SCIENCE & RESEARCH INTERNAL REPORT NO.129 KAKAPO BREEDING ACTIVITY ON LITTLE BARRIER ISLAND: OCTOBER 1990 JUNE B.D. Lloyd and R.G.
|
|
|
- Percival Fleming
- 5 years ago
- Views:
Transcription
1 SCIENCE & RESEARCH INTERNAL REPORT NO.129 KAKAPO BREEDING ACTIVITY ON LITTLE BARRIER ISLAND: OCTOBER 1990 JUNE 1991 by B.D. Lloyd and R.G. Powlesland This is an internal Department of Conservation report and must be cited as Science & Research Internal Report No Permission to use any of its contents must be obtained from the Director (Science & Research), Head Office, Department of Conservsation. Published by Head Office, Department of Conservation, P 0 Box , Wellington New Zealand ISSN ISBN
2 August 1992, Department of Conservation Keywords: kakapo, Strigops habroptilus, Little Barrier Island, breeding, booming, nesting
3 CONTENTS ABSTRACT 1 1. INTRODUCTION 2 2. METHODS Male breeding activity Monitoring of disturbance at booming sites Monitoring booming Identifying males at booming sites Feather clusters at booming sites Feather counts Female breeding activity Monitoring female movements using radiotelemetry Monitoring nesting Rodent control around nests Supplementary feeding Rat index trapping 5 3. RESULTS Male breeding activity Monitoring disturbance at booming sites Monitoring booming using tape recorders Booming heard at night Daytime booming Identifying males at booming sites Feather clusters Feather counts Female breeding activity Summary Breeding details for each female 12 Maggie 12 Heather 13 Wendy 14 John-Girl 16 Bella-rose Nest locations and descriptions Nest Attendance Incubation temperatures Egg data Period of rapid yolk deposition Chick Development Sexing chicks Potential nest problems Supplementary feeding Rat index trapping 32
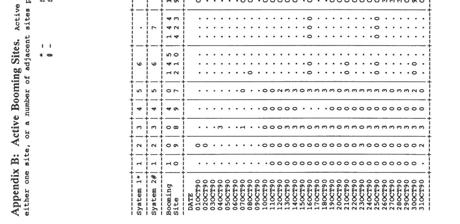
4 4. DISCUSSION Summary of breeding activity on Little Barrier Island A comparison of breeding on Little Barrier Island and Stewart Island Kakapo productivity Male breeding success CONCLUSIONS RECOMMENDATIONS 37 ACKNOWLEDGEMENTS 38 REFERENCES 39 Appendix A: Capture details for Appendix B: Active Booming Sites 42 Appendix C: Kakapo chick data (Little Barrier Island ), estimated age (days after hatching) and weight (g) 49
5 KAKAPO BREEDING ACTIVITY ON LITTLE BARRIER ISLAND: OCTOBER 1990 JUNE 1991 by B.D. Lloyd and R.G. Powlesland Science and Research Division, Department of Conservation, P.O. Box , Wellington ABSTRACT Breeding activity by kakapo on Little Barrier Island was monitored throughout the period October 1990 to June Booming began in early October, two months earlier than previously recorded, and continued until mid-april. At the height of the booming season ten males were booming. Four of the five females taking supplementary food nested during late January. Two of the nests were successful, each fledging a single chick in late May. Both fledglings survived to independence. 1
6 1. INTRODUCTION Thirteen male and nine female kakapo were transferred to Little Barrier Island between May and August One male was found dead in Male courtship displays, called booming, occurred on Little Barrier Island during five of the seven summers ( ) following the transfers, but we suspect that nesting did not occur during this period as there was no evidence of copulation at the males' booming sites (i.e. display sites, also known as track and bowl systems), and systematic surveys during the winters of 1986 and 1989, using bait lines, cage traps and trained dogs, failed to identify recruitment to the original translocated population (Veitch 1986 and Hodsell 1989). In September 1989 a programme to promote breeding by providing dietary supplements to free-living kakapo on the island was begun. During that summer (December 1989 to April 1990) male kakapo displays were the most intense and extended ever recorded, and the first two kakapo nests recorded on the island were discovered (Lloyd and Powlesland 1990). This report describes the results of a programme to monitor breeding behaviour of kakapo on Little Barrier Island during the period October 1990 to June METHODS 2.1. Male breeding activity The methods used to monitor male kakapo courtship behaviour at booming sites on Little Barrier Island have been described by Moorhouse 1986, Handford 1987, Dowding 1988, and Greene During this study these methods were used with the modifications described in Lloyd and Powlesland Monitoring of disturbance at booming sites All known booming sites were inspected repeatedly throughout the period 10 October 1990 to 19 April The regularity of inspection varied from daily, for conveniently located active sites, to monthly, for remote or inactive sites. Any kakapo sign seen at, or near, booming sites was noted, e.g. grubbing, trimming of vegetation, feeding sign, droppings or feathers. Four short (5-10 cm) upright sticks and two crossed sticks were placed in each bowl at a booming site, and subsequent disturbance to them was recorded as evidence of courtship behaviour. Details of any booming heard were also recorded Monitoring booming Small voice-activated tape recorders were left at booming sites overnight. Olympus miniature tape recorders (models L100, L200 and S930) were used with C60 tapes. A tape recorder was wrapped in two small plastic bags, switched to voice activated mode and placed in a small hole (usually dug in a peat bank) within 200 mm of a bowl. Recordings were made at a tape speed of 1.2 cm/sec in order to extend the tape duration. The tapes were played back at 2.4 cm/sec, to both speed up the playback process and make booming audible on the tape recorder's small loudspeakers. Any sounds which could be attributed to kakapo (i.e. booming, skraaking and chinging calls, and grubbing noises) were noted. A description of these calls can be found in Powlesland et al. in press. 2
7 2.1.3 Identifying males at booming sites Males were identified either by catching them at night in cage traps sited near active booming sites, or during the day by using trained dogs to track them from their booming sites to daytime roosts. Males with patterns painted on the stainless steel leg band could be identified without handling, otherwise it was necessary to handle the birds to read the band number. Identifications were undertaken at the beginning and end of the booming season to avoid interfering with actual courtship Feather clusters at booming sites Clusters of kakapo feathers found at booming sites may be the result of either copulation Powlesland (1989) or fighting. Whenever we visited booming sites we therefore examined the surrounding area for feathers. Clusters of feathers considered to be the result of copulation attempts were those in which feathers were restricted to a small area (<1 m 2 ), close to an active bowl (<5 m); the feathers were mixed into the substrate and were primarily down feathers. In contrast, clusters of feathers considered to be the result of fighting were scattered over several metres; they were usually more than five metres from active bowls; they were not mixed into the substrate and many were contour feathers Feather counts The daily schedule of supplementary feeding and booming site inspection provided a method to monitor seasonal changes in feather loss. All kakapo feathers found in the course of our work were collected and recorded. 2.2 Female breeding activity Monitoring female movements using radiotelemetry Four females were fitted with miniature radio-transmitters (supplied by Sirtrack NZ and weighing c. 30 g each) during October and November 1990, prior to this breeding season. A fifth female was caught at a nest while brooding chicks, and radiotagged on 31 March Capture details are in Appendix A. The locations of radiotagged females were monitored remotely by radio-triangulation every few days. This allowed us to identify movements to courtship areas, copulation and nesting Monitoring nesting When nesting was suspected the radiotagged females were approached both to confirm the nesting attempt and to locate the nest for intensive monitoring. Three nests belonging to radiotagged females were located during daylight when the females were sitting. Subsequent inspections were made at night when the female was off the nest. In order to do this we arrived c.100 m from a nest just before dusk and monitored the direction and strength of the female's radio-signal until she was well away from the nest. A fourth nest, which belonged to a female without a radio-transmitter, was located using a trained dog. This nest was regularly inspected during daylight with the female present until mid-way through brooding when she was radiotagged. Subsequent nest inspections were made at night as with other nests. 3
8 Nests were generally inspected four or five times during the incubation period. During the first half of the brood period nests were inspected at three day intervals and during the latter half of the brood period nests were inspected at intervals of four to five days. Egg Data Eggs were candled at the nest. Weight, length and maximum diameter were recorded. Determining egg temperatures Incubation temperatures were obtained from two nests using a temperature sensitive artificial egg. A temperature sensitive transmitter (obtained from Sirtrack, NZ) was embedded in paraffin wax in an artificial bantam's egg. The transmitter's pulse period, which varied with temperature, was measured remotely at distances of up to 50 m using a Telonics TDP Data Processor connected to the output from a Telonics TR2 receiver with a standard hand-held Yagi antenna. The system was calibrated by placing the egg in a controlled temperature incubator with a thermometer and recording pulse rates over a range of temperatures. The egg was placed in the nest with the real clutch at night the female was away from the nest. Chick development Because access to one of the two successful nests was difficult, most observations of chicks were made at only one nest. During each inspection the chick was weighed and a written description made of its plumage, mobility, physical co-ordination and vocalisation. Photographs were taken regularly. Tarsus length, culmen length and width were all recorded occasionally. Primary feather development was monitored by recording the lengths, in millimetres, of both the sheathed region and the unfurled vane of the ninth primary from the inside (that is fourth from the outside) of the right wing. The movement of fledglings Miniature radio-transmitters, weighing c. 30 g, were fitted to the chicks a few days before they left the nest. The location of both fledglings and the females was then determined every few days by remote radio-triangulation. 2.3 Rodent control around nests To reduce the possibility of the predation of chicks by kiore (Rattus exulans), rodents control was undertaken around nests. The control method was previously tested around kakapo feeding stations on Little Barrier and is fully described in Powlesland and Lloyd Six covered bait silos (McFadden 1984) were placed at equal distances on a circle of 50 m radius around each nest during the incubation period. Initially non-toxic prebait (green-dyed kibbled maize) was placed in the silos hut after c. 10 days the pre-bait was replaced by toxic bait (0.005% by weight in green-dyed kibbled maize). The toxic bait was replenished regularly until the silos were removed just before the kakapo chicks left the nests. 2.4 Supplementary As part of a concurrent supplementary feeding programme, described in Powlesland and Lloyd (1990, 1992), kakapo were provided with dietary supplements. Feeding stations were placed within the home ranges of the five known females and within 50 m of several active booming sites (Sites 7,8,9 and 21). Evidence of feeding by kakapo at these stations was recorded daily. A full account of this work can be found in Powlesland and Lloyd (1992). 4
9 2.5 Rat index trapping Changes in the abundance of rats were assessed by using a rat index line operated for three consecutive nights at two-month intervals (Lloyd and Powlesland, 1990). Two Ezeset rat raps were placed under aluminium covers at each of 45 sites located at 50 m intervals on Track 20 from approximately 150 m a.s.1. to 650 m a.s.1.. Traps were baited with cheese. 3. RESULTS 3.1 Male breeding activity Monitoring disturbance at booming sites At the end of the breeding season there were 50 known booming sites on Little Barrier Island, including one new site found in December Disturbance attributed to kakapo was detected at 31 of the 50 known sites during the period 1 October 1990 to 30 April The results of monitoring disturbance at the 31 active sites during this period are provided in Appendix B. Figure 1 is a graph of the numbers of systems used by kakapo each night during the period 1 October to 30 April. We estimate that during the booming season up to ten males were active at booming sites during a 190-day period from 1 October 1990 to 8 April 1991, and that there were 1357 active system-nights. In comparison, we estimate that during the season there were 11 to 13 males active during a 140 day period from 21 November 1989 to 10 April 1990, and that there were 964 active system-nights. Activity was first noted on 1 October 1990 during our first inspection of booming sites when one site (Site 21) was found to have been disturbed. By mid-october four systems were regularly active. A fifth system became active towards the end of October. Sometime between 20 November and 3 December a further 5 systems became active, bringing the number of regularly active systems to 10. During the period 3 December 1990 to 1 March 1991 activity was recorded at between 7 and 12 systems each night, the modal value being 10. It is probable that there were only 10 males active, the extra two systems being the result of individuals being active at more than one system in one night. From 1 March onwards the number of active systems declined steadily until by 8 April activity had ceased almost entirely, though there was sporadic activity at three systems between 8 and 19 April. Seasonal changes in the patterns of disturbance observed at booming sites were similar to those described for the season in Lloyd and Powlesland (1990). 5


10 Figure 1. The number of booming systems used each night on Little Barrier Island during the period 1 Oct April The upper boundary to the grey area is the minimum numbers of systems used each night. These numbers comprise results only from those systems inspected on consecutive days. The upper boundary to the black area is the best estimates of the numbers of systems used each night. These estimates were obtained by interpolating between inspections for those systems not inspected on consecutive days. 6
11 3.1.2 Monitoring booming using tape recorders Tape recorders were placed at booming sites during the period 9 October 1990 to 8 April 1991, most commonly at Sites 7, 8 and 21. Tape recorders were placed successfully (i.e without malfunction) close to an active bowl on 60 occasions, and kakapo vocalisations were recorded on 47 occasions. Booming was first recorded at Site 21 during October. Initially on 9 and 11 October booming was soft and intermittent but on 21 October intense and persistent booming was recorded. Skraaking but not booming was recorded at Site 7 on 17 and 18 November. Skraaking with some booming was recorded at Site 8 on 28 October but it was not until 23 November that strong and persistent booming was recorded from sites other than Site 21. From 23 November 1990 to 9 March 1991 strong and persistent booming was recorded on 33 of 35 recordings made at active sites. During this period short bursts of skraaking in the middle of booming were recorded on three occasions and chinging was recorded once (at Site 24 on 7 January 1991). Booming seemed to subside during mid-march; the last booming recorded (occasional short bouts) being on 27 March at Site Booming heard at night 24 February: An observer 50 m south of Site 7 saw a kakapo go past at Subsequently booming was heard from both Sites 7 and 8 from to Skraaking and one bout of chinging was heard from Site February: An observer 50 m south of Site 7 heard several males booming from to It seemed as if one male boomed at Site to and then moved to Site 7 where it boomed for the rest of the night. 5 March: An observer 100 m east of Site 24 heard booming begin at Site 24 at Booming was still strong at There was no activity at Site 24 after March: An observer 100 m east of Site 24 heard skraaking between There was intermittent booming from surrounding booming sites from onwards all night Daytime booming Booming was heard during the day by observers inspecting booming sites and replenishing feeding stations. It may have been prompted by observer disturbance. Daytime booming generally occurred close to active booming sites, was subdued and lasted only a few minutes. It was heard on 43 occasions on a total of 32 days over a 132 day period lasting from 24 November 1990 to 5 April Most occurrences were between Booming Sites 7 and 21 where observer activity was concentrated. 7
12 Table 1. The frequency of occurrence of daytime booming, Fortnight Starting No. of occurrences 20 Nov 3 4 Dec 2 18 Dec 6 1 Jan Jan 7 29 Jan 4 12 Feb 0 26 Feb 4 12 Mar 1 26 Mar 2 8
13 Table 1 shows a pronounced peak in daytime booming frequency during early January, which probably reflects a peak in courtship activity. Daytime booming was heard at all hours between and 18.00; the actual distribution of time of observations probably reflects observer opportunity. It was heard in all weather conditions; including hot, sunny and dry days Identifying males at booming sites During the breeding season five males were caught near booming sites (Table 2). Capture details of all birds caught and handled during the season are in Appendix A. Research on Stewart Island (Powlesland et al. in press) indicates that male kakapo show considerable site fidelity throughout most of a booming season and between seasons, but that at the beginning and towards the end of booming seasons males may move between booming sites Feather clusters Large clusters of feathers were found at active booming sites on seven occasions between 31 October 1990 and 23 February 1991 (Table 3). Three clusters found between 3 and 20 January were interpreted as being conclusive evidence of copulation; a fourth cluster found on 23 January 1991 may also have been the result of a copulation. 9
14 Table 3. Feather cluster details for the period 31 October February October. 50 m north of Site 7: Evidence of a fight. 50 down and contour feathers, some of then damaged, were found in two clusters ten metres apart. The two clusters were scattered over areas of 100x250mm and 50x50 mm. 3 January, Site 24: Evidence of copulation. More than 70 down feathers, large quantity of finely shredded fragments of down, and 10 contour feathers in a patch on the track 4 m west of the bowl. Much of the feather material was ground into the leaf litter. 6 January, Site 21: Not significant 15 down feathers were found caught on a root by the bowl. 14 January. Site 24: Evidence of copulation 35 down feathers and a large quantity of finely shredded fragments of down feathers were found ground into piles of leaf litter on the track 1-2 m east of the bowl. Between these piles of leaves and feathers there was an area, 140x300 mm and 800 mm from the bowl, swept clear of leaves. Heather was roosting near the booming site. 20 January. Site 7: Evidence of copulation. 67 down feathers, a large quantity of finely shredded fragments of down feathers, and 1 contour feather were found in three patches along a section of track from 1 to 4 m south of the bowl. The feather material was mixed with leaf litter. In the largest of the three patches there was a distinct ball of leaf litter and feather material. John-Girl was roosting near the booming site. January 23, Site 49: Possibly a fight but may have been a copulation. 20 down feathers and 76 contour feathers found on the track 2 m from the bowl mixed into the soil and leaf litter. No radiotagged female known to be close by. 23 February. Site 9: Evidence of a fight. A large number of down feathers and 6 contour feathers were found 25 m north of the bowl Feather counts A small number of down and contour feathers were found daily throughout the booming season. In addition large clusters of feathers were found on several occasions as described in the preceding section. From mid-march onwards there was a marked increase in the numbers of feathers (both down and contour) found on the tracks near booming sites. Most feathers were found individually or in small groups but occasionally groups of feathers were found. This increase in the number of feathers found corresponded closely with the decline in booming activity and resulted from the onset of post-nuptial moult. 10
15 3.2 Female breeding activity Summary Four of the five females monitored this season nested during late January 1991 (Maggie, John-Girl, Heather and Wendy). The chronology of the four nests is shown in Table 4 and a summary of estimates of the length of the different stages of nesting is shown in Table 5. The fifth female did not attempt to nest. Two of the four nests were unsuccessful. One of these unsuccessful nests contained a single infertile egg, the other contained one infertile egg and one fertile egg, but the fertile egg died at about 5-7 days, probably as a result of chilling. In an attempt to promote re-nesting we removed these two unsuccessful clutches during early incubation. Re-nesting did not occur. One chick fledged from each of the two successful nests during mid-may. At the time of writing both chicks survive as independent juveniles. A single fertile egg was removed from one of the successful nests during February and taken to Auckland Zoo to be reared artificially. The egg hatched successfully, but the chick died four days later as a result of a combination of digestive and respiratory infections. All five females were feeding at supplementary feeding stations regularly for many months before the breeding season and continued to feed at the stations during the breeding season (Powlesland and Lloyd 1992). Brooding females took large quantities of supplementary food and some water, presumably to moisten food for regurgitation. After fledging the females and at least one of the dependent juveniles fed regularly at supplementary feeding stations. Examination of juveniles's droppings and the discovery of natural feeding sign around their roost sites indicated that the juveniles were taking natural foods as well as the supplementary food. 11
16 3.2.2 Breeding details for each female Maggie Summary Maggie was caught and radiotagged on 24 October She moved towards unoccupied booming sites on 10 January but probably didn't visit an active site and there is no evidence that she copulated. We found her nest on 21 January, but did not inspect the nest until some nights later, while she was absent. There was one infertile egg laid on c. 21 January. On 29 January we replaced this infertile egg with two artificial eggs which we removed two nights later. Maggie abandoned the nest a day later on 1 February. Her location was monitored regularly throughout the rest of the breeding season but there was no evidence of further breeding attempts. Weight Mean weights prior to supplementary feeding = 1.15 kg (n=3 sd=0.200) Weights recorded since supplementary feeding began: 10 Oct kg 24 Oct kg 13 Aug kg 12 Oct kg Previous breeding history During the breeding season Maggie laid one egg which was probably infertile. There was no evidence that she copulated. Supplementary feeding Maggie had been taking supplementary food consistently since late During the period 1-21 January 1991 she fed at the feeding station irregularly: only 13 nights out of 20, compared with 84-97% throughout the preceeding 6 months. From 22 January to 2 February, when Maggie was nesting, she fed on only 3 nights out of 11. After abandoning the nest on 1 February she fed almost nightly. Pre-nesting movements Maggie roosted in the area around her feeding station and subsequent nest site until 9 January Generally she roosted within a hundred metres of the feeding station. On 10 January she roosted below Booming Site 5, on the slopes of Bald Rock, approximately 2 km from her feeding station. On 12 January she was once again close to the feeding station and from then onwards she roosted in the area in which she subsequently nested. Copulation Maggie moved towards the old unused Booming Sites 1-6 between 9 and 11 January but probably did not approach any active sites. She may never have been close enough to any booming males to hear them. There is no indication that she copulated. Nest log 21 January, We found Maggie's nest but did not inspect it. During the next two nights we attempted to visit the nest while Maggie was absent, but remote monitoring (from 100 m away) indicated that Maggie stayed close to her nest and we therefore did not approach. 12
17 23 January, 21.00: While Maggie was off the nest the nest contents were inspected but not handled. There was one egg 1.2 m into the cavity, behind a 30 mm high wall of leaves. 27 January 200: While Maggie was away from the nest the egg was removed from the nest, measured, and candled. The egg was replaced together with a temperature sensor egg. At 2.20 Maggie returned to the nest and resumed incubation. 29 January, 1220: Maggie's egg and the temperature sensitive egg were removed from under her and replaced by two artificial eggs. Maggie immediately resumed brooding. Her egg was placed in a portable incubator at 95 F and flown to Auckland Zoo by helicopter. 30 January: The artificial eggs were removed from her nest at night while she was absent. 31 January: Maggie was still on her nest. 1 February: Maggie was roosting away from her nest. Heather Summary Heather was caught and radiotagged on 26 November She spent five nights (12 to 17 January) close to the active booming sites 7, 8, 9, 49, 21 and 24. She copulated at Site 24 on the night of 13 January, presumably with Bill, who was captured at this site later in the season. Heather laid two eggs sometime during the period January. One of the eggs was infertile and the embryo of the other died at about 5 days, probably as a result of chilling during a severe storm on 2 February. We removed both eggs on 3 February and Heather deserted the nest on 5 February. She did not attempt to breed again during the season. Weight Mean weights prior to supplementary feeding = 1.24 kg (n=13 sd=0.121) Weights recorded since supplementary feeding began: 15 Nov kg 26 Nov kg 11 Sep kg 12 Sep kg 11 Oct kg Previous breeding history During the breeding season Heather nested, but the nest failed when a single nestling died at about six days old during a wet, cold period. Supplementary feeding Heather had been taking supplementary food consistently since late 1989, but it was difficult to be certain when Heather had fed because she used two feeding stations which were both also used by other kakapo. During December 1990 she probably fed nightly at a feeding station close to her subsequent nest site. In early January she fed at this same feeding station irregularly. She ceased feeding during the period January, when she visited males at their booming sites, but returned to feed irregularly during the pre-lay and early incubation periods. On 3 February, when we removed the two eggs from her nest, she resumed feeding nightly, probably moving between two nearby feeding stations. 13
18 Pre-nesting movements During the period 1-8 January Heather roosted in the area around her feeding stations. On 10 January she roosted near the subsequent nest site. From 12 to 17 January she roosted in the vicinity of active booming sites: 12 January 30 m east of Site 8, 13 January close to Site 7, 14 January 10 m from Site 24, 15 January close to Site 49, 17 January 100 m below Site 49. On 18 and 20 January she had returned to Feeding Station 9 and roosted close by. From 21 January until the nest was abandoned on 5 February she roosted at her nest site. Copulation Heather spent five nights (12 to 17 January) close to the active Booming Sites 7, 8, 9, 49, 21 and 24. She copulated at Site 24 on the night of 13 January, presumably with Bill, who was captured in a cage trap while still booming at this site on 6 April. Nest Log (Laying date: January) 24 January, midday: We approached to within 5 m of the nest but did not inspect it. Heather growled as we approached. 28 January, We inspected the nest contents. Initially we used a fibrescope but Heather attacked it, she then moved to one side revealing two eggs. 31 January, February 2.30: We monitored Heather's movements from about 70 m away. Heather stayed on, or close to the nest throughout. We approached to within 20 m of the nest briefly to confirm her location. 3 February, 14.2: The two eggs were removed and candled; one was infertile, the other was fertile but had died 18 to 24 hours earlier on 2 February (a cold squall and a violent thunderstorm occurred at about that time). Both eggs were placed in a portable incubator at 95 F and transferred to Auckland Zoo by helicopter. 4 January: Heather was roosting in the nest 5 January: Heather was roosting away from the nest. Wendy Summary Wendy's nest was found by a trained dog on 25 January Wendy was not radiotagged until 31 March when she was caught by us on the nest and radiotagged. Because she was not radiotagged early in the season we can not be sure of the details of copulation but we suspect she copulated at Site 24 on the night of 2 January with Bill. Wendy laid three eggs, probably during the period January. Two eggs hatched c. 21 February, but the third failed to hatch and we removed it on 17 March. One chick aged c. 30 days disappeared between 22 and 25 March, when there was a period of heavy rain. The other chick (subsequently named Dobby) left the nest around 29 April approximately 68 days after hatching. Before Dobby fledged he was banded and tagged. Dobby remained in or close to his natal home-range and was observed both being fed by Wendy, and feeding from a feeding station with Wendy, during September 1991, four months after leaving the nest. 14
19 Weight Mean weights prior to supplementary feeding = 1.31 kg (n=10 sd=0.152) Weights recorded since supplementary feeding began: 14 Oct kg 31 Mar kg 14 Jul kg 06 Oct kg Previous breeding history There is no information on whether Wendy attempted to breed in the breeding season. Supplementary feeding We can not be sure whether Wendy was taking supplementary food before 31 March, when she was radio-tagged, but we believe that she was taking supplementary food irregularly through the period December to mid-february and then regularly from the final week of incubation (c. 15 February) onwards. The quantities of food taken increased for the first few weeks of chick rearing until mid-march, when one of the two chicks died; quantities then declined. Water was taken regularly from mid-april, soon after it was first provided at her feeding station. Presumably the water moistened food for regurgitation to her chick. Pre-nest movements There is no reliable information on her movements before nesting as she was not radiotagged until the middle of the nest period. Copulation details Copulation feathers were found when there was no radiotagged female nearby on two occasions: 3 January at Booming Site 24, and 23 January at Site 49. The copulation on 3 January fits Wendy's nest chronology better than the later copulation but would have entailed an unusually long pre-lay period of days. The male at Site 24 was probably Bill, captured there in a cage trap while still booming on 6 April. Site 49 was used only occasionally during the season. The reduction in activity found at the closest booming site (No. 7) on 23 January indicates that the male at this site (probably Pegasus) may have moved temporarily to Site 49. Nest log Incubation Five nest visits undertaken by us before hatching were all made during the daytime while Wendy was on the nest: 25 Jan: Wendy and the nest were found. No eggs were visible. 26 Jan: Nest inspected; no eggs were visible but there was a dead petrel in the nest. 27 Jan: Nest inspected with a fibrescope; one egg was visible. 29 Jan: Nest inspected with a fibrescope; two eggs visible, we removed the dead petrel. 31 Jan: Nest inspected with fibrescope; the female sat tight and we were unable to see eggs. 15
20 Brooding (Hatching date: c. 21 February; Fledging date: 29 April, Day 67 Chick name: Dobby) Until 31 March (Day 38) the nest was visited in daylight at approximately three day intervals. From 31 March (when Wendy was radiotagged) the nest was visited at night at intervals of 4-5 days. Because the chick was difficult to reach it was only handled three times, when the opportunities arose: Days 35 (28 Mar), 60 (22 April) and 67 (29 April). Key visits were: 25 Feb, Day 4: One chick and one egg visible. 4 Mar, Day 11: Two chicks and one egg. 17 Mar, Day 24: Two chicks; we removed the egg. 22 Mar, Day 29: Two chicks; one slightly more advanced. 25 Mar, Day 32: Only one chick visible. 31 Mar, Day 38: Wendy was caught on the nest and radiotagged. 17 Apr, Day 55: The chick was not in main nest chamber, but close to the main (back) entrance. 22 Apr, Day 60: The chick was sitting at the back entrance. He was caught and banded KA Apr, Day 63: The chick was sitting at the back entrance to the nest. 29 Apr, Day 67: The chick was one metre outside the nest entrance. He was caught and a radio-tagged. 6 May, Day 74: The chick was sitting five metres outside the nest entrance. Post-fledging For the first 16 days after leaving the nest the fledgling (Dobby) roosted close to the female, less than 20 m from the nest. During June Wendy and Dobby generally roosted m apart in the vicinity of a feeding station. Subsequently, although Dobby did not move away from his natal territory, there was little overlap in the areas he and his mother roosted in. On several occasions during September Dobby was at the feeding station together with Wendy (observations were made from a hide, using night vision equipment; the birds' identities were confirmed by radiotelemetry). Dobby was being fed by Wendy, and taking food from the feeding station. Towards the end of September although Wendy still fed Dobby occasionally, she was often aggressive towards him. John-Girl Summary John-Girl was caught and radiotagged on 28 November She spent four nights 21 January) close to the active Booming Sites 7, 8 and 9. On the night of 19 January she copulated at Site 7, presumably with Pegasus, who was caught at this site later in the season. John-Girl laid two fertile eggs. The first egg was laid on, or before, 28 January. On 20 February we removed one egg to Auckland Zoo. This egg hatched on 27 February, but the chick died when it was four days old, on 3 March, as a result of infections. The egg left in the nest hatched successfully on 1 or 2 March. The chick (subsequently named Stumpy) developed normally and left the nest between 9 and 15 May, 69 to 75 days after hatching. Before Stumpy left the nest he was banded and radiotagged. Stumpy remained in the John-Girl's home range until shortly after she died in mid-september He then 16
21 shifted 1 km to the north and we lost contact with him soon after, probably as a consequence of his radio-transmitter failing. The cause of John-Girl's death could not be determined as her carcass was in an advanced state of decomposition when found on 30 September. Weight Mean weight prior to supplementary feeding = 1.23 kg (n=16, sd=0.110) Weights recorded since supplementary feeding began: 08 Oct kg 03 May kg 28 Nov kg 04 Aug kg 06 Aug kg Previous breeding history John-Girl probably didn't attempt to breed during the breeding season. She was captured and examined on 3 April 1990, at which time she didn't have a nest or brood patch. Supplementary feeding John-Girl was taking supplementary food consistently from early 1990 onwards. Between 25 December 1990 and 16 January 1991 she fed at one of the feeding stations nightly. She visited booming sites for copulation during the period January, and when she returned she fed nightly for five nights, January, before settling on a nest. During the first three weeks of incubation she fed irregularly (on 14 nights out of 24 during the period 29 January - 21 February) but during the final week of incubation she began feeding nightly and continued to do so. The quantities of foods taken increased dramatically during brooding and again after fledging. From mid-march onwards Girl also took water at the feeding stations, presumably to moisten food for regurgitation to her chick. See Powlesland and Lloyd (1992) for further details. Pre-nesting movements Until 17 January John-Girl routinely roosted in the vicinity of her feeding stations, then for the four days January she roosted close to active booming sites: 18 January 200 m west of Booming Site 9, 19 January 200 m south-west of Site 7, 20 January 50 m south of Site 7, 21 January 20 m west of Site 9. From 22 January onwards she roosted at her nest site. Copulation John-Girl spent four nights (17-21 January) close to the active Booming Sites 7, 8 and 9. On the night of 19 January she copulated at Site 7, presumably with Pegasus, who was caught at this site on 25 February in a cage trap. Nest log Incubation (Laying date: January 1991) 25 Jan, 16.00: We approached to within 2 m of the nest, John-Girl could be seen in the nest, but was not disturbed. 17
22 28 Jan, 16.05: The nest was inspected, John-Girl didn't respond; we lifted her to one side to reveal one egg, but there could have been more. 15 Feb, : John-Girl's movements were monitored remotely but because she remained in or near the nest we didn't inspect it. 17 Feb, : John-Girl's movements were monitored remotely until she left the nest. We then inspected the nest contents, there were two eggs which we numbered and candled. We replaced the eggs together with a temperature sensitive artificial egg. 20 February, 10.40: Both eggs were removed and candled. Egg #2 was placed in a portable incubator at 95 F and taken to Auckland Zoo by helicopter. Throughout the visit John-Girl continued incubating the artificial egg. Brooding (Hatching date: c. 2 March; Fledging date: c.11 May, aged 70 days; Chick name: Stumpy) During the nestling period all nest visits were made at night while John-Girl was absent from the nest. From Day 1 on 3 March to Day 45 on 16 April the nest was visited by us every three days. Subsequently the interval between visits increased to between five and ten days. During visits the chick was weighed, measured, and photographed; developmental details were also noted (see Section for details). Key visits were: 03 Mar, Day 1: The chick was first seen, probably 1 day old. No egg shell fragments were found in the nest. 21 Apr, Day 50: The chick was banded KA May, Day 60: The chick was radiotagged. 09 May, Day 68: This is the last time the chick was seen in the nest; he sat close to the entrance. 15 May, Day 74: The fledgling was 5 m from the nest, perched on a stump above the nest. The nest chamber looked as if it had not been used for several days. Post-fledging John-Girl and her fledgling (Stumpy) roosted together within a few metres of the nest for the first 16 days after fledging. During June they generally roosted separately, up to 100 m apart, away-from the nest, and close to a feeding station. Throughout July, August and early September the areas John-Girl and Stumpy roosted in were adjacent with some overlap. Generally Stumpy was within 200 m of the nest site and well within John-Girl's traditional home range. Although he may have fed at the two supplementary feeding stations in the female's home range there is no proof of this. John-Girl died in mid- September and soon afterwards Stumpy left the natal home-range and shifted 1 km to the north. We lost contact with him soon after, probably as a consequence of his radiotransmitter failing. Artificial rearing On 20 February at one egg (#2) was removed from the nest, candled, placed in a incubator at 95 F, and transferred to Auckland Zoo by helicopter. Details of its subsequent fate were provided by Dr. R. Jacob-Hoff, Senior Curator at the Auckland Zoo. 18
23 The egg hatched successfully on 27 February in an incubator at the zoo. The chick was held at standard brooding temperatures and fed with a commercial parrot formula using a syringe. The chick developed well, appearing vigorous and healthy. During the four days following hatching it increased its weight by 39%. On 3 March, at age 4 days, the chick died after an illness of 3-4 hours characterised by signs of digestive disorder and respiratory distress. Autopsy results indicate that the chick suffered a digestive disorder caused by a Klebsiella infection which led to regurgitation; food particles were inhaled and these induced a severe pneumonia, physiological shock and death. Bella-Rose Summary Bella-Rose was caught and radiotagged on the 19 October Although she moved widely during late January and early February 1991 she remained on the lower half of the island, below 400 m a.s.l and we do not believe that she visited any active booming sites. Subsequent regular monitoring did not provide any evidence of her having a nest. Weight Mean weights prior to supplementary feeding = 1.42 kg (n=3 sd=0.153) Weights recorded since supplementary feeding began: 17 Sep kg 12 Oct kg 19 Oct kg 30 Sep kg Weight Bella-rose probably didn t attempt to breed during the breeding season. She was captured on 19 February 1990 at which time she did not have a nest or brood patch. Supplementary feeding Bella-Rose fed almost nightly except for the 62-day period 16 December February 1991 at the beginning of the breeding season. During this period she fed irregularly during the period 27 December -10 January; and on the nights 20, 26, 27, 28 January & 6,11,12 February. Pre-nesting movements Up until 22 December 1990 Bella-Rose routinely roosted close to her feeding station. From then until 21 February 1991 she moved widely, only returning to her feeding station for a few days at a time. Between 13 and 19 January 1991 she roosted c. 1.5 to the east of her feeding station in the mid-altitude region on Track 17 and 19. She returned to her feeding station briefly and then moved c. 1 km north-west to Tracks 2 and 3 where she roosted from January. She returned briefly to her feeding station before again moving northwest, c. 1.5 km across to Track 3 and then Track 5 between 1-5 February After another brief visit to the feeding station area she moved about 500 m to the east and she roosted there from 8-10 February. From 21 February onwards she resumed roosting close to her feeding station. Copulation Despite her extensive wandering early on in the booming season when copulations 19
24 usually occur, Bella-Rose did not appear to climb above about 400 m a.s.l and probably did not visit an active booming site even though she was almost certainly able to hear booming. There was no sign of copulations at the booming sites nearest to her. Nesting On 25 January we approached Bella-Rose closely during the day to see whether she was nesting. She was roosting close to her feeding station in the open and moved away when approached. There was no indication of a nest at this time and throughout the rest of the breeding season her daytime roosts shifted frequently indicating that she was not on a nest Nest locations and descriptions The heights above sea level are in imperial units as used on the map in Hamilton (1961). Maggie The nest (150 ft a.s.l.) was near the bottom of a gully, 10 m above a stream which retained water during the nest period. The surrounding forest had a high, closed canopy of pohutakawa (Metrosideros excelsa), puriri (Vitex lucens), and taraire (Beilshmiedia taraire); and a relatively open understory. The nest was in a 4 m long cavity, 180 mm high and 200 mm wide, in the horizontal trunk of a pohutakawa. The entrance faced uphill and, although it was 570 mm above ground level, there were gently sloping roots leading up to it. The nest was 450 mm in from the entrance and was a bowl shaped depression lined with bark, dried leaves and wood fragments. Heather The nest (680 ft a.s.l) was located 30 m from one of our tracks on an exposed cool ridge facing south east. The surrounding forest had a scattered, open canopy of kauri (Agathis australis) and kanuka (Kunzea ericoides); and a scattered, low undergrowth of Gahnia sp., Blechnum sp., Astelia trinerva and ponga (Cyathea dealbata). The nest was on the ground, under the leaves of a clump of Gahnia sp. A 300 mm high bank ran along the back edge of the nest bowl. The entrance faced downhill to the SSE. Although the Gahnia leaves concealed the nest well, they did not provide an effective protection from wind and rain. John-Girl The nest (1100 ft a.s.l.) was located 10 m from a track on a broad ridge with a southerly aspect. The surrounding forest was mature, with a high closed canopy of tawaroa (Beilschmiedia tawaroa) and taraire (B. taraire). Undergrowth was varied and generally dense, including thickets of kiekie (Freycinetia baueriana). The nest was in a large chamber in the side of a 3 m hillock formed by the root of a dead tree. The entrance was on the downhill side of the hillock and faced south-west. The entrance aperture was 300 x 300 mm, 120 mm above floor of nest cavity. The nest cavity extended back 1160 mm from entrance and was 840 mm across at the widest point; a 500 x 500 mm annex to the left of the entrance gave the nest cavity a total length 20
25 of c mm. The maximum height of the cavity was 530 mm. The floor was flat and consisted of dry humus or peat. The ceiling and walls were rotted wood. The nest was very dry and sheltered. It remained dry after very heavy rain. Wendy The nest (1300 ft a.s.l.) was located on a sunny west facing slope deep in a large valley. The surrounding forest was mature, with a high and complete canopy of hard beech (Nothofagus truncata), tawaroa (Beilschmeidia tawaroa), rata (Metrosideros robusta). The undergrowth was varied and dense with supplejack (Ripogonum scandens), kiekie (Freycinetia baueriana), mapou (Myrsine australis), kohurangi (Brachyglottis kirkii), mingimingi (Cyathodes fasciculata), Astelia sp. The nest was in a large chamber among the buttressed roots of a large (d.b.h. 2.0 m) hard beech (Nothofagus truncata). There were two separate entrances, each c. 200 mm diameter, on opposite sides of the tree facing west and north. There was more than 3 m between the two entrances and there seemed to be a massive chamber in the centre more than 1 m in from both entrances. The eggs were incubated within 800 mm of the western entrance but later Wendy and the nestlings moved deep into the main chamber. The floor was flat and consisted of dry humus or peat. The ceiling and walls were rotted wood. The nest was well concealed and very sheltered but during a period of heavy rain some of the nest floor became wet Nest attendance Incubation Remote radio-monitoring of three of the incubating females (Maggie, Heather, and John- Girl) provided some information on nest attendance during incubation. Remote monitoring comprised both occasional instantaneous monitoring during the day, and prolonged monitoring during the first 4-5 hours of darkness on a number of nights. The information is not comprehensive but provides some indication of the pattern of attendance: Daytime: Females were on the nest on all occasions that nests with eggs were monitored during daylight. Nighttime: At night the females were active for 5-10 minutes shortly after dark, usually c. 30 minutes after official sunset. During this activity bout the females remain either in or close to the nest. There were similar short bouts of activity at approximately hourly intervals throughout the rest of the night. Longer periods off the nest to feed (at feeding stations) did not happen nightly. Early in incubation such feeding periods lasted only c. 30 minutes but towards the end of incubation a female was recorded as being away from the nest for 70 minutes. Brooding Our daytime visits to Wendy's nest throughout most of the brooding period indicate that she spent most daylight hours in the nest. Our regular night-time visits to John-Girl's nest provided reliable information on night-time nest attendance during brooding (Table 6). 21
26 22
27 The normal daytime temperature of eggs being incubated The normal daytime temperature of eggs being incubated was determined from the series of temperature obtained from two nests shown in Table 7. Data obtained during periods following disturbance was excluded (i.e. 20 Feb Feb 15.45). The average temperature of eggs during normal daytime incubation was C, varying from 36.2 to 37.2 o C. The effect of nest disturbance on egg temperatures Results of monitoring egg temperature during and following our visit to the nest show that disturbance at the nest was followed by an initial minor cooling of about half a degree (presumably a result of the eggs being exposed) followed by an increase to 37.9 C one hour after disturbance, more than one degree above normal temperature (probably a consequence of the female's increased metabolism due to agitation). Higher temperatures were also observed on the day following disturbance but normal, lower, temperatures were observed two days after the disturbance Egg data Morphometric data Table 8 is morphometric data on eggs handled during the breeding season: weight (g), length (mm), breadth, i.e. the greatest diameter (mm) and the volume and density of each egg computed using the formulae (Jordan and Mattachione 1989): Volume = Length x (Breadth) 2 x Density = Weight/Volume Egg density provides a good indication of viability (Jordan and Mattachione 1989). 23

28 Information from candling Maggie's egg One egg laid c. 21 January. The egg was candled on 27 January. There was a poorly defined central mass 20 mm across with no venation or embryo apparent but the size of the air cell indicated that the egg had been incubated for 5 days. It was concluded that the egg was infertile or had died during early development. It was transferred to Auckland Zoo on 29 January in an incubator at 95 F. Examination by experts at Auckland Zoo and Animal Health Laboratories confirmed that there was no evidence of an embryo. Heather's eggs Two eggs laid c January. The two eggs were transferred to Auckland Zoo on 3 February, 7-13 days after laying. They were candled when first removed: Egg #1-There was a 7 mm long embryo surrounded by a blood ring. (In domestic poultry a 7 mm embryo would be approximately 5 days old.) The egg was fertile but had died 24 hours before candling. Egg #2 -There was a poorly defined central mass with no venation or embryo apparent. The egg was infertile or died during early development. John-Girl's eggs Two eggs laid c January. 17 Feb, days after laying. The eggs were both well vasculated with medium to large air spaces. A small amount of albumin was visible at the small end of Egg but none was apparent in Egg #2. Egg appears to be 6-8 days before hatching, Egg looked 2-3 days more advanced. 20 Feb, days after laying. Egg had not developed noticeably since 17 February. Egg #2 had a very dark embryo occupying the entire egg except for a small well defined air cell. There was no sign of the egg tooth breaking through the air cell membrane. Egg was removed to Auckland Zoo on 20 Feb and hatched on 27 February. 24
29 Wendy's eggs Three eggs laid c. 19 January. One inviable egg was retrieved from nest on 17 March after c. 45 days incubation. Two other eggs in the nest hatched around 1 March Period of rapid yolk deposition J. Cockrem of Massey University determined the period of rapid yolk deposition for the infertile egg taken from Heather's nest (Cockrem, in prep). There were 7 growth rings, indicating that the likely period of rapid yolk deposition was 8-10 days, which is not unusually long for an egg of this size Chick Development Wendy's nest - Surviving chick: Dobby - Estimated hatching date 21 February. John Girl's nest - Chick name: Stumpy - Estimated hatching date 2 March. Most of the observations on chick development were made at John-Girl's nest because access to Wendy's nest was difficult and prevented regular and detailed observations. The few close inspections of Wendy's chicks confirmed the general pattern of development recorded at John-Girl's nest but Wendy's surviving chick seemed to develop more slowly than John-Girl's. Towards the end of the nestling period the plumage development of Wendy's chick was 10 days behind that of John-Girl's chick. Weight Figure 2 is the weight, in grams, of the chicks plotted against their estimated age, in days after hatching. The data are provided in Appendix C. Tarsus Length Tarsus length, culmen width and culmen length were all recorded occasionally and can be found in Table 12. Eyes The eyes began to open about 13 days after hatching, but were not fully open until 18 days. Feet Nineteen days after hatching the outside toe was noted curling around to the back. By 22 days zygodactyl feet were fully developed. Vocalisation On the first day after hatching the chick made a low churring noise when picked up. Subsequently it vocalised quietly throughout handling until about 30 days when it began squealing or grunting loudly. It was not until 50 days after hatching that the undisturbed chick was heard vocalising in the nest. Mobility During the first few days after hatching the chicks were weak and uncoordinated, unable to sit up. Twenty days after hatching they were they able to sit up on their haunches with their head up but they were still docile when handled. By 30 days the chicks were quite 25
30 active and difficult to handle. At days after hatching the chicks sat quite upright and moved around easily in the nest chamber. Fledging and post-fledging As early as 55 days after hatching the chicks were sitting outside the main nest chamber, close to the nest entrance. From days the chicks sat in or just outside the nest entrance. For a further c. 15 days after leaving the nest the chicks roosted with the female, ten to twenty metres from the nest. Subsequently they roosted up to 100 m from both the nest and the female but close to feeding stations. Plumage Plumage descriptions for 5-day age periods Age (days) Plumage description 0-5 The chick was covered with thick white down The chick was still covered by thick white down, but feather tracts were apparent under the skin The sheathed feathers of both body contour plumage and secondaries erupted through the skin. There was a small brown tassel at the tip of each sheath At the start of this period the brown tips of the sheathed contour feathers and grey down gave an overall brown/grey appearance to the plumage though there was green feathering on the thighs where the vanes of contour feathers were beginning to unfurl. By 25 days the green feathering was extensive on the thighs, breast, wing coverts, and secondaries. The primary feather sheaths and then the tail feather sheaths erupted through the skin The crown and rump were covered in thick dark grey down without any contour feathers. Green feathering was thick on the thighs but still sparse on the breast. The vanes of contour feathers on the thighs, wing coverts and secondaries were mm unfurled. Green feathering of the forehead and nape began. Primary and tail feather vanes were still not unfurled The rump was dark grey down, and there was now a hint of green feathering on the crown and nape as a few feather vanes unfurled 2-4 mm. Green feathering on the breast was sparse with the unfurled vanes mm long. The length of unfurled vane on the secondaries increased from about 10 mm to 30 mm. The primary and tail feathers started to unfurl The rump was still dark grey down. Feather vanes unfurling on the thighs, crown, belly, secondaries, primaries and tail. 26
31 41-45 Rump still entirely dark grey down; green feathers unfurling everywhere else. The crown and chest were almost completely feathered Rump still entirely dark grey down; green feathers unfurling everywhere else. Crown, face, chest, and wing coverts almost completely feathered with just a few wisps of down poking out. The primary and secondary and tail feathers were still unfurling. The tip mm of the inner web of primaries was now formed: - dark with bold yellow reticulations mm wide The contour plumage was almost complete except for small amounts of down on the lower rump, flanks, and underside of wings. Primary, secondary, and tail feathers were half unfurled. The green plumage was noticeably paler than adult plumage Down was still apparent on the flanks and under the wings, otherwise the green contour plumage was complete. Primary, secondary, and tail feathers were approximately two-thirds unfurled There were small patches of down on the flanks and under the wings, only visible when the wings were raised, otherwise the contour plumage was completely feathered. The longest feather in the tail was 130 mm long with pieces of sheath on the rachis. The primary feathers were approximately three-quarters unfurled The contour plumage was complete though the pattern on the back seemed finer than in adults. The primary feathers were completely unfurled, with only a few flakes of sheath at the base of rachii. Stumpy's tail was completely grown but only 170 mm long, whereas Dobby's tail at 185 mm appeared to be still growing. The trailing edges at the tip of the primaries were dark with bold yellow reticulations. There were some faint watermarks or dappling present on Stumpy but not on Dobby. All primary and secondary feathers were pointed. Growth of ninth primary Figure 3 is a plot of the length (mm) of the sheathed region and the total length (mm) of the ninth primary (counted from the inside), of the right wing, against age (days since hatching) for the two chicks. 27
32 28
33 29
34 3.2.9 Sexing chicks Feather pattern Powlesland (1989) describes a method to determine the gender of adult kakapo from the pattern at the tip of the inner or trailing vane of the outer primaries. At days after hatching the distal 20 mm of the inner vane of the outer primaries were sufficiently well formed on both chicks for us to attempt to use this method to determine their gender. At this time the trailing vanes were dark brown or black with bold yellow reticulations mm wide. The patterns on the trailing vanes were unchanged 30 to 40 days after fledging. The patterns did not correspond to either of the patterns described by Powlesland. (A very feint dappling was discernible on the vane of Stumpy 40 days after fledging but it was not felt to be sufficiently well developed to determine the bird s gender.) Thus it seems that kakapo's juvenile plumage can not be used reliably to determine gender. The gender of adult kakapo can also be determined using tarsus length and culmen width. There was almost no overlap in the ranges of values of these two measurements from samples of male and female kakapo on Stewart Island (Table 11). Values of tarsus length, and culmen width measured at irregular intervals from 42 days after hatching are shown for both chicks in Table 12. All values for both measurements for Stumpy are outside the ranges of values for females and within the ranges of values for males. For Dobby the first value for tarsus length, measured 60 days after hatching, was within the range of values for females, all subsequent values of both measurements were outside the ranges for females and within the ranges for males. We concluded that both chicks were male. 30
35 Potential nest problems Kiore Rodent control was undertaken around both John Girl's and Wendy's nests. Non-toxic pre-bait was placed in silos around John-Girl's nest on 10 February, and Wendy's nest on 6 February. Ten days later the pre-bait was replaced by toxic bait which was replenished regularly until the silos were removed (3 April, John-Girl's nest; 20 April Wendy's nest), just before the kakapo chicks left the nests. No kiore activity was noted around, or in, any nests. Mites Nest material from each of the nests was regularly inspected closely for mites. No mites were found. Bacterial infections On the 18 March, sixteen days after hatching, a swab and faecal sample were obtained from John-Girl's chick and sent to Auckland Zoo for a bacterial check. No pathogens were identified. Petrels When Wendy's nest was first inspected on 26 January a dead Cook's petrel (Petrodroma cookii) could be seen at the back of the nest chamber. When we removed it three days later it was maggot-ridden. A fresh dead petrel was also found outside the nest on 25 February. Both petrels may have been killed by Wendy. Soiled feathers During the first 4 weeks after hatching, down on John-Girl's chick repeatedly became encrusted with food material regurgitated by the mother. On the first day after hatching, down feathers under the wing and at base of bill were matted with nest material. At four days it was noted that food material encrusted one eye and feathers at the base of the bill. This material was washed off by us with warm water during our next nest visit, three days later, but fresh material again encrusted the facial 31
36 feathers at ten days. The feathers remained encrusted until 25 days when they were again cleaned by us. Feather encrustation was not subsequently observed. Malformed feather When Dobby was handled on 11 June, 40 days after he left the nest, it was noted that the outer primary of the left wing was bent outwards. 3.3 Supplementary feeding Details of the supplementary feeding programme can be found in Powlesland and Lloyd (1992). All five females had been feeding regularly at supplementary feeding stations for at least 6 months prior to January Several of the males were also taking food regularly prior to the booming season at feeding stations placed either near booming sites or within female's home ranges. Information on supplementary feeding by individual males is of poor quality as the males were not radio-tagged. 3.4 Rat index trapping Table 13. Results from rat index trapping, Little Barrier Island,
37 4 DISCUSSION 4.1 A summary of breeding activity on Little Barrier Island Table 14 is a summary of breeding activity on Little Barrier Island since kakapo were released on the island. Since supplementary feeding was begun on Little Barrier Island in September 1989: The booming season was the most intensive since kakapo were transferred to Little Barrier Island. The booming season began earlier than has ever been previously recorded for any kakapo population. Nesting has occurred in both summers, whereas there is no evidence of nesting during the seven summers prior to supplementary feeding. Two of the females attempted to nest in consecutive years. This has not previously been recorded for kakapo. It therefore seems reasonable to conclude that on Little Barrier Island supplementary feeding has promoted breeding by both male and female kakapo. 4.2 A comparison of breeding on Little Barrier Island and Stewart Island Booming The booming season on Little Barrier Island was more extended than any recorded on Stewart Island. Several males were booming by mid-october and booming continued until early April, whereas during the most intensive booming seasons monitored on Stewart Island ( and ) booming only began in mid- December and ended in April (Powlesland et al. in press). 33
38 Clutch size Table 15 shows the sizes of clutches on Stewart Island (Powlesland et al. in press) and Little Barrier Island. Table 15. Clutch sizes on Stewart Island and Little Barrier Island. Island Year Clutch Sizes Stewart Is or more, & 3 or more ,3 & 4 Little Barrier Is & ,2,2 & 3 Clutch sizes on Little Barrier Island in 1990 were smaller than those recorded on Stewart Island whereas clutch sizes on Little Barrier Island in 1991 were similar to those recorded on Stewart Island. Chick weights The two chicks on Little Barrier Island both weighed approximately 1.7 kg at fledging. Succesful fledging was only observed on Stewart Island during In that year a male weighing 1.5 kg fledged from one nest and a male weighing 2.2kg and a female weighing 1.7 kg fledged from a second nest. Thus during the 1981 breeding season on Stewart Island the diet of one of the two females whose nests were monitored was probably superior to the supplemented diet of the two females with fledglings on Little Barrier Island in Nest attendance A comparison of data on nest attendance for the first few days of brooding presented here and in Powlesland et al. (in press) indicates that the female kakapo monitored on Little Barrier Island left the nest earlier but stayed away for shorter periods (33 minutes after sunset and 63 minutes away) than the female on Stewart Island during 1985 (60 minutes after sunset and 165 minutes away). The long periods spent away from the nest on Stewart Island were probably a consequence of the shortage of natural foods on Stewart Island in
39 4.3 Kakapo productivity Nesting has occurred in only four years during the eleven years, , that kakapo populations have been monitored (Table 16). In these four years the monitored females nested a total of 12 times and produced at least 25 eggs. Seven eggs were infertile, 13 hatched, 6 young fledged, but only 5 to the sub-adult stage. Two other young were probably produced on Stewart Island in 1981 by females not being monitored. The frequency of nesting attempts, and the success rate of nests in the wild is very low. This is true even under what appear to be ideal circumstances on Little Barrier Island, i.e. ad libitum supplementary food, the absence of significant predators, and relatively mild climate. Low reproductive rates are probably normal for large parrots in the wild. For instance, the world's second largest parrot species, the Hyacinth macaw (Anodorhynchus hyacinthinus) is reported to nest about once every three years and produce no more than one chick from a nest (Munn et al. 1991). The total number of females has declined from c. 30 in 1980 to c. 15 in During this time seven young, including only two females, have been recruited to the adult population. The kakapo population is rapidly approaching what Gilpin and Soule (1986) described as an "extinction vortex" from which it can only be rescued by prompt intervention. The results of the last two years' work on Little Barrier indicate that although intensive management of population in the wild (i.e. supplementary feeding, nest protection and rodent eradication) will improve the species productivity, the improvement may not be sufficient to reverse the species decline. Many large species of parrots have been bred successfully in captivity, usually productivity being enhanced (e.g. a pair of hyacinth macaws produce a maximum of 1 young every three years in the wild whereas in captivity they can produce up to 8 young in one year, Munn et al. 1991). It is essential that there is an increased emphasis on in the Kakapo Recovery Programme. Aviculture should include breeding from captive stock, and artificial incubation and rearing of progeny from both wild and captive birds. 35
40 4.4 Male breeding success Table 16. Copulation details and There were at least 10 males booming during both the and breeding seasons, but the six recorded copulations can be reasonably attributed to only two males: Bill and Pegasus (Although Barnard was found near Site 7 in March 1990, at this time of the year males are inclined to leave their usual systems. He was found at Site 12 in February 1990 and was probably booming there during January when copulations occurred.) Thus two males, or their booming sites, are being selected by the females. Although the two offspring from this breeding season were fathered by separate males, male mating success is heavily skewed. In small populations loss of genetic diversity can be reduced by ensuring that all males contribute equally to future generations (Frankel & Soule 1981, Shaffer 1987). This is especially important for kakapo at present because the high proportion of males in the population (c. 37 males and c. 17 females) means that most of the populations genetic variation resides in males. Therefore until the population has recovered to a moderate size (i.e.>50 breeding females) males that have already produced a large number of progeny and are over-represented in the succeeding generation should be removed from the breeding population. In order to manage the free-living population in this way it will be necessary to monitor paternity by either: using DNA fingerprinting techniques; or monitoring the identity of males and females at booming sites, and progeny in the nest. Critics of a management regime of this type argue that the removal of successful breeding males may: reduce the overall productivity of the population. thwart the process of natural selection. lead to a population poorly adapted for its environment. We do not agree with these assertions: Removal of males over-represented in the next generation is unlikely to reduce the overall productivity of the population because the species is polygynous and at present there is a large surplus of males in the population. Although long term studies (see Newton 1989 and Mills 1990) indicate that in natural populations breeding success is often heavily skewed, thus providing the mechanism for natural selection, in small relict population of previously widespread species changes in gene frequencies caused by natural selection are likely to be swamped by the random loss of genetic material which reduces genetic diversity in succeeding generations. 36
41 If kakapo survive in the long term progeny from Little Barrier Island will be moved to a wide variety of other environments. Thus it is important to retain the species genetic diversity to facilitate survival in diverse environments. Simberloff (1988) provides a comprehensive discussion of these issues. 5 CONCLUSIONS Supplementary feeding promoted breeding effort by both male and female kakapo. Females attempted to nest in consecutive years. Thus it is likely that the female's reproductive cycle is annual and irregular nesting is a consequence of poor nutrition. Females fitted with back-mounted radio-transmitters mated and nested successfully. This season's observations that feather clusters (as described in Section 2.1.4) at booming sites are evidence of copulation. Nest manipulation and intensive nest monitoring (including: handling eggs and chicks, and egg substitution) can be carried out without jeopardising the success of nests. Despite high numbers of kiore present throughout the latter half of the nest period none of the nest contents were preyed on by kiore. We can not be sure, but this may have been a result of the control programme carried out around the two nests with nestlings. Intensive management of wild populations may he insufficient to prevent extinction of kakapo and should be supplemented by an intensive avicultural programme. Male breeding success is likely to be highly skewed. 6 RECOMMENDATIONS Supplementary feeding should be continued on Little Barrier Island, refined, and extended to other wild populations. Intensive management of the wild population should be supplemented by an intensive avicultural programme which should include breeding from captive stock, and artificial incubation and hand-rearing of progeny from both wild and captive birds. Long term management of kakapo should include genetic management to achieve even spread of progeny from the males. Males that have a large number of progeny, and are over-represented in the succeeding generation, should be removed from the breeding population. In free-living populations this will require monitoring of paternity which may be achieved by either: using DNA 37
42 fingerprinting techniques; or monitoring the identity of males and females at booming sites, and progeny in the nest. ACKNOWLEDGEMENTS We would like to acknowledge the assistance of all the DOC staff and volunteers whose efforts have made this study possible. Special thanks go to Terry Greene, Shaarina Boyd, Chris and Robin Smuts-Kennedy. 38
43 REFERENCES Cockrem, J.F. (in prep) Yolk formation in the egg of the kakapo (Strigops habroptilus). Notornis. Dowding, J Report on kakapo booming activity, Little Barrier Island, March-April Unpublished report, Department of Conservation. Frankel, O.H. & M.E. Soule Conservation and evolution. Cambridge University Press, Cambridge. Gilpin, M.E., & M.E. Soule Minimum viable populations: processes of species extinction. Pp In: M.E. Soule (ed): Conservation Biology: The Science of Scarcity and Diversity. Sinauer Assoc. Sunderland, Mass. Greene, T.C Report on kakapo booming activity, Little Barrier Island, January - March Science and Research Internal Report No. 48, Department of Conservation, Wellington. Hamilton, W.M Little Barrier (Hauturu). NZ DSIR Research Bulletin 137. Government Printer, Wellington, NZ. Handford, P.A Report on kakapo booming activity on Little Barrier Island in January -April Unpublished report, Department of Conservation. Hodsell, C Kakapo survey Little Barrier Island 12 September - 7 November Department of Conservation, Auckland. ISSN Series 3. Jordan, R. and S. Mattachione Parrot incubation procedures: a methodical guide to incubation and problem hatches for the aviculturalist. Pickering, Ontario, Canada Lloyd, B.D. and R.G. Powlesland, Kakapo breeding activity, Little Barrier Island, 21 November April Science and Research Internal Report No. 83, Department of Conservation, Wellington. McFadden, I Composition and presentation of baits and their acceptance by New Zealand Wildlife Service Technical Report No. 7. Mills, J.A Lifetime production in the red-billed gull. Pp in Bell et al. (Eds). Acta XX Congressus Internationalis Ornithologici Vol.III. Moorhouse, R Report on kakapo booming activity on Little Barrier Island: January -April Unpublished report, Wildlife Service. Munn, C.A., J.G. Thomsen & C Yamashita The hyacinth macaw. In: Audubon Wildlife Report 1989/1990. Eds. W.J. Chandler & L. Labate. Newton, I Synthesis. Pp in Newton, I. (Ed.). Lifetime reproduction in birds. Academic Press. London. 39
44 Powlesland, R Kakapo recovery plan Department of Conservation, Wellington. Powlesland, R. and B. Lloyd, Progress report of the supplementary feeding of kakapo on Little Barrier Island; September May Science and Research Report, Department of Conservation, Wellington. Powlesland, R. and B. Lloyd, Supplementary feeding of kakapo on Little Barrier Island, May June In prep., Science and Research Internal Report, Department of Conservation, Wellington. Powlesland, R., B. Lloyd, H.A. Best and D.V. Merton. In press. Breeding biology of the Kakapo Strigops habroptilus on Stewart Island, New Zealand. Ibis. Shaffer, M Minimum viable population size; coping with uncertainty. In: Soule, M. (ed.): Viable Populations for Conservation. Cambridge University Press Simberloff, D The contribution of population and community biology to conservation science. Ann. Rev. Ecol. Syst. 19: Veitch, C.R Kakapo - Little Barrier - Winter Unpublished report, Wildlife Service. 40
45 Appendix A: Capture details for All birds were in good condition when handled. The only bird in moult was Maggie; her wing and tail feathers were in the final stages of moult. Each of the females was fitted with a radio-transmitter when captured. 41

46 42


47 43


48 44


49 45


50 46


51 47


52 48
SCIENCE & RESEARCH INTERNAL REPORT NO.160. Kakapo on Little Barrier (Hauturu) Island. Annual report for the year July June 1995
 Island. Annual report for the year July June 1995") SCIENCE & RESEARCH INTERNAL REPORT NO.160 Kakapo on Little Barrier (Hauturu) Island Annual report for the year July 1994 - June 1995 by G.A. Harper Published by Department of Conservation P.O. Box 10-420
SCIENCE & RESEARCH INTERNAL REPORT NO.160 Kakapo on Little Barrier (Hauturu) Island Annual report for the year July 1994 - June 1995 by G.A. Harper Published by Department of Conservation P.O. Box 10-420
Breeding Activity Peak Period Range Duration (days) Laying May May 2 to 26. Incubation Early May to mid June Early May to mid June 30 to 34
 Laying May May 2 to 26. Incubation Early May to mid June Early May to mid June 30 to 34") Snowy Owl Bubo scandiacus 1. INTRODUCTION s have a circumpolar distribution, breeding in Fennoscandia, Arctic Russia, Alaska, northern Canada and northeast Greenland. They are highly nomadic and may migrate
Snowy Owl Bubo scandiacus 1. INTRODUCTION s have a circumpolar distribution, breeding in Fennoscandia, Arctic Russia, Alaska, northern Canada and northeast Greenland. They are highly nomadic and may migrate
Breeding White Storks( Ciconia ciconia at Chessington World of Adventures Paul Wexler
 Breeding White Storks(Ciconia ciconia) at Chessington World of Adventures Paul Wexler The White Stork belongs to the genus Ciconia of which there are seven other species incorporated predominantly throughout
Breeding White Storks(Ciconia ciconia) at Chessington World of Adventures Paul Wexler The White Stork belongs to the genus Ciconia of which there are seven other species incorporated predominantly throughout
PROTECTING MANLY S PENGUINS
 PROTECTING MANLY S PENGUINS NATIONAL PARKS AND WILDLIFE SERVICES VOLUNTARY PENGUIN WARDEN PROGRAM CONFERENCE PRESENTATION WEDNESDAY JULY 23 RD 2008 BACKGROUND: LITTLE PENGUINS EUDYPTULA MINOR o Manly s
PROTECTING MANLY S PENGUINS NATIONAL PARKS AND WILDLIFE SERVICES VOLUNTARY PENGUIN WARDEN PROGRAM CONFERENCE PRESENTATION WEDNESDAY JULY 23 RD 2008 BACKGROUND: LITTLE PENGUINS EUDYPTULA MINOR o Manly s
Surveys of Giant Weta
 Surveys of Giant Weta Little Barrier Island, Pig Island (Foveaux Strait), and Mt Faraday and Price s Basin (Southern Alps) SCIENCE FOR CONSERVATION: 16 Mike Meads and Peter Notman Published by Department
Surveys of Giant Weta Little Barrier Island, Pig Island (Foveaux Strait), and Mt Faraday and Price s Basin (Southern Alps) SCIENCE FOR CONSERVATION: 16 Mike Meads and Peter Notman Published by Department
Ernst Rupp and Esteban Garrido Grupo Jaragua El Vergel #33, Santo Domingo Dominican Republic
 Summary of Black-capped Petrel (Pterodroma hasitata) Nesting Activity during the 2011/2012 Nesting Season at Loma del Toro and Morne Vincent, Hispaniola Introduction and Methods Ernst Rupp and Esteban
Summary of Black-capped Petrel (Pterodroma hasitata) Nesting Activity during the 2011/2012 Nesting Season at Loma del Toro and Morne Vincent, Hispaniola Introduction and Methods Ernst Rupp and Esteban
Rock Wren Nesting in an Artificial Rock Wall in Folsom, Sacramento County, California
 Rock Wren Nesting in an Artificial Rock Wall in Folsom, Sacramento County, California Dan Brown P.O. Box 277773, Sacramento, CA 95827 naturestoc@aol.com Daniel A. Airola, Northwest Hydraulic Consultants,
Rock Wren Nesting in an Artificial Rock Wall in Folsom, Sacramento County, California Dan Brown P.O. Box 277773, Sacramento, CA 95827 naturestoc@aol.com Daniel A. Airola, Northwest Hydraulic Consultants,
Red Crowned Parakeet (Cyanoramphus novaezelandiae) health, disease and nesting study on Tiritiri Matangi 2014/2015. Emma Wells on behalf of
 health, disease and nesting study on Tiritiri Matangi 2014/2015. Emma Wells on behalf of") Red Crowned Parakeet (Cyanoramphus novaezelandiae) health, disease and nesting study on Tiritiri Matangi 2014/2015 John Sibley Emma Wells on behalf of Auckland Zoo, Supporters of Tiritiri Matangi, Massey
Red Crowned Parakeet (Cyanoramphus novaezelandiae) health, disease and nesting study on Tiritiri Matangi 2014/2015 John Sibley Emma Wells on behalf of Auckland Zoo, Supporters of Tiritiri Matangi, Massey
Physical Description Meadow voles are small rodents with legs and tails, bodies, and ears.
 A Guide to Meadow Voles Identification, Biology and Control Methods Identification There are 5 species of Meadow Vole common to California. They are the California Vole, Long-tailed Vole, Creeping Vole,
A Guide to Meadow Voles Identification, Biology and Control Methods Identification There are 5 species of Meadow Vole common to California. They are the California Vole, Long-tailed Vole, Creeping Vole,
BLACK OYSTERCATCHER NEST MONITORING PROTOCOL
 BLACK OYSTERCATCHER NEST MONITORING PROTOCOL In addition to the mid-late May population survey (see Black Oystercatcher abundance survey protocol) we will attempt to continue monitoring at least 25 nests
BLACK OYSTERCATCHER NEST MONITORING PROTOCOL In addition to the mid-late May population survey (see Black Oystercatcher abundance survey protocol) we will attempt to continue monitoring at least 25 nests
Breeding Activity Peak Period Range Duration (days) Egg laying Early April Mid-March to early May 3 to 12
 Egg laying Early April Mid-March to early May 3 to 12") Accipiter gentilis 1. INTRODUCTION The (northern goshawk) stopped breeding regularly in Britain and Ireland in the 1880s. Breeding became regular again from the mid 1900s, as a result of deliberate (unauthorised)
Accipiter gentilis 1. INTRODUCTION The (northern goshawk) stopped breeding regularly in Britain and Ireland in the 1880s. Breeding became regular again from the mid 1900s, as a result of deliberate (unauthorised)
Multiple broods from a hole in the wall: breeding Red-and-yellow Barbets Trachyphonus erythrocephalus in southeast Sudan
 Scopus 29: 11 15, December 2009 Multiple broods from a hole in the wall: breeding Red-and-yellow Barbets Trachyphonus erythrocephalus in southeast Sudan Marc de Bont Summary Nesting and breeding behaviour
Scopus 29: 11 15, December 2009 Multiple broods from a hole in the wall: breeding Red-and-yellow Barbets Trachyphonus erythrocephalus in southeast Sudan Marc de Bont Summary Nesting and breeding behaviour
I will post a pdf at the end of the presentation with some additional details and references so there is no need to try to copy it all.
 I will post a pdf at the end of the presentation with some additional details and references so there is no need to try to copy it all. The West End is a historic nest. Here's the photo of the 1929 West
I will post a pdf at the end of the presentation with some additional details and references so there is no need to try to copy it all. The West End is a historic nest. Here's the photo of the 1929 West
by A.P. Ballance c/- 7 Florida Place, Auckland 5. ABSTRACT
 PARYPHANTA AT PAWAKATUTU by A.P. Ballance c/- 7 Florida Place, Auckland 5. ABSTRACT During a 2 day collection in a stand of 26 year old pines in the Waipoua Forest, Northland, 18 live Paryphanta busbyi
PARYPHANTA AT PAWAKATUTU by A.P. Ballance c/- 7 Florida Place, Auckland 5. ABSTRACT During a 2 day collection in a stand of 26 year old pines in the Waipoua Forest, Northland, 18 live Paryphanta busbyi
2009 Eagle Nest News from Duke Farms eagle nest Written by Larissa Smith, Assistant Biologist
 2009 Eagle Nest News from Duke Farms eagle nest Written by Larissa Smith, Assistant Biologist July 7 - The youngest chick was gone from the nest this morning but has returned to the nest several times
2009 Eagle Nest News from Duke Farms eagle nest Written by Larissa Smith, Assistant Biologist July 7 - The youngest chick was gone from the nest this morning but has returned to the nest several times
For further information on the biology and ecology of this species, Clarke (1995) provides a comprehensive account.
 provides a comprehensive account.") Circus aeruginosus 1. INTRODUCTION The marsh harrier (western marsh harrier) is increasing as a breeding species in Great Britain (Gibbons et al., 1993; Underhill-Day, 1998; Holling & RBBP, 2008) with
Circus aeruginosus 1. INTRODUCTION The marsh harrier (western marsh harrier) is increasing as a breeding species in Great Britain (Gibbons et al., 1993; Underhill-Day, 1998; Holling & RBBP, 2008) with
Record of Predation by Sugar Glider on Breeding Eastern Rosellas 33Km NE of Melbourne in November 2016
 Record of Predation by Sugar Glider on Breeding Eastern Rosellas 33Km NE of Melbourne in November 2016 By Frank Pierce [email - jmandfp@bigpond.com.au ] 18/01/2016 SUMMARY Eastern Rosellas nested in a
Record of Predation by Sugar Glider on Breeding Eastern Rosellas 33Km NE of Melbourne in November 2016 By Frank Pierce [email - jmandfp@bigpond.com.au ] 18/01/2016 SUMMARY Eastern Rosellas nested in a
Husbandry Guidelines Name Species Prepared by
 Husbandry Guidelines Name Species Prepared by 1. ACQUISITION AND ACCLIMATIZATION Status of wild population Status current captive population Sources of birds Acclimatization procedures Weighing Feeding
Husbandry Guidelines Name Species Prepared by 1. ACQUISITION AND ACCLIMATIZATION Status of wild population Status current captive population Sources of birds Acclimatization procedures Weighing Feeding
OBSERVATIONS OF HAWAIIAN
 - - - - ------ - - - - - OBSERVATIONS OF HAWAIIAN HAWKACTIV ltv Spring 1985 Jack Jeffries P. O. Box 518 Volcano, HI 96785 .. INTRODUCTION This report is part of a continuing study to provide baseline data
- - - - ------ - - - - - OBSERVATIONS OF HAWAIIAN HAWKACTIV ltv Spring 1985 Jack Jeffries P. O. Box 518 Volcano, HI 96785 .. INTRODUCTION This report is part of a continuing study to provide baseline data
OBSERVATIONS OF PEMBROKE PINES BALD EAGLE NEST - FWC ID# BO-002
 OBSERVATIONS OF PEMBROKE PINES BALD EAGLE NEST - FWC ID# BO-002 DATE EGG DAY HATCH DAY FLEDGE DAY ADULTS IN VIEW NESTLNGS FLEDGLNGS ADULTS ON NEST FEEDINGS NOTES 2008-2009 Nesting Season 20081202 1 1 One
OBSERVATIONS OF PEMBROKE PINES BALD EAGLE NEST - FWC ID# BO-002 DATE EGG DAY HATCH DAY FLEDGE DAY ADULTS IN VIEW NESTLNGS FLEDGLNGS ADULTS ON NEST FEEDINGS NOTES 2008-2009 Nesting Season 20081202 1 1 One
A Case Study of the Effectiveness of TNR on a Feral Cat Colony
 Compiled and Written By: Triple R Pets PO Box 86, Western Springs, IL 60558 708.738.1438 phone 630.214.3603 fax www.triplerpets.org QUESTION Does a TNR trap/neuter/return program stabilize or reduce the
Compiled and Written By: Triple R Pets PO Box 86, Western Springs, IL 60558 708.738.1438 phone 630.214.3603 fax www.triplerpets.org QUESTION Does a TNR trap/neuter/return program stabilize or reduce the
12 The Pest Status and Biology of the Red-billed Quelea in the Bergville-Winterton Area of South Africa
 Workshop on Research Priorities for Migrant Pests of Agriculture in Southern Africa, Plant Protection Research Institute, Pretoria, South Africa, 24 26 March 1999. R. A. Cheke, L. J. Rosenberg and M. E.
Workshop on Research Priorities for Migrant Pests of Agriculture in Southern Africa, Plant Protection Research Institute, Pretoria, South Africa, 24 26 March 1999. R. A. Cheke, L. J. Rosenberg and M. E.
Seven Nests of Rufescent Tiger-Heron (Tigrisoma lineatum)
") Seven Nests of Rufescent Tiger-Heron (Tigrisoma lineatum) Steven Furino and Mario Garcia Quesada Little is known about the nesting or breeding behaviour of Rufescent Tiger-Heron (Tigrisoma lineatum). Observations
Seven Nests of Rufescent Tiger-Heron (Tigrisoma lineatum) Steven Furino and Mario Garcia Quesada Little is known about the nesting or breeding behaviour of Rufescent Tiger-Heron (Tigrisoma lineatum). Observations
Egyptian vulture (Neophron percnopterus) research & monitoring Breeding Season Report- Beypazarı, Turkey
 research & monitoring Breeding Season Report- Beypazarı, Turkey") Egyptian vulture (Neophron percnopterus) research & monitoring - 2011 Breeding Season Report- Beypazarı, Turkey October 2011 1 Cover photograph: Egyptian vulture landing in Beypazarı dump site, photographed
Egyptian vulture (Neophron percnopterus) research & monitoring - 2011 Breeding Season Report- Beypazarı, Turkey October 2011 1 Cover photograph: Egyptian vulture landing in Beypazarı dump site, photographed
Breeding biology of kakapo (Strigops habroptilus) on offshore island sanctuaries,
 on offshore island sanctuaries,") 27 Breeding biology of kakapo (Strigops habroptilus) on offshore island sanctuaries, 1990-2002 DARYL K. EASON Research Development and Improvement Division, Department of Conservation, Private Bag 5, Nelson,
27 Breeding biology of kakapo (Strigops habroptilus) on offshore island sanctuaries, 1990-2002 DARYL K. EASON Research Development and Improvement Division, Department of Conservation, Private Bag 5, Nelson,
Scholarship 2017 Biology
 93101Q 931012 S Scholarship 2017 Biology 9.30 a.m. Monday 20 November 2017 Time allowed: Three hours Total marks: 24 QUESTION BOOKLET There are THREE questions in this booklet. Answer ALL questions. Write
93101Q 931012 S Scholarship 2017 Biology 9.30 a.m. Monday 20 November 2017 Time allowed: Three hours Total marks: 24 QUESTION BOOKLET There are THREE questions in this booklet. Answer ALL questions. Write
Steggles Sydney Royal School Meat Bird Pairs Competition Support Guide
 Steggles Sydney Royal School Meat Bird Pairs Competition Support Guide 1 Contents Introduction Setting up On arrival of your day-old chicks Monitoring Weighing and assessing growth Temperature control
Steggles Sydney Royal School Meat Bird Pairs Competition Support Guide 1 Contents Introduction Setting up On arrival of your day-old chicks Monitoring Weighing and assessing growth Temperature control
Hole-nesting birds. In natural conditions great and blue tits breed in holes that are made by e.g. woodpeckers
 Hole-nesting birds In natural conditions great and blue tits breed in holes that are made by e.g. woodpeckers Norhern willow tits excavate their own holes in rotten trees and do not accept old holes or
Hole-nesting birds In natural conditions great and blue tits breed in holes that are made by e.g. woodpeckers Norhern willow tits excavate their own holes in rotten trees and do not accept old holes or
ACTIVITY PATTERNS AND HOME-RANGE USE OF NESTING LONG-EARED OWLS
 Wilson Bull., 100(2), 1988, pp. 204-213 ACTIVITY PATTERNS AND HOME-RANGE USE OF NESTING LONG-EARED OWLS E. H. CRAIG, T. H. CRAIG, AND LEON R. POWERS ABSTRACT.-A study of the movements of two pairs of nesting
Wilson Bull., 100(2), 1988, pp. 204-213 ACTIVITY PATTERNS AND HOME-RANGE USE OF NESTING LONG-EARED OWLS E. H. CRAIG, T. H. CRAIG, AND LEON R. POWERS ABSTRACT.-A study of the movements of two pairs of nesting
What is the date at which most chicks would have been expected to fledge?
 CURLEW FAQs FACTS AND FIGURES AND ADVICE FOR THOSE WANTING TO HELP SUPPORT NESTING CURLEW ON THEIR LAND The Eurasian Curlew or, Numenius arquata, spends much of the year on coasts or estuaries, but migrates
CURLEW FAQs FACTS AND FIGURES AND ADVICE FOR THOSE WANTING TO HELP SUPPORT NESTING CURLEW ON THEIR LAND The Eurasian Curlew or, Numenius arquata, spends much of the year on coasts or estuaries, but migrates
Eastern Bluebird Early Egg Viability Outcomes- A Mini- Study. By Penny Brandau and Paula Ziebarth
 Eastern Bluebird Early Egg Viability Outcomes- A Mini- Study By Penny Brandau and Paula Ziebarth Ask Madame WingNut for this issue of the OBS newsletter is coauthored by two Madame WingNuts: Penny Brandau
Eastern Bluebird Early Egg Viability Outcomes- A Mini- Study By Penny Brandau and Paula Ziebarth Ask Madame WingNut for this issue of the OBS newsletter is coauthored by two Madame WingNuts: Penny Brandau
THE BLUE PENGUIN (Eudyptula minor) AT TAIAROA HEAD, OTAGO,
 AT TAIAROA HEAD, OTAGO,") SCIENCE & RESEARCH SERIES NO.86 THE BLUE PENGUIN (Eudyptula minor) AT TAIAROA HEAD, OTAGO, 1992-1993 by Lyndon Perriman and Bruce McKinlay Published by Head Office, Department of Conservation, P 0 Box
SCIENCE & RESEARCH SERIES NO.86 THE BLUE PENGUIN (Eudyptula minor) AT TAIAROA HEAD, OTAGO, 1992-1993 by Lyndon Perriman and Bruce McKinlay Published by Head Office, Department of Conservation, P 0 Box
CAPTIVE HUSBANDRY AND REPRODUCTION OF THE LEOPARD SNAKE ELAPHE SITULA
 Captive husbandry of Elaphe situla I 123 CAPTIVE HUSBANDRY AND REPRODUCTION OF THE LEOPARD SNAKE ELAPHE SITULA By: Kevin J. Hingley, 22 Busheyfields Road, Dudley, West Midlands, DYl 2LP, England. Contents:
Captive husbandry of Elaphe situla I 123 CAPTIVE HUSBANDRY AND REPRODUCTION OF THE LEOPARD SNAKE ELAPHE SITULA By: Kevin J. Hingley, 22 Busheyfields Road, Dudley, West Midlands, DYl 2LP, England. Contents:
SAMPLE LLI RED SYSTEM BOOK. Intriguing Animals SERIES
 Protecting the Kakapo SAMPLE LLI RED SYSTEM BOOK by Bob Mor rrow Intriguing Animals SERIES Text Elements Genre: Expository Text Structures Main: Chronological Sequence Embedded: Cause/Effect, Problem/
Protecting the Kakapo SAMPLE LLI RED SYSTEM BOOK by Bob Mor rrow Intriguing Animals SERIES Text Elements Genre: Expository Text Structures Main: Chronological Sequence Embedded: Cause/Effect, Problem/
Barn Swallow Nest Monitoring Methods
 Introduction These methods have been developed to guide volunteers in collecting data on the activities and productivity of Barn Swallow nest sites. Effort has been made to standardize these methods for
Introduction These methods have been developed to guide volunteers in collecting data on the activities and productivity of Barn Swallow nest sites. Effort has been made to standardize these methods for
Birds Birds are vertebrates (animals with backbones) with wings and feathers. Most birds can fly, using powerful muscles to flap their wings.
 with wings and feathers. Most birds can fly, using powerful muscles to flap their wings.") Birds Birds are vertebrates (animals with backbones) with wings and feathers. Most birds can fly, using powerful muscles to flap their wings. But a few bird speces do not have strong enough wings to fly,
Birds Birds are vertebrates (animals with backbones) with wings and feathers. Most birds can fly, using powerful muscles to flap their wings. But a few bird speces do not have strong enough wings to fly,
Intraspecific relationships extra questions and answers (Extension material for Level 3 Biology Study Guide, ISBN , page 153)
") i Intraspecific relationships extra questions and answers (Extension material for Level 3 Biology Study Guide, ISBN 978-1-927194-58-4, page 153) Activity 9: Intraspecific relationships extra questions
i Intraspecific relationships extra questions and answers (Extension material for Level 3 Biology Study Guide, ISBN 978-1-927194-58-4, page 153) Activity 9: Intraspecific relationships extra questions
NOTES ON BREEDING SATYR TRAGOPAN TRAGOPAN SATYRA AT SRI CHAMARAJENDRA ZOOLOGICAL GARDENS, MYSORE
 NOTES ON BREEDING SATYR TRAGOPAN TRAGOPAN SATYRA AT SRI CHAMARAJENDRA ZOOLOGICAL GARDENS, MYSORE ABSTRACT Satyr tragopan Tragopan satyra has been classified as Near Threatened in the IUCN Red List of Threatened
NOTES ON BREEDING SATYR TRAGOPAN TRAGOPAN SATYRA AT SRI CHAMARAJENDRA ZOOLOGICAL GARDENS, MYSORE ABSTRACT Satyr tragopan Tragopan satyra has been classified as Near Threatened in the IUCN Red List of Threatened
Breeding Activity Peak Period Range Duration (days) Egg laying Late May to early June Mid-May to mid-july 3 to 10
 Egg laying Late May to early June Mid-May to mid-july 3 to 10") Pernis apivorus 1. INTRODUCTION The honey-buzzard (European honey buzzard) was traditionally regarded as breeding mainly in southern and southwest England, but breeding pairs have been found increasingly
Pernis apivorus 1. INTRODUCTION The honey-buzzard (European honey buzzard) was traditionally regarded as breeding mainly in southern and southwest England, but breeding pairs have been found increasingly
( 162 ) SOME BREEDING-HABITS OF THE LAPWING.
 SOME BREEDING-HABITS OF THE LAPWING.") ( 162 ) SOME BREEDING-HABITS OF THE LAPWING. BY R. H. BROWN. THESE notes on certain breeding-habits of the Lapwing (Vanettus vanellus) are based on observations made during the past three years in Cumberland,
( 162 ) SOME BREEDING-HABITS OF THE LAPWING. BY R. H. BROWN. THESE notes on certain breeding-habits of the Lapwing (Vanettus vanellus) are based on observations made during the past three years in Cumberland,
Species Fact Sheets. Order: Gruiformes Family: Cariamidae Scientific Name: Cariama cristata Common Name: Red-legged seriema
 Order: Gruiformes Family: Cariamidae Scientific Name: Cariama cristata Common Name: Red-legged seriema AZA Management: Green Yellow Red None Photo (Male): Red-legged seriemas are identical in plumage although
Order: Gruiformes Family: Cariamidae Scientific Name: Cariama cristata Common Name: Red-legged seriema AZA Management: Green Yellow Red None Photo (Male): Red-legged seriemas are identical in plumage although
Best Practice in the Breeder House
 Best Practice in the Breeder House Preventing Floor Eggs Best Practice in the Breeder House Preventing Floor Eggs Why are floor eggs a problem? Eggs laid on the floor (floor eggs) have a significantly
Best Practice in the Breeder House Preventing Floor Eggs Best Practice in the Breeder House Preventing Floor Eggs Why are floor eggs a problem? Eggs laid on the floor (floor eggs) have a significantly
Table1. Target lamb pre-weaning daily live weight gain from grazed pasture
 Grassland Management for High Lamb Performance Tim Keady and Noel McNamara Animal & Grassland Research & Innovation Centre, Teagasc, Mellows Campus, Athenry, Co. Galway. To improve the financial margin
Grassland Management for High Lamb Performance Tim Keady and Noel McNamara Animal & Grassland Research & Innovation Centre, Teagasc, Mellows Campus, Athenry, Co. Galway. To improve the financial margin
ROYAL SWAN UPPING The Queen ueen s Diamond Jubilee Edition
 ROYAL SWAN UPPING The Queen s Diamond Jubilee Edition The History of Swan Upping Historically, the reigning King or Queen was entitled to claim ownership of any unmarked mute swans swimming in open water
ROYAL SWAN UPPING The Queen s Diamond Jubilee Edition The History of Swan Upping Historically, the reigning King or Queen was entitled to claim ownership of any unmarked mute swans swimming in open water
This Coloring Book has been adapted for the Wildlife of the Table Rocks
 This Coloring Book has been adapted for the Wildlife of the Table Rocks All images and some writing belong to: Additional writing by: The Table Rocks Environmental Education Program I became the national
This Coloring Book has been adapted for the Wildlife of the Table Rocks All images and some writing belong to: Additional writing by: The Table Rocks Environmental Education Program I became the national
OBSERVATIONS ON A PAIR OF NIGHTJARS AT THE NEST
 OBSERVATIONS ON A PAIR OF NIGHTJARS AT THE NEST By H. R. TUTT INTRODUCTION IN 1952 observations were made at the nest-site of a pair of Nightjars (Caprimulgus europceus) in Essex from the time the young
OBSERVATIONS ON A PAIR OF NIGHTJARS AT THE NEST By H. R. TUTT INTRODUCTION IN 1952 observations were made at the nest-site of a pair of Nightjars (Caprimulgus europceus) in Essex from the time the young
PORTRAIT OF THE AMERICAN BALD EAGLE
 PORTRAIT OF THE AMERICAN BALD EAGLE Objectives: To know the history of the bald eagle and the cause of it's decline. To understand what has been done to improve Bald Eagle habitat. To know the characteristics
PORTRAIT OF THE AMERICAN BALD EAGLE Objectives: To know the history of the bald eagle and the cause of it's decline. To understand what has been done to improve Bald Eagle habitat. To know the characteristics
Breeding Activity Peak Period Range Duration (days) Site occupation and territorial display Early April Mid-March to early May
 Site occupation and territorial display Early April Mid-March to early May") Pandion haliaetus 1. INTRODUCTION The osprey (western osprey) is generally considered to have recolonised Scotland in 1954, after ceasing to breed about 1916 (Thom, 1986). Recently, however, it has been
Pandion haliaetus 1. INTRODUCTION The osprey (western osprey) is generally considered to have recolonised Scotland in 1954, after ceasing to breed about 1916 (Thom, 1986). Recently, however, it has been
THE production of turkey hatching
 The Use of Artificial Lights for Turkeys* H. L. WlLCKE Iowa Agricultural Experiment Station, Ames, Iowa (Presented at Annual Meeting, August 1938; received for publication September 22, 1938) THE production
The Use of Artificial Lights for Turkeys* H. L. WlLCKE Iowa Agricultural Experiment Station, Ames, Iowa (Presented at Annual Meeting, August 1938; received for publication September 22, 1938) THE production
The behaviour of a pair of House Sparrows while rearing young
 The behaviour of a pair of House Sparrows while rearing young By David C. Seel INTRODUCTION IN 1959 OBSERVATIONS were made on the behaviour of a pair of House Sparrows (Passer domesticus) rearing their
The behaviour of a pair of House Sparrows while rearing young By David C. Seel INTRODUCTION IN 1959 OBSERVATIONS were made on the behaviour of a pair of House Sparrows (Passer domesticus) rearing their
Lynx Update May 25, 2009 INTRODUCTION
 Lynx Update May 25, 2009 INTRODUCTION In an effort to establish a viable population of Canada lynx (Lynx canadensis) in Colorado, the Colorado Division of Wildlife (CDOW) initiated a reintroduction effort
Lynx Update May 25, 2009 INTRODUCTION In an effort to establish a viable population of Canada lynx (Lynx canadensis) in Colorado, the Colorado Division of Wildlife (CDOW) initiated a reintroduction effort
Spotlight on rearing:apantesis nais (Drury) (Lepidoptera: Arctiidae) in Louisiana by
 (Lepidoptera: Arctiidae) in Louisiana by") Spotlight on rearing:apantesis nais (Drury) (Lepidoptera: Arctiidae) in Louisiana by Vernon Antoine Brou Jr., 7 Jack Loyd Road, Abita Springs, Louisiana, 7 email: vabrou@bellsouth.net a e j o b c f g k
Spotlight on rearing:apantesis nais (Drury) (Lepidoptera: Arctiidae) in Louisiana by Vernon Antoine Brou Jr., 7 Jack Loyd Road, Abita Springs, Louisiana, 7 email: vabrou@bellsouth.net a e j o b c f g k
Great Horned Owl (Bubo virginianus) Productivity and Home Range Characteristics in a Shortgrass Prairie. Rosemary A. Frank and R.
 Productivity and Home Range Characteristics in a Shortgrass Prairie. Rosemary A. Frank and R.") Great Horned Owl (Bubo virginianus) Productivity and Home Range Characteristics in a Shortgrass Prairie Rosemary A. Frank and R. Scott Lutz 1 Abstract. We studied movements and breeding success of resident
Great Horned Owl (Bubo virginianus) Productivity and Home Range Characteristics in a Shortgrass Prairie Rosemary A. Frank and R. Scott Lutz 1 Abstract. We studied movements and breeding success of resident
The Heartfelt Story of our Backyard Bluebirds
 The Heartfelt Story of our Backyard Bluebirds My husband and I have had the privilege of being landlords to bluebirds for several years and we also monitor bluebird trails. We learn new things about these
The Heartfelt Story of our Backyard Bluebirds My husband and I have had the privilege of being landlords to bluebirds for several years and we also monitor bluebird trails. We learn new things about these
For further information on the biology and ecology of this species, Chapman (1999) provides a comprehensive account.
 provides a comprehensive account.") Falco subbuteo 1. INTRODUCTION The main breeding range of the hobby (Eurasian hobby) in Britain and Ireland lies in England, south of the Mersey/Humber line and extending into the borders of Wales. The
Falco subbuteo 1. INTRODUCTION The main breeding range of the hobby (Eurasian hobby) in Britain and Ireland lies in England, south of the Mersey/Humber line and extending into the borders of Wales. The
Long-eared Owl. For further information on the biology and ecology of this species, Scott (1997) provides a comprehensive account.
 provides a comprehensive account.") Long-eared Owl Asio otus 1. INTRODUCTION The long-eared owl is a widespread but scarce breeding bird in Britain. It was probably more common in the late 19th century, but numbers declined during the 20th
Long-eared Owl Asio otus 1. INTRODUCTION The long-eared owl is a widespread but scarce breeding bird in Britain. It was probably more common in the late 19th century, but numbers declined during the 20th
Activity 4 Building Bird Nests
 Activity 4 Building Bird Nests Created By Point Reyes Bird Observatory Education Program Building Bird Nests Activity 4 Objective: To teach students about songbird nests, the different types, placement
Activity 4 Building Bird Nests Created By Point Reyes Bird Observatory Education Program Building Bird Nests Activity 4 Objective: To teach students about songbird nests, the different types, placement
Beefy and the beast Special edition, March 2010
 Department of Employment, Economic Development and Innovation Biosecurity Queensland Beefy and the beast Special edition, March 2010 This special edition of Beefy and the beast summarises the findings
Department of Employment, Economic Development and Innovation Biosecurity Queensland Beefy and the beast Special edition, March 2010 This special edition of Beefy and the beast summarises the findings
Red-Tailed Hawk Buteo jamaicensis
 Red-Tailed Hawk Buteo jamaicensis This large, dark headed, broad-shouldered hawk is one of the most common and widespread hawks in North America. The Red-tailed hawk belongs to the genus (family) Buteo,
Red-Tailed Hawk Buteo jamaicensis This large, dark headed, broad-shouldered hawk is one of the most common and widespread hawks in North America. The Red-tailed hawk belongs to the genus (family) Buteo,
Koala Monitoring Program
 Koala Monitoring Program Yarrabilba Priority Development Area Koala Capture / Monitoring Event October 2017 Summary Report Female 13486 (Jean) with her back young Prepared by: Koala Ecology Group University
Koala Monitoring Program Yarrabilba Priority Development Area Koala Capture / Monitoring Event October 2017 Summary Report Female 13486 (Jean) with her back young Prepared by: Koala Ecology Group University
BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS
 Nov., 1965 505 BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS Lack ( 1954; 40-41) has pointed out that in species of birds which have asynchronous hatching, brood size may be adjusted
Nov., 1965 505 BROOD REDUCTION IN THE CURVE-BILLED THRASHER By ROBERTE.RICKLEFS Lack ( 1954; 40-41) has pointed out that in species of birds which have asynchronous hatching, brood size may be adjusted
APPENDICES APPENDIX 1: TERRITORIALITY AND AGE AT FIRST BREEDING FOR O.N.E KIWI Age of first breeding (years)
") APPENDICES APPENDIX 1: TERRITORIALITY AND AGE AT FIRST BREEDING FOR O.N.E KIWI 2002-2012 Gender Age of territoriality (years) Age of first breeding (years) Cohort Outcome or Current age (years) male (pair
APPENDICES APPENDIX 1: TERRITORIALITY AND AGE AT FIRST BREEDING FOR O.N.E KIWI 2002-2012 Gender Age of territoriality (years) Age of first breeding (years) Cohort Outcome or Current age (years) male (pair
Managing a Small Poultry Flock
 Managing a Small Poultry Flock Commitment Caring for chickens can be rewarding, but poultry are fragile and need to be well cared for at all times. Breed Selection Cornish are recommended for meat and
Managing a Small Poultry Flock Commitment Caring for chickens can be rewarding, but poultry are fragile and need to be well cared for at all times. Breed Selection Cornish are recommended for meat and
PEREGRINE FALCON HABITAT MANAGEMENT GUIDELINES ONTARIO MINISTRY OF NATURAL RESOURCES
 PEREGRINE FALCON HABITAT MANAGEMENT GUIDELINES ONTARIO MINISTRY OF NATURAL RESOURCES December 1987 2 Table of Contents Page Introduction...3 Guidelines...4 References...7 Peregrine Falcon Nest Site Management
PEREGRINE FALCON HABITAT MANAGEMENT GUIDELINES ONTARIO MINISTRY OF NATURAL RESOURCES December 1987 2 Table of Contents Page Introduction...3 Guidelines...4 References...7 Peregrine Falcon Nest Site Management
EGG production of turkeys is not important
 A Study of Egg Production in Bronze Turkeys S. J. MAESDEN National Agricultural Research Center, Beltsville, Maryland EGG production of turkeys is not important commercially but good egg production during
A Study of Egg Production in Bronze Turkeys S. J. MAESDEN National Agricultural Research Center, Beltsville, Maryland EGG production of turkeys is not important commercially but good egg production during
(170) COURTSHIP AND DISPLAY OF THE SLAVONIAN GREBE.
 COURTSHIP AND DISPLAY OF THE SLAVONIAN GREBE.") (170) COURTSHIP AND DISPLAY OF THE SLAVONIAN GREBE. BY ERIC J. HOSKING, F.R.P.S., M.B.O.U. (Plates 4 and 5.) DURING the nesting season of 1939 I was staying in Scotland and had the opportunity of witnessing
(170) COURTSHIP AND DISPLAY OF THE SLAVONIAN GREBE. BY ERIC J. HOSKING, F.R.P.S., M.B.O.U. (Plates 4 and 5.) DURING the nesting season of 1939 I was staying in Scotland and had the opportunity of witnessing
POULTRY STANDARDS The focus of PROOF certification is the on. farm management of livestock in a farming
 The focus of PROOF certification is the on farm management of livestock in a farming system that provides unrestricted daytime access to actively managed, pastured range areas in an environment that encourages
The focus of PROOF certification is the on farm management of livestock in a farming system that provides unrestricted daytime access to actively managed, pastured range areas in an environment that encourages
Trunk Contents. Crane Flight Feathers (3)
") Trunk Contents Learning occurs not only with the mind, but also with the eyes, the hands the whole child (or adult!). Items contained in the trunk are meant to be examined, handled, and shared with your
Trunk Contents Learning occurs not only with the mind, but also with the eyes, the hands the whole child (or adult!). Items contained in the trunk are meant to be examined, handled, and shared with your
COLORADO LYNX DEN SITE HABITAT PROGRESS REPORT 2006
 COLORADO LYNX DEN SITE HABITAT PROGRESS REPORT 2006 by Grant Merrill Tanya Shenk U.S. Forest Service and Colorado Division of Wildlife Cooperative Effort September 30, 2006 INTRODUCTION Lynx (Lynx canadensis)
COLORADO LYNX DEN SITE HABITAT PROGRESS REPORT 2006 by Grant Merrill Tanya Shenk U.S. Forest Service and Colorado Division of Wildlife Cooperative Effort September 30, 2006 INTRODUCTION Lynx (Lynx canadensis)
By Hans Frey ¹ ² & Alex Llopis ²
 1/7 By Hans Frey ¹ ² & Alex Llopis ² ¹ Verein EGS-Eulen und Greifvogelschutz, Untere Hauptstraße 34, 2286 Haringsee, Austria. Phone number +43 2214 84014 h.frey@4vultures.org ² Vulture Conservation Foundation
1/7 By Hans Frey ¹ ² & Alex Llopis ² ¹ Verein EGS-Eulen und Greifvogelschutz, Untere Hauptstraße 34, 2286 Haringsee, Austria. Phone number +43 2214 84014 h.frey@4vultures.org ² Vulture Conservation Foundation
Reproductive physiology and eggs
 Reproductive physiology and eggs Class Business Reading for this lecture Required. Gill: Chapter 14 1. Reproductive physiology In lecture I will only have time to go over reproductive physiology briefly,
Reproductive physiology and eggs Class Business Reading for this lecture Required. Gill: Chapter 14 1. Reproductive physiology In lecture I will only have time to go over reproductive physiology briefly,
DIARY OF A COUGAR/MULE DEER ENCOUNTER
 DIARY OF A COUGAR/MULE DEER ENCOUNTER September 7, 2006. Setting: west-facing slope at elevation 7000 feet in the foothills west of Denver, Colorado. Sunny day, warm. several mule deer browsing in Mahogany
DIARY OF A COUGAR/MULE DEER ENCOUNTER September 7, 2006. Setting: west-facing slope at elevation 7000 feet in the foothills west of Denver, Colorado. Sunny day, warm. several mule deer browsing in Mahogany
Project Snip and Tip
 Project Snip and Tip Guidelines for Trap Neuter - Return Caloosa Humane Society is the administrator of this program. We will organize the trapping, transport, vetting and release of feral cats. Cats must
Project Snip and Tip Guidelines for Trap Neuter - Return Caloosa Humane Society is the administrator of this program. We will organize the trapping, transport, vetting and release of feral cats. Cats must
LEVEL 2 AWARD IN THE SAFE USE OF RODENTICIDES
 LEVEL 2 AWARD IN THE SAFE USE OF RODENTICIDES Specimen Paper PAPER NUMBER: 0000 INSTRUCTIONS TO CANDIDATES 1 Attempt to answer every question. 2. Enter your answers on the accompanying answer sheet. Each
LEVEL 2 AWARD IN THE SAFE USE OF RODENTICIDES Specimen Paper PAPER NUMBER: 0000 INSTRUCTIONS TO CANDIDATES 1 Attempt to answer every question. 2. Enter your answers on the accompanying answer sheet. Each
Emergency Below the Ice Shelf. Narrative (Imaginative) Presenting a School Speech Procedure (Informative) The School of the Air
 Presenting a School Speech Procedure (Informative) The School of the Air") guided reading cards About Level 23 Cards Our Superhero Chickens Recount (Imaginative) Travelling to School Description (Informative) Level 24 Cards Join the Tree-Planting Project! Exposition (Persuasive)
guided reading cards About Level 23 Cards Our Superhero Chickens Recount (Imaginative) Travelling to School Description (Informative) Level 24 Cards Join the Tree-Planting Project! Exposition (Persuasive)
Mauritania. 1 May 25 October Desert Locust Information Service FAO, Rome outbreak
 Mauritania 1 May 25 October 2016 outbreak Desert Locust Information Service FAO, Rome www.fao.org/ag/locusts Keith Cressman (Senior Locust Forecasting Officer) updated: 26 Oct 2016 Zouerate oases May 2016
Mauritania 1 May 25 October 2016 outbreak Desert Locust Information Service FAO, Rome www.fao.org/ag/locusts Keith Cressman (Senior Locust Forecasting Officer) updated: 26 Oct 2016 Zouerate oases May 2016
Slide 1 NO NOTES. Slide 2 NO NOTES. Slide 3 NO NOTES. Slide 4 NO NOTES. Slide 5
 Slide 1 Slide 2 Slide 3 Slide 4 Slide 5 Left is broiler (for meat) bird (Cobb/Ross), have different nutritional needs to layers. From chick to kill can be as little as 34 days. Commercial layer (ends up
Slide 1 Slide 2 Slide 3 Slide 4 Slide 5 Left is broiler (for meat) bird (Cobb/Ross), have different nutritional needs to layers. From chick to kill can be as little as 34 days. Commercial layer (ends up
Vancouver Bald Eagle Report 2013
 Vancouver Bald Eagle Report 2013 August 2013 Eagle perches unabashedly despite approaching gull Photo by: Martin Passchier Stanley Park Ecology Society has monitored bald eagle nests during the breeding
Vancouver Bald Eagle Report 2013 August 2013 Eagle perches unabashedly despite approaching gull Photo by: Martin Passchier Stanley Park Ecology Society has monitored bald eagle nests during the breeding
Pup 61 within seconds of being born in South Haven, 22 September 2012
 Pup 61 within seconds of being born in South Haven, 22 September 212 SUMMARY 182 pups were born on Skomer in 212, the highest total ever recorded, 18 in 1993 being the previous best. 31 pups were born
Pup 61 within seconds of being born in South Haven, 22 September 212 SUMMARY 182 pups were born on Skomer in 212, the highest total ever recorded, 18 in 1993 being the previous best. 31 pups were born
Breeding the Common Golden-Backed Woodpecker in Captivity Dinopium javanense
 Breeding the Common Golden-Backed Woodpecker in Captivity Dinopium javanense Michelene M. O Connor, Zookeeper-Aviary Milwaukee County Zoological Gardens 10001 W. Bluemound Rd. Milwaukee, WI 53226 Sharpbill@aol.com
Breeding the Common Golden-Backed Woodpecker in Captivity Dinopium javanense Michelene M. O Connor, Zookeeper-Aviary Milwaukee County Zoological Gardens 10001 W. Bluemound Rd. Milwaukee, WI 53226 Sharpbill@aol.com
SEASONAL CHANGES IN A POPULATION OF DESERT HARVESTMEN, TRACHYRHINUS MARMORATUS (ARACHNIDA: OPILIONES), FROM WESTERN TEXAS
, FROM WESTERN TEXAS") Reprinted from PSYCHE, Vol 99, No. 23, 1992 SEASONAL CHANGES IN A POPULATION OF DESERT HARVESTMEN, TRACHYRHINUS MARMORATUS (ARACHNIDA: OPILIONES), FROM WESTERN TEXAS BY WILLIAM P. MACKAY l, CHE'REE AND
Reprinted from PSYCHE, Vol 99, No. 23, 1992 SEASONAL CHANGES IN A POPULATION OF DESERT HARVESTMEN, TRACHYRHINUS MARMORATUS (ARACHNIDA: OPILIONES), FROM WESTERN TEXAS BY WILLIAM P. MACKAY l, CHE'REE AND
Growth and Development. Embryonic development 2/22/2018. Timing of hatching. Hatching. Young birds and their parents
 Growth and Development Young birds and their parents Embryonic development From fertilization to hatching, the embryo undergoes sequence of 42 distinct developmental stages The first 33 stages vary little
Growth and Development Young birds and their parents Embryonic development From fertilization to hatching, the embryo undergoes sequence of 42 distinct developmental stages The first 33 stages vary little
FALL 2015 BLACK-FOOTED FERRET SURVEY LOGAN COUNTY, KANSAS DAN MULHERN; U.S. FISH AND WILDLIFE SERVICE
 INTRODUCTION FALL 2015 BLACK-FOOTED FERRET SURVEY LOGAN COUNTY, KANSAS DAN MULHERN; U.S. FISH AND WILDLIFE SERVICE As part of ongoing efforts to monitor the status of reintroduced endangered black-footed
INTRODUCTION FALL 2015 BLACK-FOOTED FERRET SURVEY LOGAN COUNTY, KANSAS DAN MULHERN; U.S. FISH AND WILDLIFE SERVICE As part of ongoing efforts to monitor the status of reintroduced endangered black-footed
Chatham Island Mollymawk research on Te Tara Koi Koia: November 2016
 Chatham Island Mollymawk research on Te Tara Koi Koia: November 2016 1 Chatham Island Mollymawk research on Te Tara Koi Koia: November 2016 Mike Bell, Dave Bell and Dave Boyle Wildlife Management International
Chatham Island Mollymawk research on Te Tara Koi Koia: November 2016 1 Chatham Island Mollymawk research on Te Tara Koi Koia: November 2016 Mike Bell, Dave Bell and Dave Boyle Wildlife Management International
Ames, IA Ames, IA (515)
") BENEFITS OF A CONSERVATION BUFFER-BASED CONSERVATION MANAGEMENT SYSTEM FOR NORTHERN BOBWHITE AND GRASSLAND SONGBIRDS IN AN INTENSIVE PRODUCTION AGRICULTURAL LANDSCAPE IN THE LOWER MISSISSIPPI ALLUVIAL
BENEFITS OF A CONSERVATION BUFFER-BASED CONSERVATION MANAGEMENT SYSTEM FOR NORTHERN BOBWHITE AND GRASSLAND SONGBIRDS IN AN INTENSIVE PRODUCTION AGRICULTURAL LANDSCAPE IN THE LOWER MISSISSIPPI ALLUVIAL
Kori Bustard Husbandry. Sara Hallager, Biologist, Smithsonian National Zoological Park
 Kori Bustard Husbandry Sara Hallager, Biologist, Smithsonian National Zoological Park Ardeotis kori 2 subspecies [?] Africa s largest flying bird Captive males: 12-19kg Seasonal weight gain up to 4kg Captive
Kori Bustard Husbandry Sara Hallager, Biologist, Smithsonian National Zoological Park Ardeotis kori 2 subspecies [?] Africa s largest flying bird Captive males: 12-19kg Seasonal weight gain up to 4kg Captive
Ciccaba virgata (Mottled Owl)
") Ciccaba virgata (Mottled Owl) Family: Strigidae (Typical Owls) Order: Strigiformes (Owls) Class: Aves (Birds) Fig. 1. Mottled owl, Ciccaba virgata. [http://www.owling.com/mottled13.htm, downloaded 12 November
Ciccaba virgata (Mottled Owl) Family: Strigidae (Typical Owls) Order: Strigiformes (Owls) Class: Aves (Birds) Fig. 1. Mottled owl, Ciccaba virgata. [http://www.owling.com/mottled13.htm, downloaded 12 November
Bald Eagles in the Yukon. Wildlife in our backyard
 Bald Eagles in the Yukon Wildlife in our backyard The Bald Eagle at a glance Both male and female adult Bald Eagles have a dark brown body and wings with a white head, neck and tail. They have a yellow
Bald Eagles in the Yukon Wildlife in our backyard The Bald Eagle at a glance Both male and female adult Bald Eagles have a dark brown body and wings with a white head, neck and tail. They have a yellow
Sparrowhawks & Goshawks and the Gymnogene
 1 Module # 6 Component # 7 Sparrowhawks & Goshawks and the Gymnogene Sparrowhawks and Goshawks There are nine Southern African species in this group, these are the: Ovambo Sparrowhawk Little Sparrowhawk
1 Module # 6 Component # 7 Sparrowhawks & Goshawks and the Gymnogene Sparrowhawks and Goshawks There are nine Southern African species in this group, these are the: Ovambo Sparrowhawk Little Sparrowhawk
Glossy Black Conservancy News No. 15 November 2015
 New Arrival at Currumbin Sanctuary In June this year Currumbin Wildlife Sanctuary, a friend of the Conservancy, successfully bred their first Glossy Black- Cockatoo chick! The parents had previously had
New Arrival at Currumbin Sanctuary In June this year Currumbin Wildlife Sanctuary, a friend of the Conservancy, successfully bred their first Glossy Black- Cockatoo chick! The parents had previously had
SEABIRD, SHARK, AND MARINE MAMMAL RESEARCH PLANS AND PROTOCOLS FOR SOUTHEAST FARALLON ISLAND
 1 SEABIRD, SHARK, AND MARINE MAMMAL RESEARCH PLANS AND PROTOCOLS FOR SOUTHEAST FARALLON ISLAND Seabirds Ashy Storm-Petrel: 1. Nest Site Maintenance After 15 March, check the status and condition of all
1 SEABIRD, SHARK, AND MARINE MAMMAL RESEARCH PLANS AND PROTOCOLS FOR SOUTHEAST FARALLON ISLAND Seabirds Ashy Storm-Petrel: 1. Nest Site Maintenance After 15 March, check the status and condition of all
Research report. Population study of the North Island Robin (Petroica australis longipes) in the Waitakere ranges in New Zealand.
 in the Waitakere ranges in New Zealand.") Research report Population study of the North Island Robin (Petroica australis longipes) in the Waitakere ranges in New Zealand Masha Leenen Population study of the North Island Robin (Petroica australis
Research report Population study of the North Island Robin (Petroica australis longipes) in the Waitakere ranges in New Zealand Masha Leenen Population study of the North Island Robin (Petroica australis
The Essex County Field Naturalists' Club's BLUEBIRD COMMITTEE REPORT FOR 2017
 The Essex County Field Naturalists' Club's BLUEBIRD COMMITTEE REPORT FOR 2017 The Bluebirds had a fair year, in 2017. We counted 22 successful pairs of Bluebirds which produced 101 fledglings. This is
The Essex County Field Naturalists' Club's BLUEBIRD COMMITTEE REPORT FOR 2017 The Bluebirds had a fair year, in 2017. We counted 22 successful pairs of Bluebirds which produced 101 fledglings. This is
DISEASE MONITORING AND EXTENSION SYSTEM FOR THE SOUTH AFRICAN DAIRY INDUSTRY
 DISEASE MONITORING AND EXTENSION SYSTEM FOR THE SOUTH AFRICAN DAIRY INDUSTRY Disease Trend Report: July 2014 IN THIS ISSUE: 1. Preface Importance of disease monitoring. 2. Get the vaccination plan in place
DISEASE MONITORING AND EXTENSION SYSTEM FOR THE SOUTH AFRICAN DAIRY INDUSTRY Disease Trend Report: July 2014 IN THIS ISSUE: 1. Preface Importance of disease monitoring. 2. Get the vaccination plan in place
Thefirst attempt at Brood Manipulation of the Golden Eagle Aquila chrysaetos in Japan
 Meyburg. B-U. & R. D. Chancellor eds. 19% Eagle Studies World Working CJroup on Birds of Prey (WW(JBP) Berlin, London & Paris Thefirst attempt at Brood Manipulation of the Golden Eagle Aquila chrysaetos
Meyburg. B-U. & R. D. Chancellor eds. 19% Eagle Studies World Working CJroup on Birds of Prey (WW(JBP) Berlin, London & Paris Thefirst attempt at Brood Manipulation of the Golden Eagle Aquila chrysaetos
A Survey of Recent Husbandry and Breeding Techniques of Curassows at the Houston Zoo.
 WWW.AVIANSAG.ORG A Survey of Recent Husbandry and Breeding Techniques of Curassows at the Houston Zoo by Rene Ryan Bird Keeper, Houston Zoo rene.m.ryan@gmail.com Introduction We ll be covering activity
WWW.AVIANSAG.ORG A Survey of Recent Husbandry and Breeding Techniques of Curassows at the Houston Zoo by Rene Ryan Bird Keeper, Houston Zoo rene.m.ryan@gmail.com Introduction We ll be covering activity
The grey partridges of Nine Wells: A five-year study of a square kilometre of arable land south of Addenbrooke s Hospital in Cambridge
 The grey partridges of Nine Wells: 2012 2016 A five-year study of a square kilometre of arable land south of Addenbrooke s Hospital in Cambridge John Meed, January 2017 1 Introduction Grey partridge populations
The grey partridges of Nine Wells: 2012 2016 A five-year study of a square kilometre of arable land south of Addenbrooke s Hospital in Cambridge John Meed, January 2017 1 Introduction Grey partridge populations
Yemen. JUNE Present. Desert Locust Information Service FAO, Rome 21 Feb 2014 update
 Yemen JUNE 0 - Present Feb 0 update Desert Locust Information Service FAO, Rome www.fao.org/ag/locusts spring-bred (June) SAUDI ARABIA W. Dawasir swarm invasion (June) Shehan Thamud Sayun Marib summer
Yemen JUNE 0 - Present Feb 0 update Desert Locust Information Service FAO, Rome www.fao.org/ag/locusts spring-bred (June) SAUDI ARABIA W. Dawasir swarm invasion (June) Shehan Thamud Sayun Marib summer
Guide for Incubation of Eggs and Care of Newborn Livestock
 Guide for Incubation of Eggs and Care of Newborn Livestock Goffle Road Poultry Farm 549 Goffle Road, Wyckoff, N.J. 07481 www.gofflepoultry.com Version_020518 Roadmap This presentation will cover how to
Guide for Incubation of Eggs and Care of Newborn Livestock Goffle Road Poultry Farm 549 Goffle Road, Wyckoff, N.J. 07481 www.gofflepoultry.com Version_020518 Roadmap This presentation will cover how to
Montgomery County Animal Care and Control
 Montgomery County Animal Care and Control 2018 Annual Report Field and Shelter Activity 1 Montgomery County Animal Care & Control 2018 Field and Shelter Activity Report Table of Contents Introduction 3
Montgomery County Animal Care and Control 2018 Annual Report Field and Shelter Activity 1 Montgomery County Animal Care & Control 2018 Field and Shelter Activity Report Table of Contents Introduction 3