming is the integration of poultry animals like chicken, duck and geese with
Simplified Rations for Farm Chickens
 CIRCULAR 66 (Reprinted August 936) JUNE 934 Simplified Rations for Farm Chickens By D. F. KING Assistant Professor Poultry Husbandry G. A. TROLLOPE Professor Poultry Husbandry AGRICULTURAL EXPERIMENT STATION
CIRCULAR 66 (Reprinted August 936) JUNE 934 Simplified Rations for Farm Chickens By D. F. KING Assistant Professor Poultry Husbandry G. A. TROLLOPE Professor Poultry Husbandry AGRICULTURAL EXPERIMENT STATION
MAXIMIZING FERTILITY WITH ROOSTER MANAGEMENT. Jeanna L. Wilson University of Georgia
 MAXIMIZING FERTILITY WITH ROOSTER MANAGEMENT Jeanna L. Wilson University of Georgia Some might ask why we are having issues with fertility? To answer that question let s take a look back 6 week old broilers,
MAXIMIZING FERTILITY WITH ROOSTER MANAGEMENT Jeanna L. Wilson University of Georgia Some might ask why we are having issues with fertility? To answer that question let s take a look back 6 week old broilers,
SUMMARY OF THESIS. Chapter VIII "The place of research, its purpose, the biological material and method"
 SUMMARY OF THESIS Raising Japanese quail is a global activity still limited compared with growth of hens and broilers, but with great prospects for the development of characteristics and adaptability of
SUMMARY OF THESIS Raising Japanese quail is a global activity still limited compared with growth of hens and broilers, but with great prospects for the development of characteristics and adaptability of
Unit E: Other Poultry. Lesson 2: Exploring the Duck Industry
 Unit E: Other Poultry Lesson 2: Exploring the Duck Industry 1 1 2 I. There are many types of ducks throughout the world and in Afghanistan. A. Both domesticated and wild ducks exist throughout the world.
Unit E: Other Poultry Lesson 2: Exploring the Duck Industry 1 1 2 I. There are many types of ducks throughout the world and in Afghanistan. A. Both domesticated and wild ducks exist throughout the world.
Performance of Sudanese native Dwarf and Bare Neck Chicken raised under improved traditional production system
 AGRICULTURE AND BIOLOGY JOURNAL OF NORTH AMERICA ISSN Print: 2151-7517, ISSN Online: 2151-7525, doi:10.5251/abjna.2011.2.5.860.866 2011, ScienceHuβ, http://www.scihub.org/abjna Performance of Sudanese
AGRICULTURE AND BIOLOGY JOURNAL OF NORTH AMERICA ISSN Print: 2151-7517, ISSN Online: 2151-7525, doi:10.5251/abjna.2011.2.5.860.866 2011, ScienceHuβ, http://www.scihub.org/abjna Performance of Sudanese
1 of 9 7/1/10 2:08 PM
 LIFETIME LAMB AND WOOL PRODUCTION OF TARGHEE OR FINN-DORSET- TARGHEE EWES MANAGED AS A FARM OR RANGE FLOCK N. Y. Iman and A. L. Slyter Department of Animal and Range Sciences SHEEP 95-4 Summary Lifetime
LIFETIME LAMB AND WOOL PRODUCTION OF TARGHEE OR FINN-DORSET- TARGHEE EWES MANAGED AS A FARM OR RANGE FLOCK N. Y. Iman and A. L. Slyter Department of Animal and Range Sciences SHEEP 95-4 Summary Lifetime
A STUDY ON EGG QUALITY TRAITS IN JAPANESE QUAILS * (Coturnix coturnix japonica)
") A STUDY ON EGG QUALITY TRAITS IN JAPANESE QUAILS * (Coturnix coturnix japonica) B. Punya Kumari 1, B. Ramesh Gupta 2, M. Gnana Prakash 3 and A. Rajasekhar Reddy 4 Sri Venkateswara Veterinary University
A STUDY ON EGG QUALITY TRAITS IN JAPANESE QUAILS * (Coturnix coturnix japonica) B. Punya Kumari 1, B. Ramesh Gupta 2, M. Gnana Prakash 3 and A. Rajasekhar Reddy 4 Sri Venkateswara Veterinary University
History of the North Carolina Layer Tests. Detailed Description of Housing and Husbandry Changes Made From through 2009
 August 7, 2012 History of the North Carolina Layer Tests Detailed Description of Housing and Husbandry Changes Made From 1958 through 2009 Research conducted at the: North Carolina Department of Agriculture
August 7, 2012 History of the North Carolina Layer Tests Detailed Description of Housing and Husbandry Changes Made From 1958 through 2009 Research conducted at the: North Carolina Department of Agriculture
CHICKENS 101 BIOLOGY (ANATOMY, BREEDS, DEVELOPMENT, & REPRODUCTION)
") CHICKENS 101 BIOLOGY (ANATOMY, BREEDS, DEVELOPMENT, & REPRODUCTION) THE BASICS: TERMS 1 Chicken Anatomy 3 THE BASICS: TERMS 1 Chicken Wing & Feather Anatomy 4 THE BASICS - TERMS 1 Chick a new hatch or
CHICKENS 101 BIOLOGY (ANATOMY, BREEDS, DEVELOPMENT, & REPRODUCTION) THE BASICS: TERMS 1 Chicken Anatomy 3 THE BASICS: TERMS 1 Chicken Wing & Feather Anatomy 4 THE BASICS - TERMS 1 Chick a new hatch or
ON COMMERCIAL poultry farms during
 Effect of Date of Hatch on Weight F. P. JEFFREY Department of Poultry Husbandry, Rutgers University, New Brunswick, New Jersey (Presented at annual meeting June, 1940; received for publication May 23,
Effect of Date of Hatch on Weight F. P. JEFFREY Department of Poultry Husbandry, Rutgers University, New Brunswick, New Jersey (Presented at annual meeting June, 1940; received for publication May 23,
THE production of turkey hatching
 The Use of Artificial Lights for Turkeys* H. L. WlLCKE Iowa Agricultural Experiment Station, Ames, Iowa (Presented at Annual Meeting, August 1938; received for publication September 22, 1938) THE production
The Use of Artificial Lights for Turkeys* H. L. WlLCKE Iowa Agricultural Experiment Station, Ames, Iowa (Presented at Annual Meeting, August 1938; received for publication September 22, 1938) THE production
TECHNICAL BULLETIN Claude Toudic Broiler Specialist June 2006
 Evaluating uniformity in broilers factors affecting variation During a technical visit to a broiler farm the topic of uniformity is generally assessed visually and subjectively, as to do the job properly
Evaluating uniformity in broilers factors affecting variation During a technical visit to a broiler farm the topic of uniformity is generally assessed visually and subjectively, as to do the job properly
THE LAYING FLOCK VIRGINIA 4-H CLUB SERIES. AGIUCU LTUJiAL EXTENSION SERVICE OF V. P. I., BLACKSBURG, VA.
 LD 51c!95 Al~' c_...\
LD 51c!95 Al~' c_...\
. California Poultry Letter
 COOPERATIVE EXTENSION UNIVERSITY OF CALIFORNIA I. California Poultry Letter FIRST AND SECOND CYCLE EGG PRODUCTION RELATIONSHIPS A study of 1231 first cycle and 887 second cycle table egg flocks (by Bell
COOPERATIVE EXTENSION UNIVERSITY OF CALIFORNIA I. California Poultry Letter FIRST AND SECOND CYCLE EGG PRODUCTION RELATIONSHIPS A study of 1231 first cycle and 887 second cycle table egg flocks (by Bell
Poultry Skillathon 2017
 Age Divisions: Junior (8-11) Intermediate (12-14) Senior (15-18) Exhibitors will participate in age-based Skillathons. This study guide includes all topics an exhibitor might be tested on. Youth will only
Age Divisions: Junior (8-11) Intermediate (12-14) Senior (15-18) Exhibitors will participate in age-based Skillathons. This study guide includes all topics an exhibitor might be tested on. Youth will only
Broiler Management for Birds Grown to Low Kill Weights ( lb / kg)
") Broiler Management for Birds Grown to Low Kill Weights (3.3-4.0 lb / 1.5-1.8 kg) April 2008 Michael Garden, Regional Technical Manager Turkey, Middle East & Africa, Aviagen Robin Singleton, Technical Service
Broiler Management for Birds Grown to Low Kill Weights (3.3-4.0 lb / 1.5-1.8 kg) April 2008 Michael Garden, Regional Technical Manager Turkey, Middle East & Africa, Aviagen Robin Singleton, Technical Service
The Effect of Full-Spectrum Fluorescent Lighting on Reproductive Traits of Caged Turkey Hens 1 ' 2
 The Effect of Full-Spectrum Fluorescent Lighting on Reproductive Traits of Caged Turkey Hens 1 ' 2 T. D. SIOPES Department of Poultry Science, North Carolina State University, Raleigh, North Carolina 27695-7608
The Effect of Full-Spectrum Fluorescent Lighting on Reproductive Traits of Caged Turkey Hens 1 ' 2 T. D. SIOPES Department of Poultry Science, North Carolina State University, Raleigh, North Carolina 27695-7608
4-H Poultry: Unit 1. The Egg Flock For an egg-producing flock, select one of these birds: production-type Rhode Island Red Leghorn hybrids sex-link
 4-H Poultry: Unit 1 A small flock of chickens does not require much space or money, and if you manage your flock well, you can earn some money. You might start with a small flock of 25 50 chickens, or
4-H Poultry: Unit 1 A small flock of chickens does not require much space or money, and if you manage your flock well, you can earn some money. You might start with a small flock of 25 50 chickens, or
ROSS TECH 07/46 Managing the Ross 708 Parent Stock Female
 ECH ROSS TECH 07/46 Managing the Ross 708 Parent Stock Female 1 Aviagen provides customers with detailed Product Performance Specifications, Management Manuals and Nutrition Specifications as the basis
ECH ROSS TECH 07/46 Managing the Ross 708 Parent Stock Female 1 Aviagen provides customers with detailed Product Performance Specifications, Management Manuals and Nutrition Specifications as the basis
CIWF Response to the Coalition for Sustainable Egg Supply Study April 2015
 CIWF Response to the Coalition for Sustainable Egg Supply Study April 2015 The Coalition for Sustainable Egg Supply study seeks to understand the sustainability impacts of three laying hen housing systems
CIWF Response to the Coalition for Sustainable Egg Supply Study April 2015 The Coalition for Sustainable Egg Supply study seeks to understand the sustainability impacts of three laying hen housing systems
Recommended Resources: The following resources may be useful in teaching
 Unit C: Poultry Management Lesson 2: Feeding, Management and Equipment for Poultry Student Learning Objectives: Instruction in this lesson should result in students achieving the following objectives:
Unit C: Poultry Management Lesson 2: Feeding, Management and Equipment for Poultry Student Learning Objectives: Instruction in this lesson should result in students achieving the following objectives:
INFO SHEET. Cull Eggs: What To Expect And How To Reduce The Incidence.
 INFO SHEET Cull Eggs: What To Expect And How To Reduce The Incidence info.hybrid@hendrix-genetics.com www.hybridturkeys.com Introduction Over the years, several Hybrid customers have inquired about the
INFO SHEET Cull Eggs: What To Expect And How To Reduce The Incidence info.hybrid@hendrix-genetics.com www.hybridturkeys.com Introduction Over the years, several Hybrid customers have inquired about the
Section 6. Embryonic Development and Hatchery Management Notes
 Section 6 Embryonic Development and Hatchery Management Notes Slide 2 A well run hatchery is critical for any integrated poultry company whether it be a primary breeder company or a commercial meat company.
Section 6 Embryonic Development and Hatchery Management Notes Slide 2 A well run hatchery is critical for any integrated poultry company whether it be a primary breeder company or a commercial meat company.
Factors Influencing Egg Production
 June, 1930 Research Bulletin No. 129 Factors Influencing Egg Production II. The Influence of the Date of First Egg Upon Maturity and Production By C. W. KNOX AGRICULTURAL EXPERIMENT STATION IOWA STATE
June, 1930 Research Bulletin No. 129 Factors Influencing Egg Production II. The Influence of the Date of First Egg Upon Maturity and Production By C. W. KNOX AGRICULTURAL EXPERIMENT STATION IOWA STATE
A Geneticist s Perspective from Within a Broiler Primary Breeder Company
 A Geneticist s Perspective from Within a Broiler Primary Breeder Company DAVID L. POLLOCK1 Primary Breeder Division, Perdue Farms, Salisbury, Maryland 21802 ABSTRACT Recent trends occurring in broiler
A Geneticist s Perspective from Within a Broiler Primary Breeder Company DAVID L. POLLOCK1 Primary Breeder Division, Perdue Farms, Salisbury, Maryland 21802 ABSTRACT Recent trends occurring in broiler
EMBRYO DIAGNOSIS AN IMPORTANT TOOL TO HELP THE HATCHERY MANAGER
 Issue No.14 / September 2007 EMBRYO DIAGNOSIS AN IMPORTANT TOOL TO HELP THE HATCHERY MANAGER By Avian Business Unit CEVA Santé Animale Libourne, France INTRODUCTION Chick quality is the first criterion
Issue No.14 / September 2007 EMBRYO DIAGNOSIS AN IMPORTANT TOOL TO HELP THE HATCHERY MANAGER By Avian Business Unit CEVA Santé Animale Libourne, France INTRODUCTION Chick quality is the first criterion
STATE FFA POULTRY EVALUATION KNOWLEDGE TEST
 2009-2010 STATE FFA POULTRY EVALUATION KNOWLEDGE TEST Contestant Number: Name: Chapter Name: Score: Directions: Read each question carefully and choose the best possible answer. If provided with Scantron
2009-2010 STATE FFA POULTRY EVALUATION KNOWLEDGE TEST Contestant Number: Name: Chapter Name: Score: Directions: Read each question carefully and choose the best possible answer. If provided with Scantron
Sand and Sage Round-Up MARKET CHICKEN STUDY GUIDE Junior and Intermediate Division (8-13 years of age as of December 31)
") Questions will come from: Colorado 4-H Ethical Care of Poultry handout Meat Quality Assurance Market Chicken Study Guide Sand and Sage Round-Up MARKET CHICKEN STUDY GUIDE Junior and Intermediate Division
Questions will come from: Colorado 4-H Ethical Care of Poultry handout Meat Quality Assurance Market Chicken Study Guide Sand and Sage Round-Up MARKET CHICKEN STUDY GUIDE Junior and Intermediate Division
Unit A: Introduction to Poultry Science. Lesson 1: Exploring the Poultry Industry
 Unit A: Introduction to Poultry Science Lesson 1: Exploring the Poultry Industry 1 Terms Broilers Chick Cockerels Drake Duckling Gander Goose Gosling Hen Layers Poult Poultry Pullet Producers Pullets Roosters
Unit A: Introduction to Poultry Science Lesson 1: Exploring the Poultry Industry 1 Terms Broilers Chick Cockerels Drake Duckling Gander Goose Gosling Hen Layers Poult Poultry Pullet Producers Pullets Roosters
THE INTERACTIONS BETWEEN GENOTYPES AND HOUSING ENVIRONMENTS IN THE DOMESTIC HEN
 THE INTERACTIONS BETWEEN GENOTYPES AND HOUSING ENVIRONMENTS IN THE DOMESTIC HEN B Y G. M C B RIDE * Summary A study was made of the egg production and egg weight of a flock of Australorp pullets in which
THE INTERACTIONS BETWEEN GENOTYPES AND HOUSING ENVIRONMENTS IN THE DOMESTIC HEN B Y G. M C B RIDE * Summary A study was made of the egg production and egg weight of a flock of Australorp pullets in which
Production Basics How Do I Raise Poultry for Eggs?
 Production Basics How Do I Raise Poultry for Eggs? C H U C K S C H U S T E R U N I V E R S I T Y O F M A R Y L A N D E X T E N S I O N C E N T R A L M A R Y L A N D C F S @ U M D. E D U J E S S I E F L
Production Basics How Do I Raise Poultry for Eggs? C H U C K S C H U S T E R U N I V E R S I T Y O F M A R Y L A N D E X T E N S I O N C E N T R A L M A R Y L A N D C F S @ U M D. E D U J E S S I E F L
Some Problems Concerning the Development of a Poultry Meat Industry in Australia
 Some Problems Concerning the Development of a Poultry Meat Industry in Australia by Fred. SKALLER* INTRODUCTION Poultry meat can be supplied either from culled laying birds, a by-product of the egg industry,
Some Problems Concerning the Development of a Poultry Meat Industry in Australia by Fred. SKALLER* INTRODUCTION Poultry meat can be supplied either from culled laying birds, a by-product of the egg industry,
Effects of Three Lighting Programs During Grow on the Performance of Commercial Egg Laying Varieties
 Effects of Three Lighting Programs During Grow on the Performance of Commercial Egg Laying Varieties 2. Laying Period Egg Production J. Arango, P. Settar, S. Saxena, J. Arthur, N.P. O Sullivan Hy-Line
Effects of Three Lighting Programs During Grow on the Performance of Commercial Egg Laying Varieties 2. Laying Period Egg Production J. Arango, P. Settar, S. Saxena, J. Arthur, N.P. O Sullivan Hy-Line
EFFECTS OF BODY WEIGHT UNIFORMITY AND PRE-PEAK FEEDING PROGRAMS ON BROILER BREEDER HEN PERFORMANCE
 2001 Poultry Science Association, Inc. EFFECTS OF BODY WEIGHT UNIFORMITY AND PRE-PEAK FEEDING PROGRAMS ON BROILER BREEDER HEN PERFORMANCE B. P. HUDSON, R. J. LIEN 1, and J. B. HESS Department of Poultry
2001 Poultry Science Association, Inc. EFFECTS OF BODY WEIGHT UNIFORMITY AND PRE-PEAK FEEDING PROGRAMS ON BROILER BREEDER HEN PERFORMANCE B. P. HUDSON, R. J. LIEN 1, and J. B. HESS Department of Poultry
A Guide to Commercial Poultry Production in Florida 1
 A Guide to Commercial Poultry Production in Florida 1 Carrol Douglas 2 FACTORS IN PLANNING A POULTRY OPERATION Independent or Contract Production (1) Independent egg producers have the total responsibility
A Guide to Commercial Poultry Production in Florida 1 Carrol Douglas 2 FACTORS IN PLANNING A POULTRY OPERATION Independent or Contract Production (1) Independent egg producers have the total responsibility
AviagenBrief. Spiking Programs to Improve Fertility. Summary. November 2010
 AviagenBrief November 2010 Spiking Programs to Improve Fertility North American Technical Team This article has been written specifically for poultry producers in North America. The advice provided is
AviagenBrief November 2010 Spiking Programs to Improve Fertility North American Technical Team This article has been written specifically for poultry producers in North America. The advice provided is
Reproductive physiology and eggs
 Reproductive physiology and eggs Class Business Reading for this lecture Required. Gill: Chapter 14 1. Reproductive physiology In lecture I will only have time to go over reproductive physiology briefly,
Reproductive physiology and eggs Class Business Reading for this lecture Required. Gill: Chapter 14 1. Reproductive physiology In lecture I will only have time to go over reproductive physiology briefly,
PHYSIOLOGIC AND GENETIC STUDIES OF CROOKED KEELS IN CHICKENS
 PHYSIOLOGIC AND GENETIC STUDIES OF CROOKED KEELS IN CHICKENS SUMMARY 1. In most instances, birds developing crooked keels will do so before maturity. Most of the keel deformities appear between the 6-
PHYSIOLOGIC AND GENETIC STUDIES OF CROOKED KEELS IN CHICKENS SUMMARY 1. In most instances, birds developing crooked keels will do so before maturity. Most of the keel deformities appear between the 6-
Agricultural Extensi?n Se:;ice University of Californi County of Orange
 Agricultural Extensi?n Se:;ice University of Californi County of Orange I 0 Pagel Poultry 1954 INTRODUCTION This is the first annual report of the current Orange County Poultry Management Study. This study
Agricultural Extensi?n Se:;ice University of Californi County of Orange I 0 Pagel Poultry 1954 INTRODUCTION This is the first annual report of the current Orange County Poultry Management Study. This study
UNIVERSITY OF KWAZULU-NATAL SCHOOL OF AGRICULTURAL SCIENCES & AGRIBUSINESS DISCIPLINE OF ANIMAL AND POULTRY SCIENCE EXAMINATIONS: NOVEMBER 2010
 DURATION: 3 HOURS TOTAL MARKS: 100 External Examiner: Prof M Chimonyo Internal Examiner: Dr NC Tyler STUDENTS ARE REQUESTED, IN THEIR OWN INTERESTS, TO WRITE LEGIBLY NOTE: THIS PAPER CONSISTS OF FIVE (5)
DURATION: 3 HOURS TOTAL MARKS: 100 External Examiner: Prof M Chimonyo Internal Examiner: Dr NC Tyler STUDENTS ARE REQUESTED, IN THEIR OWN INTERESTS, TO WRITE LEGIBLY NOTE: THIS PAPER CONSISTS OF FIVE (5)
SHEEP SIRE REFERENCING SCHEMES - NEW OPPORTUNITIES FOR PEDIGREE BREEDERS AND LAMB PRODUCERS a. G. Simm and N.R. Wray
 SHEEP SIRE REFERENCING SCHEMES - NEW OPPORTUNITIES FOR PEDIGREE BREEDERS AND LAMB PRODUCERS a G. Simm and N.R. Wray The Scottish Agricultural College Edinburgh, Scotland Summary Sire referencing schemes
SHEEP SIRE REFERENCING SCHEMES - NEW OPPORTUNITIES FOR PEDIGREE BREEDERS AND LAMB PRODUCERS a G. Simm and N.R. Wray The Scottish Agricultural College Edinburgh, Scotland Summary Sire referencing schemes
Optimum broiler development. A practical guide to ensure correct early broiler performance
 Optimum broiler development A practical guide to ensure correct early broiler performance 1.1. Why focus on incubation? Today s meat yield birds can produce higher embryonic temperatures and the risk of
Optimum broiler development A practical guide to ensure correct early broiler performance 1.1. Why focus on incubation? Today s meat yield birds can produce higher embryonic temperatures and the risk of
Reproduction in Turkey Hens as Influenced by Prebreeder and Breeder Protein Intake and the Environment
 Reproduction in Turkey Hens as Influenced by and Protein Intake and the Environment F. L. CHERMS, M. G. STOLLERAND J. J. MACILRAITH Nicholas Turkey Breeding Farms, Inc., Sonoma, California 95476 AND H.
Reproduction in Turkey Hens as Influenced by and Protein Intake and the Environment F. L. CHERMS, M. G. STOLLERAND J. J. MACILRAITH Nicholas Turkey Breeding Farms, Inc., Sonoma, California 95476 AND H.
RELATIONSHIPS AMONG WEIGHTS AND CALVING PERFORMANCE OF HEIFERS IN A HERD OF UNSELECTED CATTLE
 RELATIONSHIPS AMONG WEIGHTS AND CALVING PERFORMANCE OF HEIFERS IN A HERD OF UNSELECTED CATTLE T. C. NELSEN, R. E. SHORT, J. J. URICK and W. L. REYNOLDS1, USA SUMMARY Two important traits of a productive
RELATIONSHIPS AMONG WEIGHTS AND CALVING PERFORMANCE OF HEIFERS IN A HERD OF UNSELECTED CATTLE T. C. NELSEN, R. E. SHORT, J. J. URICK and W. L. REYNOLDS1, USA SUMMARY Two important traits of a productive
Genotypic and phenotypic relationships between gain, feed efficiency and backfat probe in swine
 Retrospective Theses and Dissertations 1970 Genotypic and phenotypic relationships between gain, feed efficiency and backfat probe in swine Ronald Neal Lindvall Iowa State University Follow this and additional
Retrospective Theses and Dissertations 1970 Genotypic and phenotypic relationships between gain, feed efficiency and backfat probe in swine Ronald Neal Lindvall Iowa State University Follow this and additional
Effect of Region and Stocking Density on Performance of Farm Ostriches. Mehrdad Bouyeh
 Effect of Region and Stocking Density on Performance of Farm Ostriches Mehrdad Bouyeh Department of Animal Science. Islamic Azad University Rasht branch.rasht, Iran E-mail: mbouyeh@gmail.com- booyeh@iaurasht.ac.ir
Effect of Region and Stocking Density on Performance of Farm Ostriches Mehrdad Bouyeh Department of Animal Science. Islamic Azad University Rasht branch.rasht, Iran E-mail: mbouyeh@gmail.com- booyeh@iaurasht.ac.ir
Right and next page: Brahma chicks with decent footfeathering, but with no fluff on the inner side of the legs and on the inner toes.
 FOOTFEATHERING By: Bobo Athes For the vast majority of chicken breeds, especially for the utility breeds, footfeathering is not included in the standard. Yet, in the case of ornamental breeds, it is a
FOOTFEATHERING By: Bobo Athes For the vast majority of chicken breeds, especially for the utility breeds, footfeathering is not included in the standard. Yet, in the case of ornamental breeds, it is a
CHAPTER3. Materials and methods
 CHAPTER3 Materials and methods 3.1 Experimental Site and Housing The study was conducted at the Animal Production Institute of the Agricultural Research Council (ARC) Irene, in Gauteng Province of South
CHAPTER3 Materials and methods 3.1 Experimental Site and Housing The study was conducted at the Animal Production Institute of the Agricultural Research Council (ARC) Irene, in Gauteng Province of South
206 Adopted: 4 April 1984
 OECD GUIDELINE FOR TESTING OF CHEMICALS 206 Adopted: 4 April 1984 1. I N T R O D U C T O R Y I N F O R M A T I O N P r e r e q u i s i t e s Water solubility Vapour pressure Avian dietary LC50 (See Test
OECD GUIDELINE FOR TESTING OF CHEMICALS 206 Adopted: 4 April 1984 1. I N T R O D U C T O R Y I N F O R M A T I O N P r e r e q u i s i t e s Water solubility Vapour pressure Avian dietary LC50 (See Test
2018 HY-LINE BROWN SCHOOL EGG LAYING COMPETITION INFORMATION BOOKLET. Proudly supported by
 2018 HY-LINE BROWN SCHOOL EGG LAYING COMPETITION INFORMATION BOOKLET Proudly supported by WELCOME On behalf of the RNA Poultry, Pigeons, Birds and Eggs Committee, I would like to welcome your school to
2018 HY-LINE BROWN SCHOOL EGG LAYING COMPETITION INFORMATION BOOKLET Proudly supported by WELCOME On behalf of the RNA Poultry, Pigeons, Birds and Eggs Committee, I would like to welcome your school to
PRODUCTION BASICS HOW DO I RAISE POULTRY FOR MEAT? Chuck Schuster University of Maryland Extension Central Maryland
 PRODUCTION BASICS HOW DO I RAISE POULTRY FOR MEAT? Chuck Schuster University of Maryland Extension Central Maryland cfs@umd.edu Jessie Flores University of Maryland Extension Lower Eastern Shore Maryland
PRODUCTION BASICS HOW DO I RAISE POULTRY FOR MEAT? Chuck Schuster University of Maryland Extension Central Maryland cfs@umd.edu Jessie Flores University of Maryland Extension Lower Eastern Shore Maryland
Genetic evaluation for egg production traits in Japanese quail.
 BENHA VETERINARY MEDICAL JOURNAL, VOL. 8, NO. 1:178 185, MARCH 015 Genetic evaluation for egg production traits in Japanese quail. Eman A. Manaa 1, Khairy M. ElBayomi, Gamal A. Sosa 3 1 Department of animal
BENHA VETERINARY MEDICAL JOURNAL, VOL. 8, NO. 1:178 185, MARCH 015 Genetic evaluation for egg production traits in Japanese quail. Eman A. Manaa 1, Khairy M. ElBayomi, Gamal A. Sosa 3 1 Department of animal
Factors Affecting Breast Meat Yield in Turkeys
 Management Article The premier supplier of turkey breeding stock worldwide CP01 Version 2 Factors Affecting Breast Meat Yield in Turkeys Aviagen Turkeys Ltd Introduction Breast meat, in the majority of
Management Article The premier supplier of turkey breeding stock worldwide CP01 Version 2 Factors Affecting Breast Meat Yield in Turkeys Aviagen Turkeys Ltd Introduction Breast meat, in the majority of
Local Grains and Free-Choice Feeding of Organic Layer Hens on Pasture at UBC Farm Introduction
 Local Grains and Free-Choice Feeding of Organic Layer Hens on Pasture at UBC Farm Darin C. Bennett, Avian Research Centre, Jacob Slosberg, Centre for Sustainable Food Systems, Faculty of Land Food Systems,
Local Grains and Free-Choice Feeding of Organic Layer Hens on Pasture at UBC Farm Darin C. Bennett, Avian Research Centre, Jacob Slosberg, Centre for Sustainable Food Systems, Faculty of Land Food Systems,
Hatchability and Early Chick Growth Potential of Broiler Breeder Eggs with Hairline Cracks
 2004 Poultry Science Association, Inc. Hatchability and Early Chick Growth Potential of Broiler Breeder Eggs with Hairline Cracks D. M. Barnett, B. L. Kumpula, R. L. Petryk, N. A. Robinson, R. A. Renema,
2004 Poultry Science Association, Inc. Hatchability and Early Chick Growth Potential of Broiler Breeder Eggs with Hairline Cracks D. M. Barnett, B. L. Kumpula, R. L. Petryk, N. A. Robinson, R. A. Renema,
Madison County Fair Broiler Showmanship Study Questions
 Broilers Madison County Fair Broiler Showmanship Study Questions 1. What breed are the birds you showed in the fair? My show broilers were Cornish Rock crosses. The commercial broiler meat industry primarily
Broilers Madison County Fair Broiler Showmanship Study Questions 1. What breed are the birds you showed in the fair? My show broilers were Cornish Rock crosses. The commercial broiler meat industry primarily
A-l. Students shall examine the circulatory and respiratory systems of animals.
 Animal Science A-l. Students shall examine the circulatory and respiratory systems of animals. 1. Discuss the pathway of blood through the heart and circulatory system. 2. Describe and compare the functions
Animal Science A-l. Students shall examine the circulatory and respiratory systems of animals. 1. Discuss the pathway of blood through the heart and circulatory system. 2. Describe and compare the functions
Broiler production introduction. Placement of chicks
 BROILER GUIDE Broiler production introduction Broilers are chickens that are specifically bred and raised for their meat. The goal for any broiler farmer is to successfully raise a day old chick to a market
BROILER GUIDE Broiler production introduction Broilers are chickens that are specifically bred and raised for their meat. The goal for any broiler farmer is to successfully raise a day old chick to a market
Best Practice in the Breeder House
 Best Practice in the Breeder House Preventing Floor Eggs Best Practice in the Breeder House Preventing Floor Eggs Why are floor eggs a problem? Eggs laid on the floor (floor eggs) have a significantly
Best Practice in the Breeder House Preventing Floor Eggs Best Practice in the Breeder House Preventing Floor Eggs Why are floor eggs a problem? Eggs laid on the floor (floor eggs) have a significantly